
Submitted:
02 March 2024
Posted:
04 March 2024
You are already at the latest version
Abstract
Inflammatory bowel disease (IBD), a heterogeneous group of recurring inflammatory condi-tions of the digestive system that encompass both ulcerative colitis (UC) and Crohn’s disease (CD), pose a significant public health challenge, currently lacking a definitive cure. The specif-ic etiopathogenesis of IBD is not yet fully understood, but a multifactorial interplay of genetic and environmental factors is suspected. A growing body of evidence supports the involve-ment of intestinal dysbiosis in the development of IBD, including the effects of dysbiosis on the integrity of the intestinal epithelial barrier, modulation of the host immune system, alter-ations in the enteric nervous system, and the perpetuation of chronic inflammation. A com-prehensive understanding of these mechanisms is important to define preventive measures, to develop new effective and lasting treatments and to improve disease outcome. This review examines the complex tri-directional relationship between gut microbiota, mucosal immune system, and intestinal epithelium in IBD. In addition, nonpharmacological and behavioral strategies aimed at restoring a proper microbial-immune relationship will be suggested.
Keywords:
IBD
; Crohn’s disease
; ulcerative colitis
; immunological modulation
; microbiota
; micobiome
; inflammation
; pathogenesis
; flare immunity
1. Introduction
Inflammatory bowel diseases (IBD) are chronic and recurring inflammatory conditions, encompassing Crohn’s disease (CD) and ulcerative colitis (UC) [1,2]. While both share similar clinical symptoms, CD can affect any part of the gastrointestinal tract, whereas UC is limited to the colon and rectum [1,2]. They are stratified into mild, moderate, and severe categories based on clinical and endoscopic evaluations [3]. These diseases are marked by persistent inflammation, leading to complications such as hospitalization, surgery, and disability, significantly impacting the quality of life of affected individuals [4,5]. Furthermore, colitis promotes carcinogenesis by fostering the expansion of genotoxic bacteria, with UC patients facing a 20% risk of colorectal cancer (CRC) development and CD patients a 1.2% risk compared to general population [6]. Timely and effective treatment is crucial to prevent these complications and improve the well-being of IBD patients [7].
Globally, approximately 6.8 million individuals suffer from IBD, with an increasing incidence in developing nations and a stable trend in highly prevalent developed countries, imposing significant strain on healthcare resources [8,9]. Although the etiology of IBD remains elusive, emerging evidence suggests that genetic susceptibility, defects in mucosal barrier function, immune stimulation, and alterations in microbial composition and function of the intestinal environment contribute to its pathogenesis [10]. This is supported by the identification of over 200 IBD-associated susceptibility genes involved in host-microbiota interactions [11]. Consequently, the gut microbiota has emerged as both a cause and consequence of IBD, attracting considerable attention in research on IBD pathogenesis and biological therapies [12].
This review aims to synthesize the intricate interplay between the gut microbiota and the initiation and progression of IBD, with implications for diagnosis, treatment, and prevention strategies.
2. Immune Dysregulation in IBD
The complex interaction among immune cells, cytokines, and molecular pathways lies at the heart of immune dysregulation in IBD, characterized by excessive activation of pro-inflammatory pathways and inadequate regulation of anti-inflammatory mechanisms [13,14]. Various immune cell types and signaling molecules contribute to this imbalance [13]. In CD,heightened production of pro-inflammatory cytokines like tumor necrosis factor alpha (TNF-α), interleukin (IL)-1β, and IL-6 sustains chronic inflammation, while in UC, abnormal activation of immune cells including T-cells, macrophages, and dendritic cells (DCs) leads to persistent mucosal inflammation [15,16].
Innate immune dysfunction plays a significant role, with DCs, macrophages, and epithelial cells detecting and responding to microbial components through pattern recognition receptors (PRRs) such as Toll-like receptors (TLRs) and nucleotide-binding oligomerization domain (NOD)-like receptors (NLRs) [17]. Dysregulation of these innate immune pathways can trigger an exaggerated immune response, contributing to chronic intestinal inflammation [17]. Genetic studies have identified susceptibility loci associated with CD, including NOD2/caspase recruitment domains (CARD)15, autophagy-related (ATG)16L1, and IL-23R, while genome-wide association studies in UC have identified genes like IL-23R, IL-10, and human leukocyte antigen (HLA) genes [18,19,20]. These genetic variants influence innate immune responses, autophagy, and the balance of pro- and anti-inflammatory cytokines [14,18,19,20].
Intestinal infections may also contribute to perpetuating inflammation [21]. Episodes of Salmonella/Campylobacter gastroenteritis have been linked to an increased risk of developing IBD, with alterations in the TLR4 gene potentially predisposing individuals to infections by these Gram-negative bacteria and increasing susceptibility to enteric infections in general [22,23]. Consequently, pathogenic infections may alter the composition of the commensal gut microbiota and disrupt commensal tolerance, leading to chronic inflammation associated with IBD [24].
Furthermore, environmental factors such as smoking, diet, and the gut microbiota significantly contribute to IBD pathogenesis and progression through epigenetic modifications like DNA methylation, histone modifications, and regulation of noncoding RNA [25]. These modifications alter gene expression patterns and immune responses in the intestinal mucosa, affecting epithelial barrier function and inflammatory pathways [25].
Lastly, bacterial metabolites may directly influence pro- and anti-inflammatory pathways, further contributing to the complex pathogenesis of IBD [26].
3. The Role of Gut Microbiota in IBD
The human gastrointestinal tract hosts a diverse ecosystem of microorganisms collectively known as the gut microbiota, comprising approximately 40 trillion microorganisms and containing about 150 times more genes than the human genome [27]. The gut microbiota consists of beneficial bacteria that contribute to gut mucosal homeostasis, as well as harmful bacteria, known as “pathobionts”, which can trigger gut inflammation and mucosal damage [27]. As a crucial component of the intestinal barrier, the gut microbiota has evolved alongside the host’s intestinal environment, contributing to the maintenance of epithelial mucosal homeostasis, immune regulation, metabolic balance, and nutrient provision in a healthy state [28]. It also plays a role in sustaining intestinal structure and defending against opportunistic pathogen invasion [28]. However, dysregulation of these interactions can lead to inflammation-related diseases [29].
The composition and function of the gut microbiota can be influenced by various factors such as diet, drug treatment, smoking, age, and genetics [30,31]. These factors may alter the symbiotic interplay between the microbiota and the host, contributing to the pathogenesis of inflammation-related diseases like IBD [32]. Disturbances in the balance between beneficial and harmful bacteria, as well as reductions in biodiversity and species richness within the microbial community, can disrupt physiological gut homeostasis and damage the intestinal mucosal barrier [33].
The integrity of the intestinal barrier is maintained through the interactions of various components, including the mucus layer, immunoglobulin A (IgA), antimicrobial peptides (AMPs), and intercellular tight junctions (TJs) [34]. The mucus layer serves to stabilize the intestinal lining, while AMPs, produced by intestinal epithelial cells (IECs), regulate microbial colonization in the gut lumen and prevent the infiltration of epithelial cells [34]. Intercellular TJs, comprised of proteins located near the apical membrane of epithelial cells, play a crucial role in determining the physical integrity of the intestinal barrier [34]. Claudin proteins, integral components of TJs, serve as essential defenses against pathogen invasion [35]. Abnormal expression of claudin can result in reduced cell adhesion, structural damage, and impaired function of both epithelial and endothelial cells [36].
Disruption of these interactions, commonly referred to as “leaky gut,” can exacerbate intestinal inflammation by allowing bacteria to translocate into the lamina propria, triggering an inflammatory response mediated by TLRs and NF-κB pathways [37]. This leads to the proliferation of pro-inflammatory T-cell subsets, including T helper Th1, Th2, and Th17 cells, which produce various pro-inflammatory cytokines, chemokines, and other mediators [37,38]. These molecules recruit pro-inflammatory cells to the gut mucosa, contributing to mucosal damage, while concurrently downregulating anti-inflammatory mediators such as T regulatory cells (Tregs) [33]. In this altered gut mucosa environment, “pathobionts,” thrive and further enhance the immune response [39] (Figure 1).
This complex pathological mechanism involving the immune system and gut microbiota is driven by the activation of specific receptors on immune cells, such as PRRs, which recognize pathogen-associated molecular patterns (PAMPs) and danger-associated molecular patterns (DAMPs) [40,41]. These receptors initiate innate immune responses against infectious agents [40,41] (Figure 2).
Comparing microbiota across individuals is challenging due to this complexity [42]. Additionally, while various molecular techniques have been employed to study the human gut microbiota, none can capture the full spectrum of microorganisms inhabiting the gut [43]. Nevertheless, consensus can be reached by cross-referencing studies employing different techniques and experimental protocols [44]. Through comparative analysis of cases and controls, recurring patterns emerge, offering a comprehensive insight into gut microbiota dynamics [45].
3.1. Bacteria in IBD
Escherichia coli (E. coli) is a gram-negative, facultative anaerobic bacterium commonly found in the lower intestine of warm-blooded organisms [46]. Elevated levels of E. coli have been observed in the IECs of patients with CD and UC [47]. Specifically, Adherent-invasive E. coli (AIEC) has been implicated in the early stages of IBD development [47]. AIEC can adhere to and traverse the intestinal mucosa, survive and multiply within macrophages, leading to the release of TNF, thereby increasing the permeability of the intestinal epithelium [47,48]. Additionally, it is associated with tissue damage resulting from the activation of the pro-inflammatory and tumorigenic transcription factor signal transducer and activator of transcription STAT3, along with the pro-inflammatory cytokine IL-17 [49]. Furthermore, E. coli has been shown to release colibactin, which damages DNA in IECs, potentially contributing to the increased susceptibility of IBD patients to CRC [50].
Bacteroides fragilis (B. fragilis) is a gram-negative anaerobic bacterium that typically exists as a commensal in the gut microbiota [51]. Strains of B. fragilis expressing a zinc-dependent metalloprotease known as B. fragilis toxin (BFT or fragilysin) are referred to as enterotoxigenic B. fragilis (ETBF) [51]. ETBF acts as an opportunistic pathogen with pro-inflammatory properties that contribute to IBD [52]. BFT directly impacts signalling pathways such as Wnt, NF-κB, STAT3, and MAPK, resulting in elevated levels of Th17 cells, Tregs, and pro-inflammatory mediators, thereby increasing mucosal permeability [52]. Additionally, ETBF disrupts the colonic epithelial barrier by cleaving the zonula adherens protein E-cadherin [52]. Finally, ETBF induces the production of reactive oxygen species (ROS) and DNA damage by upregulating the expression of spermine oxidase in colonocytes [53].
Other bacteria, such as Enterococcus faecalis and Streptococcus bovis, have been implicated in promoting cytokine expression and inflammation in the colon, leading to the development of IBD [54,55]. Enterococcus faecalis is known for its production of damaging reactive oxygen species (ROS) [56]. Additionally, experimental models have shown that IL-10 knock-out mice are more susceptible to developing IBD when exposed to Enterococcus faecalis [57]. Streptococcus bovis is associated with the production of pro-inflammatory and pro-angiogenic cytokines, including IL-6, IL-8, and IL-17 [54].
Fusobacterium nucleatum, another pro-inflammatory bacterium, activates epithelial TLR4, which induces inflammation [58]. It has also been found to be abundant in the colonic mucosa of UC patients [59].
Faecalibacterium prausnitzii (F. praunitzii) and Roseburia hominis has garnered significant attention in recent years due to their status of butyrate-producing bacteria inhabiting the gastrointestinal tract [60]. A decline in thier abundance has been noted in individuals with IBD [61]. In various in vivo models of chemically induced colitis, F. prausnitzii has been shown to mitigate the severity of intestinal inflammation by producing butyrate [62]. Butyrate helps to maintain a balance between Th17 and Tregs, promotes the production of anti-inflammatory cytokines such as interleukin IL-10, and inhibits NF-κB signaling, as well as the production of IL-8, IL-12, and interferon-γ [63].
Several other bacterial species within the Clostridium, Lactobacillus, and Bifidobacterium genera also appear to have beneficial effects against IBD [64,65]. For example, various strains of Clostridia can promote the development of Tregs in the colonic mucosa, potentially protecting against colitis [66]. Bifidobacteria have been shown to increase the secretion of TJs from intestinal cells, thereby improving symptoms in IBD mice with shortened intestines [67]. Last but not least, Akkermansia muciniphila, a mucophilic bacterium, can increase the numbers of goblet cells and mucin families in the intestinal epithelium, providing protection against IBD [68] (Figure 3).
3.2. Viruses in IBD
Unlike bacterial diversity, increased virome diversity has been noted in IBD patients, suggesting the involvement of the gut virome in bacterial dysbiosis [69] (Figure 4). For their evolutionary characteristics, viruses can be classified into bacteriophages and eukaryotic-targeting viruses, both of which may possess single-stranded or double-stranded RNA or DNA genomes [70]. Bacteriophages traditionally influence bacterial composition directly, while eukaryotic viruses can interact with both human host cells and other eukaryotic components of the microbiota, such as fungi [70]. Alterations in gut virome structure have been implicated in the onset and severity of IBD [69].
Specifically, metagenomics analysis has revealed an increase in bacteriophages of the family Caudovirales and a decrease in Microviridae [69]. Elevated levels of Caudovirales have been observed in patients with UC and CD,with similar findings in gut biopsies of IBD patients compared to controls [71]. The virome can impact other components of the microbiota, as reported in the study by Norman et al., which described the expansion of Caudovirales bacteriophages in IBD compared with controls, coupled with decreased bacterial richness and diversity, two hallmarks of IBD-associated intestinal dysbiosis [71]. Increased viral diversity has also been correlated with gut dysbiosis and pro-inflammatory cytokine levels in mouse models [72]. Studies have reported the enrichment of bacteriophages Caudovirales and Podoviridae in mouse models of colitis [72]. Furthermore, elevated bacteriophage abundance has been linked to exacerbated colitis in germ-free mice [72]. Similarly, the dynamics of bacteriophages-bacteria parasitism have been studied in the case of Faecalibacterium prausnitzii, a bacterium generally depleted in IBD, whose low abundance in IBD has been associated with a higher prevalence of F. prausnitzii phages compared with controls, suggesting enhanced phage-mediated mortality of F. prausnitzii in IBD [73].
Eukaryotic viruses have also been associated with early stages of intestinal inflammation, suggesting a possible role in the pathogenesis of IBD supported by their ability to interact with the host’s cells [69]. In this regard, a higher abundance of the eukaryotic Orthohepadnaviridae transcripts was found in early-diagnosed treatment-naive patients with UC, when compared with those with CD and controls [74]. Furthermore, studies have observed an increased abundance of eukaryotic viruses such as Pneumoviridae and Hepadnaviridae in UC patients, and Herpesviridae in both CD and UC patients [74]. Similarly, a study reported a positive association between the eukaryotic Anelloviridae prevalence in early-onset patients with IBD along with immunosuppressive treatment [75]. Moreover, research in mice with a CD risk gene, Atg16L1HM, has shown that Norovirus can induce intestinal pathologies [76]. Similarly, murine Norovirus has been found to induce colitis in an IL10-deficient mouse model of IBD in a microbiota-dependent manner [76].
Viruses contribute to barrier damage in various ways [77]. Phages indirectly fuel the immune response through the release of bacterial products following bacterial lysis or translocation across the epithelium (transcytosis), activating pattern recognition receptors on the intestinal epithelial cells or resident immune cells [77]. In addition, specific virome-derived factors can interfere with barrier integrity and impact intestinal physiology, often through interaction with other commensals [78]. For example, Sinha et al. pooled viral-like particles (VLPs) from three UC patients, mainly enriched in Microviridae phages and, to a lesser extent in crAss-like, Siphoviridae, and Podoviridae phages, and transplanted them into human microbiota-associated mice [79]. UC VLP transplantation exacerbated colitis severity [79]. Similarly, Adiliaghdam et al. found that the healthy virome directly elicited atypical anti-inflammatory innate immune activity, while viromes isolated from UC and CD, mainly enriched in Picornaviridae and Enterovirus B, induced inflammation, successfully reverted by non-IBD viromes [80].
Other studies pointed out the impact of viruses on innate immunity. For instance, filamentous Pf bacteriophages produced from Pseudomonas aeruginosa are internalized by DCs, macrophages, and B-cells to induce type-I interferon responses, thereby facilitating infection by related bacteria [81].
Interestingly, a recent study reported that viral infections with enteric viruses promoted the expansion of specific immune cell populations in the intestine, such as colonic and small intestinal lamina propria leucocytes, including effector memory T cells, macrophages, and plasmacytoid DCs [82].
Despite these pieces of evidence, the investigation of the virome’s impact on intestinal immunity and barrier functions is still in its infancy. Apart from the characterization of virome dysbiosis in IBD, studies describing virome-induced pathogenic events in the intestinal mucosa are still limited. However, compelling, these studies need further validation, particularly because interindividual and intercohort variabilities limit the generalizability of their results.
3.3. Archaea in IBD
Archaea are single-celled prokaryotes like bacteria but genetically closer to eukaryotes [83]. The predominant archaea in the human gut are methanogens, notably Methanobrevibacter and Methanosphaera [83]. Particularly, methanogens such as Methanobrevibacter smithii (M. smithii) and Methanosphaera stadtmanae (M. stadtmanae), play a role in IBD pathogenesis by influencing immune system dysregulation [84] (Figure 5). Decreased levels of M. smithii have been observed in individuals with IBD [85]. M. smithii contributes to digestive health by facilitating the fermentation of dietary fructans to acetate, which helps maintain gut homeostasis [85]. Reductions in M. smithii abundance may disrupt this process, leading to intestinal inflammation and contributing to IBD progression [85]. Conversely, studies have shown that the abundance of M. stadtmanae increases up to threefold in IBD patients [86]. This archaeon stimulates DCs to produce pro-inflammatory cytokines, exacerbating inflammation in the gut [86]. The elevated presence of M. stadtmanae may contribute to the dysregulation of the immune system observed in IBD [86]. Additionally, halophilic archaea may also play a role in IBD etiology [87]. However, Chehoud et al. reported no significant alterations in the archaeome associated with IBD [88]. Nevertheless, their contribution to IBD onset and progression remains contentious, likely due to insufficient research.
3.4. Fungi in IBD
Fungi constitute a small fraction, only 0.1%, of the overall microbial community in the gastrointestinal tract [89]. However, despite their low abundance, alterations in the mycobiome have been found to be closely associated with IBD [90] (Figure 6). Studies consistently report reduced fungal diversity in individuals with IBD, indicating a potential role of fungi in the pathogenesis of the disease [90,91,92]. Recent research conducted by Sokol et al. has shed light on specific changes in the mycobiome of IBD patients, revealing an elevated ratio of Basidiomycota/Ascomycota [90]. Additionally, heightened levels of Malasseziales and Filobasidiaceae, along with diminished levels of Penicillium and Kluyveromyces, have been documented in IBD patients, further highlighting the intricate relationship between fungal dysbiosis and IBD pathophysiology [90].
Among the various fungal species, Candida has garnered particular attention due to its increased prevalence in individuals with IBD [88,90]. Colonization by Candida albicans has been implicated in exacerbating inflammation through the stimulation of pro-inflammatory cytokines such as IL-17 and IL-23 [93]. This dysregulated immune response contributes to the chronic inflammation characteristic of IBD [93].
Conversely, reduced levels of Saccharomyces cerevisiae (S. cerevisiae) have been observed in IBD patients [90]. S. cerevisiae appears to play a protective role against IBD by inhibiting the binding of AIEC to intestinal mucosa, thereby reducing bacterial infiltration and subsequent inflammation [94]. Moreover, S. cerevisiae inhibits the transformation of C. albicans into its invasive hyphal form, a process mediated by the blockade of aspartyl proteases-2 and 6 [94,95]. By preventing fungal invasion and dysbiosis, S. cerevisiae contributes to maintaining intestinal homeostasis and reducing the risk of IBD development or exacerbation [94,95].
3.5. Microbiota-Derived Metabolites Involved in IBD
3.5.1. Short-Chain Fatty Acids (SCFAs)
SCFAs, produced by intestinal commensal bacteria through the fermentation of dietary fiber or other indigestible carbohydrates, play a pivotal role in maintaining intestinal health [98]. They accomplish this by modulating luminal pH, enhancing mucus production, serving as an energy source for IECs, and fortifying mucosal immune function, resulting in anti-inflammatory effects through the regulation of colonic Tregs [98]. The three primary SCFAs derived from the microbiota are acetate, propionate, and butyrate, with a ratio of approximately 3:1:1 [98]. Acetate is generated by various gut microbes, propionate is primarily produced by Bacteroidetes, Negativicutes, and Lachnospiraceae, while butyrate is predominantly synthesized by Eubacterium, Clostridium, and Fusobacterium [98].
SCFAs modulate cellular functions by interacting with G protein-coupled receptors expressed in IECs, including the orphan G protein-coupled receptor (GPR43) [99,100]. Binding of SCFAs to GPR43 mitigates inflammation, as evidenced by the heightened susceptibility to colitis observed in GPR43-deficient mice models [98,99].
Furthermore, SCFAs regulate intestinal homeostasis by stimulating the production of antimicrobial peptides and intestinal IgA, as well as promoting epithelial homeostasis through IL-18 production [101,102]. They also inhibit the expression of NF-κB and the secretion of TNF-α, while exerting anti-proliferative effects and exhibiting [100,101,102] (Figure 7).
Numerous studies have consistently reported lower levels of SCFAs in the feces of IBD patients compared to healthy individuals, indicating a potential involvement of decreased concentrations of SCFA-producing bacteria and SCFAs in the chronic intestinal inflammation and pathophysiology of IBD [99]. Additionally, research by Hu et al. has demonstrated the loss of butyrate-producing bacterial species, such as F. prausnitzii, Roseburia hominis, and Clostridium clusters IV and XIVa, as evidenced by reduced fecal butyrate levels in IBD patients [103]. This decrease in SCFA-producing bacteria is often accompanied by an increase in pathogenic bacteria like Escherichia-Shigella, which degrade SCFAs to counteract their anti-inflammatory effects, ultimately contributing to immune dysregulation in the intestinal tract of individuals with IBD [103].
3.5.2. Bile Acids (BAs)
BAs, a type of steroid acid present in bile and synthesized from cholesterol by the liver, play a crucial role in the emulsification and absorption of fats, as well as in the elimination of cholesterol [104]. They exert metabolic effects by binding to various receptors, including the farnesoid X receptor (FXR), pregnane X receptor (PXR), transmembrane G protein-coupled receptor 5 (TGR5), vitamin D receptor, and androstane [104].
There exists a bidirectional relationship between BAs and the microbiota [105]. In the intestine, the microbiota converts primary BAs, originating from the liver, into secondary BAs [105]. Conjugated BAs excreted in bile are partly deconjugated, dehydroxylated, and reduced [106,107]. Deconjugation of BAs, achieved by removing glycine and taurine, prevents their reuptake in the small intestine by the ASBT transporter [106,107]. This process relies on the activity of intestinal bacteria possessing the Bile Salt Hydrolase (BSH) enzyme, such as Lactobacilli and Bifidobacteria [106,107]. Deconjugated BAs not absorbed by the ileum via ASBT reach the colon, where they undergo dehydroxylation, generating secondary BAs: lithocholic acid and deoxycholic acid [106,107]. This dehydroxylation process involves several reactions conducted by bacteria belonging to the Clostridium (clusters XIVa and XI) and Eubacterium genera, which are part of the Firmicutes phylum [106,107]. Additionally, in humans, the microbiota can generate secondary BAs through isomerization reactions, with ursodeoxycholic acid formed by isomerization of chenodeoxycholic acid by Clostridium absonum [106,107].
BAs possess potent antimicrobial properties that may alter gut microbiota composition and density [108]. They exert direct antimicrobial effects on bacteria like Bifidobacterium breve and Lactobacillus salivarius, while their indirect effects involve stimulating the production of antimicrobial peptides from the host and activating FXR [108]. Activation of the nuclear receptor FXR has been shown to influence the composition of the microbiota [109]. Mice lacking FXR expression (Fxr-/-) and fed a high-fat diet exhibit an increase in the percentage of Firmicutes and a corresponding decrease in the percentage of Bacteroidetes compared to wild-type mice fed the same diet [110]. The accumulation of primary BAs in FXR-deficient mice suggests that the microbiota may have limited ability to metabolize BAs in the absence of this receptor [110].
Reduced levels of BAs in the intestinal lumen promote the proliferation of gram-negative bacteria, including several pathogens [111]. Additionally, decreased levels of fecal BAs are associated with an increased presence of potentially pathogenic bacteria capable of inducing inflammation, such as Enterobacteriaceae [111]. Conversely, elevated levels of BAs in the intestine promote gram-positive bacteria belonging to the Firmicutes phylum [111].
Numerous studies have reported altered bile acid profiles in fecal samples of IBD patients [112]. Concomitantly, studies have shown a significant reduction in FXR expression in CD patients [113]. Moreover, Labbé et al., analyzing metagenomics samples from the Human Microbiome Project and MetaHit, found a reduction in clusters of BSH genes associated with Firmicutes in IBD [114]. Among IBD patients, dysbiosis seems to lead to a lack of secondary BAs in the gut, and the beneficial effects of SBA supplementation on intestinal inflammation have been validated in animal models, possibly due to SBAs inhibiting Th17 cell function [112].
3.5.3. Bacterial Self-Metabolites
Tryptophan, an essential aromatic amino acid, undergoes significant alterations in concentration and metabolism in individuals with IBD, including changes in the activity of associated enzymes [115]. Dietary tryptophan follows three metabolic pathways: the kynurenine pathway, serotonin pathway, and indole pathway [115]. Microbial metabolism of tryptophan within the indole pathway yields various bioactive indole derivatives serving as agonists for the Aryl hydrocarbon receptor (AhR), a critical transcription factor regulating T cell immunity, cytokine expression, and anti-inflammatory effects via IL-22 [115]. Consistent with these findings, reduced AhR expression was noted in the inflamed mucosa of CD patients [115]. Certain Lactobacillus strains capable of AhR activation have demonstrated a reduction in severity of DSS-induced colitis [116]. Critical research has identified a significant association between tryptophan metabolism and IBD in clinical cohorts, indicating an inverse relationship between tryptophan levels and IBD severity. Similar findings in mouse models suggest that tryptophan deficiency may exacerbate colitis.
Succinate, a tricarboxylic acid (TCA) intermediate produced by both the host and the microbiota, has garnered attention due to its potential link with IBD [117]. Elevated succinate levels act as a pro-inflammatory signal, particularly increased in CD patients [117]. Fecal succinate levels are also heightened in both UC and CD patients [117]. Furthermore, reduced abundances of succinate-utilizing Phascolarctobacterium are observed in UC and CD patients compared to healthy individuals [117].
Histamine, which contributes to abdominal pain in patients with IBD, is predominantly produced by Klebsiella aerogenes, which is highly abundant in the fecal microbiota of IBD patients [118]. Elevated levels of histamine suppress the expression of tight junction and MUC2 proteins, diminish intestinal autophagy, and impair the function of colonic goblet cells in mucus secretion, ultimately resulting in compromised integrity of the intestinal mucosal barrier [118].
Desulfovibrio, a prominent genus of sulfate-reducing bacteria (SRB), can induce sulfide production, leading to symptoms such as frequent defecation, weight loss, and heightened intestinal permeability [119]. Consequently, individuals with UC typically exhibit elevated hydrogen sulfide levels in the intestines [119].
Finally, self-metabolites of bacteria, such as colibactin and indoleamine, have DNA damaging effects on epithelial cells and confer an increased risk of CRC [120].
3.5.4. Vitamins
Vitamin synthesis represents a critical metabolic function facilitated by the gut microbiota [121]. Clostridium is involved in synthesizing folate, cobalamin, niacin, and thiamine [121]. Bifidobacteria contributes to folate synthesis, while Bacteroides is implicated in producing riboflavin, niacin, pantothenate, and pyridoxine [121]. Certain intestinal bacteria rely heavily on host-supplied vitamins, suggesting that vitamin deficiency could impact bacterial growth or microbial utilization of host vitamins [121]. Inadequate dietary vitamin K disrupts the microbial community, leading to impaired blood clotting. This suggests a potential association between vitamin K deficiency and intestinal bleeding symptoms in IBD. While vitamin K1 is primarily obtained from food, vitamin K2 is synthesized by gut bacteria [122]. In neonates or healthy individuals, E. coli helps create an anaerobic environment in the intestine, facilitating the colonization of other anaerobes and vitamin K production to resist pathogenic bacteria invasion [122]. Additionally, vitamin K2 promotes the abundance of SCFAs and SCFA-producing genera in the colon [122].
Finally, while gut microbiota do not directly synthesize vitamin D, it significantly influences its metabolism and absorption [123]. Specific gut bacteria regulate the expression and activity of enzymes responsible for converting vitamin D to its active form, calcitriol, which plays vital roles in calcium and phosphorus metabolism, immune modulation, and anti-inflammatory actions [123]. Moreover, certain gut bacteria enhance the expression of genes involved in vitamin D metabolism, thereby promoting its bioavailability and activity [123]. This intricate interplay underscores the crucial role of gut microbiota in maintaining vitamin D homeostasis and its potential impact on overall health [123]. Dysbiosis, or disruption of gut microbiota composition, may compromise vitamin D absorption and metabolism, potentially leading to vitamin D deficiency [123].
4. Manipulation of Microbiota as a Treatment Strategy in IBD
4.1. Diet
A Western-style diet, characterized by elevated levels of protein, fat, and sugar, coupled with low fiber intake, has been associated with an augmented risk of IBD [124]. This dietary pattern has been demonstrated to diminish microbial diversity and compromise the integrity of the colonic mucus layer, facilitating the proliferation and heightened activity of pathogenic bacteria [124]. Consequently, this dysbiosis contributes to the accumulation of specific immune cell populations and disrupts the normal absorptive function of enterocytes [124]. Clinical investigations involving patients with UC who transitioned to a low-fat, high-fiber diet revealed a reduction in the relative abundance of Actinobacteria and an augmentation in F. prausnitzii levels [124]. Additionally, there was an increase in the concentration of anti-inflammatory metabolites such as acetate in their fecal matter [124].
Moreover, studies have highlighted a prevalent deficiency in vitamin D among individuals with IBD, with lower expression levels of the vitamin D receptor (VDR) in the intestines correlating with heightened inflammation severity [125]. The VDR pathway emerges as a promising therapeutic target for mitigating diet-induced inflammatory bowel disease [125]. Vitamin D has been shown to exert a beneficial impact on the gut microbiota composition in IBD patients, fostering the proliferation of beneficial bacterial species including Roseburia, Alistipes, Parabacteroides, and Faecalibacterium, while suppressing the abundance of pathogenic bacteria like Ruminococcus gnavus [125]. However, these effects appear to be transient, suggesting that sustained maintenance of this favorable microbial balance may necessitate additional interventions beyond long-term vitamin D supplementation alone [125].
4.2. Prebiotics
Prebiotics are non-digestible dietary fibers that are selectively fermented by the gut microflora and serve as fuel for beneficial bacteria in the gut [126]. Prebiotics mostly include fructooligosaccharides (FOS), galactooligosaccharides (GOS), and other oligosaccharides, such as pectin [126]. They can help promote the growth and activity of these beneficial microbes, which in turn can have positive effects on gut health [126]. In patients with IBD, the manipulation of the gut microbiota using prebiotics has shown promise as a therapeutic approach [126].
Many studies have shown that prebiotic supplementation may improve symptoms and quality of life in patients with IBD [127]. This may be due to the positive effects of prebiotics on gut microbial composition and function [128]. Prebiotics such as inulin, have been shown to induce the growth of SCFA-producing bacteria, including Lactobacillus, F. prausnitzii and Bifidobacterium [127]. Inulin has also been shown to improve histological lesions in patients with pouchitis [129]. FOS are known to increase the population of endogenous microflora, particularly Lactobacillus and Bifidobacterium [130]. FOS and GOS can improve the levels of F. prausnitzii [130]. By promoting the growth of these microbes, prebiotics can help restore microbial balance in the gut, which is often disrupted in patients with IBD [130]. When beneficial bacteria ferment prebiotics, they produce SCFAs such as acetate, propionate, and butyrate which have anti-inflammatory properties and help maintain gut barrier function [131,132].
While prebiotics show promise as a complementary therapy for IBD, they are typically used in conjunction with other treatments such as medication and dietary modifications [133]. The effectiveness of prebiotics may vary depending on factors such as the type of prebiotic used, the dosage, and individual differences in gut microbiota composition.
Overall, the manipulation of the gut microbiota by prebiotics represents a promising approach for managing IBD. However, more research is needed to fully understand the mechanisms of action and optimal use of prebiotics in this context. Specifically, given that prebiotics are fibers, albeit soluble, it is crucial to acknowledge the potential for poor tolerance among individuals with stricturing CD.
4.3. Probiotics
Probiotics, defined as live microorganisms offering various health benefits when consumed in adequate amounts, have emerged as a potential therapeutic option for IBD [134]. The rationale behind probiotic therapy in IBD lies in their ability to modulate gut microbiota composition, enhance intestinal barrier function, inhibit the colonization of pathogenic microbes, and regulate local and systemic immune responses [135]. Their actions, dependent on type, dose, and host interactions, range from direct antibacterial effects through substance production to non-immunological actions like nutrient competition, increased mucus production, pH alteration, tight junction formation, and tissue repair [135]. Additionally, probiotics modulate the immunological response by influencing immunoglobulin and cytokine production, regulating the NF-κB pathway, and balancing pro-inflammatory and anti-inflammatory cytokines like IL-8, TNF-α, IFN-γ, IL-10, and TGF-β [136].
While there is no established gold standard for effective probiotic dosing, most commercially available probiotics contain one to ten billion colony-forming units (CFU) per dose [137]. Certain probiotics, such as Saccharomyces boulardii, Escherichia coli Nissle 1917, and Bifidobacterium breve strain Yakult, have demonstrated efficacy and safety comparable to mesalamine in maintaining clinical remission in UC patients as assessed through quality of life measures, endoscopy and histology [97,138,139]. Notably, European Crohn’s and Colitis Organization (ECCO) guidelines endorse E. coli Nissle 1917 as a viable alternative to mesalamine for UC remission maintenance [140]. Moreover, when combined with conventional medications like mesalamine, Lactobacillus reuteri ATCC 55730 has shown improved clinical response and remission rates in children with UC [141]. Clostridium butyricum (C. butyricum) has also shown effectiveness in suppressing inflammation in experimental colitis and preventing pouchitis in UC patients [142]. Additionally, Saccharomyces boulardii has been shown to reduce recurrence rates in CD patients when combined with mesalazine [143]. This yeast strain can enhance intestinal barrier function by reducing intestinal permeability, increasing plasma levels of the anti-inflammatory cytokine IL-10 and intestinal IgA secretion, and preventing relapses in CD patients [143]. Finally, Lactobacillus plantarum has been found by Jin et al. to restore gut barrier function and reduce intestinal inflammation in a mouse model of DSS-induced colitis [144].
Various probiotic cocktails have been proposed for IBD treatment [144]. De Simone formulation (DSF) is a mixture of eight bacterial strains, including Lactobacillus acidophilus, L. plantarum, L. casei, L. delbrueckii subspecies bulgaricus, Bifidobacterium breve, B. longum, B. infantis, and Streptococcus salivarius subspecies thermophiles [145]. Studies have shown its effectiveness in inducing remission in patients with mild-to-moderately active UC, preventing or maintaining remission in chronic pouchitis following ileal pouch-anal anastomosis for UC, with lower incidence rates of acute pouchitis and higher maintenance of antibiotic-induced pouchitis remission compared to the placebo group, and preventing endoscopic recurrence after surgery for CD [146,147]. Moreover, there appears to be a synergistic effect between DSF and conventional drugs, with potential mechanisms including the enhancement of the anti-inflammatory effects of mesalazine, inhibition of free radical production, and suppression of leukotriene and IL-1 production [148]. Combining DSF with standard therapy has been proven to improve histological scores in children with UC [149]. Another study by Miele et al. showed that combining DSF with mesalamine and steroids could significantly improve the remission rate in children with UC [150].
Administration of a cocktail of L. acidophilus, L. plantarum, B. lactis, and B. breve has been shown to enhance the production of intestinal mucus and goblet cells in mice [151]. Another cocktail mixture of L. plantarum, L. acidophilus, L. rhamnosus, and E. faecium has demonstrated to increase wound healing and enhance the integrity of tight junctions of epithelial cells [152]. For patients with UC, combining mesalazine with a probiotic mixture of L. salivarius, L. acidophilus, and B. bifidum strain demonstrated beneficial effects, resulting in a shorter recovery time, lower disease activity, and improved endoscopic images [153]. Finally, Chen et al. reported that the probiotic mixture of B. infantis, L. acidophilus, and E. faecalis with or without Bacillus cereus could restore the relative abundance of Lactobacillus, Bifidobacterium, Bacteroides, and Akkermansia in a mouse model of DSS-induced chronic colitis [154].
Probiotic supplementation can not only restore intestinal microbiota depletion but also potentiate medication effectiveness. For example, the co-administration of Bifidobacterium with mesalazine not only ameliorates IBD symptoms but also reduces adverse effects [155]. Thus, prudent exploitation of the benefits conferred by gut flora may yield a synergistic effect greater than the mere sum of individual components.
The above-mentioned probiotic strains, among others, offer promising avenues for managing IBD by modulating gut microbiota, reducing inflammation, and maintaining intestinal barrier function. However, while some studies have reported positive outcomes, further research is needed to elucidate the optimal dosing, duration, and efficacy of probiotics in the context of IBD treatment. Additionally, individual responses to probiotic therapy may vary, highlighting the importance of personalized approaches and continued investigation in this field.
4.4. Next Generation Probiotics (NGPs)
NGPs represent a pioneering approach to modulating the gut microbiota and managing IBD [144]. Diverging from traditional probiotics, NGPs are sourced from human gut commensals or genetically engineered strains with enhanced functionalities, presenting potential advantages in terms of efficacy and specificity [144]. By leveraging the therapeutic potential of specific microbial strains or engineered organisms, NGPs offer a targeted and personalized approach to treating IBD [144].
One exemplar of an NGP is F. prausnitzii, a butyrate-producing bacterium renowned for its potent anti-inflammatory properties [156]. Multiple studies have underscored the therapeutic promise of F. prausnitzii in UC, demonstrating reductions in disease activity and inflammation [156].
Another NGP candidate is Akkermansia muciniphila, a bacterium specializing in mucin degradation and associated with bolstering gut barrier function and immune regulation [157]. Preclinical investigations have showcased the potential of A. muciniphila supplementation in ameliorating colitis and enhancing intestinal barrier integrity in animal models of IBD [157].
C. butyricum MIYAIRI, another NGP, is a butyrate-producing bacterium that has shown effectiveness in preventing pouchitis and mitigating alterations in the microbiota of UC patients [158]. Recent findings by Ma et al. indicate that C. butyricum MIYAIRI-II could alleviate parameters associated with colitis in a mouse model of DSS-induced colitis [158].
Additionally, genetically engineered probiotic strains offer innovative avenues in IBD therapy [159]. For instance, researchers have developed engineered strains of Escherichia coli Nissle 1917 that overexpress anti-inflammatory proteins or enzymes involved in metabolite production, such as catalase and superoxide dismutase [160]. These modified strains have exhibited efficacy in reducing inflammation and fostering mucosal healing in preclinical models of IBD [160]. Genetically engineered E. coli Nissle 1917 has also been shown to enhance the abundance of microbes crucial for maintaining intestinal homeostasis, such as Lachnospiraceae and Odoribacter [160].
Other NGPs currently under investigation encompass genetically modified Lactobacilli and Bifidobacteria engineered to produce anti-inflammatory cytokines or metabolites [159,161,162]. These tailored probiotics hold promise in modulating immune responses and reinstating gut homeostasis in individuals with IBD.
In conclusion, NGPs herald a promising frontier in IBD therapeutics, offering targeted interventions that address the underlying mechanisms of disease pathology. Further research and clinical trials are imperative to elucidate the safety, efficacy, and optimal dosing regimens of NGPs in the management of IBD.
4.5. Synbiotics
Synbiotics, a combination of probiotics and prebiotics, have garnered interest as a therapeutic approach for IBD [163]. By combining beneficial microorganisms with substrates that promote their growth and activity, synbiotics aim to synergistically enhance the efficacy of the treatment [163].
Several studies have investigated the use of synbiotics in IBD, with promising results [153]. For example, a synbiotic combination of Lactobacillus and FOS has been shown to improve clinical outcomes and reduce disease activity in patients with UC [164]. Similarly, another synbiotic formulation containing Bifidobacterium and inulin resulted in significant reductions in inflammation and improvement in symptoms in individuals with CD [165].
Furthermore, synbiotics have been found to enhance the efficacy of conventional treatments for IBD [166]. For instance, combining synbiotics with mesalazine, a commonly used medication for IBD, has been shown to improve clinical response rates and reduce the risk of relapse in patients with UC.
Overall, synbiotics represent a promising adjunctive therapy for IBD, offering a multifaceted approach to address the complex interplay between gut microbiota dysbiosis, immune dysfunction, and intestinal inflammation. Further research is needed to optimize synbiotic formulations, dosing regimens, and treatment durations to maximize their therapeutic benefits in IBD management.
4.6. Fecal Microbial Transplant (FMT)
FMT is a direct method for reshaping the intestinal microbiota by introducing a fecal suspension from a carefully chosen healthy donor into the intestines of a patient suffering from a disease [167]. Recipients of FMT may inherit crucial genes from the donor, that contribute to restore a healthy and functional gut ecosystem by enhancing the production of SCFAs and restoring immune dysregulation [168]. FMT is already approved in treating recurrent Clostridioides difficile (C. difficile) infections (CDI) resistant to antibiotic treatment [169]. While the microbial foundation of IBD proves to be considerably intricate and variable compared to relapsed/refractory CDI, therapies based on the microbiota represent a critical area of exploration for these chronic and incapacitating conditions [170]. Consequently, there has been a surge in clinical studies investigating the effectiveness of FMT in treating IBD [170].
Studies have found that FMT is effective in inducing remission in UC patients [171]. A significant enrichment of Bacteroides, Proteus, and Prevotella and a decline in Klebsiella and Streptococcus was found by Tian et al. after FMT [172].
A meta-analysis by Colman and Rubin showed a remission rate of 36.2% in IBD patients who received FMT [173]. They also showed a higher remission rate in CD patients than in UC patients. A meta-analysis on FMT for IBD by Paramsothy et al. showed a clinical remission rate of 50.5% [174]. Another meta-analysis by Caldeira et al. reported that FMT had a complete remission rate of 37% for IBD patients [175].
Numerous studies have been conducted to assess the efficacy of FMT in inducing remission in UC. For instance, Moayyedi et al. conducted a study involving 75 patients with mild to severe UC [176]. The study group received FMT via enemas from donors, while the control group underwent a placebo treatment. The study revealed that patients receiving FMT achieved clinical remission compared to the control group, with statistically significant results (p = 0.03). Another randomized placebo-controlled study, conducted by Paramsothy et al., involved 81 patients with mild to moderate UC, with 41 patients in the study group and 40 in the control group [177]. The results indicated a significantly higher rate of endoscopic remission in the study group compared to the control group at week 8 (p = 0.021). In the same way, Costello et al. documented a notably superior treatment effect in the study group, consisting of 38 patients with moderate UC who underwent FMT, in contrast to the control group with 35 patients in the placebo arm [171]. Following a two-month follow-up, 12 patients (32%) in the FMT group achieved both clinical and endoscopic remission, whereas only 3 out of 35 patients in the placebo group attained complete remission (p = 0.03). Similar results were obtained by Cui et al. showing that FMT improved clinical outcomes in 57% of patients with steroid-dependent UC [178]. These findings were further confirmed by Kunde et al. which found a significant improvement in 9 children with UC who received FMT via enema [179].
The efficacy and safety of FMT was also investigated in CD. A pilot single center trial evaluated the effect of multiple FMTs on 25 CD patients complicated with intraabdominal inflammatory mass [180]. All patients received the initial FMT followed by repeated FMTs every 3 months. Clinical response and clinical remission at 3 months post the initial FMT were achieved in 68.0% (17/25) and 52.0% (13/25) of patients, respectively. The proportion of patients at 6 months, 12 months and 18 months achieving sustained clinical remission with sequential FMTs was 48.0% (12/25), 32.0% (8/25) and 22.7% (5/22), respectively. 9.5% (2/21) of patients achieved radiological healing and 71.4% (15/21) achieved radiological improvement.
Another randomized controlled trial determined the efficacy and safety of different methods of FMT, as a potential therapy for CD [181]. A total of 27 patients with CD were randomized to receive FMT by gastroscopy or colonoscopy; a second transplantation was performed 1 week later. Clinical remission, assessed 8 weeks after FMT, was achieved in 18 (66.7%); no significant difference was seen between the two methods. Moreover, microbiota diversity analyses showed that, compared to donors, CD patients showed a significant increase in operational taxonomic units (OUT, 117 vs. 258, p < 0.05) 2 weeks after FMT. In CD patients, FMT contributed to increase species richness, raising levels of Clostridium, Cronobacter, Fusobacterium, and Streptococcus.
Finally, a randomized, single-blind, sham-controlled pilot trial of FMT in adults with colonic or ileo-colonic CD was performed [182]. Out of the enrolled patients, 8 underwent FMT, while 9 received sham transplantation. The steroid-free clinical remission rates at 10 and 24 weeks post-FMT were 44.4% (4/9) and 33.3% (3/9) in the sham transplantation group, respectively, and 87.5% (7/8) and 50.0% (4/8) in the FMT group. The Crohn’s Disease Endoscopic Index of Severity (CDEIS) exhibited a decrease six weeks after FMT (p = 0.03) but not after sham transplantation (p = 0.8). Conversely, the C-reactive protein (CRP) level increased six weeks after sham transplantation (p = 0.008) but not after FMT (p = 0.5). Higher colonization by donor microbiota correlated with the maintenance of remission.
The efficacy of FMT in CD remains less established compared to UC, with mixed results reported in various trials.
Long-term follow-up studies have assessed the durability of FMT-induced remission in IBD patients [183,184]. While some patients maintain remission following FMT, others may experience disease recurrence over time, highlighting the need for further research into optimal maintenance strategies [185].
However, various clinical studies conducted to examine the effect of FMT on IBD have obtained inconsistent results, raising doubts about its effectiveness. Available data suggest that the efficacy of FMT in treating IBD is not predictable. After 12 weeks of FMT, only one UC patient showed some improvement, as reported by Angelberger et al. [186]. Likewise, Suskind et al. observed no notable improvement in four children who underwent a single FMT administered via a nasogastric tube [187]. The variations in outcomes across clinical trials may stem from differences in disease pathology, donor selection criteria, FMT protocols, and individual responses to treatment. Concerns regarding the safety and effectiveness of FMT have hindered its widespread adoption for IBD management. FMT carries inherent risks, including the potential transmission of infectious agents from donor to recipient. Instances of severe infections, such as bacteremia and sepsis, have been documented post-FMT, underscoring the necessity for stringent donor screening and safety measures [188]. While most adverse events linked to FMT are mild and transient, such as gastrointestinal discomforts like bloating, diarrhea, and abdominal pain, more severe complications can arise, especially among immunocompromised individuals or those with underlying health conditions.
FMT holds promise as a potential therapeutic intervention for IBD, particularly in UC, where it has demonstrated efficacy in inducing remission in some patients. However, further research is needed to elucidate optimal protocols regarding donor selection, administration route, dosing, and long-term maintenance strategies. Additionally, safety considerations and potential adverse events must be carefully evaluated to ensure the overall benefit-risk profile of FMT in IBD patients.
4.7. Fecal Virome Transplant (FVT)
In contrast to conventional FMT, FVT entails transferring only gut viruses from healthy donors to diseased patients [189]. Most FVT investigations have been conducted using in vitro mouse models of diseases lacking clear biomarkers, such as obesity and antibiotic-induced dysbiosis [189,190]. Scientists are investigating whether FVT could be an effective treatment for IBD [191]. The rationale behind this approach lies in the idea that viruses in the gut microbiota, particularly bacteriophages, notably impact overall bacteriome compositions, altering Firmicutes–Bacteroidetes ratios, diversity, and specific bacterial abundances, albeit the latter contributing minimally to the bacteriome.
Research on FVT in inflammatory bowel disease IBD is still in its early stages, but there is growing interest in exploring its potential therapeutic applications. FVT offers advantages over FMT by reducing the risk of transferring unknown pathogens or bacteria with undesirable functionalities.
In a study by Ott et al., FVT preparations, which underwent sterile filtration, were administered to five patients with CDI, including three who had failed FMT and/or antibiotic treatments and one deemed ineligible for FMT due to infectious risks [192]. All five patients recovered from CDI infection post-FVT and remained symptom-free for at least 6 months. Although virome analysis was conducted on only one patient, significant changes were observed in the patient’s phageome, resembling that of the donor. Nonetheless, due to the study’s focus on the efficacy of fecal filtrates rather than the virome specifically, and the limited sample size, no definitive causal links between the virome and patient recovery could be established, nor were specific beneficial phages identified. Therefore, while the therapeutic potential of the virome has been demonstrated, progress in developing virome-based therapies is contingent upon a better understanding of the taxa and mechanisms by which viruses impact host metabolism, influencing both diseased and healthy gut states.
Safety is a critical consideration in FVT, as with any transplantation procedure. There are concerns about the potential transfer of harmful viruses or genetic elements from donors to recipients. Therefore, stringent donor screening protocols and safety measures are essential to minimize the risk of adverse effects. Additionally, researchers are exploring methods to isolate and characterize specific beneficial phages that could be used in targeted therapies, thereby reducing the need for whole-virome transplants.
4.8. Phage Therapy
Bacteriophages, or phages, have garnered attention as a potential alternative to antibiotics for combating multidrug-resistant bacteria [193]. In the realm of gastrointestinal disorders, phage therapy, which involves administering cocktails of bacteriophages, shows promise for conditions associated with specific bacterial colonization or infection, such as IBD with AIEC, Klebsiella pneumoniae and C. difficile [194,195].
Compared to fecal transplants or antibiotics, phage therapies offer advantages by enabling the targeting of specific commensal bacteria, including drug-resistant strains [196]. This targeted approach helps minimize unintended alterations in the gut microbiota without the transfer of live bacteria.
In experimental murine models of IBD deliberately colonized with pathobionts linked to IBD, researchers have examined diverse combinations of bacteriophages. Although these studies have typically demonstrated efficacy in eliminating targeted bacteria, assessing their clinical implications on disease activity beyond infection resolution presents challenges [197]. Moreover, the limitations of animal models hinder the direct extrapolation of these outcomes to human cases of IBD. Additionally, numerous investigations into phage therapy for IBD have concentrated on targeting individual bacterial strains, thereby neglecting the complex microbial diversity characteristic of IBD.
4.9. Targeting Archaeome
Targeting the archaeome presents significant challenges, as human methanogenic archaea have demonstrated high resistance to antibiotics, rendering them susceptible only to compounds that also affect bacteria and eukarya [198]. This limitation complicates the development of specific therapies. However, recent discoveries have indicated that statins can inhibit archaeal cell membrane biosynthesis without affecting bacterial populations, suggesting a potential avenue for targeted therapeutic intervention while preserving the integrity of the intestinal microbiota [199]. This finding offers a potential starting point for modulating the archaeome in patients with IBD. However, achieving success in this endeavor will require extensive further research and dedicated efforts to refine IBD treatment strategies.
4.10. Targeting Micobiome
When it comes to modifying fungal composition, interventions may entail the administration of specific antifungal medications. For example, a recent small-scale pilot study, NCT03476317, has completed patient enrollment to evaluate the effects of a novel therapeutic regimen targeting the gut microbiota. This regimen involves bowel lavage and antibiotic treatment, with or without the inclusion of the antifungal drug fluconazole. The study aims to evaluate the efficacy of this approach in treating active CD or indeterminate colitis (IBDU) that has shown resistance to conventional immunosuppressive therapy.
In forthcoming clinical trials, such as NCT05049525, which is currently not enrolling participants, the objective is to evaluate the effectiveness of combined antifungal therapies, including itraconazole and terbinafine, compared to a placebo in patients diagnosed with CD. This trial aims to furnish additional evidence supporting the idea that targeting fungal components in these patients may contribute to achieving remission. Similarly, the pilot study NCT04966585, also not recruiting yet, aims to investigate whether the changes in microbial composition induced by antifungal treatment are associated with reduced downstream immune responses in CD patients who possess a genetic predisposition to mounting robust immune reactions against Malassezia.
In the realm of fungal-derived factors, a randomized clinical trial examining the efficacy of Saccharomyces boulardii conducted by Plein et al. revealed heightened disease activity index scores among a cohort of CD patients [200]. Following studies in CD patients have similarly indicated enhancements in relapse rates and intestinal permeability.
5. Conclusions
In summary, the interaction between the gut microbiota and the immune system plays a pivotal role in the pathophysiology of IBD. Both UC and CD stem from a complex interplay of genetic predisposition and environmental factors, with dysbiosis serving as a central contributor to disease onset and progression. The stability of the gut microbial structure is crucial for numerous physiological processes, including direct interaction with the intestinal epithelium and modulation of mucus secretion and mucosal immunity. Understanding these intricate dynamics and the specific role of each microbiota component implicated in IBD is essential for elucidating disease mechanisms and developing innovative therapeutic strategies.
Manipulating the microbiota-immune axis holds promise for more effective IBD management and improved patient outcomes, spanning from preventive measures to novel treatments. Changes in gut microbiota structure and metabolites are emerging as key parameters for drug development and mechanistic studies. Restoring the gut microbiota structure can alleviate IBD exacerbations, with potential interventions including the colonization of beneficial bacteria or supplementation of commensal-produced metabolites to enhance intestinal barrier integrity in IBD patients. Advanced metagenomic and metabolomic analyses provide profound insights into the complex relationships among gut microbes, metabolites, and hosts.
Continued research efforts are necessary to explore nonpharmacological and behavioural interventions aimed at restoring gut ecosystem homeostasis and immune function, ultimately offering relief to individuals with IBD. Additionally, standardization of research methods for studying gut microbiota is imperative to ensure reproducible data and robust knowledge. Establishing guidelines and achieving consensus on research protocols will facilitate the generation of reliable data. Furthermore, large sample-sized cohorts are crucial to address the inherent variability in microbiota-related research, necessitating patient stratification based on geographical origin, sex, clinical characteristics, and lifestyle habits to mitigate study inconsistencies.
Author Contributions
EB wrote the article, created the figures and the tables. DGR critically reviewed the content of the paper and supervised the project. GPC, GMS and EB participated in the supervision of the project. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gomollón, F.; Dignass, A.; Annese, V.; et al. 3rd European Evidence-based Consensus on the Diagnosis and Management of Crohn’s Disease 2016: Part 1: Diagnosis and Medical Management. J Crohns Colitis 2017, 11, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Magro, F.; Gionchetti, P.; Eliakim, R.; et al. Third European Evidence-based Consensus on Diagnosis and Management of Ulcerative Colitis. Part 1: Definitions, Diagnosis, Extra-intestinal Manifestations, Pregnancy, Cancer Surveillance, Surgery, and Ileo-anal Pouch Disorders. J Crohns Colitis 2017, 11, 649–670. [Google Scholar] [CrossRef] [PubMed]
- Peyrin-Biroulet, L.; Panés, J.; Sandborn, W.J.; et al. Defining Disease Severity in Inflammatory Bowel Diseases: Current and Future Directions. Clin Gastroenterol Hepatol Off Clin Pract J Am Gastroenterol Assoc 2016, 14, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Knowles, S.R.; Graff, L.A.; Wilding, H.; et al. Quality of Life in Inflammatory Bowel Disease: A Systematic Review and Meta-analyses-Part I. Inflamm Bowel Dis 2018, 24, 742–751. [Google Scholar] [CrossRef]
- Buie, M.J.; Quan, J.; Windsor, J.W.; et al. Global Hospitalization Trends for Crohn’s Disease and Ulcerative Colitis in the 21st Century: A Systematic Review With Temporal Analyses. Clin Gastroenterol Hepatol Off Clin Pract J Am Gastroenterol Assoc 2023, 21, 2211–2221. [Google Scholar] [CrossRef]
- Sebastian, S.; Hernández, V.; Myrelid, P.; et al. Colorectal cancer in inflammatory bowel disease: results of the 3rd ECCO pathogenesis scientific workshop (I). J Crohns Colitis 2014, 8, 5–18. [Google Scholar] [CrossRef]
- Annese, V.; Duricova, D.; Gower-Rousseau, C.; et al. Impact of New Treatments on Hospitalisation, Surgery, Infection, and Mortality in IBD: a Focus Paper by the Epidemiology Committee of ECCO. J Crohns Colitis 2016, 10, 216–225. [Google Scholar] [CrossRef]
- Zhao, M.; Gönczi, L.; Lakatos, P.L.; et al. The Burden of Inflammatory Bowel Disease in Europe in 2020. J Crohns Colitis 2021, 15, 1573–1587. [Google Scholar] [CrossRef]
- Burisch, J.; Jess, T.; Martinato, M.; et al. The burden of inflammatory bowel disease in Europe. J Crohns Colitis 2013, 7, 322–337. [Google Scholar] [CrossRef]
- Guan, Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. J Immunol Res 2019, 2019, 7247238. [Google Scholar] [CrossRef]
- Frenkel, S.; Bernstein, C.N.; Sargent, M.; et al. Genome-wide analysis identifies rare copy number variations associated with inflammatory bowel disease. PloS One 2019, 14, e0217846. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wu, G.D.; Albenberg, L.; et al. Gut microbiota and IBD: causation or correlation? Nat Rev Gastroenterol Hepatol 2017, 14, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Geremia, A.; Biancheri, P.; Allan, P.; et al. Innate and adaptive immunity in inflammatory bowel disease. Autoimmun Rev 2014, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Mitsialis, V.; Wall, S.; Liu, P.; et al. Single-Cell Analyses of Colon and Blood Reveal Distinct Immune Cell Signatures of Ulcerative Colitis and Crohn’s Disease. Gastroenterology 2020, 159, 591–608. [Google Scholar] [CrossRef]
- Martin, J.C.; Chang, C.; Boschetti, G.; et al. Single-Cell Analysis of Crohn’s Disease Lesions Identifies a Pathogenic Cellular Module Associated with Resistance to Anti-TNF Therapy. Cell 2019, 178, 1493–1508. [Google Scholar] [CrossRef] [PubMed]
- Nakase, H.; Sato, N.; Mizuno, N.; et al. The influence of cytokines on the complex pathology of ulcerative colitis. Autoimmun Rev 2022, 21, 103017. [Google Scholar] [CrossRef] [PubMed]
- Saez, A.; Herrero-Fernandez, B.; Gomez-Bris, R.; et al. Pathophysiology of Inflammatory Bowel Disease: Innate Immune System. Int J Mol Sci 2023, 24, 1526. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.K.; Glapa-Nowak, A.; Banaszkiewicz, A.; et al. HLA-DQA1*05 Associates with Extensive Ulcerative Colitis at Diagnosis: An Observational Study in Children. Genes 2021, 12, 1934. [Google Scholar] [CrossRef]
- Ogura, Y.; Bonen, D.K.; Inohara, N.; et al. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 2001, 411, 603–606. [Google Scholar] [CrossRef]
- Salem, M.; Ammitzboell, M.; Nys, K.; et al. ATG16L1: A multifunctional susceptibility factor in Crohn disease. Autophagy 2015, 11, 585–594. [Google Scholar] [CrossRef]
- Kalischuk, L.D.; Buret, AG. A role for Campylobacter jejuni-induced enteritis in inflammatory bowel disease? Am J Physiol Gastrointest Liver Physiol 2010, 298, G1–9. [Google Scholar] [CrossRef] [PubMed]
- Schultz, B.M.; Paduro, C.A.; Salazar, G.A.; et al. A Potential Role of Salmonella Infection in the Onset of Inflammatory Bowel Diseases. Front Immunol 2017, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhu, Y.; Huang, X.; et al. Association between TLR2 and TLR4 Gene Polymorphisms and the Susceptibility to Inflammatory Bowel Disease: A Meta-Analysis. PloS One 2015, 10, e0126803. [Google Scholar] [CrossRef] [PubMed]
- Lidar, M.; Langevitz, P.; Shoenfeld, Y. The role of infection in inflammatory bowel disease: initiation, exacerbation and protection. Isr Med Assoc J IMAJ 2009, 11, 558–563. [Google Scholar] [PubMed]
- Ananthakrishnan, A.N.; Bernstein, C.N.; Iliopoulos, D.; et al. Environmental triggers in IBD: a review of progress and evidence. Nat Rev Gastroenterol Hepatol 2018, 15, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Gasaly, N.; Gasaly, N.; de Hermoso, M.A.V.P. Impact of Bacterial Metabolites on Gut Barrier Function and Host Immunity: A Focus on Bacterial Metabolism and Its Relevance for Intestinal Inflammation. Front Immunol 2021, 12, 658354. [Google Scholar] [CrossRef]
- Adak, A.; Khan, MR. An insight into gut microbiota and its functionalities. Cell Mol Life Sci CMLS 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; et al. Role of the normal gut microbiota. World J Gastroenterol 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Amoroso, C.; Perillo, F.; Strati, F.; et al. The Role of Gut Microbiota Biomodulators on Mucosal Immunity and Intestinal Inflammation. Cells 2020, 9, 1234. [Google Scholar] [CrossRef]
- Merra, G.; Noce, A.; Marrone, G.; et al. Influence of Mediterranean Diet on Human Gut Microbiota. Nutrients 2020, 13, 7. [Google Scholar] [CrossRef]
- Strasser, B.; Wolters, M.; Weyh, C.; et al. The Effects of Lifestyle and Diet on Gut Microbiota Composition, Inflammation and Muscle Performance in Our Aging Society. Nutrients 2021, 13, 2045. [Google Scholar] [CrossRef] [PubMed]
- Quaglio, A.E.V.; Grillo, T.G.; De Oliveira, E.C.S.; et al. Gut microbiota, inflammatory bowel disease and colorectal cancer. World J Gastroenterol 2022, 28, 4053–4060. [Google Scholar] [CrossRef]
- Di Tommaso, N.; Gasbarrini, A.; Ponziani, FR. Intestinal Barrier in Human Health and Disease. Int J Environ Res Public Health 2021, 18, 12836. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, SH. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp Mol Med 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Wang, K.; Xu, C.; et al. Intestinal Claudin-7 deficiency impacts the intestinal microbiota in mice with colitis. BMC Gastroenterol 2022, 22, 24. [Google Scholar] [CrossRef] [PubMed]
- Barmeyer, C.; Schulzke, J.D.; Fromm, M. Claudin-related intestinal diseases. Semin Cell Dev Biol 2015, 42, 30–38. [Google Scholar] [CrossRef]
- Christovich, A.; Luo, XM. Gut Microbiota, Leaky Gut, and Autoimmune Diseases. Front Immunol 2022, 13, 946248. [Google Scholar] [CrossRef] [PubMed]
- Martel, J.; Chang S-H, Ko Y-F, et al.; et al. Gut barrier disruption and chronic disease. Trends Endocrinol Metab TEM 2022, 33, 247–265. [CrossRef]
- Giambra, V.; Pagliari, D.; Rio, P.; et al. Gut Microbiota, Inflammatory Bowel Disease, and Cancer: The Role of Guardians of Innate Immunity. Cells 2023, 12, 2654. [Google Scholar] [CrossRef]
- Boyapati, R.K.; Rossi, A.G.; Satsangi, J.; et al. Gut mucosal DAMPs in IBD: from mechanisms to therapeutic implications. Mucosal Immunol 2016, 9, 567–582. [Google Scholar] [CrossRef]
- Walsh, D.; McCarthy, J.; O’Driscoll, C.; et al. Pattern recognition receptors--molecular orchestrators of inflammation in inflammatory bowel disease. Cytokine Growth Factor Rev 2013, 24, 91–104. [Google Scholar] [CrossRef]
- McIntyre, A.B.R.; Ounit, R.; Afshinnekoo, E.; et al. Comprehensive benchmarking and ensemble approaches for metagenomic classifiers. Genome Biol 2017, 18, 182. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, F.; Ghosh, T.S.; O’Toole, PW. The Healthy Microbiome-What Is the Definition of a Healthy Gut Microbiome? Gastroenterology 2021, 160, 483–494. [Google Scholar] [CrossRef] [PubMed]
- McBurney, M.I.; Davis, C.; Fraser, C.M.; et al. Establishing What Constitutes a Healthy Human Gut Microbiome: State of the Science, Regulatory Considerations, and Future Directions. J Nutr 2019, 149, 1882–1895. [Google Scholar] [CrossRef] [PubMed]
- McCallum, G.; Tropini, C. The gut microbiota and its biogeography. Nat Rev Microbiol 2024, 22, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Leimbach, A.; Hacker, J.; Dobrindt U., E. coli as an all-rounder: the thin line between commensalism and pathogenicity. Curr Top Microbiol Immunol 2013, 358, 3–32. [Google Scholar] [CrossRef]
- Palmela, C.; Chevarin, C.; Xu, Z.; et al. Adherent-invasive Escherichia coli in inflammatory bowel disease. Gut 2018, 67, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi-Lauridsen, H.C.; Vallance, B.A.; Krogfelt, K.A.; et al. Escherichia coli Pathobionts Associated with Inflammatory Bowel Disease. Clin Microbiol Rev 2019, 32, e00060–18. [Google Scholar] [CrossRef]
- Wittkopf, N.; Pickert, G.; Billmeier, U.; et al. Activation of intestinal epithelial Stat3 orchestrates tissue defense during gastrointestinal infection. PloS One 2015, 10, e0118401. [Google Scholar] [CrossRef]
- Appunni, S.; Rubens, M.; Ramamoorthy, V.; et al. Emerging Evidence on the Effects of Dietary Factors on the Gut Microbiome in Colorectal Cancer. Front Nutr 2021, 8, 718389. [Google Scholar] [CrossRef]
- Patrick, S. A tale of two habitats: Bacteroides fragilis, a lethal pathogen and resident in the human gastrointestinal microbiome. Microbiol Read Engl 2022, 168. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, Z.; Yan, Y.; et al. Enterotoxigenic Bacteroidesfragilis Promotes Intestinal Inflammation and Malignancy by Inhibiting Exosome-Packaged miR-149-3p. Gastroenterology 2021, 161, 1552–1566. [Google Scholar] [CrossRef] [PubMed]
- Haghi, F.; Goli, E.; Mirzaei, B.; et al. The association between fecal enterotoxigenic B. fragilis with colorectal cancer. BMC Cancer 2019, 19, 879. [Google Scholar] [CrossRef]
- Teitelbaum, J.E.; Triantafyllopoulou, M. Inflammatory bowel disease and Streptococcus bovis. Dig Dis Sci 2006, 51, 1439–1442. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nakagawasai, O.; Nemoto, W.; et al. Effect of Enterococcus faecalis 2001 on colitis and depressive-like behavior in dextran sulfate sodium-treated mice: involvement of the brain-gut axis. J Neuroinflammation 2019, 16, 201. [Google Scholar] [CrossRef] [PubMed]
- Strickertsson, J.A.B.; Desler, C.; Martin-Bertelsen, T.; et al. Enterococcus faecalis infection causes inflammation, intracellular oxphos-independent ROS production, and DNA damage in human gastric cancer cells. PloS One 2013, 8, e63147. [Google Scholar] [CrossRef] [PubMed]
- Fan T-J, Goeser, L.; Lu, K.; et al. Enterococcus faecalis Glucosamine Metabolism Exacerbates Experimental Colitis. Cell Mol Gastroenterol Hepatol 2021, 12, 1373–1389. [Google Scholar] [CrossRef]
- Bashir, A.; Miskeen, A.Y.; Hazari, Y.M.; et al. Fusobacterium nucleatum, inflammation, and immunity: the fire within human gut. Tumour Biol J Int Soc Oncodevelopmental Biol Med 2016, 37, 2805–2810. [Google Scholar] [CrossRef]
- Su, W.; Chen, Y.; Cao, P.; et al. Fusobacterium nucleatum Promotes the Development of Ulcerative Colitis by Inducing the Autophagic Cell Death of Intestinal Epithelial. Front Cell Infect Microbiol 2020, 10, 594806. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; et al. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front Microbiol 2016, 7, 979. [Google Scholar] [CrossRef]
- Machiels, K.; Joossens, M.; Sabino, J.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, H.; Xu, J.; et al. prausnitzii and its supernatant increase SCFAs-producing bacteria to restore gut dysbiosis in TNBS-induced colitis. AMB Express 2021, 11, 33. [Google Scholar] [CrossRef]
- Facchin, S.; Vitulo, N.; Calgaro, M.; et al. Microbiota changes induced by microencapsulated sodium butyrate in patients with inflammatory bowel disease. Neurogastroenterol Motil 2020, 32, e13914. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Dharmaprakash, V.; Nighot, P.; et al. Bifidobacterium bifidum Enhances the Intestinal Epithelial Tight Junction Barrier and Protects against Intestinal Inflammation by Targeting the Toll-like Receptor-2 Pathway in an NF-κB-Independent Manner. Int J Mol Sci 2021, 22, 8070. [Google Scholar] [CrossRef] [PubMed]
- Miri, S.T.; Sotoodehnejadnematalahi, F.; Amiri, M.M.; et al. Comparison of the prevalence of bacteriocin encoding genes in Lactobacillus spp. isolated from fecal samples of healthy volunteers, IBD-patient and IBD-recovered. Iran J Microbiol 2022, 14, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Shima, T.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, K.R.; Liu, S.X.L.; Tian, R.; et al. Bifidobacteria stabilize claudins at tight junctions and prevent intestinal barrier dysfunction in mouse necrotizing enterocolitis. Am J Pathol 2013, 182, 1595–1606. [Google Scholar] [CrossRef]
- Rodrigues, V.F.; Elias-Oliveira, J.; Pereira, Í.S.; et al. Akkermansia muciniphila and Gut Immune System: A Good Friendship That Attenuates Inflammatory Bowel Disease, Obesity, and Diabetes. Front Immunol 2022, 13, 934695. [Google Scholar] [CrossRef]
- Liang, G.; Cobián-Güemes, A.G.; Albenberg, L.; et al. The gut virome in inflammatory bowel diseases. Curr Opin Virol 2021, 51, 190–198. [Google Scholar] [CrossRef]
- Simmonds, P.; Aiewsakun, P. Virus classification - where do you draw the line? Arch Virol 2018, 163, 2037–2046. [Google Scholar] [CrossRef]
- Norman, J.M.; Handley, S.A.; Baldridge, M.T.; et al. Disease-specific alterations in the enteric virome in inflammatory bowel disease. Cell 2015, 160, 447–460. [Google Scholar] [CrossRef]
- Cao, Z.; Fan, D.; Sun, Y.; et al. The gut ileal mucosal virome is disturbed in patients with Crohn’s disease and exacerbates intestinal inflammation in mice. Nat Commun 2024, 15, 1638. [Google Scholar] [CrossRef] [PubMed]
- Cornuault, J.K.; Petit M-A, Mariadassou, M.; et al. Phages infecting Faecalibacterium prausnitzii belong to novel viral genera that help to decipher intestinal viromes. Microbiome 2018, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, F.; Massimino, L.; Furfaro, F.; et al. Metagenomic analysis of intestinal mucosa revealed a specific eukaryotic gut virome signature in early-diagnosed inflammatory bowel disease. Gut Microbes 2019, 10, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Conrad, M.A.; Kelsen, J.R.; et al. Dynamics of the Stool Virome in Very Early-Onset Inflammatory Bowel Disease. J Crohns Colitis 2020, 14, 1600–1610. [Google Scholar] [CrossRef] [PubMed]
- Parkes, M. Evidence from genetics for a role of autophagy and innate immunity in IBD pathogenesis. Dig Dis Basel Switz 2012, 30, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Tiamani, K.; Luo, S.; Schulz, S.; et al. The role of virome in the gastrointestinal tract and beyond. FEMS Microbiol Rev 2022, 46, fuac027. [Google Scholar] [CrossRef]
- Clinton, N.A.; Hameed, S.A.; Agyei, E.K.; et al. Crosstalk between the Intestinal Virome and Other Components of the Microbiota, and Its Effect on Intestinal Mucosal Response and Diseases. J Immunol Res 2022, 2022, 7883945. [Google Scholar] [CrossRef]
- Sinha, A.; Li, Y.; Mirzaei, M.K.; et al. Transplantation of bacteriophages from ulcerative colitis patients shifts the gut bacteriome and exacerbates the severity of DSS colitis. Microbiome 2022, 10, 105. [Google Scholar] [CrossRef]
- Adiliaghdam, F.; Amatullah, H.; Digumarthi, S.; et al. Human enteric viruses autonomously shape inflammatory bowel disease phenotype through divergent innate immunomodulation. Sci Immunol 2022, 7, eabn6660. [Google Scholar] [CrossRef]
- Sweere, J.M.; Van Belleghem, J.D.; Ishak, H.; et al. Bacteriophage trigger antiviral immunity and prevent clearance of bacterial infection. Science 2019, 363, eaat9691. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Feng, N.; Narváez, C.F.; et al. The influence of CD4+ CD25+ Foxp3+ regulatory T cells on the immune response to rotavirus infection. Vaccine 2008, 26, 5601–5611. [Google Scholar] [CrossRef] [PubMed]
- Eme, L.; Doolittle, WF. Archaea. Curr Biol CB 2015, 25, R851–855. [Google Scholar] [CrossRef] [PubMed]
- Raymann, K.; Moeller, A.H.; Goodman, A.L.; et al. Unexplored Archaeal Diversity in the Great Ape Gut Microbiome. mSphere 2017, 2, e00026–17. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, S.B.; Rostami, E.; Sephay, A.A.; et al. Alterations of the human gut Methanobrevibacter smithii as a biomarker for inflammatory bowel diseases. Microb Pathog 2018, 117, 285–289. [Google Scholar] [CrossRef]
- Blais Lecours, P.; Marsolais, D.; Cormier, Y.; et al. Increased prevalence of Methanosphaera stadtmanae in inflammatory bowel diseases. PloS One 2014, 9, e87734. [Google Scholar] [CrossRef] [PubMed]
- Oxley, A.P.A.; Lanfranconi, M.P.; Würdemann, D.; et al. Halophilic archaea in the human intestinal mucosa. Environ Microbiol 2010, 12, 2398–2410. [Google Scholar] [CrossRef]
- Chehoud, C.; Albenberg, L.G.; Judge, C.; et al. Fungal Signature in the Gut Microbiota of Pediatric Patients With Inflammatory Bowel Disease. Inflamm Bowel Dis 2015, 21, 1948–1956. [Google Scholar] [CrossRef]
- Hallen-Adams, H.E.; Suhr, MJ. Fungi in the healthy human gastrointestinal tract. Virulence 2017, 8, 352–358. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef]
- Qiu, X.; Zhao, X.; Cui, X.; et al. Characterization of fungal and bacterial dysbiosis in young adult Chinese patients with Crohn’s disease. Ther Adv Gastroenterol 2020, 13, 1756284820971202. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, C.; Tang, C.; et al. Dysbiosis of Gut Fungal Microbiota is Associated With Mucosal Inflammation in Crohn’s Disease. J Clin Gastroenterol 2014, 48, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Xu, X.; Liang, L.; et al. Lactic Acid-Producing Probiotic Saccharomyces cerevisiae Attenuates Ulcerative Colitis via Suppressing Macrophage Pyroptosis and Modulating Gut Microbiota. Front Immunol 2021, 12, 777665. [Google Scholar] [CrossRef] [PubMed]
- Anon. Comparative immunophenotyping of Saccharomyces cerevisiae and Candida spp. strains from Crohn’s disease patients and their interactions with the gut microbiome - PMC. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7388382/ (accessed on 27 February 2024).
- Li, B.; Zhang, H.; Shi, L.; et al. Saccharomyces boulardii alleviates DSS-induced intestinal barrier dysfunction and inflammation in humanized mice. Food Funct 2022, 13, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Sivananthan, K.; Petersen, AM. Review of Saccharomyces boulardii as a treatment option in IBD. Immunopharmacol Immunotoxicol 2018, 40, 465–475. [Google Scholar] [CrossRef]
- Anon. Dietary fiber and SCFAs in the regulation of mucosal immunity - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/36543697/ (accessed on 27 February 2024).
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; et al. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front Immunol 2019, 10, 277. [Google Scholar] [CrossRef]
- Li, Y.J.; Chen, X.; Kwan, T.K.; et al. Dietary Fiber Protects against Diabetic Nephropathy through Short-Chain Fatty Acid-Mediated Activation of G Protein-Coupled Receptors GPR43 and GPR109A. J Am Soc Nephrol JASN 2020, 31, 1267–1281. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; et al. The role of short-chain fatty acids in health and disease. Adv Immunol 2014, 121, 91–119. [Google Scholar] [CrossRef]
- Li, M.; Esch BCAM van, Wagenaar, G.T.M.; et al. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur J Pharmacol 2018, 831, 52–59. [CrossRef]
- Hu, J.; Cheng, S.; Yao, J.; et al. Correlation between altered gut microbiota and elevated inflammation markers in patients with Crohn’s disease. Front Immunol 2022, 13, 947313. [Google Scholar] [CrossRef]
- Wang, Y.; Yutuc, E.; Griffiths, WJ. Cholesterol metabolism pathways - are the intermediates more important than the products? FEBS J 2021, 288, 3727–3745. [Google Scholar] [CrossRef]
- Cai, J.; Sun, L.; Gonzalez, FJ. Gut microbiota-derived bile acids in intestinal immunity, inflammation, and tumorigenesis. Cell Host Microbe 2022, 30, 289–300. [Google Scholar] [CrossRef]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; et al. Bile acids and the gut microbiota: metabolic interactions and impacts on disease. Nat Rev Microbiol 2023, 21, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Wahlström, A.; Sayin, S.I.; Marschall H-U, et al.; et al. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab 2016, 24, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Darkoh, C.; Lichtenberger, L.M.; Ajami, N.; et al. Bile acids improve the antimicrobial effect of rifaximin. Antimicrob Agents Chemother 2010, 54, 3618–3624. [Google Scholar] [CrossRef] [PubMed]
- Bajor, A.; Gillberg P-G, Abrahamsson H. Bile acids: short and long term effects in the intestine. Scand J Gastroenterol 2010, 45, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.Y.; Aleksunes, L.M.; Tanaka, Y.; et al. Bile acids via FXR initiate the expression of major transporters involved in the enterohepatic circulation of bile acids in newborn mice. Am J Physiol Gastrointest Liver Physiol 2012, 302, G979–996. [Google Scholar] [CrossRef]
- Nie, Y.; Hu, J.; Yan, X. Cross-talk between bile acids and intestinal microbiota in host metabolism and health. J Zhejiang Univ Sci B 2015, 16, 436–446. [Google Scholar] [CrossRef]
- Lavelle, A.; Nancey, S.; Reimund J-M, et al.; et al. Fecal microbiota and bile acids in IBD patients undergoing screening for colorectal cancer. Gut Microbes 2022, 14, 2078620. [Google Scholar] [CrossRef]
- Nijmeijer, R.M.; Gadaleta, R.M.; Mil SWC van, et al. Farnesoid X receptor (FXR) activation and FXR genetic variation in inflammatory bowel disease. PloS One 2011, 6, e23745. [CrossRef]
- Labbé, A.; Ganopolsky, J.G.; Martoni, C.J.; et al. Bacterial bile metabolising gene abundance in Crohn’s, ulcerative colitis and type 2 diabetes metagenomes. PloS One 2014, 9, e115175. [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ma, N.; He, T.; et al. Tryptophan (Trp) modulates gut homeostasis via aryl hydrocarbon receptor (AhR). Crit Rev Food Sci Nutr 2020, 60, 1760–1768. [Google Scholar] [CrossRef] [PubMed]
- Connors, J.; Dawe, N.; Van Limbergen, J. The Role of Succinate in the Regulation of Intestinal Inflammation. Nutrients 2018, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Dvornikova, K.A.; Platonova, O.N.; Bystrova, EY. Inflammatory Bowel Disease: Crosstalk between Histamine, Immunity, and Disease. Int J Mol Sci 2023, 24, 9937. [Google Scholar] [CrossRef] [PubMed]
- Figliuolo, V.R.; Coutinho-Silva, R.; Coutinho, C.M.L.M. Contribution of sulfate-reducing bacteria to homeostasis disruption during intestinal inflammation. Life Sci 2018, 215, 145–151. [Google Scholar] [CrossRef]
- Dubinsky, V.; Dotan, I.; Gophna, U. Carriage of Colibactin-producing Bacteria and Colorectal Cancer Risk. Trends Microbiol 2020, 28, 874–876. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; et al. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef]
- Ellis, J.L.; Karl, J.P.; Oliverio, A.M.; et al. Dietary vitamin K is remodeled by gut microbiota and influences community composition. Gut Microbes 2021, 13, 1–16. [Google Scholar] [CrossRef]
- Pham, V.T.; Dold, S.; Rehman, A.; et al. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr Res N Y N 2021, 95, 35–53. [Google Scholar] [CrossRef]
- Rizzello, F.; Spisni, E.; Giovanardi, E.; et al. Implications of the Westernized Diet in the Onset and Progression of IBD. Nutrients 2019, 11, 1033. [Google Scholar] [CrossRef] [PubMed]
- Vernia, F.; Valvano, M.; Longo, S.; et al. Vitamin D in Inflammatory Bowel Diseases. Mechanisms of Action and Therapeutic Implications. Nutrients 2022, 14, 269. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Akram, W.; Garud, N.; Joshi, R. Role of inulin as prebiotics on inflammatory bowel disease. Drug Discov Ther 2019, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; et al. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat Rev Gastroenterol Hepatol 2019, 16, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Drzymała-Czyż S, Banasiewicz, T.; Tubacka, M.; et al. Inulin supplementation in rat model of pouchitis. Acta Biochim Pol 2011, 58, 381–384. [Google Scholar] [CrossRef]
- Dou, Y.; Yu, X.; Luo, Y.; et al. Effect of Fructooligosaccharides Supplementation on the Gut Microbiota in Human: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 3298. [Google Scholar] [CrossRef]
- Holscher, HD. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Zheng D-W, Li R-Q, An J-X, et al. Prebiotics-Encapsulated Probiotic Spores Regulate Gut Microbiota and Suppress Colon Cancer. Adv Mater Deerfield Beach Fla 2020, 32, e2004529. [CrossRef]
- Matijašić, M.; Meštrović, T.; Perić, M.; et al. Modulating Composition and Metabolic Activity of the Gut Microbiota in IBD Patients. Int J Mol Sci 2016, 17, 578. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- De Greef, E.; Vandenplas, Y.; Hauser, B.; et al. Probiotics and IBD. Acta Gastro-Enterol Belg 2013, 76, 15–19. [Google Scholar]
- La Fata, G.; Weber, P.; Mohajeri, MH. Probiotics and the Gut Immune System: Indirect Regulation. Probiotics Antimicrob Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, AC. A review of dose-responses of probiotics in human studies. Benef Microbes 2017, 8, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xu, S.; Zhang, W.; et al. Probiotic Escherichia coli NISSLE 1917 for inflammatory bowel disease applications. Food Funct 2022, 13, 5914–5924. [Google Scholar] [CrossRef] [PubMed]
- Anon. Probiotics for induction of remission in Crohn’s disease - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/32678465/ (accessed on 27 February 2024).
- Scaldaferri, F.; Gerardi, V.; Mangiola, F.; et al. Role and mechanisms of action of Escherichia coli Nissle 1917 in the maintenance of remission in ulcerative colitis patients: An update. World J Gastroenterol 2016, 22, 5505–5511. [Google Scholar] [CrossRef]
- Oliva, S.; Di Nardo, G.; Ferrari, F.; et al. Randomised clinical trial: the effectiveness of Lactobacillus reuteri ATCC 55730 rectal enema in children with active distal ulcerative colitis. Aliment Pharmacol Ther 2012, 35, 327–334. [Google Scholar] [CrossRef]
- Ma, L.; Lyu, W.; Song, Y.; et al. Anti-Inflammatory Effect of Clostridium butyricum-Derived Extracellular Vesicles in Ulcerative Colitis: Impact on Host microRNAs Expressions and Gut Microbiome Profiles. Mol Nutr Food Res 2023, 67, e2200884. [Google Scholar] [CrossRef]
- Anon. Saccharomyces boulardii in maintenance treatment of Crohn’s disease - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/10961730/ (accessed on 27 February 2024).
- Pan, Y.; Ning, Y.; Hu, J.; et al. The Preventive Effect of Lactobacillus plantarum ZS62 on DSS-Induced IBD by Regulating Oxidative Stress and the Immune Response. Oxid Med Cell Longev 2021, 2021, 9416794. [Google Scholar] [CrossRef]
- Cheng F-S, Pan, D.; Chang, B.; et al. Probiotic mixture VSL#3: An overview of basic and clinical studies in chronic diseases. World J Clin Cases 2020, 8, 1361–1384. [Google Scholar] [CrossRef] [PubMed]
- Tursi, A.; Brandimarte, G.; Papa, A.; et al. Treatment of relapsing mild-to-moderate ulcerative colitis with the probiotic VSL#3 as adjunctive to a standard pharmaceutical treatment: a double-blind, randomized, placebo-controlled study. Am J Gastroenterol 2010, 105, 2218–2227. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.; Midha, V.; Makharia, G.K.; et al. The probiotic preparation, VSL#3 induces remission in patients with mild-to-moderately active ulcerative colitis. Clin Gastroenterol Hepatol Off Clin Pract J Am Gastroenterol Assoc 2009, 7, 1202–1209. [Google Scholar] [CrossRef]
- Mardini, H.E.; Grigorian, AY. Probiotic mix VSL#3 is effective adjunctive therapy for mild to moderately active ulcerative colitis: a meta-analysis. Inflamm Bowel Dis 2014, 20, 1562–1567. [Google Scholar] [PubMed]
- Huynh, H.Q.; deBruyn, J.; Guan, L.; et al. Probiotic preparation VSL#3 induces remission in children with mild to moderate acute ulcerative colitis: a pilot study. Inflamm Bowel Dis 2009, 15, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Miele, E.; Pascarella, F.; Giannetti, E.; et al. Effect of a probiotic preparation (VSL#3) on induction and maintenance of remission in children with ulcerative colitis. Am J Gastroenterol 2009, 104, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Toumi, R.; Abdelouhab, K.; Rafa, H.; et al. Beneficial role of the probiotic mixture Ultrabiotique on maintaining the integrity of intestinal mucosal barrier in DSS-induced experimental colitis. Immunopharmacol Immunotoxicol 2013, 35, 403–409. [Google Scholar] [CrossRef]
- Ghyselinck, J.; Verstrepen, L.; Moens, F.; et al. A 4-strain probiotic supplement influences gut microbiota composition and gut wall function in patients with ulcerative colitis. Int J Pharm 2020, 587, 119648. [Google Scholar] [CrossRef]
- Palumbo, V.D.; Romeo, M.; Marino Gammazza, A.; et al. The long-term effects of probiotics in the therapy of ulcerative colitis: A clinical study. Biomed Pap Med Fac Univ Palacky Olomouc Czechoslov 2016, 160, 372–377. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, L.; Hong, G.; et al. Probiotic mixtures with aerobic constituent promoted the recovery of multi-barriers in DSS-induced chronic colitis. Life Sci 2020, 240, 117089. [Google Scholar] [CrossRef]
- Tan, F.; Deng, Y.; Guo, J.; et al. Effect of mesalazine combined with probiotics on inflammation and immune function of patients with inflammatory bowel disease. Am J Transl Res 2022, 14, 8234. [Google Scholar] [PubMed]
- Martín, R.; Miquel, S.; Benevides, L.; et al. Functional Characterization of Novel Faecalibacterium prausnitzii Strains Isolated from Healthy Volunteers: A Step Forward in the Use of F. prausnitzii as a Next-Generation Probiotic. Front Microbiol 2017, 8, 1226. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Feng, S.; Arjan, N.; et al. A next generation probiotic, Akkermansia muciniphila. Crit Rev Food Sci Nutr 2019, 59, 3227–3236. [Google Scholar] [CrossRef] [PubMed]
- Yasueda, A.; Mizushima, T.; Nezu, R.; et al. The effect of Clostridium butyricum MIYAIRI on the prevention of pouchitis and alteration of the microbiota profile in patients with ulcerative colitis. Surg Today 2016, 46, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Bravo, D.; Landete, JM. Genetic engineering as a powerful tool to improve probiotic strains. Biotechnol Genet Eng Rev 2017, 33, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Goers, L.; Lesser, CF. Emerging strategies for engineering Escherichia coli Nissle 1917-based therapeutics. Trends Pharmacol Sci 2022, 43, 772–786. [Google Scholar] [CrossRef] [PubMed]
- Chappell, T.C.; Nair, NU. Engineered lactobacilli display anti-biofilm and growth suppressing activities against Pseudomonas aeruginosa. NPJ Biofilms Microbiomes 2020, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Zuo, F.; Chen, S.; Marcotte, H. Engineer probiotic bifidobacteria for food and biomedical applications - Current status and future prospective. Biotechnol Adv 2020, 45, 107654. [Google Scholar] [CrossRef]
- Darb Emamie, A.; Rajabpour, M.; Ghanavati, R.; et al. The effects of probiotics, prebiotics and synbiotics on the reduction of IBD complications, a periodic review during 2009-2020. J Appl Microbiol 2021, 130, 1823–1838. [Google Scholar] [CrossRef]
- Haskey, N.; Dahl, WJ. Synbiotic therapy: a promising new adjunctive therapy for ulcerative colitis. Nutr Rev 2006, 64, 132–138. [Google Scholar] [CrossRef]
- Steed, H.; Macfarlane, G.T.; Blackett, K.L.; et al. Clinical trial: the microbiological and immunological effects of synbiotic consumption - a randomized double-blind placebo-controlled study in active Crohn’s disease. Aliment Pharmacol Ther 2010, 32, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Liu, M.; Pu, J.; et al. Probiotics and Their Metabolites Ameliorate Inflammatory Bowel Disease: A Critical Review. Infect Microbes Dis 2021, 3, 4–13. [Google Scholar] [CrossRef]
- Cheng Y-W, Fischer M. Fecal Microbiota Transplantation. Clin Colon Rectal Surg 2023, 36, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Weingarden, A.R.; Vaughn, BP. Intestinal microbiota, fecal microbiota transplantation, and inflammatory bowel disease. Gut Microbes 2017, 8, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Hvas, C.L.; Dahl Jørgensen, S.M.; Jørgensen, S.P.; et al. Fecal Microbiota Transplantation Is Superior to Fidaxomicin for Treatment of Recurrent Clostridium difficile Infection. Gastroenterology 2019, 156, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Anon. Fecal Microbiota Transplantation in Inflammatory Bowel Disease - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/37189634/ (accessed on 28 February 2024).
- Costello, S.P.; Hughes, P.A.; Waters, O.; et al. Effect of Fecal Microbiota Transplantation on 8-Week Remission in Patients With Ulcerative Colitis: A Randomized Clinical Trial. JAMA 2019, 321, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhou, Y.; Huang, S.; et al. Fecal microbiota transplantation for ulcerative colitis: a prospective clinical study. BMC Gastroenterol 2019, 19, 116. [Google Scholar] [CrossRef]
- Colman, R.J.; Rubin, DT. Fecal microbiota transplantation as therapy for inflammatory bowel disease: a systematic review and meta-analysis. J Crohns Colitis 2014, 8, 1569–1581. [Google Scholar] [CrossRef]
- Paramsothy, S.; Paramsothy, R.; Rubin, D.T.; et al. Faecal Microbiota Transplantation for Inflammatory Bowel Disease: A Systematic Review and Meta-analysis. J Crohns Colitis 2017, 11, 1180–1199. [Google Scholar] [CrossRef]
- Caldeira L de, F.; Borba, H.H.; Tonin, F.S.; et al. Fecal microbiota transplantation in inflammatory bowel disease patients: A systematic review and meta-analysis. PloS One 2020, 15, e0238910. [Google Scholar] [CrossRef]
- Moayyedi, P.; Surette, M.G.; Kim, P.T.; et al. Fecal Microbiota Transplantation Induces Remission in Patients With Active Ulcerative Colitis in a Randomized Controlled Trial. Gastroenterology 2015, 149, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Paramsothy, S.; Kamm, M.A.; Kaakoush, N.O.; et al. Multidonor intensive faecal microbiota transplantation for active ulcerative colitis: a randomised placebo-controlled trial. Lancet Lond Engl 2017, 389, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Li, P.; Xu, L.; et al. Step-up fecal microbiota transplantation strategy: a pilot study for steroid-dependent ulcerative colitis. J Transl Med 2015, 13, 298. [Google Scholar] [CrossRef] [PubMed]
- Anon. Safety, tolerability, and clinical response after fecal transplantation in children and young adults with ulcerative colitis - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/23542823/ (accessed on 28 February 2024).
- He, Z.; Li, P.; Zhu, J.; et al. Multiple fresh fecal microbiota transplants induces and maintains clinical remission in Crohn’s disease complicated with inflammatory mass. Sci Rep 2017, 7, 4753. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Bu, C.; Yuan, W.; et al. Fecal Microbiota Transplant via Endoscopic Delivering Through Small Intestine and Colon: No Difference for Crohn’s Disease. Dig Dis Sci 2020, 65, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Landman, C.; Seksik, P.; et al. Fecal microbiota transplantation to maintain remission in Crohn’s disease: a pilot randomized controlled study. Microbiome 2020, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Kellermayer, R.; Nagy-Szakal, D.; Harris, R.A.; et al. Serial fecal microbiota transplantation alters mucosal gene expression in pediatric ulcerative colitis. Am J Gastroenterol 2015, 110, 604–606. [Google Scholar] [CrossRef]
- Kedia, S.; Virmani, S.; K Vuyyuru, S.; et al. Faecal microbiota transplantation with anti-inflammatory diet (FMT-AID) followed by anti-inflammatory diet alone is effective in inducing and maintaining remission over 1 year in mild to moderate ulcerative colitis: a randomised controlled trial. Gut 2022, 71, 2401–2413. [Google Scholar] [CrossRef]
- Okahara, K.; Ishikawa, D.; Nomura, K.; et al. Matching between Donors and Ulcerative Colitis Patients Is Important for Long-Term Maintenance after Fecal Microbiota Transplantation. J Clin Med 2020, 9, 1650. [Google Scholar] [CrossRef]
- Angelberger, S.; Reinisch, W.; Makristathis, A.; et al. Temporal bacterial community dynamics vary among ulcerative colitis patients after fecal microbiota transplantation. Am J Gastroenterol 2013, 108, 1620–1630. [Google Scholar] [CrossRef]
- Suskind, D.L.; Singh, N.; Nielson, H.; et al. Fecal microbial transplant via nasogastric tube for active pediatric ulcerative colitis. J Pediatr Gastroenterol Nutr 2015, 60, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Baxter, M.; Colville, A. Adverse events in faecal microbiota transplant: a review of the literature. J Hosp Infect 2016, 92, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.S.; Mentzel, C.M.J.; Kot, W.; et al. Faecal virome transplantation decreases symptoms of type 2 diabetes and obesity in a murine model. Gut 2020, 69, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Draper, L.A.; Ryan, F.J.; Dalmasso, M.; et al. Autochthonous faecal viral transfer (FVT) impacts the murine microbiome after antibiotic perturbation. BMC Biol 2020, 18, 173. [Google Scholar] [CrossRef] [PubMed]
- Jansen, D.; Falony, G.; Vieira-Silva, S.; et al. Community Types of the Human Gut Virome are Associated with Endoscopic Outcome in Ulcerative Colitis. J Crohns Colitis 2023, 17, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Ott, S.J.; Waetzig, G.H.; Rehman, A.; et al. Efficacy of Sterile Fecal Filtrate Transfer for Treating Patients With Clostridium difficile Infection. Gastroenterology 2017, 152, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, F.; Massimino, L.; D’Alessio, S.; et al. The gut virome in inflammatory bowel disease pathogenesis: From metagenomics to novel therapeutic approaches. United Eur Gastroenterol J 2019, 7, 999–1007. [Google Scholar] [CrossRef]
- Yu, T.; Guo, F.; Yu, Y.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563. [Google Scholar] [CrossRef]
- Dong, X.; Pan, P.; Zheng D-W, et al; et al. Bioinorganic hybrid bacteriophage for modulation of intestinal microbiota to remodel tumor-immune microenvironment against colorectal cancer. Sci Adv 2020, 6, eaba1590. [Google Scholar] [CrossRef]
- Fuerte-Stone, J.; Mimee, M. Host happy hour: Phage cocktail targets IBD-associated microbes. Cell Host Microbe 2022, 30, 1352–1353. [Google Scholar] [CrossRef]
- Federici, S.; Kredo-Russo, S.; Valdés-Mas, R.; et al. Targeted suppression of human IBD-associated gut microbiota commensals by phage consortia for treatment of intestinal inflammation. Cell 2022, 185, 2879–2898. [Google Scholar] [CrossRef]
- Dridi, B.; Fardeau M-L, Ollivier, B.; et al. The antimicrobial resistance pattern of cultured human methanogens reflects the unique phylogenetic position of archaea. J Antimicrob Chemother 2011, 66, 2038–2044. [Google Scholar] [CrossRef]
- Gottlieb, K.; Wacher, V.; Sliman, J.; et al. Review article: inhibition of methanogenic archaea by statins as a targeted management strategy for constipation and related disorders. Aliment Pharmacol Ther 2016, 43, 197–212. [Google Scholar] [CrossRef]
- Plein, K.; Hotz, J. Therapeutic effects of Saccharomyces boulardii on mild residual symptoms in a stable phase of Crohn’s disease with special respect to chronic diarrhea--a pilot study. Z Gastroenterol 1993, 31, 129–134. [Google Scholar]
Figure 1.
The gut microbiota of a healthy individual (left) compared to that of a patient with inflammatory bowel disease (IBD) (right).
Figure 1.
The gut microbiota of a healthy individual (left) compared to that of a patient with inflammatory bowel disease (IBD) (right).
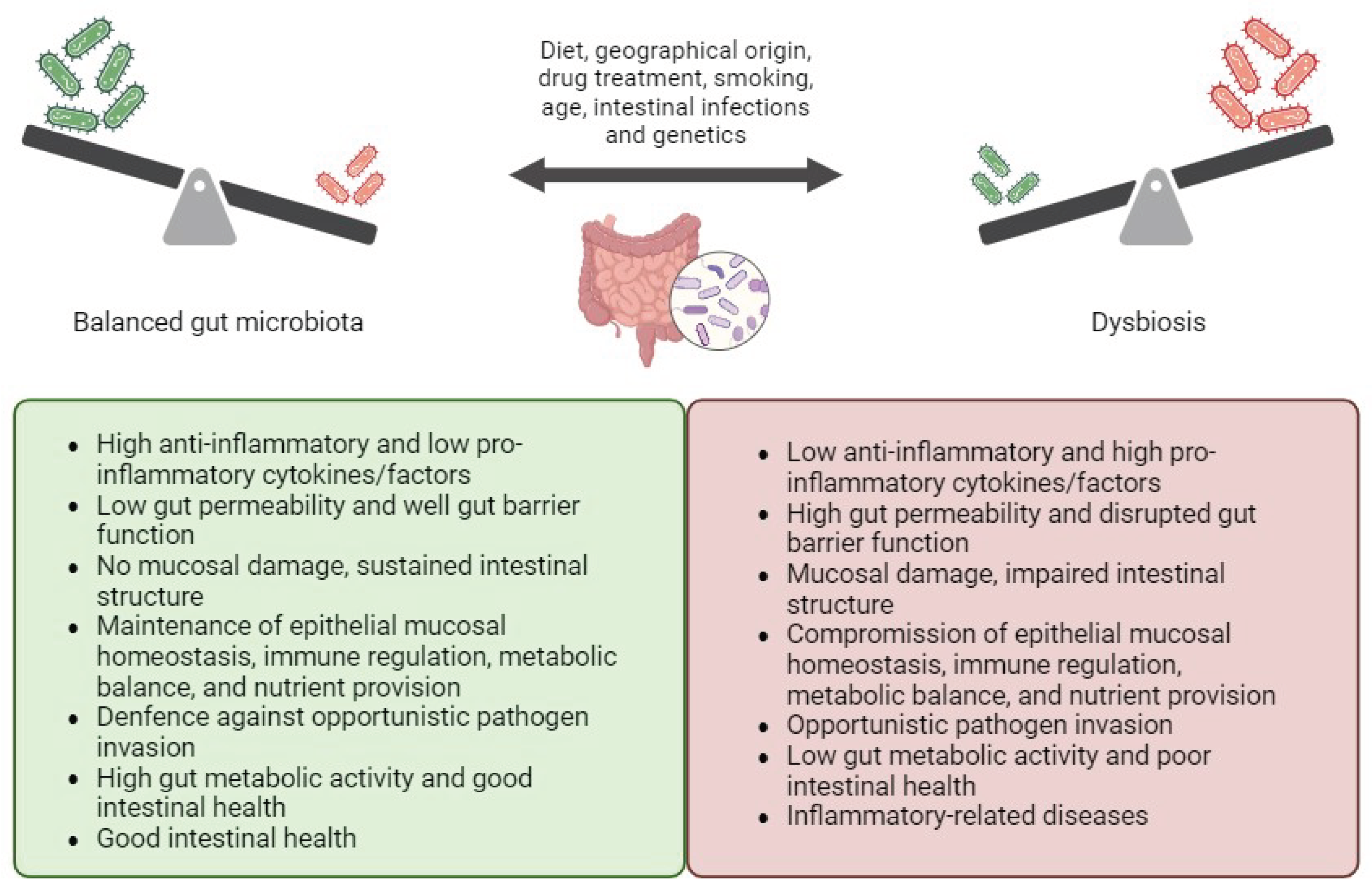
Figure 2.
Dysbiosis, leaky gut and proinflammatory intestinal and systemic response.
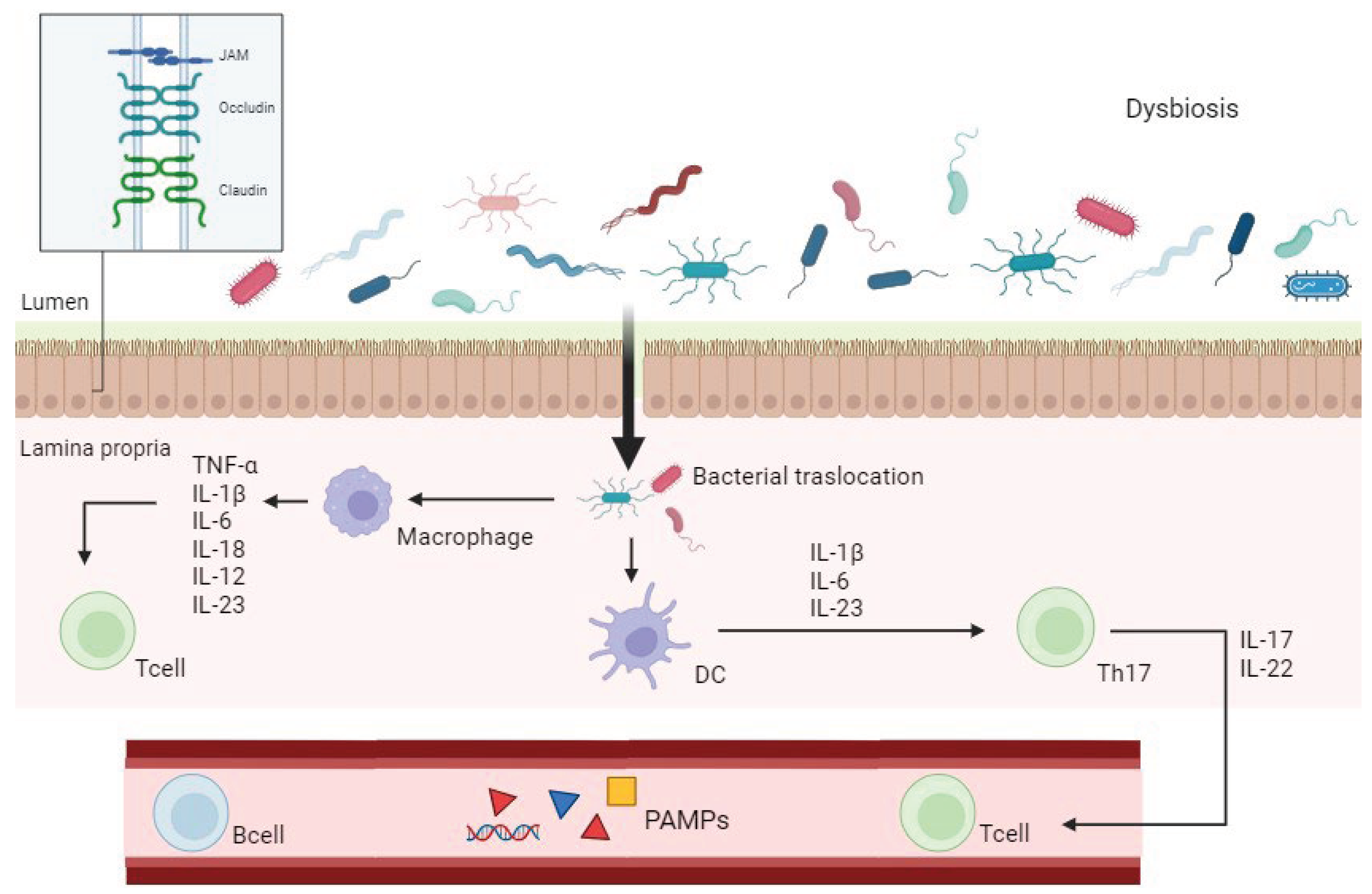
Figure 3.
Diagram of bacteria increased and decreased in inflammatory bowel disease (IBD) along with their respective actions on the immune system.
Figure 3.
Diagram of bacteria increased and decreased in inflammatory bowel disease (IBD) along with their respective actions on the immune system.
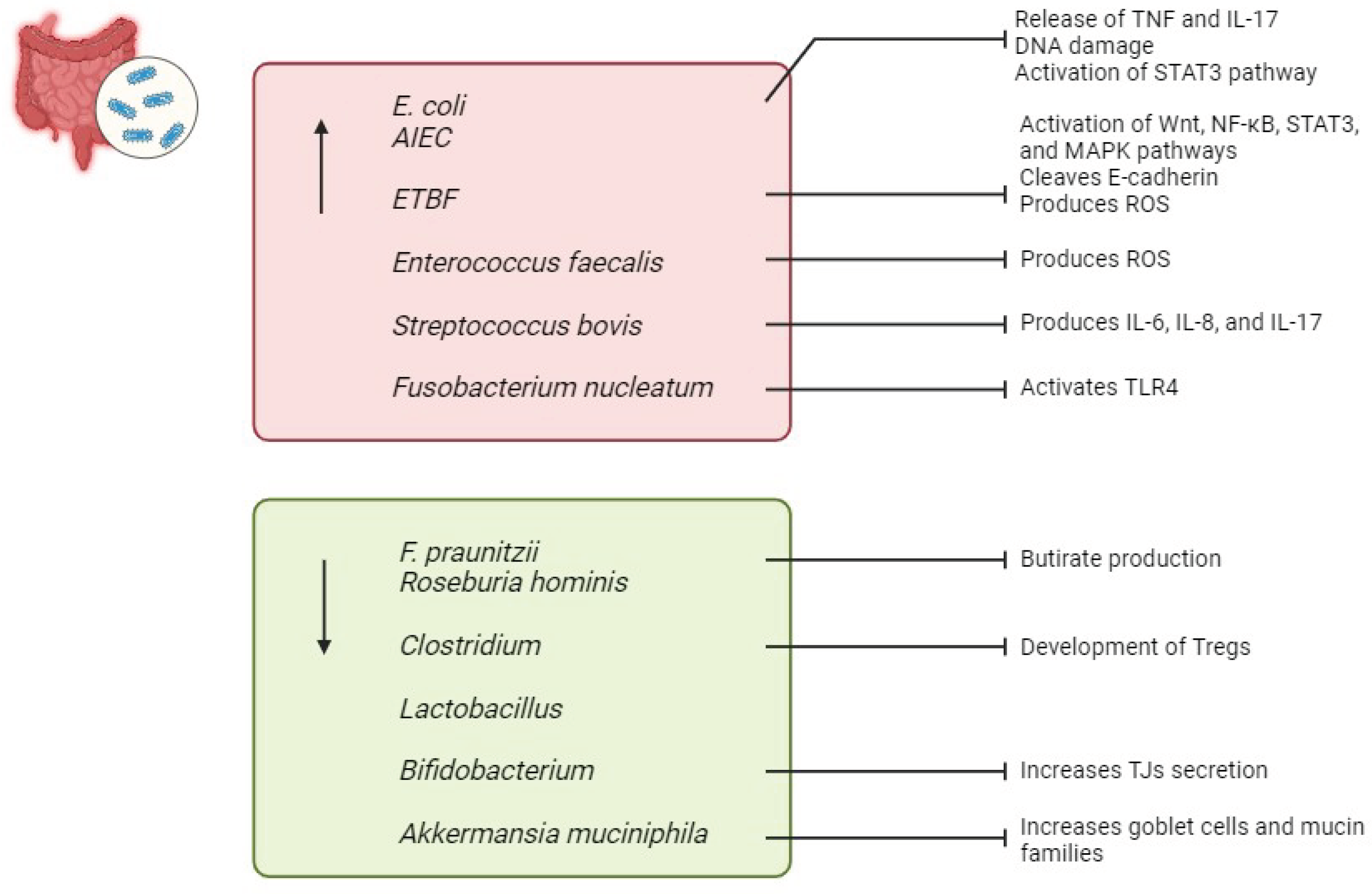
Figure 4.
Diagram of viruses increased and decreased in inflammatory bowel disease (IBD) along with their respective actions on the intestinal environment and immune system.
Figure 4.
Diagram of viruses increased and decreased in inflammatory bowel disease (IBD) along with their respective actions on the intestinal environment and immune system.
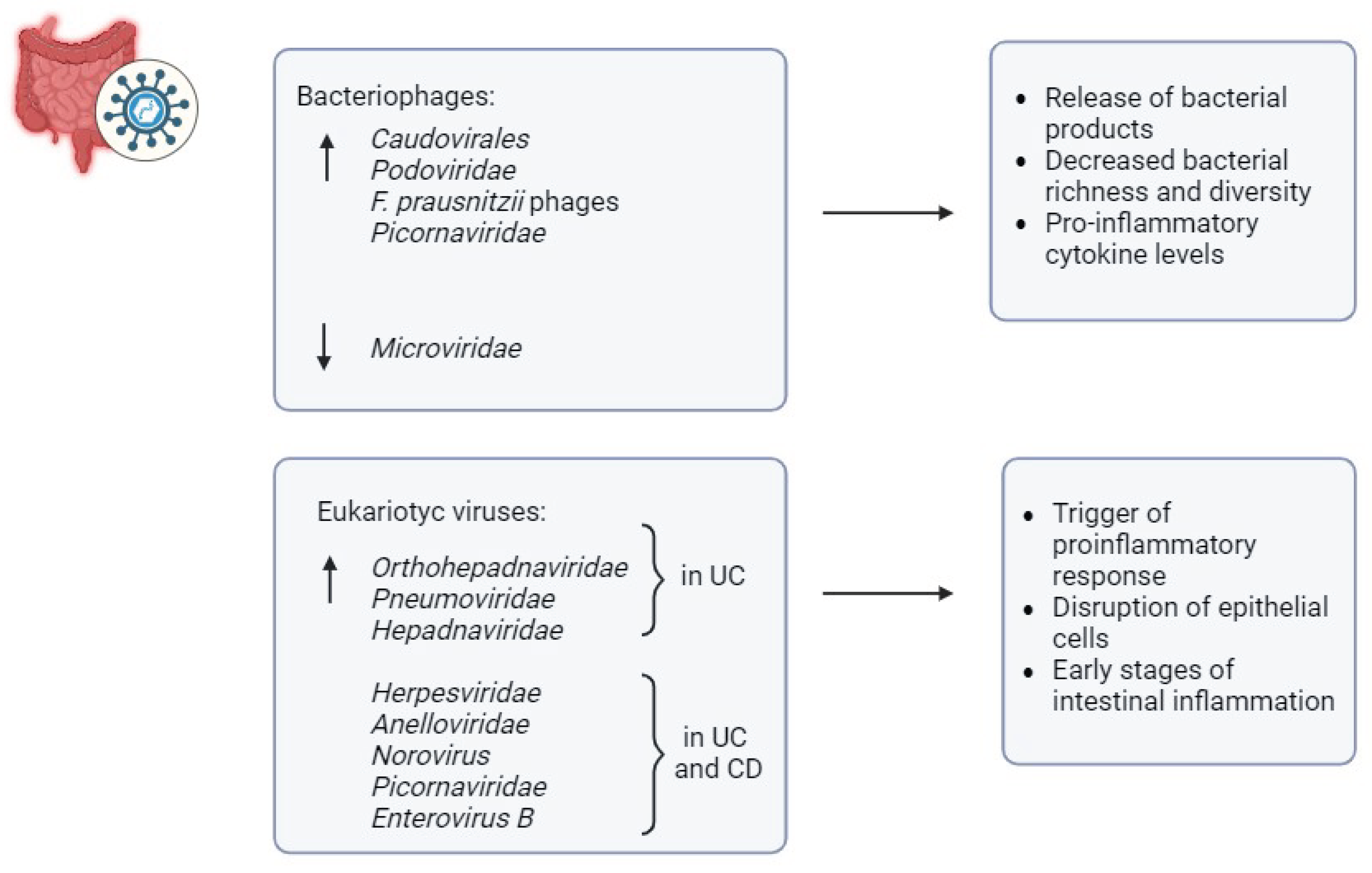
Figure 5.
Diagram of archaea increased and decreased in inflammatory bowel disease (IBD) along with their respective actions.
Figure 5.
Diagram of archaea increased and decreased in inflammatory bowel disease (IBD) along with their respective actions.
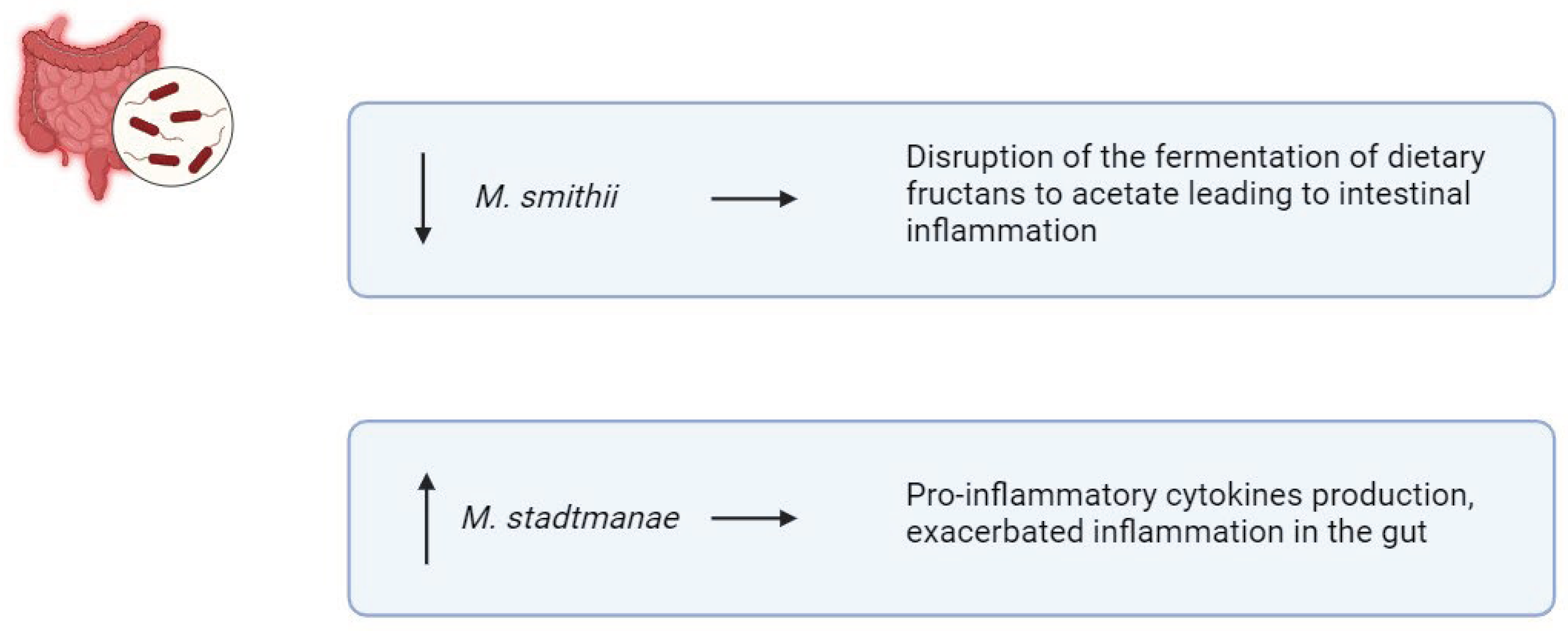
Figure 6.
Diagram of fungi increased and decreased in inflammatory bowel disease (IBD) along with their respective actions.
Figure 6.
Diagram of fungi increased and decreased in inflammatory bowel disease (IBD) along with their respective actions.
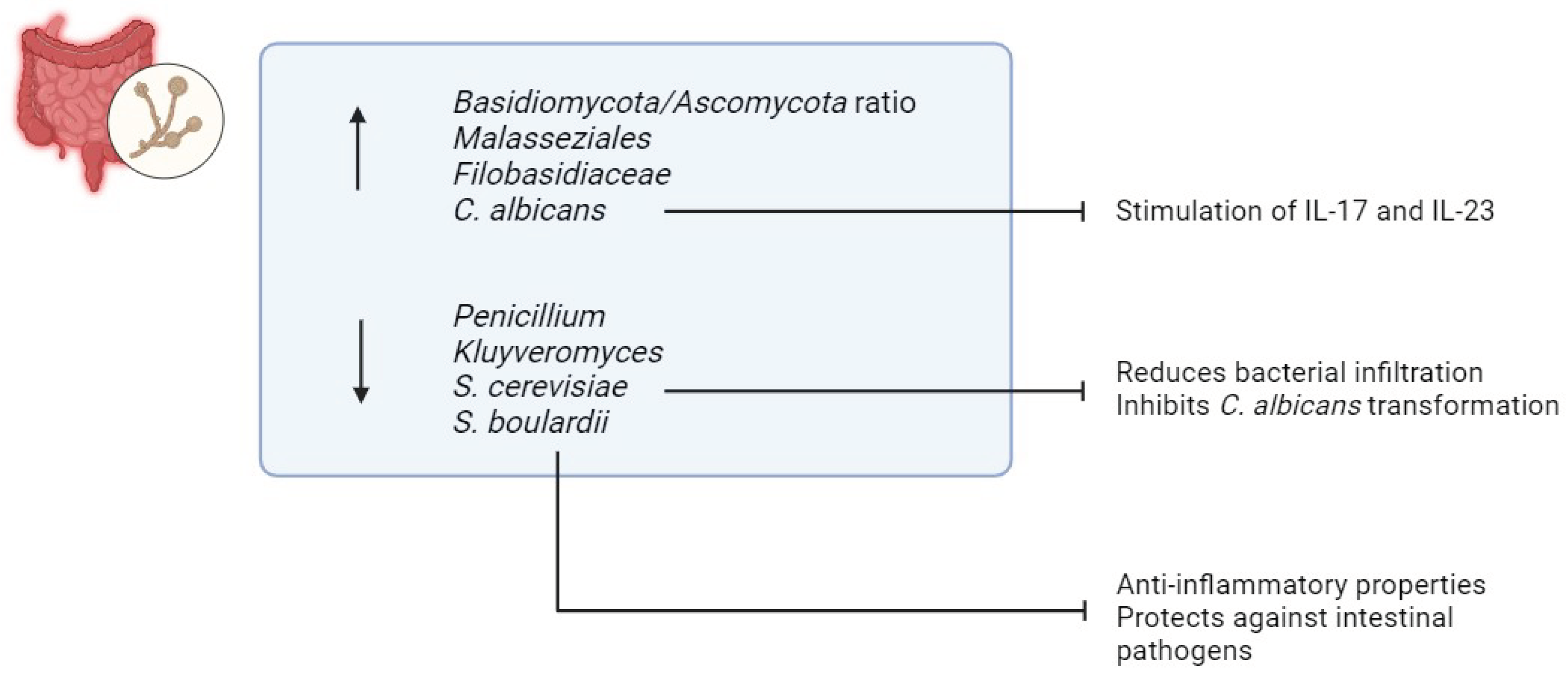
Figure 7.
Short-chain fatty acids (SCFAs) and their effects on the host.
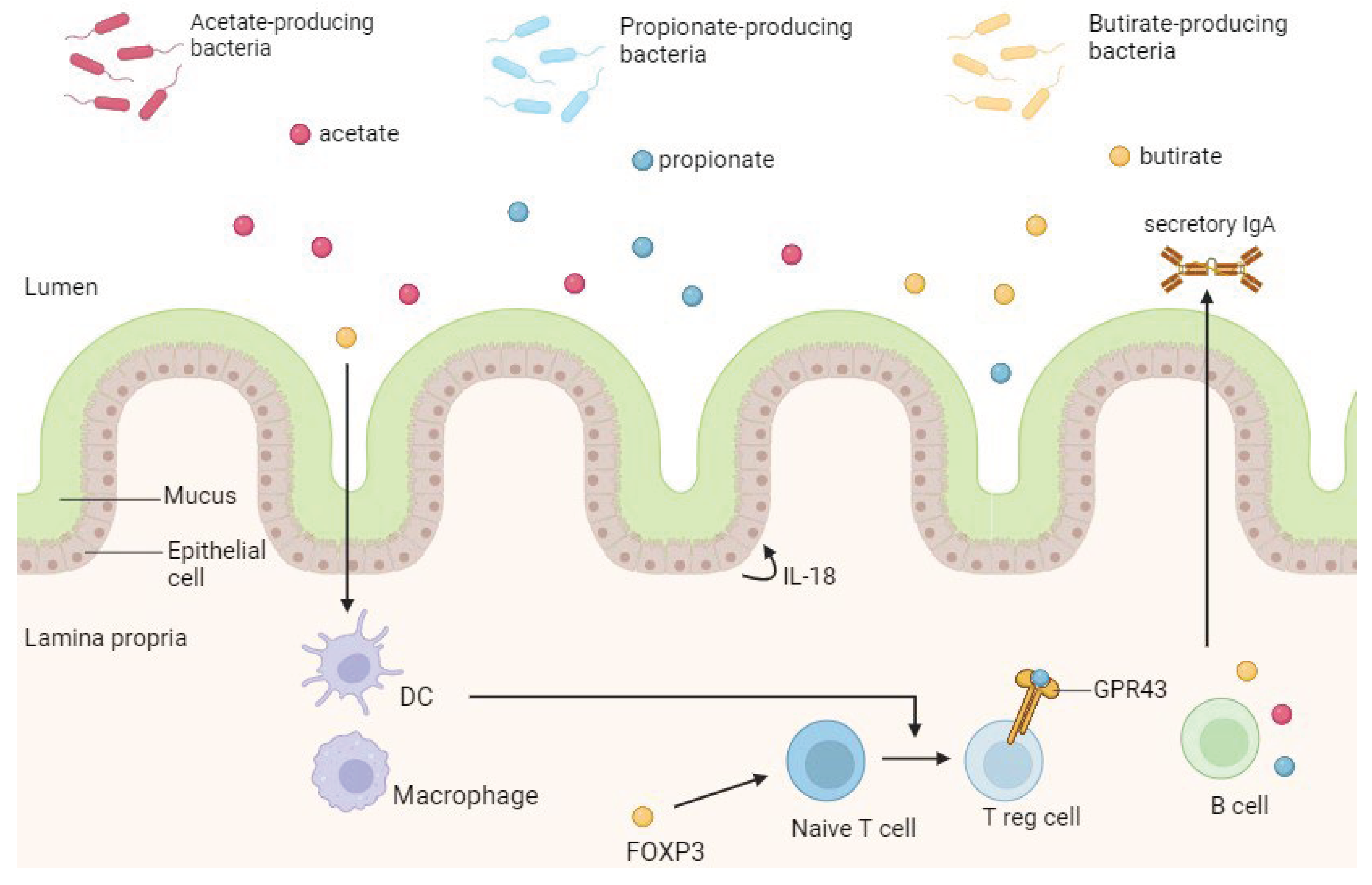
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



