
Submitted:
04 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
The global craft beer market is witnessing substantial growth, with a particular spotlight on sour beers. These unique brews are created through lactic fermentation initiated by lactic acid bacteria (LAB). However, the presence of hops, renowned for their antimicrobial properties, can pose a limitation within the realm of sour beer production. This research aims to explore and assess the potential of sour beer as a probiotic beverage. It involves a comparative analysis of the viability of Lacticaseibacillus paracasei subsp. paracasei F19 (F19) and 431 (L431) in a sour beer with an International Bitterness Unit (IBU) value of 26.65, alongside examining the ex-pression of genes such as horA, horB, horC, hitA, bsrA, and recA, which are associated with hop resistance. The study encompasses the development of two formulations of sour beers characterized by both high hop content and a robust probiotic count (8.44 – 8.77 log CFU/mL). The findings suggest that both probiotic strains of F19 and L431, are well-suited to produce sour beers with elevated hop levels, demonstrating excellent stability. Notably, the expression of genes responsible for hops resistance exhibited distinct modulation patterns among the two strains. It appears that a higher concentration of bsrA, as observed in strain L431, may be more effective in mitigating the effects of hop-related stress, potentially surpassing the combined expression of horA and bsrA. Nonetheless, further research is essential to validate this observation in gene expression. F19 and L431 emerge as promising candidates for the development of sour beers with increased hop intensity.
Keywords:
craft beer
; probiotic
; gene expression
; PMA-qPCR
; RT-qPCR
; microbiological resistance to hops.
1. Introduction
Beer is one the most popular beverages in the world. The boom of craft breweries, which began in the 1980s in the United States, reached its pinnacle worldwide in 2016 and has since continued to thrive, fostering competition both among large breweries and within the realm of smaller ones. In 2019, the United States achieved a significant milestone by hosting the largest number of small breweries, boasting a total of 8,386 microbreweries, brewpubs, and regional craft breweries, each contributing to a rich tapestry of over 20,000 distinct craft beer brands [1]. This remarkable feat solidified the United States' position as the epicenter of microbrewery culture. Moreover, in this country, the craft beer market was reported as representing a 13.6% market share in sales volumes and a 25.2% market share in dollar sales [2]. In 2019, Americans spent $ 116 billion on beer, with $ 29.3 billion on craft beers [3].
Over the past years, Brazil has witnessed a remarkable 91% surge in the number of breweries [4]. Among the diverse array of craft beers, sour beers emerge as a standout category, primarily due to their distinctive rustic charm. These beers are characterized by their extraordinary complexity, brought about by lactic fermentation driven by lactic acid bacteria (LAB) [5]. Moreover, numerous species of Lactobacillus are recognized for their probiotic qualities [6]. Probiotics are defined as "live microorganisms that bestow health benefits upon the host" [7], and their well-established potential for promoting health is widely acknowledged [6]. Consequently, sour beers represent a compelling choice for incorporating probiotic strains into their fermentation process.
Most sour beers are also known for their low hop (Humulus lupulus) content, and hops are known to have antimicrobial properties [5]. They are also considered as beneficial to health [8], mainly because of polyphenols in various chronic diseases, including insomnia and type II diabetes, and xanthohumol (XN), which is being explored as an antimicrobial, anticarcinogenic, antidiabetic, and anti-inflammatory agent [9].
Some sours beers with probiotics have been developed. Alcine Chan et al [10] promoted the first study demonstrating the feasibility of using the probiotic Lacticaseibacillus paracasei L26 strain as initial culture in the manufacture of beer. Silva et al [11] developed another sour beer with the probiotic strain Lacticaseibacillus paracasei DTA-81. More recently, it has been demonstrated that probiotic strains of Lacticaseibacillus paracasei subsp. paracasei F19 [12,13] and 431 [13] are strong contenders for utilization in sour beer production, given their impressive survival rates. However, these studies used normal amounts of hops, typically, not exceeding 10 International Bitterness Units (IBU). According to the American Society of Brewing Chemists [14], one IBU is equivalent to 1 mg of iso-α-acids per liter.
Although α-acids are harmful to lactic acid bacteria (LABs), many of these bacteria have mechanisms to for self-protection against these compounds. Among them, there are some genes that are related to Hop-Resistance by the bacteria that grow in beer, which includes horA, horB, horC, hitA, bsrA, and recA [5].
Therefore, this study aimed to evaluate the production of a sour beer with high hop, according to Beer Judge Certification Program (BJCP) (https://www.bjcp.org/) for the specifications of a sour beer content, using the probiotic Lacticaseibacillus paracasei subsp. paracasei F19 (F19) and 431 (L431) strains for the lactic acid fermentation. Moreover, to evaluate the viability of the microorganisms during the manufacturing process and storage, as well as to determine gene expression of genes associated with hops resistance.
2. Materials and Methods
2.1. Cultures Employed
For the fermentation and brewery of sour beers, any bacteria or yeast that impart an acidic flavor can be used. However, when using LAB, whether probiotic or not, to produce a sour beer, this beer does not have a high quantity of hops, as its α-acids have antimicrobial effects [5]. Based on their abilities and survival in other food matrices, the known probiotic strains of Lacticaseibacillus paracasei subsp. paracasei F19 (F19) [12,13,15,16] and 431 (L431) [13,17], from Chr. Hansen (Hørsholm, Denmark), were chosen. The yeast Saccharomyces cerevisiae strain Safale US-05 (US-05) (Fermentis, Marq en Baroeul, France) was used due to its features, including dry and balanced pure profile brewery capacity with low amounts of diacetyl under an ideal temperature (18 to 28 °C) [18].
The strains activation was performed with two successive transfers from frozen probiotics (−80 °C) to the following media: DeMan, Rogosa and Sharpe (MRS) broth (Oxoid, Thermo Fisher Scientific, Waltham, Massachusetts, USA) for F19 and 431 at 37 °C for 24 hours in anaerobiosis (AnaeroGen Anaerobic System, Oxoid).
2.2. Formulation, and Brewery of the Sour Beer
Sour beers must not exceed 10 IBU (International Bitterness Units). However, as our objective was to evaluate the survival of probiotic strains in high hop wort, a recipe with 29.65 IBU was formulated. The beer formulations were designed by the BeerSmithTM software, with the following parameters: final pH of 3.9, SRM (Standard Reference Method) 4, OG (Initial Gravity) 1.045, FG (Final Gravity) 1.008, IBU (International Bitterness Unit) 29.65, and ABV (Alcohol by Volume) 4.0%. To achieve these characteristics in two duplicate formulations (n=4), plus a control formulation (n=1 5 liters of must (1 L for each n) 50% (0.73 kg for 5 L) of Chateau Pilsen malt and 50% (0.73 kg for 5 L) of Chateau wheat malt were used (both malt from Castle Malting, Belgian), with Saaz hop (BarthHaas Group®, Nürnberg, Germany) (3.0% α-acid) in the amount of 16 g to reach the IBU of 29.65 to 5L of wort. The brewing process was divided in three steps: 1) 50 ºC for 12 minutes; 2) 72 ºC for 60 minutes; 3) 76 ºC for 10 minutes (Figure 1).
After mashing out and filtration, the wort was boiled for one hour with the addition of hops, and then cooled to 37 °C. After cooling, the following strains were inoculated: one with F19 and other with 431, reaching a pH of approximately 3.9. After the initial lactic fermentation, which lasted 24 hours, US-05 was inoculated, and the second fermentation carried out for 8 days at 18 ºC, followed by the maturation (2 days at 13 ºC, 3 days at 4 ºC, and 4 days at 0 ºC) and carbonation (11 days at 24 ºC) steps. During fermentation and other production processes, the pH was monitored by the equipment Orion Three Stars (Thermo Fisher Scientific).
2.3. Determination of pH and Microbiological Analyses
The pH values were assessed with the Orion Three Stars equipment (Thermo Fisher Scientific), using a penetration electrode, model 2A04-GF (Analyzer), in triplicate. The viability of the probiotic strains employed in the sour beer was determined by plate-count and PMA-qPCR techniques on day 1 (24 h after the first fermentation), on days 8, 10, 13, 17, and 28, and after 30 days (58 days) of storage at 4 ºC, which are critical periods due to changes in the brewery process. The S. cerevisiae US-05 viability was determined only by PMA-qPCR.
Each formulation was submitted to successive serial decimal dilutions in sterile saline solution (NaCl, 0.85 g/100 mL), not only as a counter-test to PMA-qPCR, as well as to assess the strain stress. The plates were incubated at 37 °C for 48 h in anaerobic conditions (AnaeroGenTM Anaerobic System, Oxoid), after 1 mL aliquots of each dilution were pour plated in acidified MRS agar (Oxoid).
2.4. PMA Treatment and DNA and RNA Extraction
The Before performing DNA extraction, and so that only DNA from living cells would be extracted, the samples were treated with PMA (propidium monoazide). PMA is an acid nucleic intercalator which penetrates the damaged cells and form stable covalent high-affinity bonds with DNA, following photo-activation exposure to strong visible light and, in this way, differentiate between live and dead or membrane-damaged bacteria [19].
Cell pellets were collected by centrifugation (9000 g/10 min/4°C), washed twice with Tris EDTA (TE) buffer (10 mM Tris-HCL, 1 mM EDTA, pH 8). Samples were treated with PMA in order not to have an overestimated number of cells in the quantitative PCR (qPCR) by amplification of dead cell DNA [20]. In addition to binding to dead cell DNA, PMA also binds to sample RNA as it binds to any nucleic acid [19]. Thus, for gene expression analysis, the samples were not treated with PMA.
DNA and RNA extraction were performed by the MagMAXTM kit (Thermo Fisher Scientific) and DynaMag™-2 Magnet (Thermo Fisher Scientific) rack was used. Purity quality and concentration of the nucleic acids were measured using a NanoPhotometer® N60 spectrophotometer (Implen, Munich, Germany), and the integrity analyzed using a 1% agarose gel.
2.5. PMA-qPCR
The qPCR was performed using an ABI real-time system 7500™ (Thermo Fisher Scientific) thermocycler with the followed amplification reactions: 12.5 µL of 2X Power SYBR® Green PCR Master Mix Applied Biosystems™ (Thermo Fisher Scientific); 5 µL of the DNA sample, each primer at the appropriate concentration of 100 nM, and q.s. ultrapure water to complete 25 µL. The primers cycling conditions were as follows: initial step at 50 °C for 2 min and 95 °C for 10 min, and 40 cycles at 95 °C for 15 sec and 60 °C for 30 sec. For Saccharomyces cerevisiae (0.7 mM of each respective primer), the reaction conditions were as follows: an initial step at 50 °C for 2 min and 95 °C for 10 min and 40 cycles of 95 °C for 15 s, 60 °C for 1 min and 72 °C for 30 s. Primers for F19 were designed according to Sieuwerts and Håkansson [21], for L431 according to Byun et al. [22], and for the yeast according to Zott et al. [23].
The standard curves were built with 10-fold dilution series of the genomic DNA isolated from the pure cultures of F19 (genome size: 3,063,698 bp; GenBank: CP016355.1), L431 (genome size: 3,063,698 bp; GenBank: CP016355.1), and S. cerevisiae (genome size: 12,165,468 bp; GenBank: ASM308665v1) from 100 to 1×108 genome copies per amplification reaction. The viable cell number were determined by the comparison of the threshold cycle (Ct) of each sample with the standard curves [20].
2.6. RT-qPCR for Expression of Hop Resistance Genes
For gene expression, Reverse Transcription followed by quantitative Polymerase Chain Reaction (RT-qPCR) were used with samples collected during day 2 of fermentation, where there would be greater activity of probiotic strains. RNA was treatment using DNase I (Thermo Fisher Scientific) and reverse transcription was performed using the High-Capacity RNA-to-cDNA Master Mix (Thermo Fisher Scientific), following the manufacturer’s instructions. Next, qPCR was performed using the ABI real-time system 7500™ (Thermo Fisher Scientific, Waltham) thermocycler and the SYBR® Green PCR Master Mix (Thermo Fisher Scientific). The primers for the target messenger RNAs were described by Bergsveinson et al [24]. The housekeeping gene 16S rRNA was used as an internal control.
The results were expressed as mean ± standard deviation (SD). To quantify the viable cell number, the Ct values were automatically converted to CFU equivalent/mL by the 7500 Real-Time PCR System Software Applied Biosystems™ (Thermo Fisher Scientific).
2.7. Statistical Analysis
The homogeneity and normal distribution of the collected data were evaluated. Once a normal distribution was found, the Student’s t-test was applied to compare two samples and, for three or more samples, the Analysis of Variance (ANOVA) followed by Tuckey test was applied. Results were considered as significantly different when p ≤ 0.05 (significance level of 5%). Minitab software, version 19, was used for the statistical analysis.
3. Results and Discussion
3.1. pH and Microbial Populations
The initial pH of the wort was 5.9 and it was reduced to 4.6 - 4.7 after the first fermentation (Table 1). To evaluate the pH, a control sample was used, in which no probiotic strain was inoculated, but only the yeast US-05. At the end of brewery, this control formulation reached a pH of 4.2, while the formulations with probiotic strains had a pH of 3.9 (Table 1).
In general, beer is an inhospitable environment to bacteria, due to its compounds and characteristics, such as the presence of iso-α-acids, ethanol, low pH, high CO2, and low O2 [5]. Using ANOVA and comparing counts obtained by pour plating and by PMA-qPCR (Table 1), the PMA-qPCR revealed much higher populations (p<0.05), indicating that, even with a satisfactory count, the LABs were under stress, which is understandable, due to the high quantity of hops in the formulations. This difference between pour-plate and PMA-qPCR counts is not surprising, as beer is a stressful vehicle for bacteria.
The daily amount of probiotic ingested is still a matter of debate among scientific sources. According to the government of Canada [25], a food product must contain a minimum level of 1.0 x 109 CFU/mL of one or more probiotic microorganisms.
In our study, probiotic cultures were stable throughout the production process and storage, showing a population above 10 log CFU per daily portion of 350 mL in almost all steps. These formulations not only achieved the requirements to be considered beverage with probiotic potential, but also show stability in the cultures used. There were no differences in populations between the F19 and L431 strains by PMA-qPCR. Therefore, both strains have the same growth advantage and are good candidates for fermenting hop beers.
Similarly, Alcine Chan et al [10] reported stability in yeast and LAB probiotic populations. The authors concluded that co-culture of the probiotic strain Lacticaseibacillus paracasei L26 with S. cerevisiae S-04 in unhopped wort showed excellent growth and stability and the yeast performance was not affected by the probiotic strain. Likewise, the growth of S. cerevisiae strain US-05 (US-05) in our study was not affected. In addition, like the study performed by Dysvik et al [26], our data showed that the LAB strains did not interference in the yeast growth.
3.2. A-Acids Stress and Hop Resistance Genes Expression
The presence of iso-α-acids was reported as inhibitory for bacterial growth [5]. The mechanism of action of these acids is simple. As they are weak acids, they have an affinity for the cytoplasm of bacterial cells and undissociated can cross the cell membranes of bacteria. Once in their cytoplasm, they promote the release of protons and, consequently, the drop in intracellular pH, affecting the metabolism of the entire cell [5].
Some strains are resistant to the antimicrobial actions of the hops [27]. This ability is due the action of the specific genes here analyzed. In the gene expression analysis, Recombination Factor A (recA) did not show any expression in both formulations. The expression of the other genes was well evidenced in the two beer samples (Figure 2). However, with some differences among them: the expression of Hop-Resistant B (horB) was observed to be absent in F19, while the expression of Hop-Resistant A (horA) was absent and, of Beer-Spoilage Related A (bsrA) was significantly higher in 431 (p<0.05).
It is known that hops provide protection to beer due to the concentration of iso-α-acids, which have a bacteriostatic effect [5,27]. Despite this imposed barrier, beer spoilage is not uncommon. These genes, which confer hops or α-acids resistance, for our study are of great interest, even as genetic molecular markers, since the strains here studied bring health benefits [16,17,28,29]. Among these genes, horA and horC are considered the most important ones.
The horA gene encodes an ABC transporter capable of expelling hop bitter acids from cells [30]. The horC gene encodes a proton motive force (PMF)-dependent multidrug effluence pump [31], and bsrA as multidrug ABC transporters [32], that is, they are involved in the transport of compounds or substances. On the other hand, horB is proposed as the transcriptional regulator of horC [24], but evidence to confirm this function is lacking. Besides, the influx of hop bitter acids into the cell is diminished by the contribution of horA and horC [5].
Despite several evidence and studies that point out their functions, the importance of these genes is still discussed and questioned, and still not well understood [33]. For example, in Lactobacillus brevi NBC strain UCCLB556, horA was found. However, this strain was described as unable to grow with hops and the strain UCCLBBS124, which, on the contrary, was reported as growing in the presence of hops, had plasmids with horA and horC, and not hitA [34]. Gene horA was not found in any of the isolates in Lactobacillus rossiae strains isolated from beer, while horC was found in four isolates and hitA in two [35].
There were no statistical differences in the populations comparing each group in the two methodologies, the traditional plate-count method and PMA-qPCR (Table 1), at the end of the production process (day 28). However, the plate-count method applied during the storage period evaluated, showed a lower population for the F19 strain (p < 0.05), which may be evidence of stress. Thus, the difference in the modulation of the expression of these genes may not only be associated with the survival of the strains in the wort with hops, but the different genes expressed may have an influence on the bacteria balance. Further studies are necessary to better understand the role of these genes.
Despite this, as mentioned previously, the expression of bsrA is higher in L431, and horA is present only in F19. The horA gene encodes an ABC transporter [30] and bsrA a multidrug ABC transporter [32]. ATP-Binding Cassette (ABC) transporters are responsible for expelling toxins from cells, and when found in bacteria they can confer antimicrobial resistance [36]. Interestingly, the α-acids found in hops confer antimicrobial properties [5]. Thus, both horA and bsrA seem to perform the same function, since they translate the same class of proteins. Both proteins play an important role in expelling alpha acids from the bacterial cell. The expression of bsrA is practically double in L431 compared to the same expression in F19, and horA expressed in F19. About this, we can think in two hypotheses (Figure 3).
It seems that F19 can not sufficiently express bsrA and, therefore, it expresses horA, which has a protein with the same function, to compensate for this bsrA insufficiency (Figure 3). The stimulus for the expression of horA and bsrA happened simultaneously in F19 and, thus, shows that a higher expression of bsrA, as in the case of 431, is more efficient in eliminating hop bitter acids. This can be corroborated by the fact that the F19 population determined by the agar-plating method is lower than that of 431 (Table 1) on day 2, and the PMA-qPCR counting method is the same between the two strains (Table 1) in the same period, which can evidence that the F19 is more stressed.
3.3. A new Hop Sour Beer Style?
According to the Beer Judge Certification Program (BJCP) (https://www.bjcp.org/), sour beers are characterized by having a freshness and lactic acid flavor, low pH, and the vast majority do not have much hop content. Most of them have an IBU up to 10 or 12. However, Flanders Red Ale and Oud Bruin are the only ones that can reach an IBU of 25. So far, there is no sour registered that has an IBU above 25 (Table 2). In this way, a beer with an IBU equal to 29.65 can be considered a very hop beer, although it does not have the same content as most IPAs do, which are considered the most hop beers. Even among the IPAs, there are those for which an IBU of 25 is acceptable, as in the case of Brut IPA and Hazy IPA. However, it has a value that is within the acceptable range for a German Pilsen and might be regarded as a beer with a pronounced hop flavor (Table 2) (BJCP).
As far as we know, this is the first study and the first evidence that bacteria can survive in beers with high quantity of hops. It is known that the survival of bacteria together with hops depends on their ability to expel α-acids from their cytoplasm, which are involved in membrane transport mechanisms and pumps [5]. Thus, perhaps due to their ability to survive in the gastro-intensive tract, some strains of probiotic bacteria may be the best candidates to survive in beers with more hops.
3.4. Final Considerations
In the production of a sour beer, therefore, little or no hops are used in its production. Some specific styles, as demonstrated earlier, may contain a larger amount of these ingredients. This is mainly due to the difficulty that most microorganisms have in growing together with hops. An alternative would be the production of sour beers with hops added late, that is, during the fermentation process. In this way, the release of α-acids during boiling is avoided [37]. However, this methodology demands an increased maturation time and, therefore, a higher cost in each bottle that is taken to the consumer.
The use of hop-resistant strains, in this case the probiotic strains Lacticaseibacillus paracasei subsp. paracasei F19 and L431, not only can provide functional properties to the beer, but could also accelerate the brewery of sour beers with more hops, and possibly making these beers cheaper to the consumer. These strains are strong candidates to be used in the production of other sour beers, as well as in the development of other beer styles in this category. In addition, the data presented here, as far we know, is the first evidence of bacterial survival in formulations with an IBU greater than 29, and the beer formulations here presented do not have parameters for any of the beer styles found in the BJCP database. As a consequence, they are possibly new styles of sour beer.
It is a consensus that moderate consumption of beer brings health benefits [38]. Beer contains components associated with health benefits, such as melatonin [39], hop xanthohumol (XN) [9] and iso-α-acid [40], vitamin B, minerals, and flavonoids [41]. Hops also have medical properties. Several studies have demonstrated the beneficial effects of molecules present in hops on weight gain, lipid metabolism, glucose homeostasis, insulin sensitivity, and inflammation, acting on different targets [42]. They have also been shown to improve gastrointestinal mucosal integrity and reduce systemic endotoxemia in high-fat diet mouse models [42]. Allied to these benefits that a beer with a high quantity of hops can provide, a beer with probiotics can provide even more benefits in the case of moderate or low beer consumption.
5. Conclusions
This Two formulations of sour beers with both high hop and probiotic populations were studied and we can conclude that the probiotic strains of Lacticaseibacillus paracasei subsp. paracasei F19 and L431 are suitable to produce this high hop content beer, showing good stability. The expression of the genes involved in hops resistance showed a different modulation between the strains. Possibly, a higher amount of bsrA, which was found in L431, is more effective in reducing the effect of hop stress than a combined expression of horA and bsrA. In this way, L. paracasei subsp. paracasei F19 and L431 are both good candidates for developing higher hop sour beers.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (#2019/02583-0).
Data Availability Statement
All relevant data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding author.
Acknowledgments
The author wish to thank Chr. Hansen for providing the cultures, to Dr Gabriela Müller from Higher School of Beer and Malt (Escola Superior de Cerveja e Malte – ESCM, Blumenau, Santa Catarina, Brazil) and Levteck (Florianópolis, Santa Catarina, Brazil), the brewmaster Emmanuel Nascimento Lima de Souza (Natal, Rio Grande do Norte, Brazil).
Conflicts of Interest
The author declares no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Callejo, M.J.; Tesfaye, W.; González, M.C.; Morata, A. Craft beers: Current situation and future trends. IntechOpen 2019. [Google Scholar] [CrossRef]
- Watson, B. State of the industry. 2020. Available online: https://www.brewersassociation.org/wp-content/uploads/2020/04/CBC-Online-2020-State-of-the-Craft-Brewing-Industry.pdf (accessed on 16 October 2023).
- Codifava, N. Craft beer revolution: Appropriability regime of the innovation and business model archetypes adopted by most growing craft breweries. 2018; Dissertation, Politecnico di Milano. [Google Scholar]
- Neves, V. Ramo de cervejarias artesanais cresceu 91% nos últimos três anos. Jornal da USP. 2019. Available online: https://jornal.usp.br/atualidades/ramo-de-cervejarias-artesanais-cresce-91-nos-ultimos-tres-anos (accessed on 16 October 2019).
- Dysvik, A.; La Rosa, S.L.; De Rouck, G.; Rukke, E.O.; Westereng, B.; Wicklund, T. Microbial Dynamics in Traditional and Modern Sour Beer Production. Appl Environ Microbiol. 2020, 86(14), e00566-20. [Google Scholar] [CrossRef]
- Ballan, R.; Battistini, C.; Xavier-Santos, D.; Saad, S.M.I. Interactions of probiotics and prebiotics with the gut microbiota. Prog Mol Biol Transl Sci. 2020, 171, 265–300. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H. J.; Salminen, S.; Calder, P.C.; Sanders, M.E. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Rev Gastroent Hepatol 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Knez Hrnčič, M.; Španinger, E.; Košir, I.; Knez, Ž.; Bren, U. Hop Compounds: Extraction Techniques, Chemical Analyses, Antioxidative, Antimicrobial, and Anticarcinogenic Effects. Nutrients 2019, 11(2), 257. [Google Scholar] [CrossRef]
- Aggarwal, D.; Upadhyay, S.K.; Singh, R.; Tuli, H.S. Recent patents on therapeutic activities of xanthohumol: a prenylated chalconoid from hops (Humulus lupulus L.). Pharm Pat Anal 2021. [Google Scholar] [CrossRef]
- Alcine Chan, M.Z.; Chua, J.Y.; Toh, M.; Liu, S.Q. Survival of probiotic strain Lactobacillus paracasei L26 during co-fermentation with S. cerevisiae for the development of a novel beer beverage. Food Microbiol. 2019, 82, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.C.; Schmidt, G.B.; Alves, L.G.O.; Oliveira, V.S.; Laureano-Melo, R.; Stutz, E.; Martins, J.F.P.; Paula, B.P.; Luchese, R.H.; Guerra, A.F.; Rodriguesa, R. Use of probiotic strains to produce beers by axenic or semi-separated co-culture system. Food Bioprod Process 2020, 124, 408–418. [Google Scholar] [CrossRef]
- Praia, A.B.; Herkenhoff, M.E.; Broedel, O.; Frohme, M.; Saad, S.M.I. Sour Beer with Lacticaseibacillus paracasei subsp. paracasei F19: Feasibility and Influence of Supplementation with Spondias mombin L. Juice and/or By-Product. Foods 2022, 11(24), 4068. [Google Scholar] [CrossRef] [PubMed]
- Herkenhoff, M.E.; Battistini, C.; Praia, A.B.; Rossini, B.C.; Dos Santos, L.D.; Brödel, O.; Frohme, M.; Saad, S.M.I. The combination of omics strategies to evaluate starter and probiotic strains in the Catharina sour Brazilian-style beer. Food Res Int. 2023, 167, 112704. [Google Scholar] [CrossRef] [PubMed]
- ASBC - American Society of Brewing Chemists - ASBC Methods of Analysis - Beer Bitterness. Published in March 1 2015st and updated in March 31st 2022. 2022. Available online: http://methods.asbcnet.org/about.aspx (accessed on 4 October 2022).
- Herkenhoff, M.E.; de Medeiros, I.U.D.; Garutti, L.H.G.; Salgaço, M.K.; Sivieri, K.; Saad, S.M.I. Cashew By-Product as a Functional Substrate for the Development of Probiotic Fermented Milk. Foods 2023, 12(18), 3383. [Google Scholar] [CrossRef]
- Shahrokhi, M.; Nagalli, S. Probiotics. In: StatPearls [Internet], Treasure Island (FL): StatPearls Publishing; 2020 Jan–. PMID: 31985927. [PubMed]
- Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paul, J.; Perdigón, G.D.V. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity. Frontiers in Microbiology. 2018, 9, 736. [Google Scholar] [CrossRef]
- Kawa-Rygielska, J.; Adamenko, K.; Pietrzak, W.; Paszkot, J.; Głowacki, A.; Gasiński, A.; Leszczyński, P. The Potential of Traditional Norwegian KVEIK Yeast for Brewing Novel Beer on the Example of Foreign Extra Stout. Biomolecules 2021, 11(12), 1778. [Google Scholar] [CrossRef] [PubMed]
- Nocker, A.; Mazza, A.; Masson, L.; Camper, A.K.; Brousseau, R. Selective detection of live bacteria combining propidium monoazide sample treatment with microarray technology. J Microbiol Methods 2009, 76((3)), 253–261. [Google Scholar] [CrossRef] [PubMed]
- Padilha, M.; Morales, M.L.V.; Vieira, A.D.S.; Costa, M.G.M.; Saad, S.M.I. A prebiotic mixture improved Lactobacillus acidophilus and Bifidobacterium animalis gastrointestinal in vitro resistance in petit-suisse. Food Funct 2016, 7, 2312–2319. [Google Scholar] [CrossRef] [PubMed]
- Sieuwerts, A.; Håkansson, J. Development of a standardized method for the quantification of Lactobacillus paracasei F19 in stoll samples of various ages. EC Nutrition. 2016, 3, 633–642. [Google Scholar] [CrossRef]
- Byun, R.; Nadkarni, M.A.; Chhour, K.-L.; Martins, F.E.; Jacques, N.A.; Hunter, N. Quantitative analysis of diverse Lactobacillus species present in advanced dental caries. Journal of Clinical Microbiology. 2004, 42(7), 3128–3136. [Google Scholar] [CrossRef]
- Zott, K.; Claisse, O.; Lucas, P.; Coulon, J.; Lonvaud-Funel, A.; Masneuf Pomarede, I. Characterization of the yeast ecosystem in grape must and wine using real-time PCR. Food Microbiol. 2010, 27, 559–567. [Google Scholar] [CrossRef]
- Bergsveinson, J.; Pittet, V.; Ziola, B. RT-qPCR analysis of putative beer-spoilage gene expression during growth of Lactobacillus brevis BSO 464 and Pediococcus claussenii ATCC BAA-344(T) in beer. Appl Microbiol Biotechnol. 2012, 96(2), 461–470. [Google Scholar] [CrossRef]
- Health Canada. Probiotic Claims. 2019. Available online: https://www.inspection.gc.ca/food-label-requirements/labelling/-f-for-industry/former-health-claims/eng/1514559099172/1514559100331?chap=9#s21c9 (accessed on 15 September 2020).
- Dysvik, A.; La Rosa, S.L.; Liland, K.H.; Myhrer, K.S.; Østlie, H.M.; De Rouck, G.; Rukke, E.O.; Westereng, B.; Wicklund, T. Co-fermentation Involving Saccharomyces cerevisiae and Lactobacillus Species Tolerant to Brewing-Related Stress Factors for Controlled and Rapid Production of Sour Beer. Front Microbiol. 2020, 11, 279. [Google Scholar] [CrossRef]
- Bergsveinson, J.; Ziola, B. Investigation of beer spoilage lactic acid bacteria using omic approaches, 2017. p 245–274. In Bokulich NA, Bamforth CW (ed), Brewing microbiology: current research, omics and microbial ecology. Caister Academic Press, Norfolk, UK. [CrossRef]
- Cuevas-González, P.F.; Aguilar-Toalá, J.E.; García, H.S.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Protective Effect of the Intracellular Content from Potential Probiotic Bacteria against Oxidative Damage Induced by Acrylamide in Human Erythrocytes. Probiotics and Antimicrobial Proteins. 2020, 12(4), 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yeh, C.; Jin, Z.; Ding, L.; Liu, B.Y.; Zhang, L.; Dannelly, H.K. Prospective study of probiotic supplementation results in immune stimulation and improvement of upper respiratory infection rate. Synthetic and Systems Biotechnology. 2018, 3(2), 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Margolles, A.; van Veen, H.W.; Konings, W.N. (2001). Hop resistance in the beer spoilage bacterium Lactobacillus brevis is mediated by the ATP-binding cassette multidrug transporter HorA. Journal of Bacteriology 2001, 183, 5371–5375. [Google Scholar] [CrossRef] [PubMed]
- Iijima, K.; Suzuki, K.; Asano, S.; Ogata, T.; Kitagawa, Y. HorC, a hopresistance related protein, presumably functions in homodimer form. Bioscience, Biotechnology, Biochemistry 2009, 73, 1880–1882. [Google Scholar] [CrossRef]
- Haakensen, M.; Vickers, D.M.; Ziola, B. Susceptibility of Pediococcus isolates to antimicrobial compounds in relation to hop-resistance and beer spoilage. BMC Microbiol. 2009, 9, 190. [Google Scholar] [CrossRef]
- Feyereisen, M.; Mahony, J.; O'Sullivan, T.; Boer, V.; van Sinderen, D. (2020). A Plasmid-Encoded Putative Glycosyltransferase Is Involved in Hop Tolerance and Beer Spoilage in Lactobacillus brevis. Applied Environmental Microbiology. 2020, 86(3), e02268–19. [Google Scholar] [CrossRef]
- Feyereisen, M.; Mahony, J.; Kelleher, P.; Roberts, R.J.; O’Sullivan, T.; Geertman, J.-M.A.; van Sinderen, D. Comparative genome analysis of the Lactobacillus brevis species. BMC Genomics. 2019, 20, 416. [Google Scholar] [CrossRef]
- Schneiderbanger, J.; Jacob, F.; Hutzler, M. Genotypic and phenotypic diversity of Lactobacillus rossiae isolated from beer. J Appl Microbiol. 2019, 126(4), 1187–1198. [Google Scholar] [CrossRef]
- Yadav, P.; Ambudkar, S.V.; Rajendra Prasad, N. Emerging nanotechnology-based therapeutics to combat multidrug-resistant cancer. J Nanobiotechnology. 2022, 20(1), 423. [Google Scholar] [CrossRef]
- Tonsmeire, M. American sour beer: innovative techniques for mixed fermentations. Brewers Publications 2014. Maston. [Google Scholar]
- de Gaetano, G.; Costanzo, S.; Di Castelnuovo, A.; Badimon, L.; Bejko, D.; Alkerwi, A.; Chiva-Blanch, G.; Estruch, R.; La Vecchia, C.; Panico, S.; Pounis, G.; Sofi, F.; Stranges, S.; Trevisan, M.; Ursini, F.; Cerletti, C.; Donati, M.B.; Iacoviello, L. Effects of moderate beer consumption on health and disease: A consensus document. Nutr Metab Cardiovasc Dis. 2016; 26(6), 443–67. [Google Scholar] [CrossRef]
- Marhuenda, J.; Villaño, D.; Arcusa, R.; Zafrilla, P. (2021). Melatonin in Wine and Beer: Beneficial Effects. Molecules, 2021; 26, (2), E343. [Google Scholar] [CrossRef]
- Takase, T.; Toyoda, T.; Kobayashi, N.; Inoue, T.; Ishijima, T.; Abe, K.; Kinoshita, H.; Tsuchiya, Y.; Okada, S. Dietary iso-α-acids prevent acetaldehyde-induced liver injury through Nrf2-mediated gene expression. PLoS One. 2021, 16(2), e0246327. [Google Scholar] [CrossRef]
- Sánchez-Muniz, F.J.; Macho-González, A.; Garcimartín, A.; Santos-López, J.A.; Benedí, J.; Bastida, S.; González-Muñoz, M.J. The Nutritional Components of Beer and Its Relationship with Neurodegeneration and Alzheimer's Disease. Nutrients. 2019, 11(7), 1558. [Google Scholar] [CrossRef] [PubMed]
- Ponticelli, M.; Russo, D.; Faraone, I.; Sinisgalli, C.; Labanca, F.; Lela, L.; Milella, L.; The Promising Ability of Humulus lupulus, L. (2021) Iso-α-acids vs. Diabetes, Inflammation, and Metabolic Syndrome: A Systematic Review. Molecules 2021, 26(4), 954. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Layout showing the brewing stages, which includes wort preparation, mashing, fermentation and maturation and carbonation steps.
Figure 1.
Layout showing the brewing stages, which includes wort preparation, mashing, fermentation and maturation and carbonation steps.
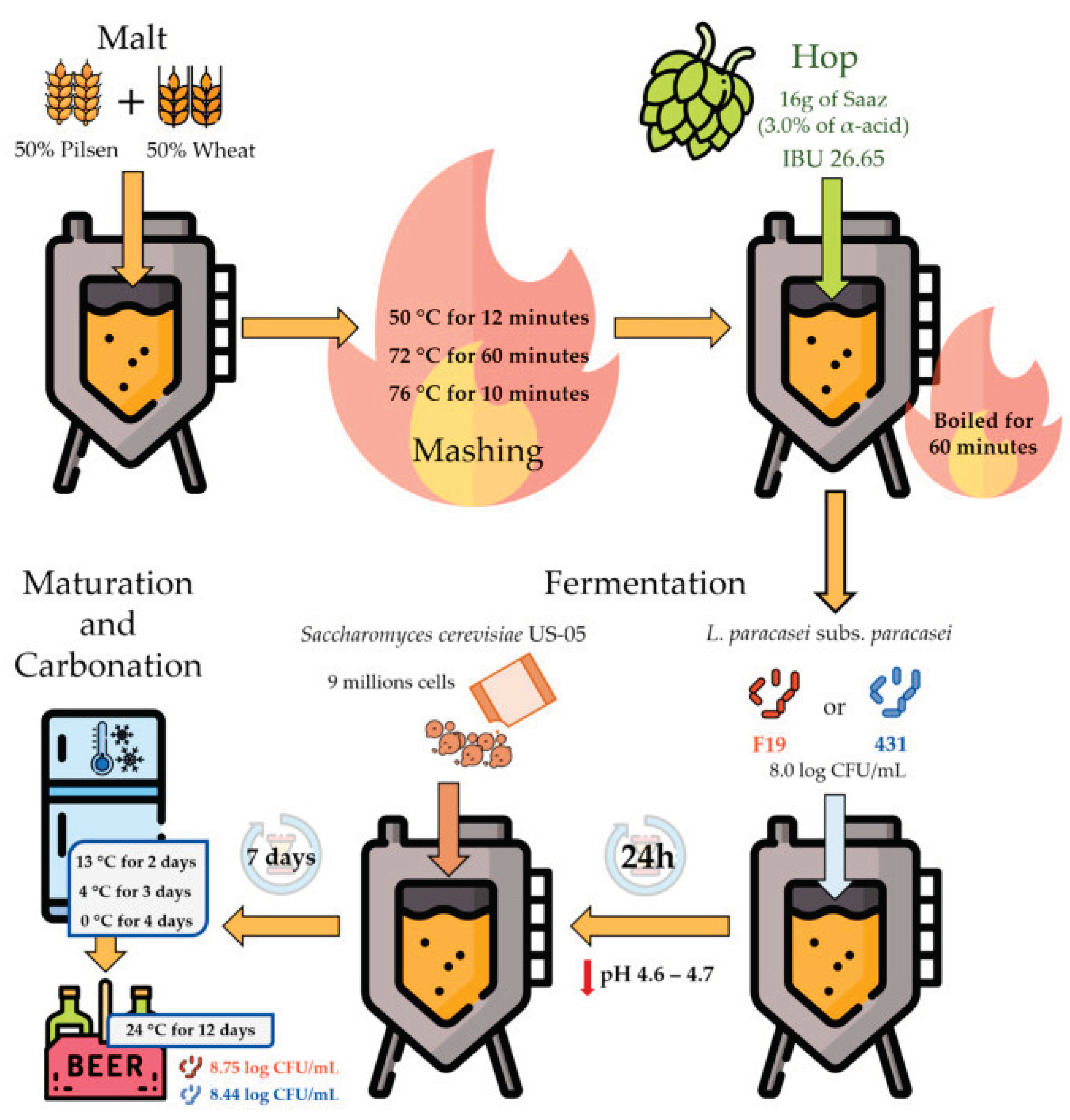
Figure 2.
Relative expression of gene mRNAs related to hop resistance of Lacticaseibacillus paracasei F19 and Lacticaseibacillus paracasei 431. A-C Different superscript between the two formulations (p < 0.05).
Figure 2.
Relative expression of gene mRNAs related to hop resistance of Lacticaseibacillus paracasei F19 and Lacticaseibacillus paracasei 431. A-C Different superscript between the two formulations (p < 0.05).
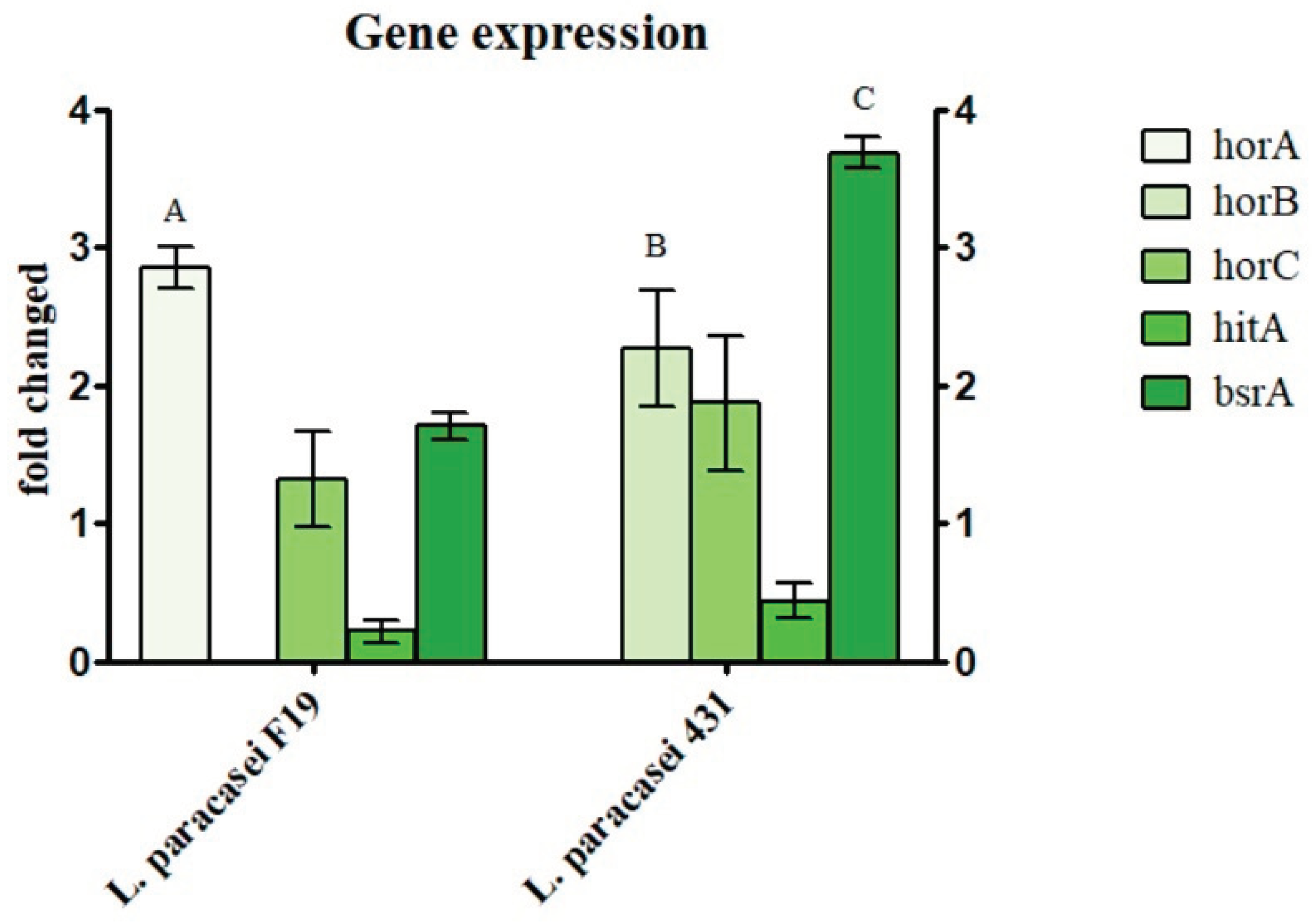
Figure 3.
Graphic presentation of differentiated gene expression between Lacticaseibacillus paracasei F19 and Lacticaseibacillus paracasei 431 strains, showing that increased expression of bsrA in L431 is more efficient to eliminate hop bitter acids from bacterial cytoplasm than expression of horA and bsrA miner expression in F19.
Figure 3.
Graphic presentation of differentiated gene expression between Lacticaseibacillus paracasei F19 and Lacticaseibacillus paracasei 431 strains, showing that increased expression of bsrA in L431 is more efficient to eliminate hop bitter acids from bacterial cytoplasm than expression of horA and bsrA miner expression in F19.
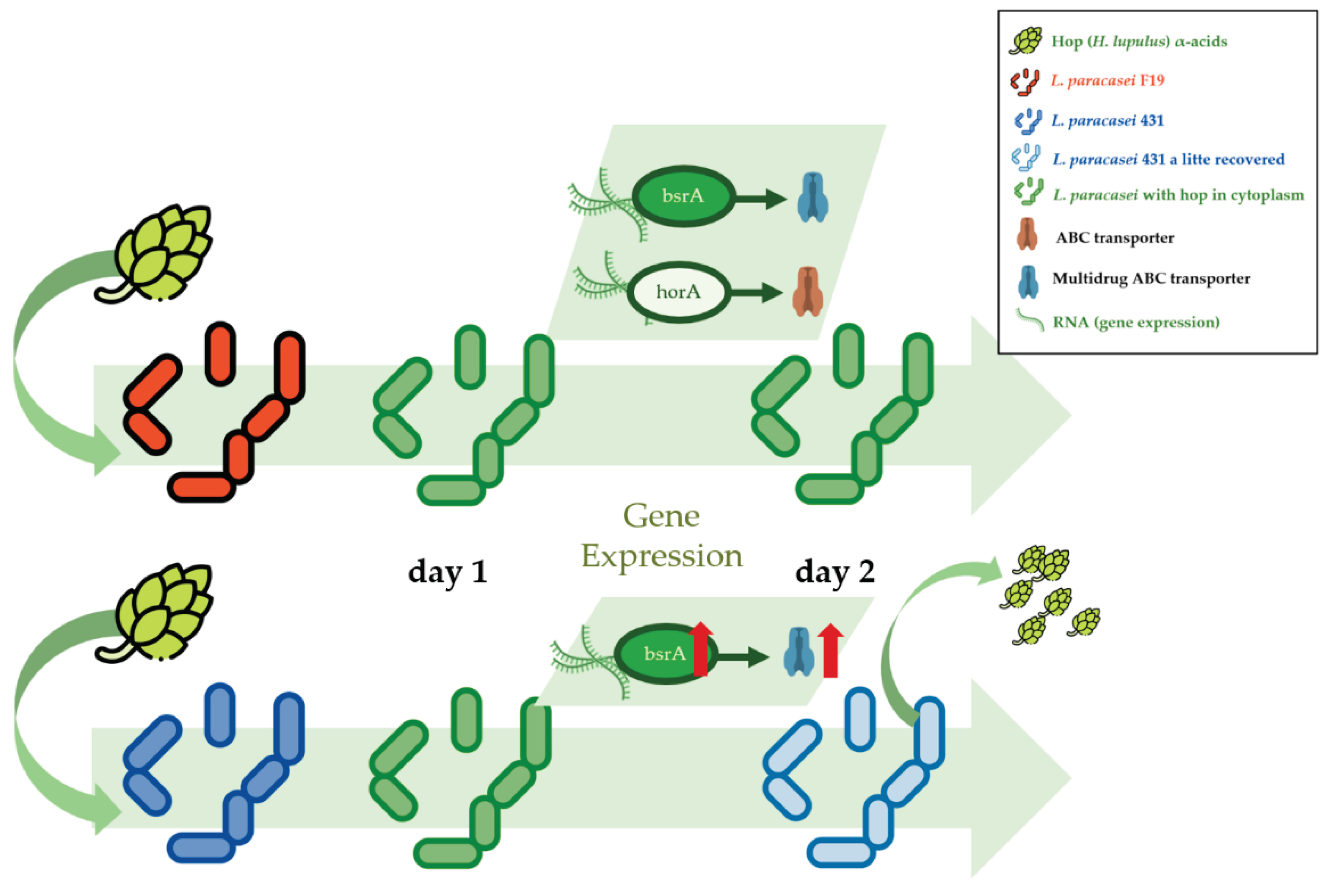

Table 1.
Variations of pH, microbiological count of lactic acid bacteria by MRS pour-plate agar (log CFU/1 mL) and PMA (propidium monoazide) – qPCR (log equivalents CFU/mL), as well as yeast Saccharomyces cerevisiae US-05 populations (also by PMA-qPCR) throughout all production steps and after one month (58 days) of storage.
Table 1.
Variations of pH, microbiological count of lactic acid bacteria by MRS pour-plate agar (log CFU/1 mL) and PMA (propidium monoazide) – qPCR (log equivalents CFU/mL), as well as yeast Saccharomyces cerevisiae US-05 populations (also by PMA-qPCR) throughout all production steps and after one month (58 days) of storage.
| Period (day)\ Formulation | Control | with Lacticaseibacillus paracasei F19 | with Lacticaseibacillus paracasei 431 | ||||||||
| pH | pH | pour-plate | PMA-qPCR | Yeast count | pH | pour-plate | PMA-qPCR | Yeast count | |||
| 0 | 5.9 | 5.9 | 0 | 0 | 0 | 5.9 | 0 | 0 | 0 | ||
| 1 | 5.9 | 4.6 | 5.57 ±0.21 A | 8.15 ±0.08 A,a | 0 | 4.7 | 5.11 ±0.38 A | 8.60 ±0.03 A,a | 0 | ||
| 2 | 4.8 | 4.3 | 3.65 ±0.74 A | 8.91 ±0.09 A,a | 0 | 4.4 | 5.00 ±0.13 B | 8.55 ±0.11 A,a | 0 | ||
| 8 | 4.2 | 3.9 | 3.63 ±1.07 A | 8.93 ±0.07A,a | 7.21 ±0.20 A,b | 3.9 | 3.66 ±0.46 A | 8.24 ±0.34 B,b | 6.47 ±0.23 B,c | ||
| 17 | 4.2 | 3.9 | 5.51 ±0.45 A | 8.90 ±0.15 A,a | 6.22 ±0.24 A,c | 3.9 | 5.73 ±0.08 A | 8.47 ±0.06 A,a | 5.77 ±0.15A,b | ||
| 28 | 4.2 | 3.9 | 6.52 ±0.13 A | 8.80 ±0.13 A,b | 5.87 ±0.34 A,c | 3.9 | 6.46 ±0.13 A | 8.31 ±0.04 B,a | 5.81 ±0.05 A,b | ||
| 58 | 4.2 | 3.9 | 5.39 ±0.26 A | 8.75 ±0.14 A,a | 6.42 ±0.07 A,c | 3.9 | 4.91 ±0.30 A | 8.44 ±0.05 A,a | 5.19 ±0.06 A,a | ||
A- F Different superscript capital letters in the same column denote significant differences (p < 0.05), comparing the formulations with the same matrices, control (without juice), with passion fruit juice and peach juice. a-b Different superscript small letters in the same row denote significant differences for each beverage between the end of the fermentation (day 28) and storage (day 58) periods for a same viability counting procedure (p < 0.05).
Table 2.
Some beer styles recognized by the Beer Judge Certification Program (BJCP), separated by category, and their parameters demonstrated. This table was formulated with data from the BJCP (https://www.bjcp.org/), with the beer formulated in this study, which is not yet recognized.
Table 2.
Some beer styles recognized by the Beer Judge Certification Program (BJCP), separated by category, and their parameters demonstrated. This table was formulated with data from the BJCP (https://www.bjcp.org/), with the beer formulated in this study, which is not yet recognized.
| Category | Beer style | IBU | SRM | OG | FG | ABV (%) |
| Sour | Berliner Weisse | 3.0 - 8.0 | 2.0 - 3.0 | 1.028 - 1.032 | 1.003 - 1.006 | 2.8 - 3.8 |
| Catharina sour | 2.0 - 8.0 | 2.0 - 6.0 | 1.039 - 1.048 | 1.002 - 1.008 | 4.0 - 5.5 | |
| Lambic | 0.0 - 10.0 | 3.0 - 6.0 | 1.040 - 1.054 | 1.001 - 1.010 | 5.0 - 6.5 | |
| Fruit Lambic | 0.0 - 10.0 | 3.0 - 7.0 | 1.040 - 1.060 | 1.000 - 1.010 | 5.0 - 7.0 | |
| Gueuze | 0.0 - 10.0 | 5.0 - 6.0 | 1.040 - 1.054 | 1.000 - 1.006 | 5.0 - 8.0 | |
| Gose | 5.0 - 12.0 | 3.0 - 4.0 | 1.036 - 1.056 | 1.006 - 1.010 | 4.2 - 4.8 | |
| Flanders Red Ale | 10.0 - 25.0 | 10.0 - 17.0 | 1.048 - 1.057 | 1.002 - 1.012 | 4.6 - 6.5 | |
| Oud Bruin | 20.0 - 25.0 | 17.0 - 22.0 | 1.040 - 1.074 | 1.008 - 1.074 | 4.0 - 8.0 | |
| Pilsen | German Pilsen | 25.0 - 45.0 | 2.0 - 5.0 | 1.044 - 1.050 | 1.008 - 1.013 | 4.4 - 5.2 |
| Bohemian Pilsen | 35.0 - 45.0 | 3.5 - 6.0 | 1.044 - 1.056 | 1.013 - 1.017 | 4.2 - 5.4 | |
| Indian Pale Ale (IPA) | Brut IPA | 20.0 - 30.0 | 2.0 - 4.0 | 1.046 - 1.057 | 0.990 - 1.004 | 6.0 - 7.5 |
| Hazy IPA | 25.0 - 60.0 | 3.0 - 7.0 | 1.060 - 1.085 | 1.010 - 1.015 | 6.0 - 9.0 | |
| American IPA | 40.0 - 70.0 | 6.0 - 14.0 | 1.056 - 1.070 | 1.008 - 1.014 | 5.5 - 7.5 | |
| Black IPA | 50.0 - 90.0 | 25.0 - 40.0 | 1.050 - 1.085 | 1.010 - 1.018 | 5.5 - 9.0 | |
| Belgian IPA | 50.0 - 100.0 | 5.0 - 8.0 | 1.058 - 1.080 | 1.008 - 1.016 | 6.2 - 9.5 | |
| Our study | Brazilian sour hop* | 29.65 | 3.9 | 1.045 | 1.008 | 4.0 |
* Name suggested by the authors for the hypothetical style of sour beer developed in this study. Source: https://www.bjcp.org/.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



