
Submitted:
02 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
The over use of pharmaceutical compounds, essentially antibiotics, led to an increase of their concentrations in aquatic ecosystems. In this study, the white-rot fungus Coriolopsis gallica (high-laccase-producing fungus) was investigated for the biodegradation of ampicillin (AMP) under different cultivation conditions. The biotransformation of the antibiotic was confirmed by means of high-performance liquid chromatography (HPLC), and the antibacterial activity was evaluated via the bacterial growth inhibition agar well diffusion method using Escherichia coli as an ampicillin-sensitive test strain. The results showed that AMP (50 mg L-1) was totally removed by C. gallica after 6 days of incubation in a liquid medium. The antibiotic removal rate was maximal with a fungal culture aged 9 days. Such a culture achieved the removal of a higher concentration of up to 500 mg L-1 of ampicillin in 3 days. This higher antibiotic removal rate was concomitant with the maximal laccase production in culture supernatant. In addition, four consecutive doses of 500 mg L-1 of ampicillin were transformed by the same fungal culture in 24 days. After that, the fungus was unable to remove the antibiotic. The measurement of ligninolytic enzyme activity showed that laccase of C. gallica might participate in the biotransformation of AMP.
Keywords:
Antibiotics
; Ampicillin
; β-lactam
; Coriolopsis gallica
; Biotransformation
; Laccase.
1. Introduction
Over the past few years, there has been increasing attention towards the existence of new emerging pollutants such as pesticides, drugs, and endocrine-disrupting chemicals (EDC) in the aquatic environment [1,2,3]. Antibiotics are a group of pharmaceuticals widely used in human medicine [4] and amongst farm animal populations as veterinary medicine and growth promoters [5]. The use of antibiotics has increased and new ones have been developed because of the proliferation of antibiotic-resistant pathogens [6]. It was reported that more than 200,000 tonnes of antibiotics had been consumed annually worldwide [7], with the most common class, β-lactam antibiotics, constituting 50–70% of sales [8]. This increase in the consumption of antibiotics has been associated with an increase in their irrational use, from 28 to 65% [9]. In Tunisia, 61% of consumers obtained antibiotics directly from a pharmacist without a medical prescription. In 2015, it was defined as a low- and middle-income country with the highest consumption rates [10]. Moreover, modern animal production practices are associated with regular use of antimicrobials, with the estimation that between 2010 and 2030, global consumption of antimicrobials will increase by 67% from 63,151 ± 1,560 tonnes to 105,596 ± 3,605 tonnes [11]. In this regard, uncontrolled use of antibiotics can directly affect the health of humans by generating pathogens resistant to antibiotics [12,13], promoting antibiotic resistance genes [14] and indirectly affecting the environment [15,16,17] through effluents from urban wastewater treatment plants (WWTP) containing antibiotics and their residues, since they are not designed to eliminate them [18]. Therefore, an urgent universal effort needs to be made to control the concentration of antibiotics, and multiple antibiotic-resistant bacteria [19]. Ampicillin is semi-synthetic β-lactam belonging to the group of isoxazolyl penicillins (PI), which obtain their antimicrobial properties from the presence of a beta-lactam ring [20]. Thus, their structures give them resistance to degradation via conventional biological and chemical methods [21]. This antibiotic is widely used in human and veterinary medicine for the treatment of infections. After administration, approximately 30% of ampicillin is excreted when taken orally and 75% is excreted after intravenous use [22]. In wastewater, the concentration of β-lactam antibiotics was about 2.1–3.5 μg L-1 in a swine lagoon, which was near the detection limit (2 μg L-1) [23]. They were also detected in natural waters at concentrations of around mg L-1 [24] and in raw wastewater from the Sfax treatment plant at a concentration greater than 75.40 ng L-1 [25]. For these reasons, the treatment for removing the β-lactam antibiotics will be a challenge in the future [26].
Many studies have been carried out to remove antibiotics from aqueous solutions [27]. Processes such as Fenton reaction [28], UV/ZnO degradation [29,30], advanced oxidation [31,32,33] and adsorption [34,35] were designed to degrade pharmaceutical waste in water matrices. However, biological methods are supposed to be the best for antibiotic removal because they represent an eco-friendly process [36]. In fact, most antibiotics tested are known to be biorecalcitrant under aerobic conditions [37], thus escaping intact from conventional wastewater treatment plants. In this light, non-biological methods have been employed to treat antibiotics (and other pharmaceuticals, too), such as advanced oxidation processes, membrane separation, adsorption, coagulation, as well as various combinations of them [31,33,38,39]. White-rot fungi (WRF) have been identified for their ability to degrade aromatic molecules due to their capability of producing extracellular enzymes, essentially laccase (which oxidizes a wide spectrum of organic pollutants) [40]. This oxidizing property suggests its use in the removal of micropollutants (which are usually persistent to biodegradation) and seems to be a very promising approach for improving water effluent quality at WWTPs [41,42].
The aim of this study was to investigate the potential for the WRF C. gallica to remove ampicillin under different operational conditions. Since the transformation of the molecules does not necessarily imply a decrease of its activity, this parameter was also investigated by means of determining the residual antibacterial activity of treated solutions to ensure the efficiency of the treatment. To the best of our knowledge, this is the first study in which C. gallica has been used for the transformation of ampicillin.
2. Results
2.1. Transformation of AMP in Liquid Media
Residual concentration of ampicillin was estimated directly in the supernatant using HPLC–UV analysis (204 nm) after 6 and 12 days of treatment by C. gallica culture. The HPLC chromatograms (Figure 1) showed that untreated AMP (control) was eluted from the column at 4.97 min. However, those corresponding to C. gallica-treated AMP after 6 and 12 days showed the disappearance of the initial peak and the appearance of a new one at 6.78 min (Figure 1b,c). These results indicate that the C. gallica strain was able to transform ampicillin during 6 days of incubation.
2.2. Monitoring of Laccase and Antibacterial Activities at Different Conditions
2.2.1. Effect of AMP Concentration
The effect of the initial antibiotic concentration on the removal of antibacterial activity was studied at AMP concentrations ranging from 25 to 500 mg L-1. The E. coli growth inhibition zone related to Ampicillin treated by the fungus or not (negative control) was measured. The evolution of laccase activity during the treatment was also evaluated (Figure 2). Different concentrations of AMP showed variable effects on the antibacterial activity removal and laccase production. C. gallica was able to remove the antibacterial activity corresponding to a wide range of AMP concentrations ranging from 25 mg L-1 up to 500 mg L-1 after 2, 6 and 9 days of treatment (Figure 2a). In fact, the ABA corresponding to 25 mg L-1 of AMP was removed after 2 days of treatment in the presence of 0.28 U mL-1 of laccase, and the ABAs corresponding to 50 and 100 mg L-1 of AMP were removed after 6 days of treatment in the presence of 0.8 and 2.33 U mL-1 of laccase, respectively. Meanwhile, the ABAs corresponding to 200 and 500 mg L-1 of AMP were removed after 9 days of treatment in the presence of 3.94 and 10 U mL-1 of laccase, respectively (Figure 2b). Hence, the concentration 500 mg L-1 of AMP was retained and used for further experiments.
2.2.2. Effect of the Age of the Culture on AMP Removal
To evaluate the effect of the age of the culture on the removal of the antibacterial activity of AMP, different fungal cultures aged 0, 3, 7, 9 and 12 days were supplemented with 500 mg L-1 of ampicillin and investigated for laccase production and the removal of antibacterial activity (Figure 3). The results showed that cultures of the C. gallica were able to eliminate the antibacterial activity of AMP (500 mg L-1) regardless of their ages. The highest removal rate was obtained by a 9-day-old culture after 4 days of incubation with AMP, whereas a 12-day-old culture was able to transform only 50% of the initial ABA after 7 days of treatment (Figure 3a). Moreover, the addition of AMP re-stimulated laccase production regardless of the age of the culture (Figure 3b).
2.2.3. Reusability of the Same Culture
To study the reusability of C. gallica (4-day-old culture) for more than one cycle of AMP treatment, consecutive additions of AMP were performed after the degradation of the first dose (500 mg L-1). When the antibacterial activity of this dose was removed, a second dose was added, and so on. The experiment was carried out without the addition of any nutrients. Obtained results show that four successive doses of 500 mg L-1 of AMP could be transformed by the same culture of C. gallica after 26 days of cultivation. After that (fourth dose), this transformation became impossible and the production of laccase decreased. The medium became poor in nutrients, and lysis of mycelia was observed. Each time that AMP was added to the culture medium, laccase was induced and the antibiotic was degraded. C. gallica can achieve efficient degradation of AMP four successive times in the same culture (Figure 4).
3. Discussion
The objective of this work was to evaluate the potential of the strain C. gallica for the degradation of a representative of the -lactams already used for the biotransformation of fluoroquinolones [51]. Firstly, the transformation of ampicillin in time course degradation experiments was estimated by comparing their chromatograms during the fungal treatment to ensure that the loss of the antibacterial activity of AMP was due to the degradation/transformation process of the antibiotic molecule. After 6 days of treatment, AMP was completely removed. Compared to previous works, C. gallica is more efficient in terms of the initial antibiotic concentration, the percentage of degradation, and the rate of transformation. Many studies have reported the transformation of beta-lactam antibiotics by ligninolytic fungi and their enzymes. Lucas et al. (2016) reported the elimination of 96% of β-lactams (initial concentration 10 µg L-1) by Trametes versicolor after 15 days of treatment [43]. Copete-Pertuz et al. (2018) also reported the elimination of 100% of the β-lactams oxacillin (16 mg L-1), cloxacillin (17.5 mg L-1) and dicloxacillin (19 mg L-1) by Leptosphaerulina sp. after 6, 7 and 8 days of treatment, respectively, under the action of laccase and versatile peroxidase [45].
After that, the ability of C. gallica to remove the antibacterial activity of AMP under different culture conditions was evaluated. In each condition, laccase activity was also measured. There is scarce in depth- studies available on the correlation between antibiotic biotransformation and laccase production, but recent papers have reported the involvement of extracellular enzymes in antibiotic degradation, with a putative major role for laccases [51]. Here we focused on laccase, as it could be easily repurposed as free or grafted systems to support sustainable processes. Removal rate of AMP was affected by the initial concentration of the antibiotic and showed variable effects on the antibacterial activity removal and laccase production. It could be noticed that the presence of AMP in the culture media induced laccase production. Further increasing the concentrations of AMP led to an increase in laccase activity. However, high initial concentrations of AMP (important ABA) required more time to be removed. These results are in agreement with those reported by Dhawan et al. (2005) where nine different antibiotics affected fungal growth, protein release and laccase production from Cyathus bulleri (5.3 U mL-1) and Pycnoporus cinnabarinus (10.9 U mL-1) to different extents. Interestingly, apramycin sulphate (500 mg L-1) stimulated maximum laccase production (23.3 U mL-1) from P. cinnabarinus. However, ampicillin trihydrate (200 mg L-1) stimulated laccase production from C. bulleri from 5.5 to 10.6 U mL-1 [46]. Praveen and Reddy (2012) also reported the role of nine antibiotics in laccase stimulation of Stereum ostrea (27.48 U mL-1) and Phanerochaete chrysosporium (1.3 U mL-1). In this study, tetracyclin (500 mg L-1) stimulated maximum laccase production (33.4 and 4 U mL-1) and ampicillin (200 mg L-1) increased laccase production (27.48 and 1.742 U mL-1) from S. ostrea and P. chrysosporium, respectively [47].
Different fungal cultures were investigated to evaluate the effect of the age of the culture on the removal of the antibacterial activity of AMP. The addition of AMP (500 mg L-1) in different C. gallica cultures affected the kinetics of the laccase production and those of the ABA removal. In the absence of AMP and under the same conditions of cultivation (M7, 30°C, 150 rpm, + Cu2+), C. gallica produced the maximum laccase quantity on day 9, after which the production of laccase declined [48]. In this study, the addition of AMP re-stimulated laccase production regardless of the age of the culture. This finding could be explained by the fact that fungi might be mimicking AMP similarly to phenolic substrates. In the same case, Sandhu and Arora (1985) observed the induction of laccase production in Polyporus sanguineus in the presence of different phenolic compounds. Furthermore, they proposed the possibility that the white-rot fungi sense the antibiotic to be a phenolic substrate to attack and to detoxify by means of enzymatic transformation [49]. Similarly, Phlebia radiata has been shown to produce lignin-modifying enzymes for detoxification purposes when toxic compounds have been present in its environment [50].
The reusability of a treatment process is a very important factor in industrial applications because its reuse feasibility is a criterion sought by manufacturers for economic purposes. Four successive doses of AMP (500 mg L-1) were transformed by the same culture of C. gallica. Laccase was re-stimulated after each add of AMP in the same culture. It could be concluded that even if laccase is not the key enzyme responsible for AMP degradation, it could be involved in its biotransformation reaction. Yang et al. (2017) investigated ampicillin degradation by immobilised Cerrena laccase. In the absence of a redox mediator, the degradation efficiency was < 40%. The mediator ABTS increased degradation efficiency to 55% [52]. Furthermore, Zhang et al. (2020a) reported effective degradation of ampicillin (100%) by free and immobilized laccase in different waters and proposed two degradation pathways involved in the ampicillin oxidized by laccase [53]. In the first degradation pathway, the process began with oxidation of the sulphur atom of ampicillin to generate a sulphur–oxygen bond in the so-called molecules (TP365 and TP397). In the second pathway, however, the β-lactam ring of ampicillin was directly opened and oxidized to generate TP366. Thus, the loss of antibiotic activity could be the result of the cleavage of the β-lactam ring which happened during the degradation of ampicillin by laccase.
4. Materials and Methods
4.1. Chemicals and Reagents
Ampicillin sodium salt (CAS No. 69-52-3, ≥ 98.0%) and 2,6-dimethoxyphenol (2,6-DMP, 99%) were obtained from Sigma-Aldrich. All other chemicals and solvents used in this study were of an HPLC or reagent grade.
Different concentrations of AMP ranging from 25 to 500 mg L-1 were obtained through appropriate dilution of the stock solution in distilled water. The maximum absorbance (λmax) of AMP was determined by means of UV-visible spectrophotometry (JENWAY 7315 Spectrophotometer). The chemical structure and some characteristics of AMP are shown in Table 1.
4.2. Microorganisms
The fungal strain used in this study was C. gallica CLBE55, a white-rot fungus isolated from a Tunisian forest biotope in the north-west and maintained via sub-culturing every 30 days on 2% malt extract agar slants at pH 5 and 30°C [54].
Escherichia coli (ATCC 25922) was used as a test strain for the measurement of the residual antibacterial activity of treated solutions.
4.3. Experimental Procedures
4.3.1. Follow-Up of AMP Concentration Time-Course in the Culture Medium
All transformation experiments were performed in 500 ml Erlenmeyer flasks containing 150 ml of the M7 medium and inoculated with 1% of homogenised mycelium. The M7 medium contained (per litre): glucose, 10 g; peptone, 5 g; yeast extract, 1 g; ammonium tartrate, 2 g; KH2PO4, 1 g; MgSO4·7H2O, 0.5 g; KCl, 0.5 g; trace element solution, 1 ml. The composition of the trace element solution per litre was as follows: B4O7Na2·10H2O, 0.1 g; CuSO4·5H2O, 0.01 g; FeSO4·7H2O, 0.05 g; MnSO4·7H2O; 0.01; ZnSO4·7H2O, 0.07 g; (NH4)6Mo7O24·4H2O, 0.01 g. The pH of the solution was adjusted to 5.5. Cultures were incubated at 30°C on a rotary shaker (160 rpm). M7 was supplemented with CuSO4·5H2O (150 µM) after 3 days of cultivation as an inducer of laccase [48]. Each experiment was conducted in triplicate and included non-inoculated controls containing 150 ml of the same medium. After 4 days of cultivation, ampicillin was added to the flasks to give the desired concentration from a stock solution in water. Flasks were incubated in the dark, to exclude the influence of light on ampicillin stability, on an orbital checker in the same conditions as mentioned. In time course experiments, 1.5ml samples were periodically withdrawn, filtered and used for assessing the laccase and antibacterial activity. To follow the residual AMP during cultivation, samples were kept at -20°C until HPLC analysis.
4.3.2. In Vitro Analysis of Residual AMP
Effect of the concentration of the antibiotic
To study the ability of C. gallica to remove the antibacterial activity of high concentrations of antibiotics, different doses of ampicillin were added to the culture medium at a final concentration ranging between 25 and 500 mg L-1 on the fourth day of cultivation. Samples were withdrawn periodically, centrifuged and used to measure residual antibacterial and laccase activity.
Effect of the age of the mycelia
To investigate the influence of the age of mycelia on the removal of the antibacterial activity of ampicillin, the antibiotic solution was injected in cultures at different ages of fungi growing and of laccase production levels. The final concentration of ampicillin in different cultures was 500 mg L-1, and the ages of tested cultures were 0, 3, 7, 9 and 12 days of cultivation.
Effect of the consecutive addition of ampicillin
The following experiment studied the potential of a C. gallica culture for consecutive use in the treatment of the antibacterial activity of ampicillin. In this study, the antibiotic was re-injected in the same culture when the antibacterial activity of the previous concentration decreased to an undetectable level of antibacterial activity. The concentration of the antibiotic was the same as the first addition, which was 500 mg L-1.
4.4. Analytical Procedures
4.4.1. HPLC analysis of Ampicillin in C. gallica Culture Filtrate
The concentration of AMP in the tested culture was measured by means of HPLC UV (Agilent 1100 Series) equipped with a micro-vacuum degasser (Agilent 1100 Series), quaternary pump, diode array, and mass detector (Agilent Technologies 61120 Quadrupole LC/MS) at a wavelength of 204 nm. The separation was performed on a ZORBAX SB-C18 (150 mm × 4.6 mm, 5 µm) column. The mobile phase was a mixture of A (H2O + 0.1% formic acid) and B (acetonitrile + 0.1% formic acid) at a flow rate of 1 ml min-1 (initial, 10% B; 15 min, 90% B; 25 min, 90% B; 26 min, 10% B; 36 min 10% B). The column temperature was set at 35°C, and 10 µl of each sample was injected.
4.4.2. Antibacterial Activity Assay
The antibacterial activity (ABA) of ampicillin before and after treatment was also evaluated via the agar well diffusion method [55]. E. coli cells were cultured overnight at 37°C with shaking (150 rpm) in a lysogeny broth (LB) medium. Petri dishes containing an LB agar medium were inoculated aseptically with a suspension of 106 cells per mL from the young culture. After drying, agar was perforated with the upper part of a Pasteur pipette. The cavities thus formed were filled with the samples taken at different times of treatment (50 µl per well). The Petri dishes were incubated at 37°C for 24 hours. Growth inhibition was calculated by measuring the diameter of growth inhibition against the control as follows:
where D0 and Dt are the diameters of the growth inhibition zone (mm) corresponding to AMP injected on day 4 in the culture and the residual AMP at culture time t, respectively.
4.4.3. Laccase Activity Assay
Laccase activity was assayed by monitoring the oxidation of 10 mM of 2,6-dimethoxyphenol (DMP) (469 nm, ε469 = 27,500 M-1cm-1) in a reaction mixture solution containing 100 mM of citrate buffer at pH 5. One unit of enzyme activity was defined as the amount of enzyme oxidizing 1 µM of substrate min-1 [56].
5. Conclusions
This study investigated the biotransformation and detoxification of ampicillin the by C. gallica under different operational conditions. The selected fungus was able to transform AMP in a liquid medium after 6 days of treatment. Based on activities assays, laccase of C. gallica could be involved in enzymatic degradation of ampicillin and contribute, among other mechanisms, to the removal of antibacterial activity. The loss of antibacterial activity could be attributed to the cleavage of the β-lactam ring under the action of laccase. Further experiments should be performed to investigate enzymes potentially involved in ampicillin degradation, such as proteomic analysis, and to identify transformation products generated during the treatment process by C. gallica.
The good performance of C. gallica in the effective removal and detoxification of ampicillin makes it a promising candidate for environmental recovery as well as further prospects for eco-friendly biological treatment processes to remove antibiotics from wastewater.
Author Contributions
Conceptualization, B.G. and T.M.; methodology, B.G.; validation, A.H.A., I.B.A. and H.Z.M.; investigation, B.G.; resources, T.M. and H.Z.M.; data curation, I.L.; writing—original draft preparation, B.G.; writing—review and editing, B.G., I.B.A., A.H.A. and A.A.A.; visualization, B.G. and I.L..; supervision, T.M.; project administration, T.M.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors are grateful to the Tunisian Ministry of Higher Education and Scientific Research for providing part of the financial support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Imai, S.; Shiraishi, A.; Gamo, K.; Watanabe, I.; Okuhata, H.; Miyasaka, H.; Ikeda, K.; Bamba, T.; Hirata, K. Removal of Phenolic Endocrine Disruptors by Portulaca Oleracea. J. Biosci. Bioeng. 2007, 103, 420–426. [Google Scholar] [CrossRef]
- Tan, B.L.L.; Hawker, D.W.; Müller, J.F.; Tremblay, L.A.; Chapman, H.F. Stir Bar Sorptive Extraction and Trace Analysis of Selected Endocrine Disruptors in Water, Biosolids and Sludge Samples by Thermal Desorption with Gas Chromatography-Mass Spectrometry. Water Res. 2008, 42, 404–412. [Google Scholar] [CrossRef]
- García, J.; García-Galán, M.J.; Day, J.W.; Boopathy, R.; White, J.R.; Wallace, S.; Hunter, R.G. A Review of Emerging Organic Contaminants (EOCs), Antibiotic Resistant Bacteria (ARB), and Antibiotic Resistance Genes (ARGs) in the Environment: Increasing Removal with Wetlands and Reducing Environmental Impacts. Bioresour. Technol. 2020, 307, 123228. [Google Scholar] [CrossRef]
- Schwartz, T.; Kohnen, W.; Jansen, B.; Obst, U. Detection of Antibiotic-Resistant Bacteria and Their Resistance Genes in Wastewater, Surface Water, and Drinking Water Biofilms. FEMS Microbiol. Ecol. 2003, 43, 325–335. [Google Scholar] [CrossRef]
- Mai, Z.; Xiong, X.; Hu, H.; Jia, J.; Wu, C.; Wang, G. Occurrence, Distribution, and Ecological Risks of Antibiotics in Honghu Lake and Surrounding Aquaculture Ponds, China. Environ. Sci. Pollut. Res. 2023, 30, 50732–50742. [Google Scholar] [CrossRef]
- Zinner, S.H. Overview of Antibiotic Use and Resistance: Setting the Stage for Tigecycline. Clin. Infect. Dis. 2005, 41, S289–S292. [Google Scholar] [CrossRef]
- Sun, K.; Huang, Q.; Li, S. Transformation and Toxicity Evaluation of Tetracycline in Humic Acid Solution by Laccase Coupled with 1-Hydroxybenzotriazole. J. Hazard. Mater. 2017, 331, 182–188. [Google Scholar] [CrossRef]
- Kümmerer, K. Antibiotics in the Aquatic Environment--a Review--Part I. Chemosphere 2009, 75, 417–434. [Google Scholar] [CrossRef]
- Erbay, A.; Bodur, H.; Akıncı, E.; Çolpan, A. Evaluation of Antibiotic Use in Intensive Care Units of a Tertiary Care Hospital in Turkey. J. Hosp. Infect. 2005, 59, 53–61. [Google Scholar] [CrossRef]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global Increase and Geographic Convergence in Antibiotic Consumption between 2000 and 2015. Proc. Natl. Acad. Sci. 2018, 115, E3463–E3470. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Carlet, J.; Rambaud, C.; Pulcini, C. Alliance contre les bactéries multirésistantes: sauvons les antibiotiques ! Ann. Fr. Anesth. Réanimation 2012, 31, 704–708. [Google Scholar] [CrossRef]
- Yuan, W.; Zhang, Y.; Riaz, L.; Yang, Q.; Du, B.; Wang, R. Multiple Antibiotic Resistance and DNA Methylation in Enterobacteriaceae Isolates from Different Environments. J. Hazard. Mater. 2021, 402, 123822. [Google Scholar] [CrossRef]
- Zhang, G.; Lu, S.; Wang, Y.; Liu, X.; Liu, Y.; Xu, J.; Zhang, T.; Wang, Z.; Yang, Y. Occurrence of Antibiotics and Antibiotic Resistance Genes and Their Correlations in Lower Yangtze River, China. Environ. Pollut. 2020, 257, 113365. [Google Scholar] [CrossRef]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.-N.; et al. Tackling Antibiotic Resistance: The Environmental Framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Dong, H.; Chen, Y.; Wang, J.; Zhang, Y.; Zhang, P.; Li, X.; Zou, J.; Zhou, A. Interactions of Microplastics and Antibiotic Resistance Genes and Their Effects on the Aquaculture Environments. J. Hazard. Mater. 2021, 403, 123961. [Google Scholar] [CrossRef]
- Feng, Y.; Hu, J.; Chen, Y.; Xu, J.; Yang, B.; Jiang, J. Ecological Response to Antibiotics Re-Entering the Aquaculture Environment with Possible Long-Term Antibiotics Selection Based on Enzyme Activity in Sediment. Environ. Sci. Pollut. Res. 2022, 29, 19033–19044. [Google Scholar] [CrossRef]
- Rodriguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sànchez-Melsió, A.; Borrego, C.M.; Barceló, D.; Balcázar, J.L. Occurrence of Antibiotics and Antibiotic Resistance Genes in Hospital and Urban Wastewaters and Their Impact on the Receiving River. Water Res. 2015, 69, 234–242. [Google Scholar] [CrossRef]
- Hossain, A.; Al Mamun, M.H.; Nagano, I.; Kitazawa, D.; Masunaga, S.; Matsuda, H. Antibiotics, Antibiotic-Resistant Bacteria, and Resistance Genes in Aquaculture: Risks, Current Concern, and Future Thinking. Environ. Sci. Pollut. Res. 2022, 29. [Google Scholar] [CrossRef]
- Kong, K.-F.; Schneper, L.; Mathee, K. Beta-Lactam Antibiotics: From Antibiosis to Resistance and Bacteriology. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2010, 118, 1–36. [Google Scholar] [CrossRef]
- Fernández-Fernández, M.; Sanromán, M.Á.; Moldes, D. Recent Developments and Applications of Immobilized Laccase. Biotechnol. Adv. 2013, 31, 1808–1825. [Google Scholar] [CrossRef]
- Foulds, G. Pharmacokinetics of Sulbactam/Ampicillin in Humans: A Review. Rev. Infect. Dis. 1986, 8, S503–S511. [Google Scholar] [CrossRef]
- Campagnolo, E.R.; Johnson, K.R.; Karpati, A.; Rubin, C.S.; Kolpin, D.W.; Meyer, M.T.; Esteban, J.E.; Currier, R.W.; Smith, K.; Thu, K.M.; et al. Antimicrobial Residues in Animal Waste and Water Resources Proximal to Large-Scale Swine and Poultry Feeding Operations. Sci. Total Environ. 2002, 299, 89–95. [Google Scholar] [CrossRef]
- Serna-Galvis, E.A.; Jojoa-Sierra, S.D.; Berrio-Perlaza, K.E.; Ferraro, F.; Torres-Palma, R.A. Structure-Reactivity Relationship in the Degradation of Three Representative Fluoroquinolone Antibiotics in Water by Electrogenerated Active Chlorine. Chem. Eng. J. 2017, 315, 552–561. [Google Scholar] [CrossRef]
- Harrabi, M.; Varela Della Giustina, S.; Aloulou, F.; Rodriguez-Mozaz, S.; Barceló, D.; Elleuch, B. Analysis of Multiclass Antibiotic Residues in Urban Wastewater in Tunisia. Environ. Nanotechnol. Monit. Manag. 2018, 10, 163–170. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Giri, B.S.; Shukla, P.; Gupta, P. Recent Advancement in Remediation of Synthetic Organic Antibiotics from Environmental Matrices: Challenges and Perspective. Bioresour. Technol. 2021, 319, 124161. [Google Scholar] [CrossRef]
- Alegbeleye, O.; Daramola, O.B.; Adetunji, A.T.; Ore, O.T.; Ayantunji, Y.J.; Omole, R.K.; Ajagbe, D.; Adekoya, S.O. Efficient Removal of Antibiotics from Water Resources Is a Public Health Priority: A Critical Assessment of the Efficacy of Some Remediation Strategies for Antibiotics in Water. Environ. Sci. Pollut. Res. 2022, 29, 56948–57020. [Google Scholar] [CrossRef]
- Ioannou-Ttofa, L.; Raj, S.; Prakash, H.; Fatta-Kassinos, D. Solar Photo-Fenton Oxidation for the Removal of Ampicillin, Total Cultivable and Resistant E. Coli and Ecotoxicity from Secondary-Treated Wastewater Effluents. Chem. Eng. J. 2019, 355, 91–102. [Google Scholar] [CrossRef]
- Al Abri, R.; Al Marzouqi, F.; Kuvarega, A.T.; Meetani, M.A.; Al Kindy, S.M.Z.; Karthikeyan, S.; Kim, Y.; Selvaraj, R. Nanostructured Cerium-Doped ZnO for Photocatalytic Degradation of Pharmaceuticals in Aqueous Solution. J. Photochem. Photobiol. Chem. 2019, 384, 112065. [Google Scholar] [CrossRef]
- Mu, X.; Huang, Z.; Ohore, O.E.; Yang, J.; Peng, K.; Li, S.; Li, X. Impact of Antibiotics on Microbial Community in Aquatic Environment and Biodegradation Mechanism: A Review and Bibliometric Analysis. Environ. Sci. Pollut. Res. 2023, 30, 66431–66444. [Google Scholar] [CrossRef]
- Yabalak, E. An Approach to Apply Eco-Friendly Subcritical Water Oxidation Method in the Mineralization of the Antibiotic Ampicillin. J. Environ. Chem. Eng. 2018, 6, 7132–7137. [Google Scholar] [CrossRef]
- Montoya-Rodríguez, D.M.; Serna-Galvis, E.A.; Ferraro, F.; Torres-Palma, R.A. Degradation of the Emerging Concern Pollutant Ampicillin in Aqueous Media by Sonochemical Advanced Oxidation Processes - Parameters Effect, Removal of Antimicrobial Activity and Pollutant Treatment in Hydrolyzed Urine. J. Environ. Manage. 2020, 261, 110224. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, B.-T.; Teng, Y.; Zhao, J. Activated Carbon Supported Nanoscale Zero Valent Iron for Cooperative Adsorption and Persulfate-Driven Oxidation of Ampicillin. Environ. Technol. Innov. 2020, 19, 100956. [Google Scholar] [CrossRef]
- Eniola, J.O.; Kumar, R.; Barakat, M.A. Adsorptive Removal of Antibiotics from Water over Natural and Modified Adsorbents. Environ. Sci. Pollut. Res. 2019, 26, 34775–34788. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Q.; Zhu, Y.; Zhang, M.; Zhu, Y.; Farooq, U.; Lu, T.; Qi, Z.; Chen, W. Adsorption of Fluoroquinolone Antibiotics onto Ferrihydrite under Different Anionic Surfactants and Solution PH. Environ. Sci. Pollut. Res. 2023, 30, 78229–78242. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Adeel, M.; Rasheed, T.; Zhao, Y.; Iqbal, H.M.N. Emerging Contaminants of High Concern and Their Enzyme-Assisted Biodegradation – A Review. Environ. Int. 2019, 124, 336–353. [Google Scholar] [CrossRef]
- Kümmerer, K. The Presence of Pharmaceuticals in the Environment Due to Human Use – Present Knowledge and Future Challenges. J. Environ. Manage. 2009, 90, 2354–2366. [Google Scholar] [CrossRef]
- Homem, V.; Santos, L. Degradation and Removal Methods of Antibiotics from Aqueous Matrices – A Review. J. Environ. Manage. 2011, 92, 2304–2347. [Google Scholar] [CrossRef]
- Pirsaheb, M.; Mohamadisorkali, H.; Hossaini, H.; Hossini, H.; Makhdoumi, P. The Hybrid System Successfully to Consisting of Activated Sludge and Biofilter Process from Hospital Wastewater: Ecotoxicological Study. J. Environ. Manage. 2020, 276, 111098. [Google Scholar] [CrossRef]
- Morsi, R.; Bilal, M.; Iqbal, H.M.N.; Ashraf, S.S. Laccases and Peroxidases: The Smart, Greener and Futuristic Biocatalytic Tools to Mitigate Recalcitrant Emerging Pollutants. Sci. Total Environ. 2020, 714, 136572. [Google Scholar] [CrossRef]
- Spina, F.; Gea, M.; Bicchi, C.; Cordero, C.; Schilirò, T.; Varese, G.C. Ecofriendly Laccases Treatment to Challenge Micropollutants Issue in Municipal Wastewaters. Environ. Pollut. 2020, 257, 113579. [Google Scholar] [CrossRef]
- Russell, J.N.; Yost, C.K. Alternative, Environmentally Conscious Approaches for Removing Antibiotics from Wastewater Treatment Systems. Chemosphere 2021, 263, 128177. [Google Scholar] [CrossRef]
- Lucas, D.; Badia-Fabregat, M.; Vicent, T.; Caminal, G.; Rodríguez-Mozaz, S.; Balcázar, J.L.; Barceló, D. Fungal Treatment for the Removal of Antibiotics and Antibiotic Resistance Genes in Veterinary Hospital Wastewater. Chemosphere 2016, 152, 301–308. [Google Scholar] [CrossRef]
- Copete-Pertuz, L.S.; Plácido, J.; Serna-Galvis, E.A.; Torres-Palma, R.A.; Mora, A. Elimination of Isoxazolyl-Penicillins Antibiotics in Waters by the Ligninolytic Native Colombian Strain Leptosphaerulina Sp. Considerations on Biodegradation Process and Antimicrobial Activity Removal. Sci. Total Environ. 2018, 630, 1195–1204. [Google Scholar] [CrossRef]
- Copete-Pertuz, L.S.; Plácido, J.; Serna-Galvis, E.A.; Torres-Palma, R.A.; Mora, A. Elimination of Isoxazolyl-Penicillins Antibiotics in Waters by the Ligninolytic Native Colombian Strain Leptosphaerulina Sp. Considerations on Biodegradation Process and Antimicrobial Activity Removal. Sci. Total Environ. 2018, 630, 1195–1204. [Google Scholar] [CrossRef]
- Dhawan, S.; Lal, R.; Hanspal, M.; Kuhad, R.C. Effect of Antibiotics on Growth and Laccase Production from Cyathus Bulleri and Pycnoporus Cinnabarinus. Bioresour. Technol. 2005, 96, 1415–1418. [Google Scholar] [CrossRef]
- Praveen, K.; Reddy, B.R. Effect of antibiotics on ligninolytic enzymes production from stereum ostrea and phanerochaete chrysosporium under submerged fermentation. 2012. [Google Scholar]
- Zouari-Mechichi, H.; Mechichi, T.; Dhouib, A.; Sayadi, S.; Martínez, A.T.; Martínez, M.J. Laccase Purification and Characterization from Trametes Trogii Isolated in Tunisia: Decolorization of Textile Dyes by the Purified Enzyme. Enzyme Microb. Technol. 2006, 39, 141–148. [Google Scholar] [CrossRef]
- Sandhu, D.K.; Arora, D.S. Laccase Production ByPolyporus Sanguineus under Different Nutritional and Environmental Conditions. Experientia 1985, 41, 355–356. [Google Scholar] [CrossRef]
- Rogalski, J.; Lundell, T.K.; Leonowicz, A.; Hatakka, A.I. Influence of Aromatic Compounds and Lignin on Production of Ligninolytic Enzymes by Phlebia Radiata. Phytochemistry 1991, 30, 2869–2872. [Google Scholar] [CrossRef]
- Ben Ayed, A.; Akrout, I.; Albert, Q.; Greff, S.; Simmler, C.; Armengaud, J.; Kielbasa, M.; Turbé-Doan, A.; Chaduli, D.; Navarro, D.; et al. Biotransformation of the Fluoroquinolone, Levofloxacin, by the White-Rot Fungus Coriolopsis Gallica. J. Fungi 2022, 8, 965. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lin, Y.; Yang, X.; Ng, T.B.; Ye, X.; Lin, J. Degradation of Tetracycline by Immobilized Laccase and the Proposed Transformation Pathway. J. Hazard. Mater. 2017, 322, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; You, S.; Zhang, J.; Qi, W.; Su, R.; He, Z. An Effective In-Situ Method for Laccase Immobilization: Excellent Activity, Effective Antibiotic Removal Rate and Low Potential Ecological Risk for Degradation Products. Bioresour. Technol. 2020, 308, 123271. [Google Scholar] [CrossRef] [PubMed]
- Daâssi, D.; Zouari-Mechichi, H.; Belbahri, L.; Barriuso, J.; Martínez, M.J.; Nasri, M.; Mechichi, T. Phylogenetic and Metabolic Diversity of Tunisian Forest Wood-Degrading Fungi: A Wealth of Novelties and Opportunities for Biotechnology. 3 Biotech 2016, 6, 46. [Google Scholar] [CrossRef]
- Meenupriya, J.; Thangaraj, M. Isolation and Molecular Characterization of Bioactive Secondary Metabolites from Callyspongia Spp. Associated Fungi. Asian Pac. J. Trop. Med. 2010, 3, 738–740. [Google Scholar] [CrossRef]
- Yaropolov, A.I.; Skorobogat’ko, O.V.; Vartanov, S.S.; Varfolomeyev, S.D. Laccase. Appl. Biochem. Biotechnol. 1994, 49, 257–280. [Google Scholar] [CrossRef]
Figure 1.
HPLC chromatograms of (a) the control (AMP at 50 mg L-1 in M7 medium) and treated AMP after (b) 6 and (c) 12 days.
Figure 1.
HPLC chromatograms of (a) the control (AMP at 50 mg L-1 in M7 medium) and treated AMP after (b) 6 and (c) 12 days.
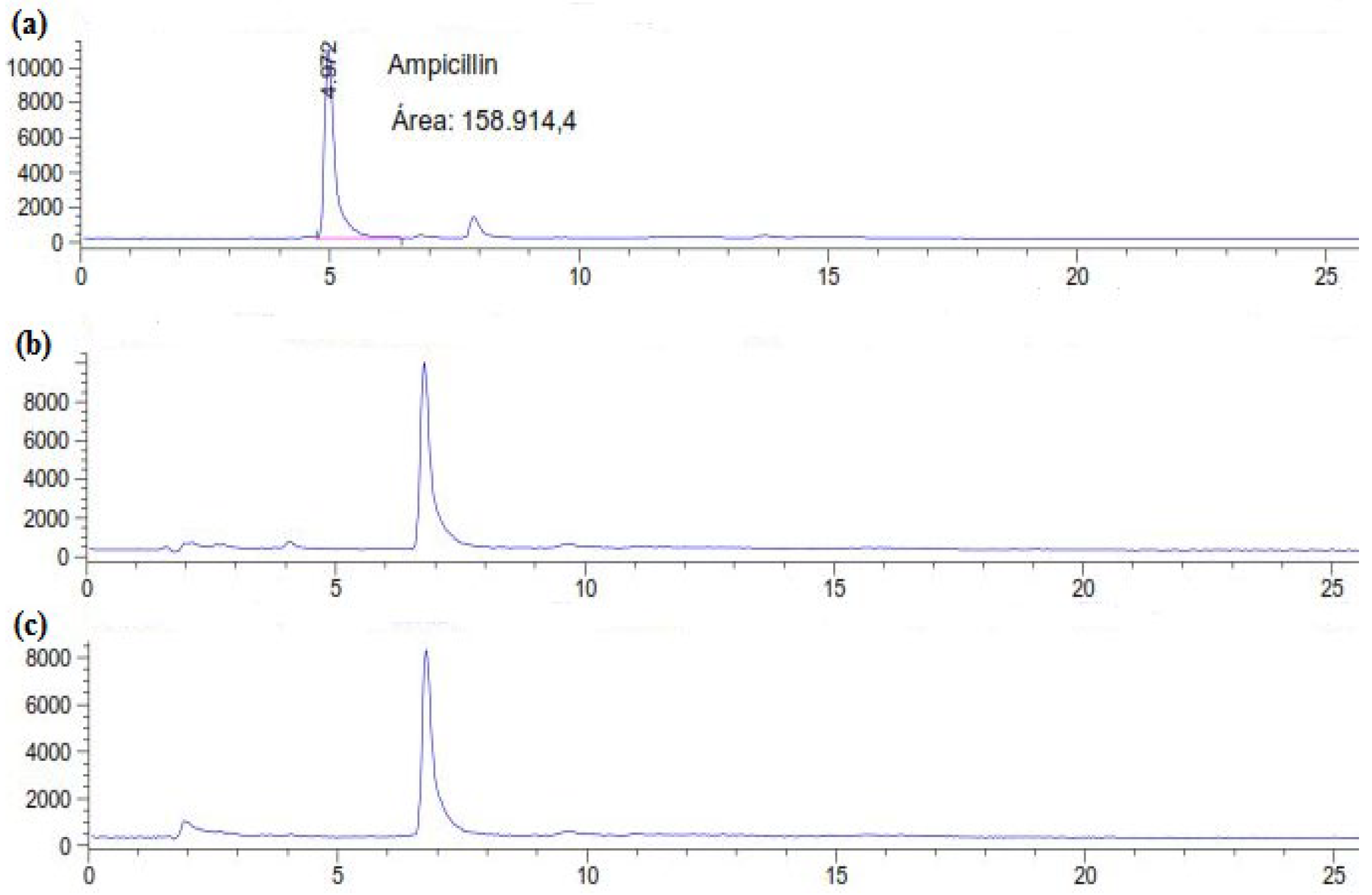
Figure 2.
Effect of initial antibiotic concentration on (a) the antibacterial activity removal and (b) laccase activity evolution (0.15 mM Cu2+, 30°C, 150 rpm). “♦”: 25 mg L-1; “■”: 50 mg L-1; “▲”: 100 mg L-1; “X”: 200 mg L-1; “*”: 500 mg L-1. Each datapoint (mean ± standard deviation) is the result of triplicate experiments.
Figure 2.
Effect of initial antibiotic concentration on (a) the antibacterial activity removal and (b) laccase activity evolution (0.15 mM Cu2+, 30°C, 150 rpm). “♦”: 25 mg L-1; “■”: 50 mg L-1; “▲”: 100 mg L-1; “X”: 200 mg L-1; “*”: 500 mg L-1. Each datapoint (mean ± standard deviation) is the result of triplicate experiments.
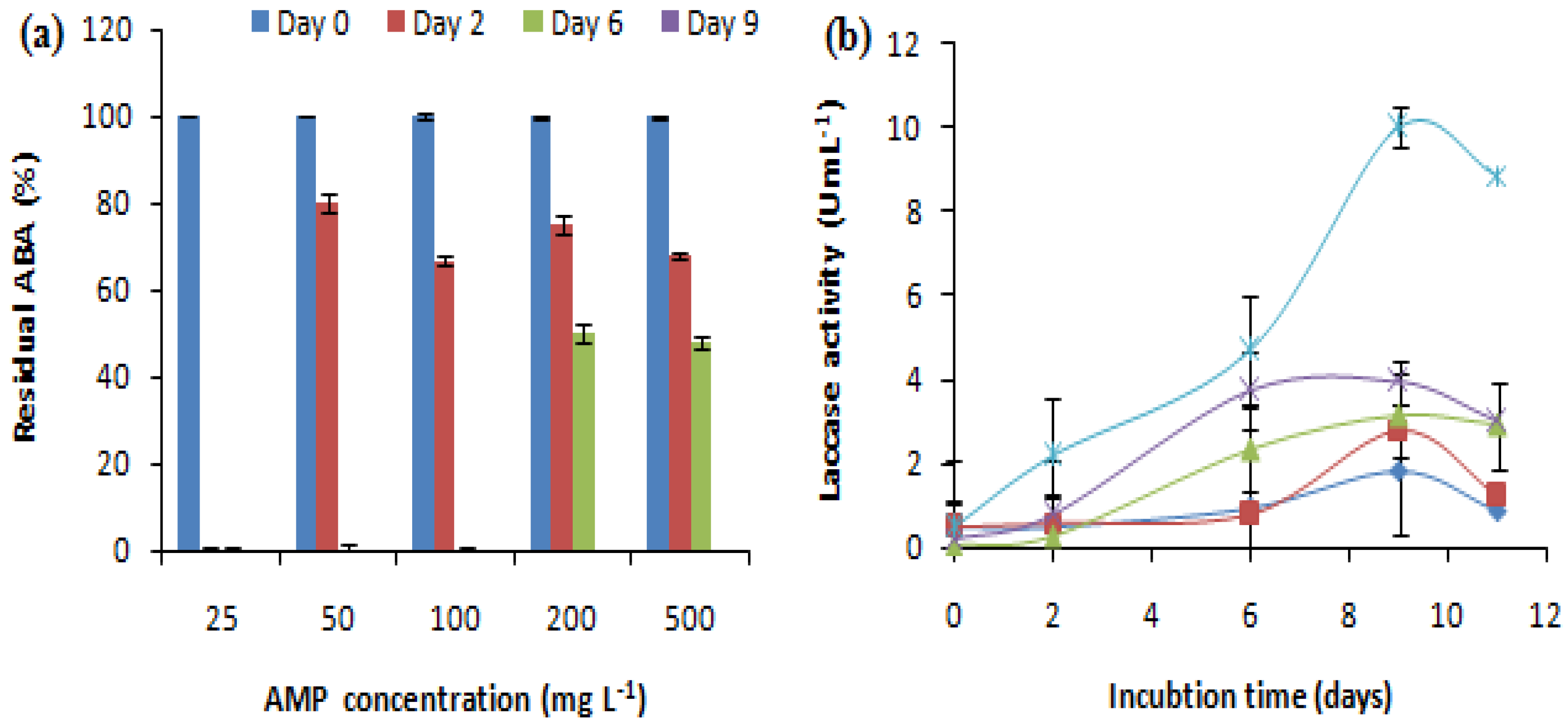
Figure 3.
Effect of the age of the fungal culture on (a) the antibacterial activity removal and (b) the laccase activity evolution. “♦”: 0 days; “■”: 3 days; “▲”: 7 days; “X”: 9 days; “*” 12 days. Each datapoint (mean ± standard deviation) is the result of triplicate experiments.
Figure 3.
Effect of the age of the fungal culture on (a) the antibacterial activity removal and (b) the laccase activity evolution. “♦”: 0 days; “■”: 3 days; “▲”: 7 days; “X”: 9 days; “*” 12 days. Each datapoint (mean ± standard deviation) is the result of triplicate experiments.
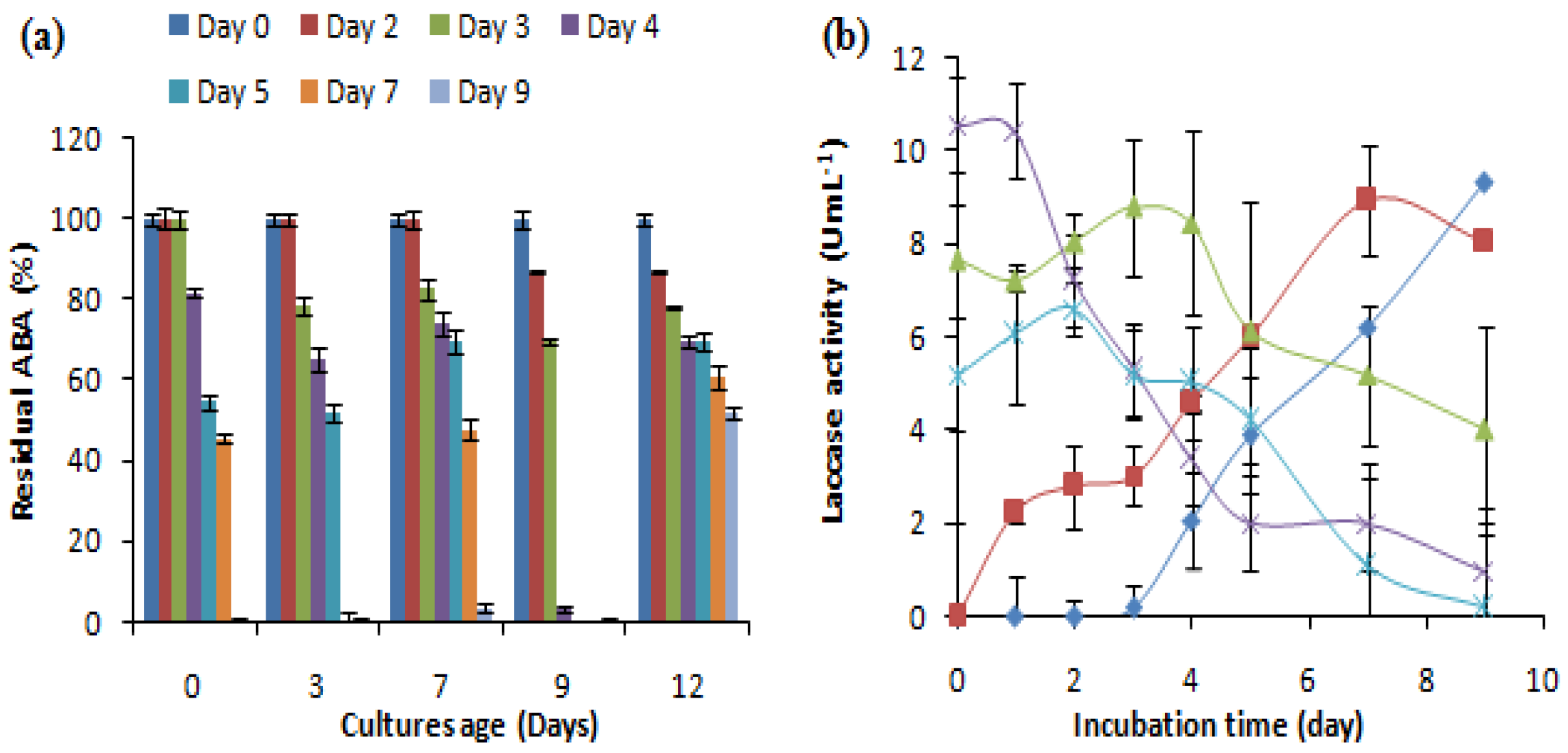
Figure 4.
Effect of the cumulative addition of AMP
(500 mg L-1) on the antibacterial activity removal “♦” and the laccase activity evolution “■”. : AMP
injection in the culture medium. Each datapoint (mean ± standard deviation) is
the result of triplicate experiments.
: AMP
injection in the culture medium. Each datapoint (mean ± standard deviation) is
the result of triplicate experiments.
: AMP
injection in the culture medium. Each datapoint (mean ± standard deviation) is
the result of triplicate experiments.
Figure 4.
Effect of the cumulative addition of AMP
(500 mg L-1) on the antibacterial activity removal “♦” and the laccase activity evolution “■”.: AMP
injection in the culture medium. Each datapoint (mean ± standard deviation) is
the result of triplicate experiments.
: AMP
injection in the culture medium. Each datapoint (mean ± standard deviation) is
the result of triplicate experiments.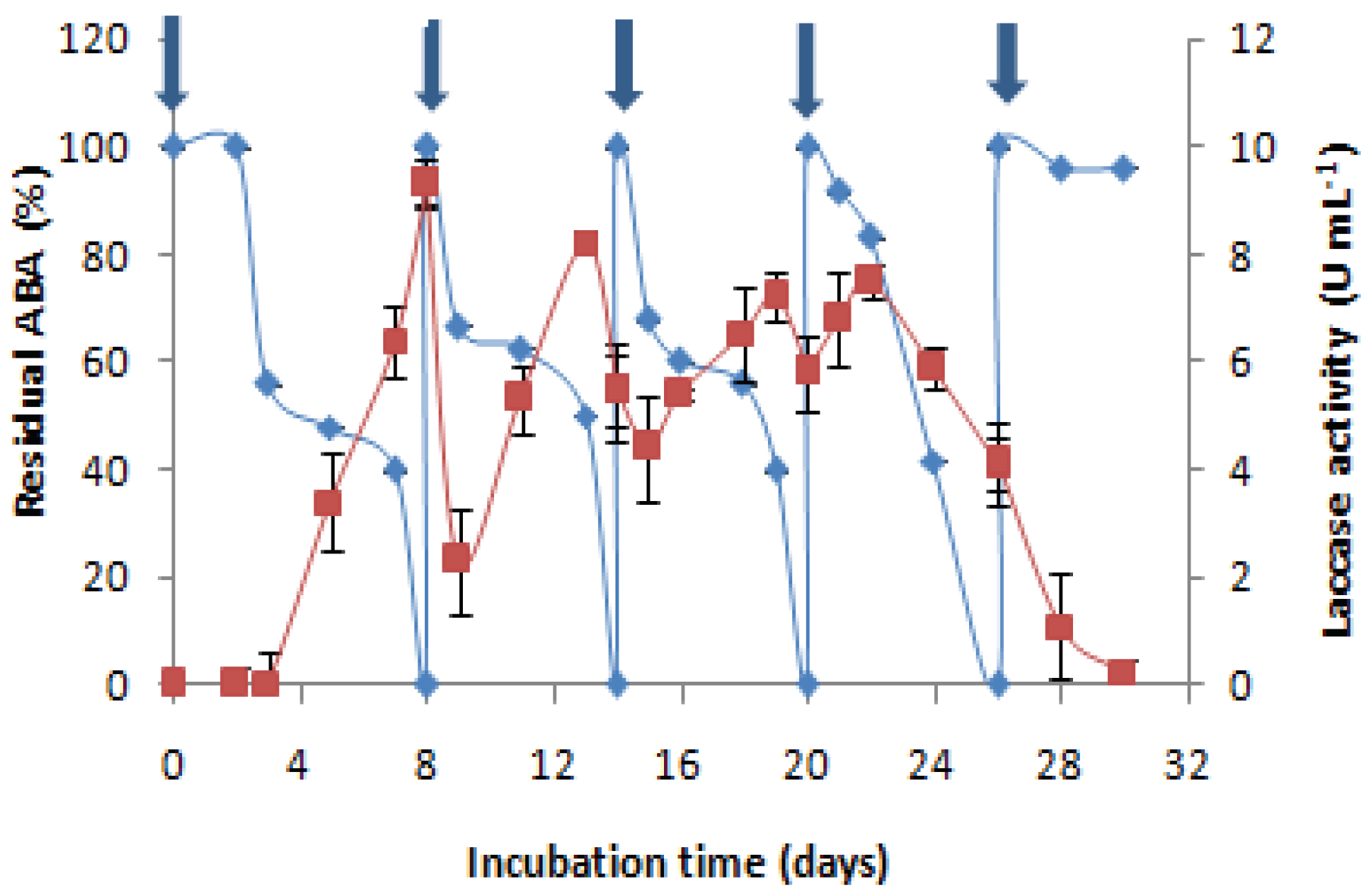

Table 1.
Physico-chemical characteristics of AMP.
| Antibiotic | Class | λmax (nm) | Chemical structure |
|---|---|---|---|
| Ampicillin | β-lactam | 204 | 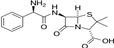 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



