
Submitted:
02 April 2024
Posted:
03 April 2024
You are already at the latest version
Abstract
L-Theanine, a unique non-protein amino acid, is an important bioactive component of green tea. Previous studies have shown that L-theanine has many potent health benefits, such as anti-anxiety effect, regulation of immune response, relaxing neural tension, reducing oxidative damage. However, little is known concerning whether L-theanine can improve the clearance of mitochondrial DNA (mtDNA) damage in organisms. Here, we reported that L-theanine treatment increased ATP production and improved mitochondrial morphology to extend the lifespan of UVC-exposed nematodes. Mechanistic investigations showed that L-theanine treatment enhanced the removal of mtDNA damage and extended lifespan by activating autophagy, mitophagy, mitochondrial dynamics, and mitochondrial unfolded protein response (UPRmt) in UVC-exposed nematodes. In addition, L-theanine treatment also up-regulated the expression of genes related to mitochondrial energy metabolism in UVC-exposed nematodes. Our study provided a theoretical basis for the possibility that tea drinking may prevent mitochondrial related diseases.
Keywords:
L-theanine
; UVC
; mitochondrial DNA damage
; C. elegans
; lifespan
1. Introduction
Energy animates life. Mitochondria is a multi-functional organelles within eukaryotic cell, and generates most energy by producing adenosine 5'-triphosphate (ATP) via oxidative phosphorylation (OxPHOs), which functions in fatty acid oxidation, apoptosis, the cell cycle and cell signaling [1]. Most enzymes required for OxPHOs are encoded by nuclear DNA (nDNA). However a small subset of 13 essential protein subunits of respiratory complexes I, III, IV and V plus 2 rRNAs and 22 tRNAs, are only be encoded by mitochondrial DNA (mtDNA) [2]. Compared with nDNA, mtDNA is more vulnerable to damage due to lacking nucleotide excision repair (NER), which repairs helix-distorting damage caused by common environmental factors, such as polycyclic aromatic hydrocarbons (PAH), mycotoxins, and ultraviolet C radiation (UVC) [3]. Previous studies reported that mtDNA is dozens of times sensitive to PAH than nDNA [4], and helix-distorting mtDNA lesions are persistent and cause a decrease in mtDNA replication and transcription [5]. Natarelli et al. have shown that persistent mtDNA damage can disrupt mitochondrial function [6]. An increasing number of studies suggested that mitochondrial dysfunction and mtDNA mutation have been associated with various human pathologies, such as cancer [7], type 2 diabetes mellitus [8], neurodegenerative conditions [9], mitochondrial diseases [10], and aging [11]. Therefore, the integrity of mtDNA is very important to the entire organism.
The nematode Caenorhabditis elegans (C. elegans), which lives primarily in decaying organic matter such as leaf litter, is well known to the scientific world because of Sydney Brenner's research in its development and neurobiology in 1965 [12]. It is widely used as one of the first-choice model organisms to study aging, stress resistance, mitochondrial biology, and apoptosis at the molecular level [13,14], mainly due to the short life span for approximately 15-21 days, as well as its completely sequencing and annotation of all the genes [15], the abundance of mutant strains [16]. The most important fact that results of trials on C. elegans can be predictive of outcomes in higher organisms [17]. There is little difference of mitochondrial biologybetween C. elegans and humans [14]. The mitochondrial genome of C. elegans is 13,794 base pairs in size, compared to 16,649 in humans [18]. The encoded genes are very similar except that the atp-8 gene has not been clearly identified in C.elegans [19]. In order to determine the long-term fate of helix-distorting mtDNA damage, Bess AS et al. used UVC to induce mtDNA damage of C. elegans to establish a system in which only mtDNA damage was detected, while nDNA damage was repaired by NER [3]. At the same time, they also found high levels of sustainability mtDNA damage could cause L3 larval arrest of C. elegans, and the degree of mtDNA damage was positively correlated with the rate of arrest [3]. Therefore, C. elegans provides a useful model for studying persistent mtDNA damage in vivo.
Previous researches showed that psychological stress very closely linked to depression, mood swings, immune and age-related diseases, cardiovascular disorders and different types of cancer [20]. To cope with the negative effects of psychological stress, people are increasingly turning to natural health supplements as preferred choice to synthetic products. Green tea is a traditional drink globally, due to its favourable taste and relaxing effects on the body [21]. Decades of scientific experiments have shown that the consumption of green tea is associated with a variety of health benefits. Therefore, the health functions of various characteristic components of green tea have become the main research topics [22]. L-Theanine (L-γ-glutamylethylamide), a unique non-protein amino acid, is an important bioactive component of green tea [23], which constitutes around 50% of total free amino acids in green tea, and accounts for 1%-2% of the weight of the dry tea [24]. In addition, L-theanine is certified as a safe, non-toxic food additive by United States Food and Drug Administration (US FDA) [25]. Previous reports have shown that L-theanine exibited potent health benefits, including neuroprotective and anxiolytic effects, regulating immune response, relaxing neural tension [26]. Hence, L-theanine is usually used as a common ingredient in functional beverages and food supplements. Pretreatment of mice with L-theanine significantly reduces irinotecan-induced genomic damage in the bone marrow cells [27]. However, little is known whether L-theanine can improve the clearance of mtDNA damage in organisms. Therefore, the current study investigated the effect of L-theanine treatment on UVC-induced mtDNA damage and its molecular mechanism in C. elegans.
2. Results
2.1. L-Theanine Enhanced the Removal of UVC-Induced mtDNA Damage
Bess et al. reported that UVC-induced mtDNA damage leads to L3 larval arrest and the proportion of L3 larval arrest was positively correlated with mtDNA damage in C. elegans [3]. Therefore, the L3 arrest phenotype serves as an important indicator of mtDNA damage. Our results showed that UVC dose-dependent increase in L3 arrest was observed in the UVC-exposed L1 nematodes grew on the plates with food for 48 h at 7.5 J/m2 start dosage (Figure 1A). When the UVC exposure dose was set-up at 12.5 J/m2, mtDNA damage was detected, while nDNA damage can not be detected in nematodes (Figure 1B). Therefore, we used 12.5 J/m2 as a dose that only induced mtDNA damage with UVC exposure in the experiment. Previously published studies found that pretreatment of mice with L-theanine significantly reduced irinotecan-induced genomic damage in the bone marrow cells [27]. This suggests that L-theanine mayhave chemoprotective activity on DNA damage in organisms. However, little is known whether L-theanine can improve the removal of UVC-induced mtDNA damage in organisms. To assess the role of L-theanine in the removal of UVC-induced mtDNA damage, we treated UVC-exposed (12.5 J/m2) L1 nematodes with various concentrations of L-theanine for 48h, and L3 arrest was measured under dissection microscope. Our results showed that L-theanine treatment in the range of 10-400 µg/ml significantly reduced L3 arrest, of which 50 µg/ml L-theanine is the most effective (Figure 1C). At the same time, the mtDNA damage was obviously reduced by treatment of 50 µg/ml L-theanine (Figure 1D). Therefore, 50 µg/ml of the L-theanine concentration was applied in the following experiments unless stated otherwise. The above results suggested that L-theanine treatment may have a role in removing UVC-induced mtDNA damage in C. elegans. In order to rule out the effect of L-theanine on L3 arrest and the development of nematodes, L1 nematodes were placed in petri dishes with various concentrations of L-theanine for 48 h, and then to detect L3 arrest and body length of nematodes. L-theanine treatment in the range of 10-400 µg/ml had no effect on L3 arrest and body length of nematodes (Figure 1E,F). The L1 nematodes were exposed to different doses of UVC and then were transferred to 50 µg/ml L-theanine treatment for 48 h to count the ratio of L3 arrest. Figure A1 showed that L3 arrest of L1 nematodes with 50 µg/ml L-theanine treatment was significantly reduced at 10 J/m2 and 12.5 J/m2 UVC, in which the more significant reduction was observed at 12.5 J/m2 UVC. Whereas L3 arrest of L1 nematodes did not significantly change at 15 J/m2 and 20 J/m2 UVC. Therefore, the combination of UVC (12.5 J/m2) with L-theanine (50 µg/ml) was used in all subsequent experiments unless stated otherwise. In order to elucidate whether L-theanine is involved in enhance the ability of nucleotide excision repair (NER) in nDNA to reduce UVC-induced L3 arrest in C. elegans, several mutant worms impaired in the nucleotide excision repair (NER) pathway were used as the UVC-exposed L1 nematodes. As shown in Figure A2, L3 arrest of L1 nematodes with L-theanine treatment was significantly reduced in the nucleotide excision repair (NER) pathway mutants (xpa-1, polh-1, and xpf-1). This suggested that L-theanine treatment had no effect on NER in nDNA in reducing UVC-induced L3 arrest in C. elegans. Taken together, these results demonstrated that L-theanine treatment enhances the removal of UVC-induced mtDNA damage in C. elegans.
2.2. L-Theanine Extended the Lifespan and Enhanced the Heat-Stress Resistance of UVC-Exposed Nematodes
Mitochondria are considered to play a key role in aging, and the aging process is accompanied by the decline of mitochondrial function [11]. The integrity of mtDNA is crucial to mitochondrial function, and persistent mtDNA damage can result in mitochondrial dysfunction [3]. Research found that the lifespan was shorten in C. elegans with dysfunctional mitochondria [11]. In order to evaluate the effect of L-theanine on the longevity of UVC-exposed nematodes, we treated UVC-exposed L1 nematodes with or without L-theanine, then the lifespan was measured . Compared to the UVC-exposed group, the L-theanine-treated group significantly increased longevity (Figure 2A). This suggested L-theanine treatment enhanced the lifespan in UVC-exposed C. elegans.
Persistent mtDNA damage causes the stalling of mtDNA replication and transcription, which lead to depletion of mtDNA and mtDNA-encoded proteins, and also induce mitochondrial dysfunction [28]. Palikaras et al. reported that the heat-stress resistance was diminished in C. elegans of mitochondrial dysfunction [29]. To detect the effect of L-theanine treatment on the heat-stress resistance of UVC-exposed nematodes, assays were carried out under heat stress. In the heat-stress resistance assaying, the UVC-exposed L1 nematodes were treated with or without L-theanine for 24 h or 48 h, and then were exposed to heat shock at 35 ℃ for 7 h. Our results showed that L-theanine treatment significantly enhanced the heat-stress resistance of UVC-exposed nematodes at 24h or 48h (Figure 2B,C). This suggested L-theanine treatment enhanced the heat-stress resistance in UVC-exposed C. elegans.
2.3. L-Theanine Improved Mitochondrial Morphology of UVC-Exposed Nematodes
Mitochondria are quite dynamic tubular organelles. They continuously were reshaped by biogenesis, fission, fusion, mitophagy, and motility in a regulated manner. These dynamic processes determine the morphology of the mitochondrial network. Changes in mitochondrial morphology are considered to be related with cellular responses to stress and pathological conditions in worms [30]. Given the requirement for mitochondrial dynamics in removal of mtDNA damage [3], we tested the effect of L-theanine on mitochondrial morphological of UVC-exposed L1 nematodes. To evaluate mitochondrial morphology, UVC-exposed L1 nematodes, which contain an extrachromosomal Pmyo-3::matrixGFP and express mito-GFP in muscle cells, were transferred to dish plates containing 50 µg/ml L-theanine. After 2, 4 and 6 days, the treated nematodes were taken pictures with confocal fluorescence microscopy. We observed that with the passage of time, the proportion of mitochondrial fragmentation increased and the proportion of tubular mitochondria decreased in UVC-exposed nematodes (Figure 3A–D). However, we found that L-theanine treatment effectively attenuated the decline of the tubular mitochondria portion in UVC-exposed C. elegans from the fourth day compared with UVC treated worms (Figure 3A–D). These results indicated that L-theanine treatment greatly improve mitochondrial morphology in UVC-exposed C. elegans.
2.4. L-Theanine Elevated Steady-State ATP Level in UVC-Exposed Nematodes
Energy production via oxidative phosphorylation (OxPHOS) is one of the main functions of mitochondria. The essential protein subunits of respiratory complexes I, III, IV and V in the OxPHOS system are encoded by mtDNA. So mtDNA damage will reduce steady-state ATP level in cell [3]. Research by Leung MC et al. has showed that the steady-state ATP level of UVC-exposured L1 nematodes was reduced at later larval development in C.elegans [31]. To test whether L-theanine treatment can improve the steady-state ATP level of UVC-exposured L1 at later larval development, we used the transgenic C. elegans (PE255) expressing GFP as a reporter transgene for inducible sur-5p expression. The result showed that ompared with UVC-exposed group, L-theanine-treated group demonstrated higher sur-5p::GFP intensity in pictures taken with confocal laser scanning microscopy at the 24 h and 48h time-point, respectively (Figure 4A–H). With quantification by Photoshop CS5, the data showed that L-theanine treatment significantly up-regulate the sur-5p::GFP expression in mutant PE255 (Figure 4G,H). This indicated that L-theanine treatment increased the steady-state ATP level of UVC-exposured L1 at later larval development.
2.5. Autophagy, Mitochondrial Dynamics, and UPRmt Mediated the Reduction of mtDNA Damage and Extension of Lifespan in UVC-Exposed C. elegans Treated by L-Theanine
MtDNA integrity plays a very important role in mitochondrial function and is extremely vulnerable to damage induced by common environmental agents including PAHs, mycotoxins, and UVC [3,25]. Previously study showed that mitochondrial dynamics, mitophagy and autophagy all contributed to removal of persistent mtDNA damage in the UVC-exposed C. elegans [3]. Therefore, we suspected that the removal of mtDNA damage by L-theanine treatment is due to the contribution of mitochondrial dynamics, mitophagy and autophagy. To verify this possibility, L3 arrest of several UVC-exposed mutant worms defective for autophagy (unc-51), mitophagy (pink-1 and pdr-1), mitochondrial dynamics ( fzo-1, eat-3, and drp-1) were treated by L-theanine. The result showed that L3 arrest was significantly inhibited by L-theanine treatment (Figure 5A,B). This suggested that the reduction of mtDNA damage by L-theanine may through mitochondrial dynamics, mitophagy and autophagy. Studies have found that unfolded protein response is a key stress response pathway that safeguards the mitochondria. Recent research has shown that mitochondrial unfolded protein response (UPRmt) plays an important role in rescuing mtDNA disease in C. elegans [32]. So, does UPRmt contribute to the removal of mtDNA damage by L-theanine treatment in UVC-exposed C. elegans? To test this hypothesis, we examined the effect of L-theanine treatment on L3 arrest of several UVC-exposed mutant worms defective for the UPRmt (atfs-1, haf-1, and ubl-5), and found that the effect of L-theanine treatment on L3 arrest was inhibited (Figure 5C), indicating the UPRmt plays a key role in the reduction of mtDNA damage by L-theanine treatment. The previous researches showed that L-theanine treatment prolongs the lifespan of wild-type nematodes exposed to UVC (Figure 2A). The life span of nematodes is closely related to the damage degree of mtDNA [11]. To explore the role of mitophagy, mitochondrial dynamics and UPRmt in prolonging the life span of nematodes exposed to UVC by L-theanine treatment, we examined the effect of L-theanine treatment on lifespan of several UVC-exposed mutant worms defective for mitophagy (pink-1), mitochondrial dynamics (drp-1) , UPRmt (atfs-1 and haf-1) and found that the effect of L-theanine treatment on lifespan extension was inhibited (Figure 5D–H). The above results indicated that L-theanine treatment prolonged the lifespan of nematodes exposed to UVC, most likely by activating multipal pathways, including autophagy, mitochondrial autophagy, mitochondrial dynamics and UPRmt to enhance the clearance ability of mtDNA damage.
The previous results showed the reduction of mtDNA damage by L-theanine treatment depended on autophagy, mitophagy, mitochondrial dynamics, and UPRmt. Therefore, the following study was focused on the effect of L-theanine treatment on the gene expression of autophagy, mitophagy, and UPRmt in UVC-exposed C. elegans. The results showed that compared to UVC treatment group, L-theanine treatment group significantly up-regulated the gene expression of bec-1,dct-1 and hsp-60, without causing any changes to the gene expression of lgg-1 and atg-18 (Figure 6). These results further confirmed that autophagy, mitophagy and UPRmt play a key role in the reduction of mtDNA damage and prolonging lifespan of nematodes exposed to UVC by L-theanine treatment. Persistent mtDNA damage causes mitochondrial dysfunction, which ultimately impairs energy metabolism [33]. Therefore, the expression of mitochondrial energy metabolism related genes were tested in UVC-exposed nematodes with or without L-theanine treatment. As shown in (Figure A3),compared to UVC treatment group, L-theanine treatment group significantly up-regulated the gene expression of pcy-1 and cox-4 , while the gene expression of cts-1 and hxk-1 were not changed. These results indicated that L-theanine treatment improve the energy metabolism damage caused by mtDNA damage.
3. Discussion
A growing number of studies have shown that the integrity of mtDNA plays an important role in human aging and disease [11,29,32]. Due to the lack of nucleotide excision repair (NER), which is used to repair helix-distorting DNA damage caused by common environmental factors, mitochondria is easy to produce persistent mtDNA damage [3,25]. Bess AS et al. established a C. elegans model of UVC induced mtDNA damage, and found that the proportion of L3 larval arrest was positively proportional to the degree of mtDNA damage [3]. Therefore, L3 larval arrest is an important phenotypic indicator of mtDNA damage. Consistent with the Bess et al research, our results showed that the L3 larval arrest was proportional to the dose of UVC in C. elegans (Figure 1A). Mitochondria are highly dynamic tubular organelles that are constantly remodeled by fusion and division, and their morphology is closely related to mtDNA rescue [3,34]. Research by Bess AS et al. also showed that young adult nematodes were exposed to UVC with a single dose of 50 J/m2 and mitochondrial morphology was not significant variability at 24 h and 48 h post exposure [3]. It is worth noting that we observed the mitochondrial morphology of UVC-exposed L1 nematodes became fragmented and disorganized on the 4th day after exposure (Figure 3A). We speculated that the above results may be mainly due to the different periods of nematodes exposed to UVC and the longer observation period than previous report. Rsearch by Shokolenko IN et al. showed that despite the rapid loss of mtDNA in Hela/2641 cells transduced with either exoIII or mUNG1 construct, changes in mitochondrial morphology were not observed on the first 4 days, but only after 120 h [35]. Their results suggested that it takes time for mtDNA damage to cause morphological changes in mitochondria.
The integrity of mtDNA is crucial to the proper function of the mitochondrial respiratory chain. Most of ATP in organism is produced by oxidative phosphorylation conducted by the mitochondrial respiratory chain in mitochondria. Persistent mtDNA damage will lead to mitochondrial dysfunction, which further leads to a decreased capacity to produced ATP [35]. Leung MC et al research showed that the production of ATP in the development of UVC-exposed L1 nematodes was significantly lower than that of the nematodes without UVC treatment [18]. Our results also found that the ATP production of UVC-exposed group was significantly lower than that of un-exposed group at the 24 h and 48 h time-point (Figure 4A–H). Thus, we concluded that L1 nematodes exposed to UVC are an excellent animal model for studying mtDNA damage, which can cause changes in mitochondrial morphology and reduce ATP content.
Mitochondria are the main sites for oxidative phosphorylation, supplying more than 90% of cellular ATP [36]. The few small subunits required for oxidative phosphorylation can only be encoded by mtDNA [37]. Reactive oxygen species (ROS) originate from a range of cellular processes, external factors, and/or various diseases [38]. ROS can cause oxidative damage to biological molecules including nucleic acids, proteins, and lipids, in which mitochondrial DNA is the most sensitive [38]. Damaged mtDNA increases the production of oxygen free radicals and further exacerbates the mitochondrial dysfunction. Therefore, some researchers pay attention to some natural active substances which can protect mtDNA as well as have antioxidant effect. A variety of antioxidant substances have been found to improve mtDNA damage caused by oxidative stress, such as curcumin [39], resveratrol [40], vitamins C [41], vitamins E [41], Phellinus linteus [42], and green tea extract [43]. Oxidative stress-induced mtDNA damage can be repaired by base excision repair pathway to remove oxidized bases [44], but it is difficult to repair helix-distorting mtDNA damage caused by common environmental factors due to the lack of nucleotide excision repair. To date, there are few reports that natural active substances are helpful to remove the helix-distorting mtDNA damage. L-theanine (L-γ-glutamylethylamide), a unique non-protein amino acid, is an important bioactive component of green tea [26]. It has been reported that L-theanine has numerous potent health benefits, including neuroprotective effects, anxiolytic effects, regulating immune response, relaxing neural tension [26]. But so far, there is no report about the effect of L-theanine on mtDNA damage. In the present study, we found that L-theanine treatment significantly reduced the UVC-induced L3 larval arrest, which indicates that L-theanine treatment is helpful to the removal of UVC-induced mtDNA damage in C. elegans (Figure 1B). Furthermore, the study also showed that L-theanine treatment improve the mitochondria morphology and increase the content of ATP in UVC-exposed C. elegans. Therefore, we concluded that L-theanine treatment enhance the ability to remove mtDNA damage and improve mitochondrial function in UVC-exposed C. elegans.
Autophagy and mitochondrial dynamics (fission, fusion, and mitophay) play important roles in eradicating damaged mitochondria, promoting overall mitochondrial function and ensuring the continuity of mitochondrial function [45]. Research by Bess AS et al. showed that mitochondrial dynamics and autophagy contribute to the removal of UVC-induced mtDNA in C. elegans [3]. UNC51 is a serine/threonine kinase with a role in autophagy induction. PINK1 is a serine/threonine-protein kinase which is a critical conserved component of mitophagy DRP1 is an important protein required for mitochondrial fission. In addition, FZO1 is a necessary protein to control outer mitochondrial membrane fusion event. Research found that mutation of these genes results in loss of autophagy, fission, and fusion in C. elegans [46]. The present study found that all mutants related to autophagy (unc-51), mitophagy (pink-1, pdr-1), fusion (fzo-1, eat-3), and fission (drp-1) have resulted in diminishing effect of L-theanine treatment on enhancing the removal of UVC-induced mtDNA damage (Figure 5A,B), indicating that autophagy and mitochondrial dynamics mediated in the reduction of mtDNA damage by L-theanine treatment. Furthermore, quantitative realtime-PCR results suggested that L-theanine treatment significantly up-regulated the mRNA expression of autophagy genes (bec-1, lgg-1) and mitophagy gene (dct-1) (Figure 6) in UVC-exposed C. elegans. Similar results were obtained in a recent study demonstrated that resveratrol alleviation of rotenone-induced mtDNA damage could be mediated through regulating the balance of mitochondrial dynamics [40]. Therefore, we concluded that L-theanine treatment improves mitochondrial dysfunction caused by mtDNA damage through augmenting autophagy and mitochondrial dynamics to enhance the clearance of mtDNA damage in UVC-exposed C. elegans.
It was reported that stresses that cause mitophagy can also activate mitochondrial unfolded protein response (UPRmt) [47]. The UPRmt is a mitochondria-to-nuclear signal transduction pathway, which is caused by the accumulation of unfolded proteins in the mitochondria, resulting in the induction of mitochondrial protective genes including mitochondrial molecular chaperones and proteases [48,49]. Previous research showed that activation of UPRmt rescue the neuromuscular defect of the polg-1(srh1) worms, which suffer from mtDNA depletion, indicating that UPRmt is helpful to improve mitochondrial dysfunction [32]. Multiple factors required to induce UPRmt have been reported, including the HAF-1 peptide exporter, the CLPP-1 protease, a ubiquitin-like protein UBL-5, and two transcription factors, DVE-1, and ATFS-1 (ZC376.7) [50]. Thus, mutants related to UPRmt (atfs-1, haf-1, and ubl-5) have been used in this study and it was found that the effect of L-theanine treatment on enhancing the removal of UVC-induced mtDNA damage was inhibited (Figure 5C). Moreover, the mRNA expression of UPRmt gene (hsp-60) was significantly up-regulated in L-theanine treatment group (Figure 6). Hence, we concluded that L-theanine treatment enhance UPRmt to improve mitochondrial dysfunction induced by mtDNA damage in UVC-exposed C. elegans.
Aging is a degenerative process caused by the accumulation of damaged lipid and protein that leads to cellular dysfunction, tissue and organ failure, and death [11]. Most human pathologies and aging have a common sign: impaired mitochondrial maintenance in disparate cell types [51]. A study showed the accumulation of mtDNA damage changes with the degree of tissue aging in mammals [11]. In this study, we found that L-theanine treatment not only increase the survival of UVC-exposed nematodes under heat stress, but also extend the life of UVC-exposed nematodes under normal culturing conditions (Figure 2). Therefore, we hypothesized that the extended lifespan of UVC-exposed nematodes was mainly due to L-theanine's ability to remove UVC-induced mtDNA damage and improve mitochondrial dysfunction induced by mtDNA damage in UVC-exposed C. elegans.
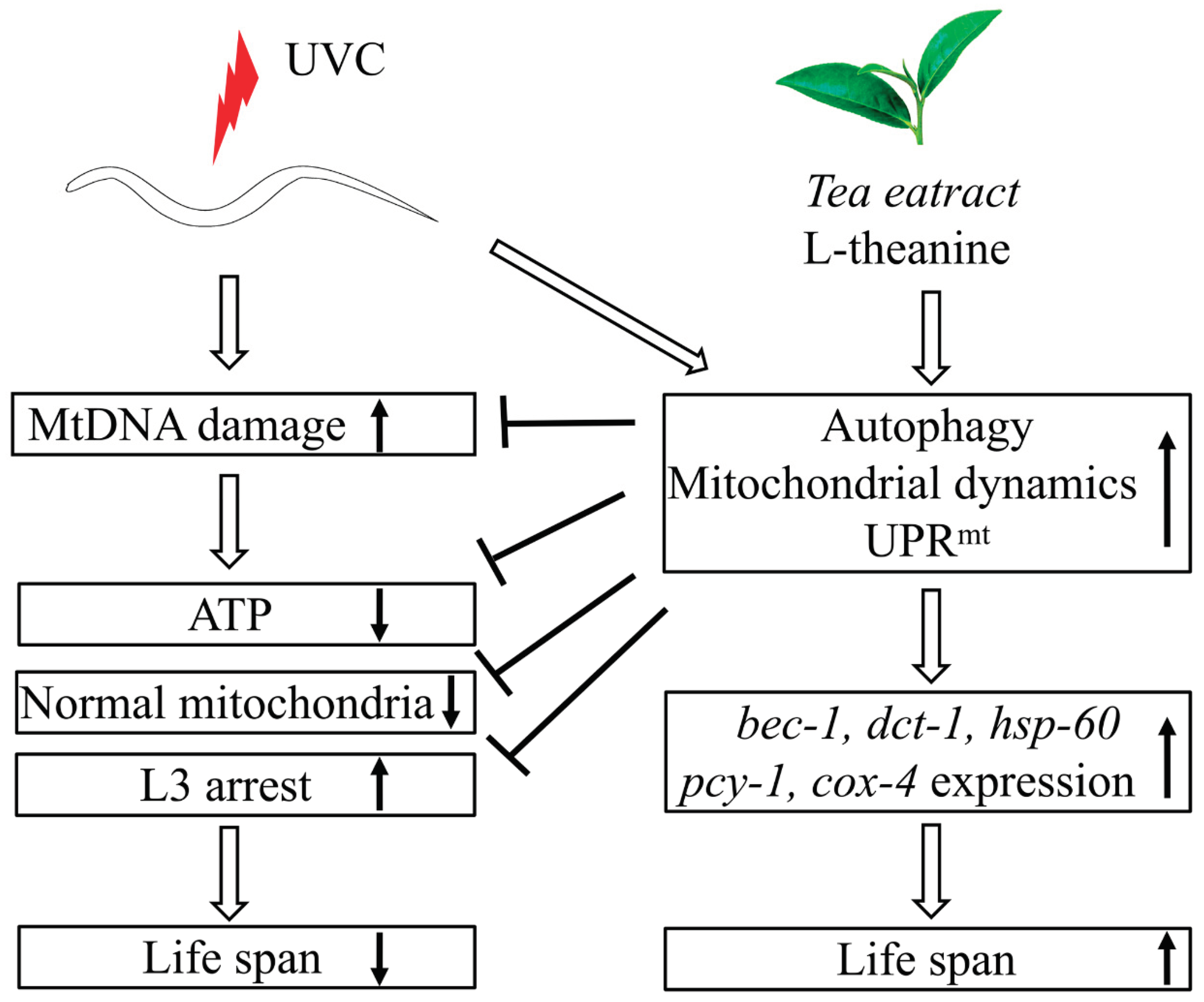
Generally, here we canonically demonstrated that L-theanine treatment extended the lifespan of UVC-exposed nematodes mainly by enhancing the ability of eliminating mtDNA damage to increase ATP production and improve mitochondrial morphology in UVC-exposed nematodes. While the primary mechanism of L-theanine action is preliminarily clarified, as shown in Figure 7. Our data provide theoretical basis for the potential of tea drinking to improve mitochondrial related diseases.
4. Materials and Methods
4.1. Reagents
L-theanine, 98% purity (Sigma, St. Louis, MO, USA), was dissolved in distilled water, sterilized by filtration through 0.2 μm pore size membranes, and then stored at - 20 ℃ for later use. All other chemicals and reagents were also purchased from Sigma-Aldrich unless otherwise stated.
4.2. C. elegans Strains and Culture Conditions
C. elegans strains were maintained at 20 ℃ on K agar plates seeded with OP50 strain Escherichia coli unless stated otherwise [52]. However, JK1107 ( glp-1(q224) III was maintained at 15 ℃. All C. elegans strains were obtained from the Caenorhabditis Genetics Center (University of Minnesota, MN, USA). They are N2 (Bristol, wild type), CB369: unc-51(e369)V, RB2547: pink-1(ok3538)II, VC1024: pdr-1(gk448) III, CU5991: fzo-1(tm1133) II, DA631: eat-3(ad426) II, CU6372: drp-1(tm1108) IV, QC115: atfs-1(et15) V, VC2654: ubl-5(ok3389) I, RB867: haf-1(ok705) IV, PE255: feIs5 [sur-5p::luciferase::GFP + rol-6(su1006)] X, SJ4103: zcIs14 [myo-3::GFP(mit)], respectively.
4.3. UVC Exposure
UVC exposures were conducted in a custom-built exposure cabinet. Dose intensity of radiation was measured with a UVX digital radiometer (UVP, Upland, CA) and was used to calculate the exposure time required to reach the desired dose. Synchronized L1 worms were placed on plates that lacked bacterial food and exposed with the desired dose. Finally, under aseptic conditions, L1 larvae exposed by UVC were evenly distributed to culture plates containing food at a density of at least 300 worms per plate.
4.4. Lifespan Experiments and Heat-Stress Resistance Assays
Lifespan assays were performed at 20 ℃ with wild type. L1 larvae exposed to UVC were placed on culture plate containing L-theanine at a density of at least 40 worms per plate. To avoid the influence of offspring, worms were transferred to new plates every day until all the nematodes died. The viability of worms was checked by touching their pharynx with picking needles made of eyebrows.
Heat-stress resistance assays were performed at 35 ℃ with wild type [55]. The L1 nematodes treated with UVC were placed on culture plates (Φ=6 cm) containing L-theanine of various concentrations at 20 ℃ for 24 h and 48 h, then exposed to heat shock at 35 ℃ for 6 h, and finally transferred to the incubator at 20 ℃ for 10 h before the number of nematode deaths was recorded. Under each concentration of L-theanine, 4 independent plates at a density of around 45 worms per plate were used and then repeated 3 times. The standard for nematode death is no response to a gentle touch.
4.5. L3 Arrest Analysis
The L1 nematodes treated with UVC were placed on culture plates (Φ=6 cm) containing L-theanine of various concentrations at a density of about 400 worms per plate. After culturing UVC-exposed L1 nematodes at 20 ℃ for 48 h, each worm was staged, and the proportion of worms that entered L4 was determined based on the presence of the vulval crescent [3]. Unstressed wild type nematodes will be in the young adult stage for approximate 2 h after 48 h of growth at 20 ℃, while some mutants will suffer from delayed growth due to the lack of some genes. In order to eliminate the influence of delaying development on the proportion of L3 arrest, the unstressed mutants enter the young adult stage for around 2 h as a reference for the counting time.
4.6. MtDNA and nDNA Damage Measurements
All the specific steps of the experiment are followed by Gonzalez-Hunt CP et al protocol [56]. The DNA damage measurement is quantified by measuring the number of polymerase-inhibiting lesions present based on the amount of amplification obtained from a long amplicon quantitative PCR (LA-QPCR). The lesion frequency was measured based on decreases in amplification efficiency relative to controls, which are assumed to be undamaged. The DNA template for long-amplicon quantitative polymerase chain reaction (LA-QPCR) was directly from the lysate of nematodes (N2, six worms). Primers used for LA-QPCR were as follows, mtDNA primers, forward-5΄-CCA TCA ATT GCC CAA AGG GGA GT-3΄, reverse-5΄-TGT CCT CAA GGC TAC CAC CTT CTT CA-3΄ (10.9 kb); nDNA primers, forward-5΄-TGG CTG GAA CGA ACC GAA CCA T-3΄, reverse-5΄-GGC GGT TGT GGA GTG TGG GAA G-3΄ (9.3 kb) [56]. To calculate the copy number of mitochondria and nuclear genome , we used worm glp-1 lysate to replace the plasmid with the known copy number to create a standard curve. Because glp-1 worms do not develop a germline when grow at 25 ℃.The concentration of 40 μl young adult glp-1 worm (20 worms) lysate is 1567 copies/µl. Primers used for Real-Time-PCR were as follows. The mtDNA primers are forward-5΄-AGC GTC ATT TAT TGG GAA GAA GAC-3΄, reverse-5΄-AAG CTT GTG CTA ATC CCA TAA ATG T-3΄ (75 bp); and nDNA primers are forward-5΄-GCC GAC TGG AAG AAC TTG TC-3΄, reverse-5΄-GCG GAG A TC ACC TTC CAG TA-3΄ (164 bp) [56]. The amount of amplified long PCR product provides a measurement of lesion frequency, while the amount of short PCR product provides normalization to DNA concentration and genome copy number.
4.7. Body Length
The L1 nematodes were cultured on K agar plates (Φ=6 cm) containing L-theanine of various concentrations indicated at 20 ℃. After 48 h, 50 worms were randomly selected from each group and anesthetized with 200 µl of 10 mM sodium azide solution for 5 minutes. The anesthetized nematodes were photographed by a stereomicroscope fitted with a camera and the length was measured using Image J software. The body length was calculated by measuring the tip-to-tail length of the worm [57].
4.8. Steady-State ATP Level Analysis
The luciferase-expressing nematode strain (PE255 glp-4) was used to investigate relatively steady state ATP levels in live worms. The treated and synchronized strain was cultured at 20℃ for 24 h and 48 h, and then mounted on 2% agarose pads dripping with 3% sodium azide for observing the fluorescence of the reporter GFP. Epifluorescence images were acquired at the same exposure parameter using the 20×objective of a microscope (BX 60, Olympus, Tokyo, Japan) equipped with a digital camera (Micropublisher 5.0, QIMAGING, Burnaby, BC, Canada). For quantifying a population of GFP reporter animals, each 20×image was analyzed using Photoshop CS5. For each experiment, multiple worms (n≥30) were imaged and blindly scored.
4.9. Quantitative Realtime-PCR
L1 stage worms exposured to UVC were treated with or without 50 µg/ml L-theanine for 48 h. Total RNA was isolated using Trizol reagent (Invitrogen, Carlsbad, CA, USA) from approximate 3000 worms, and cDNA was synthesized by oligo dT priming. The tba-1 expression level was taken as the internal control to normalize the amount of mRNA obtained from target genes. Samples were run in triplicate, and primers of target genes were listed in Table A1.
4.10. Confocal Microscopy and Image Processing
Worms were immobilized with levamisole and mounted on 2% agarose pads for microscopic examination with a Zeiss LSM 710 upright confocal microscope (Carl Zeiss AG, Jena, Germany). Worms with mitochondria-tagged GFPs in body wall muscle (zcIs14 [myo-3::GFP(mit)]) were used to assess alterations in mitochondrial morphology. The images of mitochondrial morphology were taken on confocal microscope at 60× magnification. For each experiment, multiple worms (n≥30) were imaged and blindly scored. The morphological categories of mitochondria were defined according to a previous study report [51]. The tubular was used as a highly connected network of long/filamentous mitochondria like tube shape, intermediate as a combination of interconnected and fragmented mitochondria and fragmented indicated a small round mitochondria in the taken images.
4.11. Statistical Analysis
The results of all experiments were verified at least three times. All data were presented as means ± standard deviations. Statistical comparisons of all data were performed by Student’s t-test. P < 0.05 or less was considered to be significant.
Author Contributions
Formal analysis, L.C. and Z.X.; Funding acquisition, Z.X. and Y.W.; Software, X.Z. and J.Z.; Investigation, L.C., R.W., X.W., and Y.G.; Writing—original draft, L.C., T.G., and G.C.; Writing—review and editing, Z.X. and Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Open Fund of State Key Laboratory of Tea Plant Biology and Utilization (SKLTOF20220129), the major Project of Higher Education Natural Science Research of Anhui Province (2023AH051553), the major Project of Higher Education Natural Science Research of Anhui Province (2022AH040226), Natural Science Foundation of Anhui Province (2208085MB37).
Data Availability Statement
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
Acknowledgments
We thank the Caenorhabditis Genetics Center (CGC). Some strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440).
Conflicts of Interest
The authors declare that there are no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
Appendix A

Table A1.
List of primers used for the quantitative real-time reverse transcription-polymerase chain reaction.
Table A1.
List of primers used for the quantitative real-time reverse transcription-polymerase chain reaction.
| Gene name | Primer sequence |
|---|---|
|
bec-1 lgg-1 atg-18 dct-1 hsp-60 cox-4 cts-1 pyc-1 hxk-1 tba-1 |
Fw (5΄TGATCTCTGCTGACAAGGCTT3΄) Rv (5΄CCGACCTTGAATCCAGTTGG3΄) Fw (5΄GCACCAAAGTCAAAGCTCCA3΄) Rv (5΄CCTCGTGATGGTCCTGGTAG3΄) Fw (5΄TGGGGCACAAAGATGGCTA3΄) Rv (5΄CCAAGATGTGTAAGATTTTCGCC3΄) Fw (5΄ATCGCACAATCTCCTCACGT3΄) Rv (5΄GGACAGTCTTTGGAGGTGTATT3΄) Fw (5΄GGGGAAGCCCAAAGATCACA3΄) Rv (5΄TCCAGCCTCCTCATTAGCCT3΄) Fw (5΄GCCCCAATTCGCGCCAAGGA3΄) Rv (5΄AGGTTGGCGGCAGTTCTGGG3΄) Fw (5΄CTCGACAACTTCCCAGATAACC3΄) Rv (5΄GGTACAGGTTGCGATAGATGATAGC3΄) Fw (5΄TCCAACTACTCCTCTTGCTACTGAC3΄) Rv (5΄GTGATCATACATCCTGGTCTACTGC3΄) Fw (5΄GTGCGACGAGTACTTTCTCAACTG3΄) Rv (5΄CTAGAGATGACGTCACACACTTCTC3΄) Fw (5΄TGATCTCTGCTGACAAGGCTT3΄) Rv (5΄CCGACCTTGAATCCAGTTGG3΄) |
Figure A1.
Effect of combination of various concentrations of L-theanine treatment and different doses of UVC irradiation on L3 arrest. The most obvious reduction of L3 arrest was in the combination of UVC (12.5 J/m2) + L-theanine (50 µg/ml). Results are means ± SD (n = 5, t test, * P < 0.05, ** P < 0.01).
Figure A1.
Effect of combination of various concentrations of L-theanine treatment and different doses of UVC irradiation on L3 arrest. The most obvious reduction of L3 arrest was in the combination of UVC (12.5 J/m2) + L-theanine (50 µg/ml). Results are means ± SD (n = 5, t test, * P < 0.05, ** P < 0.01).
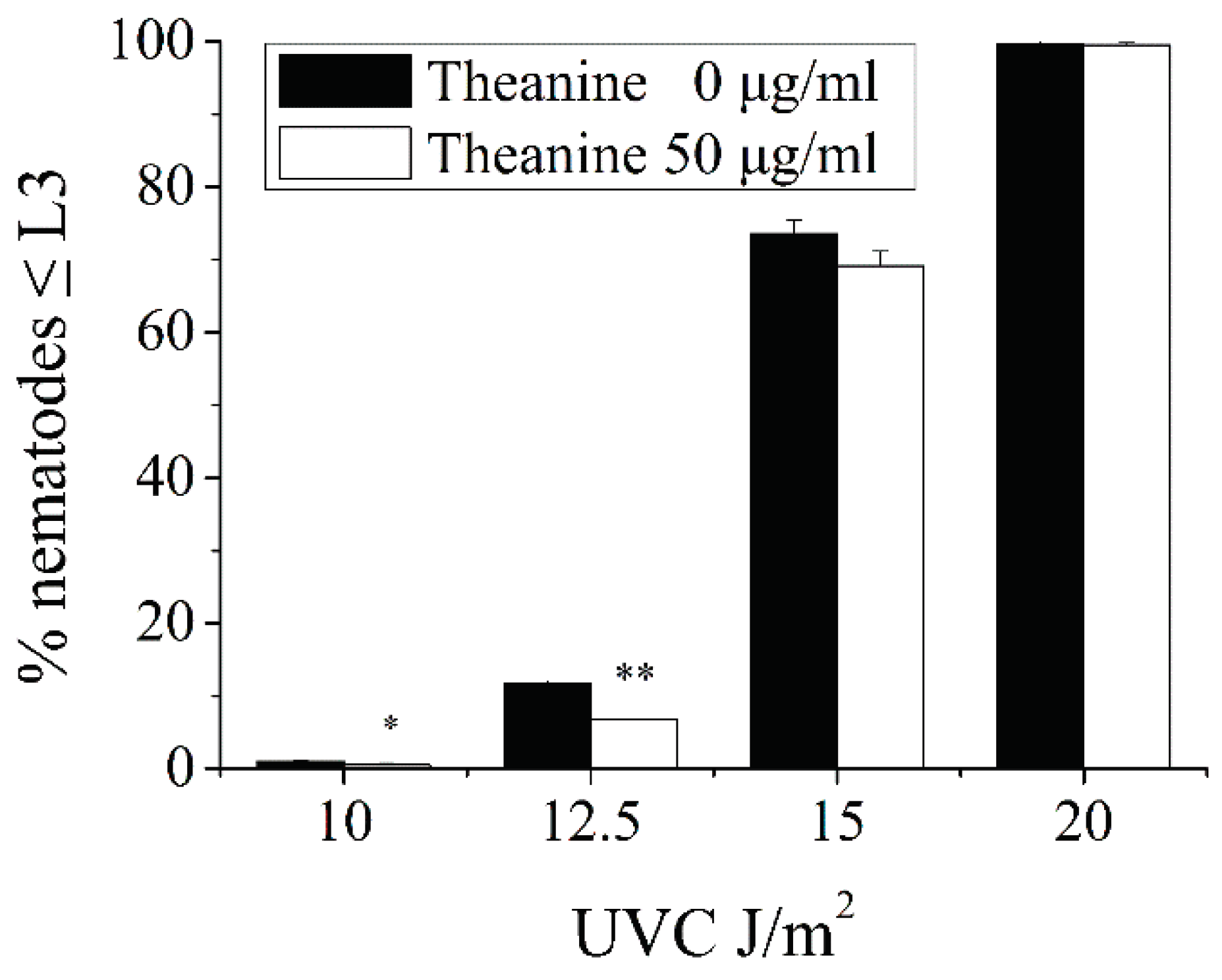
Figure A2.
Mutations in NER gene (xpa-1, polh-1, and xpf-1) did not affect L3 arrest in UVC-exposed C. elegans treated by L-theanine. Results are means ± SD (n = 5, t test, ** P < 0.01).
Figure A2.
Mutations in NER gene (xpa-1, polh-1, and xpf-1) did not affect L3 arrest in UVC-exposed C. elegans treated by L-theanine. Results are means ± SD (n = 5, t test, ** P < 0.01).
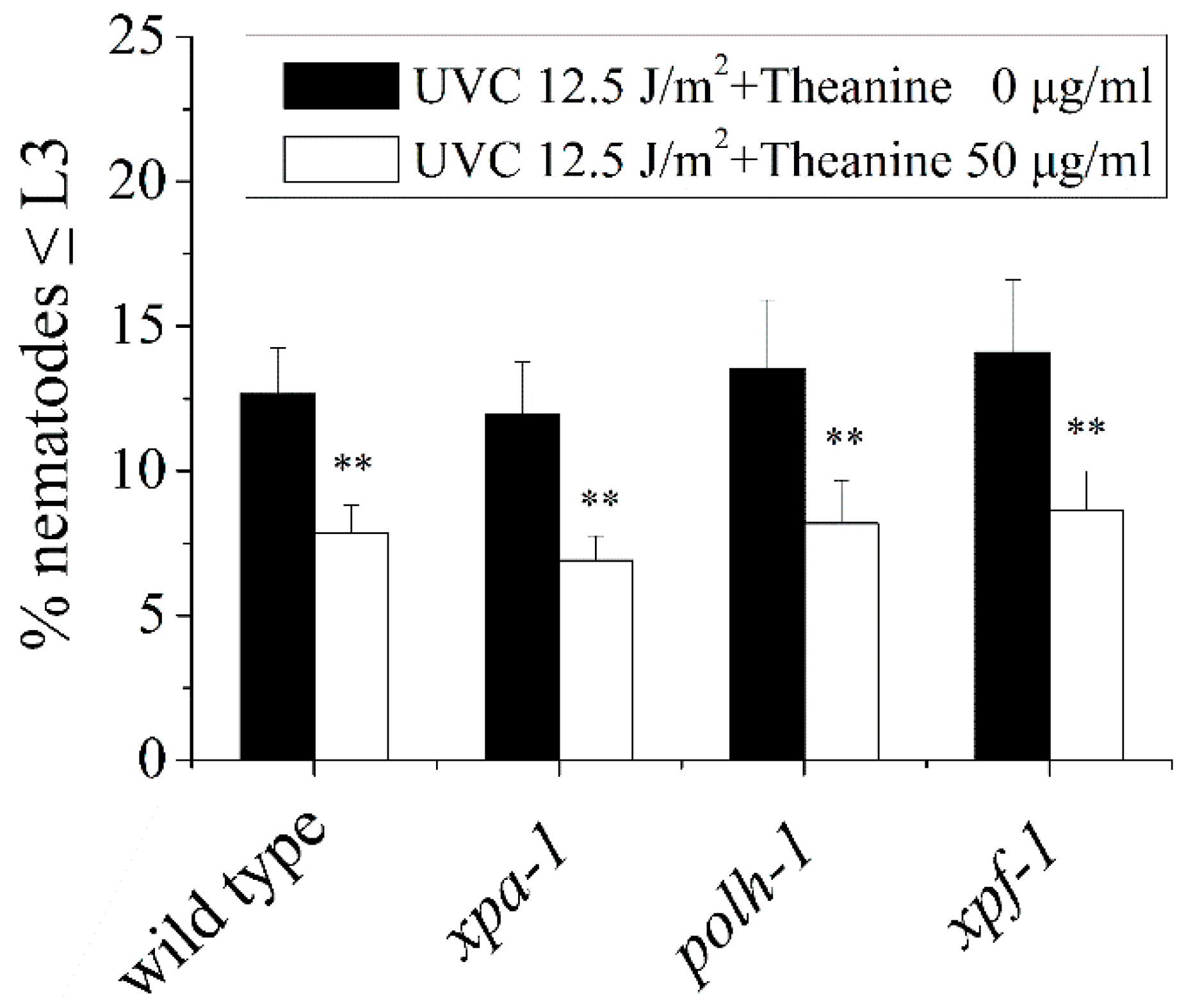
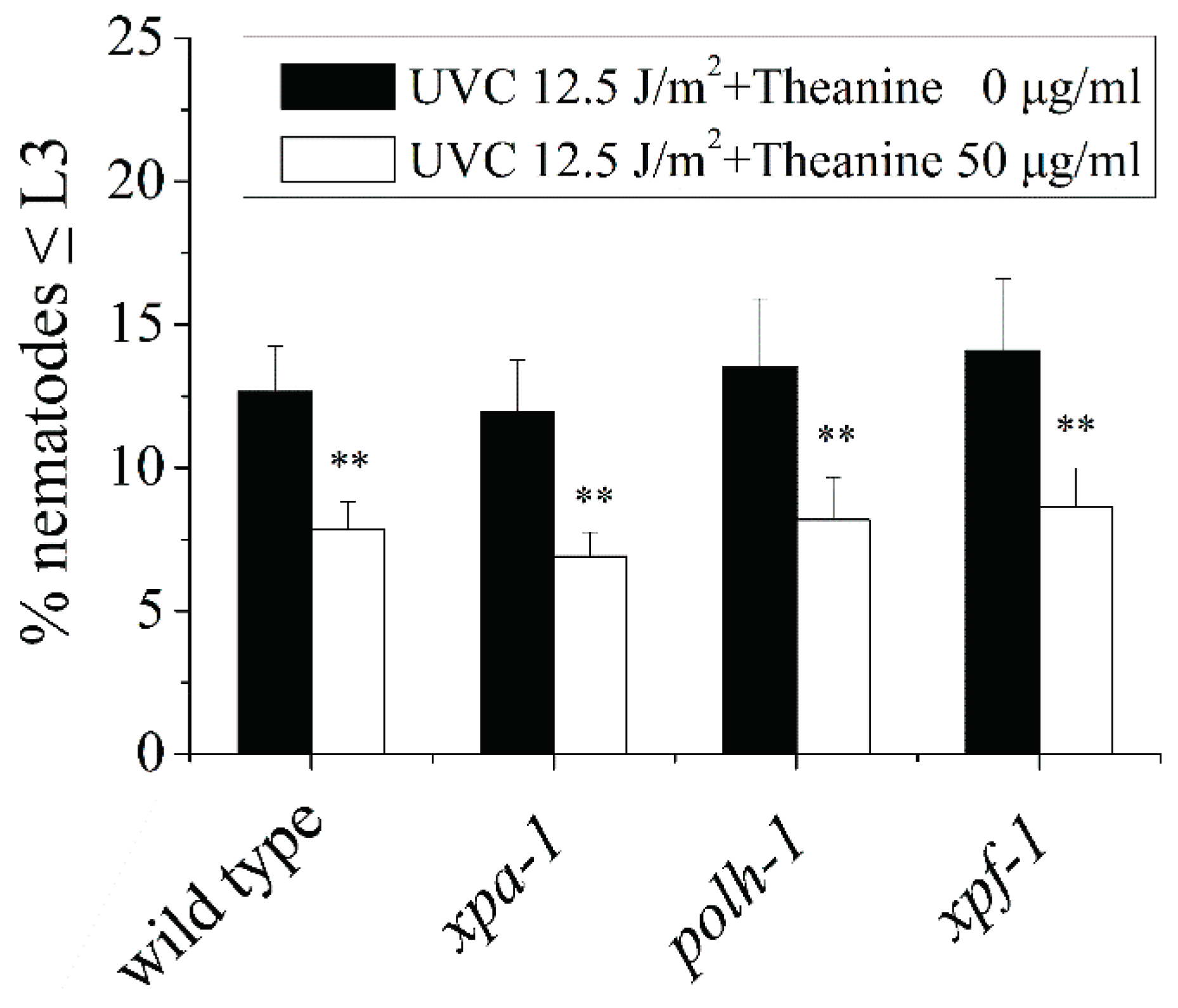
Figure A3.
Effect of L-theanine treatment on the relative expression of mitochondrial energy metabolism related genes in UVC-exposed C. elegans. L-theanine treatment did not affect hxk-1(glycolysis) and cts-1 (TCA cycle) mRNA levels in UVC-exposed C. elegans treated by L-theanine, but up-regulated pyc-1 (TCA cycle) and cox-4 (mitochondrial respiratory chain) expression. Results are means ± SD (n = 5, t test, ** P < 0.01).
Figure A3.
Effect of L-theanine treatment on the relative expression of mitochondrial energy metabolism related genes in UVC-exposed C. elegans. L-theanine treatment did not affect hxk-1(glycolysis) and cts-1 (TCA cycle) mRNA levels in UVC-exposed C. elegans treated by L-theanine, but up-regulated pyc-1 (TCA cycle) and cox-4 (mitochondrial respiratory chain) expression. Results are means ± SD (n = 5, t test, ** P < 0.01).
References
- Cunha-Oliveira, T.; Montezinho, L.; Simões, R.F.; Carvalho, M.; Ferreiro, E.; Silva, F.S.G. Mitochondria: A Promising Convergent Target for the Treatment of Amyotrophic Lateral Sclerosis. Cells 2024, 13, 248. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.N.; Lo Giudice, C.; Lavecchia, A.; Poeta, M.L.; Chiara, M.; Picardi, E.; Pesole, G. Mitochondrial and Nuclear DNA Variants in Amyotrophic Lateral Sclerosis: Enrichment in the Mitochondrial Control Region and Sirtuin Pathway Genes in Spinal Cord Tissue. Biomolecules 2024, 14, 411. [Google Scholar] [CrossRef]
- Bess, A.S.; Crocker, T.L.; Ryde, I.T.; Meyer, J.N. Mitochondrial dynamics and autophagy aid in removal of persistent mitochondrial dna damage in Caenorhabditis elegans. Nucleic acids research 2012, 40, 7916–7931. [Google Scholar] [CrossRef] [PubMed]
- Dobson, N.L.; Kleeberger, S.R.; Burkholder, A.B.; Walters, D.M.; Gladwell, W.; Gerrish, K.; Vellers, H.L. Vanadium Pentoxide Exposure Causes Strain-Dependent Changes in Mitochondrial DNA Heteroplasmy, Copy Number, and Lesions, but Not Nuclear DNA Lesions. Int. J. Mol. Sci. 2023, 24, 14507. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.A.; Gerrard, J.M.; Glover, S.M.; Ghr, R. Preferential attack of mitochondrial DNA by afiatoxin B1 during hepatocaricongenesis. Science 1982, 215, 73–75. [Google Scholar]
- Natarelli, N.; Gahoonia, N.; Aflatooni, S.; Bhatia, S.; Sivamani, R.K. Dermatologic Manifestations of Mitochondrial Dysfunction: A Review of the Literature. Int. J. Mol. Sci. 2024, 25, 3303. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, J.; Lu, W. The significance of mitochondrial dysfunction in cancer. Multidisciplinary Digital Publishing Institute 2020, 21, 5598. [Google Scholar] [CrossRef]
- Pinti, M.V.; Fink, G.K.; Hathaway, Q.A.; Durr, A.J.; Kunovac, A.; Hollander, J.M. Mitochondrial dysfunction in type 2 diabetes mellitus: an organ-based analysis. Am J Physiol Endocrinol Metab. 2019, 316, 268–285. [Google Scholar] [CrossRef]
- Ahmad, F.; Sachdeva, P. Critical appraisal on mitochondrial dysfunction in Alzheimer's disease. Aging Med (Milton). 2022, 5, 272–280. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, Y.; Xu, C.; An, P.; Luo, Y.; Jiao, L.; Luo, J.; Li, Y. Mitochondrial Dysfunction and Therapeutic Perspectives in Cardiovascular Diseases. Int J Mol Sci. 2022, 23, 16053. [Google Scholar] [CrossRef]
- Guo, Y.; Guan, T.; Shafiq, K.; Yu, Q.; Jiao, X.; Na, D.; Li, M.; Zhang, G.; Kong, J. Mitochondrial dysfunction in aging. Ageing Res Rev. 2023, 88, 101955. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, F.; Eckert, G.P. Caenorhabditis elegans as a Model for the Effects of Phytochemicals on Mitochondria and Aging. Biomolecules 2022, 12, 1550. [Google Scholar] [CrossRef] [PubMed]
- Onraet, T.; Zuryn, S. C. elegans as a model to study mitochondrial biology and disease. Semin Cell Dev Biol. 2024, 154 (Pt A), 48–58. [Google Scholar] [CrossRef]
- Consortium, C.S. Genome sequence of the nematode C. elegans: a platform for investigating biology. Science 1998, 282, 2012–2018. [Google Scholar]
- Zhang, X.; Chen, Q.; Chen, L.; Chen, X.; Ma, Z. Anti-Aging in Caenorhabditis elegans of Polysaccharides from Polygonatum cyrtonema Hua. Molecules 2024, 29, 1276. [Google Scholar] [CrossRef] [PubMed]
- Somuah-Asante, S.; Sakamoto, K. Stress Buffering and Longevity Effects of Amber Extract on Caenorhabditis elegans (C. elegans). Molecules 2022, 27, 3858. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.C.; Rooney, J.P.; Ryde, I.T.; Bernal, A.J.; Bess, A.S.; Crocker, T.L.; Ji, A.Q.; Meyer, J.N. Effects of early life exposure to ultraviolet C radiation on mitochondrial DNA content, transcription, ATP production, and oxygen consumption in developing Caenorhabditis elegans. BMC Pharmacol Toxicol. 2013, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Breton, S.; Stewart, D.T.; Hoeh, W.R. Characte.rization of a mitochondrial ORF from the gender-associated mtDNAs of Mytilus spp. (Bivalvia: Mytilidae): Identification of the “missing” ATPase 8 gene. Mar Genom. 2010, 3, 11–18. [Google Scholar] [CrossRef]
- Sharma. E.; Joshi, R.; Gulati, A. L-Theanine: An astounding sui generis integrant in tea. Food Chemistry 2018, 242, 601–610. [Google Scholar] [CrossRef]
- Ouyang, J.; Peng, Y.; Gong, Y. New Perspectives on Sleep Regulation by Tea: Harmonizing Pathological Sleep and Energy Balance under Stress. Foods 2022, 11, 3930. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.Y.; Meng, X.; Gan, R.Y.; Zhao, C.N.; Liu, Q.; Feng, Y.B. Li, S.; Wei, X.L.; Atanasov, A.G.; Corke, H.; Li, H.B. Health Functions and Related Molecular Mechanisms of Tea Components: An Update Review. Int J Mol Sci. 2019, 20, 6196. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Kang, J.; Zhu, H.; Wang, K.; Han, Z.; Wang, L.; Liu, J.; Wu, Y.; He, P.; Tu, Y.; et al. L-Theanine and Immunity: A Review. Molecules 2023, 28, 3846. [Google Scholar] [CrossRef]
- Zhu, G.; Yang, S.; Xie, Z.; Wan, X. Synaptic modification by L-theanine, a natural constituent in green tea, rescues the impairment of hippocampal long-term potentiation and memory in AD mice. Neuropharmacology 2018, 138, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.; McKune, A.J.; Georgousopoulou, E.N.; Kellett, J.; D’Cunha, N.M.; Sergi, D.; Mellor, D.; Naumovski, N. The Effect of L-Theanine Incorporated in a Functional Food Product (Mango Sorbet) on Physiological Responses in Healthy Males: A Pilot Randomised Controlled Trial. Foods 2020, 9, 371. [Google Scholar] [CrossRef]
- Li, M.Y.; Liu, H.Y.; Wu, D.T.; Kenaan, A.; Geng, F.; Li, H.B.; Gunaratne, A.; Li, H.; Gan, R.Y. L-Theanine: A Unique Functional Amino Acid in Tea (Camellia sinensis L.) With Multiple Health Benefits and Food Applications. Front Nutr. 2022, 9, 853846. [Google Scholar] [CrossRef] [PubMed]
- Attia, S. Modulation of irinotecan-induced genomic DNA damage by theanine. Food and Chemical Toxicology 2012, 50, 1749–1754. [Google Scholar] [CrossRef] [PubMed]
- Romesberg, A.; Van Houten, B. Targeting Mitochondrial Function with Chemoptogenetics. Biomedicines 2022, 10, 2459. [Google Scholar] [CrossRef] [PubMed]
- Palikaras, K.; Lionaki, E.; Tavernarakis, N. Coordination of mitophagy and mitochondrial biogenesis during ageing in C. elegans. Nature 2015, 521, 525–528. [Google Scholar] [CrossRef]
- de Boer, R.; Smith, R.L.; De Vos, W.H.; Manders, E.M.M.; van der Spek, H. In Vivo Visualization and Quantification of Mitochondrial Morphology in C. elegans. Methods Mol Biol. 2021, 2276, 397–407. [Google Scholar]
- Leung, M.C.; Rooney, J.P.; Ryde, I.T.; Bernal, A.J.; Bess, A.S.; Crocker, T.L.; Ji, A.Q.; Meyer, J.N. Effects of early life exposure to ultraviolet C radiation on mitochondrial DNA content, transcription, ATP production, and oxygen consumption in developing Caenorhabditis elegans. BMC Pharmacol Toxicol. 2013, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Haroon, S.; Li, A.; Weinert, J.L.; Fritsch, C.; Ericson, N.G.; Alexander-Floyd, J.; Braeckman, B.P.; Haynes, C.M.; Bielas, J.H.; Gidalevitz, T.; Vermulst, M. Multiple Molecular Mechanisms Rescue mtDNA Disease in C. elegans. Cell Rep. 2018, 22, 3115–3125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhao, Y.; Yu, M.; Qin, J.; Ye, B.; Wang, Q. Mitochondrial Dysfunction and Chronic Liver Disease. Curr. Issues Mol. Biol. 2022, 44, 3156–3165. [Google Scholar] [CrossRef] [PubMed]
- Dua, N.; Seshadri, A.; Badrinarayanan, A. DarT-mediated mtDNA damage induces dynamic reorganization and selective segregation of mitochondria. J Cell Biol. 2022, 221, e202205104. [Google Scholar] [CrossRef]
- Shokolenko, I.N.; Wilson, G.L.; Alexeyev, M.F. Persistent damage induces mitochondrial DNA degradation. DNA Repair 2013, 12, 488–499. [Google Scholar] [CrossRef] [PubMed]
- White, D. 3rd.; Yang, Q. Genetically Encoded ATP Biosensors for Direct Monitoring of Cellular ATP Dynamics. Cells 2022, 11, 1920. [Google Scholar] [CrossRef]
- Fernandez-Vizarra, E.; Zeviani, M. Mitochondrial disorders of the OXPHOS system. FEBS Lett. 2021, 595, 1062–1106. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, C.; Feng, C.; Yan, C.; Yu, Y.; Chen, Z.; Guo, C.; Wang, X. Role of mitochondrial reactive oxygen species in homeostasis regulation. Redox Rep. 2022, 27, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bai, K.W.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin attenuates hepatic mitochondrial dysfunction through the maintenance of thiol pool, inhibition of mtDNA damage, and stimulation of the mitochondrial thioredoxin system in heat-stressed broilers. J Anim Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef]
- Peng, K.; Tao, Y.; Zhang, J.; Wang, J.; Ye, F.; Dan, G.; Zhao, Y.; Cai, Y.; Zhao, J.; Wu, Q.; Zou, Z.; Cao, J.; Sai, Y. Resveratrol Regulates Mitochondrial Biogenesis and Fission/Fusion to Attenuate Rotenone-Induced Neurotoxicity. Oxid Med Cell Longev. 2016, 2016, 6705621. [Google Scholar] [CrossRef]
- de la Asunción, J.G.; Del Olmo, M.L.; Gómez-Cambronero, L.G.; Sastre, J.; Pallardó, F.V.; Viña, J. AZT induces oxidative damage to cardiac mitochondria: protective effect of vitamins C and E. Life Sci. 2004, 76, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Zhong, L.; Jiang, L.; Geng, C.; Yao, X.; Cao, J. Phellinus linteus mushroom protects against tacrine-induced mitochondrial impairment and oxidative stress in HepG2 cells. Phytomedicine 2013, 20, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Reddyvari, H.; Govatati, S.; Matha, S.K.; Korla, S.V.; Malempati, S.; Pasupuleti, S.R.; Bhanoori, M.; Nallanchakravarthula, V. Therapeutic effect of green tea extract on alcohol induced hepatic mitochondrial DNA damage in albino wistar rats. J Adv Res. 2017, 8, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.; Schurman, S.H.; Harboe, C.; de Souza-Pinto, N.C.; Bohr, V.A. Base excision repair of oxidative DNA damage and association with cancer and aging. Carcinogenesis 2009, 30, 2–10. [Google Scholar] [CrossRef]
- Fu W, Liu Y, Yin H. Mitochondrial Dynamics: Biogenesis, Fission, Fusion, and Mitophagy in the Regulation of Stem Cell Behaviors. Stem Cells Int. 2019, 2019, 9757201. [Google Scholar]
- Breckenridge, D.G.; Kang, B.H.; Kokel, D.; Mitani, S.; Staehelin, L.A.; Xue, D. Caenorhabditis elegans drp-1 and fis-2 regulate distinct cell-death execution pathways downstream of ced-3 and independent of ced-9. Mol Cell 2008, 31, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Zhang, Z.; Wang, R.; Bo, H.; Zhang, Y. Exercise Improves the Coordination of the Mitochondrial Unfolded Protein Response and Mitophagy in Aging Skeletal Muscle. Life (Basel) 2023, 13, 1006. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Chai, R.; Luan, Y.; Du, Y.; Xue, W.; Shi, S.; Wu, H.; Wei, Y.; Zhang, L.; Hu, Y. Trends in mitochondrial unfolded protein response research from 2004 to 2022: A bibliometric analysis. Front Cell Dev Biol. 2023, 11, 1146963. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.F.; Vander Wende, H.; Simko, M.; Klum, S.; Barfield, S.; Choi, H.; Pineda, V.V.; Kaeberlein, M. Activation of the mitochondrial unfolded protein response does not predict longevity in Caenorhabditis elegans. Nat Commun. 2014, 5, 3483. [Google Scholar] [CrossRef]
- Pinto. M.; Moraes, C.T. Mechanisms linking mtDNA damage and aging. Free Radic Biol Med. 2015, 85, 250–258. [Google Scholar] [CrossRef]
- Wang, Q.; Li, H.; Zhang, G.; Chen, X.; Wang, X. Itaconate prolongs the healthy lifespan by activating UPRmt in Caenorhabditis elegans. Eur J Pharmacol. 2022, 923, 174951. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.A.; Fleming, J.T. Basic Culture Methods. Methods in cell biology 1995, 48, 3–29. [Google Scholar] [PubMed]
- Boyd, W.A.; Crocker, T.L.; Rodriguez, A.M.; Leung, M.C.; Lehmann, D.W.; Freedman, J.H.; Van Houten, B.; Meyer, J.N. Nucleotide excision repair genes are expressed at low levels and are not detectably inducible in Caenorhabditis elegans somatic tissues, but their function is required for normal adult life after UVC exposure. Mutat Res. 2010, 683, 57–67. [Google Scholar] [CrossRef]
- Zhang. L.; Jie, G.; Zhang, J.; Zhao, B. Significant longevity-extending effects of EGCG on Caenorhabditis elegans under stress. Free Radic Biol Med. 2009, 46, 414–421. [Google Scholar] [CrossRef]
- Ermolaeva, M.A.; Segref, A.; Dakhovnik, A.; Ou, H.L.; Schneider, J.I.; Utermöhlen, O.; Hoppe, T.; Schumacher, B. DNA damage in germ cells induces an innate immune response that triggers systemic stress resistance. Nature 2013, 501, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hunt, C.P.; Rooney, J.P.; Ryde, I.T.; Anbalagan, C.; Joglekar, R.; Meyer, J.N. PCR-Based Analysis of Mitochondrial DNA Copy Number, Mitochondrial DNA Damage, and Nuclear DNA Damage. Curr Protoc Toxicol. 2016, 67, 20.11.1–20.11.25. [Google Scholar] [CrossRef]
- Wang, Y.; Gai, T.; Zhang, L.; Chen, L.; Wang, S.; Ye, T.; Zhang, W. Neurotoxicity of bisphenol A exposure on Caenorhabditis elegans induced by disturbance of neurotransmitter and oxidative damage. Ecotoxicol Environ Saf. 2023, 252, 114617. [Google Scholar] [CrossRef]
Figure 1.
Enhancement of mtDNA damage removal in UVC-exposed C. elegans treated by L-theanine. A) L3 arrest increased in a dose-dependent manner following serial UVC exposure. B) Mitochondrial and nuclear DNA damage after UVC exposure for 48 h increased in a dose-dependent manner following serial UVC exposure. C) Percent changes in L3 arrest of UVC-exposed N2 worms treated with various concentrations of L-theanine were shown. D) L-theanine treatment for 48 h reduced the frequency of mtDNA damage caused by UVC exposure. E) L-theanine treatment did not affect L3 arrest of N2 worms. F) L-theanine treatment did not change the body length of N2 worms. Results are means ± SD (results from 3 independent experiments, * P < 0.05, ** P < 0.01).
Figure 1.
Enhancement of mtDNA damage removal in UVC-exposed C. elegans treated by L-theanine. A) L3 arrest increased in a dose-dependent manner following serial UVC exposure. B) Mitochondrial and nuclear DNA damage after UVC exposure for 48 h increased in a dose-dependent manner following serial UVC exposure. C) Percent changes in L3 arrest of UVC-exposed N2 worms treated with various concentrations of L-theanine were shown. D) L-theanine treatment for 48 h reduced the frequency of mtDNA damage caused by UVC exposure. E) L-theanine treatment did not affect L3 arrest of N2 worms. F) L-theanine treatment did not change the body length of N2 worms. Results are means ± SD (results from 3 independent experiments, * P < 0.05, ** P < 0.01).
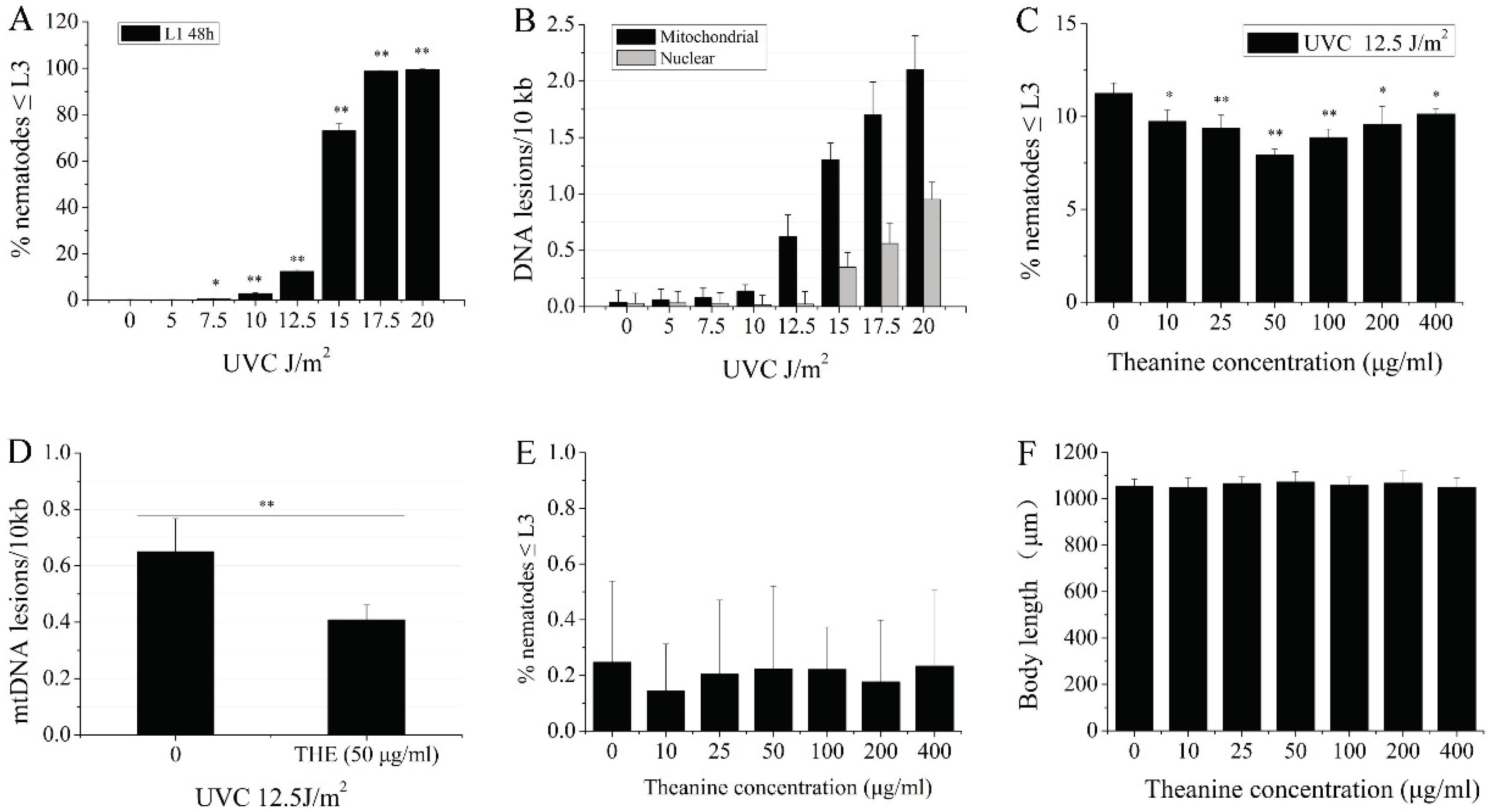
Figure 2.
Effect of L-theaninee treatment on the lifespan of UVC-exposed C. elegans. A) L-theanine treatment significantly extended the lifespan of UVC-exposed nematodes under normal culture condition. B-C) L-theanine treatment for 24 h or 48 h increased the survival rate of nematodes exposed to UVC under heat stress (35℃ for 7 h). Results are means ± SD (3 independent experiments, ** P < 0.01).
Figure 2.
Effect of L-theaninee treatment on the lifespan of UVC-exposed C. elegans. A) L-theanine treatment significantly extended the lifespan of UVC-exposed nematodes under normal culture condition. B-C) L-theanine treatment for 24 h or 48 h increased the survival rate of nematodes exposed to UVC under heat stress (35℃ for 7 h). Results are means ± SD (3 independent experiments, ** P < 0.01).
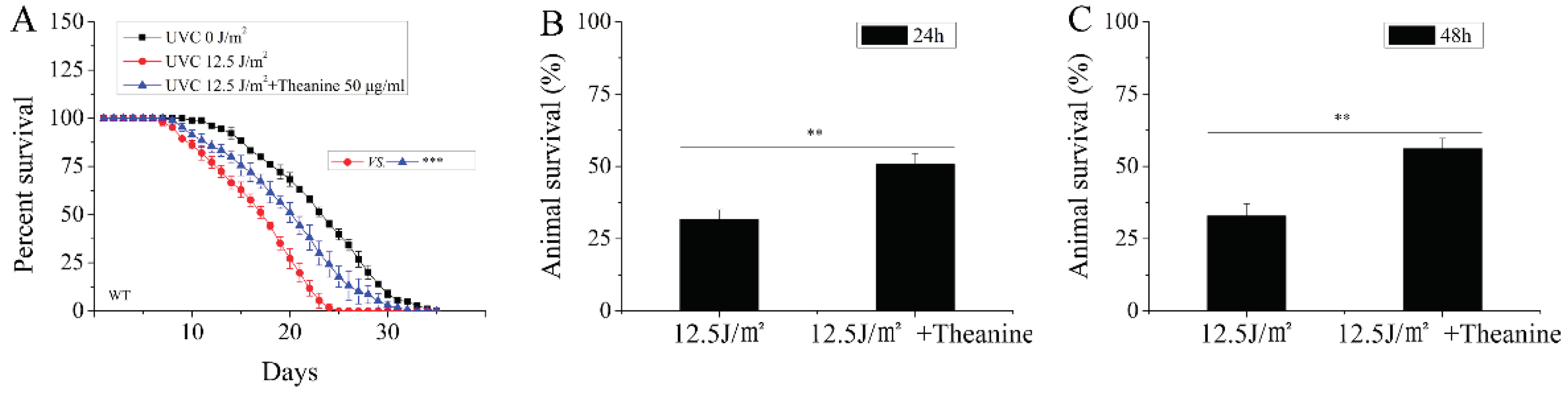
Figure 3.
Improvement of mitochondrial morphology in UVC-exposed C. elegans treated by L-theanine. A) Representative images of transgenic animals expressing myo-3::matrixGFP in body wall muscle cells. B) L-theanine treatment for 2 days did not affect the morphological categories of mitochondria in C. elegans. C-D) L-theanine treatment for 4 and 6 days improved the morphological categories of mitochondria in UVC-exposed C. elegans respectively. Results are means ± SD (at least 3 independent experiments, ** P < 0.01).
Figure 3.
Improvement of mitochondrial morphology in UVC-exposed C. elegans treated by L-theanine. A) Representative images of transgenic animals expressing myo-3::matrixGFP in body wall muscle cells. B) L-theanine treatment for 2 days did not affect the morphological categories of mitochondria in C. elegans. C-D) L-theanine treatment for 4 and 6 days improved the morphological categories of mitochondria in UVC-exposed C. elegans respectively. Results are means ± SD (at least 3 independent experiments, ** P < 0.01).
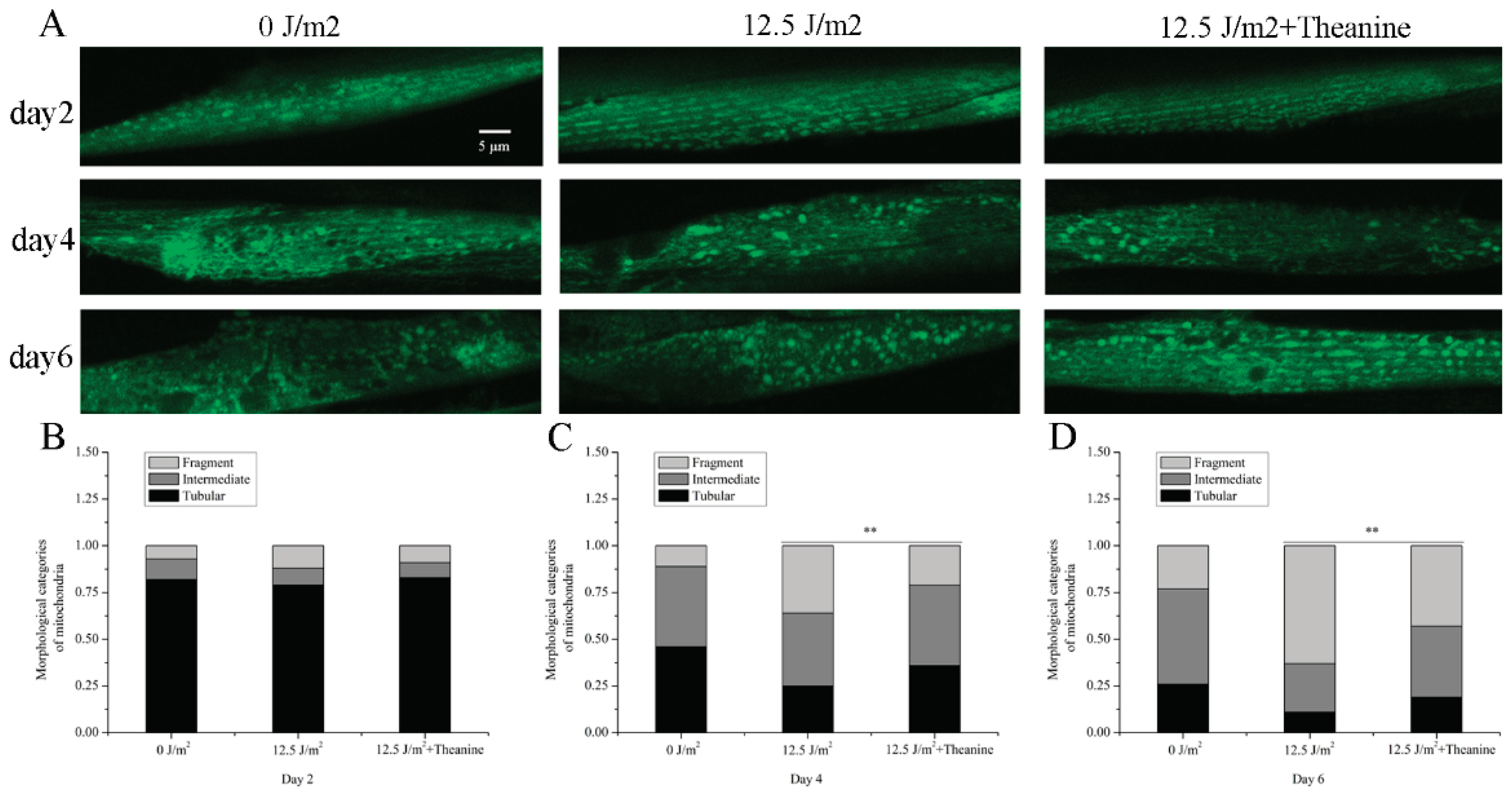
Figure 4.
Increasing steady-state ATP levels in UVC-exposed C. elegans treated by L-theanine. A-F) Representative images of transgenic animals expressing sur-5p::luciferase::GFP in the pharynx were monitored at 24 h and 48h, respectively. G-H) Steady-state ATP levels were significantly increased in UVC-exposed C. elegans treated by L-theanine at 24 h and 48 h, compared with UVC treatment, respectively. Results are means ± SD ( 3 independent experiments, t test, ** P < 0.01).
Figure 4.
Increasing steady-state ATP levels in UVC-exposed C. elegans treated by L-theanine. A-F) Representative images of transgenic animals expressing sur-5p::luciferase::GFP in the pharynx were monitored at 24 h and 48h, respectively. G-H) Steady-state ATP levels were significantly increased in UVC-exposed C. elegans treated by L-theanine at 24 h and 48 h, compared with UVC treatment, respectively. Results are means ± SD ( 3 independent experiments, t test, ** P < 0.01).
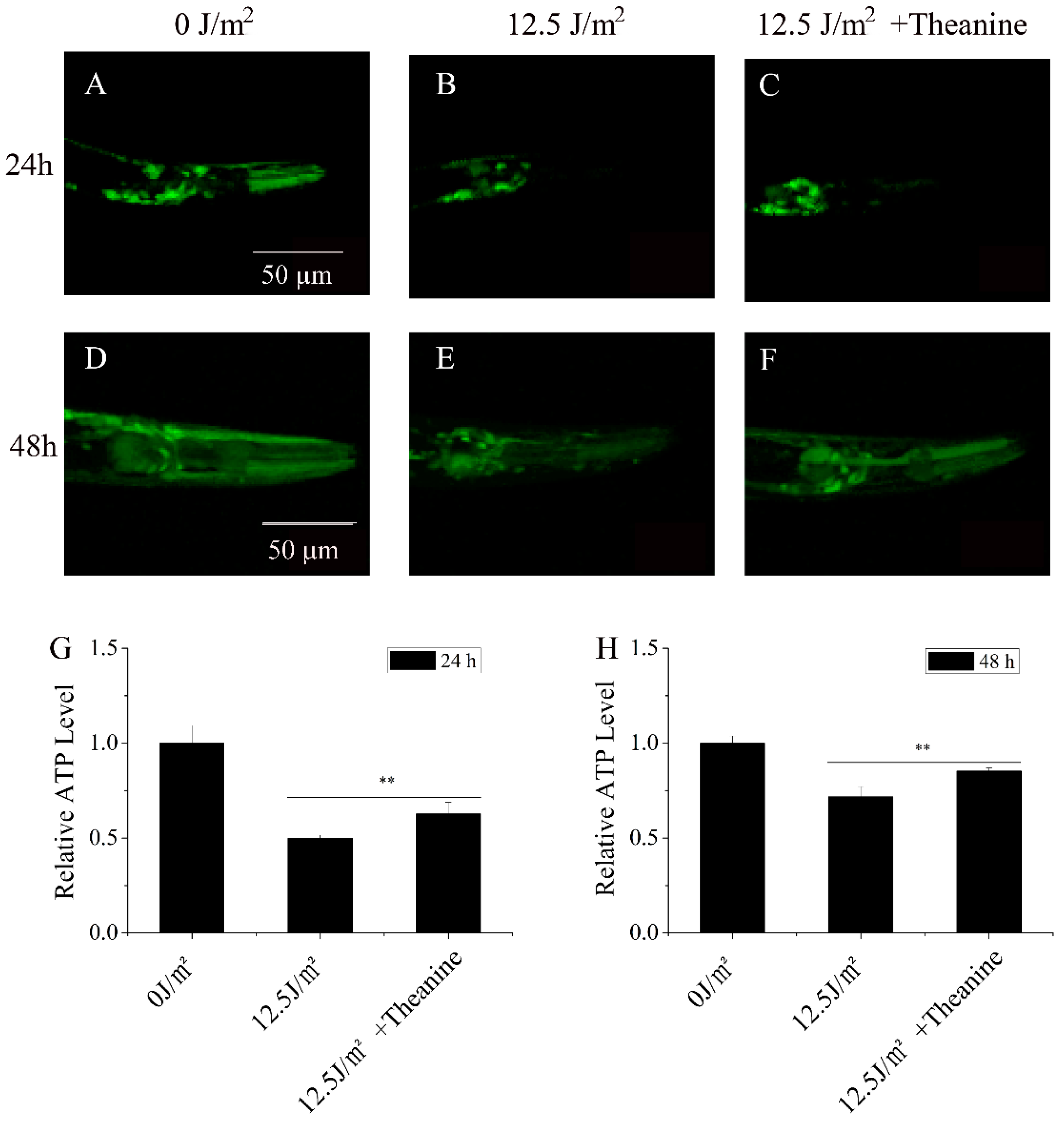
Figure 5.
Effect of L-theanine treatment on the L3 arrest and lifespan of UVC-exposed C. elegans deleted in autophagy, mitophagy, mitochondrial dynamics, and UPRmt genes. A-C) Mutations in autophagy gene (unc-51), mitophagy genes (pink-1 and pdr-1), fusion genes (fzo-1 and eat-3), fission gene (drp-1), and UPRmt genes (atfs-1, haf-1, and ubl-5) did not affect L3 arrest in UVC-exposed C. elegans treated by L-theanine, respectively. D-H) Mutations in mitophagy genes (pink-1 ), fission gene (drp-1), and UPRmt genes (atfs-1 and haf-1) did not affect the lifespan in UVC-exposed C. elegans treated by L-theanine, respectively. Results are means ± SD ( 3 independent experiments, t test, ** P < 0.01).
Figure 5.
Effect of L-theanine treatment on the L3 arrest and lifespan of UVC-exposed C. elegans deleted in autophagy, mitophagy, mitochondrial dynamics, and UPRmt genes. A-C) Mutations in autophagy gene (unc-51), mitophagy genes (pink-1 and pdr-1), fusion genes (fzo-1 and eat-3), fission gene (drp-1), and UPRmt genes (atfs-1, haf-1, and ubl-5) did not affect L3 arrest in UVC-exposed C. elegans treated by L-theanine, respectively. D-H) Mutations in mitophagy genes (pink-1 ), fission gene (drp-1), and UPRmt genes (atfs-1 and haf-1) did not affect the lifespan in UVC-exposed C. elegans treated by L-theanine, respectively. Results are means ± SD ( 3 independent experiments, t test, ** P < 0.01).
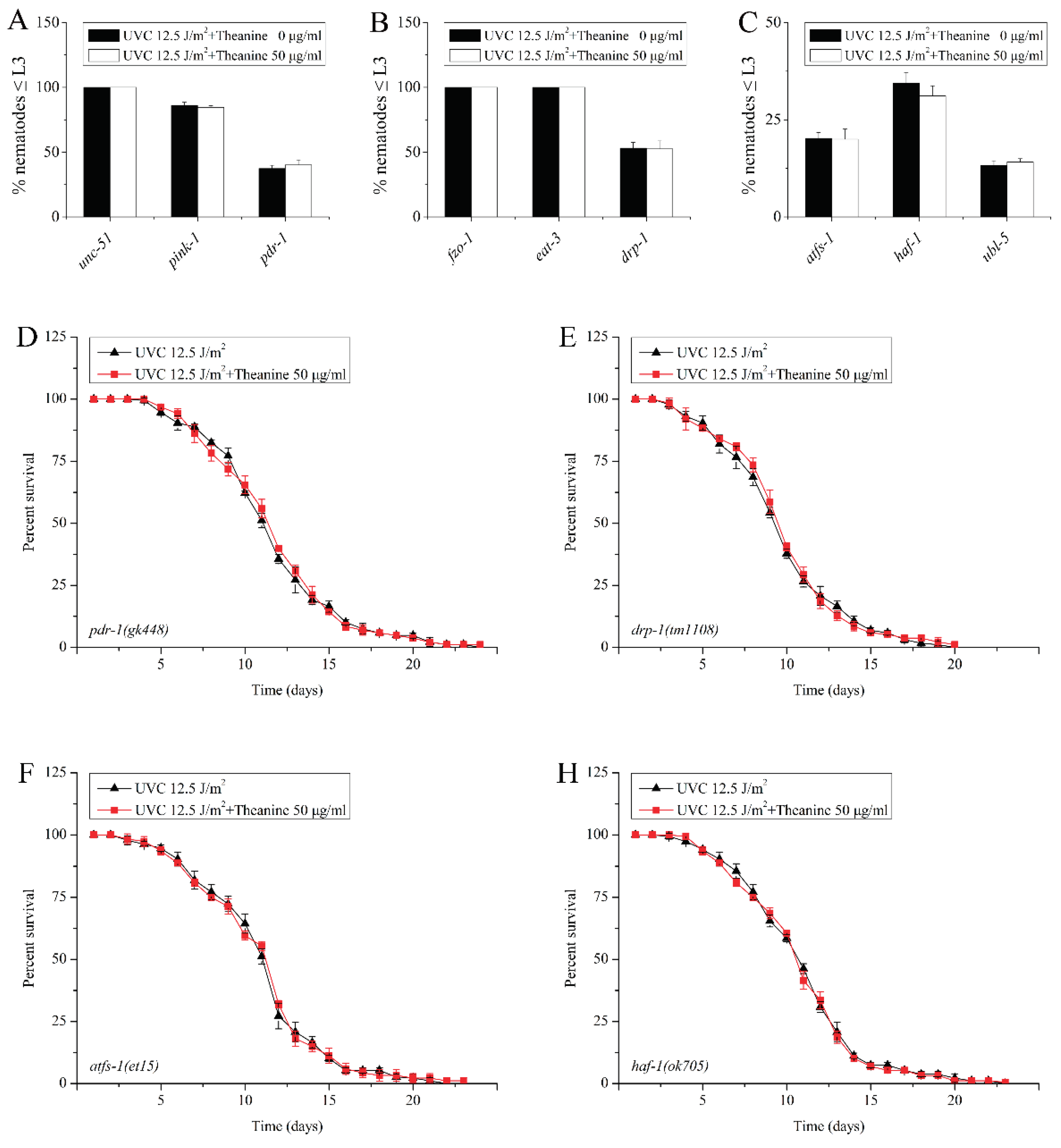
Figure 6.
Effect of L-theanine treatment on the relative expression of autophagy (bec-1, lgg-1, and atg-18), mitophagy (dct-1), and UPRmt (hsp-60) related genes in UVC-exposed C. elegans. L-theanine treatment did not affect lgg-1 and atg-18 mRNA levels in UVC-exposed C. elegans treated by L-theanine, but up-regulated bec-1, dct-1, and hsp-60 expression. Results are means ± SD (3 independent experiments, t test, ** P < 0.01).
Figure 6.
Effect of L-theanine treatment on the relative expression of autophagy (bec-1, lgg-1, and atg-18), mitophagy (dct-1), and UPRmt (hsp-60) related genes in UVC-exposed C. elegans. L-theanine treatment did not affect lgg-1 and atg-18 mRNA levels in UVC-exposed C. elegans treated by L-theanine, but up-regulated bec-1, dct-1, and hsp-60 expression. Results are means ± SD (3 independent experiments, t test, ** P < 0.01).
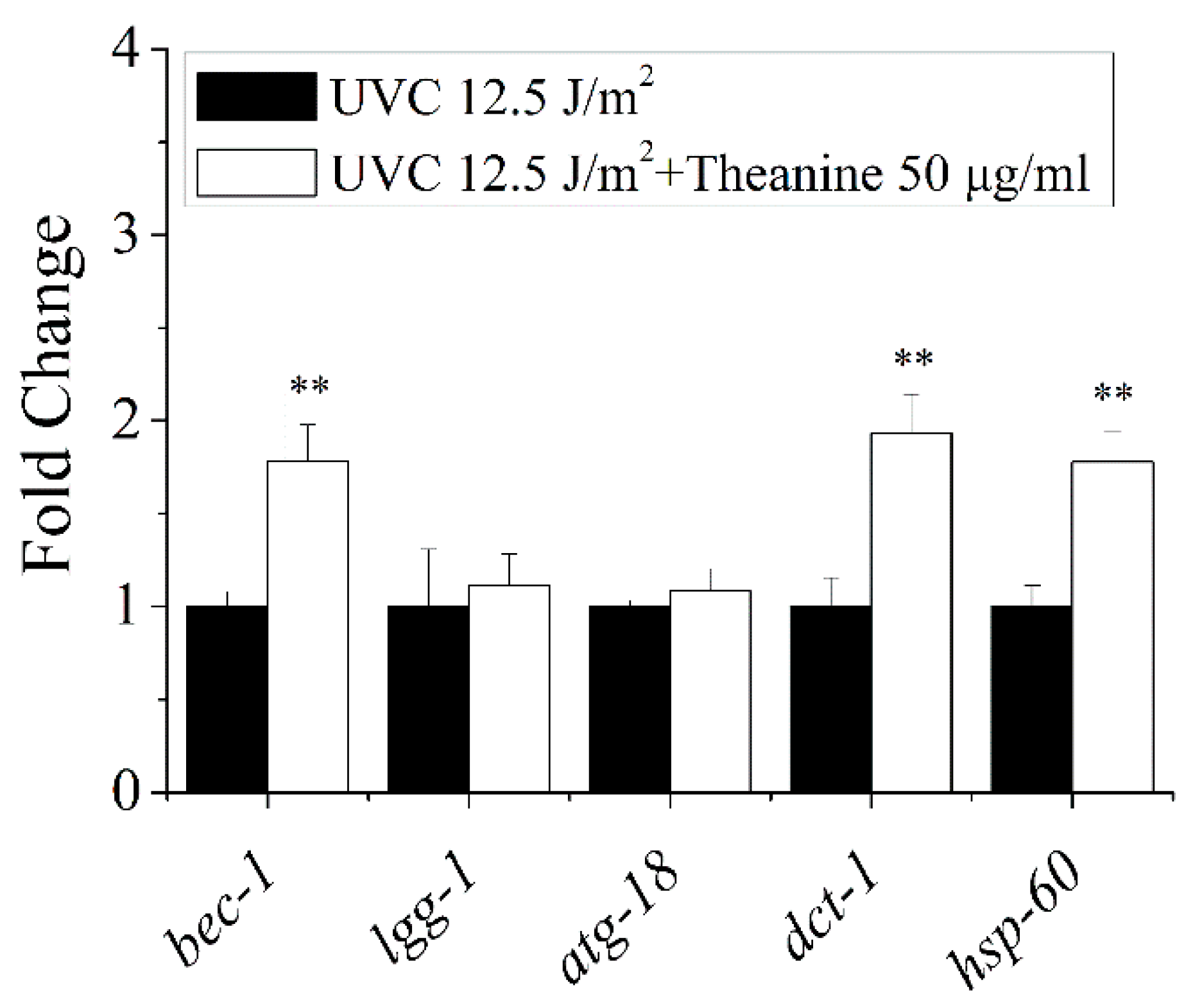
Figure 7.
Schematic diagram of postulated mechanism extending lifespan in UVC-exposed C. elegans treated by L-theanine. Persistent mtDNA damage induced by UVC irradiation causes mitochondrial dysfunction (e.g. reducing ATP levels, increasing larval arrest, and destroying mitochondrial morphology) and shortens the lifespan in C. elegans. L-theanine treatment enhances multiple molecular mechanisms (autophagy, mitochondrial dynamics, and UPRmt) to remove the mtDNA damage induced by UVC irradiation and improve mitochondrial dysfunction (e.g. increasing ATP levels, reducing larval arrest, and improving mitochondrial morphology) in UVC-exposed C. elegans, therefore increasing lifespan of UVC-exposed C. elegans.
Figure 7.
Schematic diagram of postulated mechanism extending lifespan in UVC-exposed C. elegans treated by L-theanine. Persistent mtDNA damage induced by UVC irradiation causes mitochondrial dysfunction (e.g. reducing ATP levels, increasing larval arrest, and destroying mitochondrial morphology) and shortens the lifespan in C. elegans. L-theanine treatment enhances multiple molecular mechanisms (autophagy, mitochondrial dynamics, and UPRmt) to remove the mtDNA damage induced by UVC irradiation and improve mitochondrial dysfunction (e.g. increasing ATP levels, reducing larval arrest, and improving mitochondrial morphology) in UVC-exposed C. elegans, therefore increasing lifespan of UVC-exposed C. elegans.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



