
Submitted:
05 March 2024
Posted:
06 March 2024
You are already at the latest version
Abstract
Populational aging is marked by chronic non-communicable diseases, such as metabolic syndrome (MetS). IL-10 and IL-1β are pleiotropic cytokines with multiple biological effects linked to metabolic disorders. This cross-sectional study assessed 193 participants' IL-10 and IL-1β serum levels regarding their role in developing MetS, clinical characteristics, and their IL1B rs1143627 and IL10 rs1800890 variants' genotype frequencies in a population over 60. IL-10 levels correlated weakly with HDL levels and fat mass and inversely with triglycerides, glucose, glycated hemoglobin, and estimated average blood glucose levels. IL-10 levels were also indirectly influenced by the patient's T2DM duration, lean mass amount, and bone mineral content. Participants with altered HDL, elevated serum glucose, raised HbA1c levels, or those over 80 had reduced serum IL-10 levels compared to those with normal levels or other age groups, respectively. Women also had higher serum IL-10 levels than men. Dissimilarity, IL-1β levels correlated directly only with the number of total leukocytes and segmented neutrophils, showing only significant variations with self-reported alcohol consumption. Our study also found that those with the IL10 AA genotype (lower IL-10 levels) had a significantly higher risk of developing MetS. These findings may help direct future research and more targeted therapeutic approaches in older adults.
Keywords:
Metabolic Syndrome
; Interlekine-10
; Interlekine-1β
; older population
; IL10 rs1800890
; IL1B rs1143627
1. Introduction
Population aging, or demographic transition, occurs due to a simultaneous decline in mortality and the birth rate or fertility. The Brazilian Institute of Geography and Statistics (IBGE) predicts that in 2030, Brazil will have around 40 million people aged 60 or over, surpassing the total number of children between zero and 14 years old [1,2]. This demographic transition leads to an epidemiological transition (a long-term shift in mortality and disease [3]) marked by a loss of physiological reserves and an emergence of chronic noncommunicable diseases (NCDs) that commonly come with the aging process. Consequently, aging is a risk factor for NCDs, such as metabolic syndrome (MetS), in which increased age is related to higher MetS prevalence [4,5,6].
MetS, whose base is insulin resistance, comprises a set of metabolic dysregulations, neurohormonal activations, and risk factors that manifest in an individual, increasing their risk of developing cardiovascular diseases (CVDs) and other NCDs, such as Type 2 Diabetes Mellitus (T2DM), stroke/cerebrovascular accident (CVA), and acute myocardial infarction (AMI), and even, eventually, neurological diseases [5,7,8,9,10]. Its clinical diagnosis can be defined by the presence of three or more criteria from the National Cholesterol Education Program’s Adult Treatment Panel III (NCEP-ATP III), which include increased abdominal (central) obesity, hypertension, elevated fasting blood glucose levels (hyperglycemia/dysglycemia), and atherogenic dyslipidemia, such as elevated serum triglyceride levels and reduced serum high-density lipoproteins (HDL) [5,7,8].
MetS is a low-grade (sub-clinical) chronic inflammatory disease whose pathogenesis encompasses multiple genetic, epigenetic, and acquired commodities interacting through several complex mechanisms yet to be fully elucidated [7,9]. If left untreated, the progressive increase in insulin-resistant adipose tissue, particularly visceral fat, impairs insulin-mediated inhibition of lipolysis and, thus, increases circulating free fatty acids (FFAs), which promotes gluconeogenesis and lipogenesis in the liver that ultimately results in atherogenic dyslipidemia [7,9]. Insulin resistance can also reduce the blood supply to adipocytes with consequent hypoxia partly due to the loss of insulin’s vasodilatory effect, FFA-induced vasoconstriction, and higher serum viscosity, creating a prothrombotic state that, together with increased leptins levels, increases inflammatory cytokines release from the adipose tissue [7,11].
Interleukins mediate and regulate inflammatory reactions and may differ in their immunological function. Among them, two stand out: interleukin IL-1β, a proinflammatory cytokine, and interleukin IL-10, an anti-inflammatory cytokine, both pleiotropic cytokines with multiple biological effects [7,11,12,13,14]. IL-10 is linked to metabolism as it can maintain insulin sensitivity, reduce glucose intolerance, increase whole-body lipid synthesis and skeletal muscle glycolysis, and decrease intramuscular fatty acyl-CoA levels [14,15,16,17,18,19,20,21]. Furthermore, alterations in cellular metabolism and specific metabolites, such as FFAs from the adipose tissue, regulate IL-10 production in different immune cells [19,22]. Likewise, IL-1β plays a critical role in physiological and pathological metabolism modulation (energy homeostasis), including insulin action and secretion, β-cell apoptosis, lipid metabolism, and food intake, by regulating the immune system and the neuronal and endocrine systems’ interface and, hence, in metabolic disorders, such as T2DM, MetS, and obesity [14,23,24,25,26,27,28].
Functional polymorphisms can also modify their genes’ expression and production, thus affecting their function. Both IL10 and IL1B cytokine genes are highly polymorphic [29,30]. IL1B rs1143627 (–31T>C) single nucleotide polymorphism (SNP) located in the gene’s promoter region, for instance, disrupts a TATA-box motif, markedly affecting several transcription factors’ binding affinity and thereby impacting IL1B transcription activity [29]. Another example, also located in its gene’s promoter region, is IL10 rs1800890 (-3575T>A) SNP polymorphism that affects IL-10 production levels [14,30,31]. Both polymorphisms could either protect or predispose an individual to MetS.
As various pathogenic pathways contributing to MetS development culminate in an inflammatory state, this study assessed the serum levels of IL-10 and IL-1β cytokines regarding their role in the susceptibility to developing MetS and other clinical characteristics in an older population (over 60). The study also compared the genotype frequencies of IL1B rs1143627 (–31T>C) and IL10 rs1800890 (-3575T>A) variants in these research participants to their respective serum concentrations.
2. Materials and Methods
2.1. Research Participants
This study employed a cross-sectional design with both quantitative and qualitative approaches to observe, analyze, and describe patients treated at a Basic Health Unit (UBS) in the Ceilândia Administrative Region of the Federal District—Brazil. By 2016, this region had the lowest family health coverage of the DF, with about 22% [32]. The sample consisted of older adults who voluntarily agreed to participate in the study after being invited. All participants were registered and received care from the Family Health Strategy teams at UBS No. 6 in the Ceilândia Region. A total of 193 participants were selected, taking into account a 5% margin of error and a 95% confidence level.
The inclusion criteria required participants to be 60 or older, registered with primary care services, monitored in the pre-selected unit, and capable of understanding, verbalizing, and answering the proposed questions. Concurrently, the study excluded participants if they were diagnosed with mental or psychiatric illness, undergoing cancer treatment, had cardiac (heart) surgery within the past six months, had a pacemaker or metal prosthesis, were taking hormone replacement therapy, or were taking vitamins or supplements that could interfere with the measurement of biochemical or cytokine blood levels.
Recruited elders signed the Informed Consent Form (ICF) after being fully informed about the stages of data collection. A nursing appointment was scheduled, during which the identification form was filled out with data such as weight, height, and health and socioeconomic factors. Participants also signed the Biological Material Custody Agreement, and their venous blood was collected.
This study was approved by the Research Ethics Committee (CEP) of the Health Sciences Teaching and Research Foundation (FEPECS) of the Federal District State Health Department (SES-DF) under opinion number 1.355.211/2017 with CAAE 59071116.8.0000.0030.
2.2. Sample Collection
Samples were collected from February to June 2019. The data collection process occurred during the nursing appointment at the UBS and consisted of two stages: blood collection and data collection. During the first stage, participants fasted for 8 to 12 hours before providing approximately 15 mL of venous blood from the cubital fossa region. Subsequently, they received breakfast.
In the second stage, a structured data collection questionnaire was applied to evaluate the participants’ demographic and socioeconomic profiles. The questionnaire consisted of closed questions regarding biological sex, age, education, retirement, income, lifestyle habits (smoking), clinical conditions (hypertension or diabetes), diagnosis of T2DM, and use of medications (type and route of administration).
2.3. Metabolic Syndrome Defining Condition
The Brazilian Cardiological Society (BCS) criteria used to define metabolic syndrome (MetS) include waist circumference (WC) >88 cm (female) and >102 cm (male); blood pressure (BP) ≥130 x 85 mmHg or use of antihypertensive medication; fasting blood glucose ≥110 mg/dL or use of T2DM medications; triglycerides (TG) ≥150 mg/dL or use of medications for dyslipidemia; and HDL <50 mg/dL (female) and <40 mg/dL (male); and requires for a MetS diagnosis the presence of at least three criteria. These criteria are based on the National Cholesterol Evaluation Program for Adult Treatment Panel III (NCEP-ATP III) and the Brazilian Guideline for Diagnosis and Treatment of Metabolic Syndrome, which does not require insulin measurement but allows for it to be used as a clinical criterion [5].
2.4. DNA Extraction
DNA was extracted from peripheral blood using the PureLink® Genomic DNA Mini Kit from Invitrogen (catalog #K1820-02, lot #19339891), following the manufacturer’s protocol, with an average yield of 20 ng/µL. The DNA was then stored at -70°C until further use.
2.5. Genotyping
After DNA extraction, a diluted sample underwent Polymorphism Polymerase Chain Reaction (PCR) using the Techne Thermal Cycler model TC-512 to study the distribution of single nucleotide polymorphisms (SNPs). The oligonucleotide sequences used to assess the polymorphisms were sense 5’—TAGTCCCCTCCCCTAAGAAGC—3’ and antisense 5’—CCCAGAATATTTCCCGAGTCA—3’ for IL1B −31T>C (rs1143627) SNP, and sense 5’—GGTTTTCCTTCATTTGCAGC—3’ and antisense 5’—ACACTGTGAGCTTCTTGAGG—3’ for IL10 -3575T>A (rs1800890) SNP.
Thermocycling conditions for the IL1B rs1143627 polymorphism were as follows: initial denaturation at 96˚C for 1 minute, followed by 35 cycles of denaturation at 94˚C for 50 seconds, annealing of the oligonucleotides at 50°C for 1 minute, and extension of the fragments at 72°C for 40 seconds for the extension of the fragments. The final extension was performed at 72°C for 10 minutes and cooled at 4°C for 60 minutes. Each reaction used 4.0 µL of genomic DNA at a final concentration of 2.5 ng/µL; 2.5 µL of 10x buffer (10 mM Tris and 50 mM KCl); 0.5 µL of 50 mM MgCl2 (Ludwig Biotec, Alvorada, Rio Grande do Sul, Brazil); 0.5 µL of 2.5 mM deoxyribonucleotide triphosphates (dNTPs) (Ludwig Biotec, Alvorada, Rio Grande do Sul, Brazil); 0.5 µL of 5 U/µL Taq-Polymerase, (Ludwig Biotec, Alvorada, Rio Grande do Sul, Brazil); 1.5 µL of each forward and reverse oligonucleotide (10 µM, IDT Technologies); completing with ultrapure water to a final volume of 25 µL per reaction.
The IL1B PCR product, a 421 bp fragment, was digested with the AluI restriction enzyme (New England Biolabs, Inc. Beverly, MA, USA). Allele 1 (C) creates two restriction sites, resulting in three cleavage fragments: 344, 57bp, and 20 bp, while allele 2 (T) is cleaved into four fragments (three restriction sites): 247, 97, 57 and 20 bp. This digestion system was assembled using 10.0 µL of PCR product, 2.0 µL of 10x NEB4 buffer (Biolabs), 1 µL of 10U/µL AluI enzyme, topped up with ultrapure water for a final volume of 20 µL per reaction. The system was maintained at 37°C for 3 hours.
For the IL10 rs1800890, the thermocycling conditions were 94˚C for 5 minutes (initial denaturation), followed by 35 denaturation cycles at 94˚C for 30 seconds, oligonucleotide annealing at 62˚C for 45 seconds and 72˚C per 55 seconds for the fragments’ extension. The final extension was performed at 72˚C for 8 minutes and then cooled for 4 minutes. Each reaction used 4.0 µl of genomic DNA at the final concentration of 2.5 ng/µL; 2.5 µl of 10x buffer (10 mm of Tris and 50 mm of KCl); 0.5 µL of 50 mM MgCl2 (Ludwig Biotec, Alvorada, Rio Grande do Sul, Brazil); 0.5 µL of 2.5 mM deoxyribonucleotide triphosphates (dNTPs) (Ludwig Biotec, Alvorada, Rio Grande do Sul, Brazil); 0.5 µL of 5 U/µL Taq-Polymerase, (Ludwig Biotec, Alvorada, Rio Grande do Sul, Brazil); 1.5 µL of each forward and reverse oligonucleotide (10 µM, IDT Technologies); topping with ultrapure water to a final volume of 25 µL per reaction.
The IL10 PCR product, a 228pb fragment, was digested using the ApoI enzyme (Thermo Fisher®). Allele 1 (T) creates a new restriction site, cleaving it in two fragments of 121pb and 107pb, while allele 2 (A) remains uncleaved by the ApoI enzyme. Thus, the IL10 rs1800890 SNP is divided into cleavage genotype (TT), heterozygote (TA), and non-cleavage genotype (AA). This digestion system was assembled using 10.0 μL of PCR product, 5μL of 10x UB buffer, and 1 μL of 10U/µL ApoI enzyme, completing with ultrapure water for a final volume of 50 μL per reaction. The system was kept at 37°C for 3 hours.
IL10 and IL1B digestion products underwent an electrophoretic run in a 3% agarose gel stained with 0.1% ethidium bromide at 100W power for 20 minutes.
2.6. Laboratory Analysis and Cytokine Dosage
Biochemical and Hematological tests were performed at a Federal District’s private clinical analysis laboratory funded by the research project budget, whereas IL-10 and IL-1β measurements were accomplished at our university’s clinical analysis laboratory. These measurements were carried out in triplicate using the Enzyme-Linked Immunosorbent Assay (ELISA) method, according to the specifications of the R&D Systems Quantikine® high-sensitivity ELISA kit. The sensitivity and the intraassay coefficient of variation (CV), i.e., repeatability, for these ELISA, were also determined. The control triplicate’s average determined the cutoff point, thus assuming a value of 2.0 pg/ml for IL-1β and 1.0 pg/ml for IL-10.
2.7. Statistical Analysis
For statistical analysis, absolute and relative frequency distribution was applied for categorical variables and quartiles for continuous variables, with continuous data expressed as percentiles. The study utilized Spearman’s rank correlation coefficient to examine the correlation between continuous data on anthropometric, biochemical, and immunological measurements and cytokine dosage levels. For clinical characteristics expressed as categorical data, the non-parametric statistic Kruskall-Wallis H test was used to evaluate the difference between groups’ serum cytokine levels, as the normality assumptions were unmet. The Chi-Square Test of Independence was used to determine whether there was an association between categorical variables (MetS presence versus polymorphism). Tests were conducted using SPSS software (version 28.0, SPSS Inc., Chicago, IL, USA) at a 5.0% significance level.
3. Results
3.1. IL-10 and IL-1β Serum Concentrations’ Associations with Hematological, Biochemical, and Anthropometric Measurements
Correlation analysis revealed that serum IL-10 levels weakly correlated with serum HDL cholesterol levels and fat mass. Serum IL-10 levels were also inversely associated with serum levels of triglycerides, glucose, glycated hemoglobin (HbA1c), and estimated average blood glucose. The cytokine was also indirectly influenced by three variables: the time with T2DM, the amount of lean mass, and the amount of bone mineral content. On the other hand, serum IL-1β levels correlated directly with two variables: total leukocytes and the number of segmented neutrophils (P< 0.05; Table 1).
3.2. Participants’ Serum IL-10 and IL-1β Levels and Other Clinical Signs and Symptoms
Statistically significant differences were observed between the groups with and without metabolic syndrome (MetS) regarding the research participants’ clinical status and serum IL-10 concentrations (serum cytokine levels were reduced in participants with MetS). For instance, participants over 80 exhibited a decrease in IL-10 concentration compared to the rest. Similarly, participants with altered HDL levels showed reduced serum IL-10 levels compared to the others. Participants with elevated serum glucose and HbA1c levels also exhibited lower IL-10 concentrations than those with normal levels. Regarding biological sex, women had higher IL-10 concentrations in their blood compared to men. As for serum IL-1β concentrations, the only variable that showed a statistically significant difference was alcohol consumption. Surprisingly, participants who consumed alcoholic beverages showed lower variations in serum IL-1β levels compared to those who did not (Table 2).
3.3. IL-10 and IL-1β Gene Polymorphisms’ Genotype Frequency Distribution and Their Relationship with Their Serum Concentrations
This study investigated the IL10 rs1800890 (-3575T>A) gene variant and its effect on serum IL-10 concentrations in older adults (over 60). We found that the different genotypes were associated with significant changes in serum IL-10 levels. Specifically, the presence of the ancestral genotype (TT) (56.5%) correlated with a slight increase in serum IL-10 levels compared to other genotypes (p<0.001).
Similarly, IL-1β rs1143627 (–31T>C) polymorphism also affected serum IL-1β levels. The presence of the Polymorphic Genotype (TT) (7.8%) correlated with an increase in serum IL-1β concentration compared to other genotypes (p<0.001) (Table 3).
When analyzing the IL10 -3575T>A (rs1800890) and IL1Β -31 T>C (rs1143627) SNPs’ genotypes according to whether the participants had (MetS+) or not (MetS-) metabolic syndrome, only IL10 -3575T>A (rs1800890) presented a statistically significant difference (P<0.01), with the AA genotype has been identified only in patients with metabolic syndrome (Figure 1).
4. Discussion
Our study’s results analysis revealed a series of correlations between IL-10 and IL-1β serum levels and several clinical variables, as well as with IL10 -3575T>A (rs1800890) and IL1Β -31 T>C (rs1143627) single nucleotide polymorphisms (SNPs) in older adults (over 60) with or without metabolic syndrome. These correlations offer valuable insights into the role of IL-10 and IL-1β in the disease and the aging process, although they require a cautious approach due to the complexity of the subject.
As people age, the likelihood of developing conditions such as obesity, hypertension, and other chronic noncommunicable diseases (NCDs) increases [3,12]. Our studied population covers a range of age groups over 60, with a higher prevalence of people between 60 and 65. This range allows for a deeper analysis of the aging process and variables analyzed. For example, people above 80 had lower levels of IL-10 than the rest of the groups. Notably, some participants already had chronic health conditions such as Type 2 Diabetes Mellitus (T2DM), systemic arterial hypertension (SAH), fibromyalgia, and other inflammatory diseases. Some even had multiple diseases, which is crucial to explore since the interactions between these conditions can affect the correlations found.
Metabolic syndrome (MetS) encompasses several metabolic dysregulations, including insulin resistance, atherogenic dyslipidemia, central obesity, T2DM, and hypertension [6,7,9,10,13]. Its development results from a complex interaction between genetic, epigenetic, and environmental factors [7,9,13]. Furthermore, obese women with T2DM showed an increase in the IL-10 expression and secretion in adipose tissue compared to other groups, a difference that has not been observed in men [33]. Nevertheless, another study associated low circulating IL-10 levels with MetS in obese women [17]. Our study also noticed a higher IL-10 concentration in women and an inverse correlation with the patients’ time with T2DM (T2DM duration).
Our study reveals positive and inverse correlations between serum IL-10 levels and clinical markers. Specifically, a remarkable positive correlation exists between IL-10, serum HDL levels, and fat mass. Previous studies identified significant associations between IL-10 levels and dyslipidemia; IL-10 levels have been inversely correlated with serum levels of total cholesterol, triglycerides, LDL, and VLDL and positively correlated with HDL levels [34]. Obese children with hypertriglyceridemia also presented decreased IL-10 expression in their serum and adipose tissue [18]. Moraitis et al. [35] observed an inverse relationship between IL-10 and HDL levels, indicating that IL-10 can reduce HDL and LDL cholesterol levels while increasing triglyceride levels. Similarly, Ribeiro et al. [36] found an inverse association between increased fat mass in older women and decreased serum HDL concentrations. The authors also noted that IL-10 strongly influenced body composition, as it correlated positively with waist circumference, waist-to-hip circumference ratio, and waist-to-height ratio [36]. These findings highlight the complexity of IL-10’s role in lipid and adipose regulation.
IL-10 does not directly affect human adipocyte function [37]. However, it appears to regulate lipid metabolism and inflammation in adipose tissue [37]. Acosta et al. [37] reported that proinflammatory macrophages are the ones that primarily produce IL-10 in obese and insulin-resistant individuals. Additionally, Blüher et al. [38] found that IL-10 levels are lower in individuals with impaired glucose tolerance or T2DM than those with normal glucose tolerance. We also observed that IL-10 was indirectly correlated with triglycerides, glucose, HbA1c, and estimated mean blood glucose. Collectively, these findings suggest that multiple factors may mediate complex associations and merit further investigation.
The statistically significant difference in IL-10 levels between the groups with and without metabolic syndrome suggests a possible association with this condition. However, the clinical relevance of these findings must be carefully considered, as several variables besides IL-10 influence metabolic syndrome. Low IL-10 levels are associated with metabolic syndrome, and obesity is correlated with increased levels of this cytokine [17]. In a longitudinal study of Iranian adults, a decrease in IL-10 level was observed in participants with persistent metabolic syndrome [39]. Patients with metabolic syndrome have a distinct inflammatory pattern, as the syndrome affects the expression of proinflammatory genes in adipose tissue and the circulating cytokine levels [40].
Like IL-10 implications in metabolic syndrome, IL-1β plays a role in this intricate scenario. IL-1β is associated with low-grade chronic inflammatory states, and an increase in its serum concentration often results from chronic diseases, adipose tissue inflammation, and oxidative stress [41]. Although its role, like IL-10, goes further than inflammation into the realms of a critical metabolic regulator [26], we only found its classic role in leukocyte migration.
Our study found direct correlations between IL-1β serum levels and the number of total leukocytes and segmented neutrophils. These correlations confirm IL-1β’s pivotal role in mediating the immune response and systemic inflammation, which frequently involve increased leukocyte and neutrophil counts—particularly in response to chronic inflammation. IL-1β, produced primarily by macrophages but also by other cells, including neutrophils, can induce the expression of other proinflammatory cytokines [42,43]. This expression, in turn, increases leukocyte adhesion to the endothelium and promotes leukocyte chemotaxis, increasing leukocyte and neutrophil counts at sites of inflammation [42,43]. Several other factors can notably influence the relationship between IL-1β and leukocyte and neutrophil counts, including inflammation type and duration, other cytokines and growth factors, and individual patient characteristics [42,44,45].
The discovery of significant differences in serum IL-1β levels based on alcohol consumption is intriguing. It suggests that alcohol consumption may influence a reduction in IL-1β production or release in the body by triggering inflammatory responses and raises questions about the role of alcohol in modulating systemic inflammation in older adults. In literature, chronic alcohol consumption can have substantial impacts on the immune system, leading to changes in inflammatory cells and triggering dysfunctional immune responses (inability to respond effectively to threats), which, in turn, are associated with various pathologies and an increased susceptibility to infections [46,47,48]. Our findings go against this. However, it is worth remembering that alcohol consumption in our study was self-reported.
Functional gene polymorphisms, such as IL10 -3575T>A (rs1800890) and IL1B -31 T>C (rs1143627) single nucleotide polymorphisms (SNPs), also affect these cytokines’ serum levels [14,29,30,31]. Our study likewise associated different IL10 -3575T>A (rs1800890) and IL1B -31 T>C (rs1143627) SNP genotypes with their serum levels, confirming the evidence of these cytokine’s genetic regulation. We found that the IL10 ancestral genotype (TT) correlated with IL-10 higher levels. At the same time, the IL1B mutated genotype (TT) correlated with higher IL-1β concentrations. However, only the IL10 -3575T>A (rs1800890) SNP genotypes correlated significantly with metabolic syndrome, with the AA genotype conferring a higher risk of having metabolic syndrome. This correlation makes sense as our study with older adults (over 60) found that higher IL-10 levels were inversely associated with many MetS clinical characteristics, as discussed above. In contrast, IL-1β levels in our study correlated directly with total leukocytes, the number of segmented neutrophils, and self-reported alcohol consumption. These differences in genotypes underscore the role of genetics in the inflammatory response, as specific genotypes may be associated with distinct immune response profiles, thus highlighting the significance of tailoring treatment and therapeutic approaches to a patient’s genetic characteristics.
5. Conclusions
This study’s findings highlight the complexity of the correlations between serum IL-10 and IL-1β levels and several variables in an older population that covered a range of age groups over 60. Our study also confirmed the IL10 rs1800890 (-3575T>A) and IL1B rs1143627 (–31T>C) gene variants effects on their serum concentrations in older adults (over 60), with IL10 rs1800890 ancestral genotype (TT) (56.5%) correlating with an average increase in serum IL-10 levels and IL1B rs1143627 mutated genotype (TT) (7.8%) correlating with an increase in serum IL-1β levels.
Serum IL-10 levels correlated weakly with serum HDL cholesterol levels and fat mass and inversely with serum levels of triglycerides, glucose, glycated hemoglobin (HbA1c), and estimated average blood glucose. IL-10 levels were also indirectly influenced by the patient’s time with T2DM, their amount of lean mass, and their amount of bone mineral content. In contrast, serum IL-1β levels correlated directly only with total leukocytes and the number of segmented neutrophils. These results might explain why only IL10 -3575T>A (rs1800890) SNPs correlated significantly to metabolic syndrome, with the AA genotype being a risk for metabolic syndrome.
Interestingly, participants over 80 had decreased IL-10 levels compared to other age groups. Participants with altered HDL levels or elevated serum glucose or HbA1c levels showed reduced serum IL-10 levels compared to those with normal levels. Women, in general, had higher serum IL-10 levels than men. As for serum IL-1β levels, only self-reported alcohol consumption showed statistically significant variations in serum IL-1β levels.
However, any interpretation must consider Brazilian demographic diversity, the presence of single or multiple chronic diseases, interactions between variables, and the patient’s genetic characteristics and environmental contexts. Causality cannot be inferred from our correlations as other unconsidered or uncontrolled factors may influence the observed associations. Nevertheless, our findings are crucial for comprehending the mechanisms that underlie health conditions in older adults and can direct future research and more targeted therapeutic approaches.
Author Contributions
Conceptualization, R.S.F., M.M.S., S.S.F. and I.C.R.S.; methodology, R.S.F., L.R.L., C.M.S.S., C.F.F., M.M.S., S.S.F. and I.C.R.S.; software, C.M.S.S. and I.C.R.S.; validation, C.M.S.S. and I.C.R.S.; formal analysis, R.S.F., C.M.S.S. and I.C.R.S.; investigation, R.S.F., C.M.S.S., M.M.S., S.S.F. and I.C.R.S.; resources, R.V.A., M.M.S., S.S.F. and I.C.R.S.; data curation, I.C.R.S.; writing—original draft preparation, R.S.F. and I.C.R.S.; writing—review and editing, R.S.F., C.M.S.S. and I.C.R.S.; visualization, I.C.R.S; supervision, R.V.A. and I.C.R.S.; project administration, M.M.S., S.S.F. and I.C.R.S.; funding acquisition, R.V.A., M.M.S., S.S.F. and I.C.R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001 and University of Brasília DPI/COPEI 7173/2021.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (or Research Ethics Committee—CEP) of the Health Sciences Teaching and Research Foundation (FEPECS) of the Federal District State Health Department (SES-DF) under opinion number 1.355.211/2017 with CAAE 59071116.8.0000.0030.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The research data are contained in the article’s tables.
Acknowledgments
We are grateful to the patients for participating in this study.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Instituto Brasileiro de Geografia e Estatística Projeções Da População | IBGE Censo 2022: Número de Pessoas Com 65 Anos Ou Mais de Idade Cresceu 57,4% Em 12 Anos.
- Vasconcelos, A.M.N.; Gomes, M.M.F. Transição Demográfica: A Experiência Brasileira. Epidemiologia e Serviços de Saúde 2012, 21, 539–548. [Google Scholar] [CrossRef]
- McKeown, R.E. The Epidemiologic Transition: Changing Patterns of Mortality and Population Dynamics. Am J Lifestyle Med 2009, 3, 19S–26S. [Google Scholar] [CrossRef]
- Vasconcelos, A.M.N.; Gomes, M.M.F. Transição Demográfica: A Experiência Brasileira. Epidemiologia e Serviços de Saúde 2012, 21, 539–548. [Google Scholar] [CrossRef]
- Pires Brandão, A.; Araújo Brandão, A.; da Rocha Nogueira, A.; Suplicy, H.; Ilha Guimarães, J.; de Oliveira, J.E.P. I Diretriz Brasileira de Diagnóstico e Tratamento Da Síndrome Metabólica. Arq Bras Cardiol 2005, 84, 3–28. [Google Scholar]
- Alves, R.; Santos, A.J.; Kislaya, I.; Nunes, B.; Freire, A.C. Síndrome Metabólica Em Portugal: Prevalência e Fatores Associados. Acta Med Port 2022, 35, 633–643. [Google Scholar] [CrossRef]
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int J Mol Sci 2022, 23, 786. [Google Scholar] [CrossRef]
- Simão, A.; Precoma, D.; Andrade, J.; Correa Filho, H.; Saraiva, J.; Oliveira, G.; Murro, A.; Campos, A.; Alessi, A.; Avezum Junior, A.; et al. I Diretriz Brasileira de Prevenção Cardiovascular. Arq Bras Cardiol 2013, 101, 1–63. [Google Scholar] [CrossRef]
- Kaur, J. A Comprehensive Review on Metabolic Syndrome. Cardiol Res Pract 2014, 2014, 1–21. [Google Scholar] [CrossRef]
- Maiuolo, J.; Gliozzi, M.; Musolino, V.; Carresi, C.; Scarano, F.; Nucera, S.; Scicchitano, M.; Bosco, F.; Ruga, S.; Zito, M.C.; et al. From Metabolic Syndrome to Neurological Diseases: Role of Autophagy. Front Cell Dev Biol 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Grandl, G.; Wolfrum, C. Hemostasis, Endothelial Stress, Inflammation, and the Metabolic Syndrome. Semin Immunopathol 2018, 40, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Kunnumakkara, A.B.; Aggarwal, S.; Aggarwal, B.B. Inflammation, a Double-Edge Sword for Cancer and Other Age-Related Diseases. Front Immunol 2018, 9. [Google Scholar] [CrossRef]
- Coppack, S.W. Pro-Inflammatory Cytokines and Adipose Tissue. Proceedings of the Nutrition Society 2001, 60, 349–356. [Google Scholar] [CrossRef]
- Maintinguer Norde, M.; Oki, E.; Ferreira Carioca, A.A.; Teixeira Damasceno, N.R.; Fisberg, R.M.; Lobo Marchioni, D.M.; Rogero, M.M. Influence of IL1B, IL6 and IL10 Gene Variants and Plasma Fatty Acid Interaction on Metabolic Syndrome Risk in a Cross-Sectional Population-Based Study. Clinical Nutrition 2018, 37, 659–666. [Google Scholar] [CrossRef]
- Kim, H.-J.; Higashimori, T.; Park, S.-Y.; Choi, H.; Dong, J.; Kim, Y.-J.; Noh, H.-L.; Cho, Y.-R.; Cline, G.; Kim, Y.-B.; et al. Differential Effects of Interleukin-6 and -10 on Skeletal Muscle and Liver Insulin Action In Vivo. Diabetes 2004, 53, 1060–1067. [Google Scholar] [CrossRef]
- Gao, M.; Zhang, C.; Ma, Y.; Bu, L.; Yan, L.; Liu, D. Hydrodynamic Delivery of MIL10 Gene Protects Mice From High-Fat Diet-Induced Obesity and Glucose Intolerance. Molecular Therapy 2013, 21, 1852–1861. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Pontillo, A.; Giugliano, F.; Giugliano, G.; Marfella, R.; Nicoletti, G.; Giugliano, D. Association of Low Interleukin-10 Levels with the Metabolic Syndrome in Obese Women. J Clin Endocrinol Metab 2003, 88, 1055–1058. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, D.; Yin, C.; Wang, S.; Wang, M.; Xiao, Y. IL-10/STAT3 Is Reduced in Childhood Obesity with Hypertriglyceridemia and Is Related to Triglyceride Level in Diet-Induced Obese Rats. BMC Endocr Disord 2018, 18, 39. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, L.-L.; Zhao, S.-J.; Lin, X.-X.; Liao, A.-H. IL-10: A Bridge between Immune Cells and Metabolism during Pregnancy. J Reprod Immunol 2022, 154, 103750. [Google Scholar] [CrossRef]
- York, A.G.; Skadow, M.H.; Qu, R.; Oh, J.; Mowel, W.K.; Brewer, J.R.; Kaffe, E.; Williams, K.J.; Kluger, Y.; Crawford, J.M.; et al. IL-10 Constrains Sphingolipid Metabolism via Fatty Acid Desaturation to Limit Inflammation. bioRxiv 2023. [Google Scholar] [CrossRef]
- Barcelos, A.L.V.; de Oliveira, E.A.; Haute, G.V.; Costa, B.P.; Pedrazza, L.; Donadio, M.V.F.; de Oliveira, J.R.; Bodanese, L.C. Association of IL-10 to Coronary Disease Severity in Patients with Metabolic Syndrome. Clinica Chimica Acta 2019, 495, 394–398. [Google Scholar] [CrossRef] [PubMed]
- LaMarche, N.M.; Kane, H.; Kohlgruber, A.C.; Dong, H.; Lynch, L.; Brenner, M.B. Distinct INKT Cell Populations Use IFNγ or ER Stress-Induced IL-10 to Control Adipose Tissue Homeostasis. Cell Metab 2020, 32, 243–258.e6. [Google Scholar] [CrossRef] [PubMed]
- Alfadul, H.; Sabico, S.; Al-Daghri, N.M. The Role of Interleukin-1β in Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Front Endocrinol (Lausanne) 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, T.; Horai, R.; Sudo, K.; Iwakura, Y. IL-1 Plays an Important Role in Lipid Metabolism by Regulating Insulin Levels under Physiological Conditions. J Exp Med 2003, 198, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Maedler, K.; Dharmadhikari, G.; Schumann, D.M.; Størling, J. Interleukin-Targeted Therapy for Metabolic Syndrome and Type 2 Diabetes. 2011; pp. 257–278. [Google Scholar]
- Netea, M.G.; Dinarello, C.A. More than Inflammation: Interleukin-1β Polymorphisms and the Lipid Metabolism. J Clin Endocrinol Metab 2011, 96, 1279–1281. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, S.J.; Trimigliozzi, K.; Dror, E.; Meier, D.T.; Molina-Tijeras, J.A.; Rachid, L.; Le Foll, C.; Magnan, C.; Schulze, F.; Stawiski, M.; et al. The Cephalic Phase of Insulin Release Is Modulated by IL-1β. Cell Metab 2022, 34, 991–1003.e6. [Google Scholar] [CrossRef] [PubMed]
- Ballak, D.B.; Stienstra, R.; Tack, C.J.; Dinarello, C.A.; van Diepen, J.A. IL-1 Family Members in the Pathogenesis and Treatment of Metabolic Disease: Focus on Adipose Tissue Inflammation and Insulin Resistance. Cytokine 2015, 75, 280–290. [Google Scholar] [CrossRef]
- Xu, J.; Yin, Z.; Cao, S.; Gao, W.; Liu, L.; Yin, Y.; Liu, P.; Shu, Y. Systematic Review and Meta-Analysis on the Association between IL-1B Polymorphisms and Cancer Risk. PLoS One 2013, 8, e63654. [Google Scholar] [CrossRef]
- Dai, Z.-M.; Liu, J.; Cao, X.-M.; Zhang, Y.; Wang, M.; Liu, X.-H.; Li, C.-J.; Dai, Z.-J.; Zhang, W.-G. Association Between Interleukin-10-3575T>A (Rs1800890) Polymorphism and Cancer Risk. Genet Test Mol Biomarkers 2015, 19, 324–330. [Google Scholar] [CrossRef]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and Therapeutic Potential of Interleukin-10. Journal of Experimental Medicine 2020, 217. [Google Scholar] [CrossRef]
- Fonseca, H.L.P. da A Reforma Da Saúde de Brasília, Brasil. Cien Saude Colet 2019, 24, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, N.; Tavira, B.; Hofwimmer, K.; Gutsmann, B.; Massier, L.; Abildgaard, J.; Juul, A.; Rydén, M.; Arner, P.; Laurencikiene, J. Sex-Specific Regulation of IL-10 Production in Human Adipose Tissue in Obesity. Front Endocrinol (Lausanne) 2022, 13. [Google Scholar] [CrossRef]
- Hussain, M.K. The Relationship Between IL-10 and Dislipidemia in Type 2 Diabetics. Al-Mustansiriyah Journal of Science 2011, 22, 202–213. [Google Scholar]
- Moraitis, A.G.; Freeman, L.A.; Shamburek, R.D.; Wesley, R.; Wilson, W.; Grant, C.M.; Price, S.; Demosky, S.; Thacker, S.G.; Zarzour, A.; et al. Elevated Interleukin-10: A New Cause of Dyslipidemia Leading to Severe HDL Deficiency. J Clin Lipidol 2015, 9, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.S.; Pereira, Í.S.; Santos, D.P.; Lopes, D.N.; Prado, A.O.; Calado, S.P.M.; Gonçalves, C. V.; Galantini, M.P.L.; Muniz, I.P.R.; Santos, G.S.; et al. Association between Body Composition and Inflammation: A Central Role of IL-17 and IL-10 in Diabetic and Hypertensive Elderly Women. Exp Gerontol 2019, 127, 110734. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.R.; Tavira, B.; Douagi, I.; Kulyté, A.; Arner, P.; Rydén, M.; Laurencikiene, J. Human-Specific Function of IL-10 in Adipose Tissue Linked to Insulin Resistance. J Clin Endocrinol Metab 2019, 104, 4552–4562. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M.; Fasshauer, M.; Tönjes, A.; Kratzsch, J.; Schön, M.; Paschke, R. Association of Interleukin-6, C-Reactive Protein, Interleukin-10 and Adiponectin Plasma Concentrations with Measures of Obesity, Insulin Sensitivity and Glucose Metabolism. Experimental and Clinical Endocrinology & Diabetes 2005, 113, 534–537. [Google Scholar] [CrossRef]
- Zahedi, A.S.; Daneshpour, M.S.; Akbarzadeh, M.; Hedayati, M.; Azizi, F.; Zarkesh, M. Association of Baseline and Changes in Adiponectin, Homocysteine, High-Sensitivity C-Reactive Protein, Interleukin-6, and Interleukin-10 Levels and Metabolic Syndrome Incidence: Tehran Lipid and Glucose Study. Heliyon 2023, 9, e19911. [Google Scholar] [CrossRef] [PubMed]
- Weiss, T.W.; Arnesen, H.; Seljeflot, I. Components of the Interleukin-6 Transsignalling System Are Associated with the Metabolic Syndrome, Endothelial Dysfunction and Arterial Stiffness. Metabolism 2013, 62, 1008–1013. [Google Scholar] [CrossRef]
- Silveira Rossi, J.L.; Barbalho, S.M.; Reverete de Araujo, R.; Bechara, M.D.; Sloan, K.P.; Sloan, L.A. Metabolic Syndrome and Cardiovascular Diseases: Going beyond Traditional Risk Factors. Diabetes Metab Res Rev 2022, 38. [Google Scholar] [CrossRef]
- Francischetti, I.; Moreno, J.B.; Scholz, M.; Yoshida, W.B. Os Leucócitos e a Resposta Inflamatória Na Lesão de Isquemia-Reperfusão. Revista Brasileira de Cirurgia Cardiovascular 2010, 25, 575–584. [Google Scholar] [CrossRef]
- Cruvinel, W. de M.; Mesquita Júnior, D.; Araújo, J.A.P.; Catelan, T.T.T.; Souza, A.W.S. de; Silva, N.P. da; Andrade, L.E.C. Sistema Imunitário: Parte I. Fundamentos Da Imunidade Inata Com Ênfase Nos Mecanismos Moleculares e Celulares Da Resposta Inflamatória. Rev Bras Reumatol 2010, 50, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.; Van de Water, J. Elevated Plasma Cytokines in Autism Spectrum Disorders Provide Evidence of Immune Dysfunction and Are Associated with Impaired Behavioral Outcome. Brain Behav Immun 2011, 25, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Guo, Y.; Ramos, R.I.; Hebroni, F.; Plaisier, S.B.; Xuan, C.; Granick, J.L.; Matsushima, H.; Takashima, A.; Iwakura, Y.; et al. Neutrophil-Derived IL-1β Is Sufficient for Abscess Formation in Immunity against Staphylococcus Aureus in Mice. PLoS Pathog 2012, 8, e1003047. [Google Scholar] [CrossRef]
- Garcia, L.P.; Sanchez, Z.M. Consumo de Álcool Durante a Pandemia Da COVID-19: Uma Reflexão Necessária Para o Enfrentamento Da Situação. Cad Saude Publica 2020, 36. [Google Scholar] [CrossRef]
- MAIO, R.; DICHI, J.B.; BURINI, R.C. Implicações Do Alcoolismo e Da Doença Hepática Crônica Sobre o Metabolismo de Micronutrientes. Arq Gastroenterol 2000, 37, 120–124. [Google Scholar] [CrossRef]
- Achur, R.N.; Freeman, W.M.; Vrana, K.E. Circulating Cytokines as Biomarkers of Alcohol Abuse and Alcoholism. Journal of Neuroimmune Pharmacology 2010, 5, 83–91. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
IL10 -3575T>A (rs1800890) and IL1Β -31 T>C (rs1143627) single nucleotide polymorphisms’ genotype distribution, according to the participants’ metabolic syndrome (MetS+ or -) profile.
Figure 1.
IL10 -3575T>A (rs1800890) and IL1Β -31 T>C (rs1143627) single nucleotide polymorphisms’ genotype distribution, according to the participants’ metabolic syndrome (MetS+ or -) profile.
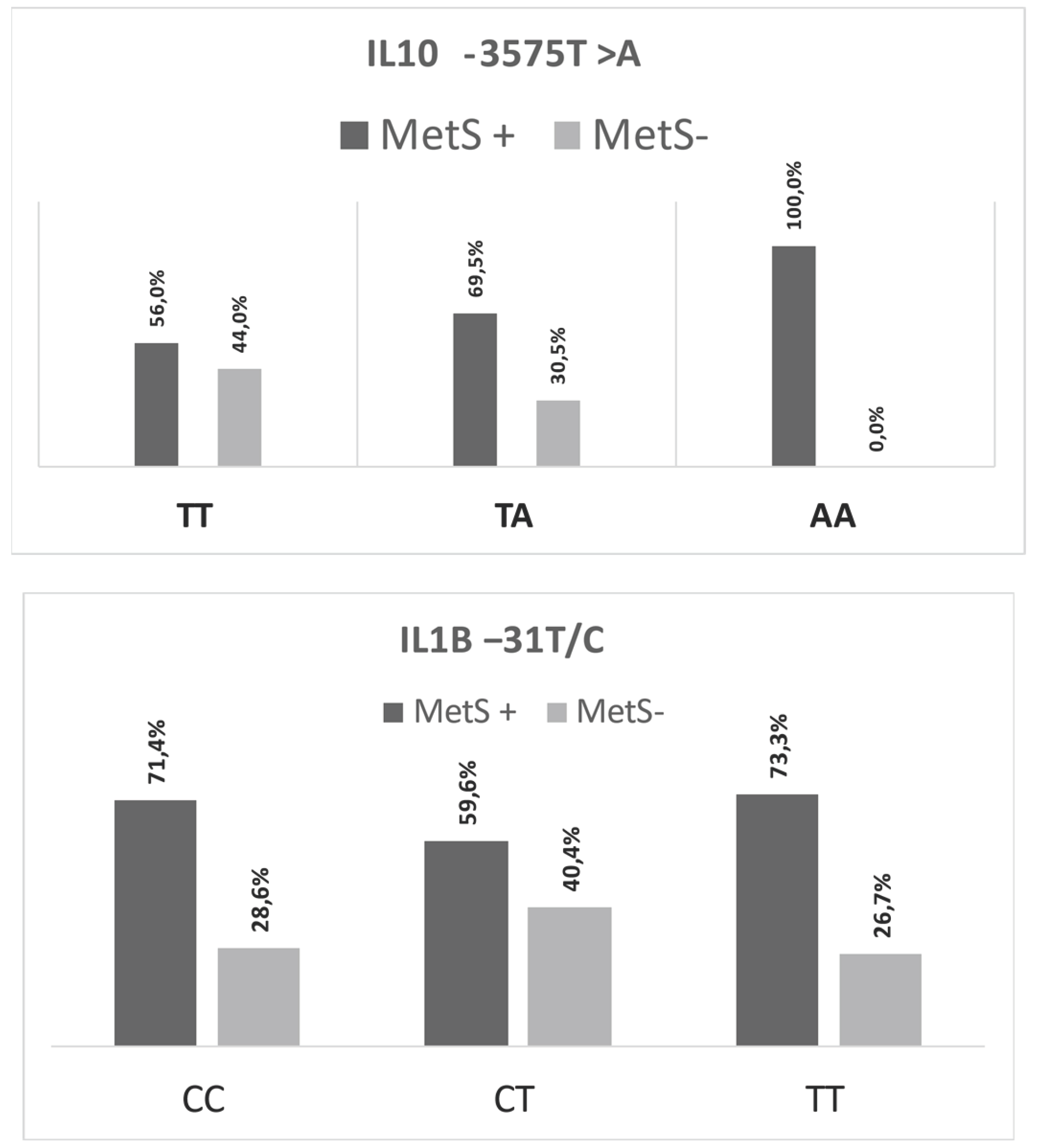

Table 1.
Correlations between IL-10 and IL-1β serum levels with biochemical, immunological, and anthropometric parameters.
Table 1.
Correlations between IL-10 and IL-1β serum levels with biochemical, immunological, and anthropometric parameters.
| Parameters | [IL-10] pg/mL | [IL-1 β] pg/mL | |
| [IL-10] pg/mL | ρ | 1.000 | 0.016 |
| P-value | - | 0.825 | |
| [IL-1 β] pg/mL | ρ | 0.016 | 1.000 |
| P-value | 0.825 | - | |
| Total leukocytes | ρ | -0.06 | 0.213** |
| P-value | 0.406 | 0.003 | |
| Rod neutrophils | ρ | 0.011 | 0.093 |
| P-value | 0.878 | 0.201 | |
| Segmented neutrophils | ρ | -0.02 | 0.163* |
| P-value | 0.781 | 0.024 | |
| Eosinophils | ρ | -0.096 | 0.046 |
| P-value | 0.187 | 0.530 | |
| Basophils | ρ | -0.104 | 0.028 |
| P-value | 0.154 | 0.703 | |
| Lymphocytes | ρ | 0.053 | -0.131 |
| P-value | 0.467 | 0.070 | |
| Monocytes | ρ | -0.068 | -0.104 |
| P-value | 0.350 | 0.153 | |
| Total cholesterol | ρ | 0.008 | 0.050 |
| P-value | 0.910 | 0.492 | |
| Triglycerides | ρ | -0.268** | 0.079 |
| P-value | <0.001 | 0.272 | |
| HDL | ρ | 0.365** | 0.038 |
| P-value | <0.001 | 0.598 | |
| LDL | ρ | 0.003 | 0.034 |
| P-value | 0.972 | 0.648 | |
| Total lipids | ρ | -0.023 | 0.106 |
| P-value | 0.830 | 0.311 | |
| Glucose | ρ | -0.370** | 0.017 |
| P-value | <0.001 | 0.817 | |
| HbA1c | ρ | -0.379** | 0.052 |
| P-value | <0.001 | 0.476 | |
| Estimated average blood glucose | ρ | -0.377** | 0.056 |
| P-value | <0.001 | 0.442 | |
| Glutamic oxaloacetic transaminase | ρ | 0.115 | -0.010 |
| P-value | 0.111 | 0.894 | |
| Pyruvic glutamic transaminase | ρ | 0.016 | -0.113 |
| P-value | 0.821 | 0.116 | |
| Time with T2DM | ρ | -0.264** | -0.031 |
| P-value | <0.001 | 0.677 | |
| Time with SAH | ρ | -0.071 | -0.053 |
| P-value | 0.332 | 0.471 | |
| BMI | ρ | 0.109 | 0.038 |
| P-value | 0.146 | 0.608 | |
| Waist Circumference | ρ | -0.150 | 0.073 |
| P-value | 0.062 | 0.366 | |
| Fat mass | ρ | 0.293** | 0.011 |
| P-value | <0.001 | 0.896 | |
| Lean Mass | ρ | -0.263** | -0.03 |
| P-value | <0.001 | 0.707 | |
| Bone mineral content | ρ | -0.328** | -0.012 |
| P-value | <0.001 | 0.879 | |
Caption: ρ: Spearman’s rank correlation coefficient; IL-10—interleukin 10; IL-1β—interleukin 1 beta; N—number of participants; *—P<0.05; **– P<0.001; <—less than; SAH—systemic arterial hypertension; T2DM—Type 2 Diabetes Mellitus; BMI—Body Mass Index; HDL—high-density lipoprotein; HbA1c—glycated hemoglobin; LDL—low-density lipoprotein.
Table 2.
Participants’ serum IL-10 and IL-1β concentration levels according to the presence/absence of clinical characteristics, signs, symptoms, or lifestyle habits.
Table 2.
Participants’ serum IL-10 and IL-1β concentration levels according to the presence/absence of clinical characteristics, signs, symptoms, or lifestyle habits.
| [IL-10] pg/mL | [IL-1β] pg/mL | |||||||||||||
| N | P25 | Median | P75 | P value | N | P25 | Median | P75 | P value | |||||
| MetS | Yes | 127 | 4.28 | 4.59 | 4.84 | <0.001* | 127 | 3.53 | 6.23 | 8.66 | 0.482 | |||
| No | 66 | 5.24 | 5.35 | 5.57 | 66 | 4.74 | 7.47 | 8.75 | ||||||
| Sarcopenia (DEXA) | Yes | 23 | 4.25 | 4.84 | 5.28 | 0.722 | 23 | 4.83 | 7.44 | 8.75 | 0.537 | |||
| No | 170 | 4.51 | 4.85 | 5.23 | 170 | 3.53 | 7.30 | 8.75 | ||||||
| SAH | Yes | 147 | 4.35 | 4.84 | 5.22 | 0.266 | 147 | 3.53 | 6.23 | 8.75 | 0.231 | |||
| No | 46 | 4.37 | 5.09 | 5.34 | 46 | 4.77 | 7.48 | 8.78 | ||||||
| T2DM | Yes | 108 | 4.34 | 4.83 | 5.12 | 0.007* | 108 | 3.53 | 7.30 | 8.68 | 0.821 | |||
| No | 85 | 4.51 | 5.02 | 5.38 | 85 | 3.54 | 7.40 | 8.76 | ||||||
| Age | 60 to 65 | 81 | 4.51 | 4.84a | 5.29 | 0.042* | 81 | 3.54 | 6.25 | 7.53 | 0.691 | |||
| 66 to 69 | 43 | 4.51 | 4.83a | 5.06 | 43 | 3.53 | 7.47 | 8.84 | ||||||
| 70 to 75 | 40 | 4.35 | 5.045a | 5.31 | 40 | 3.52 | 6.02 | 8.76 | ||||||
| 76 to 79 | 16 | 4.67 | 5.035a | 5.27 | 16 | 4.86 | 7.51 | 8.81 | ||||||
| ≥80 | 13 | 4.11 | 4.48b | 4.84 | 13 | 3.65 | 7.53 | 8.66 | ||||||
| Smoking | Yes | 14 | 4.31 | 4.94 | 5.24 | 0.808 | 14 | 3.50 | 6.85 | 8.75 | 0.644 | |||
| No | 179 | 4.37 | 4.84 | 5.24 | 179 | 3.54 | 7.40 | 8.75 | ||||||
| Use of alcoholic beverages | Yes | 9 | 4.48 | 5.01 | 5.24 | 0.709 | 9 | 3.51 | 3.53 | 5.94 | 0.037* | |||
| No | 184 | 4.37 | 4.84 | 5.24 | 184 | 3.55 | 7.42 | 8.75 | ||||||
| Perform physical exercises | Yes | 135 | 4.37 | 4.86 | 5.27 | 0.573 | 135 | 3.54 | 7.30 | 8.75 | 0.853 | |||
| No | 58 | 4.48 | 4.84 | 5.22 | 58 | 3.53 | 7.40 | 8.76 | ||||||
| Altered PA | Yes | 80 | 4.35 | 4.84 | 5.22 | 0.358 | 80 | 3.52 | 7.41 | 8.77 | 0.596 | |||
| No | 95 | 4.51 | 4.86 | 5.24 | 95 | 4.74 | 7.48 | 8.76 | ||||||
| Altered PAS | Yes | 56 | 4.33 | 4.79 | 5.16 | 0.241 | 56 | 3.52 | 6.24 | 8.73 | 0.288 | |||
| No | 116 | 4.51 | 4.86 | 5.24 | 116 | 4.71 | 7.48 | 8.77 | ||||||
| Altered PAD | No | 116 | 4.37 | 4.86 | 5.23 | 0.954 | 116 | 4.71 | 7.48 | 8.75 | 0.301 | |||
| Yes | 53 | 4.48 | 4.83 | 5.22 | 53 | 3.52 | 6.25 | 8.79 | ||||||
| HDL | Altered | 40 | 4.24 | 4.51 | 4.93 | <0.001* | 40 | 3.53 | 7.46 | 8.81 | 0.709 | |||
| Normal | 152 | 4.59 | 5.01 | 5.28 | 152 | 3.54 | 7.30 | 8.75 | ||||||
| Glucose | Altered | 98 | 4.29 | 4.76 | 5.01 | <0.001* | 98 | 3.53 | 6.27 | 7.65 | 0.567 | |||
| Normal | 95 | 4.79 | 5.22 | 5.40 | 95 | 4.68 | 7.41 | 8.76 | ||||||
| HbA1c | Altered | 110 | 4.29 | 4.77 | 5.02 | <0.001* | 110 | 3.54 | 7.46 | 8.75 | 0.341 | |||
| Normal | 82 | 4.79 | 5.19 | 5.38 | 82 | 3.52 | 6.19 | 8.69 | ||||||
| Total cholesterol | Altered | 102 | 4.48 | 4.86 | 5.24 | 0.806 | 102 | 3.54 | 7.41 | 8.76 | 0.911 | |||
| Normal | 91 | 4.35 | 4.84 | 5.22 | 91 | 3.53 | 6.29 | 8.75 | ||||||
| LDL | Altered | 100 | 4.37 | 4.84 | 5.24 | 0.945 | 100 | 3.54 | 7.46 | 8.79 | 0.891 | |||
| Normal | 87 | 4.35 | 4.84 | 5.22 | 87 | 3.53 | 6.29 | 8.75 | ||||||
| Waist Circumference | Altered | 123 | 4.51 | 4.84 | 5.07 | 0.851 | 123 | 3.53 | 7.30 | 8.75 | 0.343 | |||
| Normal | 32 | 4.28 | 4.82 | 5.30 | 32 | 3.52 | 6.23 | 7.50 | ||||||
| Biological sex | Female | 154 | 4.74 | 5.01 | 5.28 | <0.001* | 154 | 3.54 | 7.41 | 8.75 | 0.588 | |||
| Male | 39 | 4.25 | 4.31 | 5.18 | 39 | 3.53 | 6.09 | 8.79 | ||||||
Caption: IL-10—interleukin 10; IL-1β—interleukin 1 beta; N—number of participants; *—P<0.05; <—less than; MetS—metabolic syndrome; P25—25th percentile; P75—75th percentile; DEXA—dual energy X-ray absorptiometry; SAH—systemic arterial hypertension; T2DM—Type 2 Diabetes Mellitus; BP—blood pressure; SBP—systolic blood pressure; DBP—diastolic blood pressure; HDL—high-density lipoprotein; HbA1c—glycated hemoglobin; LDL—low-density lipoprotein. Note: Different letters denote statistical differences in different groups.
Table 3.
IL10 -3575T>A (rs1800890) and IL1Β -31 T>C (rs1143627) single nucleotide polymorphisms’ genotype distribution, according to the participants’ profile.
Table 3.
IL10 -3575T>A (rs1800890) and IL1Β -31 T>C (rs1143627) single nucleotide polymorphisms’ genotype distribution, according to the participants’ profile.
| IL10 -3575T >A | IL1Β −31T>C | |||||
| TT | TA | AA | CC | CT | TT | |
| [IL-10] pg/mL | [IL-10] pg/mL | [IL-10] pg/mL | [IL-1Β] pg/mL | [IL-1Β] pg/mL | [IL-1Β] pg/mL | |
| N | 109 | 59 | 25 | 84 | 94 | 15 |
| % | 56.5 | 30.6 | 12.9 | 43.5 | 48.7 | 7.8 |
| P25 | 4.76 | 4.28 | 4.02 | 3.49 | 7.48 | 17.46 |
| Median | 5.02a | 4.84b | 4.23c | 3.52a | 7.53b | 17.52c |
| P75 | 5.34 | 5.23 | 4.29 | 4.76 | 8.82 | 18.66 |
| P–value | <0.001* | <0.001* | ||||
Caption: IL10—interleukin 10 gene; IL1Β—interleukin 1 beta gene; N—number of participants; %—percentage; <—less than; *—P<0.05; P25—25th percentile; P75—75th percentile. Note: Different letters denote statistical differences in different groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



