
Submitted:
06 March 2024
Posted:
06 March 2024
You are already at the latest version
Abstract
This study aimed to elucidate the similarities and differences between amyloid-forming corpora amylacea (CA) in the prostate and lung, examine the nature of CAs in cystic tumors of the atrioventricular node (CTAVN), and clarify the distinctions between amyloid-forming CA and spheroid-type amyloid deposition. We conducted proteomics analyses using liquid chromatography–tandem mass spectrometry with laser microdissection and immunohistochemistry to validate the characteristics of CAs in the lung and prostate. Our findings revealed that CAs in these organs primarily consist of common proteins (β2-microglobulin and lysozyme) and locally produced proteins. Moreover, we observed a discrepancy between the histopathological and proteomic analysis results in CTAVN-associated CAs. In addition, while the histopathological appearance of amyloid-forming CAs and spheroid-type amyloid deposits were nearly identical, the latter deposition lacked β2-microglobulin and lysozyme, and exhibited evident destruction of surrounding tissue. A literature review further supported these findings. These results suggest that amyloid-forming CAs in the lung and prostate are formed through a shared mechanism, serving as waste containers (wasteosomes) and/or storage for excess proteins (functional amyloids). In contrast, we hypothesize that while amyloid-forming CA and spheroid-type amyloid deposits are formed, in part, through common mechanisms, the latter are pathological.
Keywords:
Amyloid
; beta-2 micro-globulin
; corpora amylacea
; functional amyloids
; lactoferrin
; lysosome
; macrophage
; pulmonary surfactant protein
1. Introduction
Corpora amylacea (CA) has been described in various human organs, including the brain, lungs, and prostate [1,2,3,4]. CA primarily involves two different types of lesions: one is a polyglucosan structure similar to starch, which is positive by periodic acid–Schiff staining, whereas the other is an aggregate of certain types of fibrillary proteins that stain positively for Congo red (CR) and thioflavin [1]. Although these are histopathologically distinct lesions, they are believed to share a common function as wastesomes [1]. Amyloid-forming CA formation is frequently observed in the prostate and lungs [1,5,6]. Several histopathological and biochemical studies have evaluated CA of the prostate [7,8], but few studies have examined CA in the lung. Furthermore, there have been no studies examining the similarities and differences between CA at the two sites.
Cystic tumor of the atrioventricular node (CTAVN) is a rare, benign lesion located at the base of the atrial septum, around the area of the AVN in the human heart. CAs are a histopathological feature of CTAVN [4,9]. These deposits may have amyloid properties because of their green birefringence under polarized light [9]; however, there has been no characterization of these deposits, particularly whether they contain amyloid-associated proteins, based on proteomic analyses.
Spheroid-type amyloid deposition (STAD) is a rare form of amyloidosis [10,11,12]. This pattern of deposition often appears as a localized form, but it can also be associated with systemic amyloidosis. Morphologically, amyloid-forming CA and STAD are very similar; however, the latter lesions often occur in association with tumors or cause damage to surrounding tissues, which suggests that they are pathologically distinct lesions. To our knowledge, no studies have thoroughly examined the differences between these lesions.
The objectives of this study were as follows: 1) elucidate the similarities and differences between amyloid-forming CAs in the prostate and lung; 2) determine the nature of CAs in CTAVN; and 3) clarify the differences between amyloid-forming CA and STAD. We conducted proteomics analyses using liquid chromatography–tandem mass spectrometry (LC-MS/MS) with laser microdissection (LMD) in a patient with pulmonary and prostatic amyloid-forming CAs. Immunohistochemistry was performed in a series of three patients with pulmonary CA and three patients with prostatic CA to validate the results of the proteomic analyses. Moreover, we performed the same proteomic analysis in a patient with CTAVN-associated CR-positive CA. To address the third objective, we re-evaluated the results of our previously reported histopathologic and proteomic analyses of a STAD patient [10]. Finally, we reviewed prior histopathological studies on STAD and compared their findings with those obtained in the present study on amyloid-forming CA.
2. Results
2.1. General Appearance of Pulmonary, Prostatic, and CTAVN-Associated CAs
We selected three cases of pulmonary CA, three cases of prostatic CA, three cases of CA associated with CTAVN, and one case of STAD from the archives of all medicolegal autopsy patients from our department between January 2008 and August 2023. A summary of the cases is presented in Table 1.
2.2. General appearance of pulmonary, prostatic, and CTAVN-associated CAs
Representative histopathological microphotographs are shown in Figure 1.
Generally, all CAs exhibited round, eosinophilic deposits with concentric laminations by H&E staining (Figure 1a–c). Pulmonary CAs exhibited a relatively uniform size and shape, with weak to moderate congophilia, and exhibited clear apple-green birefringence under polarized light observation (Figure 1a, d, g). In contrast, prostatic CAs varied in size and shape, with strong congophilia and distinct apple-green birefringence under polarized light (Figure 1b, e, h). CTAVN-associated CAs also varied in size and shape, with weak to moderate congophilia, and only some displaying apple-green birefringence under polarized light (Figure 1c, f, i). In addition, macrophages were observed around CAs in the lung (Figure 1a) and prostate (Figure 1b) but not around CTAVN-associated CAs (Figure 1c). There was no evident destruction of the surrounding tissue by CAs in all cases.
2.3. Proteomic and immunohistochemical features of the pulmonary CAs
A summary of the proteomics results and representative microphotographs of pulmonary CAs in Case 1 are shown in Figure 2. Additional microphotographs of pulmonary CAs in all three cases are provided in Supplementary Figure S1.
LC-MS/MS with LMDdetected five proteins thought to be amyloidogenic, as shown in Figure 2a. Immunohistochemical localization within the CAs was confirmed for β2-microglobulin (Figure 2b), lysozyme C (Figure 2c), pulmonary surfactant protein (PSP)-A, and PSP-B (Figure 2d, e). Immunohistochemistry for lactoferrin yielded negative results (Figure 2f), whereas focal p62 immunoreactivity was observed (Figure 2g).
2.4. Proteomic and immunohistochemical features of prostatic CAs
A summary of the proteomics results for prostatic CAs in Case 1, along with representative microphotographs of Cases 1 and 2, are shown in Figure 3. Representative microphotographs of prostatic CAs for all three cases are provided in Supplementary Figure S2.
In addition to some amyloid-associated proteins, three proteins that may be amyloidogenic were detected by LC-MS/MS with LMD(Figure 3a), which is consistent with previous studies [7,8]. Immunohistochemistry revealed that all CAs were positive for lysozyme C (Figure 3b) and lactoferrin (Figure 3c). Focally, p62 immunoreactivity was identified (Figure 3d). Interestingly, eosinophilic deposits showing moderate congophilia and apple-green birefringence under polarized light were identified within a few carcinoma glands (Figure 3e–g); however, lysozyme C, lactoferrin, and p62 were not detected in these deposits by immunohistochemistry (Figure 3h–j).
2.5. Comparison of the amino acid sequences of proteins commonly identified in pulmonary and prostatic CAs
The amino acid sequences and identified peptide sequences of β2-microglobulin, lysozyme C, and lactoferrin in the lungs and prostate are shown in Figure 4.
For β2-microglobulin and lysozyme C, nearly identical regions were identified in the lung and prostate (Figure 4a–d); however, for lactoferrin, only a small portion was identified in the lung (Figure 4e), particularly on the C-terminal end, whereas almost the entire length of lactoferrin was identified in the prostate (Figure 4f). The detected PSP-A and PSP-B peptide sequences are shown in Supplementary Figure S3.
2.6. Proteomic and immunohistochemical features of CTAVN-associated CAs
A summary of the proteomics results and representative microphotographs of immunohistochemistry in Case 1 are shown in Figure 5.
In contrast to pulmonary and prostatic CAs, we did not detect amyloid-associated or reported amyloidogenic proteins in CTAVN-associated CAs by LC-MS/MS with LMD (Case 1); however, consistent with our previous report [4], olfactomedin 4 was detected in CAs (Figure 5a). The CAs of all three cases exhibited weak to strong immunoreactivity for olfactomedin 4 (Figure 5b–d). In contrast, p62 immunoreactivity was identified only in Case 1 (Figure 5f–h). Representative findings for all three CTAVN cases are shown in Supplementary Figure S4.
2.7. Proteomic and immunohistochemical features of STAD associated with systemic amyloid immunoglobulin light chain (AL) amyloidosis
Representative microphotographs are shown in Figure 6.
In this case, the shape of the amyloid deposits was similar to that of CAs, particularly in the prostate; however, the destruction of surrounding tissue by the deposits was evident. Furthermore, although some amyloid-associated proteins and 7 peptides from the immunoglobulin light chain lambda variable region were detected, β2-microglobulin and lysozyme C were not detected. Furthermore, there was no p62 immunoreactivity in the deposits.
2.8. Literature review of previous reports on STAD
To further characterize STAD, we conducted a comprehensive literature review and summarized the results in Table 2 [10,11,12,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45].
STAD was observed in both localized and systemic amyloidosis. Although the size and morphology varied between cases and among the organs, the deposits appeared to show concentric laminations in most cases. In addition, the presence of histiocytes and multinucleated giant cells was noted around the deposits in some cases, suggesting that they may be involved in the formation of the deposits [10,11]. These features are similar to those observed in amyloid-forming CAs. Consistent with our results, immunohistochemistry and proteomics analysis did not identify β2-microglobulin or lysozyme C in the deposits, and tissue damage resulting from the deposits was believed to have occurred in almost all STAD cases.
3. Discussion
In the present study, we demonstrated that: (1) CAs in the lung and prostate are comprised of common proteins (β2-microglobulin and lysozyme C) and locally produced proteins (PSP-A and PSP-B in the lung; lactoferrin in the prostate); (2) CAs associated with CTAVN exhibit typical histopathological features of amyloid, whereas LMD with LC-MS/MS did not detect amyloid-associated or amyloidogenic proteins; (3) STAD is morphologically similar to amyloid-forming Cas, but differs because it causes tissue destruction and lacks β2-microglobulin or lysozyme C deposition (4). p62 immunoreactivity was observed in amyloid-forming Cas, but not in STAD.
Several proteomic and immunohistochemical analyses of prostatic CA have been reported [7,8,46,47,48]; however, to our knowledge, this is the first comprehensive analysis of pulmonary CA using both methods. CAs in the lung and prostate consist of common proteins (β2-microglobulin and lysozyme C) and locally produced proteins. This suggests a common mechanism underlying the formation of CAs in the lungs and prostate. CAs are frequently identified in normal prostate tissue [49], and their presence does not appear to damage the surrounding tissues in the prostate and lung. Thus, we can speculate that CAs in these organs are formed for physiological reasons rather than serving a pathological function. Riba et al. proposed that CAs act as “wasteosomes” to sequester waste products [1,50]. In contrast, certain polypeptide hormones are stored in an amyloid-like β-sheet conformation in the pituitary gland, which are considered functional amyloids [13,51]. Thus, it has been hypothesized that certain proteins, especially those prone to form amyloid fibrils, are disposed of or stored as amyloid-forming CA. Partial p62 immunoreactivity supports the role of amyloid-forming CAs as a wasteosome. β2-microglobulin and lysozyme C may be essential components for the formation of amyloid CAs.
Macrophages were observed around the CAs in both organs and were also positive for β2-microglobulin and lysozyme C, suggesting that they play an important role in this mechanism. Dobashi et al. suggested that the concentrically laminated bodies in pulmonary CAs may be formed by sequential aggregation, fusion, coalescence, and compaction of degenerated alveolar macrophages [6]; however, it remains unclear whether they remove deposits, create them, or perform both functions. Further studies are needed to elucidate this mechanism, as it may contribute to the development of amyloid removal therapy.
There are four types of PSPs: hydrophilic PSP-A and PSP-D, and hydrophobic PSP-B and PSP-C [52]. In pulmonary CAs, proteomic analysis revealed PAP-A and PAP-B, whereas PSP-C, known to form amyloid fibrils [53], was not detected. These results were confirmed by immunohistochemistry and are consistent with those of a previous report [54]. To our knowledge, however, there are no reports of PAP-A and PSP-B forming amyloid fibrils. Thus, it is unclear whether the PSP-A and PSP-B proteins within the pulmonary CA form amyloid fibrils or are merely deposited. During the metabolism of PSPs, >50% is derived from recycling or catabolic events managed by functional cross-talk between alveolar type II cells and macrophages [52]. We hypothesize that excess or denatured PSPs are converted into CAs to be discarded or stored as a stable structure. It is noteworthy that if this serves a storage function, it may represent a form of functional amyloids [55].
We observed pCR-positive deposits not only in benign acini, but also in cancer acini. Although CAs are primarily noted in benign acini, they have been identified in 0.4%–13% of cancer acini [49]. Therefore, the observation of amyloid-forming deposits in the lumen of cancer acini is not surprising; however, it is noteworthy that neither the tumor glands nor their luminal deposits were immunoreactive to lactoferrin. This is consistent with the proteomic results of Tekin et al. [47]. Thus, it is likely that the composition of amyloid deposits in cancer acini differs from those in benign acini, which may lead to the discovery of a novel amyloid precursor protein. However, it should be noted that given the presumed nonphysiological nature of secretions in cancer acini, there may be a discrepancy between the histological findings of the deposits and the results obtained from proteomic analysis, as illustrated in the case of CTAVN, which is discussed below. This is a topic for future consideration.
Interestingly, all CTAVN cases exhibited CAs positive for pCR staining and displayed apple green birefringence under polarized light; however, proteomic analysis yielded divergent results, with no accumulation of amyloid-associated proteins. The precise reason for this discrepancy remains unknown. One hypothesis is that CTAVN CAs exhibit significant variation in the degree of amyloid fibril formation, given the substantial differences in the congophilia of each deposit. Consequently, it is plausible that only deposits lacking amyloid formation were selected during the LMD process. Another possibility is that congophilia is a false positive. Because CTAVN is a non-physiologic lesion, the CAs forming within the glands of CTAVN may have different properties compared with those in the lung or prostate. The immunohistochemical characteristics of the CTAVN epithelium are similar to those of the prostate epithelium [4]; however, the proteins comprising CA are distinctly different in the two tissues. Moreover, given the significant sex-dependent differences observed in the immunohistochemical properties of the epithelium of CTAVN [4], it is important to consider that the proteomic analysis of CAs from female patients in this study may have generated results distinct from those of the prostate. It will be necessary to collect more cases of CTAVN and analyze whether amyloid fibrils are indeed formed in CAs associated with CTAVN.
Through morphological examination and a literature review, we demonstrated that amyloid-forming CAs and STADs exhibit morphological similarities; however, the former does not exhibit distinct destruction of the surrounding tissue, in contrast to STAD. Collectively, these findings suggest that the formation of characteristic deposition morphology in amyloid-forming CAs and STAD may be partially mediated by common mechanisms, whereas the former is more closely associated with physiological processes, while the latter is likely formed through pathological mechanisms. This is supported by the observation that p62 was partially positive in amyloid-forming CAs, whereas it was negative in STAD. Nevertheless, to our knowledge, there are limited studies showing the immunoreactivity of proteins related to waste substance processing and elimination, such as ubiquitin or p62, in STAD [3,23]. Additional research in this area is warranted.
In conclusion, we present the findings of a comprehensive analysis that integrates proteomic analysis and immunohistochemistry for amyloid-forming CAs. A summary of the results is presented in Table 3.
These results suggest that amyloid-forming CAs in the lung and prostate may be formed through a shared mechanism, serving as waste containers (wasteosomes) and/or storage for excess proteins (functional amyloids). It would be interesting if the human body intentionally employs structures that manifest distinctive amyloid fibril formation at the histological level. In contrast, in CAs associated with CTAVN, there was a discrepancy between the histopathological and proteomic analysis results, highlighting the presence of nonphysiological functions of the epithelium in this disease. Furthermore, while amyloid-forming CA and STAD are formed, in part, by some common mechanisms, the former may have physiological origins, whereas the latter are pathological. If this shared mechanism exists, its elucidation will contribute to the development of amyloid removal therapy.
4. Materials and Methods
4.1. Tissue samples
Tissue specimens from the lung (for evaluating pulmonary CA), prostate (for evaluating prostatic CA), heart (for evaluating CTAVN-associated CA), and kidney (for evaluating STAD) were collected, fixed in 20% buffered formalin, and routinely embedded in paraffin. Then, 4-μm-thick sections were cut and stained with hematoxylin and eosin (H&E) or were analyzed by immunohistochemistry. Furthermore, 6-μm-thick sections were cut and stained with phenol CR (pCR) [56].
4.2. Histopathological evaluation
The presence of CAs was determined using H&E-stained specimens. pCR-positive structures, which showed typical apple-green birefringence under polarized light, were histologically confirmed as amyloid deposits. Immunohistochemistry was performed to confirm the proteomics results. The immunohistochemistry methods used in this study are summarized in Supplementary Table S1. Immunostaining was performed using the Leica Bond-MAX automation system with Leica Refine detection kits (Leica Biosystems, Bannockburn, IL), following the manufacturer’s instructions. All sections were counterstained with hematoxylin.
4.3. Proteomics analysis using mass spectrometry
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Summary of the antibodies used in this study. Figure S1: Representative microphotographs of the pulmonary corpora amylacea (CA) in all three cases. Figure S2: Representative microphotographs of prostatic CA in all three cases. Figure S3: Proteomic analysis results of PSP-A and PSP-B in the pulmonary CA lesions in Case 1. Figure S4: Representative microphotographs of a cystic tumor of atrioventricular node (CTAVN)-associated CA with all three cases.
Author Contributions
Conceptualization, Shojiro Ichimata; Data curation, Shojiro Ichimata; Formal analysis, Shojiro Ichimata; Funding acquisition, Shojiro Ichimata; Investigation, Shojiro Ichimata, Yukiko Hata, Tsuneaki Yoshinaga, Nagaaki Katoh, Fuyuki Kametani, Masahide Yazaki, and Yoshiki Sekijima; Methodology, Fuyuki Kametani; Project administration, Shojiro Ichimata; Resources, Yukiko Hata, and Naoki Nishida; Supervision, Naoki Nishida; Validation, Naoki Nishida; Visualization, Shojiro Ichimata; Writing—original draft, Shojiro Ichimata; Writing—review & editing, Yukiko Hata, Tsuneaki Yoshinaga, Nagaaki Katoh, Fuyuki Kametani, Masahide Yazaki, Yoshiki Sekijima, and Naoki Nishida.
Funding
This study was supported in part by a JSPS KAKENHI grant awarded to Shojiro Ichimata (grant number JP20K18979).
Institutional Review Board Statement
This study was approved by the Ethical Committee of Toyama University (I2020006) and followed the ethical standards established in the 1964 Declaration of Helsinki and updated in 2008.
Data Availability Statement
The datasets used and analyzed in the current study are available from the corresponding authors upon request.
Acknowledgments
The authors would like to thank Ms. Miyuki Maekawa for her technical assistance. The authors would also like to thank Enago (www.enago.jp) for the English language review.
Conflicts of Interest
All authors declare no conflicts of interest.
Abbreviations
CA, corpora amylacea; CTAVN, cystic tumor of the atrioventricular node; pCR, phenol Congo red; H&E, hematoxylin and eosin; LC-MS/MS, liquid chromatography-tandem mass spectrometry; LMD, laser microdissection; PSP, pulmonary surfactant protein; STAD, spheroid-type amyloid deposition
References
- Riba, M.; Del Valle, J.; Auge, E.; Vilaplana, J.; Pelegri, C. From corpora amylacea to wasteosomes: History and perspectives. Ageing Res Rev 2021, 72, 101484. [Google Scholar] [CrossRef] [PubMed]
- Hechtman, J.F.; Gordon, R.E.; McBride, R.B.; Harpaz, N. Corpora amylacea in gastrointestinal leiomyomas: a clinical, light microscopic, ultrastructural and immunohistochemical study with comparison to hyaline globules. J Clin Pathol 2013, 66, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Hechtman, J.F.; Gordon, R.E.; Harpaz, N. Intramuscular corpora amylacea adjacent to ileal low-grade neuroendocrine tumours (typical carcinoids): a light microscopic, immunohistochemical and ultrastructural study. J Clin Pathol 2013, 66, 566–572. [Google Scholar] [CrossRef]
- Ichimata, S.; Hata, Y.; Yajima, N.; Katayama, Y.; Nomoto, K.; Nishida, N. Sex-dependent expression of prostatic markers and hormone receptors in cystic tumor of the atrioventricular node: A histopathological study of three cases. Pathol Int 2021, 71, 141–146. [Google Scholar] [CrossRef] [PubMed]
- DuPre, N.C.; Flavin, R.; Sfanos, K.S.; Unger, R.H.; To, S.; Gazeeva, E.; Fiorentino, M.; De Marzo, A.M.; Rider, J.R.; Mucci, L.A.; Transdisciplinary Prostate Cancer Partnership (ToPCaP). Corpora amylacea in prostatectomy tissue and associations with molecular, histological, and lifestyle factors. Prostate 2018, 78, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Dobashi, M.; Yuda, F.; Narabayashi, M.; Imai, Y.; Isoda, N.; Obata, K.; Umetsu, A.; Ohgushi, M. Histopathological study of corpora amylacea pulmonum. Histol Histopathol 1989, 4, 153–165. [Google Scholar] [PubMed]
- Sfanos, K.S.; Wilson, B.A.; De Marzo, A.M.; Isaacs, W.B. Acute inflammatory proteins constitute the organic matrix of prostatic corpora amylacea and calculi in men with prostate cancer. Proc Natl Acad Sci U S A 2009, 106, 3443–3448. [Google Scholar] [CrossRef]
- Kanenawa, K.; Ueda, M.; Isoguchi, A.; Nomura, T.; Tsuda, Y.; Masuda, T.; Misumi, Y.; Yamashita, T.; Ando, Y. Histopathological and biochemical analyses of prostate corpora amylacea. Amyloid 2019, 26, 160–161. [Google Scholar] [CrossRef]
- David, R.; Hiss, Y. Corpora amylacea in mesothelioma of the atrioventricular node. J Pathol 1978, 124, 111–116. [Google Scholar] [CrossRef]
- Ichimata, S.; Hata, Y.; Abe, R.; Yoshinaga, T.; Katoh, N.; Kametani, F.; Yazaki, M.; Sekijima, Y.; Ehara, T.; Nishida, N. An autopsy case of amyloid tubulopathy exhibiting characteristic spheroid-type deposition. Virchows Arch 2020, 477, 157–163. [Google Scholar] [CrossRef]
- Ichimata, S.; Aoyagi, D.; Yoshinaga, T.; Katoh, N.; Kametani, F.; Yazaki, M.; Uehara, T.; Shiozawa, S. A case of spheroid-type localized lactoferrin amyloidosis in the bronchus. Pathol Int 2019, 69, 235–240. [Google Scholar] [CrossRef]
- Kim, M.J.; McCroskey, Z.; Piao, Y.; Belcheva, A.; Truong, L.; Kurtin, P.J.; Ro, J. Y. Spheroid-type of AL amyloid deposition associated with colonic adenocarcinoma: A case report with literature review. Pathol Int 2018, 68, 123–127. [Google Scholar] [CrossRef]
- Buxbaum, J.N.; Dispenzieri, A.; Eisenberg, D.S.; Fandrich, M.; Merlini, G.; Saraiva, M.J.M.; Sekijima, Y.; Westermark, P. Amyloid nomenclature 2022: update, novel proteins, and recommendations by the International Society of Amyloidosis (ISA) Nomenclature Committee. Amyloid 2022, 29, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Demirhan, B.; Bilezikci, B.; Kiyici, H.; Boyacioglu, S. Globular amyloid deposits in the wall of the gastrointestinal tract: report of six cases. Amyloid 2002, 9, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, P.R.; Topazian, M.D.; Gertz, M.A.; Abraham, S. C. Globular amyloid deposits isolated to the small bowel: a rare association with AL amyloidosis. Am J Surg Pathol 2007, 31, 141–145. [Google Scholar] [CrossRef]
- Acebo, E.; Mayorga, M.; Fernando Val-Bernal, J. Primary amyloid tumor (amyloidoma) of the jejunum with spheroid type of amyloid. Pathology 1999, 31, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Venugopal, S.; Ravindra, S. Unique spheroid deposits of amyloid in an ampullary neuroendocrine tumour. Indian J Pathol Microbiol 2022, 65, 226–228. [Google Scholar] [PubMed]
- Diaz Del Arco, C.; Fernandez Acenero, M.J. Globular amyloidosis of the colon. Arab J Gastroenterol 2018, 19, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Martín-Arranz, E.; Pascual-Turrión, J.M.; Martín-Arranz, M.D.; Burgos, E.; Froilán-Torres, C.; Adán-Merino, L.; Lorenzo, A.; Segura-Cabral, J.M. Focal globular amyloidosis of the colon. An exceptional diagnosis. Rev Esp Enferm Dig 2010, 102, 555–556. [Google Scholar] [CrossRef]
- Harris, J.C.; Zhang, Q.; Tondon, R.; Alipour, Z.; Stashek, K. Characterization of amyloidosis in the gastrointestinal tract with an emphasis on histologically distinct interstitial patterns of deposition and misinterpretations. Am J Surg Pathol 2024, 48, 302–308. [Google Scholar] [CrossRef]
- Makhlouf, H.R.; Goodman, Z.D. Globular hepatic amyloid: an early stage in the pathway of amyloid formation: a study of 20 new cases. Am J Surg Pathol 2007, 31, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Agaram, N.; Shia, J.; Klimstra, D.S.; Lau, N.; Lin, O.; Erlandson, R.A.; Filippa, D.A.; Godwin, T.A. Globular hepatic amyloid: a diagnostic peculiarity that bears clinical significance. Hum Pathol 2005, 36, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Pilgaard, J.; Fenger, C.; Schaffalitzky de Muckadell, O.B. Globular amyloid deposits in the liver. Histopathology 1993, 23, 479–480. [Google Scholar] [CrossRef] [PubMed]
- Osick, L.A.; Lee, T.P.; Pedemonte, M.B.; Jacob, L.; Chauhan, P.; Navarro, C.; Comer, G.M. Hepatic amyloidosis in intravenous drug abusers and AIDS patients. J Hepatol 1993, 19, 79–84. [Google Scholar] [CrossRef] [PubMed]
- French, S.W.; Schloss, G.T.; Stillman, A.E. Unusual amyloid bodies in human liver. Am J Clin Pathol 1981, 75, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Chandan, V.S.; Shah, S.S.; Lam-Himlin, D.M.; Petris, G.D.; Mereuta, O.M.; Dogan, A.; Torbenson, M.S.; Wu, T.T. Globular hepatic amyloid is highly sensitive and specific for LECT2 amyloidosis. Am J Surg Pathol 2015, 39, 558–564. [Google Scholar] [CrossRef]
- Kumar, B.; Pant, B.; Kumar, V.; Negi, M. Sinonasal globular amyloidosis simulating malignancy: A rare presentation. Head Neck Pathol 2016, 10, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Drut, R.; Giménez, P.O. Acinic cell carcinoma of salivary gland with massive deposits of globular amyloid. Int J Surg Pathol 2008, 16, 202–207. [Google Scholar] [CrossRef]
- Michaels, L.; Hyams, V.J. Amyloid in localised deposits and plasmacytomas of the respiratory tract. J Pathol 1979, 128, 29–38. [Google Scholar] [CrossRef]
- Pambuccian, S.E.; Horyd, I.D.; Cawte, T.; Huvos, A.G. Amyloidoma of bone, a plasma cell/plasmacytoid neoplasm. Report of three cases and review of the literature. Am J Surg Pathol 1997, 21, 179–186. [Google Scholar] [CrossRef]
- Bauer, W.H.; Kuzma, J.F. Solitary tumors of atypical amyloid (paramyloid). Am J Clin Pathol 1949, 19, 1097–1112. [Google Scholar] [CrossRef]
- Unal, F.; Hepgül, K.; Bayindir, C.; Bilge, T.; Imer, M.; Turantan, I. Skull base amyloidoma: Case report. J Neurosurg 1992, 76, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Bommannan, B.K.K.; Sonai, M.; Sachdeva, M.U. Bone marrow amyloid spherulites in a case of AL amyloidosis. Blood Cells Mol Dis 2016, 58, 19–20. [Google Scholar] [CrossRef] [PubMed]
- Mantoo, S.; Hwang, J.S.; Chiang, G.S.; Tan, P.H. A rare case of localised AA-type amyloidosis of the ureter with spheroids of amyloid. Singapore Med J 2012, 53, e77–e79. [Google Scholar]
- Gibbons, D.; Lindberg, G.M.; Ashfaq, R.; Saboorian, M.H. Localized amyloidosis of the uterine cervix. Int J Gynecol Pathol 1998, 17, 368–371. [Google Scholar] [CrossRef]
- Gondo, T.; Ishihara, T.; Kawano, H.; Uchino, F.; Takahashi, M.; Iwata, T.; Matsumoto, N.; Yokota, T. Localized amyloidosis in squamous cell carcinoma of uterine cervix: electron microscopic features of nodular and star–like amyloid deposits. Virchows Arch A Pathol Anat Histopathol 1993, 422, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Aho, H.J.; Talve, L.; Mäenpää, J. Acantholytic squamous cell carcinoma of the uterine cervix with amyloid deposition. Int J Gynecol Pathol 1992, 11, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Nakayama, T.; Ohata, K.; Wakasa, K.; Miki, Y. Computed tomography and magnetic resonance imaging appearance of prolactinoma with spheroid-type amyloid deposition. J Comput Assist Tomogr 2011, 35, 313–315. [Google Scholar] [CrossRef]
- Gul, S.; Bahadir, B.; Dusak, A.; Kalayci, M.; Edebali, N.; Acikgoz, B. Spherical amyloid deposition in a prolactin-producing pituitary adenoma. Neuropathology 2009, 29, 81–84. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, S.H.; Lee, S.K.; Kim, T.S. Squash smear findings of spherical amyloid in pituitary prolactinoma. A case report. Acta Cytol 2004, 48, 447–450. [Google Scholar] [CrossRef]
- Hassan, T.; Ikeda, H.; Yoshimoto, T. Salmon roe-like amyloid deposition in a prolactinoma: a case report. Brain Tumor Pathol 2003, 20, 89–92. [Google Scholar] [CrossRef]
- Wiesli, P.; Brändle, M.; Brandner, S.; Kollias, S.S.; Bernays, R.L. Extensive spherical amyloid deposition presenting as a pituitary tumor. J Endocrinol Invest 2003, 26, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Hinton, D.R.; Polk, R.K.; Linse, K.D.; Weiss, M.H.; Kovacs, K.; Garner, J.A. Characterization of spherical amyloid protein from a prolactin-producing pituitary adenoma. Acta Neuropathol 1997, 93, 43–49. [Google Scholar] [CrossRef]
- Kuratsu, J.; Matsukado, Y.; Miura, M. Prolactinoma of pituitary with associated amyloid-like substances. Case report. J Neurosurg 1983, 59, 1067–1070. [Google Scholar] [CrossRef]
- Taniyama, H.; Kitamura, A.; Kagawa, Y.; Hirayama, K.; Yoshino, T.; Kamiya, S. Localized amyloidosis in canine mammary tumors. Vet Pathol 2000, 37, 104–107. [Google Scholar] [CrossRef]
- Yanamandra, K.; Alexeyev, O.; Zamotin, V.; Srivastava, V.; Shchukarev, A.; Brorsson, A.C.; Tartaglia, G. G.; Vogl, T.; Kayed, R.; Wingsle, G.; Olsson, J.; Dobson, C.M.; Bergh, A.; Elgh, F.; Morozova-Roche, L.A. Amyloid formation by the pro-inflammatory S100A8/A9 proteins in the ageing prostate. PLOS ONE 2009, 4, e5562. [Google Scholar] [CrossRef]
- Tekin, B.; Dasari, S.; Theis, J.D.; Vrana, J.A.; Murray, D.L.; Oglesbee, D.; Thompson, R.H.; Leibovich, B.C.; Boorjian, S.A.; Whaley, R.D.; Hernandez, L.H.; Jimenez, R.E.; Cheville, J.C.; Karnes, R.J.; Sukov, W.R.; Gupta, S. Mass spectrometry-based assessment of prostate cancer-associated crystalloids reveals enrichment for growth and differentiation factor 15. Hum Pathol 2023, 135, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Cross, P.A.; Bartley, C.J.; McClure, J. Amyloid in prostatic corpora amylacea. J Clin Pathol 1992, 45, 894–897. [Google Scholar] [CrossRef] [PubMed]
- Palangmonthip, W.; Wu, R.; Tarima, S.; Bobholz, S.A.; LaViolette, P.S.; Gallan, A.J.; Iczkowski, K.A. Corpora amylacea in benign prostatic acini are associated with concurrent, predominantly low-grade cancer. Prostate 2020, 80, 687–697. [Google Scholar] [CrossRef]
- Riba, M.; Campo-Sabariz, J.; Tena, I.; Molina-Porcel, L.; Ximelis, T.; Calvo, M.; Ferrer, R.; Martin-Venegas, R.; Del Valle, J.; Vilaplana, J.; Pelegri, C. Wasteosomes (corpora amylacea) of human brain can be phagocytosed and digested by macrophages. Cell Biosci 2022, 12, 177. [Google Scholar] [CrossRef]
- Maji, S.K.; Perrin, M.H.; Sawaya, M.R.; Jessberger, S.; Vadodaria, K.; Rissman, R.A.; Singru, P.S.; Nilsson, K.P.; Simon, R.; Schubert, D.; Eisenberg, D.; Rivier, J.; Sawchenko, P.; Vale, W.; Riek, R. Functional amyloids as natural storage of peptide hormones in pituitary secretory granules. Science 2009, 325, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Pioselli, B.; Salomone, F.; Mazzola, G.; Amidani, D.; Sgarbi, E.; Amadei, F.; Murgia, X.; Catinella, S.; Villetti, G.; De Luca, D.; Carnielli, V.; Civelli, M. Pulmonary surfactant: A unique biomaterial with life-saving therapeutic applications. Curr Med Chem 2022, 29, 526–590. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, M.; Thyberg, J.; Naslund, J.; Eliasson, E.; Johansson, J. Amyloid fibril formation by pulmonary surfactant protein C. FEBS Lett 1999, 464, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, H.; Yoshinouchi, T.; Watanabe, R.; Fujita, J.; Takahara, J.; Ohtsuki, Y. Immunohistochemical study of a patient with diffuse pulmonary corpora amylacea detected by open lung biopsy. Intern Med 1999, 38, 900–903. [Google Scholar] [CrossRef] [PubMed]
- Otzen, D.; Riek, R. Functional amyloids. Cold Spring Harb Perspect Biol 2019, 11, a033860. [Google Scholar] [CrossRef]
- Ishii, W.; Matsuda, M.; Nakamura, N.; Katsumata, S.; Toriumi, H.; Suzuki, A.; Ikeda, S. Phenol Congo red staining enhances the diagnostic value of abdominal fat aspiration biopsy in reactive AA amyloidosis secondary to rheumatoid arthritis. Intern Med 2003, 42, 400–405. [Google Scholar] [CrossRef]
- Kametani, F.; Haga, S. Accumulation of carboxy-terminal fragments of APP increases phosphodiesterase 8B. Neurobiol Aging 2015, 36, 634–637. [Google Scholar] [CrossRef]
- Abe, R.; Katoh, N.; Takahashi, Y.; Takasone, K.; Yoshinaga, T.; Yazaki, M.; Kametani, F.; Sekijima, Y. Distribution of amyloidosis subtypes based on tissue biopsy site—Consecutive analysis of 729 patients at a single amyloidosis center in Japan. Pathol Int 2021, 71, 70–79. [Google Scholar] [CrossRef]
Figure 1.
Representative microphotographs of pulmonary and prostatic CAs and CTAVN-associated CAs in the heart. (a, d, g) Pulmonary CA Case 1; (b, e, h) Prostatic CA Case 1; (c, f, i) CTAVN Case 1. (a–c) Hematoxylin and eosin (H&E) staining; (d–f) Phenol Congo red (pCR) staining with a bright field; (g–i) pCR staining under polarized light. (a–c) All CAs are round with eosinophilic deposits showing concentric laminations upon H&E staining. Note that macrophages are observed around CAs in the lung (a), and multinucleated giant cells are observed around CAs in the prostate (b). (d, e, g, h) Almost all pulmonary and prostatic CAs exhibit congophilia with apple-green birefringence under polarized light. Although CTAVN-associated CAs show congophilic deposits (f), some CAs (arrow) lack apple-green birefringence under polarized light (arrowheads indicate CAs showing typical apple-green birefringence). Scale bar = 200 μm (a–i).
Figure 1.
Representative microphotographs of pulmonary and prostatic CAs and CTAVN-associated CAs in the heart. (a, d, g) Pulmonary CA Case 1; (b, e, h) Prostatic CA Case 1; (c, f, i) CTAVN Case 1. (a–c) Hematoxylin and eosin (H&E) staining; (d–f) Phenol Congo red (pCR) staining with a bright field; (g–i) pCR staining under polarized light. (a–c) All CAs are round with eosinophilic deposits showing concentric laminations upon H&E staining. Note that macrophages are observed around CAs in the lung (a), and multinucleated giant cells are observed around CAs in the prostate (b). (d, e, g, h) Almost all pulmonary and prostatic CAs exhibit congophilia with apple-green birefringence under polarized light. Although CTAVN-associated CAs show congophilic deposits (f), some CAs (arrow) lack apple-green birefringence under polarized light (arrowheads indicate CAs showing typical apple-green birefringence). Scale bar = 200 μm (a–i).
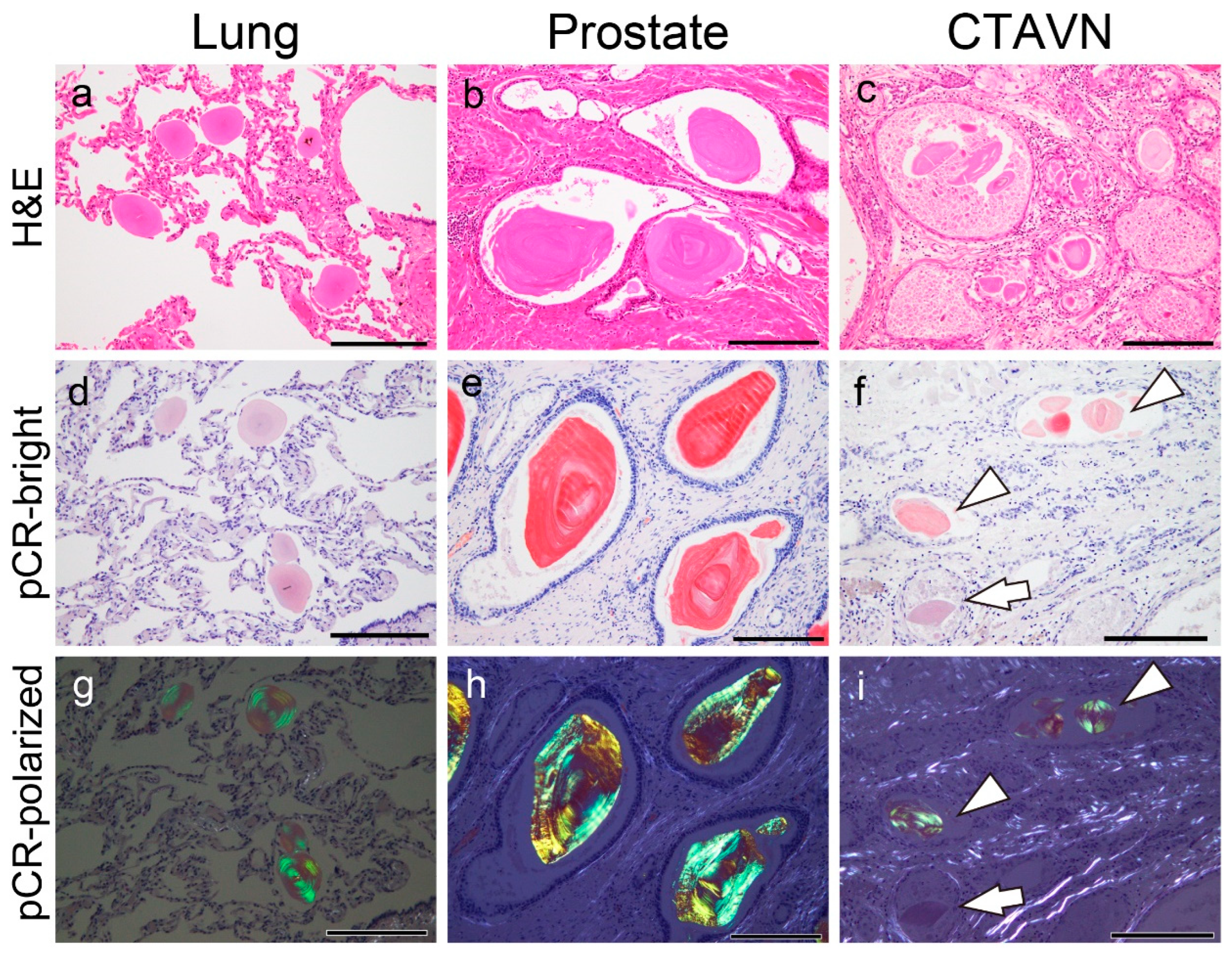
Figure 2.
Representative proteomics results and immunohistochemistry micrographs based on these results in the lung. (a) Proteins identified in the CAs using laser microdissection and liquid chromatography– tandem mass spectrometry (LMD and LC-MS/MS) in pulmonary CA Case 1; immunohistochemistry for β2-microglobulin (β2-MG) (b); lysozyme C (c); pulmonary surfactant protein A (PSP-A) (d); PSP-B (e); lactoferrin (f); and p62 (g). (a) In addition to some amyloid-associated proteins (shown in blue), five amyloidogenic proteins were detected [13]. Of these, immunoreactivity for β2-MG (b), lysozyme C, PSP-A (d), and PSP-B (e) was confirmed, whereas no immunoreactivity for lactoferrin was observed (f). The macrophages were also positive for β2-MG and lysozyme C (insets in panels b and c). Weak p62 immunoreactivity was observed in the central area of some CAs. The emPAI is the exponentially modified protein abundance index, which is used as an index for estimating relative protein quantification in mass spectrometry-based proteomic analyses. Scale bar = 200 μm (b–g).
Figure 2.
Representative proteomics results and immunohistochemistry micrographs based on these results in the lung. (a) Proteins identified in the CAs using laser microdissection and liquid chromatography– tandem mass spectrometry (LMD and LC-MS/MS) in pulmonary CA Case 1; immunohistochemistry for β2-microglobulin (β2-MG) (b); lysozyme C (c); pulmonary surfactant protein A (PSP-A) (d); PSP-B (e); lactoferrin (f); and p62 (g). (a) In addition to some amyloid-associated proteins (shown in blue), five amyloidogenic proteins were detected [13]. Of these, immunoreactivity for β2-MG (b), lysozyme C, PSP-A (d), and PSP-B (e) was confirmed, whereas no immunoreactivity for lactoferrin was observed (f). The macrophages were also positive for β2-MG and lysozyme C (insets in panels b and c). Weak p62 immunoreactivity was observed in the central area of some CAs. The emPAI is the exponentially modified protein abundance index, which is used as an index for estimating relative protein quantification in mass spectrometry-based proteomic analyses. Scale bar = 200 μm (b–g).
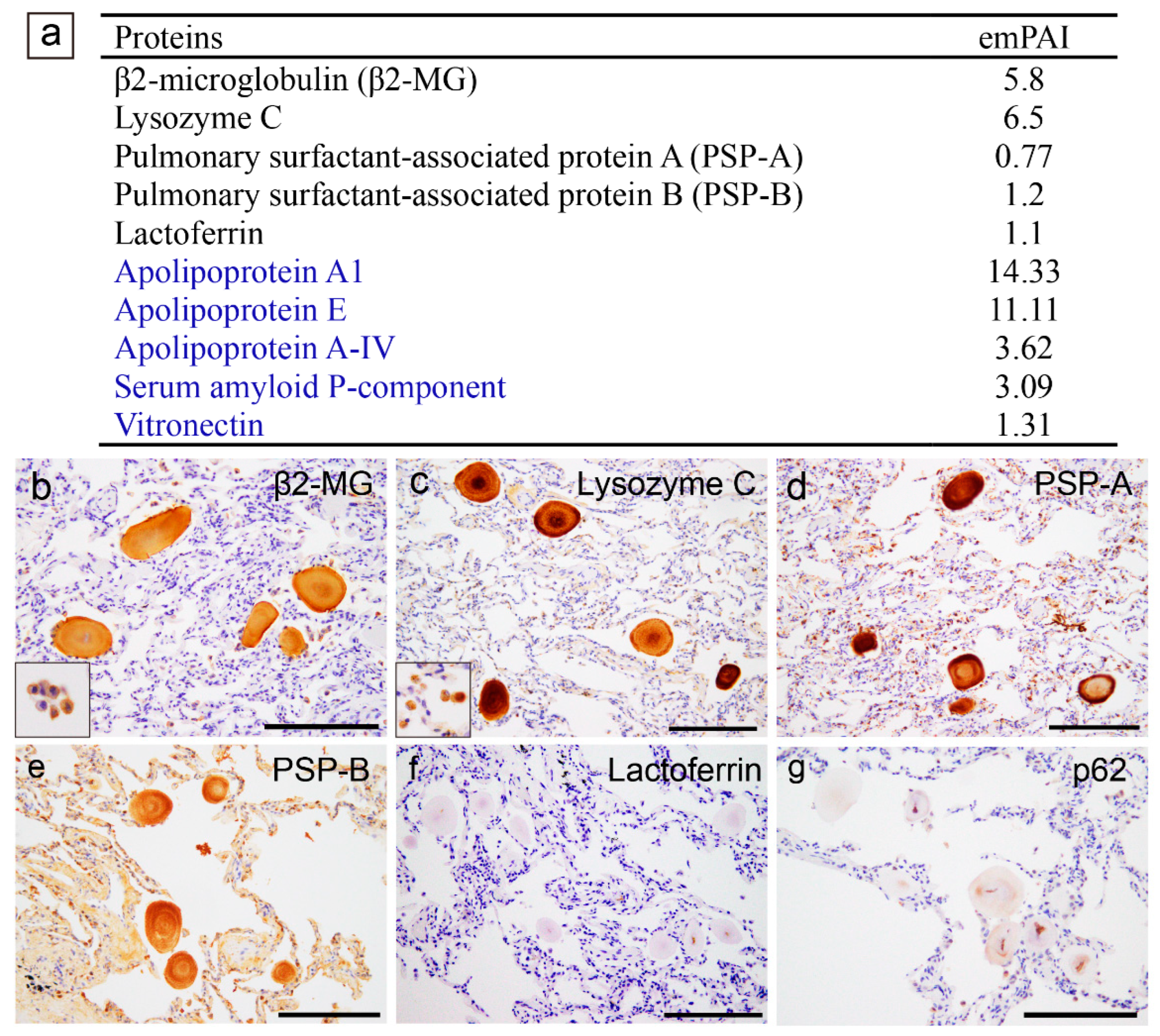
Figure 3.
Representative proteomics results and immunohistochemistry micrographs based on the results in the prostate. (b–d) CAs observed within the normal gland area; (e–j) Eosinophilic deposits observed within the acinar adenocarcinoma glands. (a) Proteins identified in the CA lesions using LMD and LC-MS/MS in prostatic CA Case 1; Immunohistochemistry for lysosome C (b, h); lactoferrin (c, i); p62 (d, j); H&E staining (e); pCR staining under a bright field (f) and a polarized field (g). (a) In addition to some amyloid-associated proteins (shown in blue), three amyloidogenic proteins were detected. VA lesions were positive for lysozyme C (b) and lactoferrin (c). the macrophages are also positive for lysozyme C (inset in panel b). p62 immunoreactivity in the central area of some CAs (d). Within the tumor glands, eosinophilic deposits, including crystalloids (arrow), are evident. (f, g) Some of the deposits show weak to moderate congophilia and exhibit apple-green birefringence under polarized light (arrowhead). Crystalloids are weakly positive for pCR, but do not exhibit apple-green birefringence under polarized light (arrowhead). In addition, these eosinophilic deposits were not immunoreactive for lysozyme C (h), lactoferrin (i), or p62 (j). Scale bar = 200 μm (b–d); 100 μm (e–j).
Figure 3.
Representative proteomics results and immunohistochemistry micrographs based on the results in the prostate. (b–d) CAs observed within the normal gland area; (e–j) Eosinophilic deposits observed within the acinar adenocarcinoma glands. (a) Proteins identified in the CA lesions using LMD and LC-MS/MS in prostatic CA Case 1; Immunohistochemistry for lysosome C (b, h); lactoferrin (c, i); p62 (d, j); H&E staining (e); pCR staining under a bright field (f) and a polarized field (g). (a) In addition to some amyloid-associated proteins (shown in blue), three amyloidogenic proteins were detected. VA lesions were positive for lysozyme C (b) and lactoferrin (c). the macrophages are also positive for lysozyme C (inset in panel b). p62 immunoreactivity in the central area of some CAs (d). Within the tumor glands, eosinophilic deposits, including crystalloids (arrow), are evident. (f, g) Some of the deposits show weak to moderate congophilia and exhibit apple-green birefringence under polarized light (arrowhead). Crystalloids are weakly positive for pCR, but do not exhibit apple-green birefringence under polarized light (arrowhead). In addition, these eosinophilic deposits were not immunoreactive for lysozyme C (h), lactoferrin (i), or p62 (j). Scale bar = 200 μm (b–d); 100 μm (e–j).
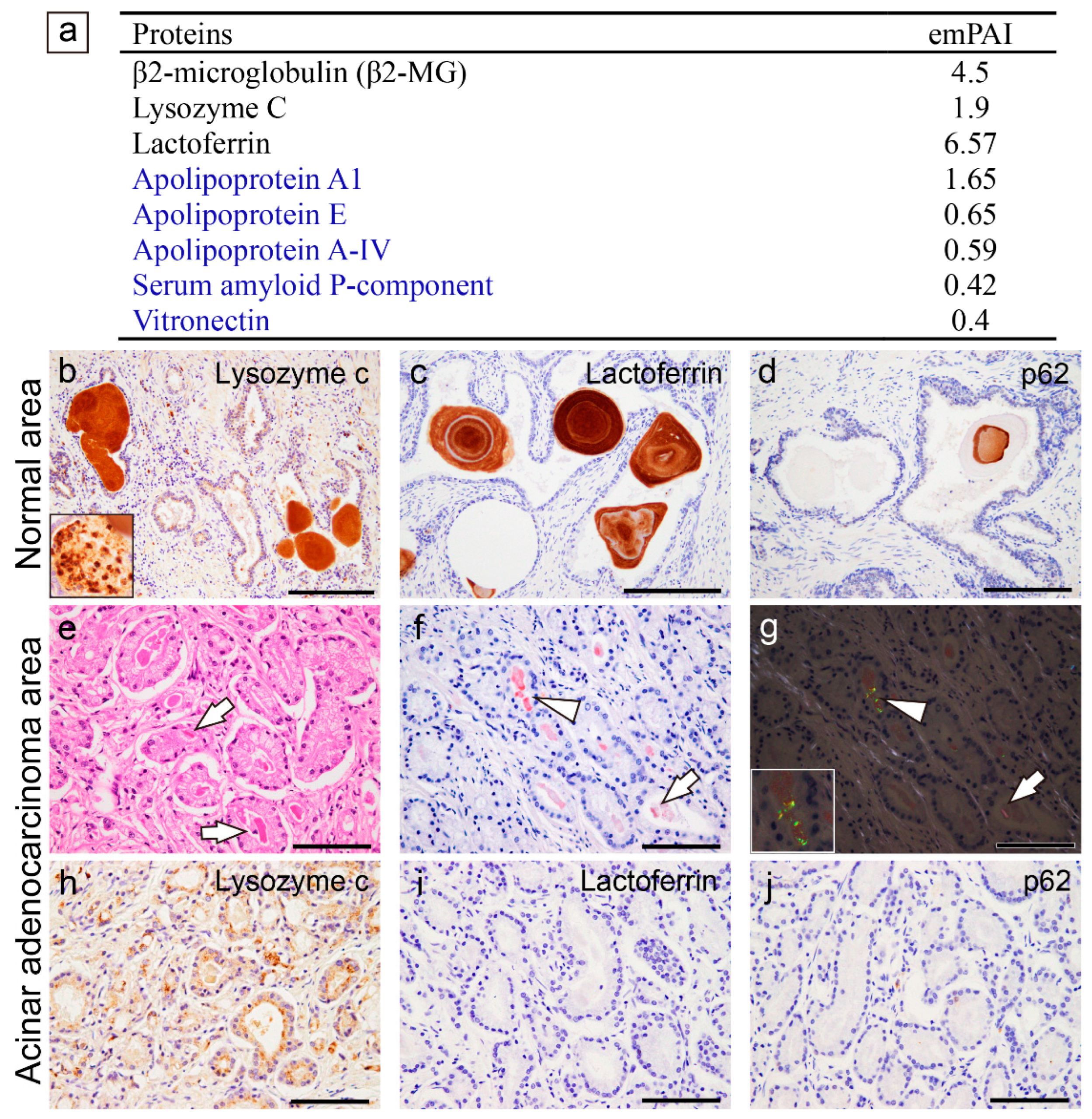
Figure 4.
Proteomics results for the amyloidogenic proteins commonly observed in pulmonary and prostatic CAs. (a, c, e) Lung; (b, d, f) prostate; (a, b) β2-MG; (c, d) lysozyme C; and (e, f) lactoferrin. Detected peptides, which have a peptide score of 30 or higher by MASCOT analysis, are shown in red. (a–d) Regarding β2-MG and lysozyme C, similar peptides were detected in the lung and prostate. (e, f) In contrast, for lactoferrin, only a few peptide sequences were identified in the lungs, whereas several sequences were identified in the prostate.
Figure 4.
Proteomics results for the amyloidogenic proteins commonly observed in pulmonary and prostatic CAs. (a, c, e) Lung; (b, d, f) prostate; (a, b) β2-MG; (c, d) lysozyme C; and (e, f) lactoferrin. Detected peptides, which have a peptide score of 30 or higher by MASCOT analysis, are shown in red. (a–d) Regarding β2-MG and lysozyme C, similar peptides were detected in the lung and prostate. (e, f) In contrast, for lactoferrin, only a few peptide sequences were identified in the lungs, whereas several sequences were identified in the prostate.
Figure 5.
Representative proteomics results and immunohistochemistry micrographs of the heart. (a–c, f) CTAVN-Case 1; (d, g) CTAVN-Case 2; (e, h) CTAVN-Case 3. (a) Proteins identified in the CAs using LMD and LC-MS/MS in CTAVN-Case 1; (b) Detected peptide sequences of olfactomedin 4; Immunohistochemistry for olfactomedin 4 (c–e); p62 (f–h). (a, b) Consistent with our previous immunohistochemistry-based study [4], olfactomedin 4 was identified in CAs and CA-like deposits within CTAVN glands. No amyloid-associated proteins were identified. Immunohistochemistry for olfactomedin 4 was strongly positive in Case 1 (c), and weakly positive in Case 2 (d) and Case 3 (e) (arrowheads indicate olfactomedin 4-positive deposits). In contrast, immunohistochemistry for p62 was negative to partially positive in Case 1 (f), whereas it was negative in Cases 2 and 3 (arrowheads indicate olfactomedin 4-positive deposits and arrows indicate negative deposits). Scale bar = 200 μm (c–h).
Figure 5.
Representative proteomics results and immunohistochemistry micrographs of the heart. (a–c, f) CTAVN-Case 1; (d, g) CTAVN-Case 2; (e, h) CTAVN-Case 3. (a) Proteins identified in the CAs using LMD and LC-MS/MS in CTAVN-Case 1; (b) Detected peptide sequences of olfactomedin 4; Immunohistochemistry for olfactomedin 4 (c–e); p62 (f–h). (a, b) Consistent with our previous immunohistochemistry-based study [4], olfactomedin 4 was identified in CAs and CA-like deposits within CTAVN glands. No amyloid-associated proteins were identified. Immunohistochemistry for olfactomedin 4 was strongly positive in Case 1 (c), and weakly positive in Case 2 (d) and Case 3 (e) (arrowheads indicate olfactomedin 4-positive deposits). In contrast, immunohistochemistry for p62 was negative to partially positive in Case 1 (f), whereas it was negative in Cases 2 and 3 (arrowheads indicate olfactomedin 4-positive deposits and arrows indicate negative deposits). Scale bar = 200 μm (c–h).
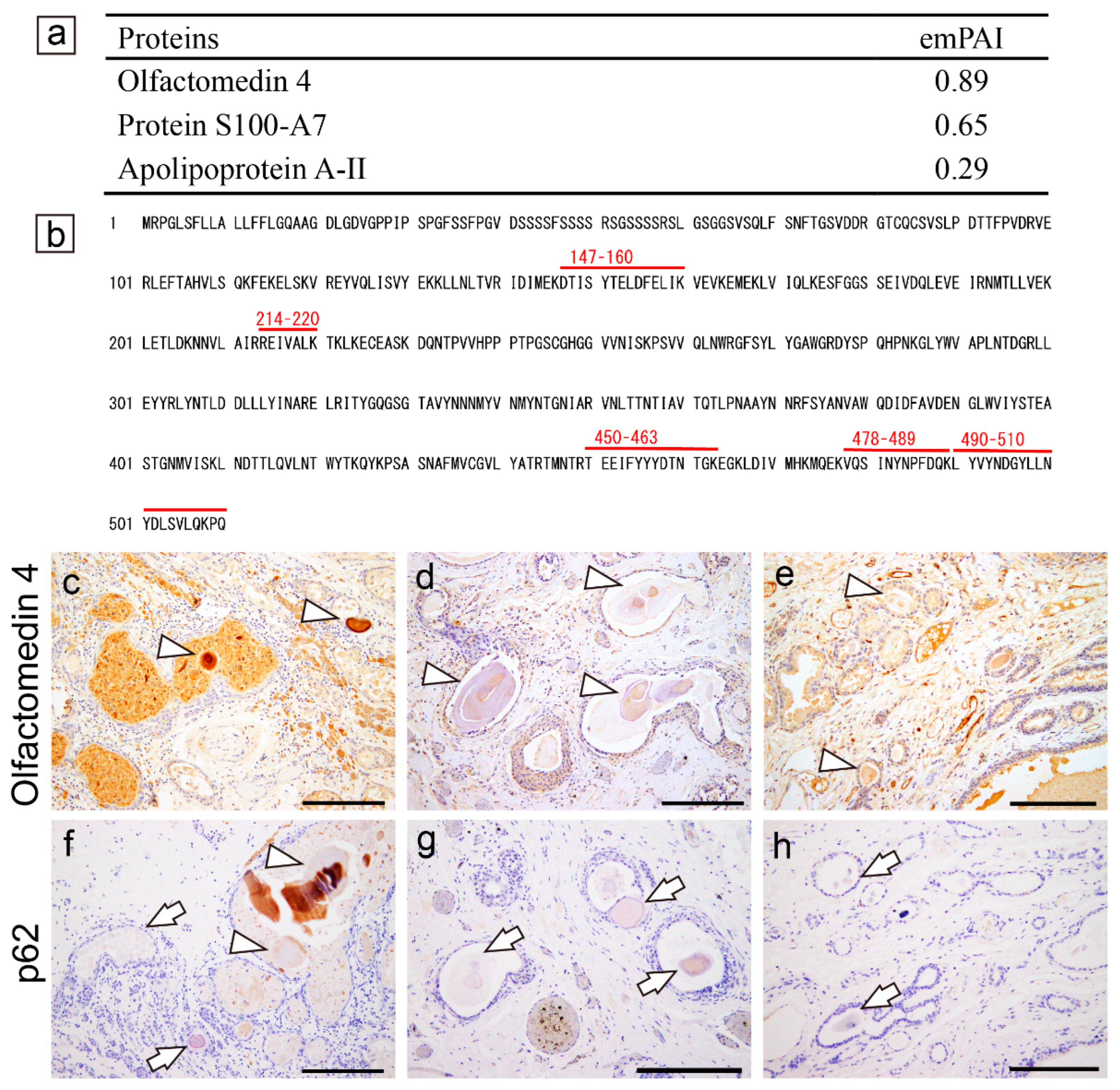
Figure 6.
Representative microphotographs of spheroid-type amyloid deposition in the kidney. (a, b) H&E staining; pCR staining under a bright field (c) and polarized light (d); immunohistochemistry for Igλ (e); p62 (f). (a, b) Spheroid-type amyloid deposits are evident in the interstitium (a) and in the tubules (b, arrow). (c, d) These deposits were positive for pCR (c) and exhibit apple-green birefringence under polarized light (d). (e) As shown in our previous report, the deposits are positive by immunohistochemistry for Igλ. (f) In contrast, immunohistochemistry for p62 was negative. Scale bar = 200 μm (e); 100 μm (a–d).
Figure 6.
Representative microphotographs of spheroid-type amyloid deposition in the kidney. (a, b) H&E staining; pCR staining under a bright field (c) and polarized light (d); immunohistochemistry for Igλ (e); p62 (f). (a, b) Spheroid-type amyloid deposits are evident in the interstitium (a) and in the tubules (b, arrow). (c, d) These deposits were positive for pCR (c) and exhibit apple-green birefringence under polarized light (d). (e) As shown in our previous report, the deposits are positive by immunohistochemistry for Igλ. (f) In contrast, immunohistochemistry for p62 was negative. Scale bar = 200 μm (e); 100 μm (a–d).
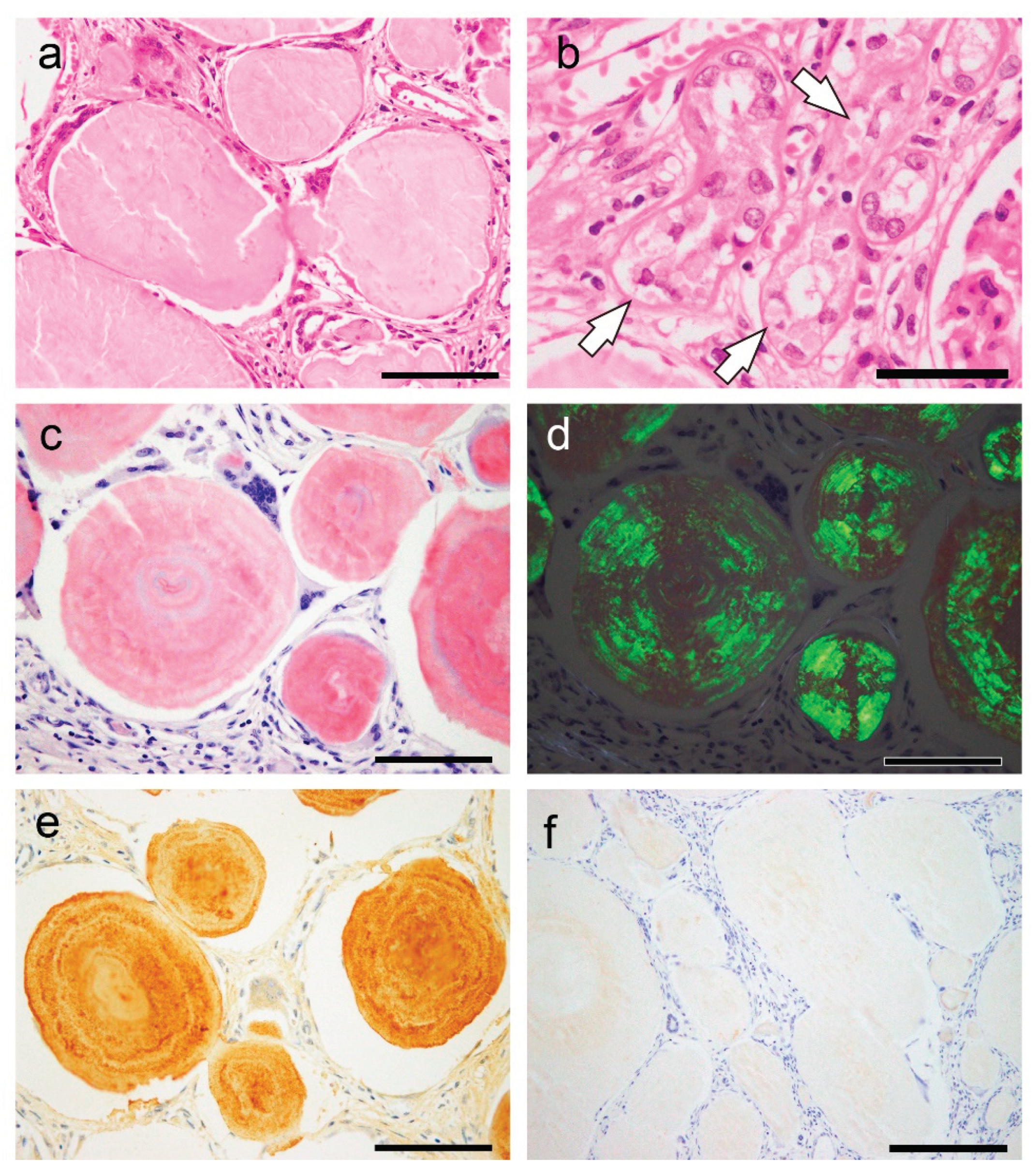

Table 1.
Demographic data associated with cases of corpora amylacea (CA).
| Pulmonary CA cases | Prostatic CA cases | CTAVN cases | |||||||
| Case # | Case 1* | Case 2 | Case 3 | Case 1* | Case 2 | Case 3 | Case 1 | Case 2 | Case 3 |
| Age | 75 | 90 | 85 | 75 | 88 | 45 | 36 | 45 | 76 |
| Sex | M | F | F | M | M | M | F | M | M |
| Cause of death | HyT | MN | Drowning | HyT | ACD | KA | SCD | SCD | SCD |
| Dialysis | None | None | None | None | None | None | None | None | None |
Abbreviations: ACD, acute cardiac dysfunction; CTAVN, cystic tumor of the atrioventricular node; F, female; HyT, hypothermia; KA, ketoacidosis; M, male; MN, malnutrition; SCD, sudden cardiac death. * Same patient.
Table 2.
Summary of previous reports of spheroid-type amyloid deposition (STAD).
| Location | S/L | Concomitant findings | Immunohistochemical analysis | Proteomic analysis |
| Stomach and small intestine [14] | S | Bronchiectasis, FMF, and renal failure | Pos: AA; Neg: β2MG, TTR, Igκ, Igλ, CD68 | NE |
| Small intestine [15,16] | L | Polypoid lesions | Pos: Igλ; Neg: AA, β2MG, TTR, Igκ [15] Pos: AP, AA, Igκ, Igλ (uneven) [16] |
NE |
| Vater ampulla [17] | L | NET | NE | NE |
| Colon, TI [12] | L | Adenocarcinoma | NE | ALλ |
| Colon [18,19,20] | L | Ulcerative lesions [18]and rounded lesions [19] | Pos: Igλ [18] | NE |
| Liver [21,22,23,24,25,26] | S/L | Various diseases (See [21,22]) | Pos: AP, AA; Neg: Igκ, Igλ [21] Pos: none; Neg: AP, AA, UB, TTR, Igκ, Ig [23] Pos: AA; Neg: β2MG, TTR, Igκ, Igλ [24] Pos: LECT2 or Ig; Neg: AA, β2MG, TTR [26] |
ALECT2 or AL [26] |
| Sino-nasal tract [27] | L | Nasal mass | Pos: Igκ and Igλ (κ>λ) | NE |
| Parotid gland [28] | L | AACC | NE | NE |
| URT [29] | S/L | Plasmacytoma | NE | NE |
| Bronchus [11] | L | Erythematous mass | Pos: Lac; Neg: AA, β2MG, TTR, Igκ, Igλ | ALac |
| Bone [30,31,32] | L | Myeloma, malignant lymphoma (see [30]) | Pos: Igκ or Igλ; Neg: AA [30] | NE |
| Bone marrow [33] | S | PCP | Pos: Igλ | NE |
| Ureter [34] | L | Hydronephrosis | NE (likely AA) | NE |
| Kidney [10] | S | PCP | Pos: Igλ; Neg: AA, β2MG, TTR, Igκ | ALλ |
| Uterine cervix [35,36,37] | L | Smooth mass [36], SCC [37],[38] | Pos: AA; Neg: CK, Igκ, Igλ [35] Pos: CK; Neg: AA, TTR, Igκ, Igλ [36,37] |
NE |
| Pituitary gland [38,39,40,41,42,43,44] | L | Prolactinoma | Pos: PRL; Neg: CK, vimentin, GFAP, GH, FSH, LH, TSH, ACTH, β-A4 [39,41,42] | NE |
| Breast [45]* | L | Mammary tumor | Pos: α-casein, Lac; Neg: AA, TTR, CK, Igκ, Igλ | NE |
Abbreviations: AA, (serum) amyloid A; ACC, acinic cell carcinoma; ACTH, adenocorticotropic hormone; AP, amyloid P component; CK, cytokeratin; FMF, familial Mediterranean fever; FSH, follicle-stimulating hormone; GFAP, glial fibrillary acidic protein; GH, growth hormone; Ig, immunoglobulin (light chain); L, localized; Lac, lactoferrin; LECT2, leukocyte chemotactic factor 2; LH, luteinizing hormone; NE, not evaluated; Neg, negative; NET, neuroendocrine tumor; PCP, plasma cell proliferation; Pos: positive; PRL, prolactin; S, systemic; SCC, squamous cell carcinoma; TI, terminal ileum; TSH, thyroid-stimulating hormone; UB, ubiquitin; URT, upper respiratory tract. * Found in 2 dogs.
Table 3.
Summary of the properties of amyloid-forming CAs and STADs.
| Prostatic-CA | Pulmonary-CA | CTAVN-CA | STAD | |
| Congophilia | Strong | Moderate–strong | Weak–moderate | Strong |
| Strength of the AGBR | Strong | Strong | Weak–moderate | Strong |
| Macrophages | Positive | Positive | Negative–positive | Positive |
| Presence of CPs | Positive | Positive | Negative | Negative |
| Presence of AAPs | Positive | Positive | Negative | Positive |
| p62-IR | Positive (focal) | Positive (focal) | Negative–Positive (focal) | Negative |
Abbreviations: AAPs, amyloid-associated proteins; AGBR, apple-green birefringence under polarized light; CPs, common proteins (β2-microglobulin and lysozyme C); IR, immunoreactivity.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



