
Submitted:
05 March 2024
Posted:
06 March 2024
You are already at the latest version
Abstract
Abstract:
Chikungunya virus (CHIKV) is an emerging pathogen with widespread distribution in regions of Africa, India, and Asia that threatens to spread into temperate climates following the introduction of its major vector, Ae. albopictus. Recent cases have been documented in Europe, the Caribbean, and the Americas. CHIKV causes a disease frequently misdiagnosed as Dengue fever, with potentially life-threatening symptoms that can result in long term debilitating arthritis. There have been ongoing investigations of possible therapeutic interventions for both acute and chronic symptoms, but to date none have proven effective in reducing the severity or lasting effects of CHIKV disease. Recently, a promising vaccine candidate has received accelerated FDA approval, indicating the importance of remedies to this emerging worldwide health threat. Nonetheless, therapeutic interventions for CHIKV and other mosquito borne virus diseases are urgently needed yet remain elusive. The increasing risk of spread from endemic regions via human travel and commerce, coupled with the absence of a vaccine or FDA approved therapeutic, puts a significant proportion of the world population at risk for this disease. In this report we explore the possibility of using SOFA (Specific On/oFf Adapter) Hepatitis Delta Virus Ribozymes (HDV-Rzs) as antivirals in cells infected with CHIKV. The results we obtained suggest there could be some role in using these SOFA HDV molecules as antiviral therapies for not only CHIKV, but potentially other viruses as well.
Keywords:
Chikungunya
; CHIKV
; Hepatitis Delta
; Ribozyme
; Antiviral
1. Introduction
Chikungunya virus (CHIKV) is a positive sense RNA virus (Family Alphaviridae) that is endemic to Asia, India, and parts of Africa, and is propagated by the bite of female Aedes mosquitoes, principally Ae. Aegypti and Ae. Albopictus. Blood-borne transmission is possible [1,2,3] including vertical transmission [4,5]. The virus produces a disease that is characterized by abrupt onset of fever, myalgia, nausea, headache, fatigue, rashes, and debilitating arthralgia [6]. Other complications associated with CHIKV include myocarditis, hepatitis, ophthalmological, and neurological disorders [7].
CHIKV was first described in 1952 during an outbreak in southern Tanzania and has been identified in almost 40 countries to date. Recent epidemics include La Reunion Island in 2005–2006, with 255,000 cases among a total population of 750,000 [8], India in 2006–2007, with 1.4 to 6.5 million estimated cases [9,10], and the Philippines in 2013, with 180 cases. Cases of CHIKV have also been reported in parts of Europe as a result of infected individuals travelling from endemic regions [11,12,13], and most recently in the Caribbean [14]. The rapid advancement of CHIKV worldwide corresponds to the spread of invasive Ae. albopictus and the advent of a CHIKV strain adapted to spread by this vector [15].
A significant amount of work is underway to develop a vaccine for CHIKV. This includes strategies such as chimeric vaccines [16], recombinant CHIKV vaccines [17], adenovirus-based vaccines [18], inactivated virus [19], virus like particles [20], and live attenuated vaccines [21]; see [22] for a detailed review). Recently, a promising vaccine candidate entered clinical trials [23], however, even with a viable vaccine there remain significant hurdles to effective prevention, containment, and control of any disease, especially in underdeveloped countries. There is a considerable amount of research devoted to the development of antivirals that may be useful as therapeutics for CHIKV disease (see [24] for review) including interferon [25], deubiquinating enzyme inhibitors [26], sphingosine kinase inhibitors [27], and radicol [28] and defective viral genomes [29].
CHIKV is a small (60-70 nm-diameter) icosahedral, enveloped virus. The nucleocapsid contains a single stranded (+) sense genomic RNA of 11.8 kb that is organized 5’ cap-nsP1-nsP2-nsP3-nsP4-(Junction region)-C-E3-E2-6K-E1-poly (A)-3’. The non-structural proteins (nsP1, nsP2, nsP3 and nsP4) are encoded by the full-length genomic RNA and are responsible for replication of the viral genome [30]. A separate sub genomic RNA is produced during replication which encodes the structural proteins capsid, E3, E2, 6k, and E1 [30].
The two RNA species that are involved in the replication of CHIKV have proven to be excellent targets for RNA inhibition strategies. Antiviral strategies attempted thus far include RNA interference (RNAi), using small interfering RNAs (siRNA) [31], or small hairpin RNAs (sh-RNA) [32] which have already demonstrated some success. While RNA interference can be an effective mechanism to suppress viral replication, this strategy requires targeting of highly conserved sequences of at least 21-23 nucleotides, which, in the case of CHIKV, limits the potential number of suitable target sites available. A single point mutation in this target site can result in decreased efficacy and escape mutants [33], and the longer the target site, the more potential for these mutations. In addition, RNAi requires continuous synthesis of large amounts of ds-RNA in order to activate and maintain the RNAi machinery and effectively suppress viral replication [34] and some viruses, like CHIKV, may replicate at a rate that allows overcoming the RNAi response [35].
Catalytic RNAs, ribozymes, can destroy viral RNAs in a targeted fashion leading to the elimination of viral genomic RNAs either entering the cell or produced during replication. Some of these catalytic RNAs are able to effectively regenerate, lessening the need for their continual production. In addition, therapeutic potential as delivered antivirals for some types of catalytic RNAs (hammerhead and hepatitis delta ribozymes, in particular) has been previously demonstrated [36,37,38]. In our lab we have been exploring various ribozymes for different applications against arboviruses [39,40,41,42,43].
This report presents evidence that the Hepatitis Delta Virus Ribozyme (HDV-Rz) may have potential as a deliverable antiviral for the control of CHIKV, and potentially other vector borne diseases. HDV-Rzs are integral in HDV replication, generating monomeric genomes and antigenomes with the antisense version capable of acting in a trans fashion (see [44] for review). This ribozyme seems to be a suitable candidate antiviral inactivation system for human cells (see [45] for review) and has previously demonstrated inhibition of gene expression in vivo [46,47,48]. In this study we explored the potential of a SOFA delta ribozyme design [49,50] to suppress the production of infectious CHIKV in Vero cells by targeting conserved CHIKV sequences, and demonstrated that transfection of these in vitro expressed effector RNAs following infection of Vero cells can significantly reduce CHIKV infection in cell cultures.
2. Materials and Methods
Virus
Due to the unavailability of BSL3 facilities we used the CHIKV 181/25 vaccine strain for these studies, which was kindly provided by Dr. Scott Weaver, University of Texas Medical Branch, Galveston, TX). The virus was propagated in Vero cell cultures originally obtained from ATCC, and maintained in DMEM (Sigma-Aldrich, USA) supplemented with 10% FBS (Atlanta Biologicals) and 1% Non-essential amino acids (Gibco, USA) at 37° C with 5% CO2 and passaged every 4 days. Virus was maintained by infection of cells at an MOI (Multiplicity of Infection) of 2 in serum free DMEM for 2 hours with gradual shaking at 37° C. After 2 hours 7 ml of DMEM with 10% FBS was added and incubated at 37° C for 2 days, and supernatants harvested and stored at -80° C. The stock virus was titered using TCID50 as described below.
Design of SOFA HDV-Rz Expression Plasmids
The SOFA HDV-Rz we employed was based upon the original construct of Bergeron and Perreault [49] and Levesque et al. [50]. Our antisense transacting SOFA HDV-Rzs modified from the published sequence with target sites selected from conserved sequences having a
-4YHRH-1 configuration, where Y=U or C, H=U,C, or A, and R=G or A (Deschenes et al., 2000; Figure 1), from CLUSTALX alignments of 100 CHIKV strains available from the Gene Bank database. Target 124 does not conform to this consensus at position -3, 694 deviates at -4 and -3, and 2504 and 7487 at position -4. Target 194 conforms at all positions (Figure 1).
Ribozyme expression plasmids were synthesized as T7-promoted expression cassettes. We also constructed inactive ribozymes having the same P1, biosensor, and blocker sequences as the active counterparts, but with a destabilized P1.1 helix through the replacement of GC nucleotides with A’s [52], and an inactive active site by deletion of C75 [53,54].
Preparation of Delta Ribozyme RNA
SOFA HDV-Rz RNA was prepared by digestion of 1µg plasmid DNA with NotI-HF (New England Biolabs) followed by purification with the E.Z.N.A. MicroElute DNA Clean-Up Kit (Omega Biotek). DNA was eluted with 20 µl nuclease free water. The in vitro transcription reactions were performed using Invitrogen’s MEGAscript T7 Transcription kit according to the manufacturer’s directions, with 2 µL RNase Inhibitor, 6 µL digested and purified DNA, a final volume of 20 µL, and incubation overnight at 37° C. Following DNase treatment for 30 min, RNA was purified using phenol:chloroform:isoamyl alcohol (25:24:1; Sigma) extraction followed by ethanol precipitation with five 70 % ethanol washes and final suspension in nuclease free water. Purified RNA was used immediately for transfection.
CHIKV Infection Inhibition Assays
The effectiveness of SOFA HDV-Rz suppression of CHIKV infections was determined using a Caspase 3/7 assay kit (Promega) as previously described (Mishra et al., 2016). Vero Cells were plated at 4 x 10⁵ in six well plates and infected with CHIKV at MOI 0.0001 followed by incubation in DMEM (Sigma) with 2 % FBS for 4 hours. The cells were then washed twice with serum free media and 2 µg of each SOFA HDV ribozyme RNA, and nuclease free water control were transfected into one infected and one uninfected well each, using Lipofectamine 2000 (Invitrogen) following the manufacture’s recommendations. The transfection media was removed and replaced with 1.5 mL DMEM 2% FBS at 7.5 hpi and cells incubated at 37° C 5% CO2. After 48 hpi, 500 µL media was removed, mixed with 500 µL FBS, and stored at -80° C for later assay. The remaining media was removed from each well and 600 µL of a 1:1 ratio PBS pH 7.4 (GIBCO) and Caspase-Glo 3/7 Reagent (Promega) was added. The plate was incubated in the dark for 10 min with gentle rocking then an additional 5min with no rocking. The 600 µL was then divided across three wells of a white walled 96-well plate and luminescence read with a LMaxII384 luminometer. The entire experiment, including RNA preparation, was repeated in triplicate. Results were expressed as relative raw Luminescence reported by the plate reader (RLU). Statistical analysis was performed within IBM SPSS via an ANOVA test followed by Tukey’s ad-hoc test to determine significance. Power was determined using a univariate analysis of variance within IBM SPSS with alpha set to 0.05 for the experiments.
TCID50-IFA Analysis of Chikungunya Viruses
Viral supernatants were 10-fold serially diluted to 10-8 in 96 well plates and trypsinised Vero cells were added (2 x 104/well) to the wells. After four days post infection (dpi), cells were fixed with Acetone: DPBS (3:1) and stained with (1:100) diluted CHIKV antibody (Catalog No: 3583 Virostat, USA). After incubation at 37° C for 40 minutes, stained cells were washed three times with 1x Dulbecco’s PBS (DPBS) and diluted (1:200) biotinylated anti-mouse antibody (RPN1001, GE Healthcare Life Sciences) was added to each well and again incubated at 37° C for 40 minutes. The cells were washed again three times to remove the unbound antibodies. Conjugated streptavidin-FITC (Catalog No: 434311, Invitrogen, USA) was added to the wells at a concentration of 1:150 and incubated for 10 minutes at 37° C. After washing two times with 1xDPBS and one time with distilled water, a drop of DABCO:glycerol (3.5 grams DABCO (Sigma- Aldrich,USA) + 10mL of 1xDPBS and 90 mL of glycerol) mix was added to each well and cytoplasmic fluorescence was observed using a Nikon Diaphot inverted fluorescent microscope. The number of positive green fluorescent wells were counted and the virus titers calculated according to Karber’s method [55]. The titer was expressed as log10TCID50/ml. Data was log10 transformed for analysis as the data was logarithmic and not evenly distributed around the mean, and after log10 transformation the data was evenly distributed. Statistical analysis was performed on log10 transformed data within IBM SPSS via an ANOVA test followed by Tukey’s ad-hoc test to determine significance. Power was determined using a univariate analysis of variance within IBM SPSS with alpha set to 0.05 for the experiments.
3. Results
- 3.1.1. Structure of SOFA HDV Ribozymes
The SOFA module of our HDV-Rzs acts as a switch to control the activity of the ribozyme by inhibiting the cleavage activity of the ribozyme with a blocker sequence that binds the P1 domain preventing formation of the catalytic configuration (Figure 2). The blocker is adjacent to the biosensor sequence, which recognizes the target sequence on the substrate RNA. Once the biosensor binds the target, the blocker sequence is released from the P1 domain, permitting formation of the active conformation and cleavage of the target RNA [56].
We synthesized T7 promoted expression cassettes of active G124, G194, G694, G2504 and G7487 SOFA HDV ribozymes, and inactive counterparts G124IN, G194IN, G694IN, G2504IN, and G7487IN. These inactive versions had the same P1, biosensor, and blocker sequences as the active counterparts, but the P1.1 catalytic helix replaced the GC nucleotides with A’s [52], and the active site was inactivated by deletion of C75 [53,54]. Lastly, the control HDV-R ribozyme retained an active P1.1 helix and active site sequence at C75, but replaced the P1, biosensor, and blocker sequences with randomly generated sequences not present in the CHIKV genome.
- 3.1.2. Assay of Caspase 3 Activity in CHIKV-Infected Vero Cells after Lipofection with SOFA HDV-Rzs
The infection of cells by CHIKV results in an increase in caspase 3 levels as a result of virus induced apoptosis. In previous analyses we have successfully used caspase 3 levels to measure relative levels of virus infection in cell cultures [43]. Caspase 3 was chosen as the benchmark for infectivity since viral infection and replication leads to caspase dependent cell death as apoptosis induction is considered a primary defense response to viral infection in an attempt to limit viral replication [57]. In this analysis, Vero cells were first infected with CHIKV 181/25, and then transfected with in vitro transcribed SOFA HDV-Rz RNA. Following a 48 hour incubation period the culture supernatants were tested for caspase 3.
Caspase 3 levels were not significantly different among all uninfected (active and inactive) SOFA HDV-Rzs when compared to each other and the controls, with p-values greater than 0.05 (Figure 3). Caspase activities of all but one of the infected samples were significantly higher compared to their uninfected controls (p<0.05). The sole exception was the infection of the G124 treatment which exhibited a slight but non-significant increase in caspase activity in the infected samples as compared with the uninfected samples (p>0.05) demonstrating greater suppressive activity than the other SOFA HDV Rzs. (Table 2 and Figure 3).
All infected active SOFA HDV-Rzs, G124, G194, G649, G2504, and G7487 exhibited statistically significant reductions in caspase 3 levels compared with the infected controls: inactive corresponding ribozymes, Mock Transfection, SOFA HDV Ribozyme without biosensor (SOFA HDV-R), T7 random RNA (T7 RNA), and Wildtype (WT, untreated Vero cells) (Table 2).
CHIKV infections treated with the five inactive control SOFA HDV-Rzs, G124IN, G194IN, G649IN, G2504IN, G7487IN, the SOFA HDV-R, T7 RNA, and Mock Transfection controls exhibited significantly increased CHIKV-induced apoptosis (as determined by the caspase 3 assay, Figure 3) compared to the WT control (p<0.05). The five inactive control SOFA HDV-Rzs, G124IN, G194IN, G649IN, G2504IN, and G7487IN, were not significantly different in caspase activity from the SOFA HDV-R control, the T7 RNA control, or the Mock Transfection control (p-values all greater than 0.05). The SOFA HDV-R, T7 RNA, and Mock Transfection controls were not significantly different from each other (p-values > 0.05) (Table 2 and Figure 3). Combined, these data demonstrate that the introduction of RNA and/or the transfection process did induce some increase in caspase 3 activity within the cells in the infected samples, but the presence of the SOFA HDV-Rz themselves did not induce an increase in the caspase 3 activity above the level of the increase due to the transfection/infection process.
SOFA HDV-Rzs G194 and G194IN both significantly decrease CHIKV-induced apoptosis compared to the CHIKV infected Mock Transfection, SOFA HDV-R, T7 RNA, and the WT controls (p-value less than 0.001 for all). The SOFA HDV-R G194 showed significantly lower caspase 3 activity when compared to its inactive control (SOFA HDV-R G194IN) p-value < 0.05) (Table 2 and Figure 3). The reduction in caspase levels with the SOFA HDV-Rz G194IN suggests the predominant mechanism of inhibition for this ribozyme may be interference with CHIKV infection through binding at the target site.
- 3.1.3. TCID50 Analysis of Infectious Virus Production in Vero Cells Treated with Anti-CHIKV SOFA HDV-Rzs
The purpose of employing these SOFA HDV-Rzs is to inhibit infectious virus production thus lessening cellular infection. TCID50 assays are an effective means of measuring the production of infectious virus. Since many of the SOFA HDV-Rzs did not reduce the caspase levels of infected cells by at least 50%, we chose only G124 and G7487 to continue our analysis. Vero cells infected with CHIKV were transfected with each of the SOFA HDV-Rzs and allowed to incubate for 48 hours. Culture supernatants were harvested and used in a dilution titration assays to measure the relative effectiveness of the ribozymes in suppressing CHIKV infection of cells.
Both active SOFA HDV-Rzs G124 and G7487 displayed significant reductions in the viral titer from the Mock with p-values less than 0.001 for both G7487 and G124 (Table 3 and Figure 4). In addition, the active SOFA HDV-Rzs significantly reduced the viral titers relative to their inactive controls. Ribozyme G124 showed a significant reduction from inactive ribozyme G124-IN with a p-value of less than 0.001 (Table 3 and Figure 4), and ribozyme G7487 showed a significant reduction from inactive ribozyme G7487-IN with a p-value of less than 0.001 (Table 3 and Figure 4). While SOFA HDV-Rzs G194, G694, and G2504 did exhibit some ribozyme activity based upon greater inhibition compared with their inactive counterparts, the level of activity was lower. We therefore did not test these ribozymes for viral inhibition and focused instead on the G124 and G7487 constructs.
4. Discussion
Chikungunya outbreaks have occurred in Africa, the Americas, Asia, Europe, the Caribbean, and islands of the Indian and Pacific Oceans, and represent an important economic burden in endemic areas. The most common symptoms of infection are fever and arthralgia, similar to those of Dengue fever. Other symptoms may include headache, myalgia, joint swelling/inflammation, or rash. No specific antiviral treatment is currently available for chikungunya; however, a number of therapeutic options are being investigated (see 24 for review). Among these are gene-inactivation approaches, including the use of antisense oligonucleotides, RNA interference (RNAi; 31), and antiviral ribozymes [43].
Our group has investigated antiviral hammerhead ribozyme transgenes for control of Dengue and Chikungunya viruses in mosquito and mammalian cell cultures [39,43]. While these ribozymes have proven effective in suppressing these viruses in cell culture, their translation to use as human antiviral therapeutics is less optimal than what may be achieved with HDV-Rz [50,57].
HDV was discovered in 1977 following the identification of a delta antigen (HDAg), in liver biopsies and sera from patients with a severe hepatitis B. This antigen was then associated with a transmissible satellite virus that was dependent on the human hepatitis B virus for packaging, release, and transmission (reviewed by 44). Rolling circle replication of the negative-sense, genomic HDV RNA produces a complementary positive-sense, antigenomic RNA as the replication intermediate for synthesis of additional genomic RNA. Both genomic and antigenomic RNA concatemers are resolved into monomers by the HDV-Rz intrinsic ribozyme structure. The identification of HDV-like agents in a diversity of vertebrate and invertebrate species, and the structural similarity of the HDV ribozyme to cellular ribozymes suggests that HDV and its ribozyme likely evolved from the cellular transcriptome (see 44 for review).
In contrast with plant-derived ribozymes like hammerhead and hairpin, the HDV-Rz offers several unique properties as a potential human therapeutic tool including the natural adaptability to function in the presence of human proteins and physiological magnesium concentrations (1 mM Mg2+), and adopting a single conformation that is resistant to human nucleases leading to a long half-life. In addition, the use of liposome reagents is an efficient means of inducing cellular uptake of HDV-Rz [58] Moreover, HDV-Rzs have demonstrated effectiveness in suppression of Hepatitis B in vitro [59], HIV in vitro [60], and Influenza infections in mice [52].
In this study we employed a SOFA HD-Rz constructs based upon the original successful construct of Bergeron and Perreault [49]. The SOFA (Specific On/Off Adapter) module of this ribozyme acts as a switch to control the activity of the ribozyme by inhibiting the cleavage activity of the ribozyme with a blocker sequence that binds the P1 domain from forming the catalytic configuration. The blocker is adjacent to the biosensor sequence that recognizes the target sequence on the substrate RNA. Once the biosensor binds the target, the blocker sequence is released from the P1 domain, permitting formation of the active conformation and cleavage of the target [56].
We employed 5 CHIKV-specific biosensor sequences targeting different G residues within the CHIKV genome for cleavage (Table 1). Of these five, two exhibited the best suppressive activity upon lipofection of CHIKV infected Vero cells (Figure 3 and Figure 4). Our controls (T7 RNA, SOFA HDV-R, and Mock Transfection) were not significantly different from each other, indicating the SOFA HDV-R lacking CHIKV targeting homologous sequences had no impact on viral induced apoptosis. These data confirm that the CHIKV targeting SOFA HDV-Rzs needs to interact with the genomic viral RNA in order to affect apoptosis and are not doing so in a non-specific manner.
While mutations in the P1 domain are not tolerated, mismatches in the biosensor region may be tolerated, depending on location relative to the P1 domain [61]. Thus, escape mutations may be somewhat less probable than would occur with RNAi or other ribozymes [33]. Certainly, the extent of homology maintenance for targeting and cleavage is considerably smaller than siRNA.
Depending upon which site is targeted, the SOFA HDV-Rz may inhibit CHIKV through simple biosensor RNA binding, or through the catalytic activity of the HDV-Rz. The fact that any reductions observed were statistically significant relative to the catalytically impaired SOFA HDV-Rz controls indicates that these reductions are at least in part due to the catalytic activity of these SOFA HDV-Rzs. Targeting of G124 resulted in significant infectivity reductions as assessed by both the caspase and TCID50 assays, effectively reducing the viral titer by 3 logs TCID50. Similarly, targeting G7487 reduced the viral titer by 1.5 logs in TCID50 assays. Such reductions could be sufficient to reduce clinical manifestation of disease.
In other systems, variability in effectiveness of HDV-Rzs is associated with several factors including substrate conformation, length and sequence of the biosensor, and cellular location of target viral genome during replication and translation [50]. These factors often cannot be reliably predicted [61]. However, there may be alternative sites in the genome that, while not necessarily consensus among all CHIKV strains, may prove more effective targets based upon these variables.
The use of many RNA species as therapeutic molecules requires engineering long term stability of these molecules. Some modifications can adversely influence their activity [62]. While the half-life of HDV-Rzs is superior to other RNA molecules, owing in part to its formation of a highly stable secondary structure [57], this stability may be improved by chemical modifications as has been done for siRNA. However, these alterations can decrease their effectiveness, and in the case of SOFA HDV-Rzs, may not offer much advantage [63,64].
While the infectiousness and productivity of the 181/25 strain in Vero cell cultures did not appear to be significantly different than virulent strains in these cells based on those obtained in prior reports (Sudeep et al., 2019), there may still be some inapparent difference in the replication efficiency of the vaccine strain that influences these results. As a consequence, we can only say that the HDV-Rz approach shows some promise that needs to be replicated with virulent strains.
5. Conclusion
We have demonstrated that SOFA HDV-Rzs can be useful as antiviral agents against CHIKV in a cell culture infection situation. We demonstrated that post infection application of these ribozymes will effectively inhibit virus production, reducing the active infection significantly. These data provide evidence that such ribozymes may be useful as therapeutic tools in the treatment of CHIKV. In fact, such ribozymes have particular advantage in the treatment of human pathogens due to their relative stability and activity compared with hairpin and hammerhead ribozymes derived from plant sources. Even if the use of these SOFA HDV-Rzs such treatments were incapable of eliminating virus infection altogether, their application could contribute to a lessening of symptoms or duration for the disease. The simplicity of SOFA HDV-Rz construction allows the rapid examination of other potential target sites. In addition, the utilization of combinations of these ribozymes may prove even more effective. Based upon our relatively positive results we believe further investigation of the therapeutic potential of HDV-Rzs for viruses like CHIKV, Dengue, and Zika does appear to be warranted.
Author Contributions
Cheryl Kucharski was responsible for the design and overseeing of the cloning, expression, transfection, and challenges of the HDV ribozymes, Erin Hanahoe participated in the cloning, expression, transfections, challenges, and TCID50 analyses. Zoe Loh was responsible for the expression, transfection, challenges, and caspase 3 assays. Mark E. Fraser made crucial analyses and interpretation of data, and was responsible for writing and final preparation of the manuscript. Malcolm J. Fraser, Jr. was the Principal Investigator and primarily responsible for all aspects of the funding, research design, interpretation, and writing of this manuscript.
Funding
This research was supported by NIH/NIAID grant 1R01AI097554 to MJF.
Acknowledgments
The authors would like to thank Dr. Scott Weaver, (university of Texas Medical Branch, Galveston, TX, USA) for providing the attenuated strain of CHIKV 181/25. We also acknowledge the invaluable instruction and assistance of Tresa Fraser in maintenance of cell cultures, and the contributions to genomic analysis of target sequences by Dr. Pruksa Nawtaisong. This research was supported by NIH/NIAID grant 1R01AI097554 to MJF.
References
- Cordel, H.; Quatresous, I.; Paquet, C.; Couturier, E. Imported cases of chikungunya in metropolitan France, April 2005–February 2006. Euro Surveill. 2006, 11, E0604203. [Google Scholar] [CrossRef]
- Bianco, C. Dengue and Chikungunya viruses in blood donations: risks to the blood supply? Transfusion. 2008, 48, 1279–81. [Google Scholar] [CrossRef]
- Appassakij, H.; Khuntikij, P.; Kemapunmanus, M.; Wutthanarungsan, R.; Silpapojakul, K. Viremic profiles in asymptomatic and symptomatic chikungunya fever: a blood transfusion threat? Transfusion 2013, 53 (Pt 2), 2567–74. [Google Scholar] [CrossRef]
- Ramful, D.; Carbonnier, M.; Pasquet, M.; Bouhmani, B.; Ghazouani, J.; Noormahomed, T.; Beullier, G.; Attali, T.; Samperiz, S.; Fourmaintraux, A.; Alessandri, J.L. Mother-to-child transmission of Chikungunya virus infection. Pediatr Infect Dis J. 2007, 26, 811–5. [Google Scholar] [CrossRef]
- Gérardin, P.; Barau, G.; Michault, A.; Bintner, M.; Randrianaivo, H.; Choker, G.; Lenglet, Y.; Touret, Y.; Bouveret, A.; Grivard, P.; Le Roux, K.; Blanc, S.; Schuffenecker, I.; Couderc, T.; Arenzana-Seisdedos, F.; Lecuit, M.; Robillard, P.Y. Multidisciplinary prospective study of mother-to-child chikungunya virus infections on the island of La Réunion. PLoS Med. 2008, 5, e60. [Google Scholar] [CrossRef]
- Pialoux, G.; Gaüzère, B.A.; Jauréguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect Dis. 2007, 7, 319–27. [Google Scholar] [CrossRef]
- Farnon, E.C.; Sejvar, J.J.; Staples, J.E. Severe disease manifestations associated with acute chikungunya virus infection. Crit Care Med. 2008, 36, 2682–3. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; Biscornet, L.; Diancourt, L.; Michel, S.; Duquerroy, S.; Guigon, G.; Frenkiel, M.P.; Bréhin, A.C.; Cubito, N.; Desprès, P.; Kunst, F.; Rey, F.A.; Zeller, H.; Brisse, S. Genome microevolution of chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.K.; Singh, M.; Mishra, N.; Lakshmi, V. Resurgence of chikungunya virus in India: an emerging threat. Euro Surveill. 2006, 11, E0608102. [Google Scholar] [CrossRef] [PubMed]
- Mavalankar, D.; Shastri, P.; Raman, P. Chikungunya epidemic in India: a major public-health disaster. Lancet Infect Dis. 2007, 7, 306–7. [Google Scholar] [CrossRef] [PubMed]
- Requena-Méndez, A.; Garcia, C.; Aldasoro, E.; Vicente, J.A.; Martínez, M.J.; Pérez-Molina, J.A.; Calvo-Cano, A.; Franco, L.; Parrón, I.; Molina, A.; Ruiz, M.; Álvarez, J.; Sánchez-Seco, M.P.; Gascón, J. Cases of chikungunya virus infection in travellers returning to Spain from Haiti or Dominican Republic, April-June 2014. Euro Surveill. 2014, 19, 20853. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya virus in US travelers returning from India, 2006. Emerg Infect Dis. 2007, 13, 764–7. [Google Scholar] [CrossRef] [PubMed]
- Hochedez, P.; Hausfater, P.; Jaureguiberry, S.; Gay, F.; Datry, A.; Danis, M.; Bricaire, F.; Bossi, P. Cases of chikungunya fever imported from the islands of the South West Indian Ocean to Paris, France. Euro Surveill. 2007, 12, 679. [Google Scholar] [CrossRef]
- Morrison, T.E. Reemergence of chikungunya virus. J Virol. 2014, 88, 11644–7. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef]
- Wang, E.; Volkova, E.; Adams, A.P.; Forrester, N.; Xiao, S.Y.; Frolov, I.; Weaver, S.C. Chimeric alphavirus vaccine candidates for chikungunya. Vaccine. 2008, 26, 5030–9. [Google Scholar] [CrossRef]
- Kumar, M.; Sudeep, A.B.; Arankalle, V.A. Evaluation of recombinant E2 protein-based and whole-virus inactivated candidate vaccines against chikungunya virus. Vaccine. 2012, 30, 6142–9. [Google Scholar] [CrossRef]
- Wang, D.; Suhrbier, A.; Penn-Nicholson, A.; Woraratanadharm, J.; Gardner, J.; Luo, M.; Le, T.T.; Anraku, I.; Sakalian, M.; Einfeld, D.; Dong, J.Y. A complex adenovirus vaccine against chikungunya virus provides complete protection against viraemia and arthritis. Vaccine. 2803. [Google Scholar] [CrossRef]
- Slifka, D.K.; Raué, H.P.; Weber, W.C.; Andoh, T.F.; Kreklywich, C.N.; DeFilippis, V.R.; Streblow, D.N.; Slifka, M.K.; Amanna, I.J. Development of a next-generation chikungunya virus vaccine based on the HydroVax platform. PLoS Pathog. 2022, 18, e1010695. [Google Scholar] [CrossRef]
- Metz, S.W.; Gardner, J.; Geertsema, C.; Le, T.T.; Goh, L.; Vlak, J.M.; Suhrbier, A.; Pijlman, G.P. Effective chikungunya virus-like particle vaccine produced in insect cells. PLoS Negl Trop Dis. 2013, 7, e2124. [Google Scholar] [CrossRef]
- Livengood, J.A.; Partidos, C.D.; Plante, K.; Seymour, R.; Gorchakov, R.; Varga, L.; Paykel, J.; Weger, J.; Haller, A.; Stinchcomb, D.T.; Osorio, J.; Weaver, S. Preclinical Evaluation of a Live Attenuated Chikungunya Vaccine, Procedia in Vaccinology. 2012, 6, 141–149. [Google Scholar] [CrossRef]
- de Lima Cavalcanti, T.Y.V.; Pereira, M.R.; de Paula, S.O.; Franca, R.F.O. A Review on Chikungunya Virus Epidemiology, Pathogenesis and Current Vaccine Development. Viruses. 2022, 14, 969. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Narciso-Abraham, M.; Hadl, S.; McMahon, R.; Toepfer, S.; Fuchs, U.; Hochreiter, R.; Bitzer, A.; Kosulin, K.; Larcher-Senn, J.; Mader, R.; Dubischar, K.; Zoihsl, O.; Jaramillo, J.C.; Eder-Lingelbach, S.; Buerger, V.; Wressnigg, N. Safety and immunogenicity of a single-shot live-attenuated chikungunya vaccine: a double-blind, multicentre, randomised, placebo-controlled, phase 3 trial. Lancet. 2023, 401, 2138–2147. [Google Scholar] [CrossRef] [PubMed]
- Battisti, V.; Urban, E.; Langer, T. Antivirals against the Chikungunya Virus. Viruses. 2021, 13, 1307. [Google Scholar] [CrossRef]
- Suzuki, Y. Interferon-induced restriction of Chikungunya virus infection. Antiviral Res. 2023, 210, 105487. [Google Scholar] [CrossRef]
- López, L.S.; Calvo, E.P.; Castellanos, J.E. Deubiquitinating Enzyme Inhibitors Block Chikungunya Virus Replication. Viruses. 2023, 15, 481. [Google Scholar] [CrossRef]
- Oyewole, O.O.; Dunnavant, K.; Bhattarai, S.; Kharel, Y.; Lynch, K.R.; Santos, W.L.; Reid, S.P. A Novel Sphingosine Kinase Inhibitor Suppresses Chikungunya Virus Infection. Viruses. 2022, 14, 1123. [Google Scholar] [CrossRef]
- Nam, S.; Ga, Y.J.; Lee, J.Y.; Hwang, W.Y.; Jung, E.; Shin, J.S.; Chen, W.; Choi, G.; Zhou, B.; Yeh, J.Y.; Go, Y.Y. Radicicol Inhibits Chikungunya Virus Replication by Targeting Nonstructural Protein 2. Antimicrob Agents Chemother. 2021, 65, e001352. [Google Scholar] [CrossRef]
- Levi, L.I.; Rezelj, V.V.; Henrion-Lacritick, A.; Erazo, D.; Boussier, J.; Vallet, T.; Bernhauerová, V.; Suzuki, Y.; Carrau, L.; Weger-Lucarelli, J.; Saleh, M.C.; Vignuzzi, M. Defective viral genomes from chikungunya virus are broad-spectrum antivirals and prevent virus dissemination in mosquitoes. PLoS Pathog. 2021, 17, e1009110. [Google Scholar] [CrossRef]
- Solignat, M.; Gay, B.; Higgs, S.; Briant, L.; Devaux, C. Replication cycle of chikungunya: a re-emerging arbovirus. Virology. 2009, 393, 183–97. [Google Scholar] [CrossRef]
- Dash, P.K.; Tiwari, M.; Santhosh, S.R.; Parida, M.; Lakshmana Rao, P.V. RNA interference mediated inhibition of Chikungunya virus replication in mammalian cells. Biochem Biophys Res Commun. 2008, 376, 718–22. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.; Chen, K.C.; Ng, M.M.; Chu, J.J. Expression of plasmid-based shRNA against the E1 and nsP1 genes effectively silenced Chikungunya virus replication. PLoS One. 2012, 7, e46396. [Google Scholar] [CrossRef]
- Bull, J.J.; Jacobson, A.; Badgett, M.R.; Molineux, I.J. Viral escape from antisense RNA. Mol Microbiol. 1998, 28, 835–46. [Google Scholar] [CrossRef]
- Cheng, G.F.; Lin, J.J.; Shi, Y.; Jin, Y.X.; Fu, Z.Q.; Jin, Y.M.; Zhou, Y.C.; Cai, Y.M. Dose-dependent inhibition of gynecophoral canal protein gene expression in vitro in the schistosome (Schistosoma japonicum) by RNA interference. Acta Biochim Biophys Sin (Shanghai). 2005, 37, 386–90. [Google Scholar] [CrossRef]
- Sanchez-Vargas, I.; Travanty, E.A.; Keene, K.M.; Franz, A.W.; Beaty, B.J.; Blair, C.D.; Olson, K.E. RNA interference, arthropod-borne viruses, and mosquitoes. Virus Res. 2004, 102, 65–74. [Google Scholar] [CrossRef]
- von Laer, D.; Hasselmann, S.; Hasselmann, K. Gene therapy for HIV infection: what does it need to make it work? J Gene Med. 2006. [Google Scholar] [CrossRef]
- Rossi, J.J.; June, C.H.; Kohn, D.B. Genetic therapies against HIV. Nat Biotechnol. 2007, 1444–54. [Google Scholar] [CrossRef]
- 38. Mitsuyasu, R.T.; Merigan, T.C.; Carr, A.; Zack, J.A.; Winters, M.A.; Workman, C.; Bloch, M.; Lalezari, J.; Becker, S.; Thornton, L.; Akil, B.; Khanlou, H.; Finlayson, R.; McFarlane, R.; Smith, D.E.; Garsia, R.; Ma, D.; Law, M.; Murray, J.M.; von Kalle, C.; Ely, J.A.; Patino, S.M.; Knop, A.E.; Wong, P.; Todd, A.V.; Haughton, M.; Fuery, C.; Macpherson, J.L.; Symonds, G.P.; Evans, L.A.; Pond, S.M.; Cooper, D.A. Phase 2 gene therapy trial of an anti-HIV ribozyme in autologous CD34+ cells. Nat Med. 2009, 15, 285–92. [Google Scholar] [CrossRef] [PubMed]
- Nawtaisong, P.; Keith, J.; Fraser, T.; Balaraman, V.; Kolokoltsov, A.; Davey, R.A.; Higgs, S.; Mohammed, A.; Rongsriyam, Y.; Komalamisra, N.; Fraser, M.J, Jr. Effective suppression of Dengue fever virus in mosquito cell cultures using retroviral transduction of hammerhead ribozymes targeting the viral genome. Virol J. 2009, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.R.; Keith, J.H.; Barde, P.V.; Fraser, T.S.; Fraser, M.J., Jr. Targeting of highly conserved Dengue virus sequences with anti-Dengue virus trans-splicing group I introns. BMC Mol Biol. 2010, 11, 84. [Google Scholar] [CrossRef]
- Carter, J.R.; Keith, J.H.; Fraser, T.S.; Dawson, J.L.; Kucharski, C.A.; Horne, K.M.; Higgs, S.; Fraser, M.J., Jr. Effective suppression of dengue virus using a novel group-I intron that induces apoptotic cell death upon infection through conditional expression of the Bax C-terminal domain. Virol J. 2014, 11, 111. [Google Scholar] [CrossRef]
- Carter, J.R.; Taylor, S.; Fraser, T.S.; Kucharski, C.A.; Dawson, J.L.; Fraser, M.J., Jr. Suppression of the Arboviruses Dengue and Chikungunya Using a Dual-Acting Group-I Intron Coupled with Conditional Expression of the Bax C-Terminal Domain. PLoS One. 2015, 10, e0139899. [Google Scholar] [CrossRef]
- Mishra, P.; Furey, C.; Balaraman, V.; Fraser, M.J. Antiviral Hammerhead Ribozymes Are Effective for Developing Transgenic Suppression of Chikungunya Virus in Aedes aegypti Mosquitoes. Viruses. 2016, 8, 163. [Google Scholar] [CrossRef]
- Netter, H.J.; Barrios, M.H.; Littlejohn, M.; Yuen, L.K.W. Hepatitis Delta Virus (HDV) and Delta-Like Agents: Insights Into Their Origin. Front Microbiol. 2021, 12, 652962. [Google Scholar] [CrossRef]
- Bergeron, L.J.; Ouellet, J.; Perreault, J.P. Ribozyme-based gene-inactivation systems require a fine comprehension of their substrate specificities; the case of delta ribozyme. Curr Med Chem. 2003, 10, 2589–97. [Google Scholar] [CrossRef]
- Kato, Y.; Kuwabara, T.; Warashina, M.; Toda, H.; Taira, K. Relationships between the activities in vitro and in vivo of various kinds of ribozyme and their intracellular localization in mammalian cells. J Biol Chem. 2001, 276, 15378-85. [Google Scholar] [CrossRef]
- D'Anjou, F.; Bergeron, L.J.; Larbi, N.B.; Fournier, I.; Salzet, M.; Perreault, J.P.; Day, R. Silencing of SPC2 expression using an engineered delta ribozyme in the mouse betaTC-3 endocrine cell line. J Biol Chem. 2004, 279, 14232-9. [Google Scholar] [CrossRef]
- Sheng, J.; Al-Anouti, F.; Ananvoranich, S. Engineered delta ribozymes can simultaneously knock down the expression of the genes encoding uracil phosphoribosyltransferase and hypoxanthine-xanthine-guanine phosphoribosyltransferase in Toxoplasma gondii. Int J Parasitol. 2004, 34, 253-63. [Google Scholar] [CrossRef]
- Bergeron, L.J.; Perreault, J.P. Target-dependent on/off switch increases ribozyme fidelity. Nucleic Acids Res. 2005, 33, 1240–8. [Google Scholar] [CrossRef]
- Lévesque, M.V.; Lévesque, D.; Brière, F.P.; Perreault, J.P. Investigating a new generation of ribozymes in order to target HCV. PLoS One. 2010, 5, e9627. [Google Scholar] [CrossRef]
- Deschênes, P.; Lafontaine, D.A.; Charland, S.; Perreault, J.P. Nucleotides -1 to -4 of hepatitis delta ribozyme substrate increase the specificity of ribozyme cleavage. Antisense Nucleic Acid Drug Dev. 2000, 10, 53–61. [Google Scholar] [CrossRef]
- Motard, J.; Rouxel, R.; Paun, A.; von Messling, V.; Bisaillon, M.; Perreault, J.P. A novel ribozyme-based prophylaxis inhibits influenza A virus replication and protects from severe disease. PLoS One. 2011, 6, e27327. [Google Scholar] [CrossRef]
- Perrotta, A.T.; Shih, I.; Been, M.D. Imidazole rescue of a cytosine mutation in a self-cleaving ribozyme. Science. 1999, 286, 123–6. [Google Scholar] [CrossRef]
- Wrzesinski, J. , Wichłacz, A., Nijakowska, D., Rebowska, B., Nawrot, B. , Ciesiołka, J. Phosphate residues of antigenomic HDV ribozyme important for catalysis that are revealed by phosphorothioate modification† New J. Chem., 2010, 34, 1018–1026. [Google Scholar]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J Virol. 2016, 5, 85–6. [Google Scholar] [CrossRef]
- Bergeron, L.J.; Reymond, C.; Perreault, J.P. Functional characterization of the SOFA delta ribozyme. RNA. 2005, 11, 1858–68. [Google Scholar] [CrossRef]
- Joubert, P.E.; Werneke, S.W.; de la Calle, C.; Guivel-Benhassine, F.; Giodini, A.; Peduto, L.; Levine, B.; Schwartz, O.; Lenschow, D.J.; Albert, M.L. Chikungunya virus-induced autophagy delays caspase-dependent cell death. J Exp Med. 2012, 209, 1029–47. [Google Scholar] [CrossRef]
- Lévesque, D.; Choufani, S.; Perreault, J.P. Delta ribozyme benefits from a good stability in vitro that becomes outstanding in vivo. RNA. 2002, 8, 464–77. [Google Scholar] [CrossRef]
- Wang, C.X.; Lu, Y.Q.; Qi, P.; Chen, L.H.; Han, J.X. Efficient inhibition of hepatitis B virus replication by hepatitis delta virus ribozymes delivered by targeting retrovirus. Virol J. 2010, 7, 61. [Google Scholar] [CrossRef]
- Lainé, S.; Scarborough, R.J.; Lévesque, D.; Didierlaurent, L.; Soye, K.J.; Mougel, M.; Perreault, J.P.; Gatignol, A. In vitro and in vivo cleavage of HIV-1 RNA by new SOFA HDV ribozymes and their potential to inhibit viral replication. RNA Biol. 2011, 8, 343–53. [Google Scholar] [CrossRef]
- Lévesque, M.V.; Rouleau, S.G.; Perreault, J.P. Selection of the most potent specific on/off adaptor-hepatitis delta virus ribozymes for use in gene targeting. Nucleic Acid Ther. 2011, 21, 241–52. [Google Scholar] [CrossRef]
- Traber, G.M.; Yu, A.M. RNAi-Based Therapeutics and Novel RNA Bioengineering Technologies. J Pharmacol Exp Ther. 2023, 384, 133–154. [Google Scholar] [CrossRef]
- Yu, A.M.; Jian, C.; Yu, A.H.; Tu, M.J. RNA therapy: Are we using the right molecules? Pharmacol Ther. 2019, 196, 91–104. [PubMed]
- Sudeep, A.B.; Vyas, P.B.; Parashar, D.; Shil, P. Differential susceptibility & replication potential of Vero E6, BHK-21, RD, A-549, C6/36 cells & Aedes aegypti mosquitoes to three strains of chikungunya virus. Indian J Med Res. 2019, 149, 771–777. [Google Scholar]
Figure 1.
CHIKV genome sequences used to construct the corresponding antisense regions in the HDV-Rz (Table 1). The target cleavage site, G, is indicated in red.
Figure 1.
CHIKV genome sequences used to construct the corresponding antisense regions in the HDV-Rz (Table 1). The target cleavage site, G, is indicated in red.

Figure 2.
Schematic of antisense SOFA HDV-Rz. The relative position of the P1 targeting sequence is indicated in blue, its complementary Blocker sequence in red, and the CHIKV complementary Biosensor in green.
Figure 2.
Schematic of antisense SOFA HDV-Rz. The relative position of the P1 targeting sequence is indicated in blue, its complementary Blocker sequence in red, and the CHIKV complementary Biosensor in green.

Figure 3.
Caspase Activity of SOFA-HDV-Rz Transfected Vero Cells Challenged with CHIKV. Controls included untransfected VERO cells (WT), T7 Megascript RNA challenged VERO cells (T7 RNA), a mock reaction of transfection conditions without nucleic acid (Mock Transfection), and an active SOFA-HDV-Rz without biosensor (SOFA-HDV-R). We also paired active with inactive SOFA-HDV-Rzs. No significant difference was found between all uninfected samples and controls (p>0.05). Uninfected controls exhibited statistically significant lower levels of caspase 3 activity from the infected counterparts (p<0.05) with the exception of G124 (p>0.05), and no significant difference was observed between all uninfected samples and controls (p>0.05). G124 and G7487 resulted in greater than 50% reduction of caspase activity compared with all infected controls, indicating effective suppression of CHIKV. There were statistically significant reductions in caspase activity for all infected SOFA-HDV-Rzs compared to their infected controls (inactive SOFA-HDV-R counterparts, Mock Transfection, T7 RNA, and WT; * = p<0.001). Mean of data are plotted with error bars representing standard deviation, with all data points individually plotted for each replicate. Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
Figure 3.
Caspase Activity of SOFA-HDV-Rz Transfected Vero Cells Challenged with CHIKV. Controls included untransfected VERO cells (WT), T7 Megascript RNA challenged VERO cells (T7 RNA), a mock reaction of transfection conditions without nucleic acid (Mock Transfection), and an active SOFA-HDV-Rz without biosensor (SOFA-HDV-R). We also paired active with inactive SOFA-HDV-Rzs. No significant difference was found between all uninfected samples and controls (p>0.05). Uninfected controls exhibited statistically significant lower levels of caspase 3 activity from the infected counterparts (p<0.05) with the exception of G124 (p>0.05), and no significant difference was observed between all uninfected samples and controls (p>0.05). G124 and G7487 resulted in greater than 50% reduction of caspase activity compared with all infected controls, indicating effective suppression of CHIKV. There were statistically significant reductions in caspase activity for all infected SOFA-HDV-Rzs compared to their infected controls (inactive SOFA-HDV-R counterparts, Mock Transfection, T7 RNA, and WT; * = p<0.001). Mean of data are plotted with error bars representing standard deviation, with all data points individually plotted for each replicate. Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
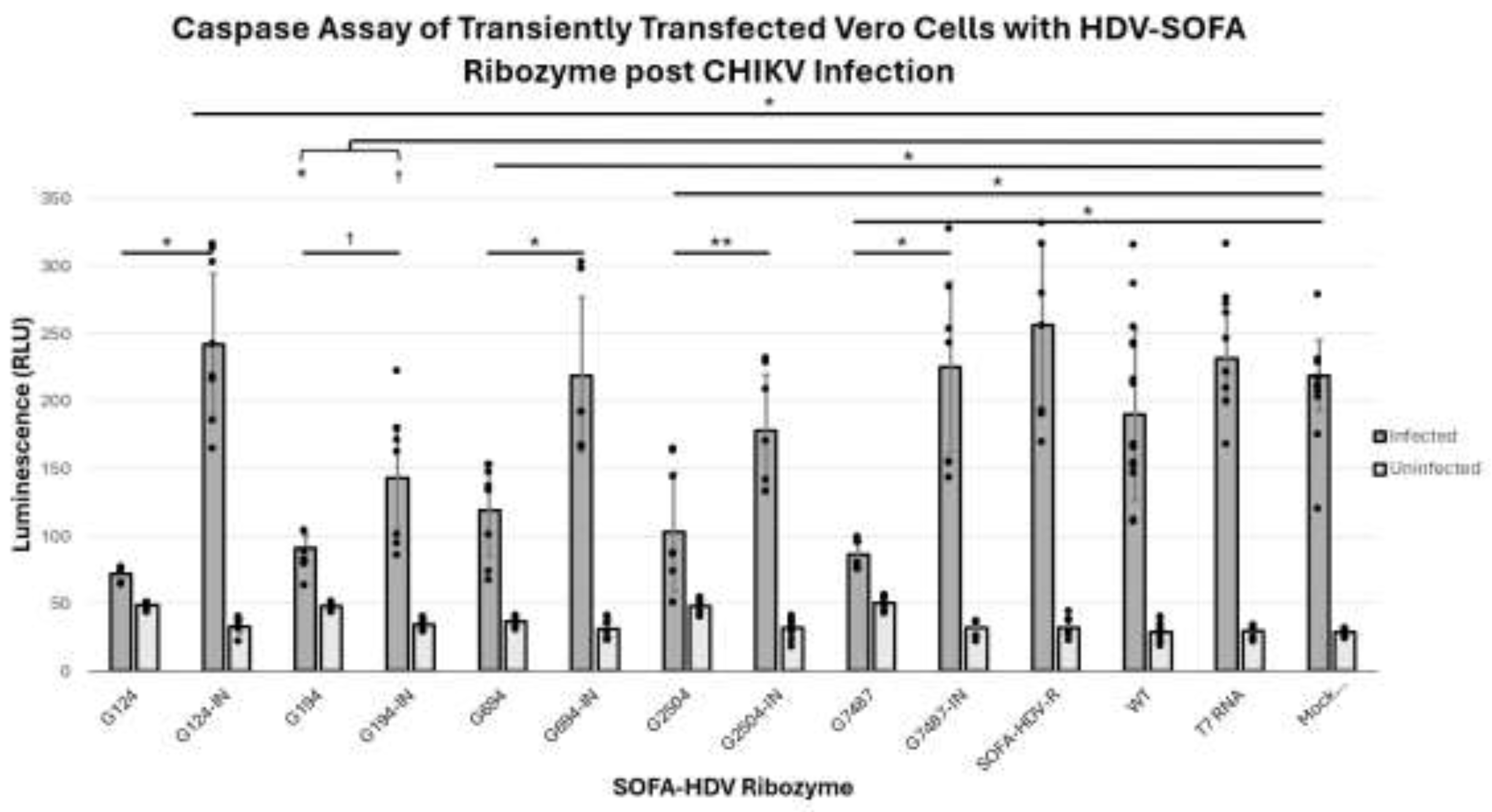
Figure 4.
Viral Titer as Determined by TCID50 Assay of Transiently Transfected Vero Cells. G124-IN and G7487-IN showed no significant reduction in viral titers from infected control (p>0.05). G7487 showed significantly reduced viral titer when compared to both G7487-IN and Mock Transfection controls by ~1.5 log10 (** = p<0.001). G124 showed significantly reduced viral titers from infected control by >3 log10 (** = p<0.001) and from G124-IN by > 2 log10 (** = p<0.001). G124 showed significantly reduced viral titer from G7487 by ~1 log10 (* = p<0.05). Mean data are plotted with error bars representing standard deviation, with all data points individually plotted for each replicate. Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
Figure 4.
Viral Titer as Determined by TCID50 Assay of Transiently Transfected Vero Cells. G124-IN and G7487-IN showed no significant reduction in viral titers from infected control (p>0.05). G7487 showed significantly reduced viral titer when compared to both G7487-IN and Mock Transfection controls by ~1.5 log10 (** = p<0.001). G124 showed significantly reduced viral titers from infected control by >3 log10 (** = p<0.001) and from G124-IN by > 2 log10 (** = p<0.001). G124 showed significantly reduced viral titer from G7487 by ~1 log10 (* = p<0.05). Mean data are plotted with error bars representing standard deviation, with all data points individually plotted for each replicate. Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
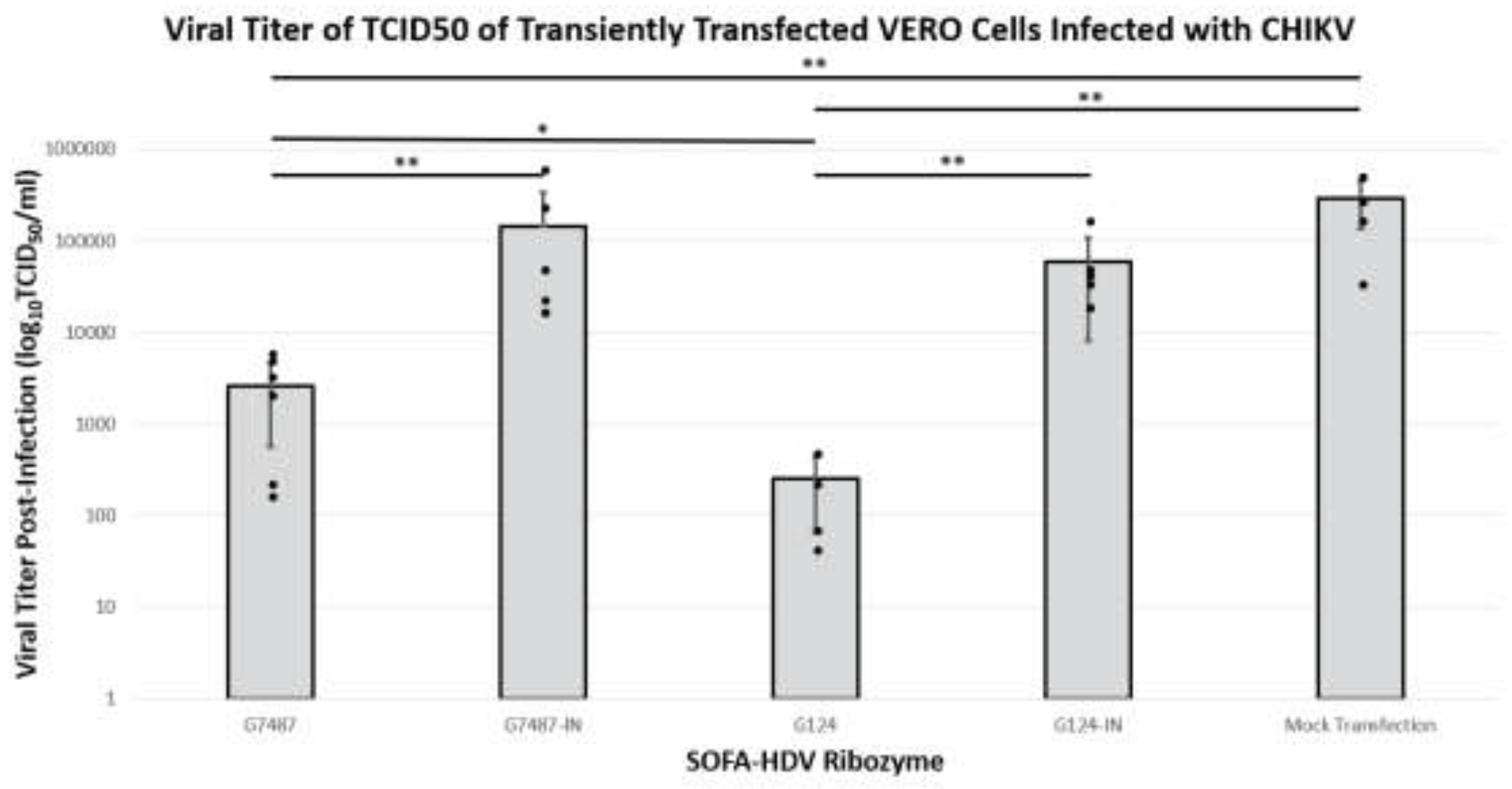

Table 1.
Sequences correspond to locations in the schematic of the SOFA HDV-Rz, Figure 1. All sequences are 5’ to 3’, with the P1 and Biosensor sequences complementary to the CHIKV genome (Figure 1), and the Blocker sequence complementary to the P1 sequence.
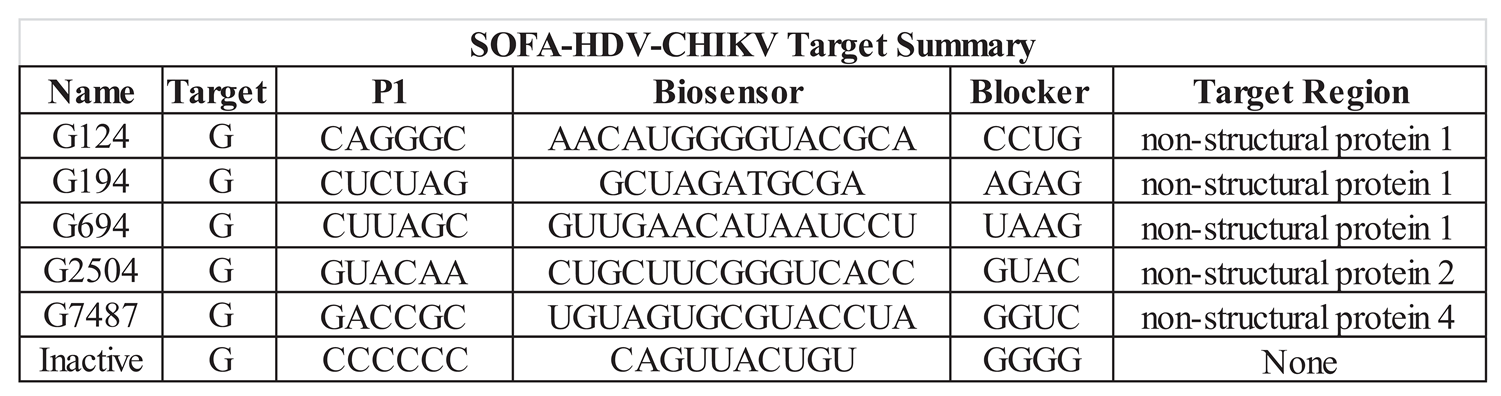 |
Table 2.
Caspase activity of CHIKV infected Vero Cells treated post infection with SOFA- HDV-Rzs. Data presented were used to prepare the graph of Figure 3. Means and Standard Deviations are in caspase-3 Relative Luciferase Units (RLU). Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
Table 2.
Caspase activity of CHIKV infected Vero Cells treated post infection with SOFA- HDV-Rzs. Data presented were used to prepare the graph of Figure 3. Means and Standard Deviations are in caspase-3 Relative Luciferase Units (RLU). Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
| Construct |
Mean (Caspase 3 RLU) |
Standard Deviation (Caspase 3 RLU) | p-value compared to Mock | p-value compared to Inactive |
| G124 | 71.67 | 5.28 | <0.001 | <0.001 |
| G124-IN | 242.10 | 52.68 | 0.202 | |
| G194 | 91.06 | 9.63 | <0.001 | 0.012 |
| G194-IN | 132.36 | 46.31 | <0.001 | |
| G694 | 119.12 | 34.15 | <0.001 | <0.001 |
| G694-IN | 219.04 | 58.13 | 1.00 | |
| G2504 | 102.92 | 44.25 | <0.001 | 0.005 |
| G2504-IN | 178.08 | 41.12 | 0.796 | |
| G7487 | 86.56 | 9.52 | <0.001 | <0.001 |
| G7487-IN | 225.08 | 63.12 | 0.849 | |
| SOFA-HDV-R | 256.27 | 60.44 | 0.136 | |
| WT | 189.67 | 63.38 | 0.024 | |
| T7 RNA | 234.73 | 34.30 | 0.419 | |
| Mock Transfection | 221.00 | 26.18 |
Table 3.
Viral Titers of CHIKV as determined TCID50 Assay of Transiently Transfected Vero Cells. Data used to prepare the graph of Figure 4. Mean and Standard Deviations are in TCID50/mL. Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
Table 3.
Viral Titers of CHIKV as determined TCID50 Assay of Transiently Transfected Vero Cells. Data used to prepare the graph of Figure 4. Mean and Standard Deviations are in TCID50/mL. Statistics were performed in IBM SPSS via ANOVA with Tukey’s post-hoc test. Power for this experiment with alpha 0.05 was determined to be 1.000 utilizing IBM SPSS via Univariate Analysis of Variance.
| Construct | Mean (TCID50/mL) | Standard Deviation (TCID50/mL) | p-value compared to Mock | p-value compared to Inactive |
| G7487 | 2632.55 | 2067.71 | <0.001 | <0.001 |
| G7487-IN | 146238.05 | 198934.72 | 0.24 | |
| G124 | 250.80 | 183.96 | <0.001 | <0.001 |
| G124-IN | 58824.37 | 50740.36 | 0.15 | |
| Mock Transfection | 292477.82 | 155664.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



