
Submitted:
06 March 2024
Posted:
07 March 2024
You are already at the latest version
Abstract
Heteroresistance (HR) to colistin is especially concerning in settings where multi-drug resistant (MDR) K. pneumoniae are prevalent and empiric use of colistin might lead to treatment failures. This study aimed to assess the frequency of occurrence of colistin HR (CHR) among (MDR) K. pneumoniae (n=676) isolated from patients hospitalized in 13 intensive care units (ICUs) in 6 European countries in a clinical trial assessing the impact of decolonization strategies. All isolates were whole-genome-sequenced and studied for in vitro colistin susceptibility. Majority were colistin-susceptible (CS) (n=597, MIC 2 µg/ml). 288 CS isolates were randomly selected for population analysis profiling (PAP) to assess CHR prevalence. CHR was detected in 108/288 CS K. pneumoniae. No significant association was found between the occurrence of CHR and country, MIC-value, K-antigen type and O-antigen type. Overall, 92% (618/671) of the K. pneumoniae were MDR with high prevalence among CS (540/592) and CR (98.7%, 78/79) isolates. In contrast, proportion of carbapenemase-producing K. pneumoniae (CP-Kpn) was higher among CR (72.2%, 57/79) than CS isolates (29.1%, 173/594). Proportions of MDR and CP-Kpn were similar among CHR (MDR: 85%, 91/107; CP-Kpn: 29.9%, 32/107) and CS isolates (MDR: 91%, 539/592; CP-Kpn: 29.1%, 153/59). WGS analysis of PAP isolates showed diverse insertion elements in mgrB or even among technical replicates underscoring the stochasticity of the CHR phenotype. CHR isolates showed high ST diversity (Simpson’s diversity index, SDI: 0.97, in 52 of the 85 STs tested). CR (SDI: 0.85) isolates were highly associated with specific STs (ST101, ST147, ST258/ST512, P ≤ 0.003). The widespread nature of CHR among MDR K. pneumoniae in our study urge the development of rapid HR detection methods to inform on the need for combination regimens.
Keywords:
colistin resistance
; mechanisms
; mgrB
; population analysis profiling
; whole genome sequencing
1. Introduction
Klebsiella pneumoniae frequently cause community-acquired and nosocomial infections such as pneumonia, urinary tract infection, liver abscesses and bloodstream infections [1]. In recent years, antimicrobial resistance in K. pneumoniae has become problematic [2,3]. In particular, resistance to carbapenems is frequently associated with resistance to multiple classes of other antibiotics which leads to limited possibilities for clinical management as alternative treatment options are limited, and lead to higher rates of treatment failures [4]. In such cases last-resort antibiotics such as colistin [5,6] may be used in association with other antibiotics. However, the high worldwide prevalence of carbapenem-resistant, multi-drug resistant (MDR) K. pneumoniae has fueled and increased the use of colistin over the last years, accelerating the emergence of isolates resistant to this compound [3]. Besides full colistin resistance (CR), colistin heteroresistance (CHR) has also been increasingly reported over the last years [7,8]. Heteroresistance (HR) is defined as a phenotype in which a bacterial isolate contains subpopulations of cells that show a substantial reduction in antibiotic susceptibility compared with the main population (minimum inhibitory concentration (MIC) increase of at least 8-fold), allowing these cells to grow in the presence of the antibiotic [9]. Detection of these subpopulations can be challenging and raise concern as their frequency may rise during antibiotic exposure and possibly lead to treatment failure [9,10,11]. However, the relevance of CHR in causing reduced clinical effectiveness and negatively affect the treatment outcome remains controversial [7]. Recent studies have highlighted the high prevalence of CHR but these were often limited by small sample size (mostly relying on local data) and limited investigation of the characteristics of the CHR K. pneumoniae strains. In this study, we took advantage of the unique availability of a large collection of clinical isolates from various intensive care units (ICUs) across Europe to gain more insight into the frequency of occurrence of CHR among colistin-sensitive (CS) K. pneumoniae and to investigate its possible association with epidemiological and clinical characteristics such as country of origin, the different therapeutic intervention strategies to which the patients were exposed during their stay in the ICU and sequence type (ST type). We also investigated CHR K. pneumoniae strains for their possible association with, capsular polysaccharide antigen types (K-antigen type), lipopolysaccharide antigen types (O-antigen type) and colistin MIC value.
2. Materials and Methods
2.1. Sample and Isolate Collection
Klebsiella pneumoniae (n=676) collected during a clinical trial as part of the R-GNOSIS project (Resistance in Gram-Negative Organisms: Studying Intervention Strategies) (NCT02208154) were utilized. Isolates were collected from patients hospitalized in ICUs from 13 sites in six European countries: Belgium (n=239), Spain (n=201), Portugal (n=61), Italy (n=143), Slovenia (n=9), and United Kingdom (n=23) between December 1,2013 till May 31, 2017. ICU patients with an expected duration of invasive mechanical ventilation of minimally 24h were included while those who were not intubated nor mechanically ventilated and those that stayed in the unit for less than 24h were not included in the study. In the primary R-GNOSIS clinical trial, participants were assigned to three different groups of intervention, namely chlorhexidine mouthwash (CHX), selective oropharyngeal decontamination (SOD) and selective digestive tract decontamination (SDD), aimed at reducing the risk of bloodstream infections due to MDR-GNB among ventilated patients in intensive care units (ICUs) with moderate to high prevalence of antibiotic resistance [12]. Both SDD and SOD topical decontamination treatments consist of an oropharyngeal paste and enteral suspension containing antimicrobials that includes colistin (as well as tobramycin and nystatin) (Supplementary Table S1). After a baseline period (6-14 months) each intervention was implemented for periods of six months in a random order for each ICU and was separated by a one-month wash-out/in period. Samples and subsequently culture-isolated microorganisms were categorized into three categories: surveillance, point prevalence survey (PPS) or clinical. Surveillance samples were from patients directly undergoing interventions (i.e. who had received the decontamination treatment) and were taken twice weekly. PPS samples were collected monthly from all patients present in the ward at that time, including those not undergoing the interventions. Clinical samples were those obtained when needed for the clinical management of the patients [12]. Isolates originated from various body sites: respiratory (more specifically aspirate, throat swab, sputum, bronchoalveolar lavage or non-directed bronchoalveolar lavage), blood, groin or rectum.
2.2. MIC Determination
All 676 first patient isolates were subjected to colistin susceptibility testing using either an E-test (bioMérieux) or automatic testing method (BD PhoenixTM, SensititreTM, Vitek®, MicroScan) [12]. Of the 676 isolates, 79 (11.7%) were determined to be fully CR. To confirm the susceptibility of the selected isolates (see population analysis profiling assay), the colistin MIC was determined by broth microdilution using the MICRONAUT MIC-Strip Colistin (MERLIN Diagnostika GmbH, Berlin, Germany). In case of one skipped well, the result was determined disregarding this well (i.e. the skipped well was not seen as the lowest concentration showing no growth). In case of multiple skipped wells, the test was repeated. An isolate was classified as susceptible (MIC ≤2 mg/L) or resistant (MIC >2 mg/L) based on the epidemiological cut-off (ECOFF) values provided by EUCAST [13]. Two CS strains Escherichia coli ATCC 25922 (MIC: 0.25-2 mg/L) and P. aeruginosa ATCC 27853 (MIC: 0.5-4 mg/L) and two CR strains K. pneumoniae 08400 (MIC: 64 mg/L) and E. coli NCTC 13846 mcr-1 positive (MIC: 4 mg/L) were used as controls [14].
2.3. Population Analysis Profiling (PAP) Assay
For HR screening, we randomly selected 288 isolates of the 597 CS (MIC ≤2 mg/L) isolates for population analysis profiling (PAP) based on the two following criteria: (1) Number of isolates selected per country had to reflect the proportion of isolates contributed by each country and also (2) to match with the distribution of isolates found during the baseline and the three intervention strategies (Supplementary Table S1).
For the PAP assay, 0.5 MacFarland (McF) bacterial suspensions were prepared using the BD PhoenixSpec™ nephelometer (BD Diagnostics, Le Pont de Claix, France) starting from an overnight culture on Columbia blood agar (Oxoid Ltd, Basingstoke, UK) with 5% defibrinated horse blood (International Medical Products, Oudergem, Belgium). A 100 µl aliquot of this suspension was spirally plated using the Eddy Jet (IUL instruments S.A., Barcelona, Spain) on a series of Mueller Hinton Agar (CAMHA) (BD Diagnostics, Le Pont de Claix, France) plates containing colistin (Sigma-Aldrich, St. Louis, MO, USA) in increasing concentrations (0 mg/L, colistin free; 1 mg/L; 2 mg/L; 4 mg/L, 8 mg/L; 16 mg/L). The number of colonies were counted after 24 h of aerobic incubation of the plates at 37°C, and a graph of the log10 CFU/mL was plotted against the increasing colistin concentrations. One CS strain E. coli ATCC 25922 (MIC: 0.25-2 mg/L), one CR strain K. pneumoniae 08400 (MIC: 64 mg/L) and two CHR strains: K. pneumoniae ATCC 13883 (MIC: 1 mg/L) and K. pneumoniae IT0244CP (MIC: 0.5 mg/L) were used as quality controls [14].
Since there is no consensus on the definition of CHR, we used two previously defined schemes (Figure 1). Classification 1 (C1) was based on the classification as used by Band et al. [15] while classification 2 (C2) was based on Andersson et al. [9] with the additional requirement that growth at a frequency of minimally 1x10-7 must be observed at least at 4 mg/L even if the 8-fold MIC of the isolate was below 4 mg/L. More specifically, this means that for a MIC of ≤0.5 mg/L, growth at a frequency of 1x10-7 must be observed on plates containing 1, 2 and 4 mg/L of colistin, for a MIC of 1 mg/L the former must be observed additionally on the plates containing 8 mg/L of colistin and for a MIC of 2 mg/L also on plates containing 16 mg/L. This additional requirement was put in place to account for the possibility of false positives occurring due to the inoculum effect. The frequency was determined for each concentration of colistin using the following calculation:
2.4. Whole Genome Sequencing
Whole genome sequencing (WGS) was employed to determine the ST type, O-antigen type and K-antigen type of the 676 K. pneumoniae isolates as well as to look for mutations in known (hetero)resistance genes for colistin. Strains were cultured on CAMHA and incubated for 16-20h at 35-37oC. After incubation, one colony of the CAMHA plate was inoculated in a polypropylene tube containing 4 ml of cation adjusted Mueller Hinton Broth (CAMHB) (BD Diagnostics, Le Pont de Claix, France) and incubated again for 16-20h at 35-37oC. A negative growth control was prepared containing only 4 ml of CAMHB. DNA extraction was performed using the MasterPureTM Complete DNA and RNA Purification Kit (Epicentre Biotechnologies, Madison, WI, USA) following manufacturer’s instructions. DNA was further purified using the DNA Clean & ConcentratorTM-10 kit (Zymo Research, Irvine, CA, USA) following instructions as provided by the manufacturer. Library preparation was performed using the Nextera® XT DNA Sample Preparation Kit and Nextera® XT Index Kit v2 Set A (Illumina, San Diego, CA, USA) in conjunction with the Zephyr® G3 NGS liquid handler (PerkinElmer, Waltham, MA, USA), containing heating and shaking modules controlled by the Inheco Multi TEC Controller (INHECO GmbH, Martinsried, Germany). Sequencing was performed with the MiSeq sequencer (Illumina, San Diego, CA, USA). Data was analyzed using BacPipe v6.0 [16] and CLC Genomics Workbench software (Qiagen, Hilden, Germany).
2.5. Statistical Analysis
To determine whether there was an association between CHR/CR and country, MIC-value, intervention strategy, ST-, O-antigen or K-antigen type and between country, ST type and MIC-value, a Pearson Chi2 test or Fisher Exact test was used. When conditions for a Chi2 test or Fisher Exact test were not met (i.e., no cells with expected values <1, and no more than 20% of cells with values <5), a Monte Carlo simulation was used. Since this is not an exact method, in contrast to the regular Fisher Exact test, the P-value was given with the 99% confidence interval (99% CI). P-values less than 0.05 were considered statistically significant. In case of a statistically significant association, a pairwise z-test with Bonferroni correction was used to assess which groups had an association. All analysis were performed using the IBM® SPSS® Statistics software version 28.0.1.1. (IBM Corp., Armonk, NY, USA).
3. Results
3.1. Population Analysis Profiling (PAP)
The 288 selected isolates were tested by PAP assay in 11 distinct runs. Results of the PAP assay for the control strains in the 11 full runs are shown in Figure 2. One of the two HR control strains, IT0244CP grew on all colistin containing plates in all 11 runs, though the frequency varied from run to run. The frequency threshold of 1x10-6 was only reached twice (classification 1), still IT0244CP did always fulfil C2. The second CHR control strain K. pneumoniae ATCC 13883 also showed some variability in the frequency of the resistant subpopulation but fulfilled C1 in all 11 runs.
Overall, out of the 288 isolates tested, 25 were classified as being CHR based on the more stringent criteria of C1 whilst 108 isolates were classified as being CHR based on the less stringent C2 criteria. All isolates that fulfilled C1 also fulfilled C2 (Table 1 and Supplementary Table S2).
3.2. High Prevalence of Multi-Drug Resistant and Carbapenemase Producing Isolates
Overall, 618 out of 671 (92%) of the K. pneumoniae isolates, for which the information was available, were classified as MDR based on Magiorakos et al. [17]. MDR prevalence was high among both CS (91.2%, 540/592) and CR (98.7%, 78/79) isolates. In contrast, the proportion of carbapenemase producing K. pneumoniae (CP-Kpn) was higher among CR isolates (72.2%, 57/79) than among CS isolates (29.1%, 173/594). On the other hand, similar proportions of MDR and of CP-Kpn were observed among CHR (MDR: 85%, 91/107; CP-Kpn: 29.9%, 32/107) and CS isolates (MDR: 91%, 539/592; CP-Kpn: 29.1%, 153/59) (Supplementary Tables S3 and S4).
3.3. Analysis of Association between Colistin Resistant K. pneumoniae, ST Type, O-Antigen Type and K-Antigen Type
To investigate the distribution of CR within various ST types, we studied those ST types that were represented at least 10 times in our collection. The 79 CR K. pneumoniae isolates in our study were distributed among 14 different ST types with three STs (ST101, ST147, ST258/512) accounting for 62% (n=49) of the total CR isolates. CR was significantly associated with specific ST types (P at least 0.003), for instance, ST101 (CR: 12.7%, 10/79, CS: 2.7%, 16/597), ST147 (CR: 30.4%, 24/79, CS: 6.9%, 41/597) and ST258/ST512 (CR: 19%, 15/79, CS: 4.9%, 29/597) (P for all < 0.05) showed higher proportions among CR strains than among CS strains. Conversely, ST15 (CR: 3.8%, 3/79, CS: 11.4%, 68/597), ST307 (CR: 3.8%, 3/79, CS: 13.6%, 81/597) and ST405 (CR: 5.1%, 4/79, CS: 10.1%, 60/597) (P for all < 0.05) had higher proportions of CS isolates compared to CR. We could not document exclusive association either with CS or with CR of any ST type that was present at least 10 times in the collection (Figure 3A).
CS isolates (n=597) were distributed over 87 ST types (Simpson’s diversity index. SDI: 0.94) while the 288 CS isolates tested on the PAP assay belonged to 85 ST types (SDI: 0.97). Of these, CHR was found among 52 different ST types and showed a remarkably higher genetic diversity (SDI: 0.97) as compared to ST distribution among CR isolates (SDI: 0.85). Ten ST types (ST11, ST15, ST45, ST101, ST147, ST258/512, ST307, ST405, ST409 and ST437) were common to both CR and CHR isolates.
Besides an association between CR and ST types, country and ST types also showed a strong association (P at least <0.001). K. pneumoniae belonging to ST15 were spread across all six countries, however, the proportion of ST15 was higher in Belgium (17.2%, 41/239) and Portugal (19.7%, 12/61) than in Italy (2.1%, 3/143) and Spain (4%, 8/201) (P < 0.05). ST307 was present in five of the six countries. In this case the proportion of ST307 isolates in Italy (29.4%, 42/143) was significantly different compared to Belgium (7.9%, 19/239) and Spain (7.5%, 15/201) (P < 0.05) but not when compared to Portugal (11.5%, 7/61) (P > 0.05). ST147 was not found in Italy and most of the isolates were from Spain (26.9%, 54/201) for which the proportion also significantly differed from both Belgium (1.7%, 4/239) and Portugal (9.8%, 6/61) (P < 0.05). In contrast to the previous ST types which were found in almost all countries, ST101 and ST258/ST512 were only found in two and three countries respectively. ST101 showed a statistically significant difference between the two countries with a higher proportion in Italy (14.7%, 21/143) compared to Spain (2.5%, 5/201) (P < 0.05). ST258/ST512 showed a statistically significant difference between Belgium (10.5%, 25/239) and Spain (2.5%, 5/201) (P < 0.05), but not between Italy (9.1%, 13/143) and the other two countries (P > 0.05). Among ST101, all CR isolates were isolated in Italy, for ST147 all but one were isolated in Spain (one CR isolate from Portugal) and for ST258/ST512 all but two from Belgium (two CR isolates from Italy) (Figure 3C).
For the K-antigen and O-antigen types, a tight association is known to exist with specific ST types and therefore the significant differences found in the proportion of CR and CS within an ST type were likewise reflected in similar differences in the proportion of K and O antigen types (P for both ≤ 0.001).
3.4. A Higher Proportion of CR among Specific ST Types Was Not Reflected in CHR Proportions
For CHR and ST type an association was found (P at least 0.021). In contrast, CHR isolates did not show any clear association with the K-antigen and O-antigen type (P ≥ 0.169). ST307 and ST405 were found to have a higher proportion of non-CHR isolates (ST307: CHR: 3.7%, 4/108, non-CHR: 12.8%, 23/180, P < 0.05 and ST405: CHR: 0.9%, 1/108, non-CHR: 5.6%, 10/180, P < 0.05) (Figure 3B). Selected isolates from both ST types were spread across different countries (Figure 3D). There was no association between ST types found to have a higher proportion of CR isolates and CHR. Of the 39 isolates tested for CHR with ST types ST101, ST147 and ST258/ST512, only two determined to be CHR based on C1 and 14 exclusively based on C2.
3.5. Analysis of Association between Colistin (Hetero)Resistance K. pneumoniae, Country, Intervention Strategy and MIC-Value
The highest proportion of CR K. pneumoniae was found in Spain (44.3%, 35/79, P < 0.05) (Figure 4A). There was also a difference in the distribution of CR across the baseline, CHX, SDD and SOD (P = 0.001) with a higher proportion of CR isolates in patients in all three intervention strategy groups as compared to the baseline period (P < 0.05) but no significant difference in distribution of CR was identified between the three intervention groups (Supplementary Figure S1). This could mainly be explained by the increase in the proportion of CR ST147 isolates in Spain during the intervention periods (for all three groups) as compared to the baseline (8.8-15.6% versus 3.8%). In contrast, no significant association was found between increase in CHR with any country (P > 0.723) (Figure 4B) or between CHR and any intervention strategy (P > 0.668). CHR isolates were present in the highest proportions in ST15 followed by ST101 (Figure 3B). Of note, CHR isolates that matched the stricter definition (i.e. C1) were found in higher proportion in ST15 followed by ST307 but they were not found at all among the ST101 and ST147 isolates (Figure 3B).
We could not find any association between colistin MIC value of the 288 selected isolates and countries (P at least 0.599) (Figure 5B) nor was an association present between ST type and MIC value (P at least 0.08) (Figure 5C). Remarkably, however, there was a trend towards higher colistin MIC values for isolates classified as CHR based on C1 compared to CHR isolates fulfilling only C2 (Figure 5A).
3.6. Colistin Resistance Mechanisms in Resistant Subpopulations
Isolates AN1505CP2 and IT0244CP were previously already determined to be CHR [14]. In this study these findings were confirmed with both isolates fulfilling classification 1, and colonies from each plate were sequenced (data not published). Mutations found in colistin resistance-associated genes (mgrB, phoP, phoQ, pmrA, pmrB, pmrD, arnA, kpnEF, kpnF, crrB and acrB) are summarized in Table 2. The resistant subpopulations of AN1505CP2 and IT0244CP both showed disruptions of the mgrB gene by various insertion sequences (IS1R, IS1X2, ISKpn34, IS903B).
For IT0244CP disruption of mgrB was found across different concentrations of colistin in the same PAP assay (but these were linked with different IS and interruption at different nucleotide positions) (Table 2). In contrast, the resistant subpopulation of AN1505CP2 did show differences in the type of mutations found in mgrB between different concentrations of colistin in the same PAP assay run. To assess whether the mechanism of resistance was ST type specific or stochastic, we looked into CR isolates with the same ST type as IT0244CP (ST409). For AN1505CP2 (ST323), we could not find in our collection any CR isolates that belong to the same ST type (Table 2). For ST409, both the CR and CHR isolates showed disruptions of mgrB by insertion sequences. However, we did not find reproducibility in the mutations selected even for the same (CHR) isolate between technical replicates of the PAP assay highlighting the stochasticity of the selection process.
4. Discussion
In general, research around HR has been proven to be challenging especially due to a lack standardization of identification methods as well as of precise classification criteria. In the past it has therefore also been difficult to come to reliable estimates on the prevalence of HR as the sample sizes of the performed studies have been generally small and, due to a lack of standardization, both in definitions of HR but also in assays, it has been difficult to compile and compare different studies [1,9]. There are various ways through which HR can be screened, including methods that are used routinely for example for MIC determination, but they have been deemed to be not performant enough [9,18,19].
A strength of this study was its large sample size compared to previous studies on CHR using the PAP assay [20,21,22,23]. Within this study two different classification schemes for CHR were used. Classification 1 was considered to be more stringent due to the higher frequency requirement and the number of isolates fulfilling this more stringent classification CHR was thus limited (n=25). The majority of isolates categorized as CHR fulfilled the second less stringent classification (n=108) which was considered to be less stringent due to a lower frequency requirement though this classification included an additional requirement with regard to the concentration at which growth should occur. It has to be acknowledged that there were also numerous isolates that could not classified as CHR by any of these two classifications but did show growth on PAP assay plates >2 mg/L colistin. Most often, this was growth at a frequency <1x10-7 (often only one colony). We cannot rule out that these colonies were spontaneous mutants and were thus not caused by the isolate being CHR or were due to an inoculum effect. They were therefore not classified as CHR.
This study was also unique in the associations investigated, which to our knowledge, have not been studied previously in K. pneumoniae. A study on CHR in A. baumannii reported the absence of association between CHR and MIC-value and clonal complexes [22], however, these results are not necessarily applicable to CHR in K. pneumoniae. In this study, we could not find any association between CHR and country, intervention strategy, K-antigen type or O-antigen type or colistin MIC value. For the ST type an association was found, however, only when classification 2 was used. In contrast, an association was found between CR and country, ST type, O-antigen type and K-type. It is interesting to note that, based on an annual report from the European Centre for Disease Prevention and Control (ECDC) on antimicrobial consumption, Spain, which was found to have a higher number of CR isolates, also had a relatively higher consumption of polymyxins in hospitals compared to other European countries at least in the last period of the trial in which these isolates were collected (2016-2017) [24]. Also during 2017, colistin consumption in Spain remained relatively much higher compared to other countries included here [24]. Unfortunately, no data was available for a larger part of the duration of the trial (2013-2016).
For the ST-, O-antigen and K-antigen type, we often saw a relationship between the different molecular indicators and the ST types found to have a statistically significant difference in the amount of CS and CR isolates e.g. a specific K-antigen type was only present in combination with a specific ST type and were both found to have a higher proportion of CR isolates. For CHR, interestingly, no such pattern was found. CHR ST types found to have an association with CR in this study are especially interesting since more than 90% of them are known to be associated with multidrug resistance [25,26,27]. In this study, MDR rates were high in CS, CHR and CR isolates (>75%) and around one third of the isolates determined to be CHR were CP-Kpn. Especially CHR in MDR CP-Kpn is especially important to report as in most sites having a historically high proportion of CP-Kpn, colistin is empirically prescribed and knowledge on the predilection towards development of CHR might facilitate the use of tailored antibiotic combinations instead.
In contrast to CS and CHR, the proportion of CR CP-Kpn isolates was relatively high (±30% vs. ±70%). Since CP-Kpn are more likely to be exposed to colistin, it is to be expected that CR rates will increase over time in this subset of isolates which is also reflected in this study with the far majority of CP-Kpn being CR. Interestingly, rates of CHR among CP-Kpn were similar to CS among CP-Kpn, and CHR rates also did not differ significantly between the baseline, CHX, SOD and SDD. This also suggests that colistin exposure through SOD and SDD had no association with selection of isolates with a CHR phenotype.
Finally, a closer look at the mechanism of colistin resistance for the resistant subpopulation of two CHR isolates showed that mutations in colistin resistance-conferring genes assessed in this study could not always be identified for each isolate. A recent study on CHR in wild-type K. pneumoniae isolates also reported that for 28% of mutants sequenced, no genetic modification was found in the panel of genes assessed [28]. Disruption of mgrB by insertion sequences was the most commonly found genetic modification in the resistant subpopulations. Additionally, a nonsense mutation and complete deletion of mgrB was found once and twice, respectively. A mutation outside of mgrB was only found once. The same study in wild-type K. pneumoniae isolates also reported a high number of mgrB genetic modifications (54%) [28]. A complete deletion was only found in 4% of the mutants while disruption by insertion sequences and other amino acid alterations were found in 28% and 22% of mutants respectively [28]. Luo et al. also reported in their study that there was a high rate of mgrB insertional mutations and no mutations in pmrAB or phoPQ and stated that this was consistent with previous findings which showed that those genes had a significantly lower mutation rate compared to mgrB. However, they also stated that the high amount of mgrB disruptions may be related to the high prevalence of ST11 since this ST type showed a significantly higher rate of mgrB disruptions compared to other ST types in their study [29].
Though this study has helped in expanding the knowledge on CHR in a clinical setting, there are also some limitations. Firstly, only limited sequencing data was available for CHR isolates. Future studies are needed to further assess the diversity of mechanisms of CHR and whether these mechanisms are ST-specific. Secondly, we cannot exclude confounding factors such as the usage of more colistin in some local settings, outbreaks with specific STs, and the prevalence of CP Kpn which may vary between sites. Thirdly, we only studied the CS population for CHR. However, CHR can also exist as a (sub)proportion of CR isolates. Additionally, we only focused on CHR as a phenomenon in which there is a minor subpopulation with a MIC above the breakpoint in a major population with a MIC below the breakpoint. However, HR may also occur in entirely susceptible populations [30].
Given the large number of isolates screened, this study is a step forward in elucidating the prevalence and burden of CHR in common ST lineages of K. pneumoniae. Our data prompt for development of more robust and simple diagnostics to enable implementation of HR detection on a larger scale, and for more structured studies to quantify the actual impact of CHR on treatment failures in patients receiving colistin.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary Table S1: Summary of interventions including amount of isolates within each intervention per sample category Table provides an overview of the contents of the different intervention strategies as well as the number of isolates sorted per sample category for both the complete collection (n=676) and the selected subset (n=288) of isolates. Surv. = surveillance, Clin. = clinical, CS = colistin sulphate, TBS = tobramycin sulphate, NYS = nystatin. 12% mouthwash replaced by 1% oral gel after reports of oral mucosal adverse events. 2Normally regiment includes four days IV cephalosporin but not included because of settings of moderate/high resistance. 3Either sampling during 1 month wash-out/in period between intervention strategies or interruption of the intervention. Supplementary Table S2: Overview of selected isolates. Table gives an overview of the selected isolates (n=288) and their characteristics. Cat. = category, Surv. = surveillance, Clin. = clinical, Undef. = undefined. Supplementary Table S3: Number of MDR and CP-Kpn isolates. Tables gives summary of the number of MDR and CP-Kpn CS and CR isolates. For CS, MDR and CP-Kpn classification was not possible for five and three isolates respectively. Supplementary Table S4: Number of MDR and CP-Kpn isolates. Tables gives summary of the number of MDR and CP-Kpn selected CS and CHR isolates. For CS, MDR and CP-Kpn classification was not possible for four and three isolates respectively of which one isolate was also CHR following C2. For CHR, C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. Supplementary Figure S1: Distribution of isolates across baseline and intervention strategies. Graphs show the number of isolates per strategy as well as the number of CR/HR and CS/non-CHR isolates. Of note C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. (A) CR per intervention strategy, (B) CHR per intervention strategy.
Author Contributions
Conceptualization and study design: S.M.-K., Y.G; Experimental execution: A. B., S.G.R., A.S., E.B.M., and C.L.; Bioinformatics: S.G.R.; Manuscript writing: S.M.-K., Y.G. and A.B.; Manuscript review and editing: all authors. All authors have read and agreed to the published version of the manuscript.
Financial Support: S.G.R. and part of this study are funded by University of Antwerp BOF Doctoral Project Funds (BOF-DOCPRO 2019–project ID 40179). S.M.-K. gratefully acknowledges University of Antwerp Methusalem funding (Vaccine & Infectious Diseases Excellence in Antwerp: Infectious disease prevention, control and management in a One Health policy context (VAX-IDEA).
Acknowledgments
We would like to thank Sabine Chapelle, Gert Leten, Liesbeth Bryssinck and Joyce Jacobs for excellent technical and administrative support.
Conflicts of Interest
We declare no other conflict of interest.
References
- Roch, M., R. Sierra, and D. O. Andrey. "Antibiotic Heteroresistance in Eskape Pathogens, from Bench to Bedside." Clin Microbiol Infect (2022). [CrossRef]
- Della Rocca, M. T., F. Foglia, V. Crudele, G. Greco, A. De Filippis, G. Franci, E. Finamore, and M. Galdiero. "Antimicrobial Resistance Changing Trends of Klebsiella Pneumoniae Isolated over the Last 5 Years." New Microbiol 45, no. 4 (2022): 338-43.
- Mohd Asri, N. A., S. Ahmad, R. Mohamud, N. Mohd Hanafi, N. F. Mohd Zaidi, A. A. Irekeola, R. H. Shueb, L. C. Yee, N. Mohd Noor, F. H. Mustafa, C. Y. Yean, and N. Y. Yusof. "Global Prevalence of Nosocomial Multidrug-Resistant Klebsiella Pneumoniae: A Systematic Review and Meta-Analysis." Antibiotics (Basel) 10, no. 12 (2021). [CrossRef]
- Wang, M. G., M. Earley, L. Chen, B. M. Hanson, Y. S. Yu, Z. Y. Liu, S. Salcedo, E. Cober, L. J. Li, S. S. Kanj, H. Gao, J. M. Munita, K. Ordoñez, G. Weston, M. J. Satlin, S. L. Valderrama-Beltrán, K. Marimuthu, M. E. Stryjewski, L. Komarow, C. Luterbach, S. H. Marshall, S. D. Rudin, C. Manca, D. L. Paterson, J. Reyes, M. V. Villegas, S. Evans, C. Hill, R. Arias, K. Baum, B. C. Fries, Y. Doi, R. Patel, B. N. Kreiswirth, R. A. Bonomo, H. F. Chambers, VG Fowler, C. A. Arias, D. van Duin, and Multi-Drug Resistant. "Clinical Outcomes and Bacterial Characteristics of Carbapenem-Resistant Complex among Patients from Different Global Regions (Crackle-2): A Prospective, Multicentre, Cohort Study." Lancet Infectious Diseases 22, no. 3 (2022): 401-12. [CrossRef]
- Ah, Y. M., A. J. Kim, and J. Y. Lee. "Colistin Resistance in Klebsiella Pneumoniae." Int J Antimicrob Agents 44, no. 1 (2014): 8-15. [CrossRef]
- Ahmed, M. A. G. E. S., L. L. Zhong, C. Shen, Y. Q. Yang, Y. Doi, and G. B. Tian. "Colistin and Its Role in the Era of Antibiotic Resistance: An Extended Review (2000-2019)." Emerging Microbes & Infections 9, no. 1 (2020): 868-85. [CrossRef]
- Band, V. I., S. W. Satola, E. M. Burd, M. M. Farley, J. T. Jacob, and D. S. Weiss. "Carbapenem-Resistant Klebsiella Pneumoniae Exhibiting Clinically Undetected Colistin Heteroresistance Leads to Treatment Failure in a Murine Model of Infection." mBio 9, no. 2 (2018). [CrossRef]
- Moosavian, M., S. Shoja, R. Nashibi, N. Ebrahimi, M. A. Tabatabaiefar, S. Rostami, and A. Peymani. "Post Neurosurgical Meningitis Due to Colistin Heteroresistant Acinetobacter Baumannii." Jundishapur J Microbiol 7, no. 10 (2014): e12287. [CrossRef]
- Andersson, D. I., H. Nicoloff, and K. Hjort. "Mechanisms and Clinical Relevance of Bacterial Heteroresistance." Nature Reviews Microbiology 17, no. 8 (2019): 479-96. [CrossRef]
- Stojowska-Swedrzynska, K., A. Lupkowska, D. Kuczynska-Wisnik, and E. Laskowska. "Antibiotic Heteroresistance in Klebsiella Pneumoniae." Int J Mol Sci 23, no. 1 (2021). [CrossRef]
- Dewachter, L., M. Fauvart, and J. Michiels. "Bacterial Heterogeneity and Antibiotic Survival: Understanding and Combatting Persistence and Heteroresistance." Mol Cell 76, no. 2 (2019): 255-67. [CrossRef]
- Wittekamp, B. H., N. L. Plantinga, B. Cooper, J. Lopez-Contreras, P. Coll, J. Mancebo, M. P. Wise, M. P. G. Morgan, P. Depuydt, J. Boelens, T. Dugernier, V. Verbelen, P. G. Jorens, W. Verbrugghe, S. Malhotra-Kumar, P. Damas, C. Meex, K. Leleu, A. M. van den Abeele, A. F. G. P. de Matos, S. F. Mendez, A. V. Gomez, V. Tomic, F. Sifrer, E. Villarreal, J. R. Ramos, I. Aragao, C. Santos, R. H. M. Sperning, P. Coppadoro, G. Nardi, C. Brun-Buisson, and M. J. M. Bonten. "Decontamination Strategies and Bloodstream Infections with Antibiotic-Resistant Microorganisms in Ventilated Patients a Randomized Clinical Trial." Jama-Journal of the American Medical Association 320, no. 20 (2018): 2087-98. [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. "The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of Mics and Zone Diameters." https://www.eucast.org, 2023.
- Rajakani, S. G., B. B. Xavier, A. Sey, E. Mariem, C. Lammens, H. Goossens, Y. Glupczynski, and S. Malhotra-Kumar. "Insight into Antibiotic Synergy Combinations for Eliminating Colistin Heteroresistant." Genes 14, no. 7 (2023). [CrossRef]
- Band, V. I., D. A. Hufnagel, S. Jaggavarapu, E. X. Sherman, J. E. Wozniak, S. W. Satola, M. M. Farley, J. T. Jacob, E. M. Burd, and D. S. Weiss. "Antibiotic Combinations That Exploit Heteroresistance to Multiple Drugs Effectively Control Infection." Nature Microbiology 4, no. 10 (2019): 1627-35. [CrossRef]
- Xavier, B. B., M. Mysara, M. Bolzan, B. Ribeiro-Goncalves, B. T. F. Alako, P. Harrison, C. Lammens, S. Kumar-Singh, H. Goossens, J. A. Carrico, G. Cochrane, and S. Malhotra-Kumar. "Bacpipe: A Rapid, User-Friendly Whole-Genome Sequencing Pipeline for Clinical Diagnostic Bacteriology." iScience 23, no. 1 (2020): 100769. [CrossRef]
- Magiorakos, A. P., A. Srinivasan, R. B. Carey, Y. Carmeli, M. E. Falagas, C. G. Giske, S. Harbarth, J. F. Hindler, G. Kahlmeter, B. Olsson-Liljequist, D. L. Paterson, L. B. Rice, J. Stelling, M. J. Struelens, A. Vatopoulos, J. T. Weber, and D. L. Monnet. "Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance." Clin Microbiol Infect 18, no. 3 (2012): 268-81. [CrossRef]
- Juhasz, E., M. Ivan, E. Pinter, J. Pongracz, and K. Kristof. "Colistin Resistance among Blood Culture Isolates at a Tertiary Care Centre in Hungary." J Glob Antimicrob Resist 11 (2017): 167-70. [CrossRef]
- Sherman, E. X., J. E. Wozniak, and D. S. Weiss. "Methods to Evaluate Colistin Heteroresistance in Acinetobacter Baumannii." Methods Mol Biol 1946 (2019): 39-50. [CrossRef]
- Morales-Leon, F., C. A. Lima, G. Gonzalez-Rocha, A. Opazo-Capurro, and H. Bello-Toledo. "Colistin Heteroresistance among Extended Spectrum Beta-Lactamases-Producing Klebsiella Pneumoniae." Microorganisms 8, no. 9 (2020). [CrossRef]
- Band, V. I., S. W. Satola, R. D. Smith, D. A. Hufnagel, C. Bower, A. B. Conley, L. Rishishwar, S. E. Dale, D. J. Hardy, R. L. Vargas, G. Dumyati, M. A. Kainer, E. C. Phipps, R. Pierce, L. E. Wilson, M. Sorensen, E. Nilsson, I. K. Jordan, E. M. Burd, M. M. Farley, J. T. Jacob, R. K. Ernst, and D. S. Weiss. "Colistin Heteroresistance Is Largely Undetected among Carbapenem-Resistant Enterobacterales in the United States." mBio 12, no. 1 (2021). [CrossRef]
- Kon, H., A. Hameir, A. Nutman, E. Temkin, A. Keren Paz, J. Lellouche, D. Schwartz, D. S. Weiss, K. S. Kaye, G. L. Daikos, A. Skiada, E. Durante-Mangoni, Y. Dishon Benattar, D. Yahav, V. Daitch, M. Bernardo, D. Iossa, L. E. Friberg, U. Theuretzbacher, L. Leibovici, Y. Dickstein, D. Pollak, S. Mendelsohn, M. Paul, and Y. Carmeli. "Prevalence and Clinical Consequences of Colistin Heteroresistance and Evolution into Full Resistance in Carbapenem-Resistant Acinetobacter Baumannii." Microbiol Spectr (2023): e0509322. [CrossRef]
- Howard-Anderson, J., M. Davis, A. M. Page, C. W. Bower, G. Smith, J. T. Jacob, D. I. Andersson, D. S. Weiss, and S. W. Satola. "Prevalence of Colistin Heteroresistance in Carbapenem-Resistant Pseudomonas Aeruginosa and Association with Clinical Outcomes in Patients: An Observational Study." J Antimicrob Chemother 77, no. 3 (2022): 793-98. [CrossRef]
- European Centre for Disease Prevention Control. "Antimicrobial Consumption in the Eu/Eea (Esac-Net) - Annual Epidemiological Report 2021." https://www.ecdc.europa.eu/, 2022.
- Bialek-Davenet, S., A. Criscuolo, F. Ailloud, V. Passet, L. Jones, A. S. Delannoy-Vieillard, B. Garin, S. Le Hello, G. Arlet, M. H. Nicolas-Chanoine, D. Decre, and S. Brisse. "Genomic Definition of Hypervirulent and Multidrug-Resistant Klebsiella Pneumoniae Clonal Groups." Emerg Infect Dis 20, no. 11 (2014): 1812-20. [CrossRef]
- Peirano, G., L. Chen, B. N. Kreiswirth, and J. D. D. Pitout. "Emerging Antimicrobial-Resistant High-Risk Klebsiella Pneumoniae Clones St307 and St147." Antimicrob Agents Chemother 64, no. 10 (2020). [CrossRef]
- Rodrigues, C., S. Desai, V. Passet, D. Gajjar, and S. Brisse. "Genomic Evolution of the Globally Disseminated Multidrug-Resistant Klebsiella Pneumoniae Clonal Group 147." Microb Genom 8, no. 1 (2022). [CrossRef]
- Sanchez-Leon, I., E. Perez-Nadales, J. A. Marin-Sanz, T. Garcia-Martinez, and L. Martinez-Martinez. "Heteroresistance to Colistin in Wild-Type Klebsiella Pneumoniae Isolates from Clinical Origin." Microbiol Spectr 11, no. 6 (2023): e0223823. [CrossRef]
- Luo, Q., L. Xu, Y. Wang, H. Fu, T. Xiao, W. Yu, W. Zhou, K. Zhang, J. Shen, J. Ji, C. Ying, and Y. Xiao. "Clinical Relevance, Mechanisms and Evolution of Polymyxin B Heteroresistance Carbapenem-Resistant Klebsiella Pneumoniae: A Genomic, Retrospective Cohort Study." Clin Microbiol Infect (2024). [CrossRef]
- El-Halfawy, O. M. , and M. A. Valvano. "Antimicrobial Heteroresistance: An Emerging Field in Need of Clarity." Clin Microbiol Rev 28, no. 1 (2015): 191-207. [CrossRef]
Figure 1.
Classification schemes for CHR. Figure describes the growth requirements, more specifically the frequency of growth required, for an isolate to be determined CHR. In case there is no frequency requirement at a specific concentration of colistin but only the requirement that there is visible growth on the plate, this has been indicated as “Growth”. For classification 2 the requirements to be fulfilled depend on the MIC for colistin of the isolates. NR = no requirement, NA = not applicable. 1 Plates containing 16 mg/L of colistin were only included for isolates with a MIC of 2 mg/L. 2 Frequency of ≥1x10-6 only required for either 4 mg/L or 8 mg/L, not for both though allowed Created with BioRender.com.
Figure 1.
Classification schemes for CHR. Figure describes the growth requirements, more specifically the frequency of growth required, for an isolate to be determined CHR. In case there is no frequency requirement at a specific concentration of colistin but only the requirement that there is visible growth on the plate, this has been indicated as “Growth”. For classification 2 the requirements to be fulfilled depend on the MIC for colistin of the isolates. NR = no requirement, NA = not applicable. 1 Plates containing 16 mg/L of colistin were only included for isolates with a MIC of 2 mg/L. 2 Frequency of ≥1x10-6 only required for either 4 mg/L or 8 mg/L, not for both though allowed Created with BioRender.com.
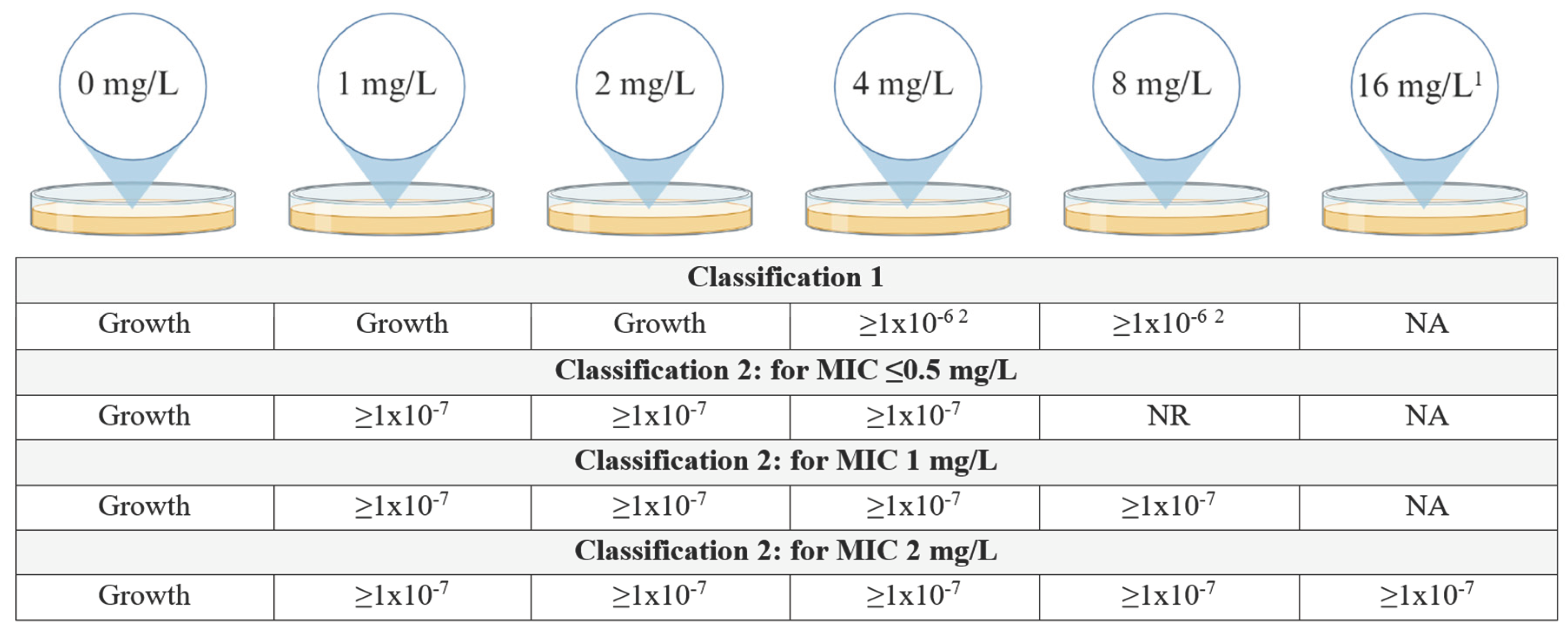
Figure 2.
PAP assay results of the control strains. Graph represents the log10 CFU/ml per concentration of colistin used in the CAMHA plates of the PAP assay. (A) Individual results for each strain for each run, graph illustrates the intra-run variation for the different control strains. (B) Average result for each control strain, graph illustrates the overall result of the control strains.
Figure 2.
PAP assay results of the control strains. Graph represents the log10 CFU/ml per concentration of colistin used in the CAMHA plates of the PAP assay. (A) Individual results for each strain for each run, graph illustrates the intra-run variation for the different control strains. (B) Average result for each control strain, graph illustrates the overall result of the control strains.
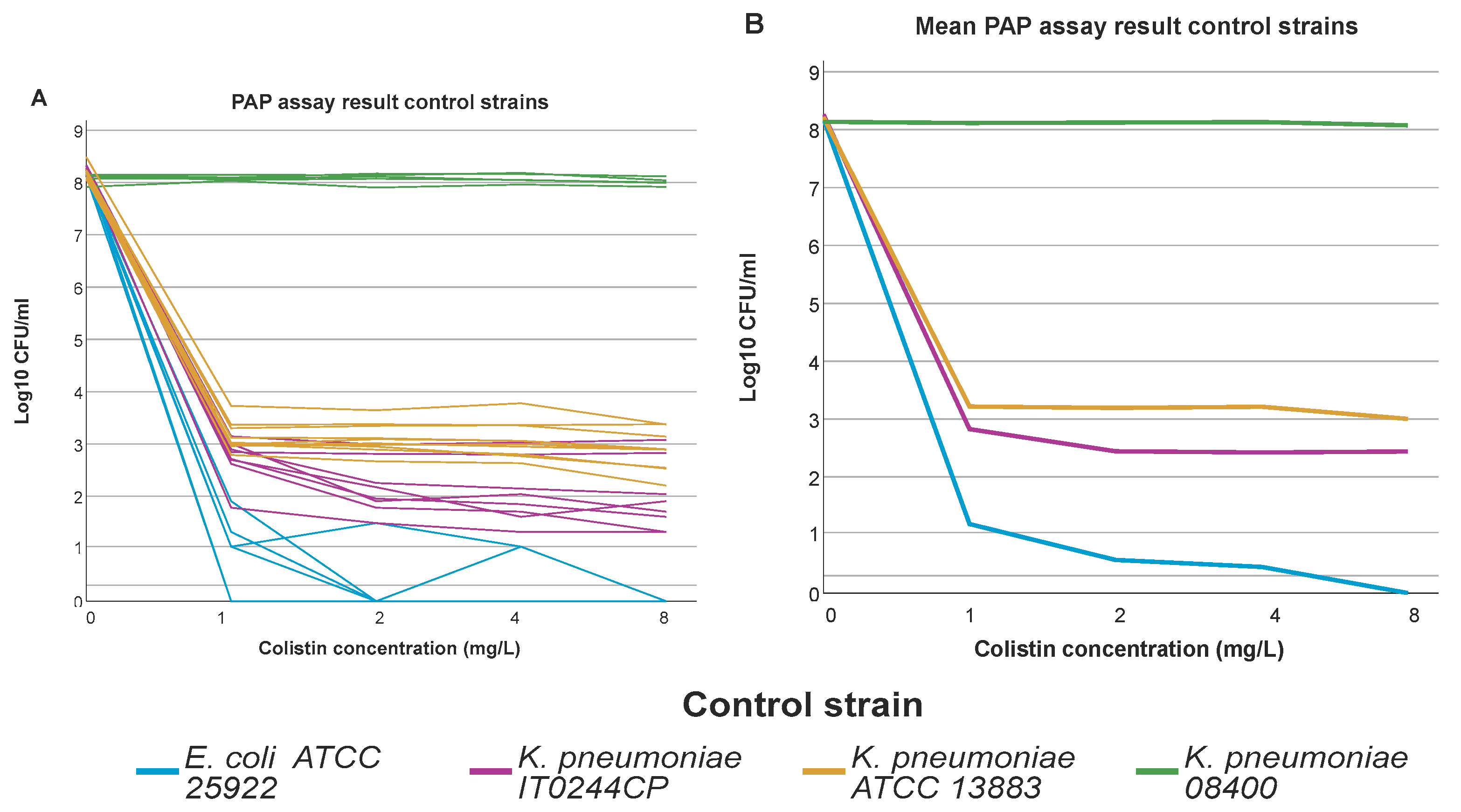
Figure 3.
Distribution of isolates across ST types. Graphs show the number of isolates per ST type as well as the number of CR/HR and CS/non-CHR isolates. Only ST types having a statistically significant association with CR and/or CHR are shown. Of note C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. (A) CR per ST type, (B) CHR per ST type, (C) CR per ST type per country, Slovenia (n=9) and United Kingdom (n=23) were not taken into further consideration due to the low number of isolates, (D) CHR per ST type per country.
Figure 3.
Distribution of isolates across ST types. Graphs show the number of isolates per ST type as well as the number of CR/HR and CS/non-CHR isolates. Only ST types having a statistically significant association with CR and/or CHR are shown. Of note C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. (A) CR per ST type, (B) CHR per ST type, (C) CR per ST type per country, Slovenia (n=9) and United Kingdom (n=23) were not taken into further consideration due to the low number of isolates, (D) CHR per ST type per country.
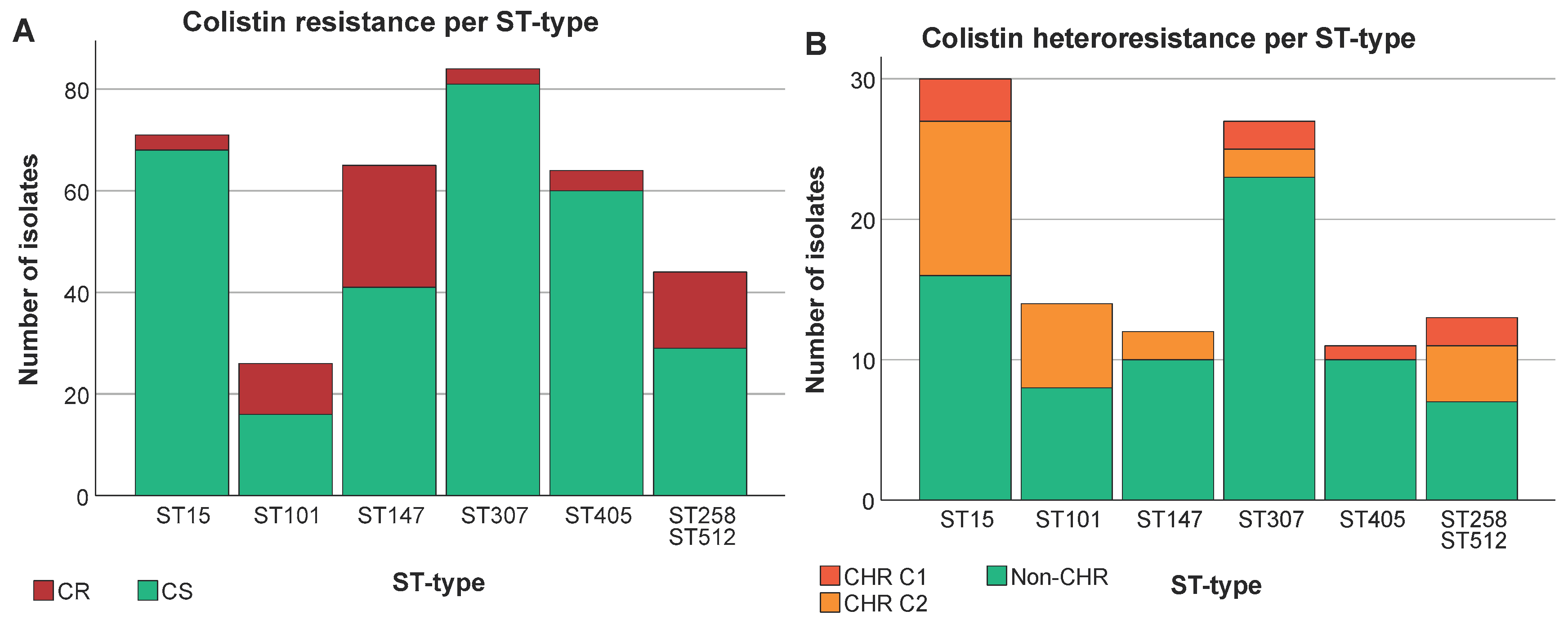
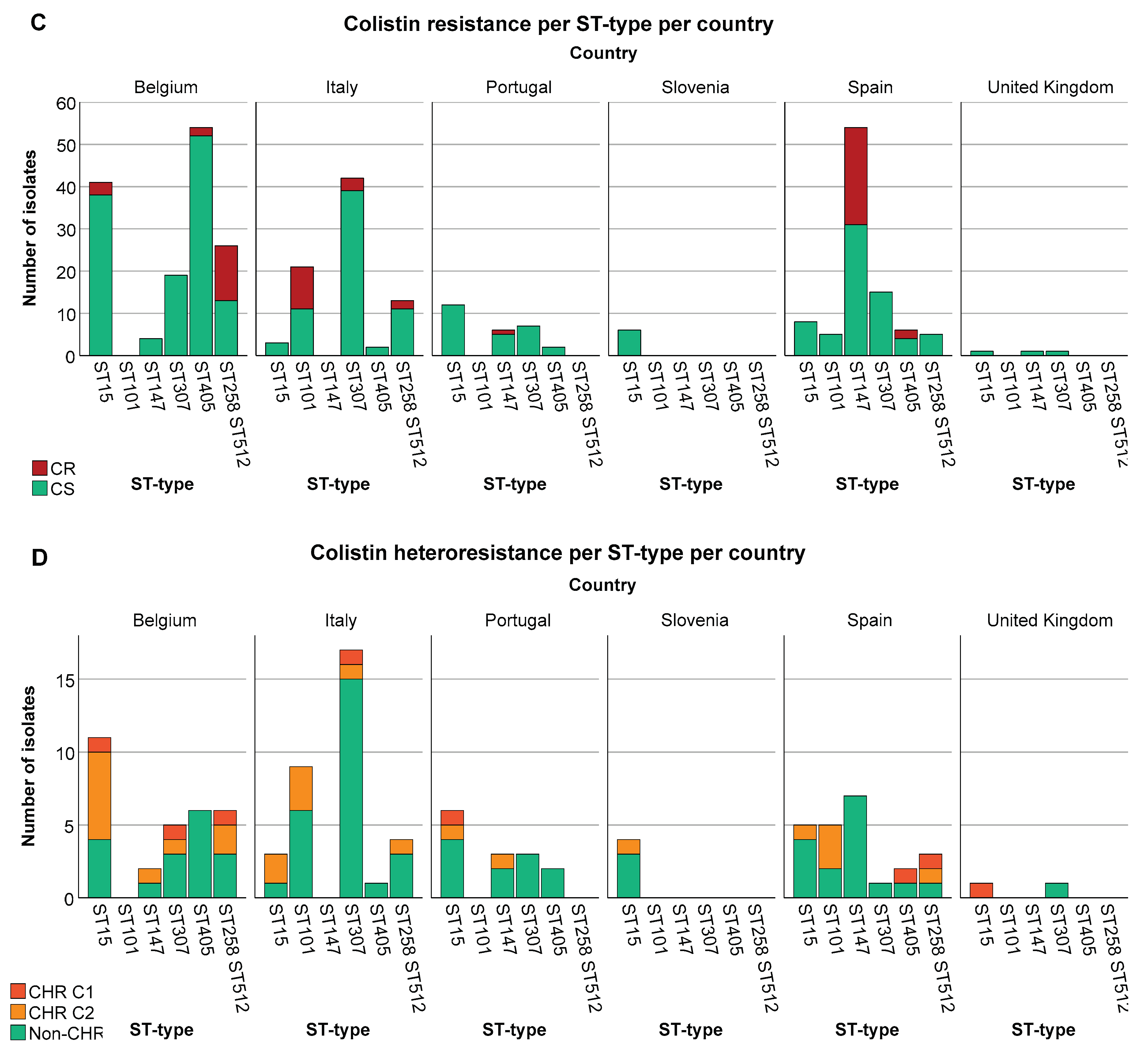
Figure 4.
Distribution of isolates across countries. Graphs show the number of isolates per country as well as the number of CR/HR and CS/non-CHR isolates. Of note C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. (A) CR per country, (B) CHR per country.
Figure 4.
Distribution of isolates across countries. Graphs show the number of isolates per country as well as the number of CR/HR and CS/non-CHR isolates. Of note C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. (A) CR per country, (B) CHR per country.
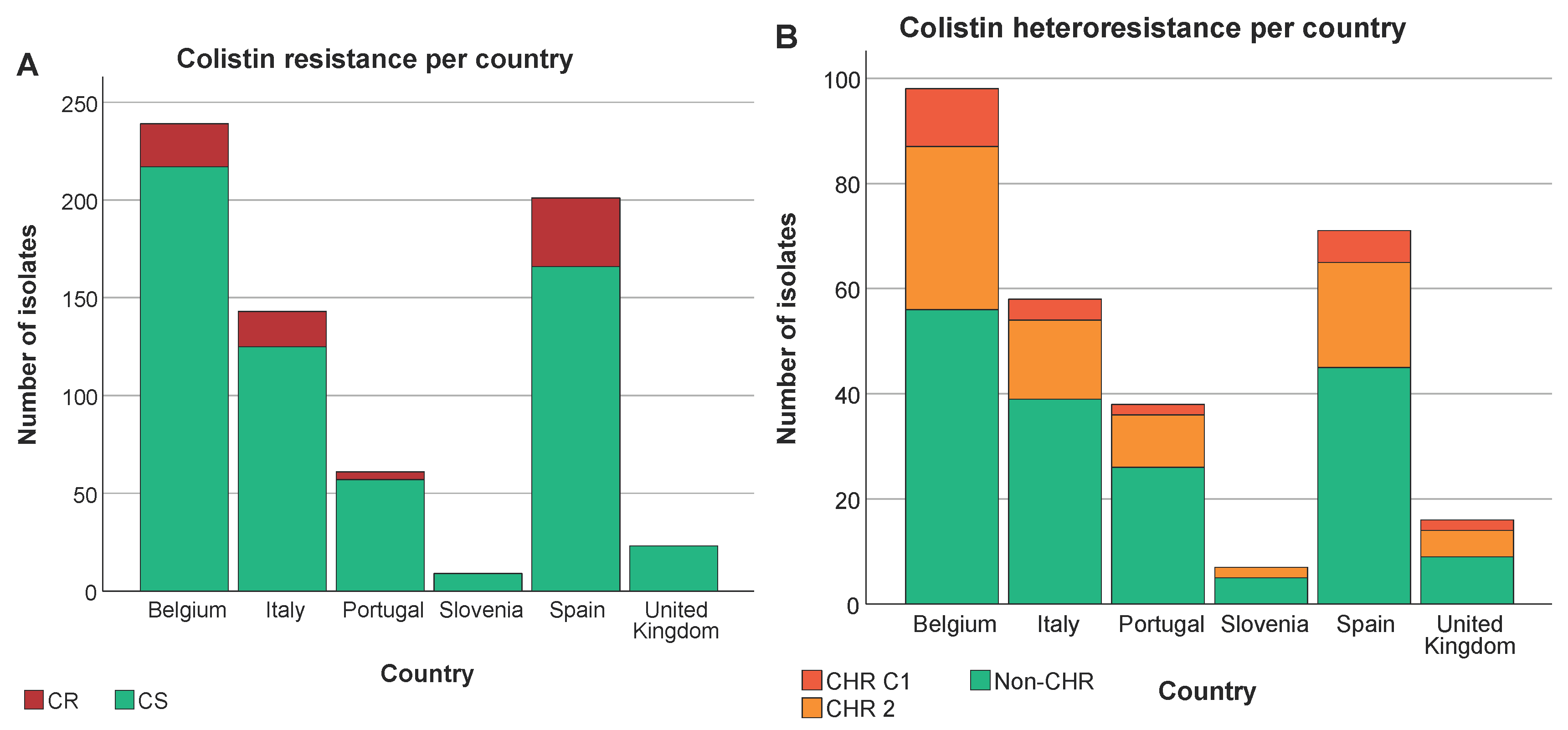
Figure 5.
Distribution of isolates across MIC values. Graphs show the number of isolates per MIC as well as the number of CHR and non-CHR isolates. Of note C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. (A) CHR per MIC value, (B) CHR per MIC value per country, (C) CHR per MIC value per ST type. Only ST types having a statistically significant association with CR and/or CHR are shown.
Figure 5.
Distribution of isolates across MIC values. Graphs show the number of isolates per MIC as well as the number of CHR and non-CHR isolates. Of note C1+C2 represents the total amount of isolates fulfilling C2 whilst C2 alone represents isolates only fulfilling C2. (A) CHR per MIC value, (B) CHR per MIC value per country, (C) CHR per MIC value per ST type. Only ST types having a statistically significant association with CR and/or CHR are shown.
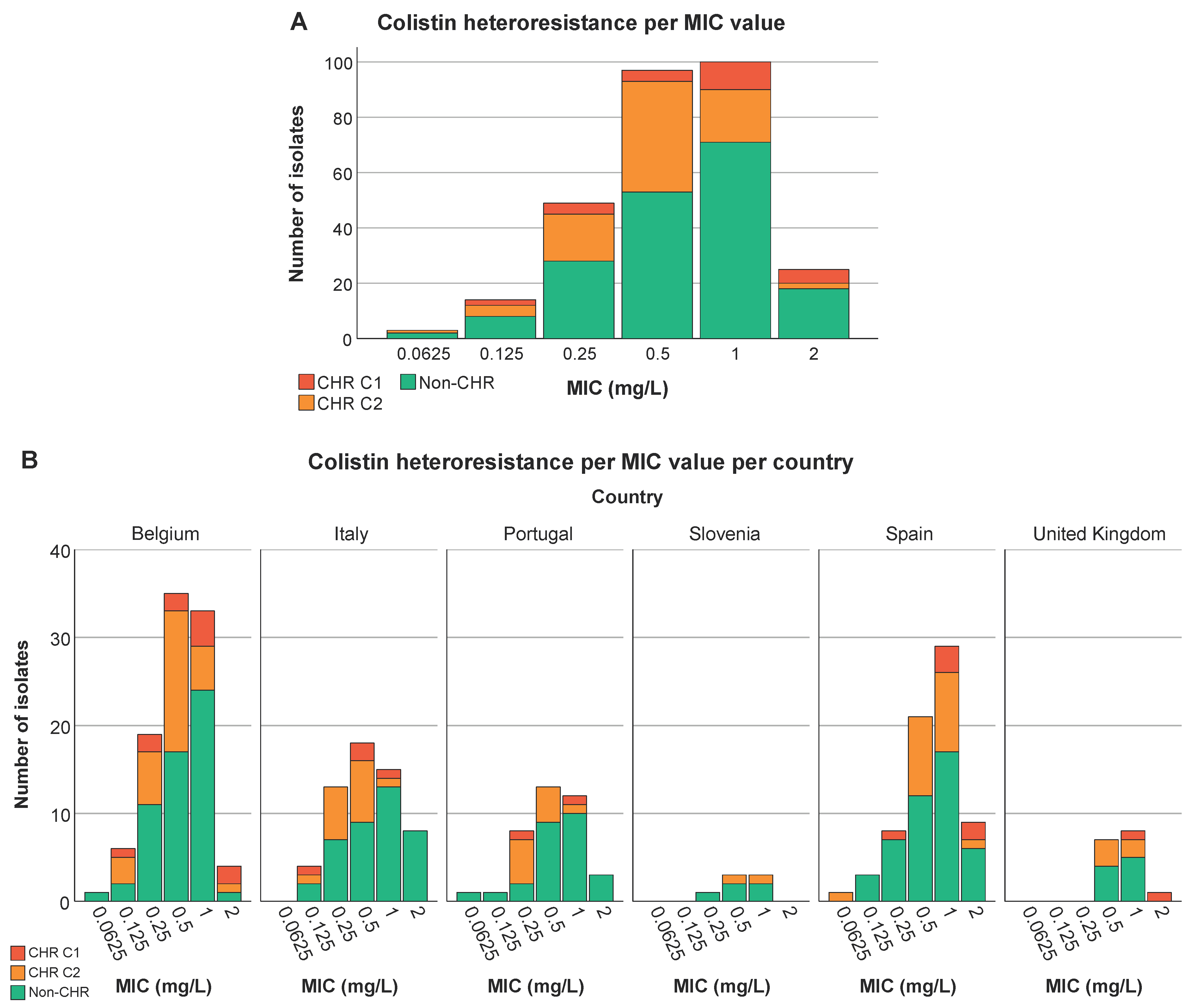
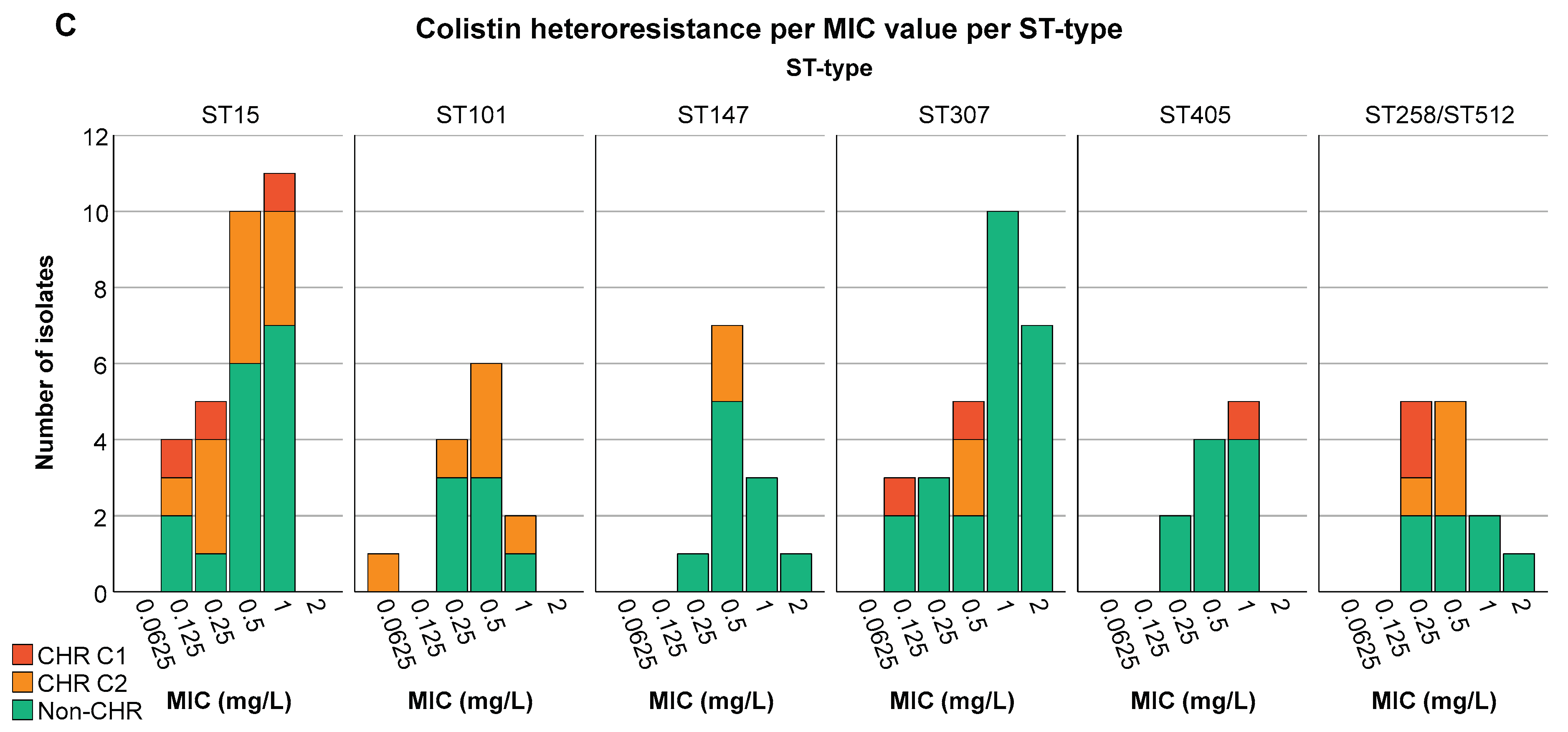

Table 1.
Table contains a detailed breakdown of the PAP assay results (n=288) including the reasons why isolates did not fulfil the definition of CHR and the number of isolates per observed result. Of note, for some isolates that did not fulfil the definition of CHR but did show growth >2 mg/L there were multiple reasons why they were not classified as CHR. Sub-reasons are listed in order of importance. Each isolate was only included once and if it fulfilled multiple sub-reasons it was only included in the sub-reason considered most important.
Table 1.
Table contains a detailed breakdown of the PAP assay results (n=288) including the reasons why isolates did not fulfil the definition of CHR and the number of isolates per observed result. Of note, for some isolates that did not fulfil the definition of CHR but did show growth >2 mg/L there were multiple reasons why they were not classified as CHR. Sub-reasons are listed in order of importance. Each isolate was only included once and if it fulfilled multiple sub-reasons it was only included in the sub-reason considered most important.
| Observed results | No. of isolates | |
|---|---|---|
| Fulfilling classification 1 | Growth on the plates containing 4 and/or 8 mg/L of colistin with a frequency of at least 1•10-6 | 25 |
| Fulfilling only classification 2 | Growth on at least all the plates containing colistin at a concentration up to and including 8-fold the MIC of the isolate at a frequency of minimally 1•10-7, minimum concentration at which there should be growth was 4 mg/L | 83 |
| Not fulfilling either classification but growth >2 mg/L | Growth on 4 and/or 8 mg/L plate but frequency <1•10-7 | 45 |
| For MIC 0.0625-0.5 mg/L: growth with frequency ≥1•10-7 on 8 mg/L plate but <1•10-7 on 4 mg/L plate | 8 | |
| For MIC 1-2 mg/L: growth with frequency ≥1•10-7 on 4 and/or 8 mg/L plate but <1•10-7 on plates ≥8-fold the MIC | 30 | |
| Growth with frequency ≥1•10-7 on plates ≥8-fold the MIC but frequency of 1•10-7 not reached on all plates <8-fold the MIC | 19 | |
| No growth at 4 and 8 mg/L | 71 | |
| No growth at 1, 2, 4 and 8 mg/L | 7 | |
Table 2.
Summary of mutations found in CR and CHR isolates. Table contains information on the mutations found in the resistant subpopulation of two confirmed CHR isolates (AN1505CP2 and IT0244CP) at different concentrations of the PAP assay plates on three separate assays. Additionally, table contains information on three CR isolates with the same ST type as IT0244CP (ST409). For AN1505CP2 there were no CR isolates with the same ST type (ST323).
Table 2.
Summary of mutations found in CR and CHR isolates. Table contains information on the mutations found in the resistant subpopulation of two confirmed CHR isolates (AN1505CP2 and IT0244CP) at different concentrations of the PAP assay plates on three separate assays. Additionally, table contains information on three CR isolates with the same ST type as IT0244CP (ST409). For AN1505CP2 there were no CR isolates with the same ST type (ST323).
| Isolate ID | MIC(mg/L) | ST type | PAP assay plate conc. (mg/L) | Mutations in mgrB | Other mutations |
|---|---|---|---|---|---|
| IT0307CP(CR) | 128 | ST409 | IS1R of IS1 family interruption at nt 107 | ||
| IT0636C(CR) | 128 | ST409 | ISKpn34 of IS3 family interruption at nt 46 | ||
| IT0915C(CR) | 64 | ST409 | IS903B of IS5 family interruption at nt 34 | ||
| IT0244CP(CHR1st PAP) | 0.5 | ST409 | 2 | ISKpn34 of IS3 family interruption at promoter | |
| 8 | IS903B of IS5 family interruption at nt 117 | ||||
| 16 | IS1S of IS1 family interruption at promoter | ||||
| IT0244CP(CHR3rd PAP) | 0.5 | ST409 | 2 | IS1X2 of IS1 family interruption at nt 123 | |
| AN1505CP2(CHR1st PAP) | 1 | ST323 | 4 | Deleted | |
| 8 | IS903B of IS5 family interruption at nt 70 | ||||
| AN1505CP2(CHR2nd PAP) | 1 | ST323 | 8 | pmrB: T157P | |
| AN1505CP2(CHR3rd PAP) | 1 | ST323 | 8 | Q30X | |
| 16 | Deleted |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



