
Submitted:
06 March 2024
Posted:
07 March 2024
You are already at the latest version
Abstract
Climate change is a big challenge for agriculture since it affects crop productivity and yield. The increase of droughts, salinity, and soil degradation are some of the main consequences of climate change. The use of microorganisms has emerged as an alternative to mitigate them. Among these microorganisms, dark septate endophytes (DSEs) have garnered more attention in recent years. Various studies show that their association with plants helps reduce the harmful effects of abiotic stresses and increases nutrient availability, allowing plants to thrive under adverse conditions. This work reviews the effect of DSEs and the subjacent mechanisms that will help plants develop a higher tolerance to climate change.
Keywords:
DSEs
; climate change
; drought
; salinity
; fertilisation
1. Introduction
The worrisome effects of climate change are causing long, severe periods of drought [1] in which agricultural production in Mediterranean regions is almost impossible for extensive farming, and in the case of intensive farming, water availability is notoriously reduced. The interactions between microorganisms and plants have also been affected by droughts and soil warming, causing a decrease in symbiotic relationships between both in various ecosystems [2,3]. Endophytic relationships between fungi and plants are being widely studied to elucidate their interactions with the hosts, the type of relationship they establish and the potential effects of that interaction. Dark septate endophytes (DSEs) (Figure 1) are conidial or sterile septate fungal endophytes, usually isolated from healthy plants, that form melanised structures including inter- and intracellular hyphae and microsclerotia in the roots. They show low host specificity and cover a wide geographical range [4]. The presence of DSEs in environments with strong abiotic stress caused by drought [5,6,7] high salinity [8,9] or the presence of heavy metals [10,11,12,13,14,15], among others, is crucial to ensure plant survival [16]. This type of fungi is less affected by long periods of drought and its colonisation rate is not reduced. DSEs do not bring as many benefits as mycorrhizal fungi [17], though they show a higher colonisation rate under abiotic stress conditions. Despite this, DSE colonisation in plants decreases when high temperature and drought phenomena occur simultaneously but does not lead to diversity loss [2]. Most studies on the interaction between DSEs and plants have been conducted in forest systems [2,18,19,20,21,22].
However, DSEs association in agriculture is becoming more relevant in the scientific sphere, particularly due to their increased tolerance under stress conditions [11], although studies on horticultural or extensive farming continue to be scarce.
For example, Andrade-Linares et al., [23] obtained a higher yield and quality in tomato fruits after applying two DSEs, and Fakhro et al., [24] obtained higher morphological parameter numbers in tomato plants. While Osuna-Avila and Barrow [25] described the increase in number and length of Daucus carota roots through the application of Aspergillus ustus. However, from a commercial standpoint, a DSE-microsclerotia-based product is not available in the market. These and other examples will be addressed throughout this review. In recent years, a patent of Rutstroemia calopus has been described as a DSE capable of promoting and increasing plant growth and development in crops, even under hydric and saline stress conditions. This bio-stimulant effect is also visible when there is an 80% reduction in the fertilisation of cucumber plants. Moreover, studies have reported a 33.8% increase in the leaf area of plants after applying Rutstroemia calopus CG11560 when fertilisation is reduced. Likewise, total dry weight has a 30.43% reported increase when compared to standard fertilisation methods [26]. Most benefits associated with DSEs are related to plant bio-stimulation which improves morphological parameters [11], stimulates plant resistance to drought, increases secondary metabolism activity [27], improves water, nutrient, and carbon absorption [11], increases antioxidant enzyme activity, or helps develop adaptation strategies against heavy metals, among other functions [28,29].
Due to the properties of endophyte fungi, whose most significant benefits are obtained under abiotic stress conditions, irrigation doses and even the use of fertilizers may be reduced. While this may not ensure a production increase, it could result in less products being used for higher yield.
2. DSEs and Their Connection to Drought and Salinity Mitigation
Drought and salinity are stressful environmental factors that negatively affect plant growth by causing reactive oxygen damage [30], as well as having an impact on the hydric potential, nutrient absorption, enzyme activity, and photosynthetic pigment content. Among other mechanisms, different studies have found that DSE can increase plant resistance by up-regulating of antioxidant enzymes, especially superoxide dismutase (SOD) activity, an important protective enzyme against reactive oxygen species (ROS) [31], as well as improving the quality of plant exudates, polysaccharide production, increased production of glutathione, proline, soluble sugar, and a large amount of melanin under stress [32]. Plant growth, development and the ability to adapt to adversities are also inseparable from the involvement of hormones and all of these actions are related to one another (Figure 2). Thus, polysaccharides have a great capacity for retaining liquids (Figure 3) through the formation of biofilms [33]. Treatment with exopolysaccharide increases the levels of abscisic acid (ABA), promotes stomatal closure, and minimises water loss [34], by acting as antiperspirant agents [35]. Likewise, cellular stability takes place due to the accumulation of rigid and soluble osmolytes in water, which increases osmotic pressure and causes minimal water loss. Galactose-Rich Heteropolysaccharide (GRH) acts as an elicitor that activates the biochemical cascade necessary to maintain hydric balance and increase antioxidant defence [34,36]. Thus, the use of GRH in rice crops improved superoxide dismutase (SOD), peroxidase activity (POD) and catalase activity (CAT) levels, while reducing malondialdehyde (MDA) contents, which indicate the level of lipid oxidation in the membrane. A variation on the levels of proline and soluble sugar was also observed, which improved the number of stress-tolerant enzymes. This included plants, adapting to drought conditions due to the role of proline as an osmoregulatory agent [34]. Similarly, inoculation with DSEs Phialophora sp., Knufia sp., Leptosphaeria sp., and Embellisia chlamydospora in Hedysarum scoparium under hydric stress conditions altered the enzyme and antioxidant activity, and increased SOD and CAT concentration [5], which enabled N and P absorption. Other studies on crops such as cowpea, soybean and rice reported that DSE responses regarding biomass depended on saline concentration [37,38,39,40]. This property of DSEs may be due to their ability to produce hormones such as indoleacetic acid and gibberellins [41,42]. Thus, the application of Alternaria alternata, Paraphoma pye, and Paraphoma radicina on wheat crops increased auxin levels which could be associated with the ability of DSEs to synthetize auxins for water intake reduction under stress conditions and activate the expression of hormone-regulated genes [43] (Li et al. 2022). Higher accumulation of SOD has also been observed in wheat, rice [43,44] Li et al. 2022; Pang et al. 2020), and citric plants [45] (Sadeghi et al. 2020) inoculated with Penicillium citrinum, Aureobasidium pullulans, and Dothideomycetes sp., individually and combined, as well as lower MDA caused by an increase in glutathione and proline content, resulting in an increased tolerance to drought in plants. Zhang et al. [7] obtained similar results after inoculating sorgo with the DSE Exophiala pisciphila. In this case, an increase in the synthesis of metabolites related to secondary metabolism was also observed. This resulted in an improved hydric state due to stomata opening which improved their transpiration rate and stomatal conductance. Likewise, the application of Neocamarosporium phragmitis, Alternaria chlamydospora, and Microascus alveolaris on Lycium ruthenicum Murr crops under drought conditions increased glutathione content, SOD activity, and soluble protein and proline content. In addition, indole acetic acid (IAA) content in plant roots also increased after inoculation with N. phragmitis compared with the control group [46]. For the first time, Isatis indigotica under stress due to drought showed structures characteristic of DSEs Acrocalymma vagum, Paraphoma chlamydocopiosa, Edenia gomezpomplae, Darksidea alpha, Brunneochlamydosporium nepalense, and Preussia terrícola, and showed an increase in IAA, proline, and chlorophyll and epigoitrin content, which promotes plant growth by improving osmotic pressure and increasing plant resistance to stress [47].
Moreover, DSEs have an impact on photosynthetic activity when plants are under stress conditions. Thus, inoculation with endophyte fungi such as P. indica, T. virens or P. indica + T. virens increased the chlorophyll content and photosynthetic activity of Stevia [48]. This may be because DSEs are capable of decomposing photosynthates; DSEs allow photosynthetic feedback inhibition and improve the physiology of host plants by increasing chlorophyll concentration and transpiration rate in stressed soils [49,50]. Furthermore, DSEs are involved in improving the ability to capture excitation energy released by chloroplasts under drought conditions, resulting in an increased photosynthetic rate and improved leaf nutrition caused by an enhanced C assimilation [48]. Therefore, the use of DSEs Alternaria alternata, Paraphoma pye, and Paraphoma radicina on wheat and rice crops led to an increase in plant height, leaf growth, chlorophyll content, and photosynthetic rate of those plants, as well as a decrease in intercellular carbon dioxide, which alleviated the damage caused to photosynthetic processes by drought [43,44]. On the other hand, the inoculation of Ormosia hosiei with Acrocalymma vagum resulted in a damage-free root cell structure and an increase in the amount of chlorophyll and carotenoids produced [51,52]. The connection between root cells and the rhizosphere through melanised hyphae can be a strategy to survive in stressful environments and to protect plants from free radicals [53]. Likewise, an increased photosynthetic activity may be directly related to the increase in leaf area, since inoculated plants have a greater capacity to hold water and show reduced evaporation [45,54,55].
Inoculation with DSEs changes the rhizosphere microbiome according to the environmental conditions to which they are subject to, as microorganisms are also very specific when going through their life cycle. For this reason, inoculation with DSEs increases the population of beneficial and neutral fungi while reducing that of pathogen fungi under drought conditions, and even displacing other endophyte fungi in some cases [56,57]. Thus, inoculation of Lycium ruthenicum Murr with A. chlamydospore and M. alveolaris under drought conditions increased arbuscular mycorrhizal fungi, fungi, bacteria, and actinomycete contents in the rhizosphere soil. These results were in accordance with those reported by Li et al. [43] in Astragalus, where an increase in bacteria and beneficial fungi was observed in the rhizosphere after inoculating a combination of DSE and Trichoderma. Therefore, microbial richness in the rhizosphere is essential for proper plant survival. However, not all obtained results were about the benefits of DSEs. The effect of DSEs depends on the type of crops they are used on, their own genotypes and the stress conditions imposed on them [58]. These microorganisms may show negative, positive, or no effects. According to Do Santos et al. [58], rice crops showed beneficial or null results in the parameters measured in their study, and negative results for some grasses under hydric stress conditions. The explanation for this phenomenon is that in this study two crops were C3 plants while the other one was C4, and they all showed different adaptative mechanisms to survive on their own under hydric stress conditions.
3. Implication of DSEs in Fertilisation Reduction
Excessive use of chemical fertilizers to furnish plants with the necessary nutritional requirements and improve the productivity of crops has caused serious environmental issues. The low efficiency and prolonged use of fertilisation has contributed to making the problem worse [59]. In addition, the use of intensive agricultural techniques, which cause soil salinisation particularly in arid and semiarid zones [60], along with climate change, which has reduced the amount of available water [61], are contributing to the continuous degradation of soils by lowering their quality and physiochemical properties and hindering nutrient availability [62]. Therefore, there is a need for developing new techniques that help reduce the use of synthetic fertilizers and maximise the utilisation of soil nutrients, in a sustainable and ecofriendly manner [63], for example, through the use of soil microorganisms capable of solubilising nutrients in the soil. Previous studies have stated that fungi are more efficient than other soil microorganisms as they solubilise nutrients that form compounds [64,65]. It has been shown that, inside fungi, DSEs can degrade complex substrates, secrete extracellular enzymes such as phosphatases and cellulases, and promote nutrient absorption and use by plants by transforming them into bioavailable forms [66].
Phosphorus is a macronutrient essential to plant metabolism [67]. Most phosphorus present in the soil can be found in the form of phosphates which form compounds together with metals, minerals (inorganic substances), or organic materials (organic substances) [68]. Although phosphorus is found abundantly in agricultural soils, most of it is found in its insoluble form, so it cannot be used directly by plants [69]. As a consequence, in order to achieve an optimal crop yield, synthetic fertilizers with phosphorus are used, but only around 30% of the added phosphorus can be used by crops, since a big portion remains immobilised once again in the form of insoluble phosphate or migrates in the soil [70]. Moreover, phosphorus is associated with various negative environmental impacts such as eutrophication [71]. Several studies show DSEs’ capacity to solubilise various phosphorus forms (organic and inorganic forms). Around 50-80% of soil organic phosphorus can be found in the form of salts of oxalic acid, called phytates [72]. A study conducted by Mikheev et al. [73] showed that the Phialocephala fortinii isolate secreted phytases, which is a special group of phosphatases that catalyses phytate hydrolysis to release phosphorus [74]. Moreover, another study shows that P. fortinii was capable of promoting Asparagus officinalis growth by using phytic acid sodium salt as the only organic source of phosphorus, which confirmed the capacity to mineralise organic phosphorus [75]. Meanwhile, other DSEs such as Exophiala pisciphila, Periconia macrospinosa, and Cadophora sp. have failed to show an ability to secrete phytases [75,76]. Regarding inorganic forms composed mainly of aluminium, iron, and calcium phosphates, various DSEs have shown a capacity for solubilising phosphorus both in vitro and in soil, with different levels of solubilisation in each compound according to the fungus species used [77,78,79]. This process also involved phosphatase enzymes that transform insoluble phosphorus into soluble substances and include both acid and alkaline phosphatases [80].
Although their capacity for solubilising phosphorus appears to be a significant contribution, some cases of DSE strains have been found to accumulate polyphosphates [73,81]. This accumulation would allow the transport of phosphorus to plants through hyphae –this interaction has been reported in AMF fungi [82,83,84]. However, more studies are needed to find out whether this process occurs or if these polyphosphates are directly profited by DSEs for their own metabolism.
Nevertheless, the effect of DSEs on phosphorus absorption by plants is ambiguous [85]. Various studies have analysed the effect of inoculation with DSEs on plant phosphorus uptake, showing mixed results based on the DSE species and study plant.
In the case of Lycium ruthenicum plants, inoculation with DSEs had a significant effect on the phosphorus found in the soil and increased the plant’s absorption of this element under hydric deficit conditions [46]. Similarly, in pine seedlings, inoculations with DSEs contributed to the absorption of bigger amounts of phosphorus [86]. In addition, inoculation with P. fortinii of Vaccinium macrocarpon plants resulted in an increased phosphorus content compared to the plants that had not been inoculated. Specifically, there was a 28% increase in the aerial part of the plant and a 61% increase in the roots after 10 months of harvesting [73]. Similar results were observed in corn after the inoculation with E. pisciphila under phosphorus deficient and sufficient conditions as a result of the DSEs presence [67,87]. In other studies, though DSEs were capable of solubilising and mineralising phosphorus in the soil by increasing phosphorus reserves easily available, (Trifolium repens) phosphorus content was only modified when DSE and AMF were applied simultaneously [88]. In a recently conducted study using sorgo and three DSE species (A. alternata, Curvularia sp. and Ophiosphaerella sp.), all three species were capable of dissolving insoluble phosphates as in vitro reagents and in the soil. However, although inoculation with Curvularia sp. produced an increase in the aerial biomass and root length of the plant, there were no statistically significant differences in the phosphorus content of inoculated plants compared to the control, except in the case of A. alternata, which had a lower P content Therefore, greater soil phosphorus solubilisation did not result in an increased absorption by the plant. A similar result was obtained in tomato plants which showed that phosphorus content in plants inoculated with Cadophora sp. and Periconia macrospinosa did not show a statistically significant difference compared to control plants [76]. In both studies, the DSE microsclerotia was not detected in the plant roots. No endophytic structures were found in the sorgo (suggesting that fungi maintained their saprophytic form) and tomatoes only showed hyphae from Cadophora sp.
Rise has also shown that some DSE isolates were capable of increasing P content in the aerial part of the plant compared to controls, while others remained the same. It should be noted that these increases took place when plants had been supplemented with an inorganic phosphorus source, which suggests that the species used had the ability to improve the absorption of phosphorus from inorganic sources [89]. These results are also supported by another case where tomato plants inoculated with Cadophora sp. and Periconia macrospinosa showed increased growth stimulation when inorganic sources were used [76]. However, other studies suggested that DSEs are more efficient in releasing phosphorus coming from organic sources instead of that from inorganic sources [90].
Potassium is an indispensable element in plant nutrition. In addition to other functions, potassium plays a significant role in regulating stomatal opening and closure in leaf plants, and a potassium deficit can affect water-plant relations. Greater resistance to drought found in plants inoculated with DSEs could be related to a higher potassium content in plant leaves [91]. Under stress conditions due to water deficit, inoculation with two strains of Paraconiothyrium, Darksidea sp., Embellisia chlamydospore, and Leptosphaeria sp. showed a significant increase in the potassium content of Ammopiptanthus mongolicus roots compared to control plants. Despite the fact that they promoted potassium acquisition in the roots, no statistically significant difference was found in the quantity of potassium in the aerial part [92]. Meanwhile, in other research works conducted on rice, tomato and corn, inoculation with DSEs did show a significant increase in potassium content in plant leaves [87,93,94].
DSEs have the ability to increase soil organic nitrogen bioavailability for the host plant, as they have the ability to mineralise it [28]. They are capable of degrading nitrogenous compounds such as proteins, ribonucleic acids, amino acids, and urea [65,66]. According to a meta-analysis by Newsham [90], this capacity would represent the biggest contribution of these fungi to the stimulation and improvement of plant growth. Several authors have assessed DSEs’ capacity for enabling nitrogen absorption and the influence of the nitrogenous compound being provided. Heteroconium chaetospira is capable of promoting the growth of its natural host, the Chinese cabbage (Brassica rapa), under limiting nitrogenous conditions. While the fungus was not capable of promoting plant growth in the presence of nitrogen available for the plant, an increase of up to four times in biomass was observed when an organic nitrogen source was used [95]. This is supported by the results of the inoculation with P. macrospinosa which increased the root biomass and aerial part of tomato plants when organic nitrogen sources were used, which means that this fungus was capable of improving organic source nitrogen intake. However, this improvement did not happen when Cadophora sp., was used, which shows the influence of the DSE species used in the obtained results [76]. Another study conducted on tomatoes using an inorganic (ammonium sulphate) or organic nitrogen source (consistent in ground Canavalia ensiformis plants) showed that, overall, both inoculated and non-inoculated tomatoes obtained nitrogen more efficiently from the inorganic source than from the organic one. However, the effects of inoculation with DSEs on nitrogen recovery was evident and significant only when the tomato plants were fertilised with C. ensiformis, as nitrogen accumulation in plants was higher with this organic form (around 24-33% based on the DSE used [94]. Similar results were obtained for rice, where plants associated with the DSE C. ensiformis exhibited a more efficient nitrogen intake, and nitrogen content increased in both the aerial part and the grains of rice, resulting in an increased accumulation of proteins and dry matter [96]. There are more examples of improved nitrogen absorption such as the case of P. fortinii, which is a DSE that promotes Asparagus officinalis growth through nitrogen mineralisation and increases N content in the host plant [90]. Nitrogen transfer between DSE fungi and the plant seems to be accompanied by an exchange of carbon from the plant. Proof of this exchange was found when tracing carbon and nitrogen in the association between H. chaetospira and B. rapa, as well as in tomato plants and Pleosporales and Calosferiales DSE [94,95].
Although nitrogen mineralisation appears to be the main method by which DSEs promote nitrogen absorption, other mechanisms that could also be helpful have been suggested. For example, DSEs in rice plants have shown higher efficiency for nitrogen intake and accumulation, resulting in a 33-47% nitrogen content increase in the plant and an increased enzyme activity of the plasma membrane proton pump H+-ATPase [97]. This protein hydrolyses ATP in the cytosol to release protons to the cell exterior. Through the plasma membrane, hydrolysis establishes an electrochemical gradient of protons necessary to regulate various physiological processes in the plant, one of them being the absorption of nutrients such as nitrogen [98]. In addition, there was an increase in the vacuolar pyrophosphatase and transcription of OsA5 and OsA genes corresponding to H+-ATPase isoforms. The authors suggested that this stimulation could be mediated by metabolites secreted by DSEs [97]. In a different study that inoculated rice seeds with DSE pleosporales, NO3 nitrogen absorption showed lower Michaelis-Menten constant (Km) values showcasing an increase in affinity which was reflected in a higher nitrogen content in the aerial part of the plant [96]. Various studies have shown that the net formed by DSE hyphae plays a significant role in water and nutrient exchange between plants and soil under stress conditions but is also dependent on the fungi species [99,100]. For example, in a study on liquorice plants (Glycyrrhiza uralensis), inoculation with DSEs increased the content of nitrogen easily available in the soil. This improvement in nitrogen availability depended on the DSE species used for inoculation and in the plant irrigation system; so, while Acrocalymma vagum, Paraphoma chrysanthemicola, Alternaria longissima, Darksidea alpha, Preussia terrícola and Alternaria chartarum showed an increase of this element when properly irrigated, under stress conditions caused by a hydric deficit, such increase was only observed with Alternaria chlamydospora and Acremonium nepalense species [101]. Similar results were observed in the case of Lycium ruthenicum [46], were the plant inoculated with DSE showed increased absorption of the nitrogen available in the soil similar to Triticum aestivum L whose association with a strain of Alternaria alternata led to an increased accumulation of nitrogen and carbon under stress due to drought [102].
DSEs can also enable the incorporation of other soil micronutrients [103]. In the case of iron, when inoculation of tomato seedlings or rice with these fungi, resulted in an increase of 72-128% in iron content [94,96]. This increased iron intake may be related to the ability of endophytic microorganisms to produce siderophores under limiting iron stress conditions [104,105,106]. In addition, these siderophores produced by endophytes tend to have more affinity for Fe3+ ions than phytosiderophores produced by plants [69]. P. fortinii is capable of synthetising hydroxamate siderophore and such synthesis has been associated to an increased incorporation of Fe3+ by the host plant [103]. The presence of these compounds has also been confirmed in UPLC-MS analyses conducted on other DSE isolates [107]. Regarding other micronutrients, increases in manganese and zinc content have been documented in DSE-inoculated tomato and rice plants, respectively [104,105,106].
Extracellular metabolites produced by DSEs can be incorporated by the plant directly as nutrients or by acting as chemical signals regulating the nutrient absorption process of plants. The use of Alternaria sp. metabolites in corn plants has been associated with a significant increase in nitrogen content in the aerial part [108]. On the other hand, a different study conducted by Wang et al. [109] on alfalfa (Medicago sativa), using the same DSE species, showed no statistically significant differences in the absorption efficiency of this element between treated and untreated plants, although there was a significant improvement in nitrogen translocation efficiency. A significant increase and higher efficiency in phosphorus intake were observed in both plant species, and the alfalfa plants showed a significant increase in the absorption and translocation efficiency of potassium. The results on nutrition improvement changed based on the culture time of microorganisms prior to metabolite extraction, with emphasis on the importance of this factor.
4. Compatibility of DSEs with Other Microorganisms
Interactions between plants and microorganisms are quite complex due to the presence of a mutual regulation, and because many factors causing their union are still unknown. Overall, studies have described that the use of DSEs modifies the soil microbial community associated with the plant [57,86,110,111,112,113]. Thus, incorporating three DSEs (Alternaria chlamydosporigena, Paraphoma chrysanthemicola, and Bipolaris sorokiniana) promoted the presence of gram-positive bacteria, gram-negative bacteria or the abundance of AM fungi in the rhizosphere of Artemisia ordosica according to the DSE type and saline concentrations applied [8]. Studies have described the natural coexistence of DSEs with other endophyte fungi and arbuscular mycorrhizal fungi in the roots of a plant [114,115,116,117,118]. Despite this, the effect of DSE use on different hosts and on rhizosphere microbiota remains unclear due to the variability observed in the study results. This can become more complicated if we add new microorganisms that are used in agriculture as potential biological control agents or bio-stimulants. Managing these microorganisms in both extensive and intensive farming could help reduce the use of agrochemicals and fertilizers [4,119]. However, more research is required to determine the behaviour of each one of these DSEs in various “agricultural ecosystems.”
Similarly, research has been conducted where the use of endophyte fungi such as mycorrhizal fungi, DSEs or various species of Trichoderma plays a significant role in the development of plants even under abiotic or biotic stress conditions [56]. In that sense, and given the current climate situation where droughts are more frequent every time and temperature is increasing, evapotransport of crops is greater, the use of microorganisms improving the intake of water available can be a solution in the face of the current climate issues, where the periods of drought are becoming more frequent and the increase in temperature is causing higher crop evapotranspiration, the use of microorganisms to improve the intake of available water can be a solution, although long period of droughts and high temperatures are also known to affect the microbiota associated with the soil and roots [120]. The soils in deserts or arid areas, where DSEs associated with plants are abundant, are a clear example of this [29]. DSE management in agriculture is not as developed as in the case of Trichoderma, but its possible use in combination with other types of microorganisms can bring multiple benefits. Co-inoculation with DSEs, Acrocalymma vagum or Paraboeremia putaminum and Trichoderma viride improved the growth of Astragalus mongholicus and caused changes in the rhizosphere microbiome according to the combination of inoculants under hydric stress conditions [56,121]. The same results were obtained by Li et al. [122] who detected an increase in plant nutrients, root activity, phosphorus availability, and phosphatase activity in the soil co-inoculated with Trichoderma koningiopsis and Amesia nigricolor, as well as changes in bacterial and fungal diversity in the soil. Co-inoculation with the genus Darksidea and the arbuscular mycorrhizal fungus (HMA) Rhizophagus irregularis of two plant species, Artemisa tridentata and native grass Poa secunda, increased the union of HMA and the formation of vesicles, but bio-stimulation did not take place when the plants were not under abiotic stress [123]. Co-inoculation with AMF Funneliformis mosseae and DSE Exophiala pisciphila improved corn yield under high Cd stress, and also reduced cadmium transfer from the roots to the stem significantly, which was closely related to changes in the photosynthesis physiology and corn roots [113]. On the other hand, an interaction between both fungi was observed, which can have a negative impact on the colonisation rate. Thus, Deram et al. [124] detected that colonisation by septate fungi was reduced in the presence of mycorrhizal fungi in non-polluted soils, but increased in polluted soils, partially due to a reduction in mycorrhisation which was impacted by the presence of heavy metals. Meanwhile, Li et al. [122] observed that co-inoculation with Trichoderma koningiopsis and DSE Amesia nigricolor reduced the DSE colonisation rate. Similarly, the level of mycorrhisation was not affected by Phialocephala fortinii s.l.-Acephala applanata species complex PAC colonisation, but the DSE complex colonisation was reduced. This negative effect on DSE colonisation has been evidenced by other authors [125,126,127]. However, positive effects showing an increase have also been detected [88,128,129], or neutral effects in which the colonisation rates of both co-inoculants have not been affected [88,130,131,132,133]. Table 1 lists various studies on the compatibility between DSEs and other microorganisms.
These effects are not only observed among fungal species, but also in bacteria. There is data that some bacteria from the Rhizobium/Agrobacterium group are endophytes of some DSE species [128,147,148]. Thus, the combined use of Agrobacterium pusense isolated from DSE Veronaeopsis simplex in tomatoes resulted in an increased number of roots colonised by the fungus. The same finding was observed by Silva et al. [135] who described that root colonisation by AMF and DSE can increase by applying Azospirillum brasilense. Wu et al. [149] found a high colonisation of mycorrhizal fungi with DSE in C. korshinskii roots, which showed the existence of symbiotic relations between them and even Rhizobium under desert conditions.
Different factors have an impact on the colonisation rates of various microorganisms. Huo et al. [136] showed that DSE and AM colonisation had different responses to the climate conditions and soil types in different geographical areas, where DSEs were more dependent on abiotic stress conditions. Lugo et al. [138] found synergy between both AM and DSE, but with a different response based on the altitude and environmental factors, as the AM colonisation rate decreased and that of DSEs did not. On the other hand, Ruotsalainen et al. [17] showed that AM and DSE colonisation depends on the season of the year, and that there is a certain degree of specificity imposed by the host. Such dependency from the host was also observed by Fernando and Currah [142] who also described a dependency on the harvesting conditions. The inoculant dose is another factor that needs to be considered regarding the root colonisation rate, as well as the potential beneficial effect on the plant. Xie et al. [150] observed that the higher the DSE Alternaria sp. inoculation dose paired with a fixed dose of AMF Diversispora epigaea, the lower the DSE colonisation, showing a competition for space between both fungal species. A bio-stimulant effect took place in corn plants under the same conditions [149].
5. Compatibility of DSEs with Active Chemical Substances
Even though they are indispensable for a comprehensive management in disease control and crop bio-stimulation, studies on the compatibility of agrochemical management with DSEs are scarce. Compatibility not only depends on microorganisms and on the chemical active substance, but also on other factors such as the amount of organic matter in the soil and the microbial community size [151]. DSEs have an increased capacity for counteracting the negative effects of heavy metal in plants due to a high antioxidant enzyme activity [4]. Furthermore, they show increased resistance to compounds such as tannins, possibly due to a physical mechanism and the production of tannase enzymes and polyphenol oxidase [152]. However, little is known about their resistance capacity against agrochemicals.
Repeated applications of herbicides are particularly harmful to soil health and microbe-plant associations. Various studies show how glyphosate produces a fast decrease in the DSE colonisation percentage in different plant species that depends on the dose and number of applications [153]. Spagnoletti and Chiocchio [154] show that DSEs Alternaria alternata and Cochliobolus sp. are tolerant to glyphosate, carbendazim, and cypermethrin, which were assessed with the recommended agronomic doses.
Mancozeb is another widely used fungicide. Manalu et al. [155] isolated DES obtained from chili pepper grown in an environment with pesticides and detected two isolates resistant to 400 ppm. In addition, the presence of Macozeb in the medium resulted in changes in the mycelium colour. This change in colour has also been described by Wyss et al. [156], who also detected a change in the shape of Phomopsis amaranthicola hyphae and conidia, in the presence of herbicides from the family of imidazolines. Widyaningsih and Triasih [157] conducted an in vitro test on the resistance of three DSEs to Propineb 70%, Mancozeb 80%, and glyphosate. All DSEs were sensitive to the different tested doses, showing a decrease in sporulation as the dose of both fungicides increased. In the case of glyphosate, they were tolerant and only one isolate had a decreased sporulation capacity. None of the cases showed changes in the mycelium shape, though a decrease in density was observed in the Petri dishes. On the other hand, Tricyclazole inhibits E. pisciphila growth by inhibiting ergosterol and melanin biosynthesis, though no toxicity in low concentrations was observed [158].
Seed treatment with systemic fungicides for controlling seed-transmitted diseases is a common practice before harvesting. However, this treatment also affects nontarget organisms such as endophytes [159], as it compromises germination and early growth of seedlings [160]. That is the case of non-pathogenic Alternaria found in seeds.
6. Conclusions
DSEs have shown a capacity for mitigating and reducing the harmful effects of climate change in agriculture, such as salinity, drought, and reduced nutrient availability in the soil. Therefore, their use is set to become a good choice for crop management in the face of the current environmental challenges, as well as an alternative that is compatible with a more sustainable agriculture. However, many obstacles need to be overcome before implementing DSEs in the field. More research is needed to shed light on the mechanisms involved and the interactions taking place between the plant and other microorganisms present in the soil, and the compatibility with active chemical substances that are frequently used. On the other hand, conducting studies is essential to allow the development of DSE formulates at an industrial scale to be used in agriculture in the future. While a lot of research is still required, interest in this field has continued to grow in recent years and the use of microorganisms in agriculture is advancing by leaps and bounds.
Author Contributions
Conceptualization, M.S. and F.D.; methodology, V.H. and A.J. software, F.D. and V.H.; validation, M.S.; writing—original draft preparation, M.S., V.H., A.J., R.C, F.D and writing—review and editing, R.C. and F.D.; supervision, M.S. and F.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The present work benefited from the input of the project TRFE-I-2022/011 and was supported by University of Almeria- Entonova S.L.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mukherjee, S.; Mishra, A.; Trenberth, K.E. Climate change and drought: A perspective on drought indices. Curr. Clim. Change Rep. 2018, 4, 2. [Google Scholar] [CrossRef]
- Gehring, C.; Sevanto, S.; Patterson, A.; Ulrich, D. E. M.; Kuske, C.R. Ectomycorrhizal and dark septate fungal associations of pinyon pine are differentially affected by experimental drought and warming. Front. Plant Sci. 2020, 11, 582574. [Google Scholar] [CrossRef]
- Perez-Ramos, I.; Álvarez-Méndez, A.; Wald, K.; Matías, L.; Hidalgo-Galvez, M. D.; Fernández, C.M. Direct and indirect effects of global change on mycorrhizal associations of savanna plant communities. Oikos 2021, 130, 1370–1384. [Google Scholar] [CrossRef]
- Santos, M.; Cesanelli, I.; Diánez, F.; Sánchez-Montesinos, B.; Moreno-Gavira, A. Advances in the role of dark septate endophytes in the plant resistance to abiotic and biotic stresses. J. Fungi 2021, 7, 939. [Google Scholar] [CrossRef]
- Li, X.; He, X.L.; Zhou, Y.; Hou, Y.T.; Zuo, Y.L. Effects of dark septate endophytes on the performance of Hedysarum scoparium under water deficit stress. Front. Plant Sci. 2019, 10, 903. [Google Scholar] [CrossRef]
- Landolt, M.; Stroheker, S.; Queloz, V.; Gall, A.; Sieber, T.N. Does water availability influence the abundance of species of the Phialocephala fortinii s.l.-Acephala applanata complex (PAC) in roots of pubescent oak (Quercus pubescens) and Scots pine (Pinus sylvestris)? Fungal Ecol. 2020, 44, 100904. [Google Scholar] [CrossRef]
- Zhang, Q.; Gong, M.; Yuan, J.; Hou, Y.; Zhang, H.; Wang, Y.; Hou, X. Dark septate endophyte improves drought tolerance in sorghum. Int. J. Agric. Biol. 2017, 19, 53–60. [Google Scholar] [CrossRef]
- Hou, L.; Li, X.; He, X. Effect of dark septate endophytes on plant performance of Artemisia ordosica and associated soil microbial functional group abundance under salt stress. Appl. Soil Ecol. 2021, 165, 103998. [Google Scholar] [CrossRef]
- Gonçalves, D.R.; Pena, R.; Zotz, G.M.; Albach, D.C. Effects of fungal inoculation on the growth of Salicornia (Amaranthaceae) under different salinity conditions. Symbiosis 2021, 84, 195–208. [Google Scholar] [CrossRef]
- Portman, T.A.; Granath, A.; Mann, M.A.; El Hayek, E.; Herzer, K.; Cerrato, J. M.; Rudgers, J. A. Characterization of root-associated fungi and reduced plant growth in soils from a New Mexico uranium mine. Mycologia 2023, 115, 165–177. [Google Scholar] [CrossRef]
- Wang, L.; Li, Z.; Zhang, G.; He, Y.; Zhan, F. Dark septate endophyte Exophiala pisciphila promotes maize growth and alleviates cadmium toxicity. Front. Microbiol. 2023, 4, 1165131. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.; Tan, J.; Xiong, Y.; (...), Ding, Y.; Xu, Z. The responses and detoxification mechanisms of dark septate endophytes (DSE), Exophiala salmonis, to CuO nanoparticles. Environ. Sci. Poll. Res. 2023, 30, 13773–13787. [Google Scholar] [CrossRef]
- Ban, Y.; Xu, Z.; Yang, Y.; Zhang, H.; Chen, H.; Tang, M. Effect of dark septate endophytic fungus Gaeumannomyces cylindrosporus on plant growth, photosynthesis and Pb tolerance of maize (Zea mays L.). Pedosphere 2017, 27, 283–292. [Google Scholar] [CrossRef]
- Guzmán-Cornejo, L.; Pacheco, L.; Camargo-Ricalde, S.L.; González-Chávez, M.D.C. Endorhizal fungal symbiosis in lycophytes and metal(loid)-accumulating ferns growing naturally in mine wastes in Mexico. Int. J. Phytoremediation 2023, 25, 538–549. [Google Scholar] [CrossRef]
- Zhu, G.S.; Yu, Z.N.; Gui, Y.; Liu, Z.Y. A novel technique for isolating orchid mycorrhizal fungi. Fungal Div. 2008, 33, 123–137. [Google Scholar]
- Knapp, D.G.; Kovács, G.M.; Zajta, E.; Groenewald, J.Z.; Crous, P.W. Dark septate endophytic pleosporalean genera from semiarid areas. Persoonia 2015, 35, 87–100. [Google Scholar] [CrossRef]
- Ruotsalainen, A.L.; Kauppinen, M.; Wäli, P.R.; Saikkonen, K.; Helander, M.; Tuomi, J. Dark septate endophytes: Mutualism from by-products? Trends Plant Sci. 2022, 27, 247–254. [Google Scholar] [CrossRef]
- Dhyani, A.; Jain, R.; Pandey, A. Contribution of root-associated microbial communities on soil quality of Oak and Pine forests in the Himalayan ecosystem. Trop. Ecol. 2019, 60, 271–280. [Google Scholar] [CrossRef]
- Chu, H.; Wang, H.; Zhang, Y.; Li, Z.; Wang, C.; Dai, D.; Tang, M. Inoculation with ectomycorrhizal fungi and dark septate endophytes contributes to the resistance of Pinus spp. in to pine wilt disease. Front. Microbiol. 2021, 12, 687304. [Google Scholar] [CrossRef]
- Chu, H; Wang, C; Li, Z.; Wang, H.; Xiao, Y.; Chen, J.; Tang, M. The dark septate endophytes and ectomycorrhizal fungi effect on Pinus tabulaeformis Carr. seedling growth and their potential effects to pine wilt disease resistance. Forests, 2019, 10, 140. [Google Scholar] [CrossRef]
- Lukešová, T.; Kohout, P.; Větrovský, T.; Vohník, M. The potential of dark septate endophytes to form root symbioses with ectomycorrhizal and ericoid mycorrhizal middle european forest plants. PLoS ONE 2015, 10, e0124752. [Google Scholar] [CrossRef]
- Diene, O.; Wang, W.; Narisawa, K. Pseudosigmoidea ibarakiensis sp. nov., a dark septate endophytic fungus from a cedar forest in Ibaraki, Japan. Microbes Environ. 2013, 28, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Linares, D.R.; Grosch, R.; Restrepo, S.; Krumbein, A.; Franken, P. Effects of dark septate endophytes on tomato plant performance. Mycorrhiza 2011, 21, 413–422. [Google Scholar] [CrossRef]
- Fakhro, A.; Andrade-Linares, D.R.; von Bargen, S.; Bandte, M.; Buttner, C.; Grosch, R.; Schwarz, D.; Franken, P. Impact of Piriformospora indica on tomato growth and on interaction with fungal and viral pathogens. Mycorrhiza 2010, 20, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Osuna-Avila, P.; Barrow, J. In vitro system to determine the role of Aspergillus ustus on Daucus carota Roots. Terra Latinoamericana 2009, 27, 363–372. [Google Scholar]
- Santos, M.; Diánez, F. Strain of Rutstroemia calopus, compositions and uses. 2017. Patent ES2907599. 7599.
- Zhang, M.; Yang, M.; Shi, Z.; Gao, J.; Wang, X. Biodiversity and variations of arbuscular mycorrhizal fungi associated with roots along elevations in Mt. Taibai of China. Diversity 2022, 14, 626. [Google Scholar] [CrossRef]
- Malicka, M.; Magurno, F.; Piotrowska-Seget, Z. Plant association with dark septate endophytes: When the going gets tough (and stressful), the tough fungi get going. Chemosphere 2022, 302, 134830. [Google Scholar] [CrossRef] [PubMed]
- Madouh, T.A.; Quoreshi, A.M. The Function of Arbuscular Mycorrhizal Fungi Associated with Drought Stress Resistance in Native Plants of Arid Desert Ecosystems: A Review. Diversity 2023, 15, 391. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Zandi, P.; Schnug, E. Reactive oxygen species, antioxidant responses and implications from a microbial modulation perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Muley, A.B.; Shingote, P.R.; Patil, A.P.; Dalvi, S.G.; Suprasanna, P. Gamma radiation degradation of chitosan for application in growth promotion and induction of stress tolerance in potato (Solanum tuberosum L). Carbohydr Polym 2019, 210, 289–301. [Google Scholar] [CrossRef]
- Santra, H.K.; Banerjee, D. Drought alleviation efficacy of a galactose rich polysaccharide isolated from endophytic Mucor sp. HELF2: A case study on rice plant. Front. Microbiol. 2023, 13, 1–15. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- He, Z.; Cui, K.; Wang, R. Multi-omics joint analysis reveals how Streptomyces albidoflavus OsiLf-2 assists Camellia oleifera to resist drought stress and improve fruit quality. Front Microbiol 2023, 14, 1–12. [Google Scholar] [CrossRef]
- Farias, G.C.; Nunes, K.G.; Soares, M.A. Dark septate endophytic fungi mitigate the effects of salt stress on cowpea plants. Brazilian. Brazilian. J. Microbiol. 2020, 51, 243–253. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kim, Y.H.; Kang, S.M.; Lee, J.H.; Lee, I.J. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Process Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Hamayun, M.; Hussain, A.; Khan, S.A. Gibberellins producing endophytic fungus Porostereum spadiceum AGH786 rescues growth of salt affected soybean. Front Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Khan, A.; Hamayun, M.; Kang, S.M. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 1–14. [Google Scholar] [CrossRef]
- Barrow, J.R. Atypical morphology of dark septate fungal root endophytes of Bouteloua in arid southwestern USA rangelands. Mycorrhiza 2003, 13, 239–247. [Google Scholar] [CrossRef]
- Sadok, W.; Schoppach, R. Potential involvement of root auxins in drought tolerance by modulating nocturnal and daytime water use in wheat. Ann. Bot. 2019, 124, 969–978. [Google Scholar] [CrossRef]
- Li, M.; Ren, Y.; He, C.; Yao, J.; Wei, M.; He., X. Complementary effects of dark septate endophytes and Trichoderma strains on growth and active ingredient accumulation of Astragalus mongholicus under drought stress. J. Fungi 2022, 8, 920. [Google Scholar] [CrossRef]
- Pang, Z.; Zhao, Y.; Xu, P.; Yu, D. Microbial diversity of upland rice roots and their influence on rice growth and drought tolerance. Microorganisms 2020, 8, 1–18. [Google Scholar] [CrossRef]
- Sadeghi, F.; Samsampour, D.; Askari Seyahooei, M.; Bagheri, A.; Soltani, J. Fungal endophytes alleviate drought-induced oxidative stress in mandarin (Citrus reticulata L.): Toward regulating the ascorbate–glutathione cycle. Sci. Hortic. 2020, 261, 108991. [Google Scholar] [CrossRef]
- He, C.; Han, T.; Tan, L.; Li, X. Effects of dark septate endophytes on the performance and soil microbia of Lycium ruthenicum under drought stress. Front. Plant Sci. 2022, 13, 898378. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hou, L.; Liu, J. Growth-promoting effects of dark septate endophytes on the non-mycorrhizal plant Isatis indigotica under different water conditions. Symbiosis 2021, 85, 291–303. [Google Scholar] [CrossRef]
- Bahari Saravi, H.; Gholami, A.; Pirdashti, H.; Baradaran Firouzabadi, M.; Asghari, H.; Yaghoubian, Y. Improvement of salt tolerance in Stevia rebaudiana by co-application of endophytic fungi and exogenous spermidine. Ind. Crops Prod. 2022, 177, 114–443. [Google Scholar] [CrossRef]
- Wu, F.L.; Li, Y.; Tian, W. A novel dark septate fungal endophyte positively affected blueberry growth and changed the expression of plant genes involved in phytohormone and flavonoid biosynthesis. Tree Physiol. 2021, 40, 1080–1094. [Google Scholar] [CrossRef]
- Likar, M.; Regvar, M. Isolates of dark septate endophytes reduce metal uptake and improve physiology of Salix caprea L. Plant Soil 2013, 370, 593–604. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, X. Dark septate endophyte improves the drought-stress resistance of Ormosia hosiei seedlings by altering leaf morphology and photosynthetic characteristics. Plant Ecol. 2021, 222, 761–771. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, X. Dark septate endophyte improves drought tolerance of Ormosia hosiei Hemsley & E.H. Wilson by modulating root morphology, ultrastructure, and the ratio of root hormones. Forests 2019, 10, 830. [Google Scholar]
- Zhan, F.; He, Y.; Zu, Y.; Li, T.; Zhao, Z. Characterization of melanin isolated from a dark septate endophyte (DSE), Exophiala pisciphila. World J. Microbiol. Biotechnol. 2011, 27, 2483–2489. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M.; Zhang, Y. The applicability of empirical vegetation indices for determining leaf chlorophyll content over different leaf and canopy structures. Ecol. Complex. 2014, 17, 119–130. [Google Scholar] [CrossRef]
- Ogburn, R.M.; Edwards, E.J. Quantifying succulence: A rapid, physiologically meaningful metric of plant water storage. Plant Cell Environ. 2012, 35, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Liu, C.; Liu, H.; Wang, W.; Hou, J.; Li, X. Dual inoculation of dark septate endophytes and Trichoderma viride drives plant performance and rhizosphere microbiome adaptations of Astragalus mongholicus to drought. Environ. Microbiol. 2022, 24, 324–340. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, T.; Vediyappan, S. Comparison of arbuscular mycorrhizal and dark septate endophyte fungal associations in soils irrigated with pulp and paper mill effluent and well-water. Eur. J. Soil Biol. 2010, 46, 157–167. [Google Scholar] [CrossRef]
- Do Santos, S.G.; Silva, P.R.A.; Garcia, A.C.; Zilli, J.É.; Berbara, R.L.L. Dark septate endophyte decreases stress on rice plants. Brazilian J. Microbiol. 2017, 48, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, D.K.; Dheeman, S. Endophytes in mineral nutrient management: Introduction in endophytes: Mineral nutrient management: Introduction. In Endophytes in mineral nutrient management: Introduction in endophytes: Mineral nutrient management. Maheshwari, D. K., Dheeman, S., Eds.; Springer International Publishing 2021, 3, 3-9.
- Khan, N.M.; Rastoskuev, V.V.; Sato, Y.; Shiozawa, S. Assessment of hydrosaline land degradation by using a simple approach of remote sensing indicators. Agric. Water Manag. 2005, 77, 96–109. [Google Scholar] [CrossRef]
- García-Latorre, C.; Rodrigo, S.; Santamaría, O. Endophytes as plant nutrient uptake-promoter in plants. In Endophytes: Mineral Nutrient Management, Maheshwari, D. K., Dheeman, S., Eds.; Springer International Publishing (Springer, Cham.) 2021, 3, 247–265. [Google Scholar]
- Kumar, S.; Diksha; Sindhu, S.S.; Kumar, R. Biofertilizers: An ecofriendly technology for nutrient recycling and environmental sustainability. Curr. Res. Microb. Sci. 2022, 3, 100094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cook, J.; Nearing, J.T.; Zhang, J.; Raudonis, R.; Glick, B.R.; Langille, M.G.I; Cheng, Z. Harnessing the plant microbiome to promote the growth of agricultural crops. Microbiol. Res. 2021, 245, 126690. [Google Scholar] [CrossRef]
- Pradhan, N.; Sukla, L. Solubilization of inorganic phosphates by fungi isolated from agriculture soil. African J. Biotechnol. 2006, 5, 850–854. [Google Scholar]
- Gupta, N.; Sabat, J.; Parida, R.; Kerkatta, D. Solubilization of tricalcium phosphate and rock phosphate by microbes isolated from chromite, iron and manganese mines. Acta Bot. Croat. 2007, 66, 197–204. [Google Scholar]
- Caldwell, B.A.; Jumpponen, A.; Trappe, J.M. Utilization of major detrital substrates by dark-septate, root endophytes. Mycologia 2000, 92, 230–232. [Google Scholar] [CrossRef]
- Xu, R.; Li, T.; Shen, M.; Yang, Z.L.; Zhao, Z.W. Evidence for a dark septate endophyte (Exophiala Pisciphila, H93) enhancing phosphorus absorption by maize seedlings. Plant Soil 2020, 452, 249–266. [Google Scholar] [CrossRef]
- Byregowda, R.; Prasad, S.R.; Oelmüller, R.; Nataraja, K.N.; Kumar, M.K. Is endophytic colonization of host plants a method of alleviating drought stress? Conceptualizing the hidden world of endophytes. Int. J. Mol. Sci. 2022, 23, 9194. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.H; Abd El-Megeed, F.H.; Hassanein, N.M.; Youseif, S.H.; Farag, P.F.; Saleh, S.A.; Abdel-Wahab, B.A.; Alsuhaibani, A.M.; Helmy, Y.A.; Abdel-Azeem, A.M. Native rhizospheric and endophytic fungi as sustainable sources of plant growth promoting traits to improve wheat growth under low nitrogen input. J. Fungi 2022, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- López-Arredondo, D.L.; Leyva-González, M.A.; González-Morales, S.I.; López-Bucio, J; Herrera-Estrella, L. Phosphate nutrition : Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.; Sharpley, A.; Pierzynski, G.; Mcdowell, R. Chemistry, cycling, and potential movement of inorganic phosphorus in soils. agriculture and the environment sims, J.T; Sharpley, A.N., Ed.; American Society of Agronomy: Madison, usa, 2005; pp. 53–86. [Google Scholar]
- Liu, X.; Han, R.; Cao, Y.; Turner, B. Enhancing phytate availability in soils and phytate-p acquisition by plants: A review. Environ. Sci. Technol. 2022, 56, 9196–9219. [Google Scholar] [CrossRef] [PubMed]
- Mikheev, V.S.; Struchkova, I.V.; Ageyeva, M.N.; Brilkina, A.A.; Berezina, E.V. The role of Phialocephala fortinii in improving plants’ phosphorus nutrition: New puzzle pieces. J. Fungi 2022, 8, 1225. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Boukhris, I.; Pragya; Kumar, V.; Yadav, A.N.; Farhat-Khemakhem, A.; Kumar, A.; Singh, D.; Blibech, M.; Chouayekh, H.; Alghamdi, O. Contribution of microbial phytases to the improvement of plant growth and nutrition: A review. Pedosphere 2020, 30, 295–313. [Google Scholar] [CrossRef]
- Surono, S.; Narisawa, K. The dark septate endophytic fungus Phialocephala fortinii is a potential decomposer of soil organic compounds and a promoter of Asparagus officinalis growth. Fungal Ecol. 2017, 28, 1–10. [Google Scholar] [CrossRef]
- Yakti, W.; Kovács, G.M.; Vági, P.; Franken, P. Impact of dark septate endophytes on tomato growth and nutrient uptake. Plant Ecol. Divers. 2018, 11, 637–648. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Tobar, N.E.; Fernández Di Pardo, A.; Chiocchio, V.M.; Lavado, R.S. Dark septate endophytes present different potential to solubilize calcium, iron and aluminum phosphates. Appl. Soil Ecol. 2017, 111, 25–32. [Google Scholar] [CrossRef]
- Barresi, O.; Lavado, R.S.; Chiocchio, V.M. Can dark septate endophytic fungi (DSE) mobilize selectively inorganic soil phosphorus thereby promoting sorghum growth? A preliminary study. Rev. Argent. Microbiol. 2022, 54, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Priyadharsini, P.; Muthukumar, T. The root endophytic fungus Curvularia geniculata from Parthenium hysterophorus roots improves plant growth through phosphate solubilization and phytohormone production. Fungal Ecol. 2017, 27, 69–77. [Google Scholar] [CrossRef]
- Jha, B.; Pragash, G.; Cletus, J.; Gurusamy, R.; Sakthivel, N. Simultaneous phosphate solubilization potential and antifungal activity of new fluorescent pseudomonad strains, Pseudomonas aeruginosa, P. plecoglossicida and P. mosselii. World J. Microbiol. Biotechnol. 2009, 25, 573–581. [Google Scholar] [CrossRef]
- Saito, K.; Kuga, Y.; Saito, M.; Peterson, R. Vacuolar localization of phosphorus in hyphae of Phialocephala fortinii, a dark septate fungal root endophyte. Can. J. Microbiol. 2006, 5, 643–650. [Google Scholar] [CrossRef]
- Nguyen, C.; Saito, K. Role of cell wall polyphosphates in phosphorus transfer at the arbuscular interface in mycorrhizas. Front. Plant Sci. 2021, 12, 725939. [Google Scholar] [CrossRef]
- Ohtomo, R.; Saito, M. Polyphosphate dynamics in mycorrhizal roots during colonization of an arbuscular mycorrhizal fungus. New Phytol. 2005, 167, 571–578. [Google Scholar] [CrossRef]
- Ferrol, N.; Azcón-Aguilar, C. , Pérez-Tienda, J. Arbuscular mycorrhizas as key players in sustainable plant phosphorus acquisition: An overview on the mechanisms involved. Plant Sci. 2019, 280, 441–447. [Google Scholar] [CrossRef]
- Bueno de Mesquita, C.; Sartwell, S.A.; Ordemann, E.V.; Porazinska, D.L.; Farrer, E.C.; King, A.J.; Spasojevic, M.J.; Smith, J.G.; Suding, K.N.; Schmidt, S.K. Patterns of root colonization by arbuscular mycorrhizal fungi and dark septate endophytes across a mostly-unvegetated, high-elevation landscape. Fungal Ecol. 2018, 36, 63–74. [Google Scholar] [CrossRef]
- Jumpponen, A.; Mattson, K.; Trappe, J. Mycorrhizal functioning of Phialocephala fortinii with Pinus contorta on glacier forefront soil: Interactions with soil nitrogen and organic matter. Mycorrhiza 1998, 7, 261–265. [Google Scholar] [CrossRef] [PubMed]
- He, Y.M.; Fan, X.M.; Zhang, G.Q.; Li, B.; Li, M.R.; Zu, Y.Q.; Zhan, F.D. Influences of arbuscular mycorrhizal fungi and dark septate endophytes on the growth, nutrition, photosynthesis, and antioxidant physiology of maize. Int. J. Agric. Biol. 2019, 22, 1071–1078. [Google Scholar]
- Della Monica, I.F.; Saparrat, M.C.N.; Godeas, A.M.; Scervino, J.M. The co-existence between DSE and AMF symbionts affects plant P pools through P mineralization and solubilization processes. Fungal Ecol. 2015, 17, 10–17. [Google Scholar] [CrossRef]
- Vergara, C.; Campos Araujo, K.M.; Soares, L.A.; de Souza, S.R.; Azevedo, L.S.; Santa-Catarina, C.; da Silva, K; Duarte, G.M.; Ribeiro, G.X.; Zilli, J.E. Contribution of dark septate fungi to the nutrient uptake and growth of rice plants. Brazilian J. Microbiol. 2018, 49, 67–78. [Google Scholar] [CrossRef]
- Newsham, K.K. A meta-analysis of plant responses to dark septate root endophytes. New Phytol 2011, 190, 3–783. [Google Scholar] [CrossRef]
- Spagnoletti, F.; Giacometti, R. Dark septate endophytic fungi (DSE) response to global change and soil contamination. In: Hasanuzzaman, M. (eds) Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II. Springer, Singapore. Springer Singapore, 2020; 629–642. [Google Scholar] [CrossRef]
- Li, X.; Hou, L.; Ren, Y.; Wang, S.; Su, F. Dark septate endophytes isolated from a xerophyte plant promote the growth of Ammopiptanthus mongolicus under drought condition. Sci. Rep. 2018, 8, 26–28. [Google Scholar] [CrossRef]
- Sodhi, G.K.; Saxena, S. Plant growth promotion and abiotic stress mitigation in rice using endophytic fungi: Advances made in the last decade. Environ. Exp. Bot. 2023, 209, 105312. [Google Scholar] [CrossRef]
- Vergara, C.; Araujo, K.E.C.; Urquiaga, S.; Schultz, N.; de Carvalho, F.B.; Medeiros, P.S.; Santos, L.A.; Xavier, G.R.; Zilli, J.E. Dark Septate endophytic fungi help tomato to acquire nutrients from ground plant material. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Usuki, F.; Narisawa, K. A mutualistic symbiosis between a dark septate endophytic fungus, Heteroconium chaetospira, and a nonmycorrhizal plant, Chinese cabbage. Mycologia 2007, 99, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Vergara, C.; Araujo, K.E.C.; Urquiaga, S.; Santa-Catarina, C.; Schultz, N.; da Silva Araújo, E.; de Carvalho Balieiro, F.; Xavier, G.R.; Zilli, J.É. Dark septate endophytic fungi increase green manure-15N recovery efficiency, N contents, and micronutrients in rice grains. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vergara, C.; Araujo, K.E.C.; Sperandio, M.V.L.; Santos, L.A.; Urquiaga, S.; Zilli, J.É. Dark septate endophytic fungi increase the activity of proton pumps, efficiency of 15N recovery from ammonium sulphate, N content, and micronutrient levels in rice plants. Brazilian J. Microbiol. 2019, 50, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Morsomme, P.; Boutry, M. The plant plasma membrane H+-ATPase: Structure, function and regulation. Biochim. Biophys. Acta - Biomembr 2000, 1465, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, K.; Jumpponen, A. Seeking the elusive function of the root-colonising dark septate endophytic fungi. Stud. Mycol. 2005, 53, 173–189. [Google Scholar] [CrossRef]
- Rodriguez, R.; White, J.; Arnold, A.E. Redman, R. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- He, C.; Zeng, Q.; Chen, Y. Colonization by dark septate endophytes improves the growth and rhizosphere soil microbiome of licorice plants under different water treatments. Appl Soil Ecol 2021, 166, 103–993. [Google Scholar] [CrossRef]
- Qiang, X.; Ding, J.; Lin, W.; Li, Q.; Xu, C.; Zheng, Q.; Li, Y. Alleviation of the detrimental effect of water deficit on wheat (Triticum aestivum L.) growth by an indole acetic acid-producing endophytic fungus. Plant Soil 2019, 439, 373–391. [Google Scholar] [CrossRef]
- Bartholdy, B.A.; Berreck, M.; Haselwandter, K. Hydroxamate siderophore synthesis by Phialocephala fortinii, a typical dark septate fungal root endophyte. Biometals 2001, 14, 33–42. [Google Scholar] [CrossRef]
- Ahmed, E.; Holmstrom, S. Siderophores in environmental research: Roles and applications. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef]
- Kajula, M.; Tejesvi, M.V.; Kolehmainen, S.; Mäkinen, A.; Hokkanen, J.; Mattila, S.; Pirttilä, A.M. The siderophore ferricrocin produced by specific foliar endophytic fungi in vitro. Fungal Biol. 2010, 114, 248–254. [Google Scholar] [CrossRef]
- Haas, H.; Eisendle, M.; Turgeon, B.G. Siderophores in fungal physiology and virulence. Annu. Rev. Phytopathol. 2008, 46, 149–187. [Google Scholar] [CrossRef] [PubMed]
- Tienaho, J.; Karonen, M.; Muilu-Mäkelä, R.; Wähälä, K.; Leon Denegri, E.; Franzén, R.; Karp, M.; Santala, V.; Sarjala, T. Metabolic profiling of water-soluble compounds from the extracts of dark septate endophytic fungi (DSE) isolated from scots pine (Pinus sylvestris L.) seedlings using UPLC-Orbitrap-MS. Molecules 2019, 24, 12–2330. [Google Scholar] [CrossRef]
- Wang, S.; Bi, Y.; Christie, P. Effects of extracellular metabolites from a dark septate endophyte at different growth stages on maize growth, root structure and root exudates. Rhizosphere 2023, 25, 100657. [Google Scholar] [CrossRef]
- Wang, S.; Bi, Y.; Quan, W.; Christie, P. Growth and metabolism of dark septate endophytes and their stimulatory effects on plant growth. Fungal Biol. 2022, 126, 674–686. [Google Scholar] [CrossRef]
- Xiang, L.; Gong, S.; Yang, L.; Hao, J.; Xue, M.; Zeng, F.; Zhang, X.; Shi, W.; Wang, H.; Yu, D. Biocontrol potential of endophytic fungi in medicinal plants from Wuhan Botanical Garden. Biol. Cont. 2016, 94, 47–55. [Google Scholar] [CrossRef]
- Scervino, J.M.; Gottlieb, A.M.; Silvani, V.; Pérgola, M. Exudates of dark septate endophyte (DSE) modulate the development of the arbuscular mycorrhizal fungus (AMF) Gigaspora rosea. Soil Biol. Biochem. 2009, 41, 1753–1756. [Google Scholar] [CrossRef]
- Li, X.; He, C.; He, X.; Su, F.; Hou, L.; Ren, Y.; Hou, Y. Dark septate endophytes improve the growth of host and non-host plants under drought stress through altered root development. Plant Soil 2019, 439, 259–272. [Google Scholar] [CrossRef]
- He, Y.M.; Fan, X.M.; Zhang, G.Q.; Li, B. Effects of arbuscular mycorrhizal fungi and dark septate endophytes on maize performance and root traits under a high cadmium stress. S. Afr. J. Bot. 2020, 134, 415–423. [Google Scholar] [CrossRef]
- Vohník, M.; Albrechtová, J. The co-occurrence and morphological continuum between ericoid mycorrhiza and dark septate endophytes in roots of six European rhododendron species. Folia Geobot. 2011, 46, 373–386. [Google Scholar] [CrossRef]
- Belfiori, B.; Rubini, A.; Riccioni, C. Diversity of endophytic and pathogenic fungi of saffron (Crocus sativus) plants from cultivation sites in Italy. Diversity 2021, 13, 535. [Google Scholar] [CrossRef]
- Lingfei, L.; Anna, Y.; Zhiwei, Z. Seasonality of arbuscular mycorrhizal symbiosis and dark septate endophytes in a grassland site in Southwest China. FEMS Microbiol. Ecol. 2005, 54, 367–373. [Google Scholar] [CrossRef]
- Chaudhry, M.S.; Rahman, S.U.; Ismaiel, M.S. Coexistence of arbuscular mycorrhizae and dark septate endophytic fungi in an undisturbed and a disturbed site of an arid ecosystem. Symbiosis 2009, 49, 19–28. [Google Scholar] [CrossRef]
- Jones, S.L.; French, K. Soil nutrients differentially influence root colonisation patterns of AMF and DSE in Australian plant species. Symbiosis 2021, 83, 209–223. [Google Scholar] [CrossRef]
- Díaz-Urbano, M.; Goicoechea, N.; Velasco, P.; Poveda, J. Development of agricultural bio-inoculants based on mycorrhizal fungi and filamentous fungi: Co-inoculants for improve plant-physiological responses in sustainable agriculture. Biol. Cont. 2023, 182, 105223. [Google Scholar] [CrossRef]
- Seaton, F.; Reinsch, S.; Goodall, T.; White, N.; Jones, D.L.; Griffiths, R.; Creer, S.; Smith, A.; Emmett, B.A.; Robinson, D.A. Long-term drought and warming alter soil bacterial and fungal communities in an upland heathland. Ecosystems 2022, 25, 1279–1294. [Google Scholar] [CrossRef]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci 2013, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Shang, X. J.; Luo, Q. X.; Yan, Q.; Hou, R. Effects of the dual inoculation of dark septate endophytes and Trichoderma koningiopsis on blueberry growth and rhizosphere soil microorganisms. FEMS Microbiol. Ecol. 2023, 99, fiad008. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, C.L.; White, M.; Serpe, M.D. Co-inoculation with a dark septate endophyte alters arbuscular mycorrhizal colonization of two widespread plants of the sagebrush steppe. Symbiosis 2021, 85, 343–357. [Google Scholar] [CrossRef]
- Deram, A.; Languereau, F.; Haluwyn, C.V. Mycorrhizal and endophytic fungal colonization in Arrhenatherum elatius L. roots according to the soil contamination in heavy metals. Soil Sediment. Contam. 2011, 20, 114–127. [Google Scholar] [CrossRef]
- Soteras, F.; Renison, D.; Becerra, A.G. Growth response, phosphorus content and root colonization of Polylepis australis Bitt. seedlings inoculated with different soil types. New For. 2013, 44, 577–589. [Google Scholar] [CrossRef]
- Berthelot, C.; Blaudez, D.; Beguiristain, T. Co-inoculation of Lolium perenne with Funneliformis mosseae and the dark septate endophyte Cadophora sp. in a trace element-polluted soil. Mycorrhiza 2018, 28, 301–314. [Google Scholar] [CrossRef]
- Reininger, V.; Sieber, T.N. Mycorrhiza reduces adverse effects of dark septate endophytes (DSE) on growth on conifers. PLoS ONE 2012, 7, e42865. [Google Scholar] [CrossRef]
- Guo, Y.; Matsuoka, Y.; Miura, T.; Nishizawa, T. Ohta, H. Complete genome sequence of Agrobacterium pusense VsBac-Y9, a bacterial symbiont of the dark septate endophytic fungus Veronaeopsis simplex Y34 with potential for improving fungal colonization in roots. J. Biotechnol, 2018, 284, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Luo, R.; Wang, S. Effect of inoculation on the mechanical properties of alfalfa root system and the shear strength of mycorrhizal composite soil. J. China Coal. Soc. 2022, 47, 2182–2192. [Google Scholar]
- Valli, P.S.; Muthukumar, T. Dark Septate Root Endophytic Fungus Nectria haematococca Improves Tomato Growth Under Water Limiting Conditions. Indian J. Microbiol. 2018, 58, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Songachan, L.S.; Kayang, H. Diversity of arbuscular mycorrhizal fungi associated with Flemingia vestita Benth ex Bak. Mycology 2013, 4, 85–95. [Google Scholar] [CrossRef]
- Saravesi, K.; Ruotsalainen, A.L.; Cahill, J.F. Contrasting impacts of defoliation on root colonization by arbuscular mycorrhizal and dark septate endophytic fungi of Medicago sativa. Mycorrhiza 2015, 24, 239–245. [Google Scholar] [CrossRef]
- De Souza, G.L.; Mendonça, J.J.; Teixeira, J.L. , Mangieri de Oliveira, Exotic arbuscular mycorrhizal fungi and native dark septate endophytes on the initial growth of Paspalum millegrana grass. Rev. Caatinga 2019, 32, 607–615. [Google Scholar]
- Guo; X., Wan; Y., Hu; J., Wang; D. Dual inoculations of dark septate endophytic and ericoid mycorrhizal fungi improved the drought resistance of blueberry seedlings. SSRN 2022. [CrossRef]
- Silva, P.S.T; Cassiolatto, A.M.R.; Galindo, F.S.; Jalal, A.; Nogueira, T.A.R.; Oliveira; C.E.; Filho, M.C. M. T. Azospirillum brasilense and zinc rates effect on fungal root colonization and yield of wheat-maize in tropical savannah conditions. Plants 2022, 11, 3154. [Google Scholar] [CrossRef]
- Huo, L.; Gao, R.; Hou; X.; Yu, X.; Yang, X. Arbuscular mycorrhizal and dark septate endophyte colonization in Artemisia roots responds differently to environmental gradients in eastern and central China. Sci ToTENV 2021, 795, 148808. [Google Scholar] [CrossRef]
- Surendirakumar, K.; Chongtham, I.; Pandey, R. R.; Muthukumar, T. Arbuscular mycorrhizal and dark septate endophytic fungal symbioses in Parkia timoriana (DC. in Merr. and Solanum betaceum Cav. plants growing in North East India. Vegetos 2021, 34, 761–774. [Google Scholar] [CrossRef]
- Lugo, M.A.; Menoyo, E.; Allione, L.R.; Negritto, M.A.; Henning, J.A.; Anton, A.M. Arbuscular mycorrhizas and dark septate endophytes associated with grasses from the Argentine Puna. Mycologia 2018, 110, 654–665. [Google Scholar] [CrossRef]
- Ruotsalainen, A.L.; Vatre, H.; Vestberg, M. Seasonality of root fungal colonization in low alpine herbs. Mycorrhiza 2002, 12, 29–36. [Google Scholar] [CrossRef]
- Cao, G.H. Colonization investigation of arbuscular mycorrhizal fungi (AMF) and dark septate endophytes (DSE) in roots of Polygonatum kingianum and their correlations with content of main functional components in rhizomes. Chinese Tradit. Herb. Drugs 2019, 24, 3930–3936. [Google Scholar]
- Gong, M.; Bai, N.; Wang, P.; Su, J.; Chang, Q.; Zhang, Q. Co-Inoculation with arbuscular mycorrhizal fungi and dark septate endophytes under drought stress: Synergistic or competitive effects on maize growth, photosynthesis, root hydraulic properties and aquaporins? Plants 2023, 12, 2596. [Google Scholar] [CrossRef] [PubMed]
- Fernando, A.A.; Currah, R. A comparative study of the effects of the root endophytes Leptodontidium orchidicola and Phialocephala fortinii (Fungi Imperfecti) on the growth of some subalpine plants in culture. Can. J. Bot. 1996, 74, 1071–1078. [Google Scholar] [CrossRef]
- Gao, C.; Li, M.; Liu, R. Combination effects of arbuscular mycorrhizal fungi and dark septate endophytes on promoting growth of cucumber plants and resistance to nematode disease. Mycosystema 2016, 35, 1208–1217. [Google Scholar]
- Reininger, V.; Sieber, T.N. Mitigation of antagonistic effects on plant growth due to root co-colonization by dark septate endophytes and ectomycorrhiza. Environ. Microbiol. 2013, 5, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubian, Y.; Goltapeh, E.M.; Pirdashti, H. Effect of Glomus mosseae and Piriformospora indica on growth and antioxidant defense responses of wheat plants under drought stress. Agric. Res. 2014, 3, 239–245. [Google Scholar] [CrossRef]
- Anith, K.N.; Faseela, K.M.; Archana, P.A.; Prathapan, K.D. Compatibility of Piriformospora indica and Trichoderma harzianum as dual inoculants in black pepper (Piper nigrum L.). Symbiosis 2011, 55, 11–17. [Google Scholar] [CrossRef]
- Baltrus, D.A.; Dougherty, K.; Arendt, K.R.; Huntermann, M; Clum, A.; Pillay, M.; Palaniappan, K; Varghese, N.; Mikhailova, N.; Stamatis, D.; Reddy, T.B.K.; Ngan, C.Y.; Daum, C.; Shapiro, N.; Markowitz, V.; Ivanova, N.; Kyrpides, N.; Woyke, T.; Arnold, A.E. Absence of genome reduction in diverse, facultative endohyphal bacteria. Microb Genom. 2017, 3, e000101. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Matsuoka, Y.; Nishizawa, T.; Ohta, H.; Narisawa, K. Effects of Rhizobium Species Living with the Dark Septate Endophytic Fungus Veronaeopsis simplex on Organic Substrate Utilization by the Host. Microbes Environ 2018, 33, 102–106. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, T.; He, X. Mycorrhizal and dark septate endophytic fungi under the canopies of desert plants in Mu Us Sandy Land of China. Front Agric China 2009, 3, 164–170. [Google Scholar] [CrossRef]
- Xie, L.; Bi, Y.; Ma, S.; Shang, J.; Hu, Q; Christie, P. Combined inoculation with dark septate endophytes and arbuscular mycorrhizal fungi: Synergistic or competitive growth effects on maize? BMC Plant Biol. 2021, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bending, G.D.; Rodriguez-Cruz, M.S.; Lincoln, S.D. Fungicide impacts on microbial communities in soils with contrasting management histories. Chemosph. 2007, 69, 82–88. [Google Scholar] [CrossRef]
- Kernaghan, G; Griffin, A.; Gailey, J. Hussain, A. Tannin tolerance and resistance in dark septate endophytes. Rhizosphere 2022, 23, 100574. [Google Scholar] [CrossRef]
- Bueno de Mesquita, C.P.; Solon, A.J.; Barfield, A.; Mastrangelo, C.F. Adverse impacts of Roundup on soil bacteria, soil chemistry and mycorrhizal fungi during restoration of a Colorado grassland. Appl. Soil Ecol. 2023, 185, 104778. [Google Scholar] [CrossRef]
- Spagnoletti, V.M.; Chiocchio, F.N. Tolerance of dark septate endophytic fungi (DSE) to agrochemicals in vitro. Rev. Argentina Microbiol. 2020, 52, 43–49. [Google Scholar] [CrossRef]
- Manalu, J.N.; Soekarno, B.P.W.; Tondok, E.T.; Surono, S. Isolation and capability of dark septate endophyte against mancozeb fungic. Ilmu Pertan Indones 2020, 25, 193–198. [Google Scholar] [CrossRef]
- Wyss, G.S.; Charudattan, R.; Rosskopf, E.N. Littell, R.C. Effects of selected pesticides and adjuvants on germination and vegetative growth of Phomopsis amaranthicola, a biocontrol agent for Amaranthus spp. Weed Res. 2002, 44, 469–482. [Google Scholar] [CrossRef]
- Widyaningsih, S.; Triasih, U. Pesticides effect on growth of dark septate endophytes in vitro IOP Conf. 012037 IOP Int. Conf Sustain. Agric. 2023, 1172, 012037. [Google Scholar]
- Zhan, F.; He, Y.; Yang, Y.; Li, Y.; Li, T.; Zhao, Z. Effects of tricyclazole on cadmium tolerance and accumulation characteristics of a Dark Septate Endophyte (DSE. in Exophiala pisciphila. Bull. Environ. Contami. Toxicol. 2016, 96, 235–41. [Google Scholar] [CrossRef] [PubMed]
- Ayesha, M. S.; Suryanarayanan, T. S.; Nataraja, K. N.; Rajendra, S.P.; Shaanker, R.U. Seed treatment with systemic fungicides. Front. Plant Sci. 2021, 2, 654512. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.; Caradus, J.R.; Johnson, L.J. Fungal endophytes for sustainable crop production. FEMS Microbiol. Ecol. 2016, 92, 194. [Google Scholar] [CrossRef]
Figure 1.
Typical characteristics of dark septate endophytes. DSE (Macrophomina phaseolina) grown in Petri dishes with PDA medium (A) and melanized hyphae (B). Microsclerotia (C, G) and microsclerotium (D) on PDA medium of Rutstroemia calopus. Scanning electron micrographs microsclerotium of Rutstroemia calopus (E). Cleistothecia of Sordaria (F). DSE root colonization (H, I).
Figure 1.
Typical characteristics of dark septate endophytes. DSE (Macrophomina phaseolina) grown in Petri dishes with PDA medium (A) and melanized hyphae (B). Microsclerotia (C, G) and microsclerotium (D) on PDA medium of Rutstroemia calopus. Scanning electron micrographs microsclerotium of Rutstroemia calopus (E). Cleistothecia of Sordaria (F). DSE root colonization (H, I).
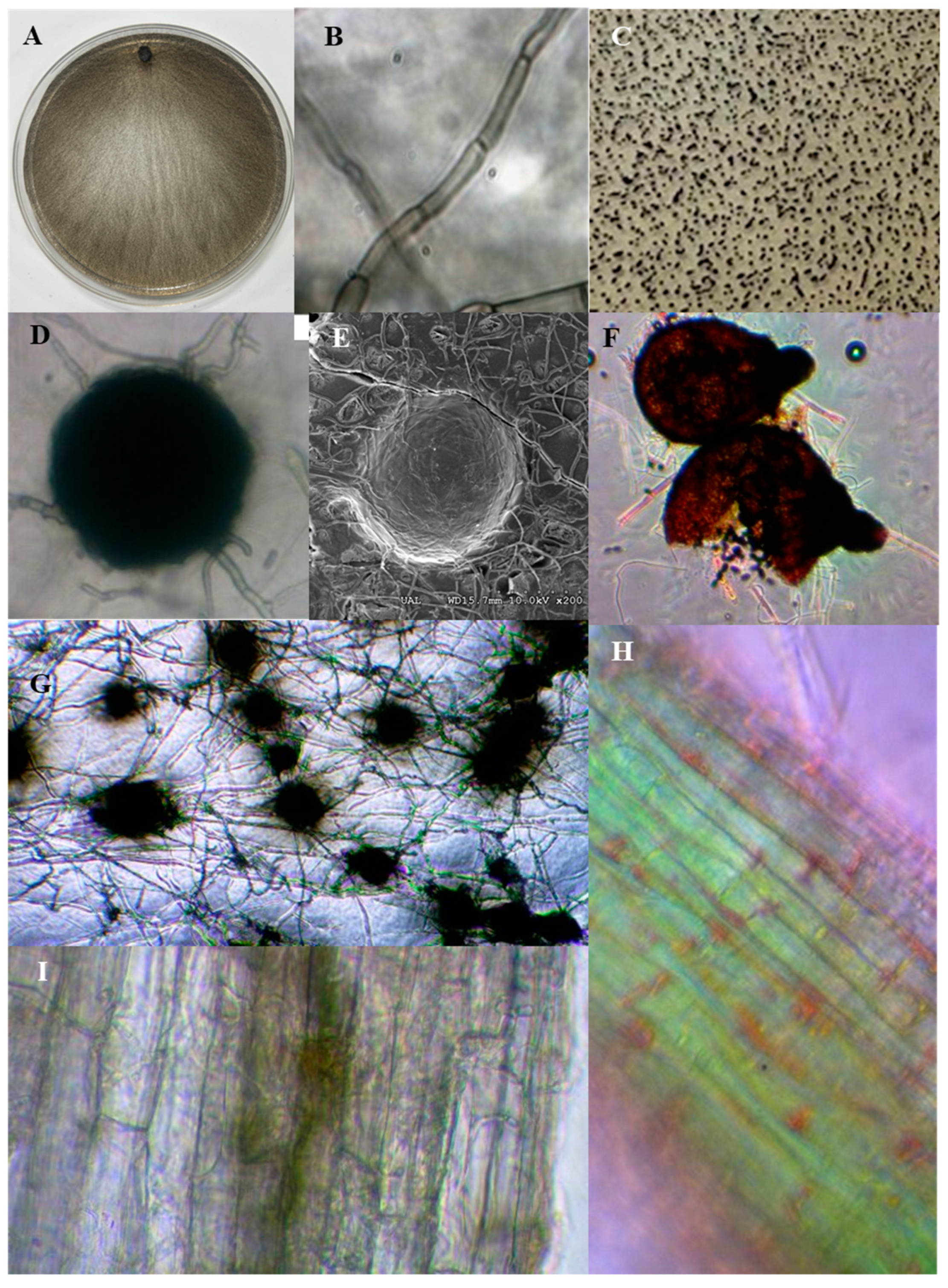
Figure 2.
Actions performed by DSEs to mitigate stress in plants. SOD: Superoxide dismutase; POD: peroxidase activity; CAT: catalase activity; MDA: malondialdehyde; IAA: Indole-3-acetic acid.
Figure 2.
Actions performed by DSEs to mitigate stress in plants. SOD: Superoxide dismutase; POD: peroxidase activity; CAT: catalase activity; MDA: malondialdehyde; IAA: Indole-3-acetic acid.

Figure 3.
Gelatinization of the liquid culture medium at 25 ºC by Stromatinia narcissi (A and B) and Macrophomina phaseolina (C).
Figure 3.
Gelatinization of the liquid culture medium at 25 ºC by Stromatinia narcissi (A and B) and Macrophomina phaseolina (C).


Table 1.
Studies on inoculant compatibility between DSEs and other microorganisms.
| DSE species | Co-inoculum | Host plant | Reference |
|---|---|---|---|
|
Acrocalymma vagum Paraboeremia putaminum |
Trichoderma viride | Astragalus mongholicus | [56] |
| Alternaria sp. | Diversispora epigaea | Zea mays | [134] |
| Amnesia nigricolor | Trichoderma koningiopsis | Vaccinium corymbosum | [122] |
| Cadophora sp. | Funneliformis mosseae | Lolium perenne | [126] |
| Cladosporium cladosporioides | Oidiodendron citrinum | Vaccinium corymbosum | [127] |
| Darksidea | Rhizophagus irregularis |
Artemisa tridentata Poa secunda |
[123] |
| DSE |
Azospirillum brasilense AMF |
Zea mays Triticum aestivum |
[135] |
| DSE | AMF | Medicago sativa | [129] |
| DSE | AM | Artemisia sp. | [136] |
| DSE | AM |
Parkia timoriana Solanum betaceum |
[137] |
| DSE | AM | Poaceae | [138] |
| DSE | AM |
Alchemilla glomerulans Carex vaginata Ranunculus acris ssp. pumilus Trollius europaeus |
[139] |
| DSE | AMF | Polygonatum kingianum | [140] |
| DSE |
Rhizoglomus clarum Claroideoglomus etunicatum Acaulospora morrowiae |
Paspallum millegrana | [133] |
| Exophiala pisciphila | Funneliformis mosseae | Zea mays | [87,113] |
| Exophiala pisciphila | Rhizophagus irregularis | Zea mays | [141] |
| Phialocephala fortinii | Glomus intraradices | Medicago sativa | [100] |
| Phialocephala fortinii | Leptodontidium orchidicola |
Potentilla fruticosa Dryas octopetala Salix glauca Picea glauca |
[142] |
| Phoma leveillei | Acaulospora laevis | Cucumis sativus | [143] |
|
Macrophomina pseudophaseolina Paraphoma radicina |
T. afroharzianum, T. longibrachiatum |
Astragalus mongholicus | [43] |
| Paraphoma chrysanthemicolaGaeumannomyces cylindrosporus |
Suillys bovinus Amanita vaginata |
Pinus tabulaeformis | [20] |
|
Phialocephala turiciensis Acephala applanata P. glacialis Phaeomollisia piceae |
Gigaspora rosea | Trifolium repens | [88] |
|
Phialocephala fortinii Phialocephala subalpina |
Laccaria bicolor | Pseudotsuga menziesii | [127] |
| Phialocephala fortiniis.l.–Acephala applanata species complex (PAC) | Hebeloma crustuliniforme | Picea abies | [144] |
| Piriformospora indica | Glomus mosseae | Triticum aestivum | [145] |
| Piriformospora indica | T. harzianum | Piper nigrum | [146] |
|
Cladosporium cladosporioides Exophiala salmonis Phialophora mustea Paraphoma chrysanthemicola Gaeumannomyces cylindrosporus |
Schizophyllum sp. Suillus laricinus Amanita vaginata Handkea utriformis Suillus tomentosus Suillus bovinus Suillus lactifluus |
Pinus tabulaeformis | [19] |
|
Acephala applanate Phialocephala europaea Phialocephala fortinii Phialocephala Helvetica Phialocephala letzii Phialocephala subalpina Phialocephala turiciensis Phialocephala uotolensis Acephala macrosclerotiorum Phialocephala glacialis |
Paxillus involutus Rhizoscyphus ericae |
Vaccinium corymbosum | [21] |
| Veronaeopsis simplex | Agrobacterium pusense | Solanum lycopersicum | [128] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



