
Submitted:
06 March 2024
Posted:
08 March 2024
You are already at the latest version
Abstract
Noninvasive brain stimulation technique can be used for modulation of motor behaviors such as reaction time (RT). The main aim of this study was to investigate whether RT during a sequential visual isometric pinch task (SVIPT) can be affected by anodal transcranial direct current stimulation (a-tDCS) over three different stimulation sites such as dorsolateral prefrontal cortex (DLPFC), primary motor cortex (M1) or posterior parietal cortex (PPC). We also aimed to assess if stimulation of these cortical sites affect the transfer of learning during SVIPT. A total of 48 right-handed healthy participants were randomly assigned to one of the four a-tDCS groups: 1) left M1, 2) left DLPFC, 3) left PPC and 4) sham. A-tDCS was applied concurrent with SVIPT in which the participants were precisely controlled their forces to reach seven different target forces from 10 to 40% of maximum voluntary contraction (MVC) presenting on a computer screen with right dominant hand. Four test blocks were randomly performed at baseline and 15 min after the intervention including sequence and random blocks with either hand. Our results showed significant differences between a-tDCS groups on ratio RT in the sequence blocks of both the right trained (P = .041) and left untrained hands (P = .023) for target forces of 15% and 30 % of MVC, respectively. In sequence blocks, M1 showed significant elongation in ratio RT for the target force of 15% and 30 % MVC compared to PPC and Sham groups in the right hand. In sequence blocks with left hand, this negative effect were found between M1-DLPFC and M1-Sham groups. In random blocks, no significant differences were found among four groups at each target forces for the left hand while there were significant differences for 15% (P =. 04) and 40% MVC (P =. 035) in the right hand. Our findings suggest that three different stimulation sites (DLPFC, M1 or PPC) were differentially affected by a single session of a-tDCS at some target forces during SVIPT. The effects of a-tDCS on ratio RT were transferred in the left untrained hand for sequence blocks but not random blocks. Further research is needed to understand fundamental aspects of these main areas of the brain on temporal processing in a precision control task such as SVIPT.
Keywords:
anodal transcranial direct current stimulation
; a-tDCS
; Reaction time
; transfer learning
; primary motor cortex
; dorsolateral prefrontal cortex
; posterior parietal cortex
1. Introduction
The ability to acquire a motor skill is expressed by a significant reduction in reaction time (RT) or error rate through practice [1]. Reduced RT after training is related to neuroplasticity in different areas of the frontoparietal cortex (FPC) [2,3,4]. Shorter RT in response to expected visual stimuli has been mainly associated with increased activation of the posterior parietal cortex (PPC) [5]. The PPC is strongly associated with sensorimotor integration for perception and action [6]. The dorsolateral prefrontal cortex (DLPFC) has been activated for inhibition of unrelated stimuli to produce the best response to stimuli in difficult task demands [7]. The primary motor cortex (M1) is a key motor area that is mainly activated in the process of acquiring a motor skill through the sustained learning of complex movements [8,9,10,11]. Although neuroimaging studies reveal important insights into brain areas involved in motor timing, further research is needed to determine the essential role of different areas of the brain in RT as one of the most important temporal variables during motor learning.
Non-invasive brain stimulation methods like transcranial direct current stimulation (tDCS) can be used to determine the specific role of different brain areas involved in the temporal processing of a certain task. TDCS is the application of a weak direct electrical current via the scalp to modulate cortical excitability in the human brain [12]. The application of anodal tDCS (a-tDCS) over cortical target areas depolarizes the resting membrane potentials of the neurons, which may cause increased excitability [13,14]. This may lead to the formation of stronger and more effective synaptic connections between activated neurons during the learning process [15,16,17]. Changes in physical performance following the application of a-tDCS over M1 have been reported in sequenced learning tasks such as a serial reaction time task (SRTT) [18,19,20,21,22] and sequential visual isometric pinch task (SVIPT) [23,24,25]. Even though there are some studies on the effects of a-tDCS on RT in SRTT, little is known about the effects of brain stimulation on SVIPT. SVIPT is a force control sequenced task with greater motor demands compared to SRTT, which is a key pressing task in which participants focus on cognitive functions rather than motor functions.
Both human and animal studies have demonstrated that the sequential knowledge acquired in one hand transfers to the other hand [26,27,28]. Such a phenomenon is called “intermanual transfer” and it reflects how unilateral hand practice affects the performance of the other hand [29,30,31,32,33]. Neuroimaging studies revealed that training with one hand led to excitatory or inhibitory activity in both hemispheres [34,35]. It is well-known that the corpus callosum is the main neural pathway that connects left and right cortical areas, including the prefrontal, motor, somatosensory, parietal, and occipital areas on either hemisphere [36]. Indeed, the corpus callosum enables the transfer of motor skills from one hand to the other hand. For example, bilateral M1 activation has been reported when participants performed SRTT training with one hand [37,38]. The transfer learning is not only observed from one hand to another hand but also seen from one task to another task. Although some studies confirmed the presence of intermanual transfer of learning in SRTT, little is known about the effects of brain stimulation on the transfer of learning in SVIPT. Therefore, in the current study, we aimed to investigate whether a-tDCS over three stimulation sites (DLPFC, M1 or PPC) could differentially affect RT during SVIPT. We also aimed to explore whether these effects are transferred to the untrained hand.
2. Methods and Materials
2.1. Participants and Study Design
A convenience sampling was employed to recruit participants in this study, which was a parallel randomized single-blind sham-controlled study. Forty-eight healthy right-handed students (34 females, 14 males; 25.83 ± 6.174) from Monash University participated in this study. For the allocation of participants in each intervention, a random numbers table was used to generate the random allocation sequence. Participants were blinded to the experimental conditions and randomly assigned to one of the four stimulation groups: 1) a-tDCS of left M1, 2) a-tDCS of left DLPFC, 3) a-tDCS of left PPC, 4) sham a-tDCS (Figure 1). All participants were right-handed, based on the Edinburgh Handedness Inventory (Oldfield, 1971). Participants were excluded if they had contraindications for receiving tDCS, a history of neurological or psychiatric diseases and significant experience with the use of musical instruments. All participants were naive to the purpose of the experiments. All participants signed a consent form before taking part in our experiment. The study was approved by the Human Ethics Committee at Monash University which follows the declaration of Helsinki (F13/3302_2013001720).
| Figure 1. | CONSORT flow diagram. |
2.2. Procedure
A force transducer (AD Instrument MLT004/ST, NSW, Australia) was used for SVIPT [39] (Figure 2). For this task, participants were instructed to squeeze a force transducer between their thumb and index finger and match their force production on the force transducer as precisely and quickly as possible to reach each target force which appeared on a computer screen. A PowerLabTM (4/35) was used and directly connected to the force transducer to convert voltage signals to digital signals. The target forces were designed from 10 to 40% of Maximum voluntary contraction (MVC) in each trial. A simple random number was employed to create the sequence order (10, 35, 20, 40, 25, 15, and 30% MVC), which was used in this study. At the beginning of each experiment, MVC was individually determined for each participant. Two trials were then given as familiarization. After familiarization, two sequences or random blocks with each hand were randomly performed as baseline measurement. For delivering visual targets in either random or sequence order, a number of macros were developed in the PowerLabTM ADInstrument 4/35 with LabChartTM. Each block consisted of eight trials and each trial included seven target forces which appeared in a sequence order (10, 35, 20, 40, 25, 15, and 30% MVC) or random order on the computer screen. During training, each participant completed eight blocks of the same sequence order with the dominant hand, except for block 6 which was set in a random order. Participants were not aware of the order of sequence during and after the training. Fifteen minutes after concurrent application of both training and brain stimulation, participants randomly completed four test blocks as a post-test assessment including sequence right (Seq.R), sequence left (Seq.L), random right (Ran.R) and random left (Ran.L) hand. RTs as behavioral outcomes were measured in each assessment block.
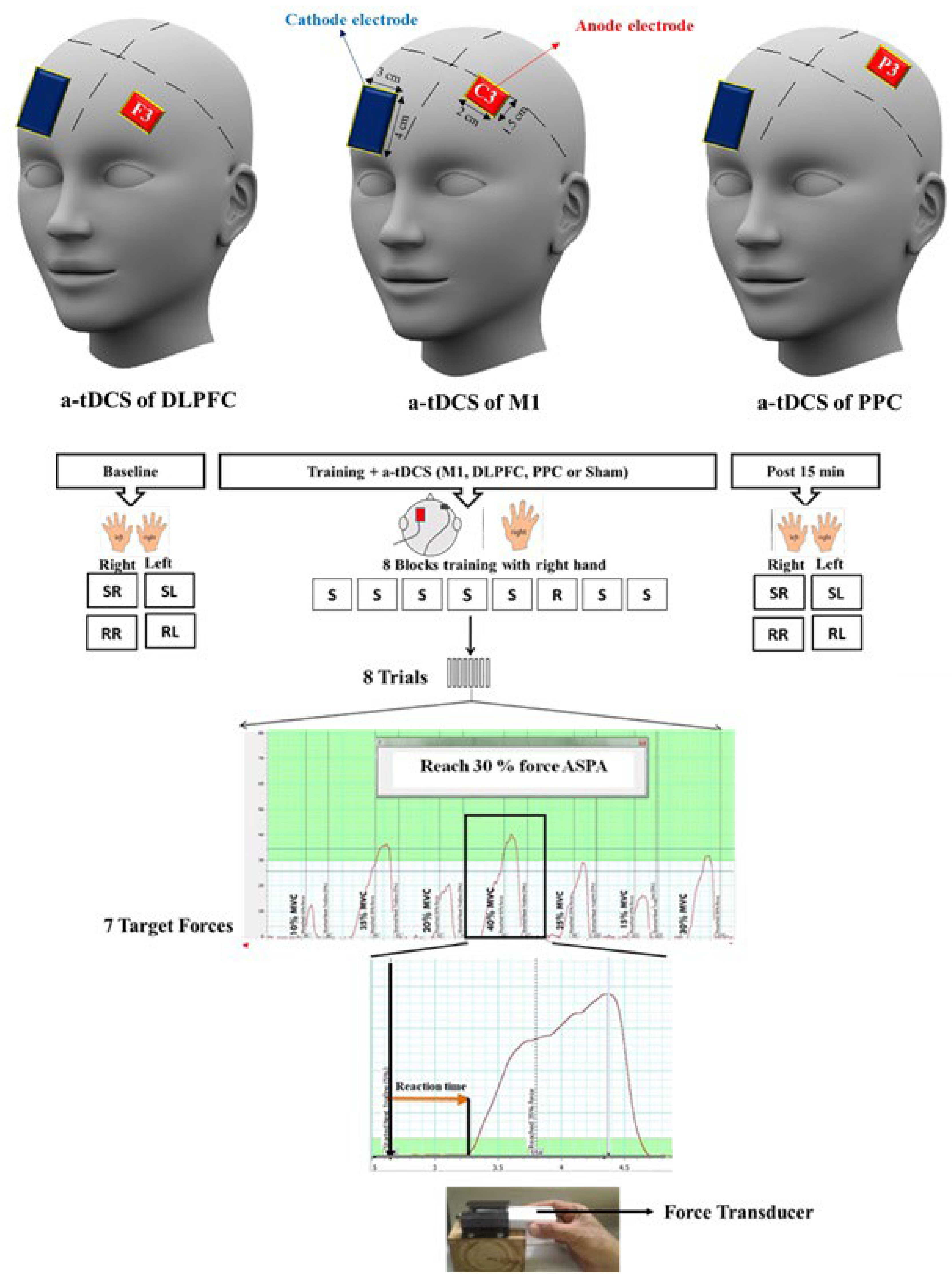
Figure 2.
Experimental set up. Participants were instructed to squeeze a force transducer as precisely as possible to reach each target force that appeared on the computer screen. Each sequence block consisted of eight trials, which included seven different target forces from 10 to 40 % of their MVC. In a sequence block, target forces appeared in a sequence order (10, 35, 20, 40, 25, 15 and 30% of MVC) while target forces were randomly presented in a random block. They were asked to complete each block as quickly and accurately as possible. RT was measured as temporal variables for each target force. SVIPT: Sequential visual isometric pinch task, A-tDCS: Anodal transcranial direct current stimulation, M1: Primary motor cortex, DLPFC: Dorsolateral prefrontal cortex, PPC: Posterior parietal cortex, S: Sequence block, R: Random block. RT: reaction time, SR: Sequence right, SL: Sequence left, RR: Random right, RL: Random left.
Figure 2.
Experimental set up. Participants were instructed to squeeze a force transducer as precisely as possible to reach each target force that appeared on the computer screen. Each sequence block consisted of eight trials, which included seven different target forces from 10 to 40 % of their MVC. In a sequence block, target forces appeared in a sequence order (10, 35, 20, 40, 25, 15 and 30% of MVC) while target forces were randomly presented in a random block. They were asked to complete each block as quickly and accurately as possible. RT was measured as temporal variables for each target force. SVIPT: Sequential visual isometric pinch task, A-tDCS: Anodal transcranial direct current stimulation, M1: Primary motor cortex, DLPFC: Dorsolateral prefrontal cortex, PPC: Posterior parietal cortex, S: Sequence block, R: Random block. RT: reaction time, SR: Sequence right, SL: Sequence left, RR: Random right, RL: Random left.
As shown in Figure 2, RT is the interval between appearances of a stimulus (force target) on the computer screen until the moment where the force response was taken above a resting range. The mean of RT for eight repetitions of the same target force across a block was calculated as RT for the given target in that block. The ratio RT [(pre-post/pre) *100] was also measured in each target force for all four test bocks (Seq.R, Seq.L, Ran.R and Ran.L).
2.3. Transcranial Direct Current Stimulation (tDCS)
A commercial stimulator (Intelect Advanced Therapy System, Chattanooga, TN, USA) was used to deliver a direct current with an intensity of 0.3 mA for 20 min during training. The active electrode (1.5 × 2 = 3 cm2) was placed over the left M1, DLPFC, and PPC and the return electrode (4 × 3 = 12 cm2) was placed over the contralateral supraorbital region. The small size of electrodes yields a highly focused direct current over the target regions, the current intensity for the small electrode size were adjusted by keeping the current density (0.1 mA/cm2) in a safe range. Two electrodes were covered by saline-soaked sponges and strapped in place by two elastic bands [40]. The location of the M1 area was identified using transcranial magnetic stimulation (TMS) and centered on the representational field of the right first interosseous muscle (FDI), which plays a dominant role during SVIPT [32]. The location of DLPFC (F3) and PPC (P3) were determined using the international 10-20 system. Participants reported the side effects under the electrodes such as itching, tingling, burning sensations, and burning pain and adverse effects such as headache [41]. If participants reported burning pain or any other side effects such as itching or burning under the electrodes, we injected some normal saline into the sponges using a syringe to keep them wet throughout the experiment [40]. For the sham stimulation group, the same procedure was performed but the current was applied for only 30 s. The active electrode was randomly positioned over the three different stimulation areas (M1, DLPFC, or PPC). The transient current was ramped up to 0.3 mA and then ramped down so that participants received an initial sensation for 30 seconds of stimulation.
In each experiment (Figure 2), the same procedure was followed: 1) baseline assessment, 2) concurrent training with anodal/sham tDCS, 3) assessment 15 min after the interventions. The participants randomly performed four blocks consisted of 7 trials in either sequence or random orders with either hand (Seq.R, Seq.L, Ran.R and Ran.L) at two time points: baseline and after intervention.
2.4. Data Analysis
Sample size calculation: A power analysis (G-Power v3.1) for a F test; ANOVA: Fixed effects, omnibus, one-way, was used to calculate the sample size for this study. In G-Power, this test can be applied for a nonparametric Kruskal Wallis Test. A total sample size of 48 participants was determined for a power of .8 with the alpha set at 0.05 and an effect size of 0.5.
The normality of data was assessed using the Kolmogorov-Smirnov (K-S) test. For normal distributed variables, a two-way ANOVA was used with two independent factors (Groups and Blocks) as between-subject factors and time (baseline and 15 min after stimulation) as within subject factor. A one-way ANOVA was conducted to examine significant differences in participants’ characteristics among the four groups at baseline. If normality was violated, the non-parametric analysis, the Kruskal Wallis Test, was used to determine differences in the mean rank of variables among four groups separate from all four assessment blocks. When appropriate, paired comparisons were carried out using the Wilcoxon signed rank test.
SPSS (version 20) and MATLAB (R2014a) were used to analyze the data in this study. Statistical significance was set at p = .05.
3. Results
The results of the one-way ANOVA showed no significant differences in participant’s characteristics among four groups (P < .05). All of the participants tolerated tDCS and reported no side-effects during or after the experimental session.
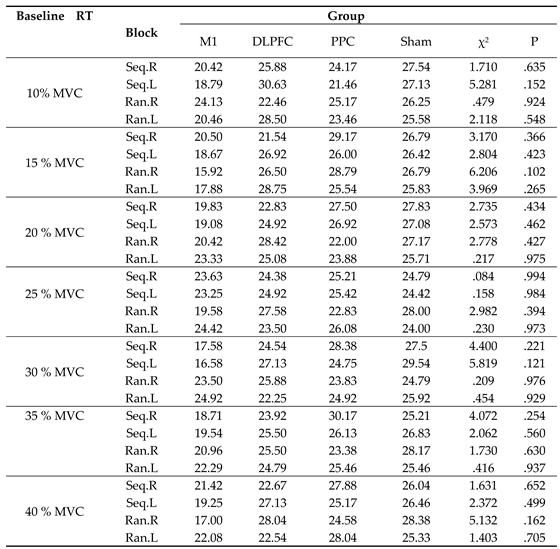
The results of the Kolmogorov-Smirnov (K-S) test revealed that the measured temporal variables i.e., RT and their ratio of RT were not normally distributed. Therefore, the Kruskal Wallis Test was used to determine the differences in these variables for each test block among four groups. As shown in Table 1, there were no significant differences in the mean rank of RTs for all target forces among four groups at the baseline (p > .05).

Table 1.
The results of the Kruskal Wallis Test for four test blocks on the mean rank of ratio RT (either sequence or random blocks with either hand) among the four stimulation groups (M1, DLPFC, PPC and sham). RT: reaction time; Seq: sequence; Ran: random; R: right; L: left.
Table 1.
The results of the Kruskal Wallis Test for four test blocks on the mean rank of ratio RT (either sequence or random blocks with either hand) among the four stimulation groups (M1, DLPFC, PPC and sham). RT: reaction time; Seq: sequence; Ran: random; R: right; L: left.
 |
3.1. Ratio RT for Sequence Blocks in Both Right and Left Hands
The results of the Kruskal Wallis Test showed that there were significant differences among a-tDCS groups in the ratio of RT at target forces of 15% and 30 % MVC for both right and left hands (P < .05) (Table 2) (Figure 3 and Figure 4).
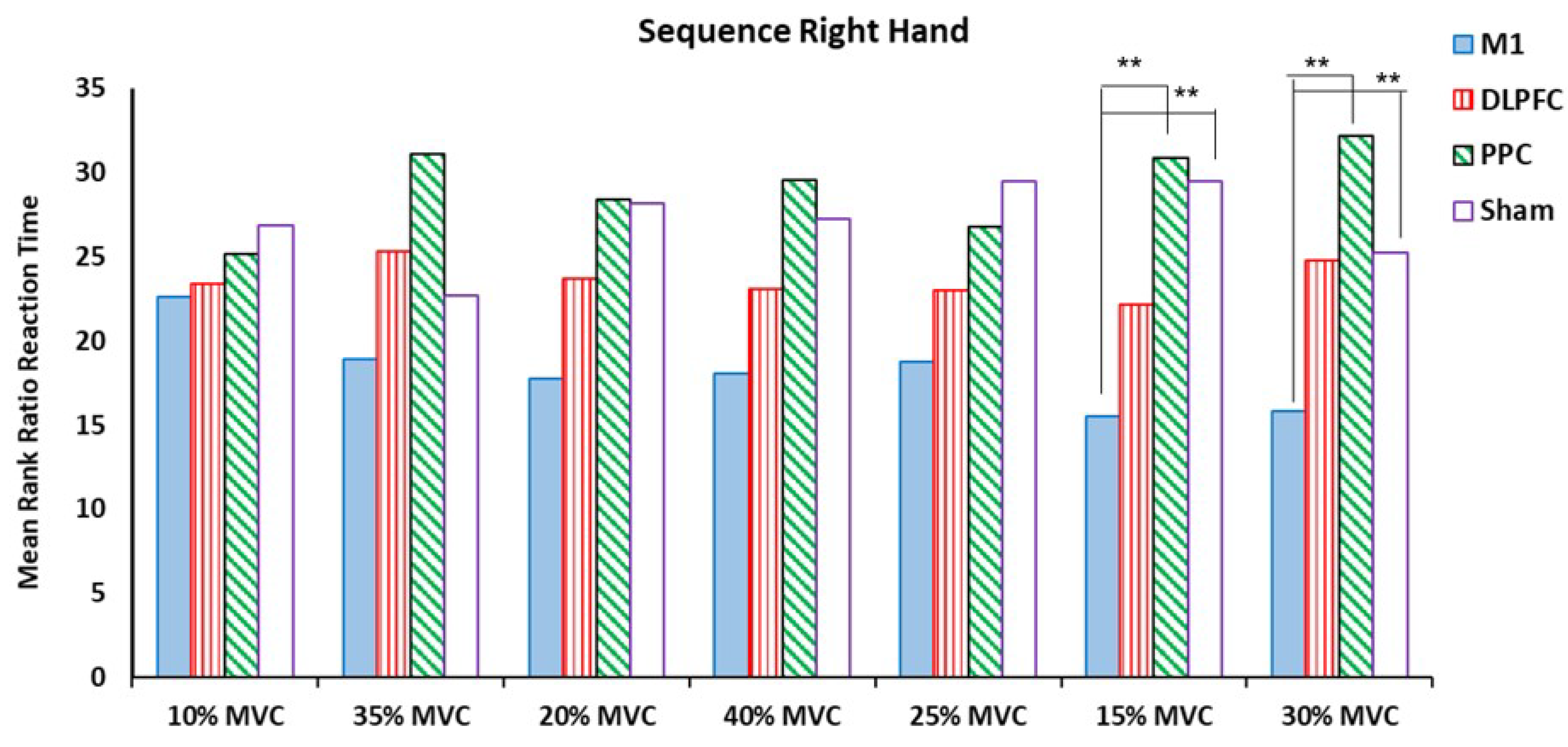
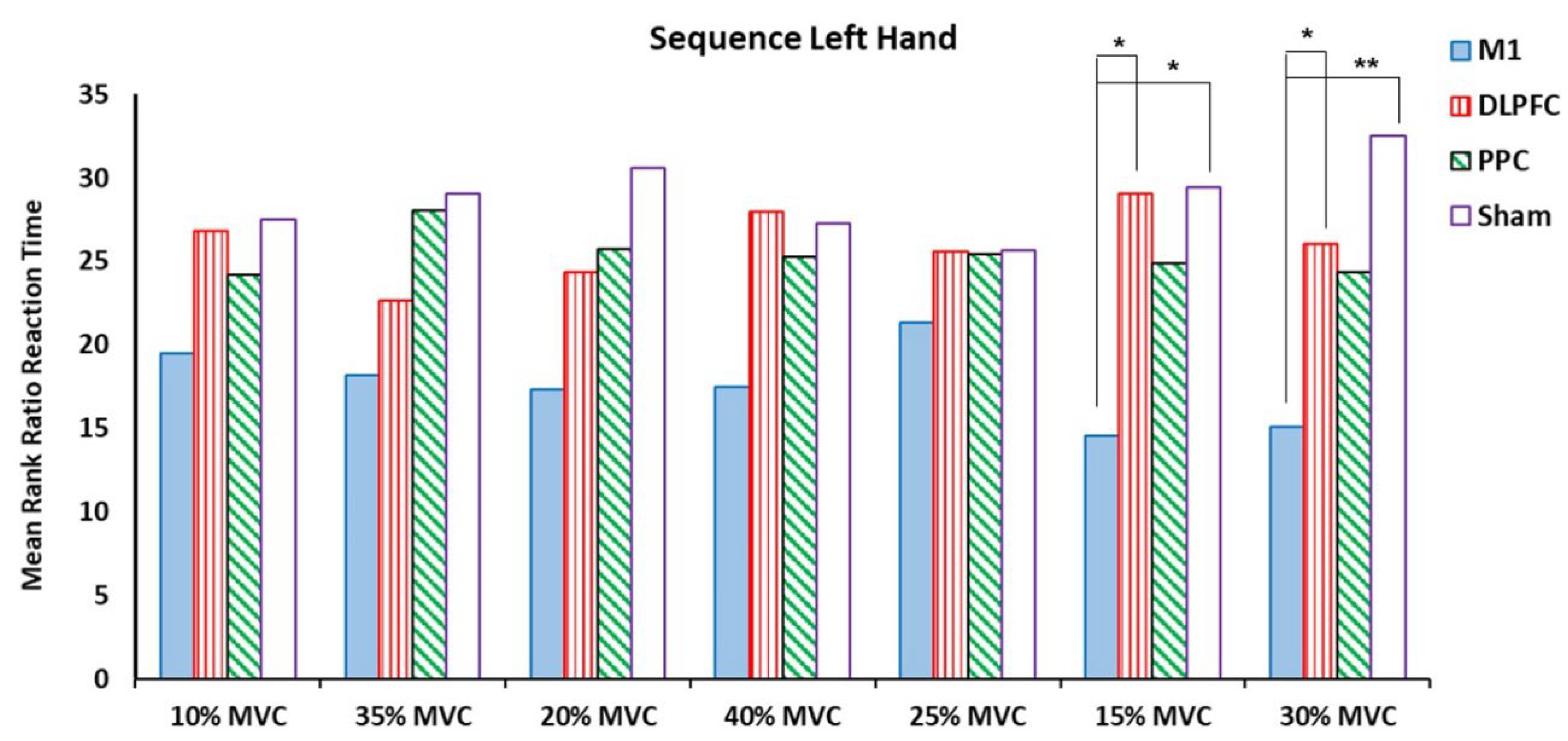
Table 2.
The results of the Kruskal Wallis Test on mean rank of ratio RT in four assessment blocks (either sequence or random blocks with either hand) among four stimulation groups (M1, DLPFC, PPC and sham).
Table 2.
The results of the Kruskal Wallis Test on mean rank of ratio RT in four assessment blocks (either sequence or random blocks with either hand) among four stimulation groups (M1, DLPFC, PPC and sham).
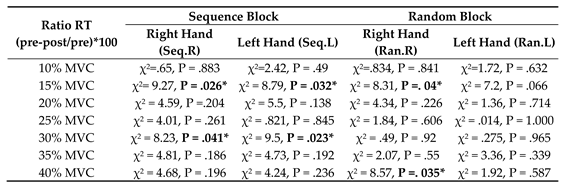 |
Figure 3.
The mean rank of RT ratio in the sequence right hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
Figure 3.
The mean rank of RT ratio in the sequence right hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
Figure 4.
The mean rank ratio RT in sequence left hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
Figure 4.
The mean rank ratio RT in sequence left hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
For the right trained hand, the results of pairwise comparison showed that participants who received a-tDCS over left M1 had significant elongation in the ratio of RT for the force 15% MVC compared to two other groups, PPC (Z = -2.59, P = .009) and sham (Z = -2.59, P = .009). For force target 30% MVC, this negative effect was also observed between M1-PPC (Z = -2.59, P = .009) and M1-sham (Z = -2.021, P = .043) groups. No significant differences were found in other target forces (Figure 3).
For the left untrained hand, the Kruskal Wallis Test showed significant differences at the same temporal measured variables, i.e., 15 % and 30% MVC (Table 2). The results of pairwise comparison showed that M1 compared to DLPFC (Z = -2.54, P = .011) and Sham (Z = -2.483, P = .013) groups showed increase in the measured variable for force target 15% MVC. Significant differences were found between M1-DLPFC (Z = -2.13, P = .033) and M1-sham (Z = -2.598, P = .009) in favors of DLPFC and sham at force target of 30% MVC (Figure 4).
3.2. Ratio RT for Random Blocks in Both Right and Left Hands
For the right trained hand, the results of the Kruskal Wallis Test showed significant differences on ratio RT for force targets of 15% and 40% of MVC (Table 2). Pairwise comparison revealed that a-tDCS on left M1 had a negative effect on the measured temporal variable at force target of 15% MVC compared to the PPC (Z = -2.598, P = .009) and sham (Z = -2.252, P = .024) groups. For the force target of 40% MVC, a-tDCS over left M1 showed significant elongation in the ratio of RT compared to three other groups, i.e., DLPFC, PPC (Z = -2.021, P = .043) and sham (Z = -2.598, P = .009) groups (Figure 5).
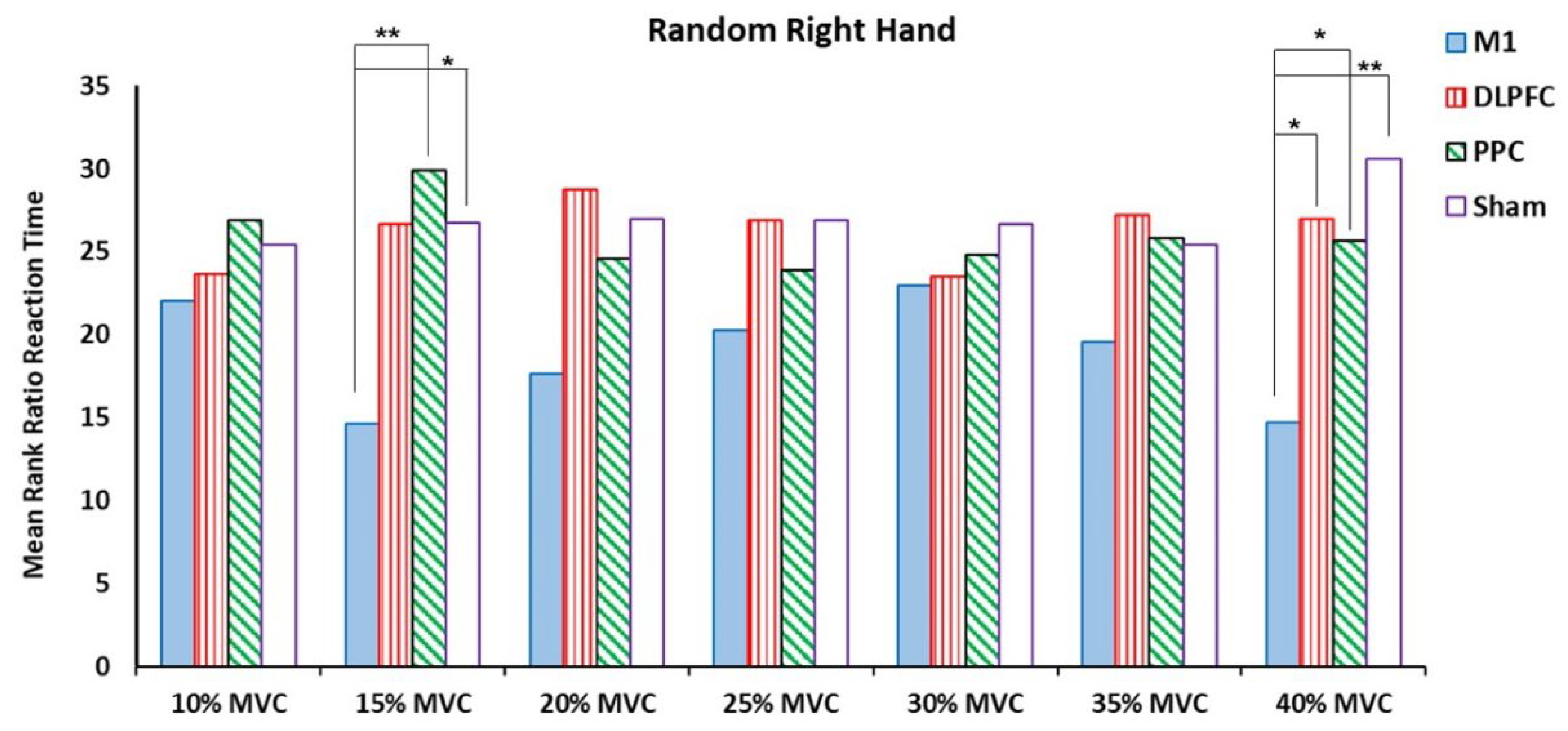
Figure 5.
The mean ranks the ratio of RT in the random right-hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
Figure 5.
The mean ranks the ratio of RT in the random right-hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
For the left untrained hand, no significant effects were found on the ratio of RT at any force target among the four tDCS stimulation sites (Table 2) (Figure 6).
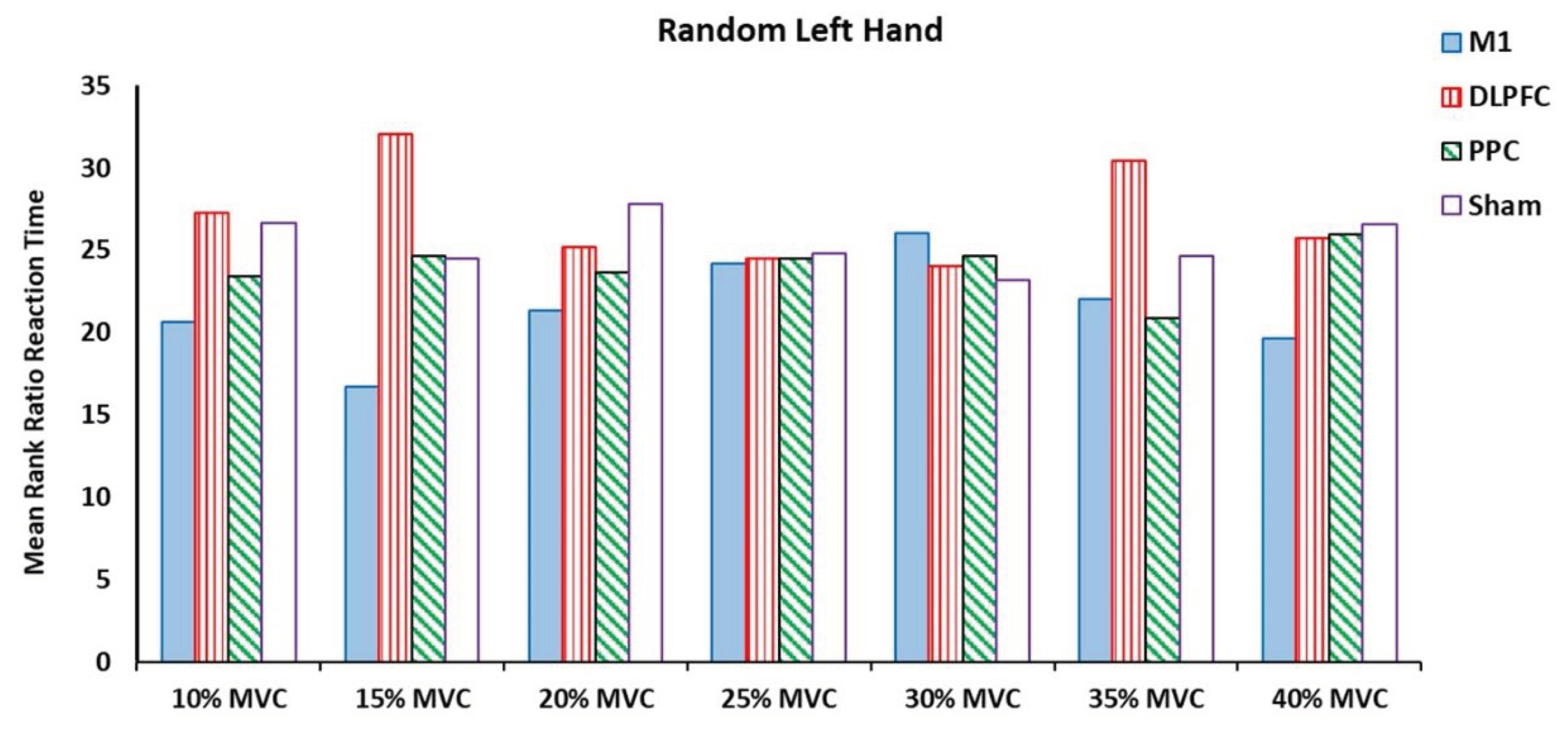
Figure 6.
Mean rank ratio RT in the random left-hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
Figure 6.
Mean rank ratio RT in the random left-hand block assessment test among four tDCS stimulation sites (M1, DLPFC, PPC and sham).
4. Discussion
Our findings showed that participants who received the left M1 stimulation showed a significant increase in the RT ratio for some target forces compared to the sham group, while DLPFC and PPC stimulation did not modify RT within the SVIPT. The observed elongations in the ratio of RT after M1-a-tDCS were transferred into the untrained hand in sequence blocks of SVIPT but not random blocks. In the current study, we aimed to assess whether RT during SVIPT was differentially affected by stimulation of three different areas of the FPC. No improvement in RT was observed after a single session of a-tDCS concurrent with SVIPT training. Unlike DLPFC and PPC a-tDCS, which resulted in no effects on RTs, significant impairments were observed in this variable after M1 stimulation.
Contrary to our results, Waters-Metenier et al. (2014) observed an enhancement in both execution time and RT following a 4-day application of bihemespheric M1 a-tDCS with an intensity of 2 mA and an electrode size of 35 cm2 during a piano-like key task [42]. In contrast, Horvath et al. (2016) found no significant effects of anodal or cathodal M1 tDCS (2 mA or 1 mA with electrode size of 35 cm2) on a simple motor reaction time task [43]. They suggested that tDCS over M1, regardless of polarity, stimulation intensity, and electrode montage, might not have a positive effect on reaction time in a relatively lower-level motor behavioral task [43]. In the current study, we applied a single session of a-tDCS with an intensity of 0.3 mA and a small electrode size of 3 cm2 over M1 during a complex sequential motor task in which participants control their forces to reach different target forces appeared on the computer screen. We used a focal small electrode size of 3 cm2 to selectively stimulate the M1 area, not nearby areas, such as the primary sensory area, premotor cortex, or supplementary motor area. It is probable in the current study, M1 representations of the specific muscles that are involved in the SVIPT task were not selectively stimulated by the 3-cm2 tDCS. Nitsche et al. (2007) showed that reducing stimulation electrode size produces changes in M1 excitability only for the muscle representation covered by the small tDCS electrode, not for the muscle representation of the adjacent areas [44]. In addition, focal stimulation of the M1 area with an electrode size of 3 cm2 can decrease stimulation in nearby areas such as the premotor, or supplementary areas, which may influence M1 excitability. Boros et al. (2008) found that anodal stimulation of the premotor (0.1 mA, 3.5 cm2, 13 min) modifies the intracortical excitability of the ipsilateral M1 [45]. Elbert and co-workers observed that application of anodal tDCS (0.26 mA, 1.5 cm2) at the vertex close to the supplementary motor area can improve RT in a tone-noise sequences task [46]. Therefore, activity modulation of adjacent interconnected areas might increase the effects of M1 a-tDCS stimulation in order to improve RT processing. In our study, a small electrode size of 3 cm2 over the left M1 area was used. The stimulation of nearby areas such as premotor or supplementary areas and their effects on RT during SVIPT should be explored in future studies.
In our study, we found no significant effects of left DLPFC stimulation on RTs within the SVIPT compared to sham stimulation. Marshall et al. (2005) showed that both anodal and cathodal stimulation (260 μA; 15 sec-on/15 sec-off; 8 mm diameter, 15 min) impaired reaction time processing in a working memory task [47]. In contrast, enhancement effects in a stop-signal reaction time were observed following the right DLPFC a-tDCS stimulation (0.5 mA, 9 cm2, 19 min) with extra cephalically montage on the contralateral deltoid [48]. They found that a-tDCS over the right DLPFC can improve cognitive inhibition processes in a stop-signal reaction time by making fewer omission errors [48]. Contrary to their results, we observed no positive effects on RTs within SVIPT in participants who received the left DLPFC a-tDCS compared to the sham group. These discrepancies can be explained by the different methodologies used in these studies. In our study, we stimulated left DLPFC with a contra-orbital montage in a constant, not intermittent, manner during a pinch-force sequential task. With regards to the positive effects observed in RT following the right DLPFC in a recognition reaction time task, it might be valuable in future studies to explore the effects of the right DLPFC tDCS on RTs within SVIPT.
In the current study, we also observed no significant effects of left PPC stimulation on RTs within SVIPT. However, the relevance of the left PPC as an anticipatory center for precise sensorimotor timing has been identified in the study by Krause et al. (2012). They showed that activity in the PPC is essential for precise execution of sensorimotor tasks, especially when quick adjustment of movements is required in response to external stimuli [49]. In addition, Heinen et al. (2016) have shown that bilateral PPC stimulation, independent of electrode configuration, can enhance visual working memory precision more than unilateral PPC stimulation [50]. They also found that cathodal but not anodal tDCS over the right PPC can improve general working memory precision [50]. Although the SVIPT task used in the current study, was not similar to theirs, bilateral PPC stimulation or cathodal PPC stimulation within SVIPT should be explored in future studies.
In this study, we also aimed to assess the differential effects of brain stimulation over three different areas of the FPC on the transfer of learning within SVIP. No transfer learning was observed in the DLPFC and PPC stimulation groups. We also observed that the impairments in the ratio of the RTs in the M1 group were transferred to the left untrained hand. The present result is in line with a study by Keitel et al. (2018) showing that a-tDCS applied to the right M1 impairs implicit motor sequence learning of both hands [51]. They applied a-tDCS (9 cm2, 0.25 mA, 10 min) over right (ipsilateral) M1 during SRTT with the right trained hand [51]. In the current study, we applied a-tDCS over left (contralateral) M1 during SVIPT training with the right hand. In both studies, participants were not aware of the underlying sequential pattern indicating implicit learning, which is mediated by a cortico-striatal-cerebellar network [52]. The observed negative interannual transfer in M1 group showed that there is an interaction between bilateral M1, which support the hypothesis of interhemispheric rivalry. Therefore, the inhibitory effect of left M1 a-tDCS on implicit motor sequence learning was seen in both hands. Regarding the fact that the a-tDCS technique used in this study showed no significant improvement on RTs, further research is needed to investigate the impact of different stimulus conditions of tDCS in terms of electrode montage, current intensity, or electrode size on RT in SVIPT.
Limitations
The findings in the current study should be interpreted in light of a number of limitations. We included healthy young individual participants, so we cannot generalize our findings to the elderly population or patients with neurological disorders. We computed the sample size required for a parametric test in G power in this study. For a non-parametric test, we need to add at least 15% to the total sample size. Therefore, the recruitment of more participants could increase the power of this study to find significant differences between groups, if any exist. This study was single-blinded, where participants were not aware of the type of stimulation while the researcher was not blinded to the intervention groups, which may increase the risk of bias. Long-term outcome measures were not evaluated in this study. Therefore, it is suggested that future studies investigate the effects of brain stimulation on behavior outcomes at longer follow-up times within SVIPT.
5. Conclusion
Our results demonstrated an elongation in the ratio of RTs following left M1 stimulation compared to the sham group. No significant effects were observed after left DLPFC and PPC stimulation on the ratio of RTs in SVIPT compared to sham group stimulation. We also found that the observed impairments in RTs in the M1 a-tDCS group were transferred into the untrained hand only for sequence blocks of SVIPT.
Acknowledgments
The authors gratefully thank Sabastian White for his help to create macros in LabChart software to measure RT and automation analysis of data obtained from the force transducer. The authors are grateful to all participants who participated in this study. We also appreciate the financial support of Monash University which made this project possible.
Conflicts of Interest statement
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
- Cohen, N.J. and L.R. Squire, Preserved learning and retention of pattern-analyzing skill in amnesia: Dissociation of knowing how and knowing that. Science, 1980, 210, 207–210. [CrossRef]
- Pecenka, N., A. Engel, and P.E. Keller, Neural correlates of auditory temporal predictions during sensorimotor synchronization. Frontiers in human neuroscience, 2013, 7, 380. [CrossRef]
- Karabanov, A., et al., The dorsal auditory pathway is involved in performance of both visual and auditory rhythms. Neuroimage, 2009, 44, 480–488. [CrossRef]
- Coull, J. and A. Nobre, Dissociating explicit timing from temporal expectation with fMRI. Current opinion in neurobiology. 2008, 18, 137–144. [CrossRef]
- Coull, J.T., J. Cotti, and F. Vidal, Differential roles for parietal and frontal cortices in fixed versus evolving temporal expectations: Dissociating prior from posterior temporal probabilities with fMRI. Neuroimage. 2016, 141, 40–51. [CrossRef]
- Rivera-Urbina, G.N., et al., Parietal transcranial direct current stimulation modulates primary motor cortex excitability. Eur J Neurosci. 2015, 41, 845–855. [CrossRef]
- Brazovskaya, F., A. Malikova, and R. Pavlygina, After-effects of anodal polarization in the cat cerebral cortex. Neurophysiology. 1972, 4, 194–199. [CrossRef]
- Hodgson, R., et al., Training-induced and electrically induced potentiation in the neocortex. Neurobiology of learning and memory. 2005, 83, 22–32. [CrossRef]
- Monfils, M.H. and G.C. Teskey, Skilled-learning-induced potentiation in rat sensorimotor cortex: a transient form of behavioural long-term potentiation. Neuroscience. 2004, 125, 329–336. [CrossRef]
- Rioult-Pedotti, M.S., D. Friedman, and J.P. Donoghue, Learning-induced LTP in neocortex. Science. 2000, 290, 533–536. [CrossRef]
- Muellbacher, W., et al., Early consolidation in human primary motor cortex. Nature. 2002, 415, 640–644. [CrossRef]
- Nitsche, M.A. and W. Paulus, Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J Physiol. 2000, 527 Pt 3, 633–639. [CrossRef]
- Nitsche, M.A., et al., Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [CrossRef]
- Priori, A., et al., Polarization of the human motor cortex through the scalp. Neuroreport. 1998, 9, 2257–2260. [CrossRef]
- Nitsche, M.A. and W. Paulus, Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology. 2001, 57, 1899–1901. [CrossRef]
- Nitsche, M.A., et al., Modulation of cortical excitability by weak direct current stimulation--technical, safety and functional aspects. Suppl Clin Neurophysiol. 2003, 56, 255–276. [CrossRef]
- Liebetanz, D., et al., Pharmacological approach to the mechanisms of transcranial DC-stimulation-induced after-effects of human motor cortex excitability. Brain. 2002, 125 Pt 10, 2238–2247. [CrossRef]
- Tecchio, F., et al., Anodal transcranial direct current stimulation enhances procedural consolidation. J Neurophysiol. 2010, 104, 1134–1140. [CrossRef]
- Nitsche, M.A., et al., Facilitation of implicit motor learning by weak transcranial direct current stimulation of the primary motor cortex in the human. Journal of Cognitive Neuroscience. 2003, 15, 619–619. [CrossRef]
- Cuypers, K., et al., Is Motor Learning Mediated by tDCS Intensity? PLoS ONE 2013, 8, e67344. [CrossRef]
- Stagg, C.J., et al., Polarity and timing-dependent effects of transcranial direct current stimulation in explicit motor learning. Neuropsychologia. 2011, 49, 800–804. [CrossRef]
- Kantak, S.S., C.K. Mummidisetty, and J.W. Stinear, Primary motor and premotor cortex in implicit sequence learning - Evidence for competition between implicit and explicit human motor memory systems. European Journal of Neuroscience. 2012, 36, 2710–2715. [CrossRef]
- Schambra, H.M., et al., Probing for hemispheric specialization for motor skill learning: a transcranial direct current stimulation study. J Neurophysiol. 2011, 106, 652–661. [CrossRef]
- Saucedo Marquez, C.M., et al., Task-specific effect of transcranial direct current stimulation on motor learning. Front Hum Neurosci. 2013, 7, 333. [CrossRef]
- Reis, J., et al., Noninvasive cortical stimulation enhances motor skill acquisition over multiple days through an effect on consolidation. Proc Natl Acad Sci U S A. 2009, 106, 1590–1595. [CrossRef]
- Parlow, S.E. and M. Kinsbourne, Asymmetrical transfer of training between hands: implications for interhemispheric communication in normal brain. Brain Cogn. 1989, 11, 98–113. [CrossRef]
- Japikse, K.C., et al., Intermanual transfer of procedural learning after extended practice of probabilistic sequences. Exp Brain Res. 2003, 148, 38–49. [CrossRef]
- Perez, M., et al., Neural substrates of intermanual transfer of a newly acquired motor skill. Current Biology. 2007, 17, 1896–1902. [CrossRef]
- Calford, M.B. and R. Tweedale, Interhemispheric transfer of plasticity in the cerebral cortex. Science. 1990, 249, 805–807. [CrossRef]
- Gordon, A.M., H. Forssberg, and N. Iwasaki, Formation and lateralization of internal representations underlying motor commands during precision grip. Neuropsychologia. 1994, 32, 555–568. [CrossRef]
- Sainburg, R.L. and J. Wang, Interlimb transfer of visuomotor rotations: independence of direction and final position information. Experimental brain research. 2002, 145, 437–447. [CrossRef]
- Liang, N., et al., Effects of intermanual transfer induced by repetitive precision grip on input–output properties of untrained contralateral limb muscles. Experimental brain research. 2007, 182, 459–467. [CrossRef]
- Criscimagna-Hemminger, S.E., et al., Learned dynamics of reaching movements generalize from dominant to nondominant arm. Journal of neurophysiology. 2003, 89, 168–176. [CrossRef]
- Almeida, R. and M. Stetter, Modeling the link between functional imaging and neuronal activity: synaptic metabolic demand and spike rates. Neuroimage. 2002, 17, 1065–1079. [CrossRef]
- Tinazzi, M. and G. Zanette, Modulation of ipsilateral motor cortex in man during unimanual finger movements of different complexities. Neuroscience letters. 1998, 244, 121–124. [CrossRef]
- Bloom, J.S. and G.W. Hynd, The role of the corpus callosum in interhemispheric transfer of information: excitation or inhibition? Neuropsychology review. 2005, 15, 59–71. [CrossRef]
- Daselaar, S.M., et al., Similar network activated by young and old adults during the acquisition of a motor sequence. Neurobiology of aging. 2003, 24, 1013–1019. [CrossRef]
- Bischoff-Grethe, A., et al., Neural substrates of response-based sequence learning using fMRI. Journal of cognitive neuroscience. 2004, 16, 127–138. [CrossRef]
- Hashemirad, F., et al., Single-session anodal tDCS with small-size stimulating electrodes over frontoparietal superficial sites does not affect motor sequence learning. Frontiers in human neuroscience. 2017, 11, 153. [CrossRef]
- Koechlin, E., et al., The role of the anterior prefrontal cortex in human cognition. Nature. 1999, 399, 148–151. [CrossRef]
- Poreisz, C., et al., Safety aspects of transcranial direct current stimulation concerning healthy subjects and patients. Brain research bulletin. 2007, 72, 208–214. [CrossRef]
- Waters-Metenier, S., et al., Bihemispheric transcranial direct current stimulation enhances effector-independent representations of motor synergy and sequence learning. Journal of Neuroscience. 2014, 34, 1037–1050. [CrossRef]
- Horvath, J.C., O. Carter, and J.D. Forte, No significant effect of transcranial direct current stimulation (tDCS) found on simple motor reaction time comparing 15 different simulation protocols. Neuropsychologi. 2016, 91, 544–552. [CrossRef]
- Nitsche, M.A., et al., Shaping the effects of transcranial direct current stimulation of the human motor cortex. Journal of neurophysiology. 2007, 97, 3109–3117. [CrossRef]
- Boros, K., et al., Premotor transcranial direct current stimulation (tDCS) affects primary motor excitability in humans. European Journal of Neuroscience. 2008, 27, 1292–1300. [CrossRef]
- Elbert, T., et al., The influence of low-level transcortical DC-currents on response speed in humans. International Journal of Neuroscience. 1981, 14, 101–114. [CrossRef]
- Marshall, L., et al., Bifrontal transcranial direct current stimulation slows reaction time in a working memory task. BMC neuroscience. 2005, 6, 1–7. [CrossRef]
- Friehs, M.A. and C. Frings, Pimping inhibition: Anodal tDCS enhances stop-signal reaction time. Journal of Experimental Psychology: Human Perception and Performance. 2018, 44, 1933.
- Krause, V., et al., 1 Hz rTMS of the left posterior parietal cortex (PPC) modifies sensorimotor timing. Neuropsychologia. 2012, 50, 3729–3735. [CrossRef]
- Heinen, K., et al., Cathodal transcranial direct current stimulation over posterior parietal cortex enhances distinct aspects of visual working memory. Neuropsychologia. 2016, 87, 35–42. [CrossRef]
- Keitel, A., et al., Anodal transcranial direct current stimulation (tDCS) over the right primary motor cortex (M1) impairs implicit motor sequence learning of the ipsilateral hand. Frontiers in Human Neuroscience. 2018, 12, 289. [CrossRef]
- Doyon, J. and H. Benali, Reorganization and plasticity in the adult brain during learning of motor skills. Current opinion in neurobiology. 2005, 15, 161–167. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



