
Submitted:
06 March 2024
Posted:
07 March 2024
You are already at the latest version
Abstract
Barcoding of DNA is a species identification technique that utilizes a small DNA fragment from a specific gene or genes known as "sequencing." The idea behind DNA barcoding is that a single sequence can be used to specifically identify an organism to species by comparison with a reference library of these DNA sections. Herbal adulteration can be monitored using DNA barcodes that utilize medicinal plants such as Atropa belladonna. Because it is challenging to distinguish between species solely based on morphology, other plants belonging to the same genus are replacing it in trade. Such replacement with plants not utilized for medical purposes could have negative side effects. Also, Atropa belladonna's positive effects can be completely lost. Taxonomists also have trouble distinguishing the genuine Atropa belladonna plant from cultivated items. As a result, DNA barcoding can now be used to overcome this challenge. The development of DNA barcodes may help with trade control, which protects and conserves endangered species from export outside of the country due to their economic and biological significance. In this review, we discuss the DNA barcoding of Atropa belladonna plant in the medicinal plants field.
Keywords:
DNA
; Atropa belladonna
; Barcoding
; Adulteration
; Sequencing.
Introduction
According to David O. Kennedy (2014) [1], the Solanaceae family of nightshades includes a poisonous perennial herbaceous plant known as Atropa belladonna, also known as deadly nightshade. It is indigenous to Western Asia, Europe, and North Africa. The plant’s range extends from Ukraine and eastern Gilan in Iran to the United Kingdom in the west, including various regions of America and Canada, where it has also become naturalized or introduced [1,2,3,4]. The foliage and berries of Atropa belladonna are highly hazardous when consumed due to the presence of tropane alkaloids [1,2,3,4]. These alkaloids, such as atropine, scopolamine, and hyoscyamine, can cause delirium and hallucinations [1,2]. They also serve as pharmacological anticholinergics [4]. Tropane alkaloids are commonly found in the Solanoideae subfamily of the Solanaceae plant family, which includes the New World tribe Datureae, as well as the tribes of the Old World Hyoscyameae and Mandragoreae, including the Atropa genus [1].
The effects of Atropa belladonna can be unpredictable [2]. Pilocarpine or physostigmine are commonly used as remedies for belladonna toxicity [5]. Atropa belladonna is a branching herbaceous perennial hemicryptophyte that often grows from rhizomes as a subshrub. It can reach a height of 1.5 meters (5 ft) and has ovate-shaped leaves that can grow up to 7 inches long [1]. The bell-like blossoms are drab purple with a base that leans yellow-green and emit a weak aroma. The fruits are berries with a diameter of about 1.5 cm (0.6 in). They start off green and turn lustrous black when fully ripe. Despite their poisonous alkaloid content, animals are attracted to the tasty berries and help disperse the seeds through their droppings [6]. Sometimes, people confuse A. belladonna with the less poisonous Solanum nigrum (black nightshade), which is related to the Solanaceae family but belongs to a separate genus [7].
A comparison between the two types of nightshade fruit reveals that the berries of black nightshade are round, dull-looking, and grow in clusters. In contrast, the berries of Atropa belladonna are glossier, twice as big, a little flattened, and borne singly. Unlike other nightshades, black nightshade flowers have white, star-shaped petals with a central cone of yellow anthers [8].
Due to the difficulties of distinguishing between species based only on morphology, other plants related to this genus are utilized in commerce in place of the traditional medicine herb Atropa belladonna. Such substitution with a plant not utilized for medical purposes could have negative repercussions. Atropa belladonna’s positive effects can also be completely lost. Taxonomists also have trouble distinguishing the genuine Atropa belladonna plant from cultivated items. As a result, DNA barcoding can now be used to overcome this challenge. The development of DNA barcodes may help with trade control, which protects and conserves endangered species like Atropa belladonna from export outside of the country due to its economic and biological significance. Even a non-specialist can use the DNA barcoding approach to identify particular species because it creates data by sequencing DNA from an organism’s genetic regions. An appropriate DNA barcode should be chosen to use this identifying technique. DNA barcodes are short sections of the gene sequence that change quickly for species differentiation. Species mixtures that work well for the trade of unprocessed drugs can be evaluated by DNA barcoding, as demonstrated by Nithaniyal et al. (2017) [9]. This demonstrates how DNA barcoding is used to track the adulteration of herbal products. The current investigation will be carried out using several potential existing barcoding options to create DNA barcodes specifically for the Atropa belladonna species. For plants, there isn’t a single DNA barcode identifier that is universal. The CBOL Consortium for the Barcoding of Life has proposed the gene ribosomal RNA maturase K (matK) of the chloroplast and the large subunit coding region RuBisCO (rbcL) as plant DNA barcode primary markers. However, certain plant species cannot be distinguished with 100% accuracy using those locations. The non-coding marker psbA-trnH in the chloroplast was suggested by CBOL as a secondary marker because it was shown to distinguish species in the family Solanaceae. Additionally, for extensive DNA barcoding, the adoption of the region marker of internal transcribed spacer (ITS2) was advised [10]. Thus, in this overview, the literature is examined on the advantages and disadvantages of DNA barcoding in studying plants with medicinal properties, with a focus on the Atropa belladonna plant.
1. An overview of Atropa belladonna’s
1.1. Genus Atropa belladonna (Table 1)
Although Atropa belladonna was introduced and grown outside of its original area, its origin can be traced back to eastern, central, and southern Eastern Europe, as well as Turkey, the Caucasus, Iran, and North Africa. It was mentioned in the Skne Flora 1870 as being grown in Malmö apothecary gardens in southern Sweden [11].
In Britain, Atropa belladonna only grows naturally on tainted ground, calcareous soils, field borders, open forests, and hedgerows. As an alien species and often a relic from its previous use, it is more commonly found as a medicinal herb. According to the “Online Flora Atlas of Britain and Ireland,” birds are the primary means of seed dispersal [11].
In some North American regions, Atropa belladonna has become naturalized and can frequently be found in damp, shaded areas with limestone-rich soils. The Plants Profile for Atropa belladonna describes it as a weed that thrives in disturbed soils around the world [12]. Table 2 shows the various names of Atropa belladonna.
1.2. Atropa belladonna Morphology:
The species Atropa belladonna belongs to the Solanaceae family and thrives in uncultivated and desolate regions. It is widespread throughout the Himalayas, Western Europe, and the Mediterranean, including Greece. According to Lee (2007) [13], it has also made its way to Northern America. The name “Atropa belladonna” is derived from the Greek word “Atropos,” which refers to one of the three mythological Fates, and “belladonna” is an Italian translation meaning “beautiful lady.” The plant acquired this name because Venetian women used Atropa belladonna for cosmetic purposes, as it causes dilation of the pupils (mydriasis). It is also commonly known as deadly nightshade. In dry quarries and plains, Atropa belladonna grows as a perennial shrub. It produces greenish-purple flowers, spherical leaves, and sweet purple-black berries. It can reach a height of 1.5 m [13].
The alkaloids scopolamine, atropine, and hyoscine are present in all parts of the plant, which gives it its toxic and hallucinatory properties. According to Zárate, el Jaber-Vazdekis, Medina, and Ravelo (2006) [14], the highest concentration of alkaloids is found in fully ripe fruits and green foliage. Atropine, primarily found in ripe fruits, is the most abundant alkaloid, constituting approximately 98% of the alkaloid content. It is estimated that the plant’s fruits contain around 2 mg of atropine, as shown in Figure 1.
1.3. Medicinal Uses of Atropa belladonna:
Belladonna is known for its extreme toxicity. It contains over 20 distinct anticholinergic alkaloids that inhibit the function of acetylcholine in the body. One of the main active molecules is hyoscyamine, which is transformed into a combination known as atropine [15,16]. While cholinergic activity is generally beneficial in the body, inhibiting cholinergic action can be helpful in certain conditions, particularly respiratory problems. Belladonna has been used to alleviate headaches, irritable bowel syndrome, and menopausal symptoms [17,18].
The alkaloids in Belladonna, its major active ingredients, serve as deterrents to prevent animals from consuming the plant. The roots contain approximately 0.7% alkaloids, while the leaves have about 0.04% alkaloids. Hyoscyamines, scopolamine, atropine, apoatropine, cuscohygrine, and aposcopolamine are among the alkaloids found in varying concentrations in the leaves and roots [19,20]. The seeds of Belladonna contain around 0.06% alkaloids, with atropine and scopolamine being the most abundant [20,21]. Each mature berry of Belladonna contains approximately 2 mg of alkaloids, with 98% being atropine [22].
Ingesting two to five berries or even a single leaf can be lethal. Most cases of poisoning occur due to the mistaken consumption of the berries, often confused with blueberries, or intentional self-harm. Unintentional poisonings have also been linked to extracts of Belladonna [23]. Due to its high toxicity, even a single leaf or a small number of berries can be fatal, and the resemblance of the berries to other fruits can lead to confusion [23].
Organophosphates are highly toxic insecticides that inhibit the acetylcholinesterase enzyme responsible for degrading acetylcholine, leading to its accumulation in the body. In cases of poisoning with organophosphates, atropine, derived from Belladonna, is commonly used to alleviate the overstimulation of cholinergic activity. Anisodamine, a less potent alkaloid, has been explored as a potentially successful alternative to atropine with lower toxicity [24].
The total extract of alkaloids, including hyoscyamine and atropine from Belladonna, has been found to reduce mortality rates in dogs and rats poisoned with the insecticide parathion [25]. In rats, a commercial extract of belladonna alkaloids, combined with ergotamine, enabled survival from otherwise fatal scorpion toxin injections [26]. In two separate modern trials involving nearly 150 individuals with irritable bowel syndrome (IBS), a 10 mg belladonna extract drug combined with 50 mg of opium papaverine alkaloid reduced discomfort, cramps, and bowel movements. However, two other commercial drugs were found to be more effective [27,28]. A study from 1979 on 12 individuals discovered that scopolamine, a belladonna alkaloid (10 mg of hyoscine butylbromide taken four times daily), provided relief for irritable bowel syndrome, particularly when combined with lorazepam and ispaghula husk laxative [29]. In conventional medicine, additional medications are often used to treat the symptoms of irritable bowel syndrome, as it is a complex disease that requires a comprehensive approach addressing various underlying imbalances [30].
In two clinical trials involving over 100 menopausal women, a 0.2 mg alkaloids drug called Bellergal was found to relieve symptoms. However, after 8 weeks, the effects of one of the trials were no longer distinct from those of the placebo [31]. Bellergal was also used to treat hot flashes among 31 postmenopausal breast cancer survivors in an observational study [32]. In three clinical studies involving more than 200 individuals undergoing prostate and urinary procedures, opium suppositories and Belladonna were found to alleviate pain, urgency of urine, and reduce the need for painkillers. However, in two clinical trials involving over 100 women, they were unable to reduce pain from bladder injections or vaginal surgery [33].
Belladonna alkaloids have been found to minimize airway obstruction by inhibiting bronchial constriction. In two investigations involving over 100 children, oral tinctures of belladonna (providing 0.01 mg per kg of atropine 1-2 times per day for 7 days) improved breathing and unblocked airways [34,35]. However, in the case of chronic obstructive pulmonary disease (COPD), obstructed airways are typically treated in conjunction with other medications, including beta-agonists and anticholinergics [36]. In an earlier clinical experiment involving 55 individuals with regular headaches, a combination of 0.2 mg belladonna alkaloids, phenobarbital, and ergotamine taken twice daily for four weeks alleviated discomfort and reduced the need for pain medication. A similar combination, known as Bellergal, achieved “good results” in reducing headaches in 73% of individuals in a case series from the 1950s [37].
A previous clinical study on 25 women taking Bellergal, which contains belladonna alkaloids at a dose of 0.2 mg three times daily for 10 days before menstruation, demonstrated alleviation of symptoms [38]. In two therapeutic trials, the alkaloid atropine derived from Belladonna was found to reduce drooling in seven adults and 19 disabled children. In a trial involving 140 children, it also alleviated drooling caused by the sedative ketamine, but this effect was not observed in an observational study of 164 individuals [39].
Pregnant women often experience increased salivation, which can be associated with nausea. Belladonna, by inhibiting the rest-and-digest function of acetylcholine, reduces salivation. In two pregnant women, the alkaloids of belladonna (administered four times daily for 5 days) in combination with phenothiazine suppositories reduced nausea and excessive drooling [40].
In an earlier clinical trial involving 75 individuals with digestive ailments, a dose of 0.1 mg of Bellergal taken four times daily for 4 weeks effectively reduced anxiety, surpassing the effects of the sedative Librium (chlordiazepoxide) [41]. However, in another earlier clinical trial with almost 1,100 participants, both 20 drops of oral tincture or atropine at a dose of 0.4 mg were unable to alleviate anxiety induced by a contrast agent used for evaluating the urinary tract [42].
1.5. Composition and Properties of Atropa belladonna Species
Chemical Composition
The following are the primary components of belladonna: Atropine (C17H23NO3) [43]. The main component of belladonna root is hyoscyamine (C17H23NO3). Heat converts it to atropine, and this conversion also occurs naturally within the plant [43]. Atropamine (C17H21NO2), which is the same as apoatropine, was produced by Pesci in 1882 through the reaction of nitric acid on atropine [43,44]. The action of hydrochloric acid (HCl) or potassium hydroxide (KOH) converts atropamine to belladonine (C17H21NO2) [43]. Belladonine, an amorphous alkaloid, is obtained by heating hyoscyamine or atropine at temperatures ranging from 120°C to 130°C for an extended period [43]. By progressively increasing the transformation and temperature, the following compounds are obtained in the listed order: (1) hyoscyamine, (2) atropine, (3) atropamine, and (4) belladonine [43].
Hyoscine (C17H17NO4), also known as scopolamine, was first identified by Ladenburg in Hyoscyamus niger. While it is found in minor amounts in the roots of belladonna, Geiger and Hesse in 1833 demonstrated its similarity to the scopolamine alkaloid isolated from the root of Scopolia atropoides. In 1892, Schmidt discovered the formula C17H21NO4 for hyoscine [43].
Previous studies have revealed additional components present in belladonna, including chrysatropic acid (Kunz), which exhibits a greenish-blue fluorescence in concentrated solutions, as well as red-coloring atrosin primarily found in the root. Other components include malates, herb succinic acid, oxalates mixed with potassium, magnesium, and sodium, gum, asparagine, salts, chlorophyll, leaf wax, albumin, and starch [43], as shown in Figure 2.
Species Related to Atropa belladonna and Adulterants
Scopolia japonica, also known as Maximowicz (Japanese belladonna), is a rhizome measuring 5-10 dm in length and 1 dm in diameter. It has a cylindrical shape, is slightly compressed, and rarely branches or forms knots. The upper surface of the rhizome features round, slightly alternating stem scars. It has a brown external appearance, a paler interior speckled with numerous white spots, and emits a musky, narcotic odor [45].
Scopolia is an interesting genus as it bridges the gap between the genera Hyoscyamus and Atropa. Its rhizome contains unique, crystal-like components similar to Atropa, although they are less prominent. The bark and fibrovascular bundles in Scopolia are not as abundant or large, and the starch grains are smaller [45].
The anatomy of these two plants is very similar, although most of the significant characteristics are less apparent in belladonna. Similar to Japanese belladonna, Belladonna scopolia refers to a rhizome that can be distinguished from true belladonna, which has a root structure. It is important to note that this medicine should not be used if a product claims to represent a Pharmacopoeia that specifically focuses on the use of authentic belladonna. According to London drug merchants Messrs. W. H. Cole & Co., manufacturers of atropine often regard Scopolia carniolica as useless [45].
In Europe, Medicago sativa (Medicago) has been identified as an adulterant, characterized by thinner bark, a solid crown, and a stiff, woody meditullium with numerous small medullary rays. Althaea officinalis (marshmallow) is also purportedly employed as an adulterant for belladonna. It resembles an uncut, young root and can be identified by its branching wood, mucilage, and numerous bast fibers. Holmes reports that up to 50% of belladonna root may consist of marshmallow root. Early reports indicate that Madridian herbalists were selling a plant as belladonna that did not belong to the Solanaceae family [45].
The roots of Lappa (Arctium lappa) may be combined with belladonna. They can be distinguished by their peculiar pith, prominent branching bark, and the fact that they turn yellow with iodine and blue with ferric chloride. Henbane, foxglove, and mullein leaves are also occasionally reported as admixtures. Both Scopolia species, Hyoscyamus Scopolia (Scopolia carniolica) and Scopolia japonica Maximowicz, contain belladonna’s main ingredients, such as hyoscine, hyoscyamine, fluorescent scopoletin, and choline. They have been used in the country to produce belladonna and atropine plasters [45].
According to E. Schmidt, these plants contain hyoscine and scopolene (scopolamine) (C17H21NO4), which are mentioned as key ingredients of belladonna. Scopolene boiling in baryta water yields methyl-aesculin, resulting in the formation of scopoline (C8H13NO2) and atropic acid (C9H8O2). Scopolamine belongs to the scopolines, which are formed when the alkaloid scopoline interacts with organic acids. Scopoline is a more potent mydriatic than atropine, causing earlier and longer-lasting dilation of the pupils. According to Sir Dyce Duckworth, it does not have internal effects on the pupil and does not cause throat dryness as belladonna does. Dr. Gordon Sharp suggests that the effects of duboisine, hyoscine, daturine, and scopolamine differ only slightly from those of atropine [45,46].
2. Detection of Medicinal Plant DNA Barcoding
2.1. Definition and Goals behind DNA Barcoding
DNA barcode is a species-specific short sequence of genes in the genome [47]. It is a valuable method for the identification, categorization, and classification of species by tracking a standardized short DNA sequence in a popular gene. Polymerase chain reaction (PCR) is used to amplify a highly variable region in the nuclear, chloroplast, or mitochondrial genomes’ DNA barcode region, such as mitochondrial DNA, chloroplast DNA, nuclear DNA (ITS), matK, trnL-F, psbA, rbcL, trnH, and psbK [47]. These regions are commonly utilized for DNA barcoding, with COI (Cytochrome C oxidase subunit l mtDNA) proposed as the worldwide barcode [47]. The usage of short-sequenced DNA for species identification was proposed in 2003, and COI has been proven to discriminate over 90% of species in most animal groupings [47]. The Consortium for the Barcode of Life (CBOL) plays a crucial role in organizing global efforts for DNA barcoding [47].
Jaakola et al. (2007) developed highly-resolution melting procedures using plastid and ITS DNA to combat market adulteration of bilberries [48]. Jian et al. (2012) discovered residues in roasted tea’s granules using DNA barcoding, highlighting its application in international trade [49]. Therodoridis et al. (2014) utilized DNA barcodes to identify Lamiaceae species epidemic in Turkey and Greece, specifically psbA-trnH, matK, and rbcL markers [50].
DNA barcoding has been applied in various studies, including the identification of toxic plant components by Bruni et al. (2010) and the determination of species in cooked plant materials in spices by Moreoer Mattia et al. (2011) [51,52]. Cornara et al. (2013) employed DNA barcoding to identify components in spice mixes for sale [53]. Several studies, such as those by Koolman et al. (2013), Newmaster et al. (2013), and Wallace et al. (2012), demonstrated the effective application of DNA barcoding in detecting adulteration in raw medicinal plant products and natural health products [54,55,56].
2.2. Unique Plant DNA Barcoding Requirement
According to Lee (2016) [57], the Barcoding of Plant has made slower progress compared to other fields. The mitochondrial DNA (mtDNA) in plants was found to evolve too slowly to distinguish between different species. Kress et al. (2005) [58] explained that the limited differentiation and frequent changes in plant mitochondrial genome organization hinder the use of mitochondrial genes for species-level classification. Erickson and Kress (2008) [59] proposed three prerequisites for a DNA barcode in the genomic region: a gene area with sufficient variation, global Primers with conserved sides, and a compact size for easy extraction and amplification.
A fixed barcoding region meeting all these requirements has not been identified, and multiple combined regions may be necessary for effective plant species discrimination (Jian, 2014) [60]. Plastid genes, including coding genes, introns, exons, and intergenic spacers, are reliable options for plant DNA barcodes due to their variability, conserved configuration, and high genotyping rates. The core barcodes consist of the coding gene Ribosomal RNA maturase K (matK) and the big subunit coding region of RuBisCO (rbcL). The Consortium for the Barcode of Life (CBOL, 2009) [61] recently selected the combination of matK and rbcL as the ideal plant barcode, although it has a discriminating efficiency of only 72%. Therefore, taxonomists suggest using a multi-loci approach to differentiate between plant species. In addition to the core barcode markers, the psbA-trnH spacer and ITS regions are widely used as plant fingerprints for the majority of plant species (Kress and Erickson, 2008; Chen et al., 2010) [59,62].
Figure 4 illustrates the DNA barcode regions in the plastid genome. According to CBOL, the rbcL and matK genes should be supplemented by other regions to distinguish between closely related species. The psbA-trnH region continues to provide new insights. Other plastid loci, such as trnL-F, rpoB2, rpoCl, and others, have been sequenced for phylogenetic purposes in plant systematics, each with varying levels of accuracy (Kress et al., 2005) [58], Figure 3 and Figure 4.
2.3. matK Locus
The rapidly changing encoding gene ribosomal RNA maturase K (matK) [63] plays a significant role in plant phylogenetics. Another region of interest is the intron of the chloroplast gene trnK, which contains a region spanning 780-1480 base pairs (bp) (Figure 5). This region has been utilized to study nucleotide variations and frequency changes in gene sequences for generating phylogenies at the divisional level up to the tribe level [64], Figure 5.
MatK, as described by Windelspecht (2007) [65], exhibits a rapid rate of evolution, has an appropriate length, demonstrates clear interspecific variation, and displays a low rate of transformation. However, the amplification of matK using currently available primer sets can be challenging. The Working Group for CBOL Plant (2009) [66] found that amplifying gymnosperms DNA using two primers alone achieved a successful rate of over 90%. Nonetheless, when multiple primer sets were used, the success rate was limited to 83% in gymnosperms and significantly lower at 10% in cryptogams. Different primer pairs were required for each taxonomic category [65]. Lahaye et al. (2008) [67] successfully amplified the matK gene in gymnosperms using specific primers. Another challenge lies in discerning the variation rates among different taxonomic groups. MatK can distinguish over 90% of Solanaceae species but only 49% of Nutmeg species (Kress & Erickson, 2007) [68]. In an attempt to identify 92 organisms from 32 taxonomic groups using the matK barcode, Fazekas et al. (2008) [69] achieved a success rate of only 56%. These findings indicate that a standalone matK barcode may not serve as a universally applicable barcoding pattern.
2.4. rbcL Locus
The coding region of RuBisCO (rbcL) in the plastid genome is approximately 600-700 base pairs (bp) long. Due to its ability to detect evolutionary links at the genus level, the rbcL region holds promise for plant barcoding, as highlighted by Windelspecht (2007) [70], Figure 6.
The locus rbcL in most terrestrial plants can be easily amplified, sequenced, and utilized as a valuable DNA barcoding region for species and family levels. However, Kress et al. (2005) discovered a gradual change pattern in rbcL, indicating a low level of divergence compared to chloroplast genes in flowering plants, making it insufficient for species-level differentiation. Another limitation is its length, which requires four primers for double-stranded sequencing. Despite these constraints, the rbcL gene is still widely employed for plant barcoding due to the abundance of available data and easy retrieval of the complete gene sequence. Although rbcL alone may not possess all the ideal characteristics of a standalone coding region for sequencing, when combined with multiple chloroplast or nuclear markers, it can provide accurate identification (Renner, 1999) [71].
2.5. The trnH-psbA Locus
Many plants possess a non-coding region known as trnH-psbA, which encompasses tRNA-His and photosystem II protein D1. This region exhibits several properties that make it an ideal barcode candidate. The intron of the plastid DNA contains the trnH-psbA gene, which spans 850-1480 base pairs (bp) in length. The coding sequence within this region is evolutionarily conserved, allowing the development of widely usable primers that can amplify the majority of flowering plants using a single primer. The 3’ untranslated region of psbA and trnH, characterized by a high frequency of insertions and deletions, makes it a promising choice for barcoding flowering plants (Shaw et al., 2007) [72], Figure 7.
2.6. The ITS Locus
From the 16S to the 23S gene sequences, the internal transcribed spacer (ITS) region is often considered. However, CBOL (Consortium for the Barcode of Life) only recommends it as a supplementary locus and not as a fundamental barcode indicator. There are several reasons for this. First, the ITS locus poses challenges in terms of sequencing and amplification. Second, it exhibits a high degree of concerted evolution, which complicates its use for species differentiation. Lastly, the ITS locus is not exclusive to plants, as it is also present in fungi. This can lead to the amplification of fungal-contaminated sequences, potentially resulting in misinterpretation of the ITS sequence (Mishra et al., 2015) [73], Figure 8.
2.6. Combination of Barcodes for Plant Identification.
Besides intensive efforts, identifying standardized barcodes for plants akin to CO1 in mammals has proven impossible because of a lack of sufficient variety inside a single locus. In 74’s suggestion, the need for a multi-loci technique to achieve effectively differentiated species. When choosing a standard plant DNA barcode, several aspects must be considered. 75 and 76 propose criteria to examine when choosing for terrestrial plants’ DNA barcoding. These characteristics include the versatility of PCR enrichment for various purposes, the capacity to distinguish between different species, the range of classification orders and applications, the complementarity of genomic regions, the utilization of computational techniques, and the process of obtaining approval [74,75,76]. The barcode area with the greatest rate of restoration by PCR is universal PCR amplification [76]. The power of species differentiation has numerous dimensions. Some Unresolvable species use any of the potential DNA barcode markers, although reference of markers can enhance recovery. The use of several barcode markers is constrained of angiosperms when addressing the spectrum of taxonomic variety and applicability [76]. This rule agreement promotes barcoding a plant with another two marker regions in terms of locus complementation [75]. The complementarity of loci has significantly improved the results [75]. Considering that DNA barcodes function by comparing a query’s genome sequences to known sequences from a voucher sample, bioinformatics is essential [77]. As a result, an independent access barcode catalog is a reference point for the sample’s unknown DNA barcode that may be compared to a database-stored known sequence. CBOL advocated rbcL and matK as a standard barcode pattern since the rbcL area may be recovered immediately and the matK sequence has excellent differentiation potency [75]. Despite a greater identifying efficiency than other variations, the matK and rbcL amalgamation falls short of the intended objectives of a universal DNA barcoding. For starters, the matK+rbcL combo has a relatively low PCR efficiency. Second, discrimination capacity in animals is lower than COI [75]. Although combined barcodes are advocated because they have various benefits, the opposite argument for their use is also being made. According to 78, combination barcode improves over a single-locus marker analytical difficulties. Furthermore, in a single study, CBOL showed using seven distinct candidate locus combinations did not increase prejudice in the level of species capacity when compared to matK and rbcL [78]. According to certain scientists, disagreement between unrelated species and the chloroplast genotype tree may fail barcodes with multiple loci. They demonstrated the inability of such numerous regional barcodes to overcome the intrinsic flaws in plant DNA barcoding currently in use [79].
IV. Conclusion
These research outcomes demonstrated primers presently used in DNA barcoding, ITS-2, rbcL, matK, and psbA-trnH, my magnify Atropa belladonna DNA loci. DNA four sections were successfully sequenced and correctly linked to an identical genus-level plant, but not in the matK locus. The findings showed ITS-2 and rbcL barcode sections may to excel as DNA barcodes for the Atropa belladonna of the four loci studied. The recognition, preservation, and quality standards of priceless materials will be facilitated by these DNA barcodes for Atropa belladonna species of medicinal plants, especially from Saudi Arabia. Finally, when the benefits and drawbacks of barcoding become obvious, it is obvious that classification techniques that incorporate sequencing DNA will attain optimum effectiveness in identifying species. DNA barcoding allows us to accelerate taxonomists’ efforts in species identification. Notwithstanding the few limitations of the barcoding of DNA, the research presented here demonstrated the effectiveness of using barcoding areas in recognizing Atropa belladonna species from Saudi Arabia and Pollution is the contamination in the sector for herbal preparations. The research advises that efforts be going forward to produce genomic barcoding to supplement the barcode loci just utilized for much more precise barcoding findings.
Author Contributions
This study is a literature review and analysis conducted by Mohammad Odah.
Funding
This research did not receive any specific funding.
Institutional Review Board Statement
This research study was conducted following the ethical guidelines and regulations set forth by the Saudi Board or Ethics Committee, but due to the nature of the study being a literature review and analysis, formal ethics approval was not required.
Informed Consent Statement
Given that this study is a literature review and analysis, individual consent for publication is not applicable.
Data Availability Statement
The data analyzed in this study were obtained from publicly available sources, including academic journals, books, reports, and official websites. No specific datasets or materials were collected or utilized in this study.
Acknowledgments
We would like to express our heartfelt appreciation and gratitude to Prince Sattam bin Abdulaziz University for their unwavering support and encouragement throughout our research project. Without their support, this study would not have been possible. We would also like to extend our sincere thanks to the faculty members and research staff at Prince Sattam bin Abdulaziz University especially Prof. Farag Elessawy, Dr. Mohammad bin Shaie Al-Matrafi, and Dr. Mohammad Mahzari for their valuable insights, suggestions, and assistance during the study. Their inputs and guidance have been instrumental in shaping our research project. I’d really want also to thank the Deanship of Scientific Research at Prince Sattam Bin Abdulaziz University, Al-Kharj, Saudi Arabia, and the Genetics and Genomics Department at King Abdullah University of Science and Technology KAUST, Riyadh, Saudi Arabia, for their generous support and assistance.
Conflicts of Interest
The authors declare no competing interests that could potentially influence the interpretation or presentation of the research findings.
References
- Kennedy, D. O. (2014). Plants and the human brain. Oxford University Press.
- Blumenthal, M. , Goldberg, A., & Brinckmann, J. (Eds.). (2000). Herbal medicine: Expanded Commission E monographs. Integrative Medicine Communications.
- Frohne, D. , & Pfänder, H. J. (2005). A colour atlas of poisonous plants: A handbook for pharmacists, doctors, toxicologists, and biologists. Manson Publishing.
- Schep, L. J., Slaughter, R. J., & Vale, J. A. (2009). Pharmacological and toxicological properties of Atropa belladonna L. Journal of Pharmacy Practice and Research, 39(2), 145-147. [CrossRef]
- Arroyo, S. , & Lavoie, M. (2015). Plant toxins in poisonings and intoxications. In J. H. Duffield, G. P. Bailey, & T. S. Hovda (Eds.), Veterinary toxicology: Basic and clinical principles (3rd ed., pp. 457-476). Academic Press.
- Jones, J. R. (2005). Deadly nightshade. In Poisonous plants of Pennsylvania and their toxins (2nd ed., pp. 44-45). Pennsylvania Department of Agriculture.
- Peterson, L. (2020). Solanum nigrum L. In M. L. Gargiullo & B. Magnuson (Eds.), Flora of North America North of Mexico (Vol. 7, pp. 326-331). Oxford University Press.
- Martin, R. F. (2004). Atropa belladonna L. In G. W. Argus (Ed.), Flora of North America North of Mexico (Vol. 7, pp. 75-78). Oxford University Press.
- Nithaniyal, S., Vassou, S. L., Poovitha, S., & Jayabalan, M. (2017). DNA barcoding as a reliable method for the authentication of medicinal plants used in the preparation of herbal products. Journal of Herbal Medicine, 9, 45-51. [CrossRef]
- CBOL Plant Working Group. (2009). A DNA barcode for land plants. Proceedings of the National Academy of Sciences, 106(31), 12794-12797. [CrossRef]
- Svensson, B. M. , & Tyler, T. (2013). Flora of Skne: The vascular plants of Skne, Sweden (2nd ed.). Sknes Flora Frlag.
- USDA, NRCS. (2021). Plants Profile for Atropa belladonna L. Retrieved from https://plants.usda.gov/core/profile?
- Lee, T. (2007). Flora of China: Solanaceae. In Z. Y. Wu, P. H. Raven, & D. Y. Hong (Eds.), Flora of China (Vol. 17, pp. 287-292). Science Press/Missouri Botanical Garden Press.
- Zárate, R. , el Jaber-Vazdekis, N., Medina, M., & Ravelo, Á. G. (2006). Tropane alkaloid profile of Atropa belladonna L. at different growth stages. Journal of Agricultural and Food Chemistry, 54(6), 2048-2052. [CrossRef]
- Riddle, J. M. (2016). Atropa belladonna. In J. Riddle (Ed.), Dioscorides on pharmacy and medicine (pp. 201-203). University of Texas Press.
- Hahn, R. A. (2002). The nocebo phenomenon: Concept, evidence, and implications for public health. Preventive Medicine, 34(6), 639-648. [CrossRef]
- Ernst, E. , & Pittler, M. H. (2000). Efficacy of homeopathic arnica: A systematic review of placebo-controlled clinical trials. Archives of Surgery, 135(11), 1321-1326. [CrossRef]
- Ernst, E. , & Pittler, M. H. (2000). Efficacy of ginger for nausea and vomiting: A systematic review of randomized clinical trials. British Journal of Anaesthesia, 84(3), 367-371. [CrossRef]
- Adams, R. P. (2007). Identification of essential oil components by gas chromatography/mass spectrometry (4th ed.). Allured Publishing Corporation.
- Perry, L. M. , & Metzger, J. (1980). Medicinal plants of East and Southeast Asia: Attributed properties and uses. MIT Press.
- Bisset, N. G. (1994). Herbal drugs and phytopharmaceuticals (1st ed.). CRC Press.
- Rätsch, C. (2005). The encyclopedia of psychoactive plants: Ethnopharmacology and its applications (3rd ed.). Park Street Press.
- Langford, N. J., & Boobis, A. R. (2002). Belladonna poisoning. Toxicological Reviews, 21(2), 89-96. [CrossRef]
- Bismuth, C. , & Hall, A. H. (2009). Anticholinergic syndrome. In A. H. Hall, L. J. Rumack, & R. L. Clark (Eds.), Antidote for poisoning by organophosphorus pesticides (pp. 57-59). CRC Press.
- Gardner, J. R. (1965). The use of alkaloids of Belladonna to counteract the toxic effects of parathion and other organophosphorus insecticides. The American Journal of the Medical Sciences, 250(2), 173-178. [CrossRef]
- Tyler, V. E. , Brady, L. R., & Robbers, J. E. (1988). Pharmacognosy (9th ed.). Lea & Febiger.
- Kruis, W., Fric, P., Pokrotnieks, J., Lukás, M., Fixa, B., Kascák, M., ... & Schreiber, I. (2004). Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut, 53(11), 1617-1623. [CrossRef]
- Lichtenstein, G. R. , Hanauer, S. B., Sandborn, W. J., & Practice Parameters Committee of American College of Gastroenterology. (2009). Management of Crohn’s disease in adults. The American Journal of Gastroenterology, 104(2), 465-483. [CrossRef]
- Maxton, D.G.; Whorwell, P.J. The treatment of irritable bowel syndrome with lorazepam, hyoscine butylbromide, and ispaghula husk. Br. J. Clin. Pract. 1979, 33, 251–253. [Google Scholar] [CrossRef]
- Quigley, E. M. M. (2016). Gastrointestinal motility disorders in the elderly. Gastroenterology Clinics of North America, 45(3), 405-418. [CrossRef]
- Bachmann, G.A.; Schaefers, M.; Nilaver, G. A comparison of the effect of Bellergal, an extract of Belladonna, Hedera helix and Rhododendron ponticum, on menopausal symptoms. Minerva Ginecol. 1983, 35, 443–446. [Google Scholar]
- Stearns, V. , Beebe, K. L., Iyengar, M., Dube, E., Paroxetine Study Group, & Gomez, H. L. (2003). Paroxetine controlled release in the treatment of menopausal hot flashes: A randomized controlled trial. JAMA, 289(21), 2827-2834. [CrossRef]
- Wiffen, P. J. , McQuay, H. J., & Moore, R. A. (2017). Obstructed labour: Medical interventions for associated short-term complications in women undergoing caesarean section. Cochrane Database of Systematic Reviews, 2017(7), CD004749. [CrossRef]
- Min, S. , & Gallardo-Casero, J. M. (2013). The effects of Chinese herbs on bronchial asthma: A review of literature. Chinese Medicine, 4(3), 99-107. [CrossRef]
- Tabarki, B. , Al-Shafi, A., & Abu Laban, A. (2008). The impact of belladonna tincture as a pre-anesthetic medication on the pediatric patient with difficult airways. Journal of Clinical Anesthesia, 20(7), 522-526. [CrossRef]
- O’Donnell, D. E. , Aaron, S., Bourbeau, J., Hernandez, P., Marciniuk, D. D., & Balter, M. (2017). Canadian Thoracic Society recommendations for management of chronic obstructive pulmonary disease—2007 update. Canadian Respiratory Journal, 14(Suppl B), 5B-32B. [CrossRef]
- Frank, R.G.; Knoedler, C.H. The treatment of vascular headache: Clinical experiences with belladonna alkaloids-phenobarbital-ergotamine combinations. J. Kans. Med. Soc. 1957, 58, 243–246. [Google Scholar]
- Hernández-Muñoz, G. , Placencio-Hickok, V. R., Hernández-Muñoz, J. M., Contreras-Ochoa, C. O., & Del Valle-Martinez, L. M. (1990). The treatment of premenstrual syndrome (PMS) with a standardized mixture of Ginkgo biloba and Belladonna extracts. Gynecologic and Obstetric Investigation, 30(2), 113-117. [CrossRef]
- Lorton, D. , & Bellinger, D. L. (2016). Molecular mechanisms underlying β-adrenergic receptor-mediated cross-talk between sympathetic neurons and immune cells. International Journal of Molecular Sciences, 17(5), 638. [CrossRef]
- Hatton, F., & Apte, S. (2018). The pharmacological management of morning sickness in pregnancy. Australian Prescriber, 41(4), 112-117. [CrossRef]
- Demiris, G. , Tsatalou, E. G., & Schulpis, K. H. (1993). Anxiety in patients with digestive problems: Its prevalence and the effectiveness of a psychotropic treatment. Journal of Psychosomatic Research, 37(3), 301-306. [CrossRef]
- Zeman, J., Kvetina, J., & Pintova, J. (1970). Atropine and Librium in urological radiography. Journal of Urology, 103(5), 616-621. [CrossRef]
- Tschirch, A. , & Rohde, A. (1922). The constituents of the belladonna root. In A. Tschirch & A. Rohde (Eds.), The chemistry of plant products (Vol. 2, pp. 370-377). Macmillan Publishers.
- Pesci, A. About atropamine. Gazz. Chim. Ital. 1882, 12, 317–319. [Google Scholar]
- Anonymous. (n.d.). Scopolia japonica Maximowicz. In H. G. A. Engler & K. Prantl (Eds.), Die natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren Arten, insbesondere den Nutzpflanzen (Vol. 4, 2, pp. 277-278).
- Duckworth, D. (n.d.). On the mydriatic effects of scopoline, duboisine, and hyoscine, with special reference to the differential action of these drugs. British Medical Journal, 1(1439), 437-440. [CrossRef]
- Hebert, P. D. N., Cywinska, A., Ball, S. L., & deWaard, J. R. (2003). Biological identifications through DNA barcodes. Proceedings. Biological Sciences, 270(1512), 313-321. [CrossRef]
- Jaakola, L., Pirttilä, A. M., Halonen, M., & Hohtola, A. (2007). Isolation of high quality DNA for genetic studies of bilberry (Vaccinium myrtillus L.) and other Vaccinium species. Molecular Ecology Notes, 7(6), 1261-1264. [CrossRef]
- ian, H., Wang, J., Yu, J., & Chen, X. (2012). Identification of roasted tea varieties using DNA barcoding. Food Research International, 48(1), 491-496. [CrossRef]
- Theodoridis, S. , Stefanaki, A., Tezcan, M., Bozdogan, B., & Goudet, J. (2014). Systematic biology in Europe: The EMBO-sponsored European conference on the barcoding of arthropods. Ecological Research, 29(6), 1159-1162. [CrossRef]
- Bruni, I., De Mattia, F., Galimberti, A., Galasso, G., & Banfi, E. (2010). Identification of poisonous plants by DNA barcoding approach. International Journal of Legal Medicine, 124(6), 595-603. [CrossRef]
- De Mattia, F., Bruni, I., Galimberti, A., Cattaneo, F., Casiraghi, M., Labra, M., & Savadori, P. (2011). A comparative study of different DNA barcoding markers for the identification of some members of Lamiaceae. Food Research International, 44(3), 693-702. [CrossRef]
- Cornara, L., La Rocca, A., Marsili, S., & Mariotti, M. G. (2013). Traditional uses of plants in the Eastern Riviera (Liguria, Italy). Journal of Ethnopharmacology, 146(2), 659-668. [CrossRef]
- Kool, A., de Boer, H. J., Krüger, Å., Rydberg, A., Abbad, A., Björk, L., Martin, G., et al. (2013). Molecular identification of commercialized medicinal plants in southern Morocco. PLoS ONE, 8(6), e60677. [CrossRef]
- Newmaster, S. G. , Grguric, M., Shanmughanandhan, D., Ramalingam, S., & Ragupathy, S. (2013). DNA barcoding detects contamination and substitution in North American herbal products. BMC Medicine, 11, 222. [CrossRef]
- Wallace, L. J., Boilard, S. M., & Eagle, S. H. C. (2012). DNA barcodes for routine identification of medicinal plants in the Canadian marketplace. Genome, 55(3), 192-200. 5. [CrossRef]
- Lee, H. J. (2016). Barcoding of Plant (RbcL, MatK) and Animal (CO1, 16S) Genes for the Authentication of Traditional Korean Medicine. Genomics & Informatics, 14(4), 196-200. [CrossRef]
- Kress, W. J., Erickson, D. L., Jones, F. A., Swenson, N. G., Perez, R., Sanjur, O., & Bermingham, E. (2009). Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama. Proceedings of the National Academy of Sciences, 106(44), 18621-18626. [CrossRef]
- Erickson, D. L. , & Kress, W. J. (2008). Assessing phylogenetic relationships of the Orchidaceae with DNA-barcoding. Molecular Phylogenetics and Evolution, 44(2), 1032-1040. [CrossRef]
- Jian, H. , et al. (2014). Can DNA Barcoding of Plant Material in Holistic Approach Make a Routine Contribution for Adulteration Control of Chinese Medicines? BioMed Research International, 2014, 1-12. [CrossRef]
- CBOL Plant Working Group. (2009). A DNA barcode for land plants. Proceedings of the National Academy of Sciences, 106(31), 12794-12797. [CrossRef]
- Chen, S., Yao, H., Han, J., Liu, C., Song, J., Shi, L., & Zhu, Y. (2010). Validation of the ITS2 Region as a Novel DNA Barcode for Identifying Medicinal Plant Species. PLoS ONE, 5(1), e8613. [CrossRef]
- Kress, W. J. , Erickson, D. L., & Jones, F. A. (2009). DNA barcodes: Genes, genomics, and bioinformatics. Proceedings of the National Academy of Sciences, 106(Supplement 2), 19675-19676. [CrossRef]
- Chase, M. W. , Salamin, N., Wilkinson, M., Dunwell, J. M., Kesanakurthi, R. P., Haider, N., & Savolainen, V. (2005). Land plants and DNA barcodes: Short-term and long-term goals. Philosophical Transactions of the Royal Society B: Biological Sciences, 360(1462), 1889-1895. [CrossRef]
- Hollingsworth, P. M., Graham, S. W., & Little, D. P. (2011). Choosing and using a plant DNA barcode. PLoS ONE, 6(5), e19254. [CrossRef]
- Lahaye, R., Van der Bank, M., Bogarin, D., Warner, J., Pupulin, F., Gigot, G., & Savolainen, V. (2008). DNA barcoding the floras of biodiversity hotspots. Proceedings of the National Academy of Sciences, 105(8), 2923-2928. [CrossRef]
- Kress, W. J. , & Erickson, D. L. (2007). A two-locus global DNA barcode for land plants: The coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS ONE, 2(6), e508. [CrossRef]
- Fazekas, A. J. , Kesanakurti, P. R., Burgess, K. S., Percy, D. M., Graham, S. W., Barrett, S. C. H., & Husband, B. C. (2008). Are plant species inherently harder to discriminate than animal species using DNA barcoding markers? Molecular Ecology Resources, 9(s1), 130-139. [CrossRef]
- Renner, S. S. (1999). Circumscription and phylogeny of the Laurales: Evidence from molecular and morphological data. American Journal of Botany, 86(9), 1301-1315. [CrossRef]
- Shaw, J. , Lickey, E. B., Schilling, E. E., & Small, R. L. (2007). Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. American Journal of Botany, 94(3), 275-288. [CrossRef]
- Mishra, P. , Kumar, A., Nagireddy, A., Mani, D. N., Shukla, A. K., & Tiwari, R. (2015). Plant barcode loci as tools for species identification: A review. Biotechnology Advances, 33(5), 1892-1907. [CrossRef]
- Hollingsworth, P. M. , Forrest, L. L., Spouge, J. L., Hajibabaei, M., Ratnasingham, S., Van der Bank, M., Chase, M. W., & Cowan, R. S. (2009). A DNA barcode for land plants. Proceedings of the National Academy of Sciences, 106(31), 12794-12797. [CrossRef]
- Kress, W. J., Wurdack, K. J., Zimmer, E. A., Weigt, L. A., & Janzen, D. H. (2005). Use of DNA barcodes to identify flowering plants. Proceedings of the National Academy of Sciences, 102(23), 8369-8374. [CrossRef]
- CBOL Plant Working Group. (2009). A DNA barcode for land plants. Proceedings of the National Academy of Sciences, 106(31), 12794-12797. [CrossRef]
- Hebert, P. D. N. , Ratnasingham, S., & de Waard, J. R. (2003). Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proceedings. Biological Sciences, 270(Supplement 1), S96-S99. [CrossRef]
- Newmaster, S. G., Fazekas, A. J., Steeves, R. A., & Janovec, J. (2008). Testing candidate plant barcode regions in the Myristicaceae. Molecular Ecology Resources, 8(3), 480-490. [CrossRef]
- Chen, S., Yao, H., Han, J., Liu, C., Song, J., Shi, L., Zhu, Y., Ma, X., Gao, T., Pang, X., Luo, K., Li, Y., Li, X., Jia, X., Lin, Y., Leon, C. (2010). Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE, 5(1), e8613. [CrossRef]
- Raza, G.; Rahman, M.; Sajjad, Y.; He, S.; Yang, M.; Zhang, H.; Zhao, M.; Zhang, X. DNA barcoding: A molecular tool to identify economically important plant species. Plant Omics 2019, 12, 95–109. [Google Scholar]
- Kool, A., de Boer, H., Krüger, Å., Rydberg, A., Abbad, A., & Björk, L. (2012). Molecular identification of commercialized medicinal plants in southern Morocco. PLoS ONE, 7(6), e39459. [CrossRef]
Figure 1.
Atropa belladonna. Source: Britannica Encyclopedia.

Figure 2.
Chemical Composition of Atropa belladonna.
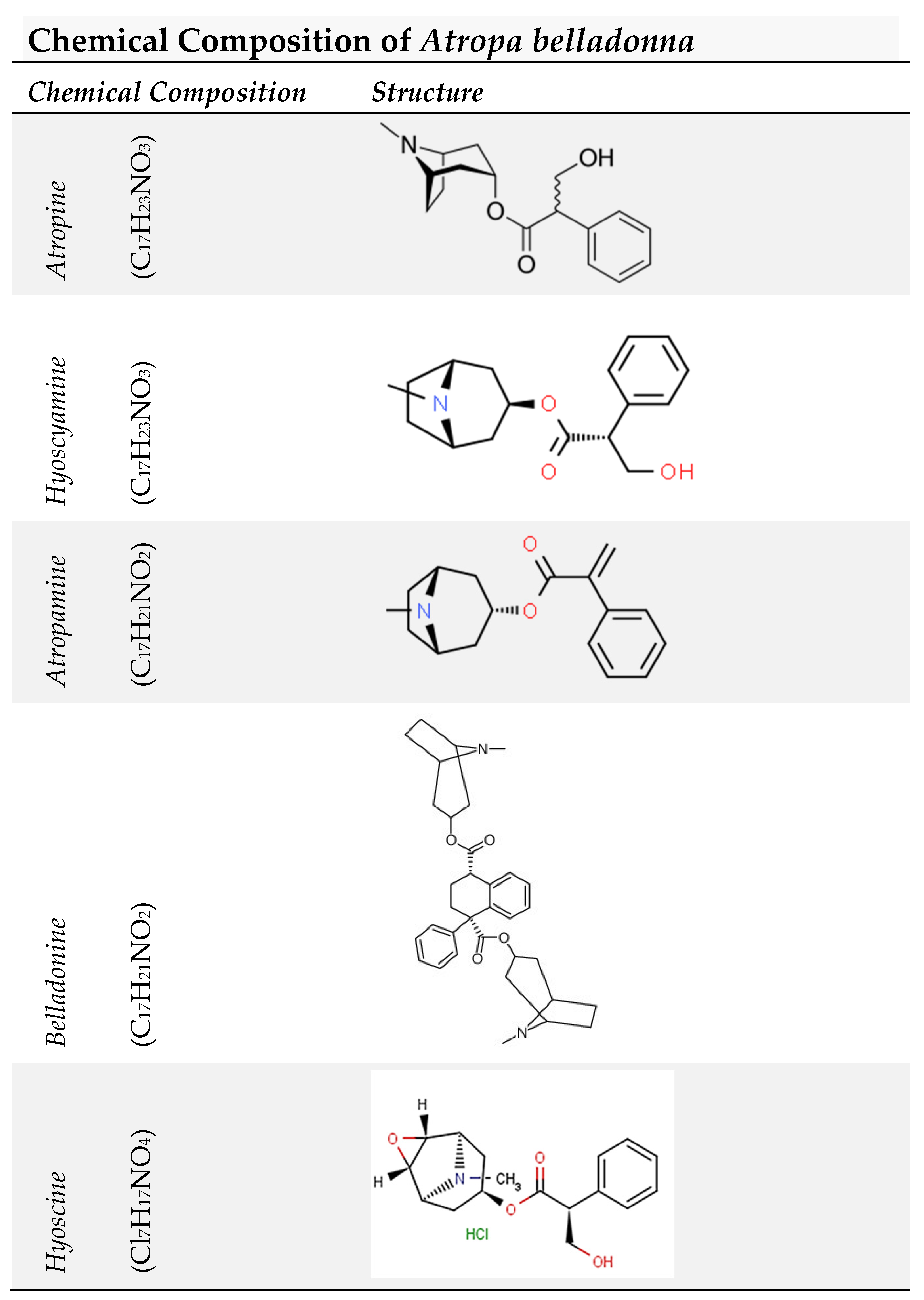
Figure 3.
Locations of Atropa belladonna DNA barcodes in the plastid genome (Source: Chen et al., 2010).
Figure 3.
Locations of Atropa belladonna DNA barcodes in the plastid genome (Source: Chen et al., 2010).
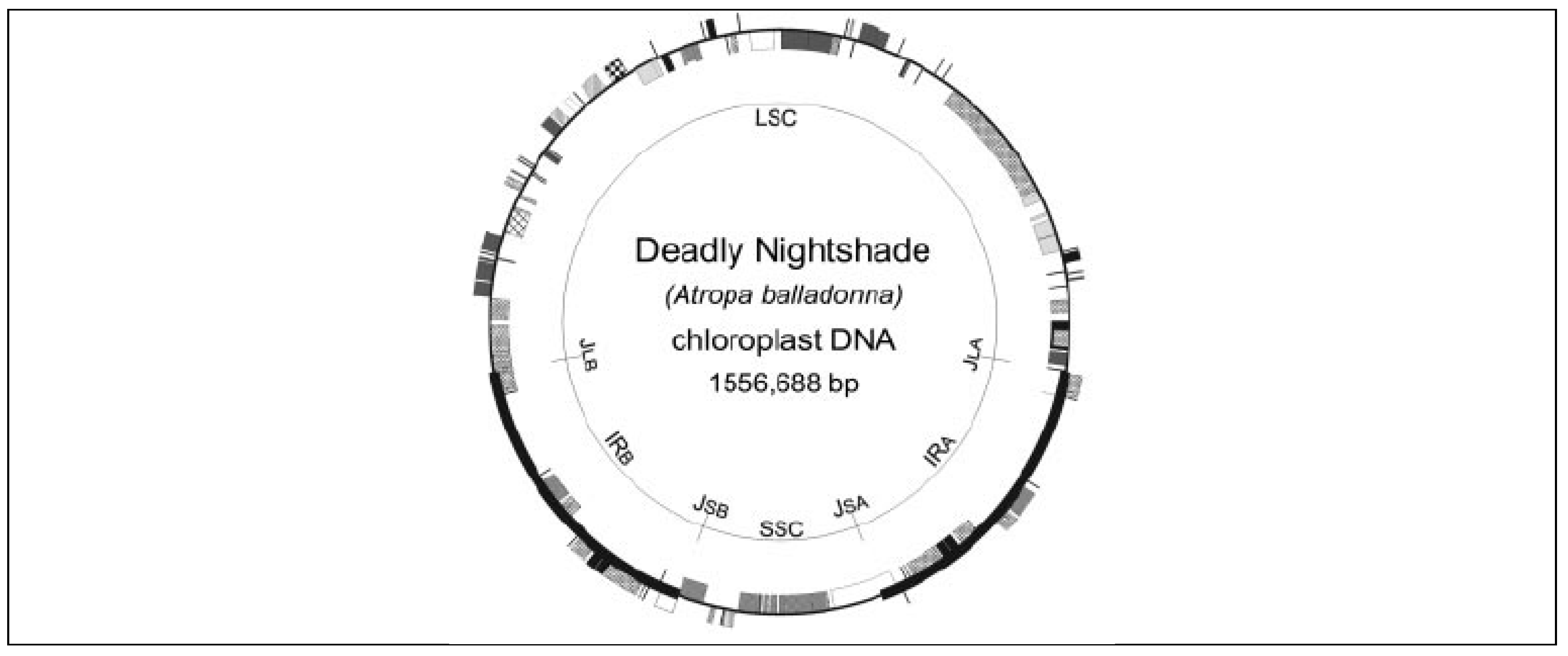
Figure 4.
Areas of potential barcodes of Atropa belladonna in the genome of chloroplast (Source: Kress et al., 2005).
Figure 4.
Areas of potential barcodes of Atropa belladonna in the genome of chloroplast (Source: Kress et al., 2005).

Figure 5.
Enhanced matK loci. Coding regions are represented by boxes, and spacer regions by connecting lines. (Source: Kress et al., 2005).
Figure 5.
Enhanced matK loci. Coding regions are represented by boxes, and spacer regions by connecting lines. (Source: Kress et al., 2005).
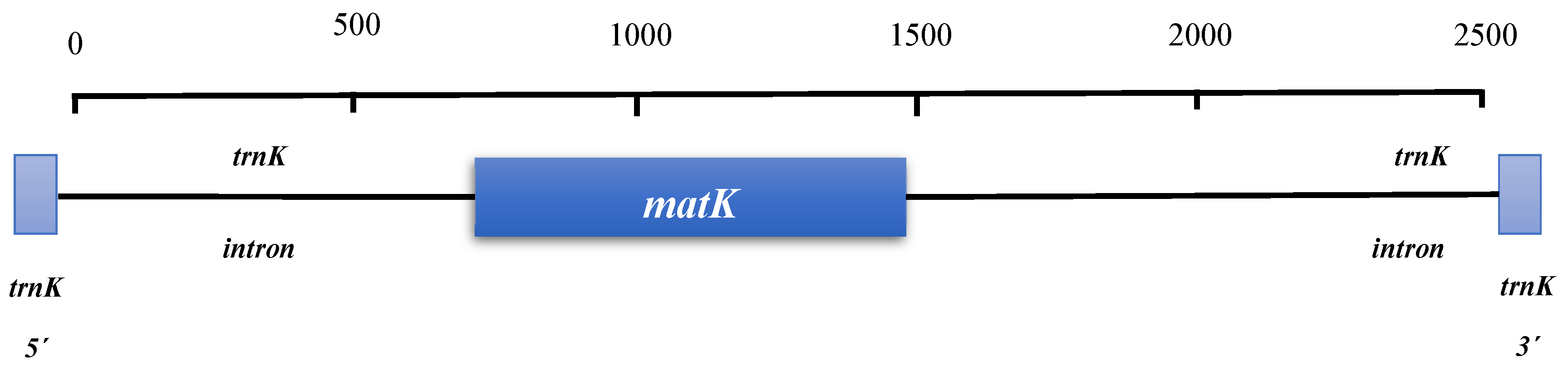
Figure 6.
The Atropa belladonna rbcL region of the plastome (Source: Chen et al., 2010).
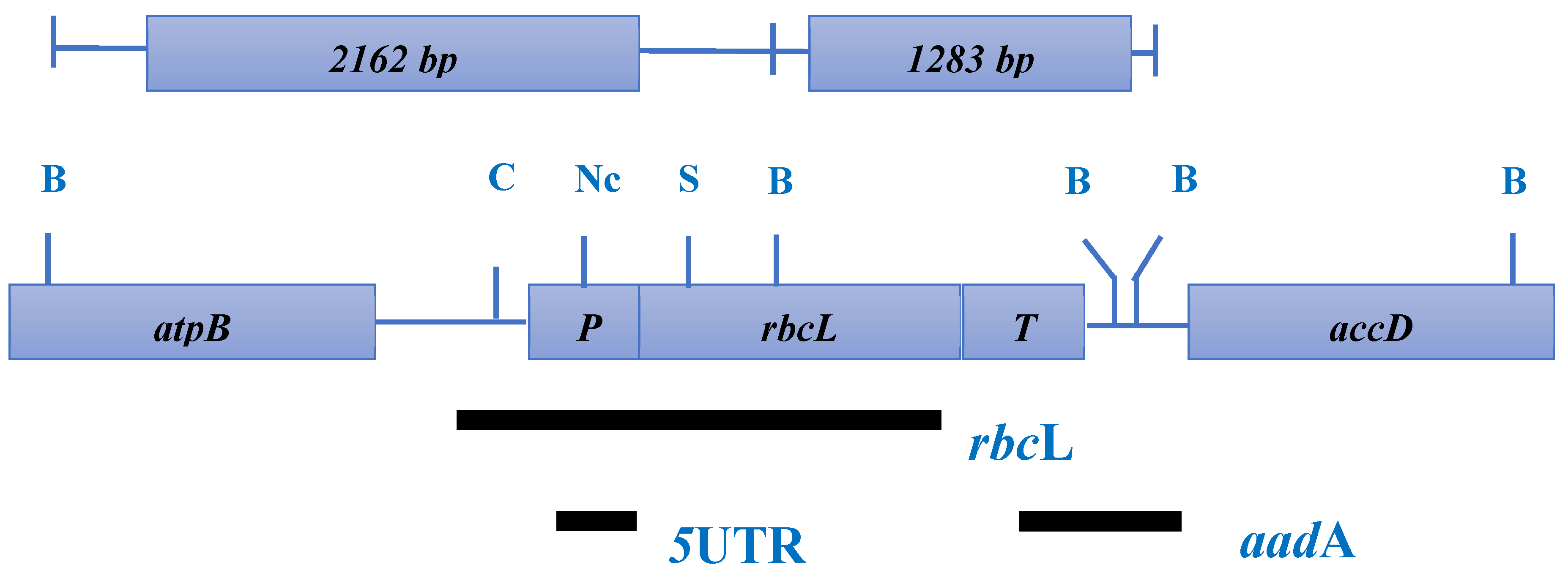
Figure 7.
A summary diagram of the psbA-trnH spacer’s structure is shown, with the fixed C areas, and attribute V areas denoted (Source: Shaw et al., 2007).
Figure 7.
A summary diagram of the psbA-trnH spacer’s structure is shown, with the fixed C areas, and attribute V areas denoted (Source: Shaw et al., 2007).
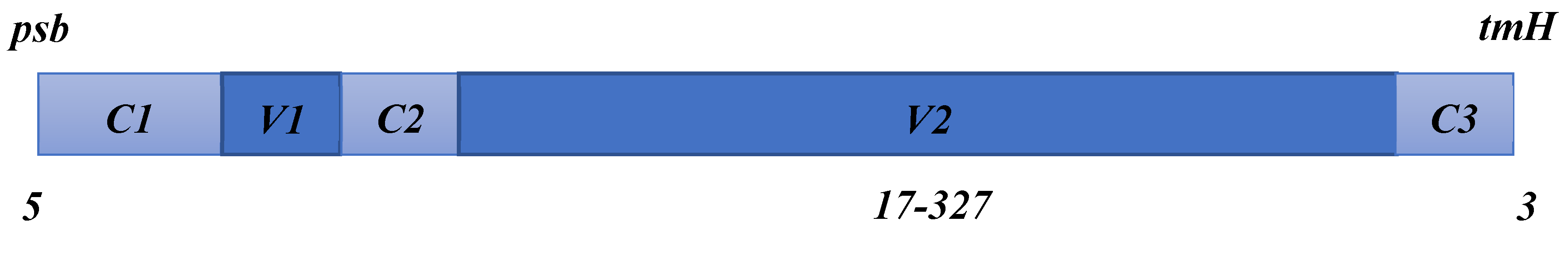
Figure 8.
Location in relation to the ITS area. Coding areas are represented by boxes, and connecting lines indicate spacer areas (Source: Kress et al., 2005).
Figure 8.
Location in relation to the ITS area. Coding areas are represented by boxes, and connecting lines indicate spacer areas (Source: Kress et al., 2005).
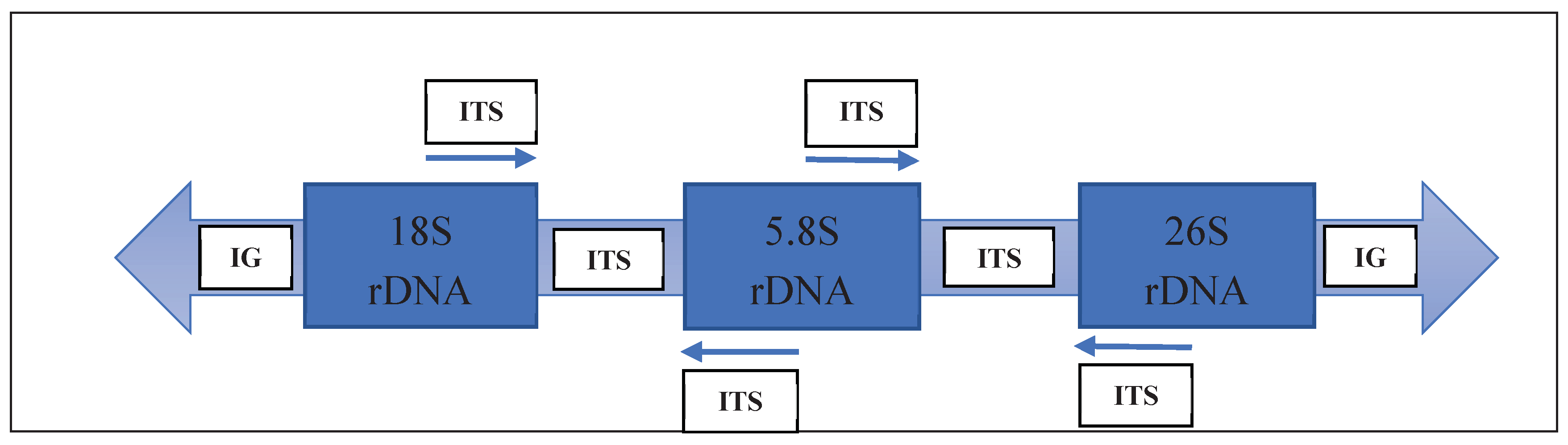

Table 1.
Classification of Atropa belladonna.
| Kingdom | Plantae |
|---|---|
| Division | Magnoliophyts |
| Class | Mangoliopsida |
| Order | Solanales |
| Family | Solanaceae |
| Genus | Atropa |
| Species | A. belladonna |
Table 2.
Atropa belladonna species.
| Belladonna sp. | Local name | Synonyms |
|---|---|---|
| Atropa belladonna | Atropa acuminate, Deadly Nightshade. | Belle-Dame, Bouton Noir, Grande Morelle, Herbe du Diable, Poison Black Cherries, Belle-Galante, Devil’s Herb, Morelle Furieuse, Baccifère, Divale, Herbe à la Mort, Cerise du Diable, Dwayberry, Naughty Man’s Cherries, Dwale, Guigne de la Côte, Devil’s Cherries. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



