
Submitted:
10 March 2024
Posted:
11 March 2024
You are already at the latest version
Abstract
Fibrosis is a ubiquitous pathology, and prior studies have indicated that various artemisinin (ART) derivatives (including artesunate (AS), artemether (AM), and dihydroartemisinin (DHA)) can reduce fibrosis in vitro and in vivo. The medicinal plant, Artemisia annua L., is the natural source of ART and is widely used, especially in underdeveloped countries, to treat a variety of diseases including malaria, while A. afra contains no ART but is also antimalarial. Using human dermal fibroblasts (CRL-2097), we compared the effects of A. annua and A. afra tea infusions, ART, AS, AM, DHA, and a liver metabolite of ART, deoxyART (dART), on fibroblast viability and expression of key fibrotic marker genes after 1 and 4 days of treatment. AS, DHA, and Artemisia tea significantly reduced fibroblast viability at 1 and 4 d post-treatment. After 4 d post treatment, AS, DHA, and A. afra tea downregulated ACTA2, and upregulated MMP3, with other genes either being unaffected or differentially affected. ART and AM had no significant effect on either fibroblast viability or fibrotic gene expression. Although A. annua contains ART, it had a significantly greater anti-fibrotic effect than ART alone. Immunofluorescent staining for smooth muscle α-actin (α-SMA) correlated well with transcriptional responses of drug-treated fibroblasts. Together, proliferation, qPCR, and immunofluorescence results show that treatment with ART, AS, DHA, and the two Artemisia teas yield differing responses, including those related to fibrosis, in human dermal fibroblasts.
Keywords:
artemisinin
; artesunate
; dihydroartemisinin
; artemether
; Artemisia annua
; Artemisia afra
; tea infusion
; α-SMA
1. Introduction
Fibrosis is a pathological healing process where the response to tissue injury occurs via non-regenerative mechanisms and leads to scar formation [1]. Fibrotic healing can affect a variety of organs including the skin, liver, kidney, lungs, and heart. Instead of replacing injured tissue with functional native cellular components and appropriate microstructure, this chronic condition is characterized by transdifferentiation of resident connective tissue fibroblasts or other progenitor cells into highly synthetic and contractile α-SMA+ myofibroblasts [2,3], which produce excessive amounts of acellular, primarily collagen I-based extracellular matrix (ECM) that lacks functional properties of the uninjured tissue. We and others have shown that multifunctional members of the transforming growth factor β (TGF-β) family act as master regulators of fibrosis [4,5]. Several signaling pathways downstream of TGF-β (e.g., SMAD, MAPK, and PI3K/Akt-mediated signaling) engage in crosstalk with other upstream cytokines, leading to complex signaling paradigms involved in fibrotic healing [6,7,8]. Although our understanding of the basic science of fibrosis has progressed significantly over the past several decades, development of clinically successful therapeutics has lagged greatly behind the elucidation of key mechanisms underlying development, progression, and maintenance of tissue fibrosis (reviewed by [9], underlying a dire need for discovery of novel anti-fibrotic therapeutics.
Artemisinin (ART) produced in the plant Artemisia annua L. (Figure 1), and ART derivatives (artesunate, AS; dihydroartemisinin, DHA; and artemether, AM), are sesquiterpene lactone antimalarials with demonstrated antiparasitic [10,11], antibacterial [10,12,13,14], antiviral [15,16,17,18], anticancer [19], and anti-fibrotic effects [20,21,22]. When compared, A. annua is equally or more efficacious in vitro than ART against many of the same diseases [23,24], e.g., tuberculosis (TB) [12,13] and Covid-19 [17,25]. ART from A. annua also distributes efficiently to many tissues and organs including lungs, liver, muscle, brain, and heart [26]. A side benefit of using ART or A. annua is their antinociceptive activity [27,28,29,30,31]. There also are no significant adverse effects with long term use of A. annua [27,28,29,32]. Though A. annua is globally used as a medicinal plant against many diseases including malaria, and despite accumulating evidence that artemisinin chemical derivatives may be effective in counteracting fibrotic tissue responses, there is a dearth of studies examining the effects of A. annua on fibrosis.
Previously we showed that AS treatment ameliorated fibrosis in human dermal fibroblasts, [21] a finding that was subsequently corroborated [33,34] and that is consistent with the literature describing anti-fibrotic effects of artemisinin compounds in fibrotic progenitor cells of other organs. Here we compared the effects of ART, AS, DHA, AM, and A. annua and A. afra hot water extracts (tea infusions) on the viability and gene expression of dermal fibroblasts. Tea infusions remain a common mode of traditional use among global populations, especially in low- and middle-income countries [35].
2. Results
2.1. Differential Effects of Artemisia Teas vs. ART Drugs on Human Dermal Fibroblast Viability
Preliminary data from viability assays suggested that A. annua tea, ART, and AS did not similarly affect viability of human dermal fibroblasts. That led us to further explore how the other ART derivatives, AS, AM, and DHA, and ART’s liver metabolite dART, affected fibroblast viability. After 1 d of treatment, fibroblast viability significantly decreased when treated with A. annua tea, AS, or DHA, with a further decline after 4 d drug exposure (Figure 2). In contrast, after 1 d exposure to ART, AM, or dART, there was no significant decrease in fibroblast viability. At 4 d post-treatment, viability modestly but significantly increased in AM-treated fibroblasts. Treatment with the liver metabolite of ART, dART, also modestly but significantly increased fibroblast viability after 1 d of treatment, but not after 4 d. Together these results showed that, while ART is a major constituent of A. annua tea, as a pure molecule at the same molar concentration as in the tea, it had either no or slight effect on fibroblast viability. In contrast, A. annua tea, AS, and DHA all significantly inhibited fibroblast viability. Futher experiments focused on the effects on expression of fibrosis-related genes by A. annua tea, ART (to compare with the ART in A. annua tea), AS, and DHA.
2.2. Differential Expression of Fibrotic Genes in ART Drug Treated Fibroblasts
Preliminary transcriptional response of eight genes (ACTA2, COL1A1, COL3A1, TGFB1, TGFB2, TGFBR1, MMP1, and MMP3) related to ECM remodeling and transition to fibrosis was measured in fibroblasts after 4 d treatment with ART, AS, or a hot water extract of A. annua. Transcriptional results showed that cells treated with ART or AS responded differently with AS having a more robust effect on transcription than ART. ART and A. annua tea treatments also yielded an unexpected dissimilar response, given that we had hypothesized that the effects of A. annua tea on fibroblasts were due at least in part to effects of ART.
Because they did not inhibit fibroblast viability (Figure 2), we eliminated AM and dART from further study, and instead focused on transcriptional analyses of the effects of ART, AS, and DHA (Figure 3). After fibroblasts were treated for 4 d with 50 µM ART, AS, and DHA, ART increased MMP1 about 2.8-fold, but otherwise lacked effects on expression of other markers. AS decreased ACTA2 expression about 3.4-fold, COL1A1 2.9-fold, COL3A1 5-fold, TGFB2 4.6-fold, and TGFBR1 2.5-fold. AS increased MMP1 and MMP3 17.5- and 32.6-fold, respectively. DHA decreased ACTA2 expression 6.2-fold, TGFB2 3.7-fold, and TGFBR1 4-fold, while increasing MMP1 and MMP3 8- and 51.8-fold, respectively. Other investigated marker genes were unaffected.
2.3. A. afra vs. A. annua.
A. afra lacks artemisinin but is closely related to A. annua, and it has shown similar therapeutic efficacy against various diseases. Therefore, to better query whether other phytochemicals may be affecting fibroblast viability and gene expression, we compared infusions of A. afra to those of A. annua (Figure 4). Surprisingly, on a dry mass basis, A. afra was about four times more potent than A. annua at inhibiting fibroblast viability (Figure 4) and, at a concentration equal to that of the A. annua infusion, completely inhibited fibroblast viability (data not shown). Transcriptional responses were also dramatically different, with A. afra tea increasing COL3A1 transcripts by about 5-fold compared to untreated cells, whereas A. annua tea did not affect COL3A1 expression. A. afra also increased MMP3 expression nearly 16-fold (versus 5-fold for A. annua) but decreased MMP1 expression 7.2-fold and TGFB1 3.5-fold.
2.4. Differential Expression of α-SMA Protein after 1 and 4 days of ART, ART Drugs and Artemisia Treatments in Fibroblasts
To determine if transcriptional changes were also accompanied by changes in protein levels, fibroblasts treated for 1 and 4 d with drugs were fixed and stained for α-SMA using immunofluorescence. DMSO (control solvent for all ART pure drugs), water (control solvent for Artemisia teas), and media with no solvent or drug showed similar staining patterns of α-SMA (Figure 5). After 4 d, α-SMA staining correlated with the qPCR data in Figure 3 and Figure 4; AS, DHA, and the two Artemisia teas considerably decreased α-SMA compared to their respective solvent controls (Figure 5). In contrast, ART showed substantial increase in α-SMA staining compared to its untreated DMSO control. Together, our qPCR and immunofluorescence results correlate, demonstrating that ART, AS, DHA, and the two Artemisia teas have different anti-fibrotic responses in human dermal fibroblasts.
2.5. A. annua, ART, and ART Drugs Show Bioconversion in Culture Medium after 4 d Incubation
Microscopic analysis suggested that differences among the effects of ART drugs may be mediated by the presence of particular artemisinic metabolic products in culture, especially in those treated with DHA and AS. We consequently extracted media from cells incubated for 1 and 4 d in media containing A. annua tea (diluted to 50 µM ART), and ART, AS, or DHA at 50 µM as for previous experiments, and then separated the extracts via thin layer chromatography (TLC) to visualize the amount of each ART drug remaining after 1 and 4 d treatment (Figure 6). When treated with A. annua tea, ART nearly disappeared from the media and dART became more apparent, but not in amounts sufficient to account entirely for the ART disappearance if converted to dART; also note that dART is already present in A. annua tea. A similar disappearance of ART from ART-treated cells occurred, accompanied by an increase in dART, by day 4. AS and DHA were nearly gone from the media after day 1 and were undetectable by day 4 (Figure 6), suggesting that any effects from these compounds were likely driven by responses initiated early in the treatment period.
Because fibroblast culture medium has a diversity of components, we wondered if the medium itself contributed to the degradation of the drugs. This was confirmed after GC-MS analysis of the culture media incubated with the same concentrations of drugs for 4 days, but without any cells (Table 1). Of the ART in the A. annua tea extract, ~9% remained on day 4, but >99% of ART in ART-containing medium disappeared. Neither AS nor DHA was detectable after 4 d incubation in cell-free media (Table 1). The CYP P450 metabolite of ART, dART, was observed in AS and DHA samples after 4 d drug treatment (Figure 6). Surprisingly, fibroblast-free media also had dART after 4 d incubation in drugs (Table 1) suggesting P450 activity in the culture media, which does contain some serum. Taken together, however, the media analyses indicate that thedramatic differences observed in viability, gene expression, and immunochemistry among cell cultures exposed to different ART drugs and Artemisia treatments are likely regulated by different metabolites of these species in culture, at least some of which occurs even in the absence of cells. This suggests that more frequent drug dosing and monitoring of media metabolites may be required in order to better understand the in vitro efficacy of ART drugs.
3. Discussion
Against malaria, the mechanism of ARTs is consistent, requiring the endoperoxide bridge, acting via several mechanisms. As a prodrug, ART is activated by heme after parasite digestion of hemoglobin, resulting in generation of reactive oxygen species (ROS) [36] that damage the parasite but not human cells (see review by [37]). In another case, mitochondria of the parasite activate ART, leading to mitochondrial damage of the parasite but not of human cells [37]. Other possible mechanisms include the inhibition of PfATP6, analogous to mammalian SERCA, and potential covalent binding of heme-activated ART to a large number of other identified proteins [37,38,39]. AS, DHA, AM, and ART all contain the endoperoxide bridge responsible for heme activation of ART (Figure 1). However, in dermal fibroblasts, our study showed that neither ART nor AM seemed to have anti-fibrotic nor potential ECM-remodeling activity, while AS, DHA, and Artemisia extracts did. Furthermore, ART-containing A. annua has in vitro anti-fibrotic activity that differs from pure ART, suggesting that the plant tissue contains other molecules with the ability to modulate fibrotic responses.
The degradation of AS to DHA and then DHA to dART reflects the instability of AS in aqueous media [40]. While dART is not therapeutically active against malaria, AS is ART acylated with succinate to make the drug more bioavailable, but it is also readily hydrolyzed to DHA, which then is the bioactive metabolite against malaria (see review by [41]). Although observed degradation of the ART drugs over 4 d may indicate a need for additional dosing in future experiments, the data showed that there were still measurable phenotypic responses of cultured fibroblasts to the drugs. Nevertheless, the observed degradation should be considered in any analysis of subsequent results, as well as a lens through which to interpret other data generated from the use of these drugs in cell culture. For example, while a variety of ART drugs including ART, DHA, AM, AS, and SM934 have demonstrated anti-fibrotic efficacy when applied to various disease models in vivo, the vast majority of in vitro data in cell types relevant to fibrosis has been generated using artesunate or DHA (reviewed in [20]). Notably, this is consistent not only with our data in this manuscript which, for example, demonstrated anti-proliferative effects towards fibroblasts from artesunate and DHA that were notably lacking in artemisinin and artemether, but also with the known susceptibility of artesunate to spontaneous (non-enzymatic) decomposition in aqueous solution [42] or (enzymatic) hydrolysis of the ester group in the presence of esterases (e.g., introduced by animal serum supplemented to culture medium). This raises the intriguing possibility that the active anti-fibrotic agent common to most (or even all) of these previous studies is DHA, regardless of the specific agent delivered, and that myriad reports of anti-fibrotic effects of ART drugs both in vivo and in vitro are actually describing delivery of pro-drugs that only ameliorate fibrosis when they result in administration of meaningful quantities of DHA (i.e., (1) via aqueous decomposition or enzymatic conversion to DHA in vitro, depending on the cell and media type, (2) via hepatic metabolism to DHA in vivo, or (3) due to direct administration of DHA as an active component). Metabolism of AS, ART, and DHA to dART in cell-free media may be from trace CYP P450 activity that is detectable in serum that was in the fibroblast culture media [43].
Fibrosis is a pathological process of staggering consequence. Development of effective treatments for tissue fibrosis has been hampered by the clinical failure of agents designed towards obvious fibrotic targets, particularly TGF-β family ligands and receptors, largely due to functional redundancy and positive feedback loops characteristic of the fibrotic response [44]. In addition, therapeutic strategies that are shown to blunt development of tissue fibrosis in pre-clinical models may translate poorly to use in humans, since human patients tend to be diagnosed with fibrotic indications at late stages of disease development, at which time fibrosis has advanced significantly and resulted in tissue damage, necessitating reversal rather than prevention of the fibrotic response in order to yield any functional benefit [45]. The observed increase in MMP3 transcripts by AS, DHA, and A. afra and increases in COL3A1 by A. afra also indicate these drugs have the potential for remodeling of the ECM. Thus, the identification and development of novel therapeutic strategies to treat fibrotic diseases in humans is of paramount importance. Extensive research has identified ART derivatives as effective therapeutics for blunting development of tissue fibrosis in both in vitro and pre-clinical animal models of disease (reviewed in [20]). However, practically none of this literature has investigated the therapeutic effects of artemisinin delivered as A. annua, nor the additional benefit of other phytochemicals introduced upon ingestion of plant material. Previously, we showed that ART is significantly more bioavailable when delivered per os via the plant vs. as a pure drug and distributes to multiple organs including heart, lungs, liver, brain, etc. [26]. This greater ART bioavailability from the plant vs. the pure drug is enhanced by the plant’s essential oils [46,47], greater intestinal transport [48], and inhibition of hepatic CYPs 3A4 and 2B6 metabolism of ART [26,49] resulting in greater levels of serum ART [26,50,51].
Limitations of this study include investigation of expression of a small number of genes and proteins, as well as analysis of only one isolate of human fibroblasts, though our data are consistent with our [21] and others’ [33,34] demonstrations of similar effects in other isolates of fibroblasts, as well as with numerous reports of anti-fibrotic activity of artesunate and DHA in other myofibroblast progenitor cells that are known to contribute to fibrosis (Cite our review and/or the other review, only if you want to). Future work will seek to characterize with greater resolution the effects of artemisinin family compounds and Artemisia extracts on pro-fibrotic phenotypes in fibroblasts, as well as assess and compare the anti-fibrotic effects of these treatments utilizing in vivo models of skin fibrosis, which will shed light on the effects of these compounds on other cells relevant to wound healing (e.g., macrophages, endothelial cells, etc.) and give evidence for or against their use as therapeutic compounds for skin fibrosis in humans.
4. Materials and Methods
4.1. Biological Materials
CRL-2097 human dermal fibroblasts were acquired from ATCC and maintained in culture in DMEM/F-12 with GlutaMAX supplement (Thermo Fisher Scientific 10565018) and 10% Fetal Clone III (Cytiva SH30109.03), and incubated at 37 °C and 5% CO2 in high humidity. Dried leaves of Artemisia annua L. cv. SAM (voucher MASS 317314; batch B#1.Sh6.01.15.20) and Artemisia afra cv. MAL Jacq. ex Willd. (voucher FTG181107; batch B#1RbA.10.12.20) were acquired from Atelier Temenos LLC in Homestead, FL. Hot water extracts (tea) at 10 g leaf dry weight per L were prepared according to Kane et al. [49]. Artemisinin content in the tea was 581.69 µM and was diluted to 50 µM for experiments. For A. afra experiments, hot water extracts were prepared in a similar manner.
4.2. Drug Treatments
Drug treatments were performed as follows. A. annua tea was diluted in growth media to 50 µM ART; A. afra tea was diluted with water to 1/2, 1/4, or 1/8 strength of the A. annua tea on a dry weight basis as needed; artemisinin (ART; Cayman Chemical 11816), artesunate (AS; Cayman Chemical 11817), artemether (AM; gift from Prof. J. Plaizier-Vercammen (Brussels, Belgium)), dihydroartemisinin (DHA; Cayman Chemical 19846) and deoxyartemisinin (dART; Toronto Research Chemicals D232150) were dissolved in DMSO as 1000X stock solutions, filter sterilized, and added to a final concentration of 50 μM. DMSO and water were the solvent controls for the artemisinic compounds and teas, respectively. Fibroblasts were seeded in 6-well plates in 2 mL medium at a density of 6250 cells/cm2. After overnight attachment, media was aspirated and replaced with test drugs at the indicated concentrations and incubated at 37 °C and 5% CO2 in high humidity. After 4 d of drug treatment, cells were harvested, washed with 1× Dulbecco’s phosphate-buffered saline (DPBS; Thermo Fisher Scientific 14190136), snap frozen in liquid nitrogen, and stored at −80 °C until time of RNA extraction for analysis.
4.3. Resazurin Assay
Fibroblasts were seeded in 96 well plates at a density of 2000 cells/well (~6060 cells/cm2) and, after overnight attachment, cells were treated with drugs as described elsewhere. After 1 or 4 d of treatment, media was aspirated and replaced with media containing resazurin (Thermo Fisher Scientific B21187.03) at a final concentration of 25 µg/mL (stock solution 0.15 mg/mL in DPBS). Cells were incubated for 2 h at 37 °C as above, and then fluorescence was read at ex/em = 544 nm/590 nm on a Perkin Elmer Victor3 1420 plate reader.
4.4. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
RNA from cells after 4 d drug treatment was isolated using the NucleoSpin RNA/protein mini kit (Macherey-Nagel 740933.50) according to the manufacturer’s instructions. The concentration of isolated RNA was measured using a Nanodrop 2000c Spectrophotometer (Thermo Scientific). Complementary DNA (cDNA) was synthesized from RNA using a QuantiTect reverse transcription kit (Qiagen 205313) following manufacturer’s instructions. SYBR-based PowerTrack master mix (Thermo Fisher Scientific A46012) was used to perform RT-qPCR on a QuantStudio 6 Pro Real-Time PCR system (Thermo Fisher Scientific) in 10 μL reactions. PCR primer sequences are listed in Table 2.
4.5. Immunocytochemistry
Fibroblasts were seeded in 48 well plates at 6060 cells/cm2 and, after overnight attachment, cells were treated with drugs as described above. After 1 and 4 d, cells were fixed with paraformaldehyde and stained for immunofluorescence using primary mouse anti-α-SMA (Santa Cruz Biotechnology sc-32251) and secondary goat anti-mouse IgG AlexaFluor488-conjugated antibodies (Thermo Fisher Scientific A-11001), and DNA was counterstained with Hoechst 33342 as previously described [8].
4.6. Analysis of Artemisinic Metabolites in Media
Media extracts were spotted on Si-gel 60 F254 plates (EMD Millipore cat #1.05735.0001) and run in a mobile phase of 5:11:4 dichloromethane:hexane:acetone plus 0.5% v/v acetic acid. Plates were stained with p-anisaldehyde reagent (75 mL ethanol, 2.5 mL sulfuric acid, 1 mL acetic acid, 2 mL p-anisaldehyde) and heated for 10 min at 105 °C. ART, dART, AS, and DHA were quantified in fibroblast media after two-phase extraction with dichloromethane using gas chromatography mass spectrometry as previously detailed in Kane et al. [49] on an Agilent GC-MS system (GC, Agilent 7890A; MS, Agilent 5975 C).
4.7. Sample Sizes and Statistical Analyses
Resazurin analyses each comprised at least 12 replicates, and Student’s t-tests were used to compare treatments versus respective controls and between the two time points. For the RT-qPCR calculations, 4–8 biological replicates were pooled from at least 2 independent experiments to create box plots. The box plots were calculated using GraphPad Prism version 10.0.0.
5. Conclusions
To our knowledge this is the first comparison of the different artemisinic drugs, ART, AS, AM, DHA, and A. annua and A. afra tea infusions on dermal fibroblasts as they shift into myofibroblasts that precede and contribute to fibrotic tissue formation.
Author Contributions
Conceptualization, P.W., D.D., and T.D.; validation, M.T.; formal analysis, M.T.; investigation, M.T. and B.H.K.; resources, P.W. and T.D.; writing—original draft preparation, P.W.; writing—review and editing, P.W., M.T., B.H.K., D.D., and T.D.; visualization, M.T.; supervision, P.W. and M.T.; project administration, P.W. and T.D.; funding acquisition, P.W. and T.D. All authors have read and agreed to the published version of the manuscript.
Funding
Award Number NIH-2R15AT008277-02 to PJW from the National Center for Complementary and Integrative Health partially and previously funded analysis of the plant material in this study. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Center for Complementary and Integrative Health or the National Institutes of Health. Atelier Temenos provided funding some supplies and a summer stipend for undergraduate, Trevor Bush.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
Authors thank undergraduates Trevor Bush (WPI), and Hadiya Giwa (Tufts University). Victoria Bicchieri (WPI) provided fluorescence microscopy assistance. Gratitude is extended also to Atelier Temenos for funding an undergraduate to participate in the project during the summer of 2023.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the plant materials used in this study are available from the authors upon reasonable request.
References
- Wynn, T. Cellular and molecular mechanisms of fibrosis. The Journal of Pathology: A Journal of the Pathological Society of Great Britain and Ireland 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.; Duffy, H.S. Fibroblasts and myofibroblasts: what are we talking about? Journal of cardiovascular pharmacology 2011, 57, 376–379. [Google Scholar] [CrossRef]
- Hinz, B. Myofibroblasts. Experimental eye research 2016, 142, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Dolivo, D.M.; Larson, S.A.; Dominko, T. Crosstalk between mitogen-activated protein kinase inhibitors and transforming growth factor-β signaling results in variable activation of human dermal fibroblasts. Int. J. Mol. Med. 2018, 43, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Common and unique mechanisms regulate fibrosis in various fibroproliferative diseases. J. Clin. Investig. 2007, 117, 524–529. [Google Scholar] [CrossRef]
- Kim, K.K.; Sheppard, D.; Chapman, H.A. TGF-β1 signaling and tissue fibrosis. Cold Spring Harbor perspectives in biology 2018, 10, a022293. [Google Scholar] [CrossRef]
- Zhang, Y.E. Non-Smad pathways in TGF-β signaling. Cell Res. 2009, 19, 128–139. [Google Scholar] [CrossRef]
- Dolivo, D.M.; Larson, S.A.; Dominko, T. FGF2-mediated attenuation of myofibroblast activation is modulated by distinct MAPK signaling pathways in human dermal fibroblasts. J. Dermatol. Sci. 2017, 88, 339–348. [Google Scholar] [CrossRef]
- Henderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: from mechanisms to medicines. Nature 2020, 587, 555–566. [Google Scholar] [CrossRef]
- Efferth, T. Artemisinin: a versatile weapon from traditional Chinese medicine. In Herbal drugs: ethnomedicine to modern medicine; Springer: 2009; pp. 173–194.
- Naß, J.; Efferth, T. The activity of Artemisia spp. and their constituents against Trypanosomiasis. Phytomedicine 2018, 47, 184–191. [Google Scholar] [CrossRef]
- Kiani, B.H.; Alonso, M.N.; Weathers, P.J.; Shell, S.S. Artemisia afra and Artemisia annua Extracts Have Bactericidal Activity against Mycobacterium tuberculosis in Physiologically Relevant Carbon Sources and Hypoxia. Pathogens 2023, 12, 227. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.C.; Zhang, T.; Williams, J.T.; Abramovitch, R.B.; Weathers, P.J.; Shell, S.S. Artemisia annua and Artemisia afra extracts exhibit strong bactericidal activity against Mycobacterium tuberculosis. J. Ethnopharmacol. 2020, 262, 113191–113191. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Williams, J.T.; Aleiwi, B.; Ellsworth, E.; Abramovitch, R.B. Inhibiting Mycobacterium tuberculosis DosRST Signaling by Targeting Response Regulator DNA Binding and Sensor Kinase Heme. ACS Chem. Biol. 2019, 15, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T. Beyond malaria: The inhibition of viruses by artemisinin-type compounds. Biotechnol. Adv. 2018, 36, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.R.; Efferth, T.; Serrano, M.A.; Castaño, B.; Macias, R.I.; Briz, O.; Marin, J.J. Effect of artemisinin/artesunate as inhibitors of hepatitis B virus production in an “in vitro” replicative system. Antivir. Res. 2005, 68, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.; Huang, Y.; Fidock, D.; Polyak, S.; Wagoner, J.; Towler, M.; Weathers, P. Artemisia annua L. extracts inhibit the in vitro replication of SARS-CoV-2 and two of its variants. J. Ethnopharmacol. 2021, 274, 114016–114016. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gilmore, K.; Ramirez, S.; Settels, E.; Gammeltoft, K.A.; Pham, L.V.; Fahnøe, U.; Feng, S.; Offersgaard, A.; Trimpert, J.; et al. In vitro efficacy of artemisinin-based treatments against SARS-CoV-2. Sci Rep 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Abba, M.L.; Patil, N.; Leupold, J.H.; Saeed, M.E.; Efferth, T.; Allgayer, H. Prevention of carcinogenesis and metastasis by Artemisinin-type drugs. Cancer Lett. 2018, 429, 11–18. [Google Scholar] [CrossRef]
- Dolivo, D.; Weathers, P.; Dominko, T. Artemisinin and artemisinin derivatives as anti-fibrotic therapeutics. Acta Pharm. Sin. B 2020, 11, 322–339. [Google Scholar] [CrossRef]
- Larson, S.A.; Dolivo, D.M.; Dominko, T. Artesunate inhibits myofibroblast formation via induction of apoptosis and antagonism of pro-fibrotic gene expression in human dermal fibroblasts. Cell Biol. Int. 2019, 43, 1317–1322. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; You, F.; Xue, J. Novel use for old drugs: The emerging role of artemisinin and its derivatives in fibrosis. Pharmacol. Res. 2020, 157, 104829. [Google Scholar] [CrossRef] [PubMed]
- Gruessner, B.M.; Cornet-Vernet, L.; Desrosiers, M.R.; Lutgen, P.; Towler, M.J.; Weathers, P.J. It is not just artemisinin: Artemisia sp. for treating diseases including malaria and schistosomiasis. Phytochem. Rev. 2019, 18, 1509–1527. [Google Scholar] [CrossRef]
- Weathers, P.J. Artemisinin as a therapeutic vs. its more complex Artemisia source material. Natural Product Reports 2023.
- Nie, C.; Trimpert, J.; Moon, S.; Haag, R.; Gilmore, K.; Kaufer, B.B.; Seeberger, P.H. In vitro efficacy of Artemisia extracts against SARS-CoV-2. Virol J 2021, 18, 182. [Google Scholar] [CrossRef] [PubMed]
- Desrosiers, M.R.; Mittleman, A.; Weathers, P.J. Dried Leaf Artemisia Annua Improves Bioavailability of Artemisinin via Cytochrome P450 Inhibition and Enhances Artemisinin Efficacy Downstream. Biomolecules 2020, 10, 254. [Google Scholar] [CrossRef]
- Hunt, S.; Stebbings, S.; McNamara, D. An open-label six-month extension study to investigate the safety and efficacy of an extract of Artemisia annua for managing pain, stiffness and functional limitation associated with osteoarthritis of the hip and knee. . 2016, 129, 97–102. [Google Scholar] [PubMed]
- Stebbings, S.; Beattie, E.; McNamara, D.; Hunt, S. A pilot randomized, placebo-controlled clinical trial to investigate the efficacy and safety of an extract of Artemisia annua administered over 12 weeks, for managing pain, stiffness, and functional limitation associated with osteoarthritis of the hip and knee. Clin. Rheumatol. 2015, 35, 1829–1836. [Google Scholar] [CrossRef]
- Yang, M.; Guo, M.-Y.; Luo, Y.; Yun, M.-D.; Yan, J.; Liu, T.; Xiao, C.-H. Effect of Artemisia annua extract on treating active rheumatoid arthritis: A randomized controlled trial. Chin. J. Integr. Med. 2016, 23, 496–503. [Google Scholar] [CrossRef]
- de Faveri Favero, F.; Grando, R.; Nonato, F.R.; Sousa, I.M.; Queiroz, N.C.; Longato, G.B.; Zafred, R.R.; Carvalho, J.E.; Spindola, H.M.; Foglio, M.A. Artemisia annua L. : evidence of sesquiterpene lactones’ fraction antinociceptive activity. BMC Complementary and Alternative Medicine 2014, 14, 1–11. [Google Scholar]
- Dehkordi, F.M.; Kaboutari, J.; Zendehdel, M.; Javdani, M. The antinociceptive effect of artemisinin on the inflammatory pain and role of GABAergic and opioidergic systems. Korean J. Pain 2019, 32, 160–167. [Google Scholar] [CrossRef]
- Han, B.; Kim, S.-M.; Nam, G.E.; Kim, S.-H.; Park, S.-J.; Park, Y.-K.; Baik, H.W. A Randomized, Double-Blind, Placebo-Controlled, Multi-Centered Clinical Study to Evaluate the Efficacy and Safety of Artemisia annua L. Extract for Improvement of Liver Function. Clinical Nutrition Research 2020, 9, 258. [Google Scholar] [PubMed]
- Li, R.; Yin, H.; Wang, J.; He, D.; Yan, Q.; Lu, L. Dihydroartemisinin alleviates skin fibrosis and endothelial dysfunction in bleomycin-induced skin fibrosis models. Clin. Rheumatol. 2021, 40, 4269–4277. [Google Scholar] [CrossRef] [PubMed]
- Nong, X.; Rajbanshi, G.; Chen, L.; Li, J.; Li, Z.; Liu, T.; Chen, S.; Wei, G.; Li, J. Effect of artesunate and relation with TGF-β1 and SMAD3 signaling on experimental hypertrophic scar model in rabbit ear. Arch. Dermatol. Res. 2019, 311, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Poswal, F.S.; Russell, G.; Mackonochie, M.; MacLennan, E.; Adukwu, E.C.; Rolfe, V. Herbal Teas and their Health Benefits: A Scoping Review. Plant Foods Hum. Nutr. 2019, 74, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Benakis, A.; Romero, M.R.; Tomicic, M.; Rauh, R.; Steinbach, D.; Häfer, R.; Stamminger, T.; Oesch, F.; Kaina, B.; et al. Enhancement of cytotoxicity of artemisinins toward cancer cells by ferrous iron. Free. Radic. Biol. Med. 2004, 37, 998–1009. [Google Scholar] [CrossRef]
- Yang, J.; He, Y.; Li, Y.; Zhang, X.; Wong, Y.-K.; Shen, S.; Zhong, T.; Zhang, J.; Liu, Q.; Wang, J. Advances in the research on the targets of anti-malaria actions of artemisinin. Pharmacol. Ther. 2020, 216, 107697–107697. [Google Scholar] [CrossRef] [PubMed]
- Karunajeewa, H.A. Artemisinins: artemisinin, dihydroartemisinin, artemether and artesunate. Treatment and prevention of malaria: antimalarial drug chemistry, action and use 2012, 157-190.
- Wang, J.; Zhang, C.-J.; Ni Chia, W.; Loh, C.C.Y.; Li, Z.; Lee, Y.M.; He, Y.; Yuan, L.-X.; Lim, T.K.; Liu, M.; et al. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef] [PubMed]
- Gashe, F.; Wynendaele, E.; De Spiegeleer, B.; Suleman, S. Degradation kinetics of artesunate for the development of an ex-tempore intravenous injection. Malar. J. 2022, 21, 1–11. [Google Scholar] [CrossRef]
- A Morris, C.; Duparc, S.; Borghini-Fuhrer, I.; Jung, D.; Shin, C.-S.; Fleckenstein, L. Review of the clinical pharmacokinetics of artesunate and its active metabolite dihydroartemisinin following intravenous, intramuscular, oral or rectal administration. Malar. J. 2011, 10, 263–263. [Google Scholar] [CrossRef]
- Bezuidenhout, J.W.; Aucamp, M.; Stieger, N.; Liebenberg, W.; Haynes, R.K. Assessment of Thermal and Hydrolytic Stabilities and Aqueous Solubility of Artesunate for Formulation Studies. Aaps Pharmscitech 2023, 24, 1–7. [Google Scholar] [CrossRef]
- Steenbergen, R.; Oti, M.; ter Horst, R.; Tat, W.; Neufeldt, C.; Belovodskiy, A.; Chua, T.T.; Cho, W.J.; Joyce, M.; Dutilh, B.E.; et al. Establishing normal metabolism and differentiation in hepatocellular carcinoma cells by culturing in adult human serum. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G. Transforming growth factor–β in tissue fibrosis. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Walraven, M.; Hinz, B. Therapeutic approaches to control tissue repair and fibrosis: Extracellular matrix as a game changer. Matrix Biol. 2018, 71-72, 205–224. [Google Scholar] [CrossRef] [PubMed]
- Desrosiers, M.R.; Weathers, P.J. Effect of leaf digestion and artemisinin solubility for use in oral consumption of dried Artemisia annua leaves to treat malaria. J. Ethnopharmacol. 2016, 190, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Desrosiers, M.R.; Towler, M.J.; Weathers, P.J. Artemisia annua and Artemisia afra Essential Oils and Their Therapeutic Potential. In Essential Oil Research: Trends in Biosynthesis, Analytics, Industrial Applications and Biotechnological Production, Malik, S., Ed.; Springer International Publishing: Cham, 2019; pp. 197–209. [Google Scholar]
- Desrosiers, M.R.; Weathers, P.J. Artemisinin permeability via Caco-2 cells increases after simulated digestion of Artemisia annua leaves. J. Ethnopharmacol. 2018, 210, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Kane, N.F.; Kiani, B.H.; Desrosiers, M.R.; Towler, M.J.; Weathers, P.J. Artemisia extracts differ from artemisinin effects on human hepatic CYP450s 2B6 and 3A4 in vitro. J. Ethnopharmacol. 2022, 298, 115587. [Google Scholar] [CrossRef]
- Weathers, P.J.; Elfawal, M.A.; Towler, M.J.; Acquaah-Mensah, G.K.; Rich, S.M. Pharmacokinetics of artemisinin delivered by oral consumption of Artemisia annua dried leaves in healthy vs. Plasmodium chabaudi-infected mice. J. Ethnopharmacol. 2014, 153, 732–736. [Google Scholar] [CrossRef]
- Weathers, P.J.; Towler, M.; Hassanali, A.; Lutgen, P.; Engeu, P.O. Dried-leaf Artemisia annua: A practical malaria therapeutic for developing countries? World journal of pharmacology 2014, 3, 39. [Google Scholar] [CrossRef]
Figure 1.
Left: artemisinin (ART), artemisinin derivatives (DHA, AS, AM), and an artemisinin liver metabolite, deoxyartemisinin (dART); right: Artemisia annua and afra.
Figure 1.
Left: artemisinin (ART), artemisinin derivatives (DHA, AS, AM), and an artemisinin liver metabolite, deoxyartemisinin (dART); right: Artemisia annua and afra.
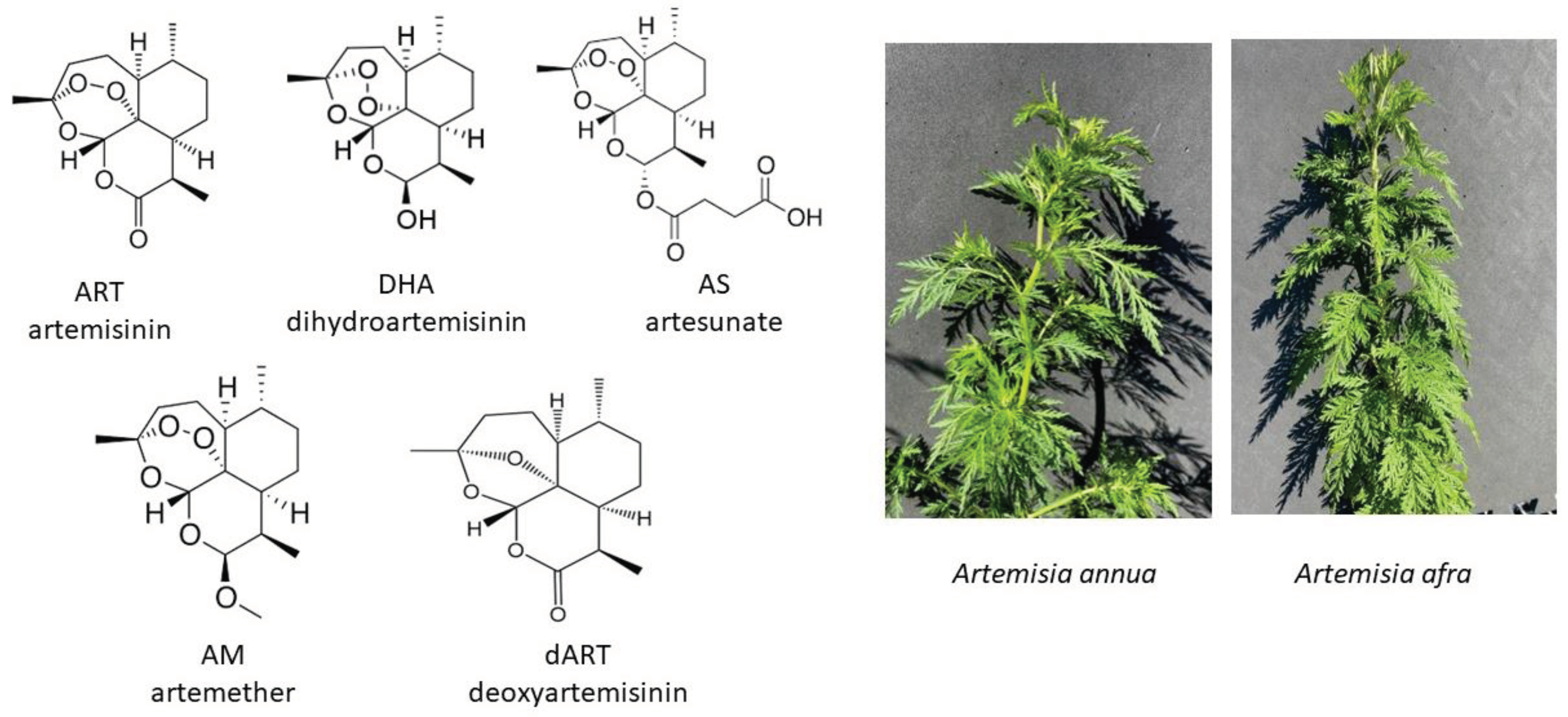
Figure 2.
Relative viability responses of human dermal fibroblasts via resazurin assay at 1 and 4 days after exposure to artemisinin, each of the artemisinic derivatives, artemisinin’s liver metabolite deoxyartemisinin, and hot water extracts of A. annua, all normalized to 50 µM of the relevant artemisinic compound. Control = 100%; n ≥ 12; * = p ≤ 0.05 for samples to solvent control; # = p ≤ 0.05 for 1 day compared to 4 days using Student’s t-test.
Figure 2.
Relative viability responses of human dermal fibroblasts via resazurin assay at 1 and 4 days after exposure to artemisinin, each of the artemisinic derivatives, artemisinin’s liver metabolite deoxyartemisinin, and hot water extracts of A. annua, all normalized to 50 µM of the relevant artemisinic compound. Control = 100%; n ≥ 12; * = p ≤ 0.05 for samples to solvent control; # = p ≤ 0.05 for 1 day compared to 4 days using Student’s t-test.
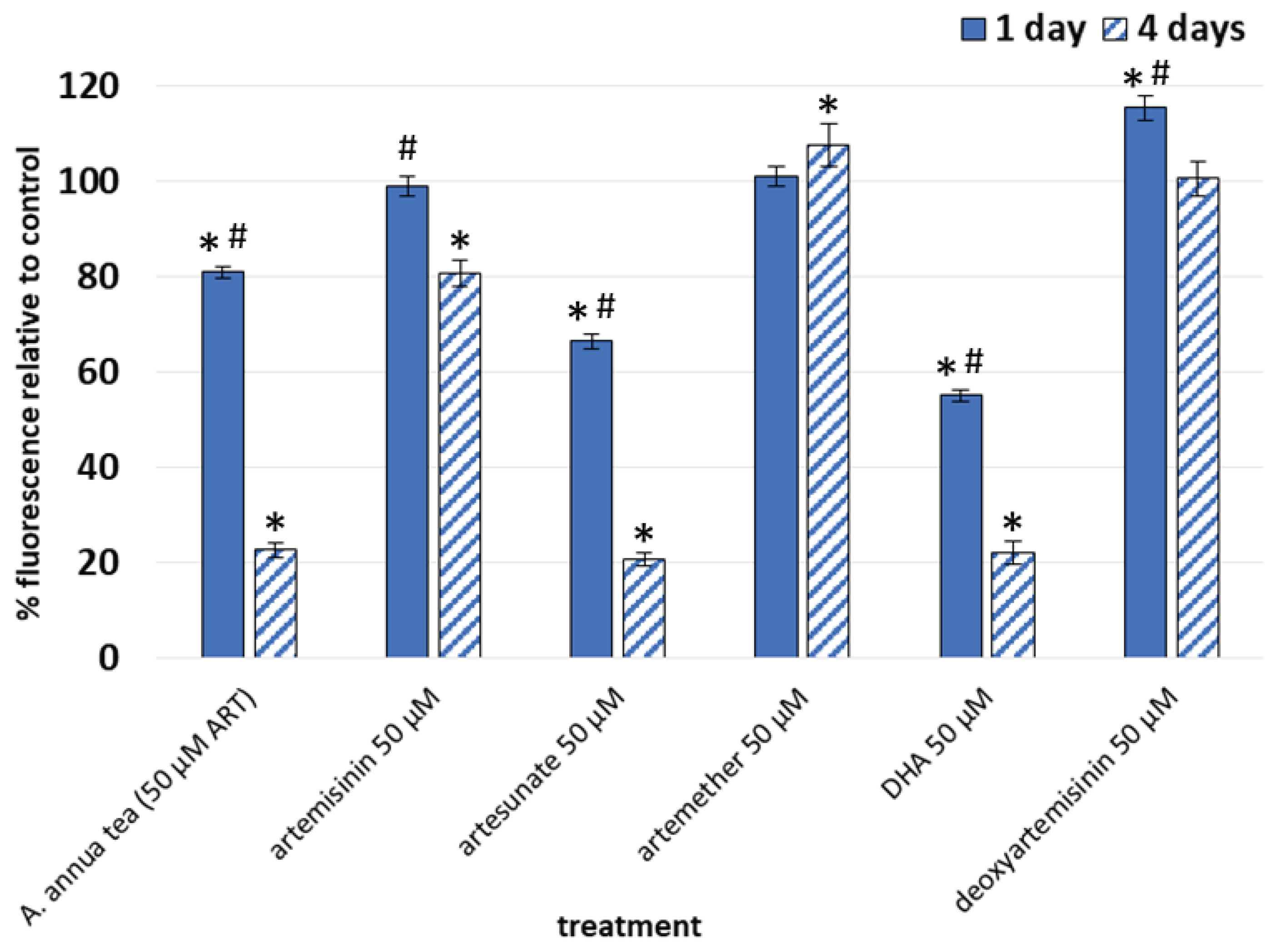
Figure 3.
Fold change vs. solvent control in gene expression in fibroblasts after 4 d treatment with 50 µM ART, AS, or DHA; GAPDH was used as the internal control; n ≥ 5.
Figure 3.
Fold change vs. solvent control in gene expression in fibroblasts after 4 d treatment with 50 µM ART, AS, or DHA; GAPDH was used as the internal control; n ≥ 5.
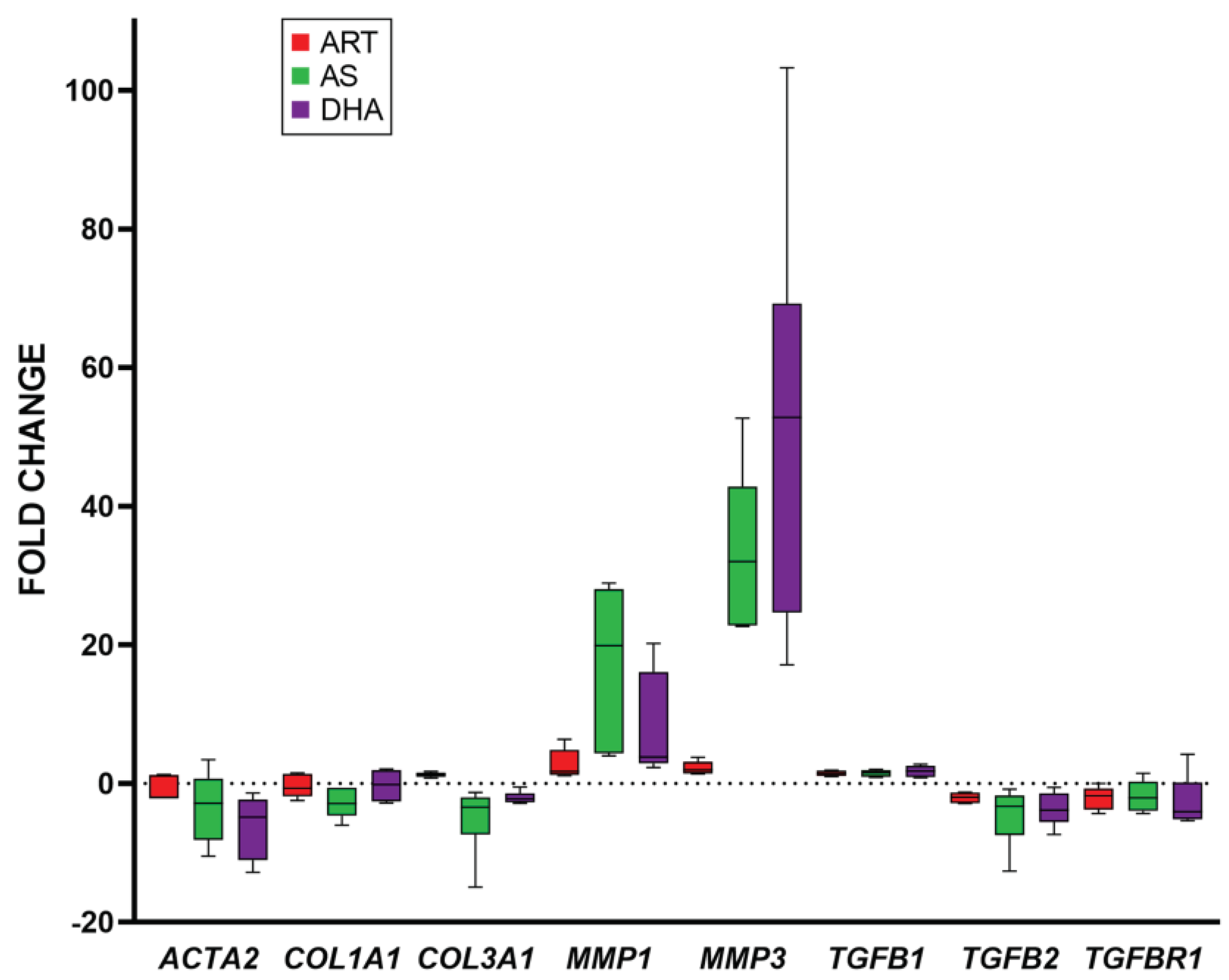
Figure 4.
Viability and transcript expression of fibroblasts after treatment with A. annua or A. afra infusions. Left: resazurin assay of fibroblast viability after 1 and 4 d treatment with A. annua and A. afra infusions. Right: Fold change vs. solvent control in gene expression in fibroblasts after 4 d treatment with infusions of A. annua (at 50 µM ART content) and A. afra diluted to ¼ of the dry mass of the A. annua tea; GAPDH was used as the internal control; n = 12 for resazurin assay (* = p ≤ 0.05 vs. solvent control; and # = p ≤ 0.05, 1 d vs 4 d); and n ≥ 4 for gene expression.
Figure 4.
Viability and transcript expression of fibroblasts after treatment with A. annua or A. afra infusions. Left: resazurin assay of fibroblast viability after 1 and 4 d treatment with A. annua and A. afra infusions. Right: Fold change vs. solvent control in gene expression in fibroblasts after 4 d treatment with infusions of A. annua (at 50 µM ART content) and A. afra diluted to ¼ of the dry mass of the A. annua tea; GAPDH was used as the internal control; n = 12 for resazurin assay (* = p ≤ 0.05 vs. solvent control; and # = p ≤ 0.05, 1 d vs 4 d); and n ≥ 4 for gene expression.
Figure 5.
Representative views of fibroblasts after 1 and 4 d treatment with ART, AS, DHA, and A. annua (AnT) and A. afra (1/4X; AfT) tea infusions (n = 3). Cells were stained for smooth muscle α-actin (α-SMA) and nuclei were counterstained with Hoechst. Scale bar = 100 μm for all images.
Figure 5.
Representative views of fibroblasts after 1 and 4 d treatment with ART, AS, DHA, and A. annua (AnT) and A. afra (1/4X; AfT) tea infusions (n = 3). Cells were stained for smooth muscle α-actin (α-SMA) and nuclei were counterstained with Hoechst. Scale bar = 100 μm for all images.

Figure 6.
Thin layer chromatography of media after 1 and 4 day A. annua tea and ART drug treatments of fibroblasts. Media was extracted with dichloromethane and then spotted on Si-gel 60 F254 plates and run in a mobile phase of 5:11:4 DCM:hexane:acetone plus 0.5% v/v acetic acid. Plates were stained with p-anisaldehyde reagent that stains all ART drugs dark blue-purple, except for ART (dark pink) and dART (brown). Image is a composite of representative media extracts of three replicates.
Figure 6.
Thin layer chromatography of media after 1 and 4 day A. annua tea and ART drug treatments of fibroblasts. Media was extracted with dichloromethane and then spotted on Si-gel 60 F254 plates and run in a mobile phase of 5:11:4 DCM:hexane:acetone plus 0.5% v/v acetic acid. Plates were stained with p-anisaldehyde reagent that stains all ART drugs dark blue-purple, except for ART (dark pink) and dART (brown). Image is a composite of representative media extracts of three replicates.
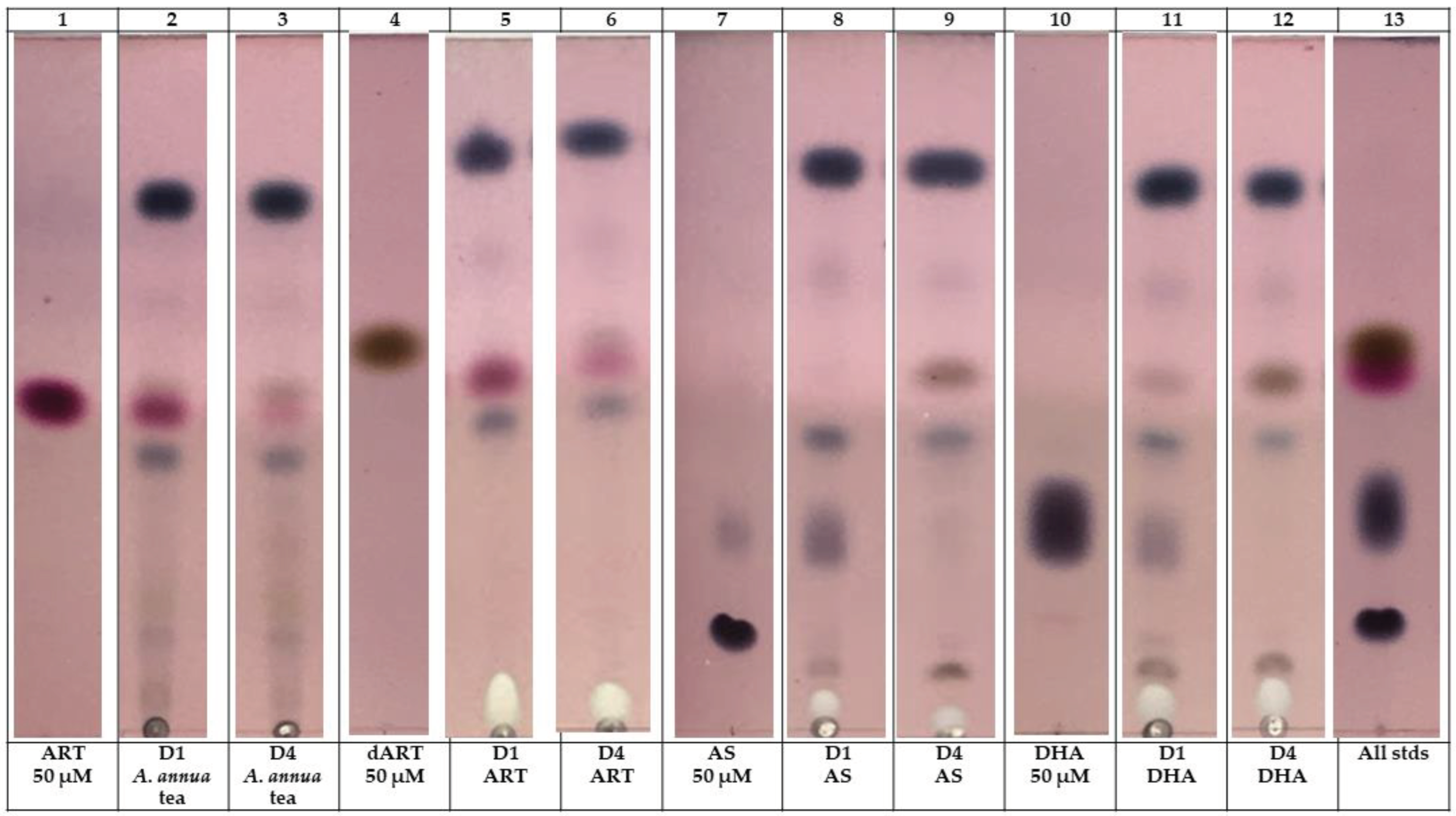

Table 1.
Culture media degrades ART drugs in vitro. GC-MS analysis of culture media incubated for 4 days with 50 µM ART drugs in the absence of cells. ND = not detected.
Table 1.
Culture media degrades ART drugs in vitro. GC-MS analysis of culture media incubated for 4 days with 50 µM ART drugs in the absence of cells. ND = not detected.
| Treatment | Day 0 (µg/mL) | Day 4 (µg/mL) |
|---|---|---|
|
A. annua tea ART dART |
141.2 14.5 |
12.0 5.1 |
| ART ART dART |
141.2 0 |
0.42 0.5 |
| AS AS dART |
192.2 0 |
ND 1.5 |
| DHA DHA dART |
142.2 0 |
ND 6.9 |
Table 2.
Primers used in the RT-qPCR analysis of drug-treated fibroblasts.
| Gene name | Protein encoded |
Forward primer (5′-3′) | Reverse primer (5′-3′) |
|---|---|---|---|
| ACTA2 | α-SMA | ACTGCCTTGGTGTGTGACAA | |
| CACCATCACCCCCTGATGTC | |||
| COL1A1 | Collagen 1 (α1) | GTCAGGCTGGTGTGATGGG | |
| GCCTTGTTCACCTCTCTCGC | |||
| COL3A1 | Collagen III (α1) | GGACACAGAGGCTTCGATGG | |
| CTCGAGCACCGTCATTACCC | |||
| MMP1 | Matrix metalloproteinase 1 | GCATATCGATGCTGCTCTTTC | |
| GATAACCTGGATCCATAGATCGTT | |||
| MMP3 | Matrix metalloproteinase 3 | ACCTGACTCGGTTCCGCCTG | |
| GTCAGGGGGAGGTCCATAGAGGG | |||
| TGFB1 | TGF-β1 | CATTGGTGATGAAATCCTGGT | |
| TGACACTCACCACATTGTTTTTC | |||
| TGFB2 | TGF-β2 | GAGCGACGAAGAGTACTACG | |
| TTGTAACAACTGGGCAGACA | |||
| TGFBR1 | TGF-βR1 | GCAGACTTAGGACTGGCAGTAAG | |
| AGAACTTCAGGGGCCATGT | |||
| GAPDH | GAPDH | GAGTCCACTGGCGTCTTCAC | |
| TTCACACCCATGACGAACAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



