
Submitted:
13 March 2024
Posted:
14 March 2024
You are already at the latest version
Abstract
Willow (Salix spp.) trees, found worldwide, contain secondary metabolites that are valuable as dietary supplements for animal feed and as anti-parasitic compounds. We quantified secondary metabolites (phenolic, flavonoids, and salicylic acid) in ethanolic extracts from leaves and branches of three Salix acmophylla genotypes and investigated their potential to inhibit Eimeria sp. sporulation, a major concern in ruminants. The total phenolic content of willow leaves and branches was similar in two of three different genotypes. The total flavonoid content of the branches was significantly higher than that of leaves of the same genotype; however, the salicylic acid content was significantly higher in leaves than in branches. Importantly, all extracts exhibited significant inhibition of Eimeria sporulation, where over 70% inhibition was obtained at concentrations as low as 750 ppm. The sporulation inhibition by branch or leaf extracts exceeded 80% for leaves and 90% for branches at concentrations above 1250 ppm. The study highlights the potential of using Salix extracts as bioactive compounds for biological control of coccidiosis in ruminants and emphasizes the importance of genotype-specific variations in secondary metabolite profiles. We conclude that all parts of S. acmophylla can provide secondary metabolites that act as a coccidiostat to treat Eimeria in goats.
Keywords:
Ethanol extracts
; Plant secondary metabolites
; Coccidia sporulation
; Eimeria sp.
; willow genotypes
1. Introduction
Willows (genus Salix), the largest woody species in the Salicaceae family, thrive in both natural and cultivated habitats across numerous countries worldwide [1]. The trees encompass up to 500 species and more than 200 hybrids, characterized by significant variability in size, growth rates, and plant architecture [2,3]. Willows have been used as a traditional remedy for more than 6,000 years [4].
Willow exhibits high rates of evapotranspiration and a robust tolerance to soil saturation, which can lead to oxygen shortages in the root zone [5]. These trees are distinguished by their rapid growth rates, substantial biomass yield across multiple harvests, adaptability to dense plantings, and ability to regenerate from the stump after numerous harvests [6]. Nutritionally, willow leaves and fine branches offer richer sustenance for grazing ruminants compared to low-quality summer pastures [5,7]. Beyond nutritious forage, the trees yield nutraceutical compounds such as anthelminthics, antioxidants, and analgesic secondary metabolites [8,9,10]. Secondary metabolites can protect plants against herbivores, bacteria, viruses, or competing plants, and act as signaling compounds to attract pollinators or facilitate seed dispersers. Thus, these secondary metabolites are not only crucial for the survival and reproductive fitness of the plant but can also serve as adaptive components subject to natural selection throughout evolution [11,12].
Secondary metabolites in willows are primarily composed of phenols and phenolic glycoside compounds. Phenolic compounds, based on their chemical structure, can be categorized into subgroups such as phenolic acids, flavonoids, tannins, coumarins, lignans, quinones, stilbenes, and curcuminoids [13]. Phenolic glycosides, especially salicylates, have been utilized by humans as remedies for pain relief, fever reduction, and the alleviation of rheumatic disorders, inflammations, and headaches [8]. Salicin, the most prevalent phenolic glucoside in willows, is converted into acetylsalicylic acid, the nonsteroidal anti-inflammatory drug commonly known as aspirin [14,15]. Salidroside, also found in willow, attenuates inflammatory responses by suppressing nuclear factor-κB and mitogen-activated protein kinase activation in lipopolysaccharide-induced mastitis in mice [16]. Additionally, administering sodium salicylate to cows in early lactation has been shown to have beneficial effects throughout the whole lactation period [17].
At least 36 different chemical compounds were identified in willow plant extracts, some of which exhibit antioxidant, antimicrobial, and cytotoxic activities against human leukemia cells (HL 60) [18,19,20,21]. While the growth of willow plants may be influenced by water sources, their chemical content remains unaffected [6]. Furthermore, secondary metabolites in willow plants offer a promising alternative to conventional drugs for treating parasitic nematodes in livestock [22,23,24,25,26]. Most research has focused on S. alba (native to Europe and Asia) and S. babylonica (native to China), with relatively little investigation into S. acmophylla, which is native to the Middle East. Four Salix species of willow were reported in Israel by Rottenberg, et al. (1999) [27]. However, Muklada et al., (2020) analyzed willow samples from both Jordan and Israel, identifying only two species: pure S. alba, pure S. acmophylla, and hybrids of S. acmophylla with S. alba and other Salix species. The current study utilized the S. acmophylla genotypes as defined by Muklada et al., (2020) [28].
The S. acmophylla biomass is composed of 45% leaves and 55% branches, on a dry matter (DM) basis [6]. The nutrient and mineral contents, as well as the concentration of secondary compounds in branches and leaves, varied between two willow phenotypes, termed “red” and “white” for their bark color. White willows contained 1.2-fold more salicin, 1.5-fold more gallic acid, and 1.8-fold more kaempferol than red willows. The two types of willow did not show differences in their concentrations of salicylic acid, hyperin, salidroside, and helicon. Short-term feeding of dairy goats with willow fodder demonstrated positive effects on somatic cell counts and might promote anti-inflammatory effects and/or delay of mammary involution [6]. Subsequent research revealed that S. acmophylla has three genotypes [29], which have yet to be investigated regarding their metabolic and coccidistatic characteristics.
In ruminants, coccidiosis is a parasitic disease caused by Eimeria spp., which has a significant economic impact due to its global distribution and high infection rates, sometimes exceeding 90% in certain areas [30,31,32,33]. The primary clinical symptom of coccidiosis is diarrhea, but under conditions that are favorable for Eimeria development, other accompanying clinical symptoms can include low feed conversion rate, weight loss, and even death, leading to substantial economic losses from increased veterinary costs and high mortality rates among infected animals [32,33]. Kids and lambs between one and four months of age are the most vulnerable compared to older animals [30,31,33]. While coccidiosis can affect all livestock species, it is important to note that coccidia is host-specific, meaning that the coccidia found in cattle or chicken are specific to those species and do not cause disease in goats and sheep, or vice versa. Rivero-Perez et al., (2019) identified a significant inhibitory effect (97% in vivo) of S. babylonica extract on Eimeria sp. coccidia in rabbits at doses ranging from 25 to 50 mg/kg body weight.
In pursuit of a natural source of bioactive compounds for the biological control of coccidiosis in goats, we extracted secondary metabolites from leaves and branches of different S. acmophylla genotypes and evaluated their potential to inhibit the sporulation of Eimeria oocysts found in goats.
2. Materials and Methods
2.1. Plant Material and Samples Preparation
Willows collected from native stands across various regions of Jordan during 2013-2017 were planted in replicated blocks in a 0.5 ha plot at the Jordan Valley research center at Deir Allah in 2021, as part of a joint Jordanian-Israeli research project funded by USAID. From a collection of more than 40 ecotypes sampled in different regions in Israel and Jordan, three genotypes of S. acmophylla were identified [28]. Genotype 1 is pure S. acmophylla, genotype 2 is a 60:40 mix of S. acmophylla and another unidentified Salix species, and genotype 3 is a 50:50 mix of S. acmophylla and S. alba. In June 2023, foliage of trees was collected from Deir Allah, and separated into leaves and thin branches up to 12 mm in thickness. This included genotype 1 (a mixture of ecotypes from Ain Fotaha, Rajab, and Shoubak), genotype 2 (a mixture of ecotypes from Ajloun and Wadi Kursi), and genotype 3 (a mixture of ecotypes from Rmimmen and Hajleh). The samples were initially dried under shade for a few days and subsequently for 48 h in an oven at 60ºC. The dried materials were ground into composite samples for each genotype, stored in paper bags at room temperature (23-26ºC), shipped to Israel, and maintained at 20-22ºC until use.
2.2. Quantity and Quality of Secondary Metabolites in Willow Leaves and Branches
2.2.1. Preparation of Plant Extracts
Samples were prepared as previously described by Azaizeh et al., (2013) [35]; and Awwad et al., (2021) [26]. Five grams of dried leaves or branches were incubated with 50 ml of 7:3 ethanol: water (v:v) for 24 h. The mixture was then filtered and the solvent evaporated under a vacuum (Rotorvapor Hie-VAP; Hiedolph, Germany) at 40ºC to remove the ethanol and water. The yield of the extract was calculated by dividing the weight of the leaf or branch extract (g) by the weight of the leaves or branches before extraction (g), and expressed as (g) of extract per (g) of DM of plant biomass, in percentages. The extract was stored at -20ºC until it was analyzed or used.
2.2.2. Phenolic Acids and Flavonoid Assays
The total phenolic acid content in plant extracts was evaluated and calculated as equivalents of gallic acid (GAE) or tannic acid (TAE) according to the Folin-Ciocalteu method as described by Zhishen et al., (1999) [36]. Salicylic acid content was determined according to Warrier et al., (2013) [37], and flavonoid content according to Zhang et al., (2011) [38].
2.2.3. Coccid Sporulation Test
The effect of plant extracts from the three different willow genotypes was tested in vitro to assess their impact on coccid sporulation. Oocysts of Eimeria sp. were isolated from feces collected from goats that had spontaneously become infected during unsupervised grazing. The flotation method of Ekawasti et al., (2019) was adopted with some modifications for oocyst purification by using a saturated sugar solution (specific gravity 1.27). A 3.5 g sample of goat feces was lightly crushed, dissolved in 50 ml of tap water, and filtered through a sieve. The filtrate was centrifuged at 1700 rpm for 3 min at 4˚C. The supernatant was discarded and the sediment was suspended in a 50 ml saturated sugar solution (454 g sugar in 355 ml distilled water) and allowed to stand for 5 min to allow coarse fecal material to sink. The suspension was again centrifuged at 1700 rpm for 3 min at 4˚C. One ml of the sample solution was placed onto a McMaster Chamber to examine at a magnification of 40X and to count the Eimeria sp. oocysts in the entire field. When oocysts were detected, approximately 7 ml was aspirated from the 50 ml test tube surface and the oocyst suspension was further washed twice in phosphate saline buffer (pH=7.3) using centrifugation at 1700 rpm for 3 min at 4˚C. The supernatant was discarded, and the oocysts were concentrated by re-suspending from the sediment in a buffered saline solution. The oocysts were stored at 4˚C to prevent sporulation from occurring until they were used for the in vitro sporulation assay.
The sporulation test was conducted in 0.5 ml Eppendorf tubes containing a total volume of 100 µl of extracts at concentrations of 250, 750, 1250, and 5000 ppm, diluted from stock extracts from each willow genotype. The 100 µl of total volume included a fixed volume of 30 µl of oocysts suspension containing approximately 15X103 oocysts. The control group consisted of oocysts suspended in buffer alone.
The tubes were perforated to facilitate oxygen delivery and were incubated for 48 h at 28°C with constant shaking at 150 rpm to promote sporulation. After the control group reached 90% sporulation (within 2-3 days), the effects of willow extracts on oocyst sporulation were examined using a light microscope (Motic Laboratory Microscope BA210E) at a magnification of 40X, using 10 µl of each suspension.
Sporulated oocysts were counted, and the percentages of sporulation inhibition (SI) results were calculated using the following two equations:
where SI (%) is the percentage of inhibition of the sporulation.
2.2.4. Data Analysis
The experiment was performed twice in its entirety, with three replicate samples each time. Statistical analysis was carried out using PRISM 8 statistical software. A two-way ANOVA was utilized to assess the significance of the data in the study (p < 0.05). Means were separated using Tukey’s HSD test.
3. Results
3.1. Yield (%) of Ethanolic Extracts from Willow Leaves and Branches
The dry extract yield of willow leaves from the three different genotypes (L1, L2, and L3) averaged 31.3%, nearly 2.5 times greater than that of the branch extracts (B1, B2, and B3) (Table 1).
3.2. Total Phenolic Content of Willow Leaves and Branch Extracts' Obtained from the Different Genotypes
The total phenolic content (TPC) of both leaf and branch extracts showed little variations whether measured as GAE (gallic acid equivalents) or as TAE (tannic acid equivalents) (Table 1). Leaves from genotype 3 contained 17% more TPC than leaves from the other two genotypes. Branches of genotype 1 had approximately 17% more TPC than those from genotype 3, with the values for genotype 2 being intermediate (Table 1).
3.3. Total Flavonoid Content of Extracts from Leaves and Branches of the Different Willow Genotypes
The total flavonoid content (TFC) of leaves from genotypes 1 and 3 was similar and was 17% greater than the TFC of genotype 2 (Table 1). Branch extracts of genotype 1 had 24% more TFC than those of the other two genotypes. Branch TFC exceeded that of leaf TFC by 63%, 50%, and 38 %, for genotypes 1, 2, and 3, respectively (Table 1).
3.4. Salicylic Acid Content of Willow Leaves and Branches
The salicylic acid (SA) content of leaves was higher than that of branches by 50%, 34%, and 44% for genotypes 1, 2, and 3, respectively (Table 1). The SA content of leaves from genotype 1 was 27% higher than that of the other two genotypes, while the SA content in branches of genotype 3 was 33% higher than that of genotype 1, with genotype 2 being statistically comparable to the other two.
3.5. Anticoccidial Activity of Willow Extracts Obtained from Leaves and Branches at Different Concentrations of the Three Genotypes.
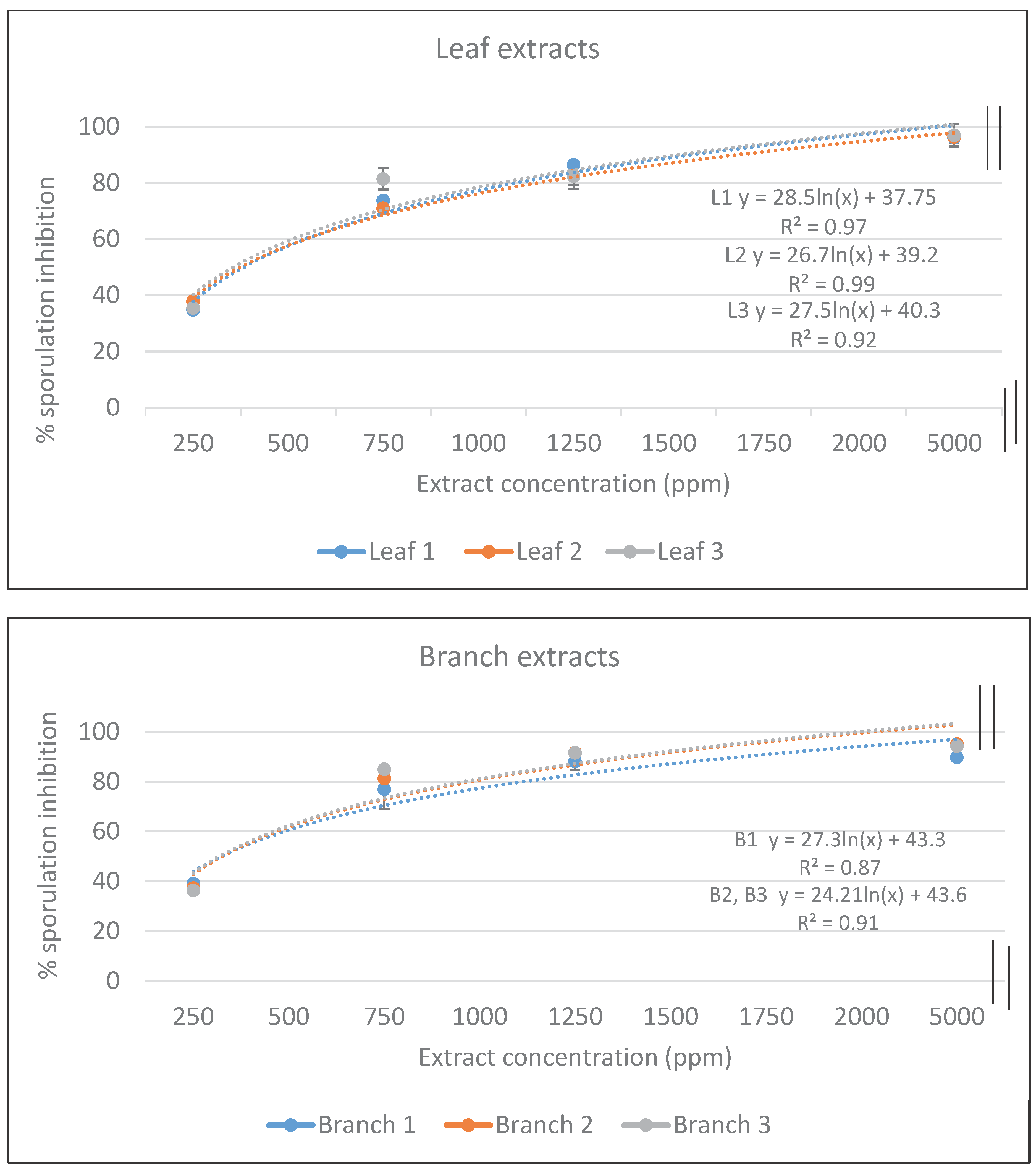
Leaf and branch extracts from the three different willow genotypes exhibited similar levels of sporulation inhibition percentages at a given concentration with no significant differences (Fig. 1). Increasing the concentration of the extracts from 250 ppm to 750 ppm, 1250 ppm, or 5000 ppm significantly increased sporulation inhibition from approximately 35% to 75% at 750 ppm and more than 90% at both 1250 and 5000 ppm.
Figure 1.
Percent inhibition of Eimeria sporulation by ethanolic extracts from leaf (above) or branch (below) tissues from the three genotypes of S. acmophylla. Each data point is the mean of two triplicate experiments. Standard deviation is shown where it is larger than the data point.
Figure 1.
Percent inhibition of Eimeria sporulation by ethanolic extracts from leaf (above) or branch (below) tissues from the three genotypes of S. acmophylla. Each data point is the mean of two triplicate experiments. Standard deviation is shown where it is larger than the data point.
4. Discussion
The phytochemistry of the Salicaceae family has been under investigation since the 19th century, initially focusing on various pharmaceutical applications, particularly related to the production of salicylic acid [13,26,40]. Subsequently, ecological reasons emerged as a significant area of study. These efforts have culminated in comprehensive knowledge regarding the secondary metabolite content of Salicaceae, including flavonoids, phenolic and nonphenolic glycosides, procyanidines, organic acids and their derivatives, simple phenolics, sterols and terpenes, lignans, volatiles, and fatty acids [8,13,26,40]. These substances have attracted considerable attention for their role in plant-herbivore interactions [9]. Willow extracts, in particular, have been recognized for their anti-inflammatory, antioxidant, anti-rheumatic, antipyretic, antibiotics, antidiabetic, anti-parasitic nematodes, and antiseptic properties, which can vary by genotype [26,40,41,42].
Our present results with S. acmophylla contain valuable data on extract yield, total phenolic content, flavonoids, salicylic acid, and the percentage of sporulation inhibition. The extract yield of willow leaf was more than 2.9-fold greater than that of branches. Salicylic acid levels in leaves exceeded those in branches by up to 1.4-fold. While branches contained as much as 2.6 times the total flavonoid content than that of leaves. However, the total phenolic content (TPC) was similar in both leaves and branches across the three tested genotypes. The source of irrigation water (treated wastewater or freshwater) did not influence the quantity and quality of secondary metabolites, such as phenols, in willow tissue, but there were differences in the chemical profile of glycoside phenols between willow genotypes [26].
Despite variations in secondary compound content between leaves and branches, sporulation inhibition percentages were similar (Fig. 1). The TPC indicates the overall phenolic content in an extract, but it does not reveal how the phenolic content varies from one extract to another, therefore, the use of HPLC becomes crucial, as it can indicate these variations. Preliminary HPLC analysis revealed significant differences in concentrations for similar phenols or flavonoids between branches or leaves, as well as some different phenolic compounds which require further analysis (data not shown). Salicylates, a group of chemically related phenolic glucosides based on the structure of saligenin (salicyl alcohol), are the most common phenolic glucosides found in the Salicaceae family, particularly in willow, aspen, and poplar [41,43]. To date, more than 20 different phenolic glycosides - glycosylated and esterified derivatives of salicyl alcohol have been described and found in variable concentrations in members of the Salicaceae [41]. Several of these compounds have shown bioactive properties against insects (Donaldson and Lindroth, 2007), and against nematodes in goats [26].
Extracts of S. babylonica have been extensively utilized as anthelmintics and were effective against gastrointestinal parasites in sheep and goats, including those of the genus Eimeria [24,45]. The extracts were orally administered to each animal weekly before the morning feeding for 60 days, during which fecal eggs or oocysts of Eimeria, Dictyocaulus, and Moniezia were counted [45]. For sheep, the administration of plant extract significantly decreased the fecal egg count. After 20 days of treatment, eggs or oocysts of most parasites were not detected. It was concluded that the weekly administration of the plant extract can be effectively used to treat gastrointestinal and lung nematodes of small ruminants in both organic and traditional farming systems in tropical regions [45], however, the efficacy in the tested sheep was higher than in goats [45].
We have previously shown that silage made from S. acmophylla irrigated with saline water does not adversely affect milk quality or chemistry in goats [46], while feeding sheep willow silage made from trees irrigated with treated wastewater led to more tender meat [47]. This report is the first to explore the potential of using S. acmophylla extracts as bioactive compounds for the biological control of coccidiosis in ruminants, highlighting the significance of genotype-specific variations in secondary metabolite profiles. It demonstrates that all parts of S. acmophylla can provide secondary metabolites that act as coccidiostats to treat Eimeria in ruminants. Future research should investigate the effect of the irrigation source of willow (saline or treated wastewater vs fresh water) on secondary metabolites and their impact on Eimeria, similar to studies conducted with nematodes [26]. Additionally, the Israeli ecotypes within each genotype warrant further examination, as well as the validation of our in vitro results in vivo. The utilization of secondary metabolites could offer an environmentally friendly approach to controlling parasitic coccidia in animals while maintaining or improving milk and meat quality.
5. Conclusions
At a concentration of 750 ppm, leaf or branch extracts from the three Jordanian S. acmophylla genotypes inhibited coccidia sporulation by more than 70%. These inhibition percentages increased to over 80% at 1250 ppm and exceeded 90% at 5000 ppm. The phenolic composition of each extract was found to be sufficient for effective sporulation inhibition. Although this study didn’t pinpoint the specific phenolic and flavonoids responsible for sporulation inhibition, it suggests that using either branches or leaves can achieve a comparable result. Leaves and branches from willow plants of the three different genotypes can be collectively utilized to derive secondary metabolites acting as a coccidiostat to treat Eimeria.
Acknowledgments
This work was partially supported by MERC (Middle East Regional Cooperation - U.S. Agency for International Development), grant M39-014.
Disclosures
No conflicts of interest are declared.
References
- Lauron-Moreau, A.; Pitre, F.E.; Argus, G.W.; Labrecque, M.; Brouillet, L. Phylogenetic relationships of American willows (Salix L., Salicaceae). PLoS One 2015, 10, e0121965. [Google Scholar]
- Argus, G.W. Salix (Salicaceae) distribution maps and a synopsis of their classification in North America, north of Mexico. Harv Pap Bot. 2007, 12, 335–369. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Richardson, J. Poplars and willows: trees for society and the environment. Boston, USA: The Food and Agriculture Organization of the United Nations and CABI, 2014, 634.
- Mahdi, J.G. Medicinal potential of willow: A chemical perspective of aspirin discovery. J Saudi Chem Soc. 2010, 14, 317–322. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Knee, M.; Quigley, M.F. Effects of soil compaction and flooding on the growth of 12 willow (Salix L.) species. J Environ. Hort. 2004, 22, 155–160. [Google Scholar] [CrossRef]
- Muklada, H.; Klein, J.; Glasser, T.; Dvash, L.; Azaizeh, H.; Halabi, N.; Davidovich-Rikanati, R.; Lewinsohn, E.; Landau, S.Y. Initial evaluation of willow (Salix acmophylla) irrigated with treated wastewater as a fodder crop for dairy goats. Small Rum Res 2017, 163, 783. [Google Scholar] [CrossRef]
- Larsen, S.U.; Lærke, P.E.; Jørgensen, U. Harvest of green willow biomass for feed - effects of harvest time and frequency on yield, nutrient concentration, silage quality and regrowth. Acta Agri Scand 2020. [Google Scholar] [CrossRef]
- Saller, R.; Melzer, J.; Felder, M. Pain relief with a proprietary extract of Willow bark in rheumatology. An Open Trial Schweiz Zschr GanzheitsMedizin 2008, 20, 156–162. [Google Scholar] [CrossRef]
- Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B. Phenolic glycosides of the Salicaceae and their role as anti-herbivore defenses. Phytochem. 2011, 72, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.R.; Aboud, A.S. Antimicrobial Activities of Aqueous and Methanolic Extracts from Salvia officinalis and Salix acmophylla Used in the treatment of wound infection isolates. Ibn AL-Haitham J.l for Pure and Appl Sci. 2017, 23, 25–39. [Google Scholar]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochem. 2003, 64, 3–19. [Google Scholar] [CrossRef]
- El-Shazly, A.; El-Sayed, A.; Fikrey, E. Bioactive secondary metabolites from Salix tetrasperma Roxb. Z Naturforsch 2012, 67, 353–359. [Google Scholar] [CrossRef]
- Gan, R.Y.; Chan, C.L.; Yang, Q.Q.; Li, H.B.; Zhang, D.; Ge, Y.Y.; Corke, H. Bioactive compounds and beneficial functions of sprouted grains. In: Sprouted Grains, 2019, 91-246. AACC International Press.
- Lindroth, R.L.; Pajutee, M.S. Chemical analysis of phenolic glycosides: art, facts, and artifacts. Oecologia 1987, 74, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, B.; Kahlich, R.; Biegert, C.; Gleiter, C.H.; Heide, L. HPLC-MS/MS analysis of willow bark extracts contained in pharmaceutical preparations. Phytochem Anal. 2005, 16, 470–478. [Google Scholar] [CrossRef]
- Li, D.; Fu, Y.; Zhang, W.; Su, G.; Liu, B.; Guo, M.; Li, F.; Liang, D.; Liu, Z.; Zhang, X.; et al. Salidroside attenuates inflammatory responses by suppressing nuclear factor-κB and mitogen-activated protein kinases activation in lipopolysaccharide-induced mastitis in mice. Inflamm Res. 2013, 62, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Farney, J.K.; Mamedova, L.K.; Coetzee, J.F.; Minton, J.E.; Hollis, L.C.; Bradford, B.J. Sodium salicylate treatment in early lactation increases whole-lactation milk and milk fat yield in mature dairy cows. J Dairy Sci. 2013, 96, 7709–7718. [Google Scholar] [CrossRef] [PubMed]
- Nyman, T.; Julkunen-Tiitto, R. Chemical variation within and among six northern willow species. Phytochem. 2005, 66, 2836–2843. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Z.; Zhang, X.F.; Wang, L.J.; Zheng, Y.N.; Yuan, C.C.; Sun, G.Z. Isolation and characterization of phenolic compounds from the leaves of Salix matsudana. Molec. 2008, 13, 1530–1537. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, G.M.; Hussien, N.N.; Marzoog, T.R.; Awad, H.A. Phenolic content, antioxidant, antimicrobial and cytotoxic activities of ethanolic extract of Salix alba. Am J Biochem Biotechnol. 2013, 9, 41–46. [Google Scholar] [CrossRef]
- El-Wakil, E.; El-Sayed, S.; El-Sayed, M.M.; Abdel-Lateef, E. Identification of the chemical composition of the methanolic extract of Salix tetrasperma Roxb. using LC-ESI-MS and evaluation its potential as antioxidant agent. Der Pharma Chem 2015, 7, 168–177. [Google Scholar]
- Hoste, H.; Jackson, F.; Athanasiadou, S.; Thamsborg, S.M.; Hoskin, S.O. The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends in parasitol. 2006, 22, 253–261. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Unraveling the conundrum of tannins in animal nutrition and health. J Sci of Food and Agri. 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Hernandez, P.M.; Salem, A.Z.M.; Elghandour, M.Y.; Cipriano-Salazar, M.; Cruz-Lagunas, B.; Camacho, L.M. Anthelmintic effects of Salix babylonica L. and Leucaena leucocephala Lam. extracts in growing lambs. Trop Animal Health and Produc. 2014, 46, 173–178. [Google Scholar] [CrossRef]
- Glazer, I.; Salame, L.; Dvash, L.; Muklada, H.; Azaizeh, H.; Mreny, R.; Landau, S. Effects of tannin-rich host plants on the infection and establishment of the entomopathogenic nematode Heterorhabditis bacteriophora. J inver pathol. 2015, 128, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Awwad, S.; Markovics, A.; Halahlih, F.; Yazbak, A.; Haj-Zaroubi, M.; Muklada, H.; Klein, J.D.; Azaizeh, H. Effect of irrigation water source on secondary metabolites in Salix acmophylla and their potential to impair exsheathment of gastro-intestinal nematodes. Ann Agric Crop Sci. 2021, 6, 1079. [Google Scholar] [CrossRef]
- Rottenberg, A.; Zohary, D.; Nevo, E. Patterns of isozyme diversity and vegetative reproduction of willows in Israel. Int J Plant Sci. 1999, 160, 561–566. [Google Scholar] [CrossRef]
- Muklada, H.; Davidovich-Rikanati, R.; Wilkerson, D.G.; Klein, J.D.; Deutch-Traubaum, T.; Zou, J.; Awabdeh, S.; Sweidan, R.; Landau, S.Y.; Schwartz, A.; et al. Genotypic diversity in willow (Salix spp.) is associated with chemical and morphological polymorphism, suggesting human-assisted dissemination in the Eastern Mediterranean. Biochem Syst Ecol 2020, 91, 104081. [Google Scholar] [CrossRef]
- Hussein Muklada, H.; Davidovich-Rikanati, R.; Awabdeh, S.; Weinberg, Z.G.; Hen, Y.; Deutch, T.; Klein, J.D.; Voet, H.; Lewinsohn, E.; Landau, S.Y. Ensiling willow (Salix acmophylla) fodder modifies the contents of plant specialized metabolites, but not nutritional attributes. Animal Feed Sci and Technol. 2021, 278, 115019. [Google Scholar] [CrossRef]
- Keeton, S.T.N.; Navarre, C.B. Coccidiosis in Large and Small Ruminants. Veterinary Clinics of North America: Food Animal Pract. 2018, 34, 201–208. [Google Scholar] [CrossRef]
- Bangoura, B.; Bardsley, K.D. Ruminant Coccidiosis. Veterinary Clinics of North America: Food Animal Pract. 2020, 36, 187–203. [Google Scholar] [CrossRef]
- Mohamaden, W.I.; Sallam, N.H.; Abouelhassan, E.M. Prevalence of Eimeria species among sheep and goats in Suez Governorate, Egypt. Int J Vet Sci and Med. 2018, S2314459918300012. [Google Scholar] [CrossRef]
- Juszczak, M.; Sadowska, N.; Udała, J. Parasites of the digestive tract of sheep and goats from organic farms in Western Pomerania. Poland Ann Parasitol. 2019, 65, 245–250. [Google Scholar] [PubMed]
- Rivero-Perez, N.; Hernández-Alvarado, J.L.; Valladares-Carranza, B.; Delgadillo-Ruiz, L.; Ojeda-Ramírez, D.; Sosa-Gutiérrez, C.G.; Morales-Ubaldo, A.L.; Vega-Sanchez, V.; Zaragoza-Bastida, A. Salix babylonica L. as a Natural Anticoccidial Alternative in Growing Rabbits. Evid Based Complement Altern Med. 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Azaizeh, H.; Halahleh, F.; Abbas, N.; Markovics, A.; Muklada, H.; Ungar, E.D.; Landau, S.Y. Polyphenols from Pistacia lentiscus and Phillyrea latifolia impair the exsheathment of gastro-intestinal nematode larvae. Vet Parasitol. 2013, 191, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. Determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Warrier, R.; Paul, M.; Vineetha, M. Estimation of salicylic acid in Eucalyptus leaves using spectrophotometric methods. Gen Plant Physiol. 2013, 3, 90–97. [Google Scholar]
- Zhang, G.; Li He, L.; Hu, M. Optimized ultrasonic-assisted extraction of flavonoids from Prunella vulgaris L. and evaluation of antioxidant activities in vitro. Innov. Food Sci. Emerg Technol. 2011, 12, 18–25. [Google Scholar] [CrossRef]
- Ekawasti, F.; Nurcahyo, W.; Wardhana, A.H.; Shibahara, T.; Tokoro, M.; Sasai, K.; Matsubayashi, M. Molecular characterization of highly pathogenic Eimeria species among beef cattle on Java Island, Indonesia. Parasitol Int. 2019, 72, 101927. [Google Scholar] [CrossRef]
- Tawfeek, N.; Mahmoud, M.F.; Hamdan, D.I.; Sobeh, M.; Farrag, N.; Wink, M.; El-Shazly, A.M. Phytochemistry, pharmacology and medicinal uses of plants of the genus Salix: An updated review. Front. Pharmacol. 2021, 12, 593856. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R. Phenolic constituents of Salix: a chemotaxonomic survey of further Finnish species. Phytochem. 1989, 28, 2115–2125. [Google Scholar] [CrossRef]
- Barnes, J.; Anderson, L.A.; Phillipson, J.D. Herbal Medicines; Royal Pharmaceutical Society: London, UK, 2002. [Google Scholar]
- Shah, Z.A.; Hameed, A.; Ahmed, A.; Simjee, S.U.; Jabeen, A.; Ullah, A.; Shaheen, F. Cytotoxic and anti-inflammatory salicin glycosides from leaves of Salix acmophylla. Phytochem Lett. 2016, 17, 107–113. [Google Scholar] [CrossRef]
- Donaldson, J.R.; Lindroth, R.L. Genetics, environment, and their interaction determine efficacy of chemical defense in trembling aspen. Ecol. 2007, 88, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.Z.M.; Elghandour, M.M.Y.; Kholif, A.E. Tree leaves of Salix babylonica extract as a natural anthelmintic for small-ruminant farms in a semiarid region in Mexico. Agrofores Syst. 2016, 91, 111–122. [Google Scholar] [CrossRef]
- Landau, S.Y.; Glasser, T.A.; Zachut, M.; Klein, J.D.; Deutch-Traubman, T.; Voet, H.; Kra, G.; Davidovich-Rikanati, R. Milking performance and plant specialized metabolites in the milk of goats fed silage from willow (Salix acmophylla) irrigated with saline water. Livestock Sci. 2023, 270, 105205. [Google Scholar] [CrossRef]
- Awabdeh, S.; Sweidan, R.; Landau, S.Y. Growth performance and carcass characteristics of fattening Awassi lambs fed willow silage. Small Rum Res. 2022, 215, 106758. [Google Scholar] [CrossRef]

Table 1.
Ethanol (70%) extraction percentage and chemical components in willow leaves and branches of different S. acmophylla genotypes.
Table 1.
Ethanol (70%) extraction percentage and chemical components in willow leaves and branches of different S. acmophylla genotypes.
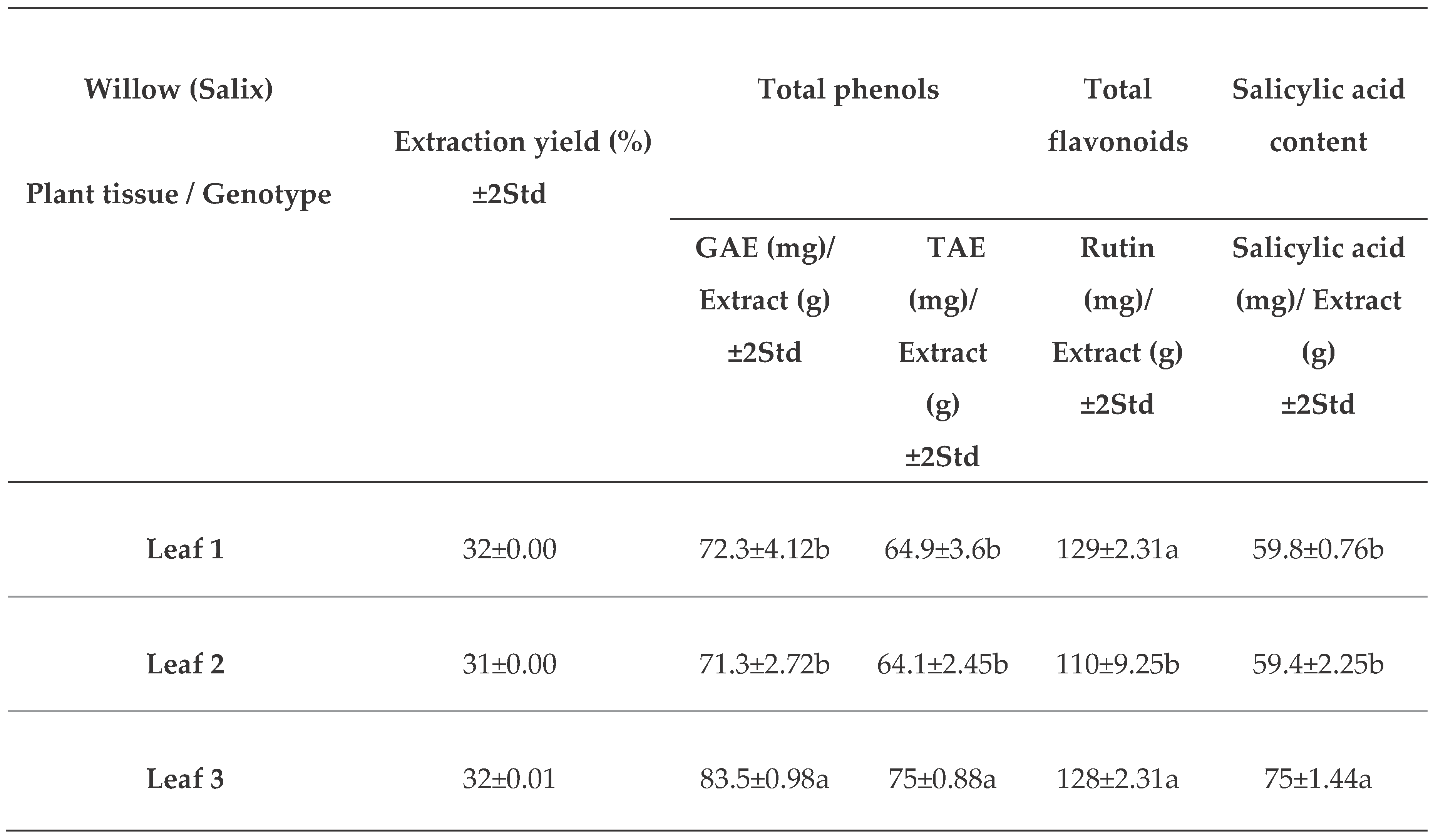

GAE - gallic acid equivalent; TAE- tannic acid equivalent. Means and standard deviation of two experiments, each with n=3, mean separation in each tissue by Tukey’s HSD test p< 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



