
Submitted:
15 March 2024
Posted:
15 March 2024
You are already at the latest version
Abstract
During choriogenesis in insects, chorion (eggshell) is formed by surrounding follicular epithelial cells in ovarioles. However, the regulatory endocrine factor(s) activating choriogenesis and the chemical components on eggshell deserve further exploration. In two representative Coleopterans, a Coccinellid Henosepilachna vigintioctopunctata and a Chrysomelid Leptinotarsa decemlineata, genes encoding the 20-hydroxyecdysone (20E) receptor heterodimer, ecdysone receptor (EcR) and ultraspiracle (USP), and two chitin biosynthesis enzymes UDP-N-acetylglucosamine pyrophosphorylase (UAP) and chitin synthase (ChS1), were highly expressed in ovaries of the young females. RNA interference (RNAi)-aided knockdown of either HvEcR or Hvusp in H. vigintioctopunctata inhibited oviposition, suppressed the expression of HvChS1, and lessened the positive signal of Calcofluor staining on the chorions, which suggests the reduction of a chitin-like substance (CLS) deposited on eggshells. Similarly, RNAi of LdEcR or Ldusp in L. decemlineata constrained oviposition, decreased the expression of LdUAP1 and LdChS1, and declined CLS contents in the resultant ovaries. Knockdown of LdUAP1 or LdChS1 caused similar defective phenotypes, i.e., reduced oviposition and CLS contents in the L. decemlineata ovaries. These results, for the first time, indicate that 20E signaling activates choriogenesis in two Coleopteran species. Moreover, our findings propose the deposition of a CLS on the chorions.
Keywords:
Leptinotarsa decemlineata
; Henosepilachna vigintioctopunctata
; choriogenesis
; ecdysone signal
; chitin-like substance
; eggshell
1. Introduction
The development of oocytes (oogenesis) in insect ovarioles consists of previtellogenesis, vitellogenesis and choriogenesis stages. Among the three successive physiological processes, choriogenesis is the final developmental period towards oocyte maturation when the chorion (eggshell) is formed by the surrounding follicular epithelial cells [1]. Up to now, however, the regulatory endocrine factor(s) activating choriogenesis and the chemical components on eggshell have not been well explored.
Endocrine control of oogenesis has widely been reported in insects [2,3,4]. Nevertheless, all these publications are focused on the stimulation of previtellogenesis and vitellogenesis. Three classical hormones, ecdysteroids (20-hydroxyecdysone, 20E, the most active form), juvenile hormone (JH) and insulin-like peptides (ILP), have been confirmed as gonadotropins [2,5]. In Tribolium castaneum, for instance, application of JH III induces vitellogenin (Vg) mRNA in the previtellogenic females, whereas suppressing JH signal impairs Vg gene expression and Vg accumulation [5]. Similarly, RNAi studies show that 20E receptor heterodimer, ecdysone receptor and ultraspiracle genes (EcR and usp), and 20E downstream genes (E75, E93, HR3, HR4 and βFTZ-F1) are required for vitellogenesis and oocyte maturation [6,7]. Moreover, bursicon neuropeptide (downstream 20E component) also triggers Vg expression, possibly through activation of JH and ILP pathways [8]. Up to now, whether the same endocrine pathways are involved in the government of choriogenesis remains unclear. In fact, some reseachers believe that insect choriogenesis is autonomously initiated when an oocyte has been developed to a certain vitellogenetic phase [9].
As for chemicals on insect eggshells, the most important component is proteins [1]. In the Drosophila melanogaster eggshells, for instance, more than 30 protein components are assembled [10,11]. Similarly, in the chorions of the superfamily Bombycoidea insects, more than 100 different polypeptides are accumulated in a stepwise fashion [1,12]. Intriguingly, a chitin-like substance (CLS) accumulates in oocytes during oogenesis, it is also present in ovaries, newly laid and dark eggs in Aedes aegypti [13]. Whether the CLS is an eggshell structural component during oogenesis needs to be established in insects other than A. aegypti [13].
Chitin biosynthesis has well been investigated in insects. UDP-N-acetylglucosamine pyrophosphorylase (UAP) and chitin synthases (ChS) catalyze the final two consecutive biochemical steps [14,15,16,17]. In most insects, ChSs have been segregated into two groups, namely, ChS1 and ChS2. ChS1 is exclusively distributed in the ectodermally-derived epidermal cells, it is responsible for chitin generation in epidermis, trachea, salivary gland, foregut and hindgut cells. ChS2 is specifically located in midgut cells and is dedicated to chitin formation in the peritrophic matrix (PM) [14,15,16,17]. In Leptinotarsa decemlineata larvae, UAPs are also encoded by UAP1 and UAP2 genes. UAP1 and UAP2 specifically catalyze chitin production in ectodermally-derived cells and midgut PM. Moreover, chitin synthesis is tightly controlled by 20E during larval development, through binding to EcR/USP heterodimer in L. decemlineata [17,18]. Providing that chitin biosynthesis is strictly coordinated within the cycle of insect molts [19,20], the biochemical process should also be governed by 20E cascade during metamorphosis in other insect species.
A Chrysomelid L. decemlineata and a Coccinellid Henosepilachna vigintioctopunctata are Coleopteran potato defoliators. Our previous results reveal that vitellogenesis occurs in the 5- to 7-day-old females, while the choriogenesis takes place in the 8- and 9-day-old adults. Copulation and oviposition happen before and at the age of 8 days old in H. vigintioctopunctata [21]. RNA interference (RNAi) of either EcR or usp inhibits oocyte development, dramatically represses the transcription of Vg in fat bodies and of VgR in ovaries in the two Coleopterans [22]. In the current paper, we performed the same RNAi experiments in the newly-emerged adult females. We intended to test two hypotheses. Firstly, 20E cascade mediates choriogenesis. Secondly, a CLS is deposited on oocyte chorion. Our findings supported the two assumptions.
2. Results
2.1. The Temporal Expression of Related Genes
Using qRT-PCR, the temporal expression patterns of two 20E receptor genes (EcR and usp) and three chitin biosynthesis genes (UAP, ChS1 and ChS2) were respectively measured in the 0- to 9-, and 0- to 13-day-old ovaries in L. decemlineata and H. vigintioctopunctata (Figure 1). All 5 genes in L. decemlineata (LdEcRA, LdEcRB1, Ldusp, LdUAP1, LdChS1, LdChS2) and 4 genes in H. vigintioctopunctata (HvEcRA, HvEcRB1, HvUAP, HvChS1, HvChS2) were highly expressed in the young adults (at the ages of 0 to 5 days old), whereas Hvusp was constantly transcribed in the 0- to 13-day-old H. vigintioctopunctata ovaries (Figure 1).
2.2. Knockdown of HvEcR Prevents Oviposition in H. vigintioctopunctata
Three days after dsHvEcR injection, the HvEcR level in the ovary was significantly reduced (Figure 2A). For the dsegfp-injected beetles, no female laid eggs until 8 days after adult emergence. They then continuously oviposited. In contrast, the dsHvEcR-injected females did not lay any eggs until 20 days post adult emergence. They then laid only turbid liquid (Figure 2B versus Figure 2C).
Consistent with failure of oviposition, the 30-day-old HvEcR RNAi females possessed larger body sizes and heavier fresh weights than the 30-day-old controls (Figure 2D–J). The mature eggs in the 30-day-old control ovaries were ellipse-shaped (Figure 2K). The average major axis and minor axis were 1.30 mm and 0.50 mm respectively. The ratio of major/minor axis was 2.7 (Figure 2M–O). The eggs in the 30-day-old HvEcR RNAi ovaries were misshapen, nearly globular (Figure 2L); the mean major axis, minor axis and ratio of major/minor axis were 0.85 mm, 0.57 mm and 1.5 respectively (Figure 2M–O).
2.3. Depletion of Hvusp Affects Oogenesis in H. vigintioctopunctata
Injection of dsHvusp into the 0-day-old females successfully knocked down the target gene in the ovary (Figure 3A). Whereas the control females more than 8 days old continuously laid eggs (Figure 3B), the Hvusp RNAi females did not lay any eggs until around 20 days post eclosion. They then laid only turbid liquid (Figure 3C). Interestingly, some Hvusp RNAi females more than 40 days old could lay a few normal eggs daily, along with turbid liquid (Figure 3D).
Similar to the 30-day-old HvEcR RNAi females, the 30-day-old Hvusp RNAi females had larger body sizes and heavier fresh weights than the 30-day-old controls (Figure 3E–G).
In the 20-day-old Hvusp RNAi ovarioles, there were two kind of mature eggs. Many misshapen eggs were accumulated in the median oviduct and the genital chamber, whereas one or two normally-shaped eggs were sited in the lateral oviduct (Figure 3H).
The mature eggs in the 20-day-old control ovaries were ellipse-shaped (Figure 3I). The average major axis and minor axis were 1.25 mm and 0.45 mm respectively. The ratio of major/minor axis was 2.8 (Figure 3K–M). Conversely, the eggs in the Hvusp RNAi ovarioles were misshapen (Figure 3J); the mean major axis, minor axis and ratio of major/minor axis were 1.30 mm, 0.70 mm and 1.8 respectively (Figure 3K–M).
2.4. RNAi of HvEcR Affected CLS Deposition on H. vigintioctopunctata Oocytes
Our previous results have documented the identification of two chitin synthase genes (HvChS1 and HvChS2) in H. vigintioctopunctata [15]. Here we detected their expression levels in the 10- and 20-day-old ovaries. In contrast to HvChS2 that is only expressed in the midgut [15], the transcript levels of HvChS1 were significantly reduced in the HvEcR silenced samples, compared with these in the control ovaries (Figure 4A–D).
HE stained sections of ovaries displayed that the maturing eggs in the control ovarioles had smooth and continuous chorions (Ch) (Figure 4E), whereas the eggshells in the HvEcR RNAi developing eggs were irregular and discontinuous (Figure 4F).
The presence of CLS was shown by Calcofluor staining [23]. The fluorescence images of frozen sections displayed that two layers of positive-stained membranes (↑) were observed in the control ovarioles, one lining the follicle cells (FC) and the other being deposited to the chorion (Ch) (Figure 4G,I). Conversely, only the positive-stained layer along the follicle cells was seen in the dsHvEcR treated ovariole samples (Figure 4H,J).
2.5. Silence of Hvusp Blocked CLS Deposition on H. vigintioctopunctata Oocytes
CLS deposition on the Hvusp RNAi H. vigintioctopunctata oocytes was also compared (Figure 5). The expression levels of HvChS1 were significantly diminished in the 10- and 20-day-old Hvusp silenced ovaries, compared with these in the control specimens (Figure 5A,B). In contrast, the transcript levels of HvChS2 varied little between Hvusp RNAi and control samples (Figure 5C,D).
From HE stained sections, it was easily noted that the normal maturing eggs had smooth and continuous chorions (Ch) (Figure 5E). Conversely, the egg shells in the Hvusp RNAi eggs were irregular and discontinuous. Some yolk substances were seeped out (^) (Figure 5F).
The fluorescence images of frozen sections stained with Calcofluor exhibited two layers of positively-stained membranes (↑) in the follicle cells (FC) and chorion (Ch) (Figure 5G). Conversely, the two positively-stained layers were almost disappeared in the dsHvusp treated ovarian specimens (Figure 5H).
2.6. Shortage of 20E Signal Prevents CLS Contents in L. decemlineata Ovaries
In the newly-eclosed L. decemlineata females, we repeated the RNAi experiments and examined the defective phenotypes (Figure 6).
Three days after dsLdEcR or dsLdusp injection, the target genes LdEcR (Figure 6A) and Ldusp (Figure 6I) in the ovaries were respectively silenced.
When the 9-day-old control L. decemlineata females normally oviposited (Figure 6B), the 15-day-old LdEcR and Ldusp RNAi females laid turbid liquid, along with a few deformed eggs (Figure 6C). These misshapen eggs had shrinked eggshells and wet surface (Figure 6C).
The expression of LdUAP1, LdUAP2, LdChS1 and LdChS2 in the 10-day-old ovaries was determined. The transcript levels of LdUAP1 and LdChS1, but not LdUAP2 and LdChS2, were significantly lowered in the LdEcR (Figure 6D–G) or Ldusp (Figure 6J–M) RNAi ovaries.
The chitin contents in the 10-day-old LdEcR (Figure 6H) or Ldusp (Figure 6N) RNAi ovaries were tested. As expected, the chitin amounts were greatly declined in the treated ovaries when evaluated by N-acetylglucosamine (GlcNAc) concentrations using the chitinase degraded method, compared with control specimens.
2.7. Chitin Defeciency Causes Similar Defects on L. decemlineata Oocytes
To determine whether chitin shortage causes similar defects on oocytes to those in the LdEcR or Ldusp RNAi ovaries, we knocked down LdUAP1, LdUAP2, LdChS1, or LdChS2 by injected corresponding dsRNA into 4-day-old female adults (Figure 7A–D). Introduction of dsLdUAP1, dsLdUAP2, dsLdChS1, or dsLdChS2 significantly lowered its target mRNA when detected 3 days post treatment, but displayed little influences on non-target transcripts (Figure 7A–D).
Depletion of LdUAP1 and LdChS1 significantly reduced chitin contents in the ovaries when measured with GlcNAc concentrations using the chitinase degraded method, in contrast to control, and LdUAP2 and LdChS2 RNAi specimens (Figure 7E). Moreover, no mature oocytes were seen in 10-day-old LdUAP1 and LdChS1 RNAi ovaries, in contrast to the control, and LdUAP2 and LdChS2 RNAi samples (Figure 7G,I versus Figure 7F,H,J). Twenty days after treatment, the LdUAP1 and LdChS1 RNAi female adults began to lay turbid liquid, along with a few deformed eggs (Figure 7K).
3. Discussion
3.1. 20 E Signal Triggers Choriogenesis
After initiation of choriogenesis, chorion proteins and other structural components are deposited on eggshells. Whether 20E signal activates choriogenesis remains an opening question [9].
In the present paper, four pieces of experiment evidence were attained to indicate that 20E signal initiates choriogenesis in the two Coleopterans. Firstly, both EcR and usp were actively expressed immidiately after adult eclosion. The accumulated EcR and USP proteins may mediate 20E signaling and activate oogenesis. As a result, oocytes are continuously matured and the oviposition persists in mature H. vigintioctopunctata females [21]. Likewise, ecdysone response gene transcripts (E75, E74, BrC, HR3, HR4, and FTZ-F1) are mostly upregulated in the ovary during the first three days post blood meal in Rhodnius prolixus [24].
Secondly, our data showed that the EcR and usp transcripts were present in the ovaries in both H. vigintioctopunctata (Figure 1, Figure 2 and Figure 3) and L. decemlineata (Figure 1 and Figure 6) females, similar to those in R. prolixus [24,25].
Thirdly, knockdown of either EcR or usp significantly repressed oviposition in both H. vigintioctopunctata (Figure 2 and Figure 3) and L. decemlineata (Figure 6) females. In accordance with our data, knockdown of EcR, usp, E75, E74 or FTZ-F1 decreases mature eggs, and severely reduces the laid eggs and their hatching rate in R. prolixus [24,25]. The reduced oviposition in 20E signal repressed females has also been documented in T. castaneum [7], D. melanogaster [26], Nilaparvata lugens [27,28], and Cimex lectularius [29].
Lastly, we discovered that RNAi of either EcR or usp destroyed the structure of eggshells and deformed oocyte shapes. Consistently, some of the eggs have an irregular shape and smaller volume laid by the EcR, usp, E75, E74 or FTZ-F1 RNAi R. prolixus females [24,25]. The deformation of chorion structure and oocyte shape gives a solid piece of experiment evidence that 20E cascade triggers choriogenesis, in addition to activation of vitellogenesis in both H. vigintioctopunctata and L. decemlineata females [22].
3.2. 20E Signal Triggers the Production of a CLS
In the current article, our findings suggest that 20E signal triggers the production of a chitin-like substance (CLS), a component critical for the formation of oocyte chorion during choriogenesis in the two Coleoptera species.
Firstly, UAP and ChA catalyze the last two biochemical steps for chitin biosynthesis in L. decemlineata [16,17] and H. vigintioctopunctata [14,15]. In the present paper, expression analysis showed that their encoding genes were actively expressed in the adult ovaries (Figure 1, Figure 4, Figure 5, Figure 6 and Figure 7). Moreover, the expression of either UAP or ChS is strictly coordinated with the transcription of 20E receptor gene EcR in both Coleopterans (Figure 1). Similar expression patterns between chitin biosynthesis and 20E cascade genes have widely been documented during larval development in representative insect species [17,18,19,20].
Secondly, biochemical analysis demonstrated that the CLS was present in L. decemlineata ovaries (Figure 6), similar to those on A. aegypti eggshells [13] and in R. prolixus ovaries [30]. Moreover, the Calcofluor stain [23] showed that two layers of CLS were observed in H. vigintioctopunctata ovaries, one lining the follicle cells and the other in the eggshells (Figure 4 and Figure 5). Follicle cells are ectodermally-derived epidermal tissue. Thus, our data are in agreement with the common notion that ChS1 locates in the ectodermally-derived epidermal cells while ChS2 in midgut cells [14,15,16,17].
Thirdly, our findings showed that knockdown of either EcR or usp significantly lowered the expression levels of UAP and ChS1, but not ChS2, in the two Coleopterans (Figure 4, Figure 5 and Figure 6). Consistently, suppression of 20E signal reduced the CLS contents in L. decemlineata ovaries (Figure 6) and decreased the positive reaction to Calcofluor stain in H. vigintioctopunctata follicle cells and eggshells (Figure 4 and Figure 5). Similarly, a CLS has been reported to be present in oocytes in a Diptera species [13]. Recently, a specific mucin protein has been documented to contribute to the formation of eggshell in N. lugens [31,32] and Spodoptera exigua [33]. Moreover, enzymes involving in chorion tanning are reported in D. melanogaster [34,35] and N. lugens [36] eggshells. Given that intestinal mucins exhibit strong association with chitin [37] and tanning process involves the oxidative conjugation and cross-linking of cuticular proteins and chitin by quinones [38], it is reasonably assumed that many insect eggshells contains chitin. Further researches will shed light on this issue.
Fourthly, knockdown of LdUAP1 and LdChS1, rather than LdUAP2 and LdChS2, caused similar defects on L. decemlineata oocytes to those in the EcR or usp RNAi beetles (Figure 7). We accordingly propose that dysfunction of EcR or USP inhibits CLS accumulation on the eggshells in L. decemlineata. As a result, the structure of eggshells is destroyed and the oocyte shapes are deformed (Figure 2, Figure 3 and Figure 6). Interestingly, dsChS treatment affects oviposition in R. prolixus [30]. Knockdown of ChS1 disrupts adult fecundity in Aphis gossypii [39]. Moreover, dietary introduction of chitin synthesis inhibitors declines fecundity in A. aegypti [13]. In addition, RNAi of LsChS1 in pre-adult I results in deformation of ovaries and oocytes in adult Lepeophtheirus salmonis [40]. This result suggests the likelihood that CLS may also be present in eggshells and deficiency of CLS causes misshapen eggs in other arthropods. Nevertheless, whether 20E regulation of CLS deposition on oocyte chorion is a widely conservative way among arthropods deserves further investigation to determine.
In the current paper, we injected dsLdUAP1 and dsLdChS1 into 4-day-old female adults, the target mRNAs were significantly reduced three days after treatment (Figure 7). Our previous observation showed that some oocytes have completed vitellogenesis and the patency among the follicle epithelium cells was closed in 5- to 7-day-old H. vigintioctopunctata females [21], similar to the oocyte maturation processes in other Coleopterans [2]. Moreover, after vitellogenesis oocytes are often arrested in prophase I of meiosis, showing very modest synthetic activity [41]. Therefore, it appears that only follicle epithelium cells can actively generate CLS in the 5- to 7-day-old H. vigintioctopunctata females and only the CLS biosynthesized by follicle epithelium cells can deposit on eggshells. Our findings support a common notion that the chorion is formed by the surrounding follicular epithelial cells during oocyte maturation [1].
3.3. Does 20E Signal Tune the Production of Chorion Proteins?
Comparison of the data in the current survey displays two dissimilarities. Firstly, even though inhibition of chitin deposition affected eggshell structure in the two Coleopterans after disruption of 20E signal, the defects of matured eggs are different. L. decemlineata eggs in treated females looked normal and could be deposited (Figure 6 and Figure 7), whereas H. vigintioctopunctata eggs were seriously deformed and failed to lay out (Figure 2 and Figure 3). Secondly, within H. vigintioctopunctata the influence of chitin deposition was more serious in the usp RNAi females (Figure 3), whereas the destruction of chorion and the delay of oviposition were more severe in the EcR RNAi females (Figure 2). The two dissimilarities suggest that 20E signal may also tune the production of chorion proteins. In accordance with the suggestion, EcRB1 isoform and USP are responsible for 20E-mediated transcriptional activation of the eggshell gene VM32E, which produces a protein component of the vitelline membrane and endochorion layers [42]. Similarly, silence of EcR, usp, or ecdysone biosynthesis gene shade significantly reduces the expression of chorion protein gene transcripts (Rp30 and Rp45) in R. prolixus [25].
If 20E signal stimulates the production of chorion proteins, as suggested above, the difference of eggshell phenotypic defects between EcR and usp RNAi females implies that EcR and USP respectively regulate a specific subset of chorion protein genes, dependent or independent of EcR/USP complex (Figure 7L). In this context, both EcR and USP exert specific atypical function independent of EcR/USP complex. For EcR, it may dimerize with other partners, form homodimers [43] and/or form protein complexes that have yet to be described [44]. In D. melanogaster male adults, for example, EcR-depleted (but not USP-depleted) accessory glands fail to make seminal proteins and have dying cells. The active receptor may be a homodimer [45]. A similar paradigm is observed in the expression of genes encoding glue proteins in the salivary glands [44]. Interestingly, EcR apparently does not require USP as a heterodimeric partner in scorpions [46]. As for USP, it is able to form homomeric complexes in living cells [43]. It is also bound to other nuclear receptors [47]. In fact, some functions are independent of EcR have been explored in D. melanogaster and A. aegypti USPs [47,48]. In A. aegypti, for instance, the interaction of Svp and USP rather than binding competition for the Vg ecdysteroid response element accounts for the inhibition of Vg expression after a batch of eggs is laid [49]. This issue deserves further investigations.
4. Materials and Methods
4.1. Insect Rearing
Both H. vigintioctopunctata and L. decemlineata beetles were cultured in laboratory at 28 ± 1°C, 16 h:8 h light–dark photoperiod and 50-60% relative humidity conditions, using the fresh potato leaves, using foliage at the vegetative growth or young tuber stages from potato plants (Solanum tuberosum L) growing in insecticide-free soil. Under this feeding protocol, the duration from laying eggs to emergence adults of the next generation was about one month.
4.2. Synthesis of dsRNAs
For H. vigintioctopunctata, a highly efficient and specific siRNA fragment originated from HvEcR or Hvusp was selected using the siRNA online design website (http://sidirect2.rnai.jp/). Then, a pair of primers was designed using the software primer premier 5.0 (Table S1) to amplify a cDNA including the siRNA fragment. The targeted cDNA sequence was further BLASTN searched against the H. vigintioctopunctata transcriptome data to identify any possible off-target sequences that had an identical match of 20 bp or more. For L. decemlineata, the same dsLdEcR and dsLdusp were used [50,51]. Moreover, a cDNA fragment was derived from enhanced green fluorescent protein (egfp) in Aequorea victoria. These fragments were respectively amplified by PCR using specific primers (Table S1) conjugated with the T7 RNA polymerase promoter. The dsRNAs were synthesized using the MEGAscript T7 High Yield Transcription Kit (Ambion, Austin, TX, USA) according to the manufacturer’s instructions. Subsequently, the synthesized dsRNA (at a concentration of 5-8 μg/μl) was determined by agarose gel electrophoresis and the Nanodrop 1000 spectrophotometer (data not shown) and kept at -80°C until use.
4.3. Injection of dsRNAs
The same method described previously was used to inject dsRNA into 0-day-old H. vigintioctopunctata and L. decemlineata female adults [52,53]. Briefly, an aliquot (0.1 μl) of the solution including 400 ng dsRNA was injected into the newly emerged female adults. Negative control newly emerged female adults were injected with the same volume of dsegfp solution.
For 0-day-old H. vigintioctopunctata female adults, two biologically independent experiments were carried out using different generations, each bioassay setting two treatments: (1) dsegfp and (2) dsHvEcR or dsHvusp. A group of 10 injected newly emerged female adults was set as a replicate. The resultant newly emerged female adults were allowed to feed potato foliage until the death. Each treatment had 24 replicates. A total of 9 replicates were harvested 3, 10 and 20 days after treatment to dissect their ovaries. Three replicates were sampled 3 days after injection for qRT-PCR to test RNAi efficacy in the ovaries. The other 6 replicates were used to measure the expression levels of genes involved in chitin biosynthesis (HvChS1 and HvChS2) in the ovaries. Three replicates were used to record oviposition, to measure body size and to detect fresh weight. Another 12 replicates were collected, dissected for observation under a microscope or stained using HE or Calcofluor 10, 20, 30 or 40 days after injection.
For 0-day-old L. decemlineata female adults, two biologically independent experiments were carried out using different generations, with two treatments: (1) dsegfp and (2) dsLdEcR or dsLdusp. A group of 10 injected newly emerged female adults was set as a replicate. The resultant newly emerged female adults were allowed to feed potato foliage until the death. Each treatment had 12 replicates. A total of 6 replicates were harvested 3 and 10 days after treatment to dissect their ovaries. Three replicates were sampled 3 days after injection for qRT-PCR to test RNAi efficacy in the ovaries. Three replicates at the age of 10 days old were used to measure the expression levels of genes involved in chitin biosynthesis (LdUAP1, LdUAP2, LdChS1 and LdChS2) in the ovaries. Three replicates were used to record oviposition. Another three replicates were collected 10 days after injection, dissected the ovaries to determine chitin content.
For 4-day-old L. decemlineata female adults, a bioassay was performed with five treatments: (1) dsegfp, (2) dsLdUAP1, (3) dsLdUAP2, (4) dsLdChS1 and (3) dsLdChS2. A group of 10 injected female adults was set as a replicate, and were allowed to feed potato foliage. Each treatment had 12 replicates. Three replicates were harvested 3 days after treatment for qRT-PCR to test RNAi efficacy, and for measurement of the expression levels of genes involved in chitin biosynthesis (LdUAP1, LdUAP2, LdChS1 and LdChS2) in the ovaries. Three replicates were used to record oviposition. Another six replicates were collected 10 days after injection, dissected the ovaries, and determined chitin content in the ovaries.
4.4. Quantitative Real-Time PCR
For temporal expression analysis, RNA templates were derived from 0-9 and 0-13 days old in L. decemlineata and H. vigintioctopunctata female adults respectively. Each sample contained 20-30 individuals and was repeated three times. For analysis of the effects of treatments, total RNA was extracted from treated female adults. Each sample contained 10 individuals and was repeated three times. The RNA was extracted using SV Total RNA Isolation System Kit (Promega, USA). Purified RNA was subjected to DNase I to remove any residual DNA according to the manufacturer’s instructions. Quantitative mRNA measurements were performed by qRT-PCR in technical triplicate, using 2 stably-expressed internal control genes (for L. decemlineata, LdRP18 and LdRP4; for H. vigintioctopunctata, HvRPS18 and HvRPL13; the primers listed in Table S1) according to the published results [54,55]. An RT negative control (without reverse transcriptase) and a non-template negative control were included for each primer set to confirm the absence of genomic DNA and to check for primer-dimer or contamination in the reactions, respectively. The primer pair for each gene was tested with a 5-fold logarithmic dilution of a cDNA mixture to generate a linear standard curve (crossing point plotted vs. log of template concentration), which was used to calculate the primer pair efficiency.
All primer pairs amplified a single PCR product with the expected sizes, showed a slope less than −3.0 and exhibited efficiency values ranging from 2.4 to 2.7. Data were analyzed by the 2−ΔΔCT method, using the geometric mean of the four internal control genes for normalization.
4.5. Hematoxylin-Eosin (HE) Staining
HE staining was performed to observe defective phenotypes in H. vigintioctopunctata ovaries. Briefly, the ovaries in the dsegfp- and dsHvusp-treated larvae were dissected 10, 20 and 30 days after the initiation of bioassay, and were then fixed in 4% paraformaldehyde and embedded in paraffin. The two embedded tissues were subsequently cut into 6-μm thick sections. The sections were stained using Mayer’s H&E (Yeasen, Shanghai, China) following a routine staining procedure and observed with an Olympus BH-2 light microscope (Olympus, Tokyo, Japan).
4.6. Chitin Staining with CALCOFLUOR
The H. vigintioctopunctata ovary sections prepared above were also stained with Calcofluor according to a previously described method [23]. Briefly, section preparations were incubated in 0.001% Calcofluor in 100 mm Tris-HCl, pH 9 for 30 min at room temperature. After washing off the excess Calcofluor, the fluorescence was recorded using light of appropriate excitation and emission wavelengths.
4.7. Chitin Analysis
The same method as described [16,17] was used to test chitin contents in 10-day-old L. decemlineata ovaries. Briefly, the samples were individually mixed with 0.5 g zirconium beads (0.7 mm diameter, BioSpec Products, Bartlesville, OK) and 0.5 ml 6% KOH, and were homogenized. The homogenates were then heated at 80 °C for 90 min, and were centrifuged at 12000 g for 20 min and the supernatants were removed. The pellets were individually suspended in 1 ml PBS, and centrifuged again at 12000 g for 20 min and the PBSs were discarded. Each pellet was then resuspended in 200 µl Mcllvaine’s buffer (0.1 M citric acid, 0.2 M NaH2PO4, pH 6) and 5 µl of Streptomyces plicatus chitinase-63 (5 mg/ml in PBS) was added to hydrolyze chitin to GlcNAc by incubation for 40 h at 37 °C.
GlcNAc concentrations were individually measured by a modified Morgan-Elson assay[56]. In a 0.2 ml PCR tube, 10 µl 0.27 M sodium borate and 10 µl of sample supernatant (12000 g, 1 min centrifugation) were combined. In a thermocycler, samples were heated to 99.9 °C for about 60 s, mixed gently, and incubated at 99.9 °C for 10 min. Immediately upon cooling to room temperature, 100 µl of diluted dimethylaminobenzaldehyde (DMAB) solution (10% w/v DMAB in 12.5 ml concentrated HCl and 87.5 ml glacial acetic acid stock, diluted 1 : 10 with glacial acetic acid) was added, followed by incubation at 37 °C for 20 min. Eighty µl of each sample was transferred to 96-well low-evaporation microtitre dishes, and the absorbance at 585 nm was recorded. Standard curves were prepared from stocks of 0.075-2.0 mM GlcNAc.
4.8. Data Analysis
The SPSS for Windows (Chicago, IL, USA) was used for statistical analyses. The averages (±SE) were compared using t-test.
5. Conclusions
Accordingly, a presumptive model is proposed for summarization of the regulation of 20E signal to choriongenesis in two representative Coleoptera species (Figure 7L). Our findings highlighted that 20E cascade triggers the deposition of chorion chemical components on the developing oocytes. Moreover, we uncovered that a CLS is critical for oocyte maturation. Our research provides a solid foundation for future studies on hormone regulation of reproduction in beetles.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Primers used in dsRNA synthesis and qRT-PCR.
Author Contributions
G.Q.L. developed the concepts. X.J.Z., L.J., K.Y.F. and W.C.G. performed the experiments. X.J.Z., K.Y.F. and L.J. analyzed the data. G.Q.L. and X.J.Z. discussed and wrote the manuscript.
Funding
This research was financially supported by the National Key Research and Development Program of China (2022YFC2601000).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data generated in association with this study are available in the Supplementary Materials published online with this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Papantonis, A.; Swevers, L.; Iatrou, K. Chorion genes: A landscape of their evolution, structure, and regulation. Annu Rev Entomol 2015, 60, 177–194. [Google Scholar] [CrossRef]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory pathways controlling female insect reproduction. Annu Rev Entomol 2018, 63, 489–511. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory mechanisms of vitellogenesis in insects. Front Cell Dev Biol 2021, 8, 593613. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.Z.; Ahmad, S.; Ngegba, P.M.; Zhong, G. Role of endocrine system in the regulation of female insect reproduction. Biology 2021, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, R.; Sun, Z.; Bai, H.; Palli, S.R. Juvenile hormone regulation of vitellogenin synthesis in the red flour beetle, Tribolium castaneum. Insect Biochem Mol Biol 2010, 40, 405–414. [Google Scholar] [CrossRef]
- Eid, D.M.; Chereddy, S.C.R.R.; Palli, S.R. The effect of E93 knockdown on female reproduction in the red flour beetle, Tribolium castaneum. Arch Insect Biochem Physiol 2020, 104, e21688. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tan, A.; Palli, S.R. The function of nuclear receptors in regulation of female reproduction and embryogenesis in the red flour beetle, Tribolium castaneum. J Insect Physiol 2010, 56, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, Z.; Bi, J.; Feng, Q.; Beerntsen, B.T.; Song, Q. Neuropeptide bursicon influences reproductive physiology in Tribolium Castaneum. Front Physiol 2021, 12, 717437. [Google Scholar] [CrossRef] [PubMed]
- Swevers, L.; Iatrou, K. Early establishment and autonomous implementation of a developmental program controlling silk moth chorion gene expression. Dev Biol 1992, 150, 12–22. [Google Scholar] [CrossRef]
- Petri, W.H.; Wyman, A.R.; Kafatos, F.C. Specific protein synthesis in cellular differentiation. III. The eggshell proteins of Drosophila melanogaster and their program of synthesis. Dev Biol 1976, 49, 185–199. [Google Scholar] [CrossRef]
- Tootle, T.L.; Williams, D.; Hubb, A.; Frederick, R.; Spradling, A.C. Drosophila eggshell production: Identification of new genes and coordination by Pxt. PLoS ONE 2011, 6, e19943. [Google Scholar] [CrossRef]
- Kafatos, F.C.; Spoerel, N.; Mitsialis, S.A.; Nguyen, H.T.; Romano, C.; Lingappa, J.R.; Mariani, B.D.; Rodakis, G.C.; Lecanidou, R.; Tsitilou, S.G. Developmental control and evolution in the chorion gene families of insects. Adv Genet 1987, 24, 223–242. [Google Scholar]
- Moreira, M.F.; Dos Santos, A.S.; Marotta, H.R.; Mansur, J.F.; Ramos, I.B.; Machado, E.A.; Souza, G.H.M.F.; Eberlin, M.N.; Kaiser, C.R.; Kramer, K.J.; et al. A chitin-like component in Aedes aegypti eggshells, eggs and ovaries. Insect Biochem Mol Biol 2007, 37, 1249–1261. [Google Scholar] [CrossRef]
- Jiang, L.-H.; Mu, L.-L.; Jin, L.; Anjum, A.A.; Li, G.-Q. Silencing uridine diphosphate N-acetylglucosamine pyrophosphorylase gene impairs larval development in Henosepilachna vigintioctopunctata. Pest Manag Sci 2022, 78, 3894–3902. [Google Scholar] [CrossRef]
- Jiang, L.-H.; Mu, L.-L.; Jin, L.; Anjum, A.A.; Li, G.-Q. RNAi for chitin synthase 1 rather than 2 causes growth delay and molting defect in Henosepilachna vigintioctopunctata. Pestic Biochem Physiol 2021, 178, 104934. [Google Scholar] [CrossRef]
- Shi, J.-F.; Mu, L.-L.; Chen, X.; Guo, W.-C.; Li, G.-Q. RNA interference of chitin synthase genes inhibits chitin biosynthesis and affects larval performance in Leptinotarsa decemlineata (Say). International Journal of Biological Sciences 2016, 12, 1319–1331. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.-F.; Fu, J.; Mu, L.-L.; Guo, W.-C.; Li, G.-Q. Two Leptinotarsa uridine diphosphate N-acetylglucosamine pyrophosphorylase genes LdUAP1 and LdUAP2 are specialized for synthesis of chitin in larval epidermal cuticle and midgut peritrophic matrix. Insect Biochemistry and Molecular Biology 2016, 68, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.-F.; Mu, L.-L.; Guo, W.-C.; Li, G.-Q. Identification and hormone induction of putative chitin synthase genes and splice variants in Leptinotarsa decemlineata (Say). Archives of Insect Biochemistry and Physiology 2016, 92, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Specht, C.A.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Chitin synthases are required for survival, fecundity and egg hatch in the red flour beetle, Tribolium castaneum. Insect Biochem Mol Biol 2008, 38, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, C.G.; Rezende, G.L.; Lamers, G.E.; van der Zee, M. The extraembryonic serosa protects the insect egg against desiccation. P Roy Soc B Biol Sci 2013, 280, 20131082. [Google Scholar] [CrossRef]
- Zhang, X.-Q.; Jin, L.; Li, G.-Q. The timings of vitellogenesis and choriogenesis in the Henosepilachna vigintioctopunctata oocytes. Journal of Asia-Pacific Entomology 2024, 27, 102183. [Google Scholar] [CrossRef]
- Zhang, X.-Q.; Jin, L.; Li, G.-Q. RNAi-mediated functional analysis reveals the regulation of oocyte vitellogenesis by ecdysone signaling in two Coleoptera species. Biology 2023, 12, 1284. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Specht, C.A.; Tomoyasu, Y.; Lorenzen, M.D.; Kanost, M.R.; Beeman, R.W. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix, respectively. Insect Mol Biol 2005, 14, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Benrabaa, S.; Orchard, I.; Lange, A.B. A critical role for ecdysone response genes in regulating egg production in adult female Rhodnius prolixus. PLoS ONE 2023, 18, e0283286. [Google Scholar] [CrossRef] [PubMed]
- Benrabaa, S.A.M.; Orchard, I.; Lange, A.B. The role of ecdysteroid in the regulation of ovarian growth and oocyte maturation in Rhodnius prolixus, a vector of Chagas disease. J Exp Biol 2022, 225, jeb244830. [Google Scholar] [CrossRef]
- Knapp, E.M.; Li, W.; Singh, V.; Sun, J. Nuclear receptor Ftz-f1 promotes follicle maturation and ovulation partly via bHLH/PAS transcription factor Sim. Elife 2020, 9, e54568. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Shi, Q.; Xu, C.; Wang, R.; Wang, H.; Song, Y.; Zeng, R. Regulation of NlE74A on vitellogenin may be mediated by angiotensin converting enzyme through a fecundity-related SNP in the brown planthopper, Nilaparvata lugens. Comp Biochem Physiol 2018, 225, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Li, Y.; Gao, H.; Lin, X. The direct interaction between E93 and Kr-h1 mediated their antagonistic effect on ovary development of the brown planthopper. Int J Mol Sci 2019, 20, 2431. [Google Scholar] [CrossRef] [PubMed]
- Gujar, H.; Palli, S.R. Juvenile hormone regulation of female reproduction in the common bed bug, Cimex lectularius. Sci Rep 2016, 6, 35546. [Google Scholar] [CrossRef]
- Mansur, J.F.; Alvarenga, E.S.; Figueira-Mansur, J.; Franco, T.A.; Ramos, I.B.; Masuda, H.; Melo, A.C.; Moreira, M.F. Effects of chitin synthase double-stranded RNA on molting and oogenesis in the Chagas disease vector Rhodnius prolixus. Insect Biochem Mol Biol 2014, 51, 110–121. [Google Scholar] [CrossRef]
- Lou, Y.H.; Pan, P.L.; Ye, Y.X.; Cheng, C.; Xu, H.J.; Zhang, C.X. Identification and functional analysis of a novel chorion protein essential for egg maturation in the brown planthopper. Insect Mol Biol 2018, 27, 393–403. [Google Scholar] [CrossRef]
- Lou, Y.H.; Shen, Y.; Li, D.T.; Huang, H.J.; Lu, J.B.; Zhang, C.X. A mucin-like protein is essential for oviposition in Nilaparvata lugens. Front Physiol 2019, 10, 551. [Google Scholar] [CrossRef]
- Ahmed, S.; Seo, K.; Kim, Y. An ovary-specific mucin is associated with choriogenesis mediated by prostaglandin signaling in Spodoptera exigua. Arch Insect Biochem Physiol 2021, 106, e21748. [Google Scholar] [CrossRef]
- Margaritis, L.H. The egg-shell of Drosophila melanogaster III. Covalent crosslinking of the chorion proteins involves endogenous hydrogen peroxide. Tissue Cell 1985, 17, 553–559. [Google Scholar] [CrossRef]
- Mindrinos, M.N.; Petri, W.H.; Galanopoulos, V.K.; Lombard, M.F.; Margaritis, L.H. Crosslinking of the Drosophila chorion involves a peroxidase. Wilehm Roux Arch Dev Biol 1980, 189, 187–196. [Google Scholar] [CrossRef]
- Lu, J.B.; Zhang, M.Q.; Li, L.C.; Zhang, C.X. DDC plays vital roles in the wing spot formation, egg production, and chorion tanning in the brown planthopper. Arch Insect Biochem Physiol 2019, 101, e21552. [Google Scholar] [CrossRef]
- Wang, P.; Granados, R.R. An intestinal mucin is the target substrate for a baculovirus enhancin. Proc Natl Acad Sci U S A 1997, 94, 6977–6982. [Google Scholar] [CrossRef] [PubMed]
- Ze, L.-J.; Wang, P.; Peng, Y.-C.; Jin, L.; Li, G.-Q. Silencing tyrosine hydroxylase or dopadecarboxylase gene disrupts cuticle tanning during larva-pupa-adult transformation in Henosepilachna vigintioctopunctata. Pest Manag Sci 2022, 78, 3880–3893. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Gul, H.; Wang, X.; Ding, Q.; Said, F.; Gao, X.; Desneux, N.; Song, D. RNAi-mediated knockdown of chitin synthase 1 (CHS1) gene causes mortality and decreased longevity and fecundity in Aphis gossypii. Insects 2019, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Harethardottir, H.M.; Male, R.; Nilsen, F.; Dalvin, S. Chitin synthases are critical for reproduction, molting, and digestion in the salmon louse (Lepeophtheirus salmonis). Life 2021, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Rios, T.; Bomfim, L.; Ramos, I. The transition from vitellogenesis to choriogenesis triggers the downregulation of the UPR sensors IRE1 and PERK and alterations in the ER architecture in the follicle cells of the vector Rhodnius prolixus. Cell Tissue Res 2022, 387, 63–74. [Google Scholar] [CrossRef]
- Bernardi, F.; Romani, P.; Tzertzinis, G.; Gargiulo, G.; Cavaliere, V. EcR-B1 and Usp nuclear hormone receptors regulate expression of the VM32E eggshell gene during Drosophila oogenesis. Dev Biol 2009, 328, 541–551. [Google Scholar] [CrossRef]
- Dutko-Gwóźdź, J.; Gwóźdź, T.; Orłowski, M.; Greb-Markiewicz, B.; Duś, D.; Dobrucki, J.; Ozyhar, A. The variety of complexes formed by EcR and Usp nuclear receptors in the nuclei of living cells. Mol Cell Endocrinol 2008, 294, 45–51. [Google Scholar] [CrossRef]
- Costantino, B.F.; Bricker, D.K.; Alexandre, K.; Shen, K.; Merriam, J.R.; Antoniewski, C.; Callender, J.L.; Henrich, V.C.; Presente, A.; Andres, A.J. A novel ecdysone receptor mediates steroid-regulated developmental events during the mid-third instar of Drosophila. PLoS Genet 2008, 4, e1000102. [Google Scholar] [CrossRef]
- Sharma, V.; Pandey, A.K.; Kumar, A.; Misra, S.; Gupta, H.P.K.; Gupta, S.; Singh, A.; Buehner, N.A.; Ravi Ram, K. Functional male accessory glands and fertility in Drosophila require novel ecdysone receptor. PLoS Genet 2017, 3, e1006788. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Sakai, A.; Magata, F.; Ogura, T.; Miyashita, M.; Miyagawa, H. Molecular cloning of the ecdysone receptor and the retinoid X receptor from the scorpion, Liocheles australasiae. FEBS J 2007, 274, 6191–6203. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Zhang, Z.L.; Robinson, G.E.; Palli, S.R. Identification and characterization of a juvenile hormone response element and its binding proteins. J Biol Chem 2007, 282, 37605–37617. [Google Scholar] [CrossRef] [PubMed]
- Bodofsky, S.; Koitz, F.; Wightman, B. Conserved and exapted functions of nuclear receptors in animal development. Nucl Receptor Res 2017, 4, 101305. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Miura, K.; Chen, L.; Raikhel, A.S. Cyclicity of mosquito vitellogenic ecdysteroid-mediated signaling is modulated by alternative dimerization of the RXR homologue Ultraspiracle. Proc Natl Acad Sci U S A 2003, 100, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.-Y.; Deng, P.; Zhang, Q.; Li, A.; Fu, K.-Y.; Guo, W.-C.; Li, G.-Q. Ecdysone receptor isoforms play distinct roles in larval-pupal-adult transition in Leptinotarsa decemlineata. Insect Sci 2020, 27, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.-Y.; Deng, P.; Li, A.; Zhang, Q.; Mu, L.-L.; Fu, K.-Y.; Guo, W.-C.; Li, G.-Q. Functional characterization of ultraspiracle in Leptinotarsa decemlineata using RNA interference assay. Insect Mol Biol 2019, 28, 676–688. [Google Scholar] [CrossRef]
- Xu, P.; Ze, L.-J.; Kang, W.-N.; Wu, J.-J.; Jin, L.; Ali, A.A.; Li, G.-Q. Functional divergence of white genes in Henosepilachna vigintioctopunctata revealed by RNA interference. Insect Molecular Biology 2020, 29, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Ze, L.-J.; Xu, P.; Kang, W.-N.; Wu, J.-J.; Jin, L.; Anjum, A.A.; Li, G.-Q. Disruption of ommochrome biosynthesis affects eye coloration, phototaxis and climbing in Henosepilachna vigintioctopunctata. Amino Acids 2021, 53, 1091–1104. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.-Q.; Guo, W.-C.; Wan, P.-J.; Zhou, L.-T.; Ren, X.-L.; Tursun, A.; Fu, K.-Y.; Li, G.-Q. Validation of reference genes for expression analysis by quantitative real-time PCR in Leptinotarsa decemlineata (Say). BMC Research Notes 2013, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Chen, S.; Guo, M.; Ye, C.; Qiu, B.; Wu, J.; Yang, C.; Pan, H. Selection and validation of reference genes for RT-qPCR analysis of the ladybird beetle Henosepilachna vigintioctomaculata. Front Physiol 2018, 9, 1614. [Google Scholar] [CrossRef]
- Reissig, J.L.; Strominger, J.L.; Leloir, L.F. A modified colorimetric method for the estimation of N-acetylamino sugars. Journal of Biological Chemistry 1955, 217, 959–966. [Google Scholar] [CrossRef]
Figure 1.
The temporal expression patterns of selected genes in the ovaries of two Coleoptera potato defoliators. The cDNA templates were derived from the ovaries of 0- to 9-day-old L. decemlineata, and 0- to 13-day-old H. vigintioctopunctata female adults respectively. The transcript levels of EcRA, EcRB1, usp, UAP, ChS1, and ChS2 were determined. For each sample, 3 independent pools of 5-10 individuals were measured in technical triplicate using real-time quantitative PCR (red dots). The bars represent 2-ΔΔCT value (±SD) normalized to the geometrical mean of house-keeping gene expression. The lowest transcript levels at a specific developmental time point were set as 1.
Figure 1.
The temporal expression patterns of selected genes in the ovaries of two Coleoptera potato defoliators. The cDNA templates were derived from the ovaries of 0- to 9-day-old L. decemlineata, and 0- to 13-day-old H. vigintioctopunctata female adults respectively. The transcript levels of EcRA, EcRB1, usp, UAP, ChS1, and ChS2 were determined. For each sample, 3 independent pools of 5-10 individuals were measured in technical triplicate using real-time quantitative PCR (red dots). The bars represent 2-ΔΔCT value (±SD) normalized to the geometrical mean of house-keeping gene expression. The lowest transcript levels at a specific developmental time point were set as 1.
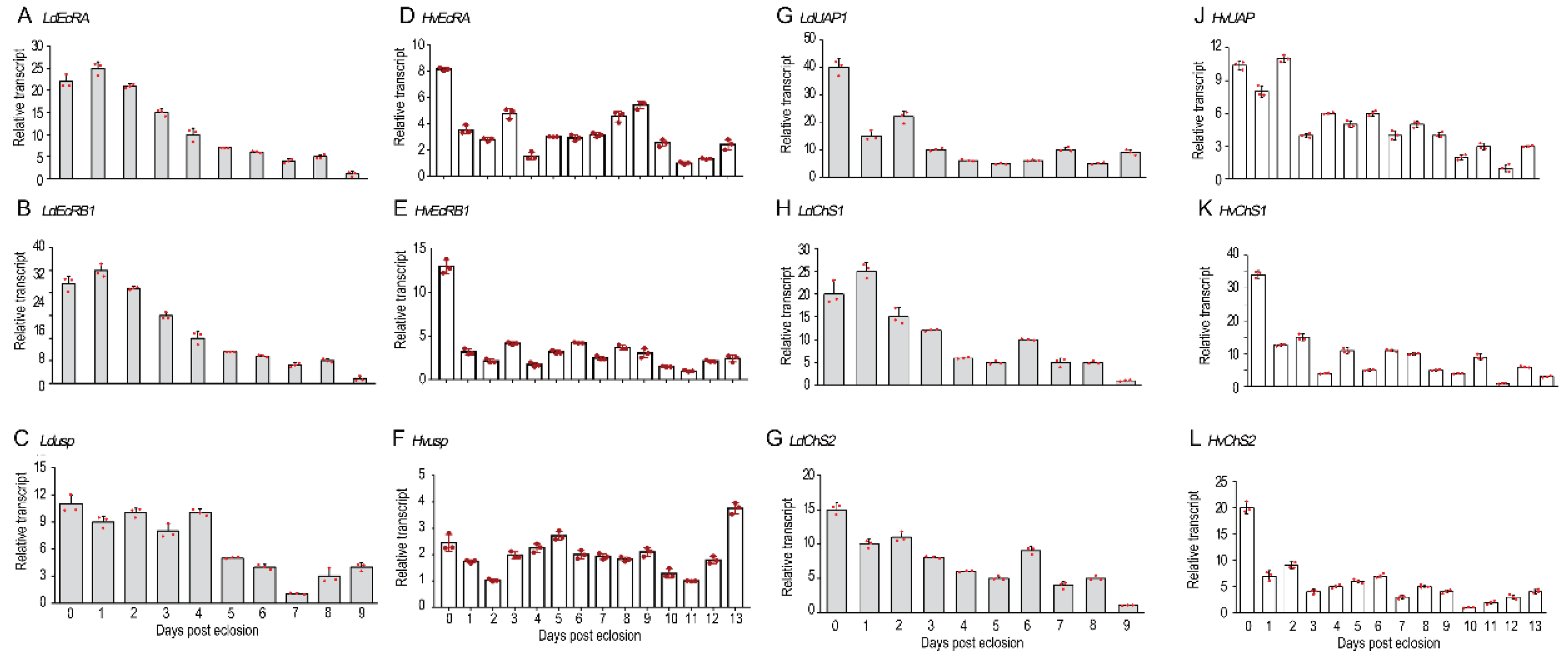
Figure 2.
Knockdown of HvEcR in the 0-day-old Henosepilachna vigintioctopunctata female adults. The 0-day-old female adults were injected with 0.1 μL dsegfp or dsHvEcR (400 ng). The treated beetles were fed on fresh potato foliage. Three days after treatment, transcript level of HvEcR was determined (n=3, A). Relative transcript is the ratio of relative copy number in treated individuals to dsegfp-treated controls, which is set as 1. The body sizes and fresh weights were recorded in the 30-day-old females (n=9-12, G-J). The eggs in the 30-day-old females were measured (n=15, M-O). The averages and SD ranges are drawn. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. The oviposition was observed 0 to 30 days post eclosion (B, C). The body sizes (D-F) and egg shapes (K, L) of 30-day-old females were shown.
Figure 2.
Knockdown of HvEcR in the 0-day-old Henosepilachna vigintioctopunctata female adults. The 0-day-old female adults were injected with 0.1 μL dsegfp or dsHvEcR (400 ng). The treated beetles were fed on fresh potato foliage. Three days after treatment, transcript level of HvEcR was determined (n=3, A). Relative transcript is the ratio of relative copy number in treated individuals to dsegfp-treated controls, which is set as 1. The body sizes and fresh weights were recorded in the 30-day-old females (n=9-12, G-J). The eggs in the 30-day-old females were measured (n=15, M-O). The averages and SD ranges are drawn. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. The oviposition was observed 0 to 30 days post eclosion (B, C). The body sizes (D-F) and egg shapes (K, L) of 30-day-old females were shown.
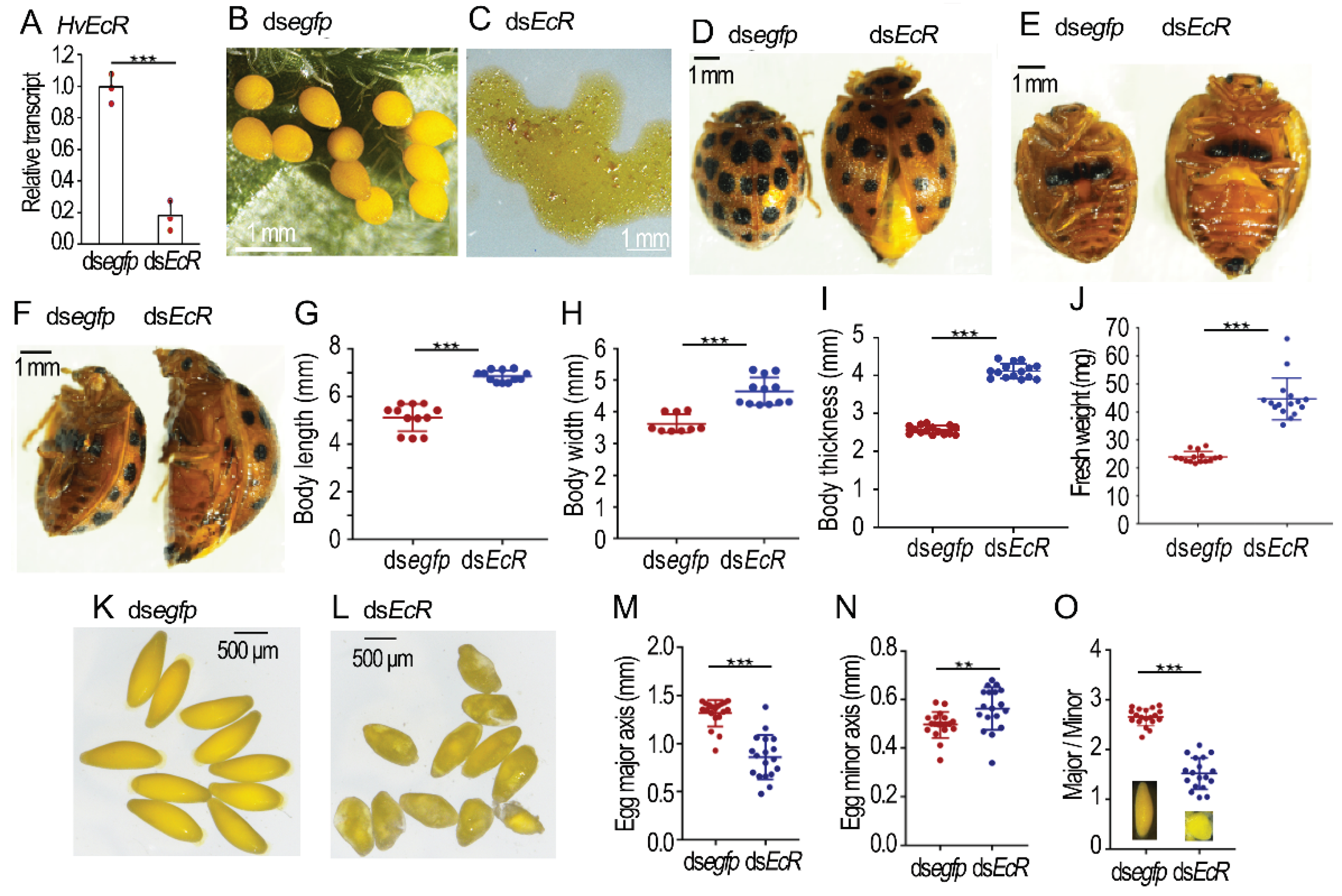
Figure 3.
Silence of Hvusp in the 0-day-old Henosepilachna vigintioctopunctata female adults. The 0-day-old female adults were injected with 0.1 μL dsegfp and dsHvusp (400 ng). The treated beetles were fed on fresh potato foliage. Three days after treatment, transcript level of Hvusp was determined (A). Relative transcript is the ratio of relative copy number in treated individuals to dsegfp-treated controls, which is set as 1. The fresh weights were recorded in the 30-day-old females (n=18, G). The eggs in the 20-day-old females were measured (n=15, K-M). The averages and SD ranges are drawn. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. NS, no significance. The oviposition was observed 0 through 40 days post eclosion (B-D). The body sizes of 30-day-old females (E, F) and egg shapes of 20-day-old females (H-J) were shown.
Figure 3.
Silence of Hvusp in the 0-day-old Henosepilachna vigintioctopunctata female adults. The 0-day-old female adults were injected with 0.1 μL dsegfp and dsHvusp (400 ng). The treated beetles were fed on fresh potato foliage. Three days after treatment, transcript level of Hvusp was determined (A). Relative transcript is the ratio of relative copy number in treated individuals to dsegfp-treated controls, which is set as 1. The fresh weights were recorded in the 30-day-old females (n=18, G). The eggs in the 20-day-old females were measured (n=15, K-M). The averages and SD ranges are drawn. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. NS, no significance. The oviposition was observed 0 through 40 days post eclosion (B-D). The body sizes of 30-day-old females (E, F) and egg shapes of 20-day-old females (H-J) were shown.
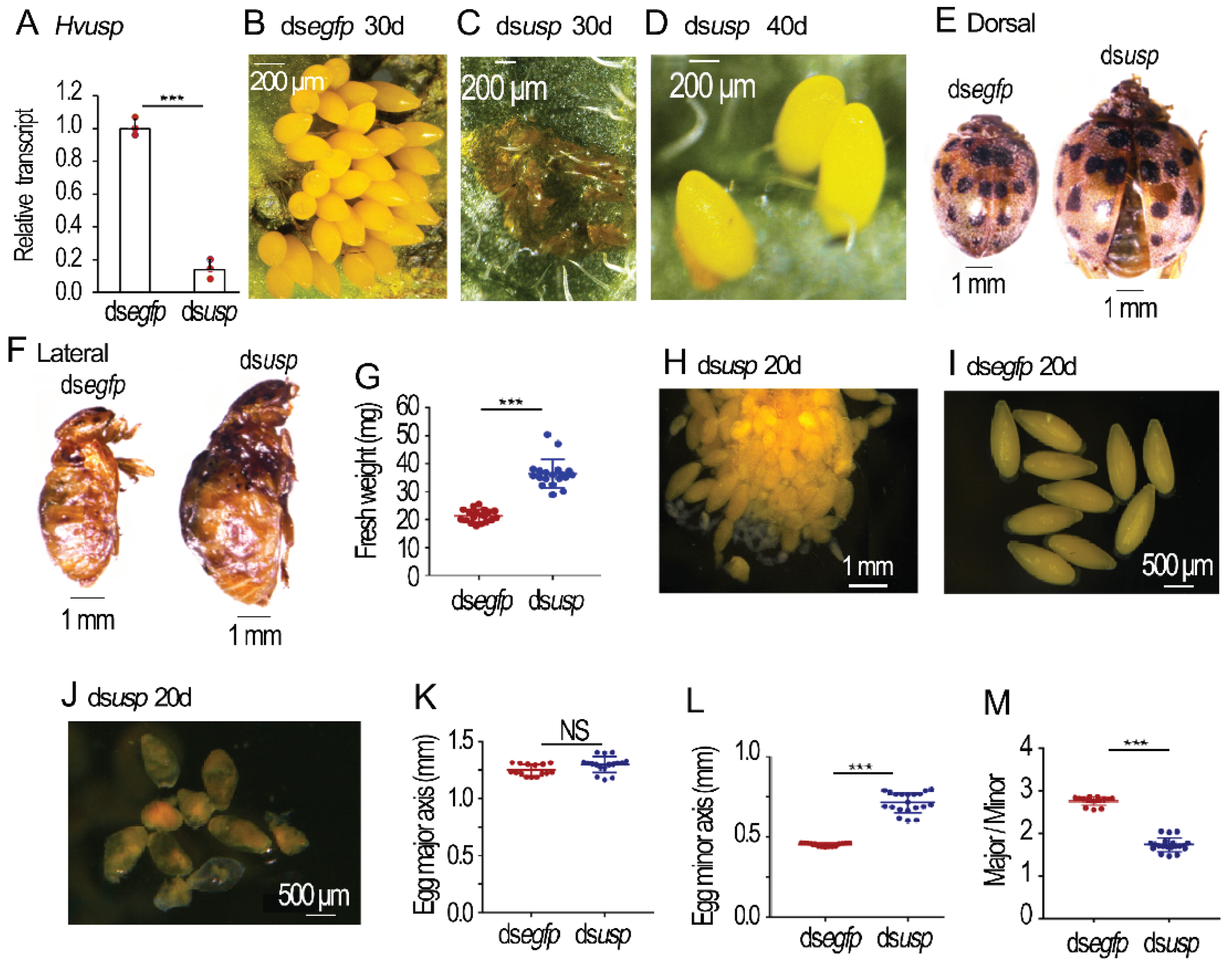
Figure 4.
RNAi of HvEcR represses choriogenesis in Henosepilachna vigintioctopunctata. Ten and twenty days post injection of dsegfp and dsHvEcR, the ovaries of resultant females were dissected. The expression levels of HvChS1 and HvChS2 from 10- and 20-day-old ovaries were determined using the 2-ΔΔCT method (A-D). Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp controls, which are set as 1. The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), 0.001 (***) or 0.0001 (****) using t test. NS, no significance. The paraffin sections of ovaries from 20-day-old treated and control female adults were treated with hematoxylin-eosin staining (E and F), or dyed with Calcofluor-White and 10% KOH (G-J). The images were showed. Blue color in G to J (shown with red arrow) marks chitin layer.
Figure 4.
RNAi of HvEcR represses choriogenesis in Henosepilachna vigintioctopunctata. Ten and twenty days post injection of dsegfp and dsHvEcR, the ovaries of resultant females were dissected. The expression levels of HvChS1 and HvChS2 from 10- and 20-day-old ovaries were determined using the 2-ΔΔCT method (A-D). Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp controls, which are set as 1. The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), 0.001 (***) or 0.0001 (****) using t test. NS, no significance. The paraffin sections of ovaries from 20-day-old treated and control female adults were treated with hematoxylin-eosin staining (E and F), or dyed with Calcofluor-White and 10% KOH (G-J). The images were showed. Blue color in G to J (shown with red arrow) marks chitin layer.
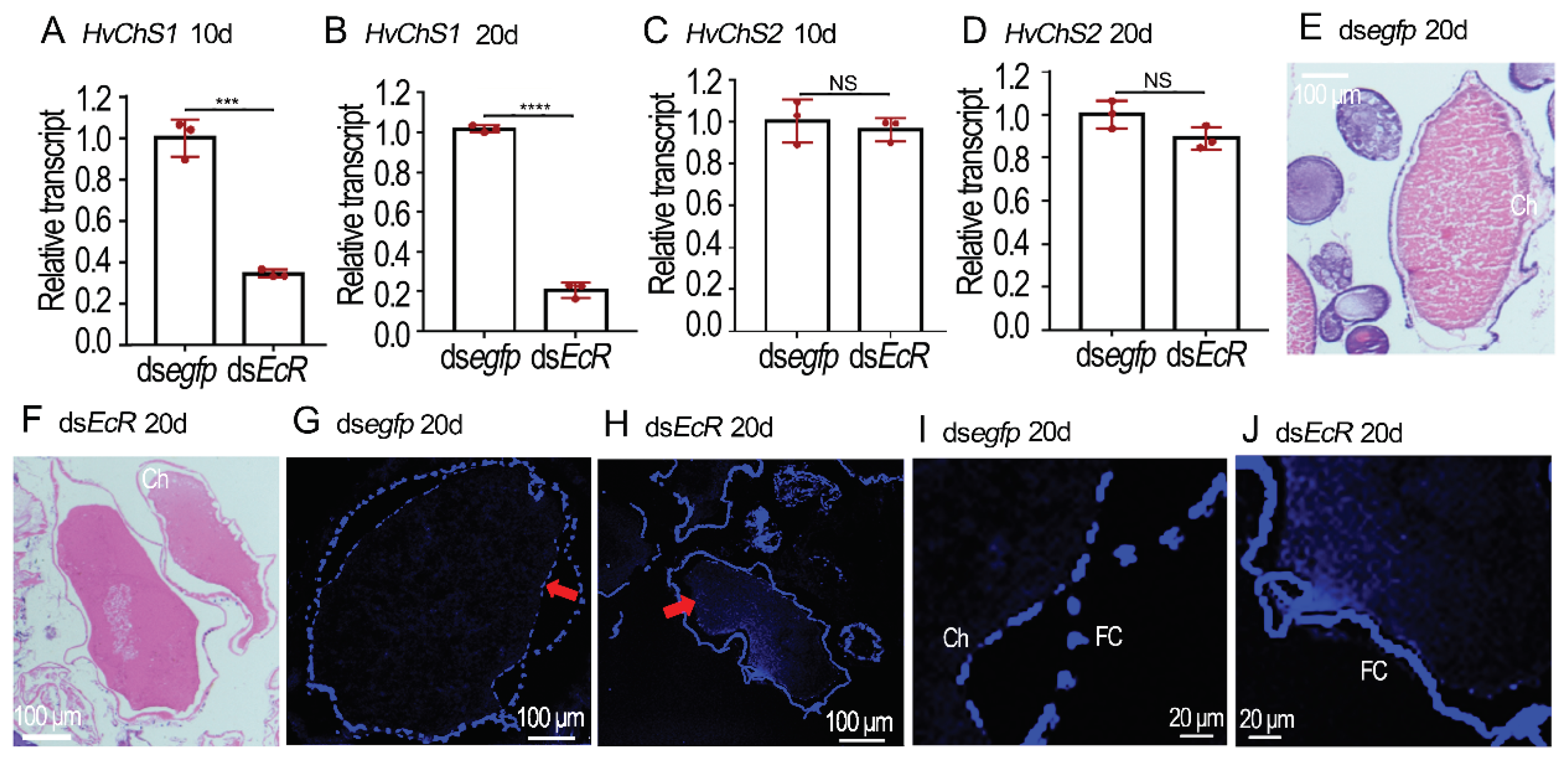
Figure 5.
Depletion of Hvusp blocks choriogenesis in Henosepilachna vigintioctopunctata. Ten and twenty days post injection of dsegfp and dsHvusp, the ovaries of resultant females were dissected. The expression levels of HvChS1 and HvChS2 from 10- and 20-day-old ovaries were determined using the 2-ΔΔCT method (A-D). Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp controls, which is set as 1. The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**) or 0.001 (***) using t test. NS, no significance. The paraffin sections of ovaries from 20-day-old treated and control female adults were treated with hematoxylin-eosin staining (E and F), or dyed with Calcofluor-White and 10% KOH (G, H). The images were showed. Blue color in G and H (shown with red arrow) marks chitin layer.
Figure 5.
Depletion of Hvusp blocks choriogenesis in Henosepilachna vigintioctopunctata. Ten and twenty days post injection of dsegfp and dsHvusp, the ovaries of resultant females were dissected. The expression levels of HvChS1 and HvChS2 from 10- and 20-day-old ovaries were determined using the 2-ΔΔCT method (A-D). Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp controls, which is set as 1. The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**) or 0.001 (***) using t test. NS, no significance. The paraffin sections of ovaries from 20-day-old treated and control female adults were treated with hematoxylin-eosin staining (E and F), or dyed with Calcofluor-White and 10% KOH (G, H). The images were showed. Blue color in G and H (shown with red arrow) marks chitin layer.
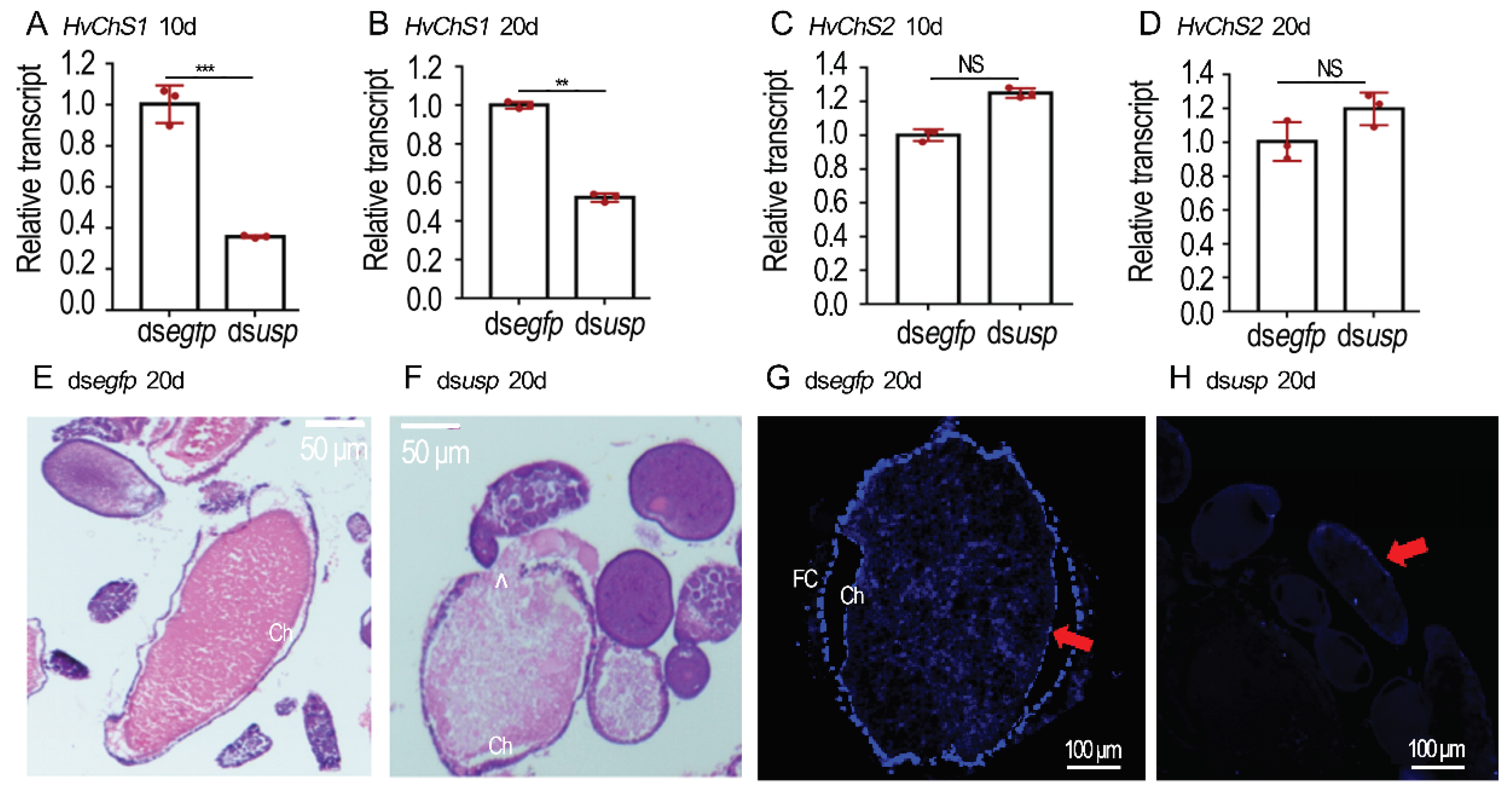
Figure 6.
RNAi of LdEcR or Ldusp in the 0-day-old Leptinotarsa decemlineata female adults. Two biologically independent experiments were carried out using different generations, with two treatments: (1) dsegfp and (2) dsLdEcR or dsLdusp. The treated beetles were fed on fresh potato foliage. Three or ten days days after treatment, transcript levels of LdEcR, LdUAP1, LdUAP2, LdChS1 and LdChS2 (A, D-G), or Ldusp, LdUAP1, LdUAP2, LdChS1 and LdChS2 (I-M) in the ovaries were determined. Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp-treated controls, which are set as 1. The chitin contents of the 10-day-old ovaries were measured by N-acetylglucosamine (GlcNAc) concentrations using the chitinase degraded method (H, N). The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. NS, no significance. The oviposition was observed 0 to 15 days post eclosion (B, C).
Figure 6.
RNAi of LdEcR or Ldusp in the 0-day-old Leptinotarsa decemlineata female adults. Two biologically independent experiments were carried out using different generations, with two treatments: (1) dsegfp and (2) dsLdEcR or dsLdusp. The treated beetles were fed on fresh potato foliage. Three or ten days days after treatment, transcript levels of LdEcR, LdUAP1, LdUAP2, LdChS1 and LdChS2 (A, D-G), or Ldusp, LdUAP1, LdUAP2, LdChS1 and LdChS2 (I-M) in the ovaries were determined. Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp-treated controls, which are set as 1. The chitin contents of the 10-day-old ovaries were measured by N-acetylglucosamine (GlcNAc) concentrations using the chitinase degraded method (H, N). The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. NS, no significance. The oviposition was observed 0 to 15 days post eclosion (B, C).
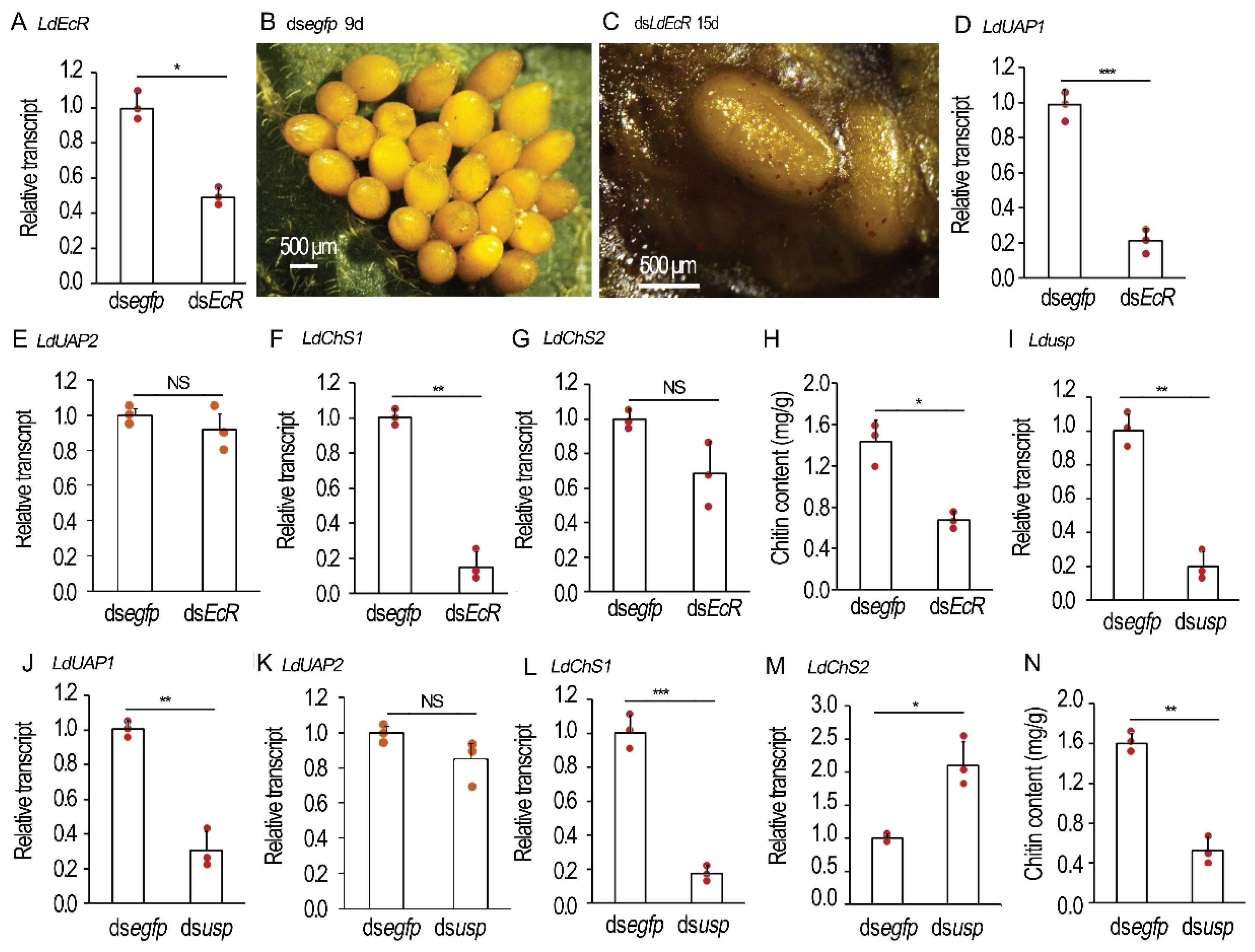
Figure 7.
Knockdown of chitin biosynthesis genes in the 4-day-old Leptinotarsa decemlineata female adults. The 4-day-old female adults were injected with 0.1 μL dsegfp, dsLdUAP1, dsLdUAP2, dsLdChS1 or dsLdChS2 (400 ng). The treated beetles were fed on fresh potato foliage. Three days after treatment, transcript levels of LdUAP1, LdUAP2, LdChS1 and LdChS2 were determined (A-D). Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp-treated controls, which are set as 1. The chitin contents of the 10-day-old ovaries were measured by N-acetylglucosamine (GlcNAc) concentrations using the chitinase degraded method (E). The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. NS, no significance. The 10-day-old ovaries were shown (F-J). The oviposition was observed 0 to 20 days post eclosion (K). A presumptive model of the molecular regulation of 20E signal for choriogenesis in the two Coleopteran species (L). EcR/USP complex activates the expression of chitin biosynthesis genes, such as UAP and ChS1, or may indirectly acts on these genes, to regulate the supply of chitin-like substance (CLS). Meanwhile, EcR and USP may respectively regulate a specific subset of chorion protein genes, dependent or independent of EcR/USP complex, to synthesize constructive proteins for the formation of eggshells (L).
Figure 7.
Knockdown of chitin biosynthesis genes in the 4-day-old Leptinotarsa decemlineata female adults. The 4-day-old female adults were injected with 0.1 μL dsegfp, dsLdUAP1, dsLdUAP2, dsLdChS1 or dsLdChS2 (400 ng). The treated beetles were fed on fresh potato foliage. Three days after treatment, transcript levels of LdUAP1, LdUAP2, LdChS1 and LdChS2 were determined (A-D). Relative transcripts are the ratios of relative copy numbers in treated individuals to dsegfp-treated controls, which are set as 1. The chitin contents of the 10-day-old ovaries were measured by N-acetylglucosamine (GlcNAc) concentrations using the chitinase degraded method (E). The columns represent averages with vertical lines indicating SD. Different stars indicate significant difference at p value < 0.05 (*), 0.01 (**), or 0.001 (***) using t test. NS, no significance. The 10-day-old ovaries were shown (F-J). The oviposition was observed 0 to 20 days post eclosion (K). A presumptive model of the molecular regulation of 20E signal for choriogenesis in the two Coleopteran species (L). EcR/USP complex activates the expression of chitin biosynthesis genes, such as UAP and ChS1, or may indirectly acts on these genes, to regulate the supply of chitin-like substance (CLS). Meanwhile, EcR and USP may respectively regulate a specific subset of chorion protein genes, dependent or independent of EcR/USP complex, to synthesize constructive proteins for the formation of eggshells (L).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



