Submitted:
19 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Globally, cancer is an important health problem worldwide, and has become one modern medicine’s most serious problems to prevent, diagnose, and treat. Many types of cancers still have non-negligible mortality rates, and new treatments need to be developed for them. This study aimed to elucidate cancer mechanisms and facilitate cancer treatment and prevention. The ideal temperature for cancer growth is 35°C; however, conditions were not consistent with chromosomal instability, loss of alleles of tumor-suppressor genes, abnormal DNA methylation, and collagen phagocytosis, which are known causes of cancer. In addition, when cancer develops, white blood cells attack the cancer tissue and remove it; however, as the decrease in body temperature drops, leads to a reduction in the immune potential of the white blood cells. Based on previous papers, this study focused on aimed to determine the effects of the low-temperature environment of 35°C, the temperature at which cancer is most likely to proliferate, and changes in adenosine triphosphate (ATP) levels. The study revealed that Therefore, the causes of cancer is caused by include chromosome fusion that occurs occurring due to the impaired DNA topoisomerase IIα proliferation and primary cilia rotation due to resulting from decreased intercellular adhesion.
Keywords:
carcinogenesis
; collagen
; metabolism
; primary cilia
; temperature sensitivity
; mitochondria
Introduction
Cancer has multiple etiologies, and although certain causes have been discovered, no clear cause has been identified in all cases. Therefore, this study summarized the typical etiologies of cancer.
Cells have cancer-suppressor genes, —p53 and Rb, which maintain their function if any one of the two alleles in the cell is normal. However, the first step in carcinogenesis is often associated with the loss of tumor-suppressor genes because this may occur through the loss of two alleles, which prevents the tumor-suppressor genes from functioning. The loss of heterozygosity and chromosome number abnormalities are caused by chromosome instability, which occurs when a part of a chromosome is torn, and disengages and fuses with another chromosome during cell division and 1%–2% of genes pass only one of the two alleles to the daughter cells, which are known to be typically involved in carcinogenesis [1,2].
DNA methylation is an important process that is involved in the regulation of gene expression and gene inactivation; however, genome-wide hypomethylation and localized DNA methylation abnormalities occur in cancer cells, and genome-wide hypomethylation induces chromosomal instability and promotes tumorigenesis such as in cases of lymphomas. In addition, in local hypermethylation, hypermethylation of the CpG island promoter region of a tumor-suppressor gene induces the silencing of the downstream genes. Therefore, hypomethylation of the entire genome and hypermethylation of the CpG island promoter region of the tumor-suppressor genes for some reasons were reported to adversely affect the chromosomes and are were involved in carcinogenesis [3,4].
Collagen is a major component of the extracellular matrix and is also involved in cell division, growth, and motility. However, cancer cell growth is impeded by collagen, which forms the tissue skeleton. Urokinase plasminogen activator receptor-associated proteins on the cell surface of fibroblasts are necessary for cellular collagen adhesion, collagen phagocytosis, and subsequent degradation by lysosomes, and cancer cells are responsible for the collagen phagocytosis via urokinase plasminogen activator receptor-associated proteins. Cancer cells have low levels of interaction with the extracellular matrix and proliferate in tissues with denatured collagen [5,6].
On the other hand, the following factors were involved in cell division.
Mitochondria play a central role in ATP production; however, the abundance of mitochondria decreases with age [7,8].Mitochondrial DNA is damaged by reactive oxygen species, and the mitochondrial respiration rate of cells in people aged >75 years is approximately half that of those aged <40 years [9]. Moreover, the mitochondrial inner membrane contains a type of heme protein, called cytochrome c. Increase in the intracellular calcium ion concentration leads to the release of cytochrome c from the mitochondria by stimulating mitochondrial permeability transition pore opening. The small amount of released cytochrome c interacts with the IP3 receptor on the endoplasmic reticulum membrane and releases calcium from the endoplasmic reticulum. Calcium, whose concentration is increased by this process, causes a massive release of cytochrome c, which forms a positive feedback loop and continues to be released from the endoplasmic reticulum until the amount of calcium becomes cytotoxic [10,11]. Cytochrome c released into the cytoplasm activates caspase-9 (cysteine protease), which in turn consequently activates caspases-3, caspase-6, and caspase-7, ultimately causing apoptosis and contributing to cell death. However, cancer cells are known to evade this pathway [12].
Oncogenes include tumor-suppressor genes such as p53 and p16, approximately 100 proto-oncogenes exist on the chromosomes of normal cells before becoming oncogenes, and these two are called cancer-related genes. Cancer cells have no mitotic limitations, stem cells are known to be activated, the cancer suppressor gene p16 maintains higher self-renewal capacity in the stem cells of p16-deficient mice than in wild-type mice of the same age, and cancer progresses when the cancer suppressor gene p53 is inactivated. Proto-oncogenes do not exist to make cells cancerous. However, the products of proto-oncogenes, along with collagen, encode proteins involved in cellular activities such as adhesion of neighboring cells in the extracellular matrix, cell cycle, apoptosis, and DNA repair, which mutate with age [13].
Mammalian cells contain primary cilia, i.e., cell organelles that control cell proliferation, but act as a site of signal transduction [14]. Primary cilia disappear in many cancer cells, which are believed to be caused by cancer cells that do not transition to the quiescent phase [15]. Conversely, primary cilia are required in medulloblastoma and basal cell carcinoma induced through the activation of mutants of Smoothened (SMO), an activator of hedgehog signaling [16]. Cancer cells have an abnormal number of centrioles, and the deletion of Plk4, STIL, and HsSAS-6—three proteins responsible for centriole replication—results in the absence of daughter centrioles, whereas the overexpression of the genes results in the formation of multiple daughter centrioles at once [17–19].
It was known that the cell membrane has two types of ion transport mechanisms: active transport, which uses ATP to transport ions against a concentration gradient to maintain a potential difference between inside and outside the cell, and passive transport, which does not use ATP and is driven by the concentration difference of substances [20].
Mouse DNA topoisomerase IIα is mutagen-treated and introduced into a yeast DNA topoisomerase IIα disruptant strain via the plasmid shuffle method, temperature sensitivity may be observed, with growth at 28°C but no growth at 35°C, and almost no growth at 37°C [21]. The temperature sensitivity of DNA topoisomerase IIα is caused by a decrease in enzyme activity, not a decrease in protein. DNA topoisomerase IIα uses its energy in the presence of ATP during DNA replication to simultaneously cleave two DNA strands and change the positive superhelix into a negative superhelix form by rotating the strongly wound DNA. In cell division too, chromosomes are separated by the simultaneous cleavage of two strands of DNA and the passage of another two strands of DNA between them as the chromosomes are separated into daughter cells [22].
Reports have indicated that carcinogenesis is caused by various factors, such as chromosomal instability and DNA replication errors, and collagen degeneration is involved in cancer growth. However, it is to be noted that they did not take into account metabolism and body temperature, even though it is known that cancer grows best at 35°C [23].
Cancer cells are known to overproduce the nuclear protein HSP90 [24]. By binding to ATP, HSP90 stabilizes cancer-associated proteins as a molecular chaperone and contributes to cancer cell proliferation.The heat that creates body temperature is generated by the process of ATP synthesis and breakdown, so at hypothermia of 35°C, the amount of ATP is low, so even taking into account the high amount of HSP90 in the nucleus during carcinogenesis, had to also consider changes in the amount of ATP because HSP90 is also present in normal cells [25,26].
Regarding cancer growth, we considered that carcinogenesis and cancer growth are related because when immune cells, such as white blood cells function normally at a body temperature of ≥36.5°C, cancer cells are attacked and cannot grow [27,28].
Therefore, if it was possible to verify how the decrease in proteins and enzymes and changes in their activity due to the decrease in body metabolism and body temperature in a low-temperature environment (35°C) could cause cell mutation and how it affects cancer, results could be useful in the development of cancer treatment.
Methods and Materials
Investigation of activity capacity and urine values using mice
By comparing the number of hamster wheel rotations, body weight, and weekly average of urinary inorganic phosphate and pH in cancer mice(BALB/c) and usually mice(BALB/c) until death, we investigated changes in the activity of and aging mitochondria to synthesize adenosine diphosphate(ADP) into ATP investigate.
Comparison of hamster wheel number of revolutions can be used as an indicator of mitochondrial activity and basal metabolism, since higher physical activity tends to lead to higher metabolism [29], while comparison of body weight can be used as an indicator of aging, since protein intake and muscle mass tend to decrease in older people [30].
Also, the kidneys are also responsible for maintaining a constant pH level by removing inorganic phosphate and pH from the blood into the urine [31,32]. It is known that kidney function declines with age, and the increase or decrease in inorganic phosphate in urine can be used as an indicator of the activity of the body's mitochondria or the synthesis of ATP from inorganic phosphate and ADP through anaerobic respiration, while an increase or decrease in urine pH can be used as an indicator of metabolism, as acid is released into the blood when nutrients are metabolized.
Ethical review of animal experiments: Even for research using mice, we conducted ethical review of animal experiments as stipulated in the Declaration of Helsinki, and prepared research plans that took into consideration the necessity of the experiments and the welfare of the animals.
(Identification code A-1, approval date December 2012 _25th, Animal Experimentation Committee)
Results
The results of an investigation of activity capacity and urine values using mice.
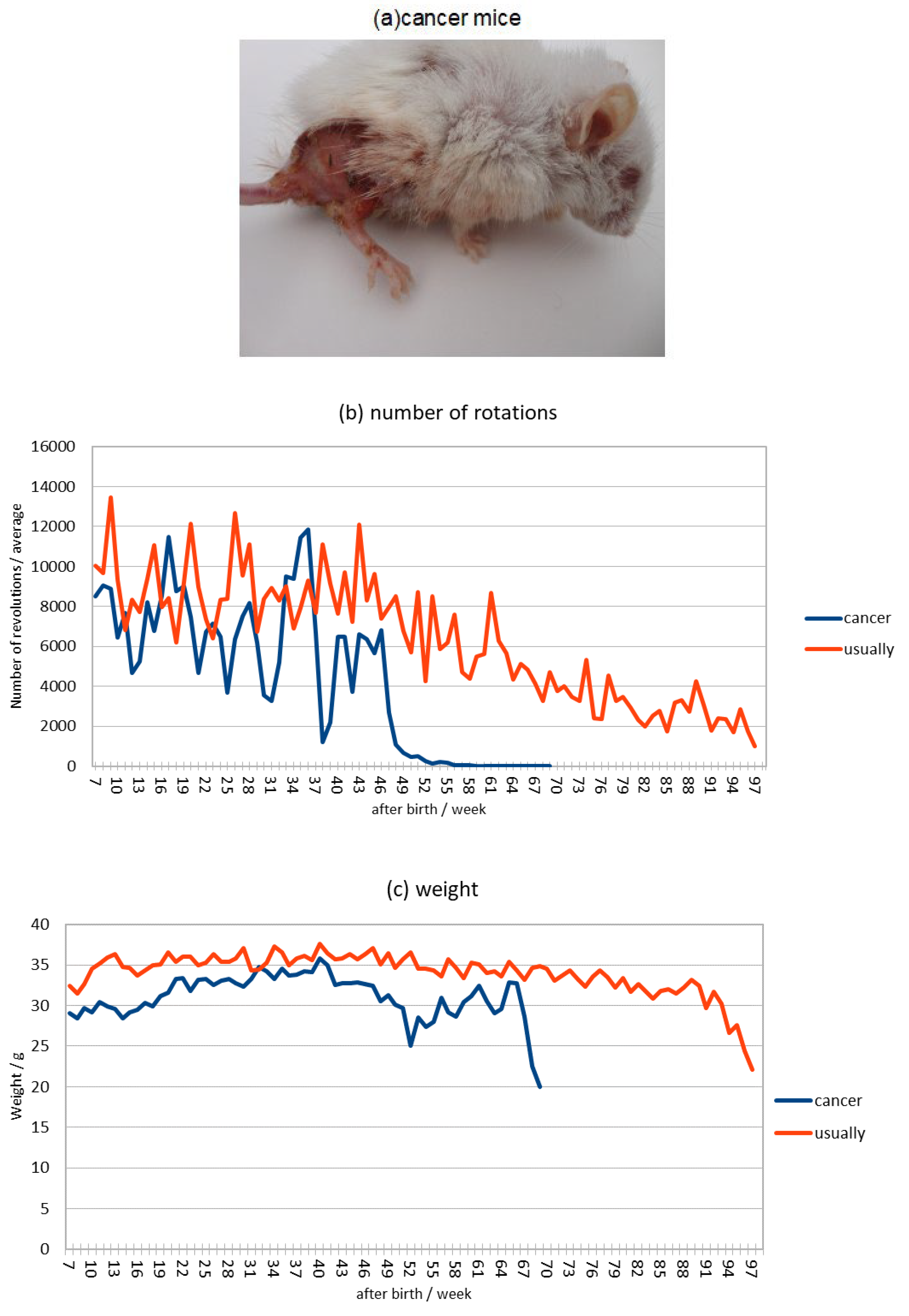
Cancer mice (Figure 1a) used individuals who developed skin cancer at 45 weeks after birth and died at 69 laps after birth, while usually mice died at 97 weeks after birth and were used for comparison.
The number of revolutions of the hamster wheel in cancer mice decreased greatly at 39 weeks, and the number of revolutions was extremely low after 49 weeks, while the number of revolutions in usually mice decreased slowly until death at 69 weeks (Figure 1b).
Comparing the weights, the cancerous mice show a severe increase or decrease in weight after skin cancer and an extreme decrease in weight just before death, while the usually mice show a gradual decrease in weight shortly before death (Figure 1c).
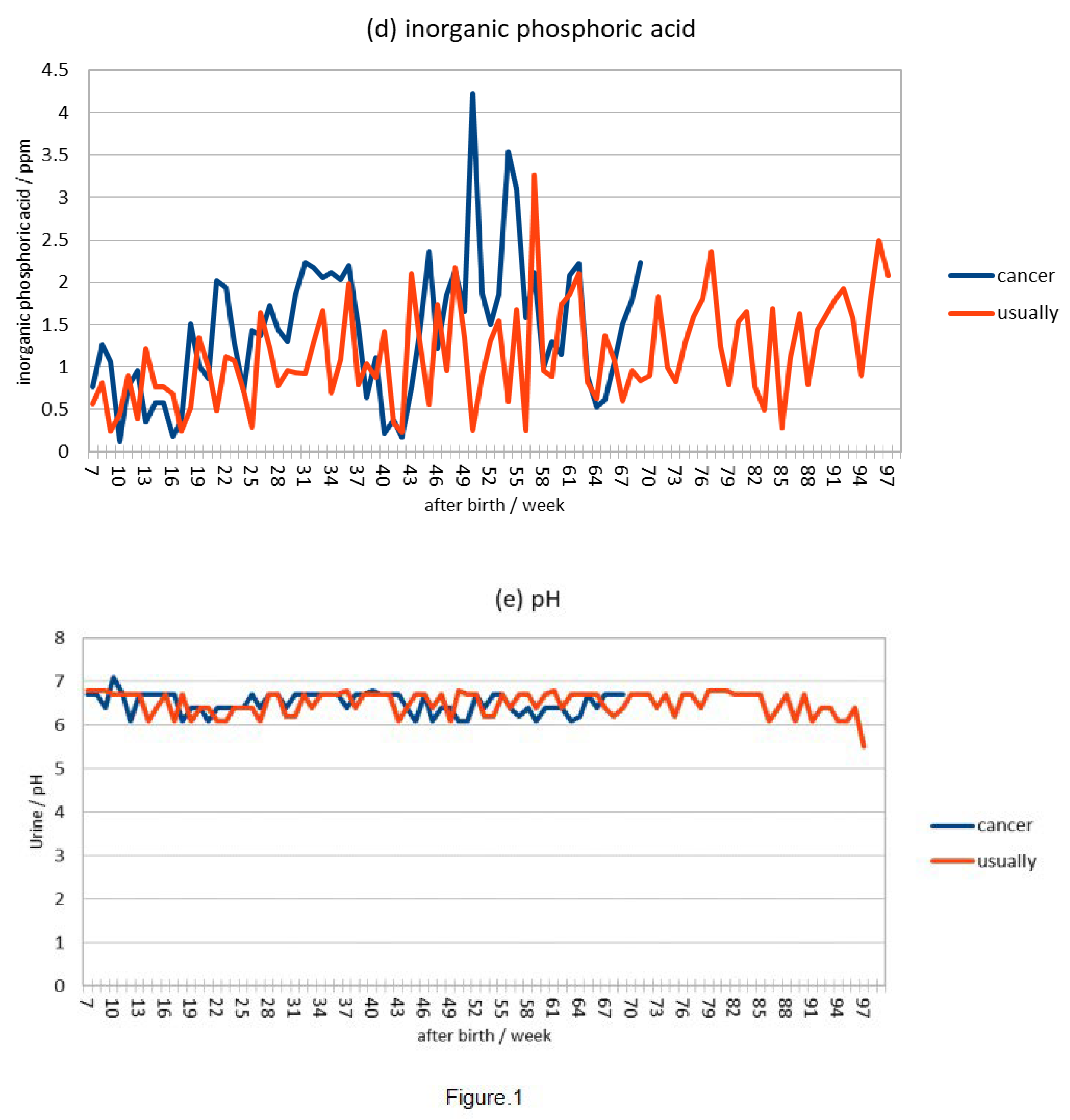
The ppm of inorganic phosphoric acid in 1 μl of urine showed that the highest value in cancer mice was measured at 50 weeks after skin cancer and was slightly higher immediately before death, but the highest value and the value immediately before death in usually mice were not much different from those in cancer mice (Figure 1d).
Comparing the pH of urine, mice with cancer maintain a pH of 6.0-7.0, which is within the standard value, until they die, but in normal mice, the pH becomes acidic at 5.0-6.0 69 weeks before death . (Figure 1e).
Discussion
When decrease in the number of mitochondria in the body occurs, it leads to a decrease in the metabolism and body temperature decrease, and consequently, changes occur in enzymes, proteins, and cell membranes. In addition, when cancer develops, there are cell organelles that undergo mutations. Therefore, the present study considered the importance of the properties of DNA topoisomerase IIα and the structure of primary cilia to elucidate the mechanism of cancer as intermediate steps in DNA replication.
The results from aging-related changes in mitochondrial function, cancer-related genes, cell organelles, and anaerobic respiration present the mechanism of cancer considering that they are also involved in the cells.
Mammalian cell membranes contain a Na+/K+-ATPase (sodium–potassium pump) that actively transports sodium ions and hydrolyzes ATP to transport three intracellular sodium ions (Na+) out of the cell and two potassium ions (K+) into the cell against a 10-fold or greater concentration difference of ≥10-fold (Figure 2).
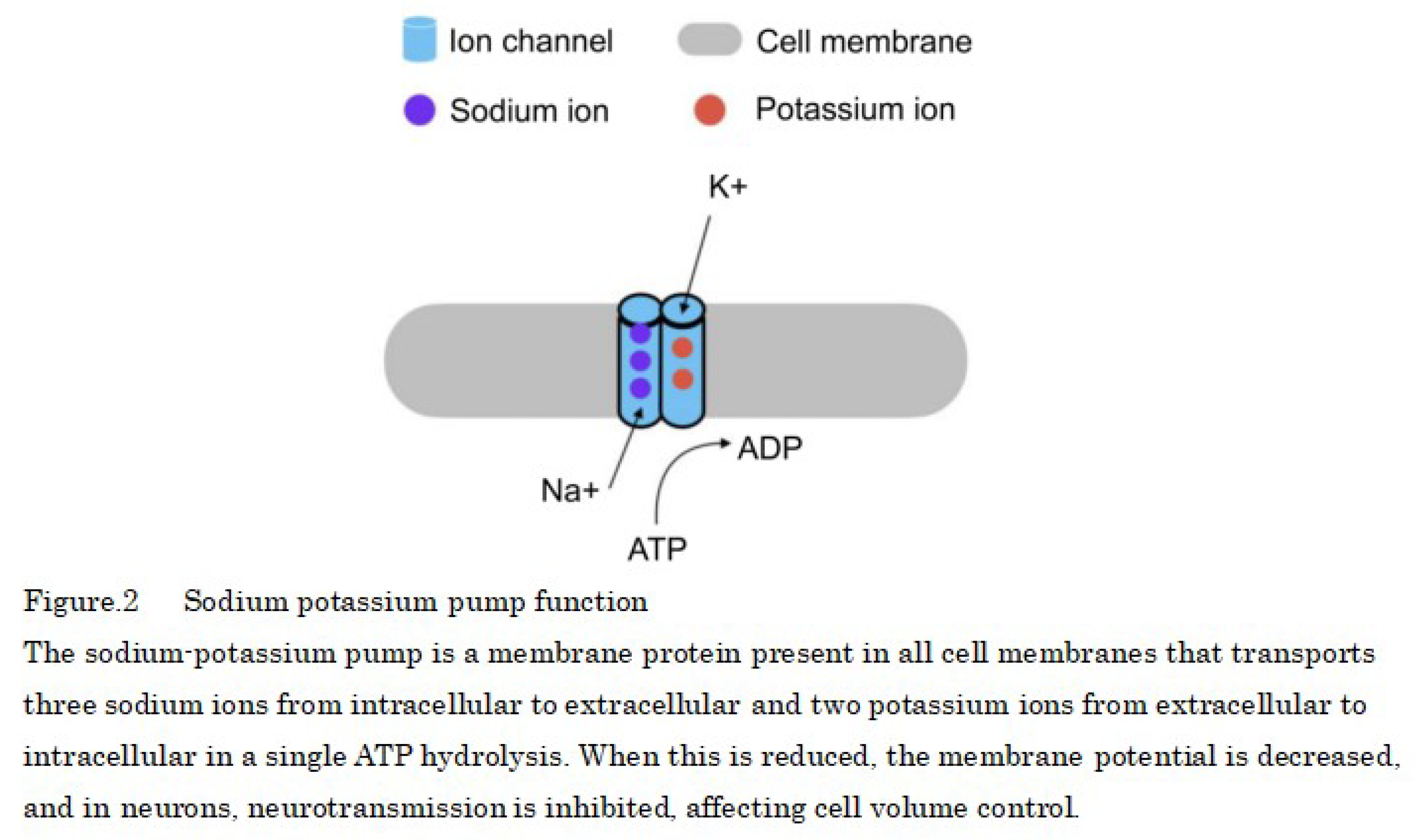
In normal cells, potassium ions are predominantly intracellular (98% intracellular, 2% extracellular), whereas sodium ions are predominantly extracellular. However, as metabolism declines and ATP is reduced, the ratio of passive transport, which is driven by differences in the concentration of substances, increases, and ions flow through the cell membrane from higher to lower concentrations.
Prokaryotic cells also have hair-like cell organelles called flagella that are propelled by hydrogen ions, and eukaryotic cells, such as animals are thought to have evolved from prokaryotic cells (Figure 3). Eukaryotic cells also have primary cilia, which are descendants of have evolved from flagella and growth only one per cell but, in animals, the primary cilia are immobile because the cells are attached to each other by via an extracellular matrix.
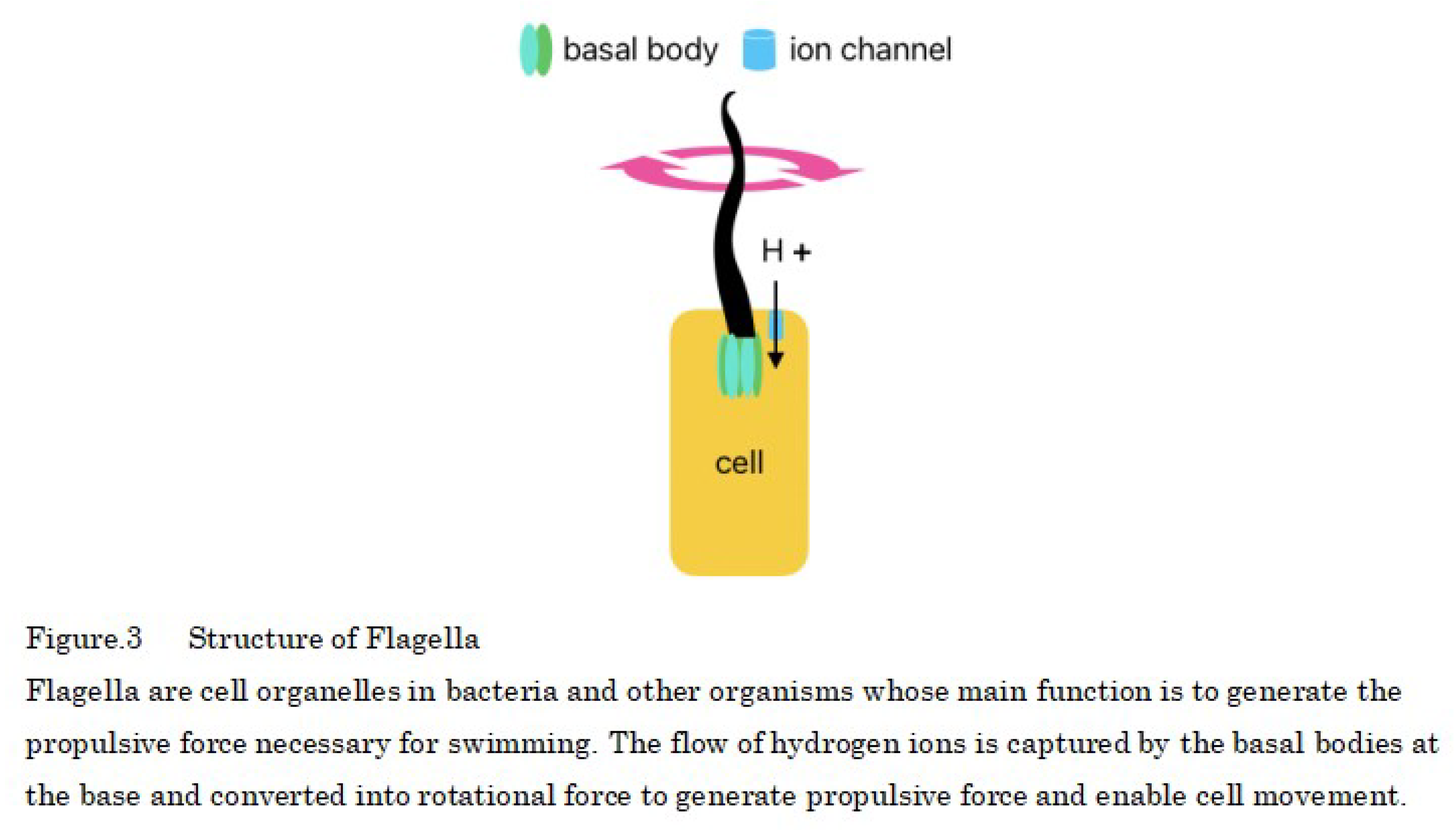
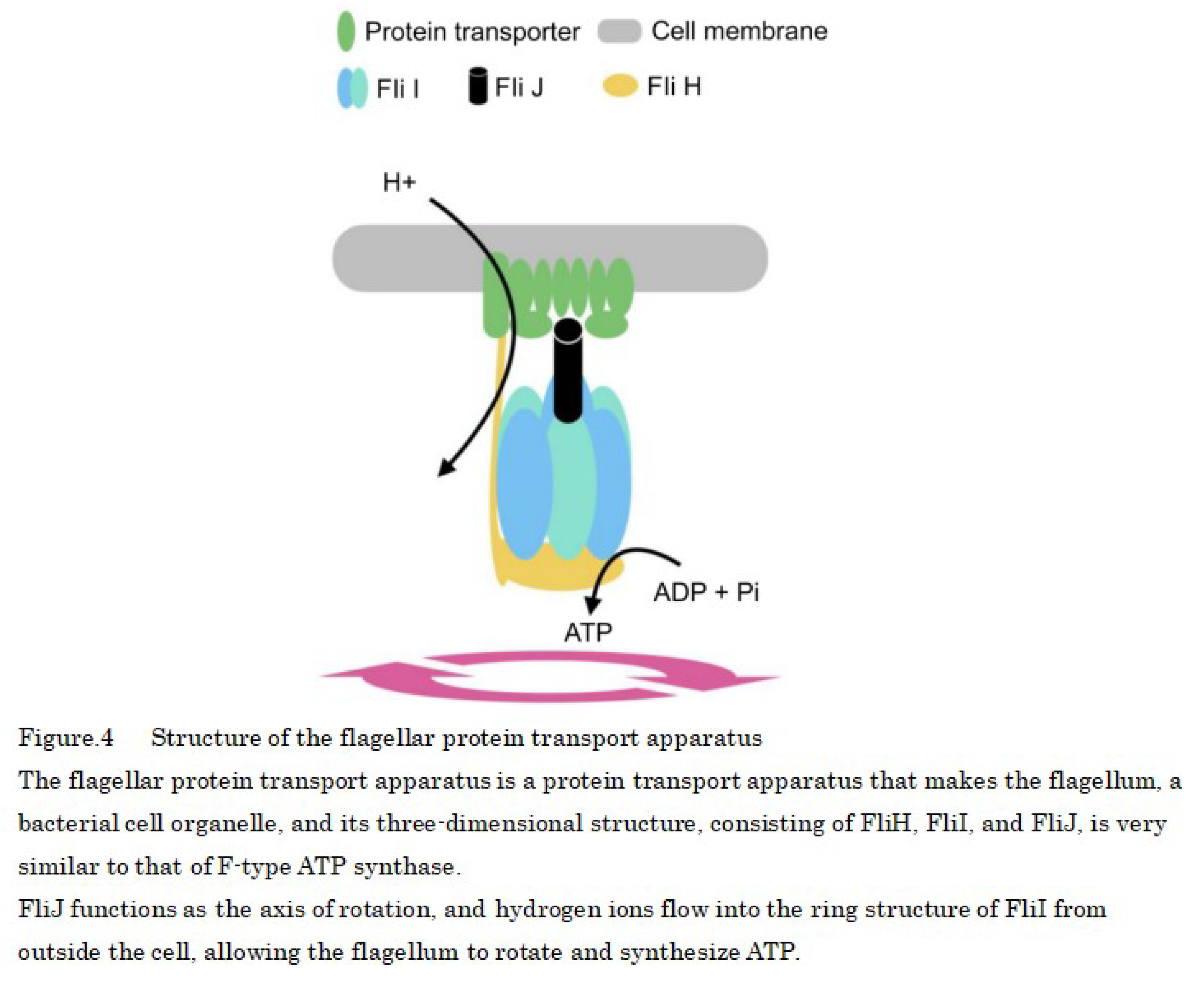
Proteins that make up the flagella of motile prokaryotes are transported from cells to the flagellar tip by a protein transport apparatus, where they bind to the flagellar tip, and the construction of the flagellum proceeds. This transport apparatus consists of six membrane proteins and three soluble proteins, among which FliJ is the soluble protein essential for transport function. The three-dimensional structure of FliJ closely resembles that of the γ-subunit of the F-type ATP synthase in all organisms, and the complex formed by FliJ and FliI is nearly identical to a part of F-type ATP synthase in all organisms [33]. The presence of motor proteins in the cell wall of prokaryotes allows them to synthesize ATP instead of moving the flagellum [34] (Figure 4).
The structure of the flagellar motor protein is as follows: The rotary portion of the flagellar motor protein contains a passageway through which hydrogen ions flow, and when hydrogen ions flow into the rotary portion because of differences in ion concentration and pass through the MotA portion—one of the proteins that form the flagellar motor—the MotA changes shape or moves [35], and this force is transmitted to the motor shaft and is used as the driving energy for ATP synthesis by rotating the shaft using the flow of hydrogen ions, similar to a water wheel using water flow (Figure 4).
Prokaryotic flagella have a 9 + 2 structure (9 doublet microtubules surround 2 singlet microtubules) and are motile, whereas the primary cilia of mammals such as humans have a 9 + 0 structure, are non-motile, and are present in all normal cells, except those of the blood cell lineage. The functions of the primary cilia range from being biochemical elements, i.e., involved in the secretion of growth factors, hormones, and neuropeptides, to being antennas for light, temperature, gravity, and cells. Also, the microtubules are highly acetylated and stabilized.
Additionally, mammalian primary cilia rotate from the right to the left in the early fetal period as a result of the water flow into a depressed area called the motile node [36], which causes the rotation of the primary cilia from the right to the left, resulting in the flow of body fluids. In response to this stimulus, the signaling factors Nodal and Lefty, which define the left side of the body, are induced, breaking symmetry and forming a left-right axis, producing asymmetrical organs [37,38,39] (Figure 5).
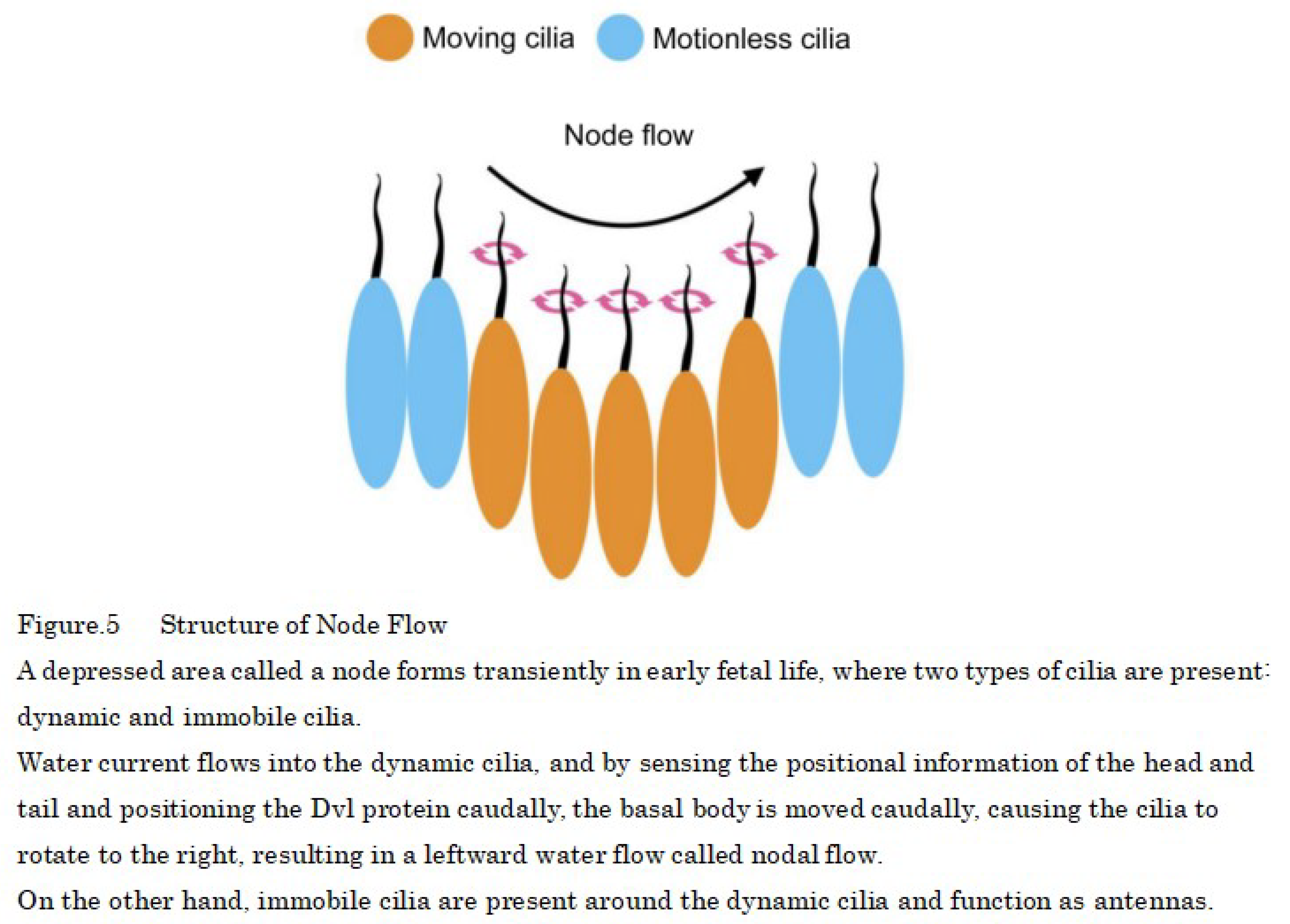
The motility of the nodal cilia is attributable to Cfap53, which is present in axonemes and binds the outer arm dynein, which drives the cilia, to the axonemes thus, Cfap53 expression is strictly limited to cells with motile cilia [40]. The rightward rotation of cilia is caused by occurs when the basal body being is biased caudally in nodal cells, and nodal cells sense the head–tail position information and position the Dvl protein caudally, moving the basal body to the caudal side, which results in the cilia caudal tilting [41].
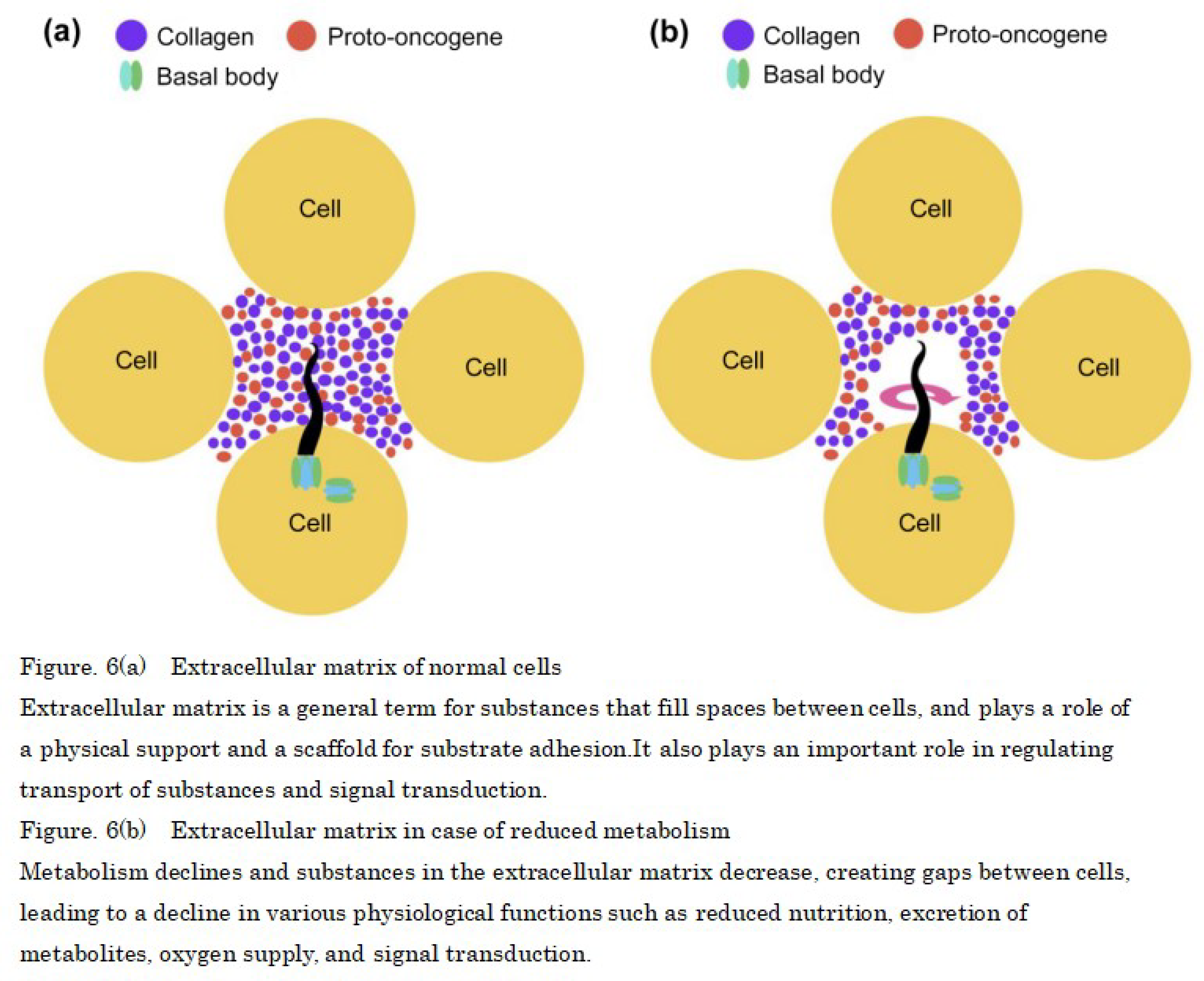
Furthermore, cells are attached to each other by extracellular matrix [42,43] (Figure 6a), but when metabolism decreases, it suggests that the slower speed of translation from the proto-oncogene reduces the amount of protein and collagen in the proto-oncogene and creates intercellular spaces in the extracellular matrix, which allows space for primary cilia to rotate, similar to the depressed state of nodal cells during the fetal stage. The basal body of the primary cilium is connected by fixing the axoneme, and the basal body has a pinwheel-like structure derived from the appendage structure of the mother centriole. When decrease in ATP decreases during active transport, which is—i.e., the normal ion transport of the cell membrane—leads to an increase in the proportion of passive transport increases, sodium ions flow in from the extracellular matrix similar to flagellum movements in prokaryotes, and rotate the basal body like a waterwheel. (Figure 6b).
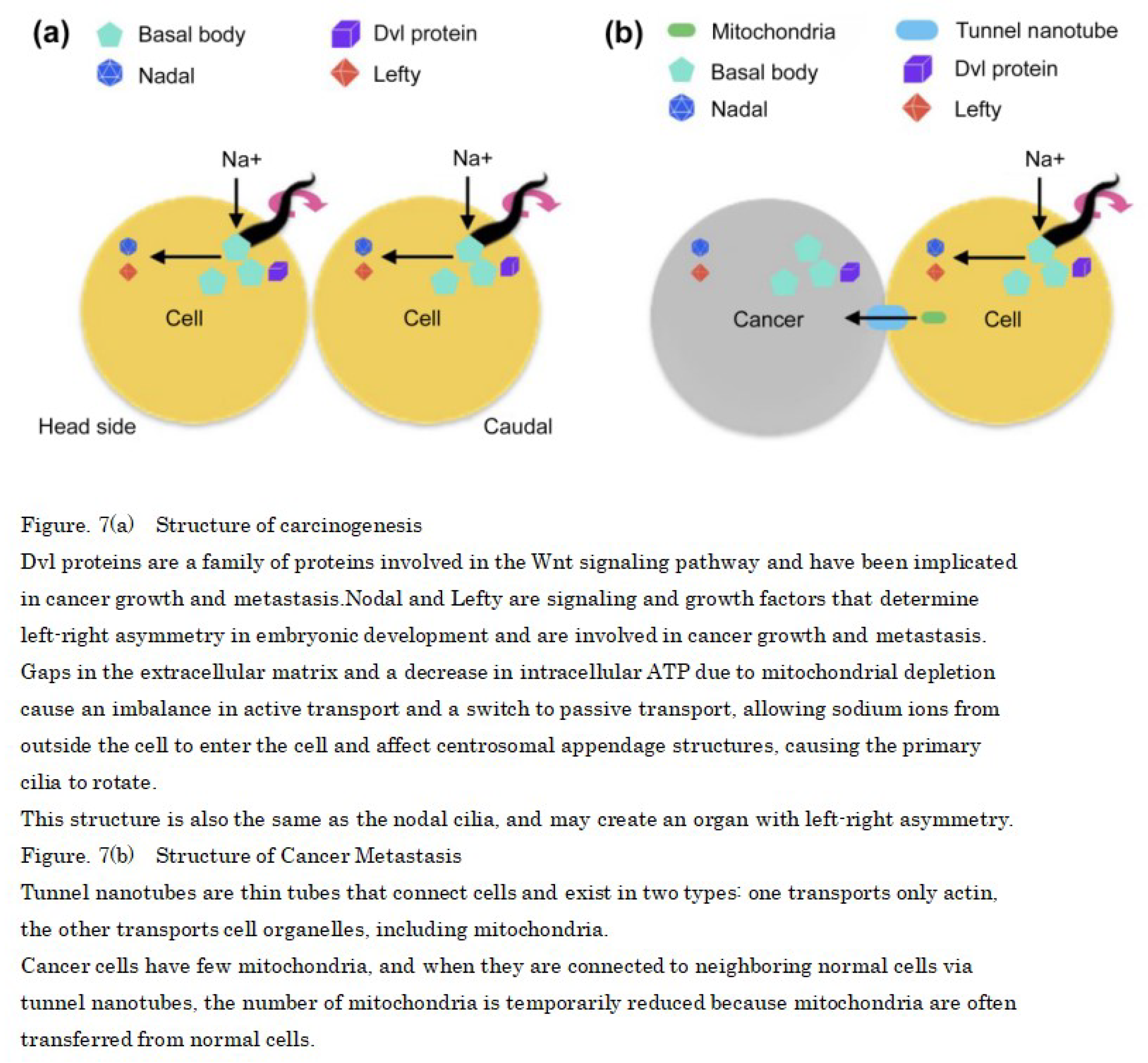
This suggests that the Dvl protein and the signaling factors Nodal and Lefty that regulate the left side are highly expressed in cancer cells, so that the primary cilia of normal cells rotate from right to left and become cancerous [44,45]. In addition, overexpression of Plk4, STIL, and HsSAS-6, which control centrosome replication in cancer cells, results in the concomitant formation of multiple daughter centrioles, which implies that primary cilia rotation is involved in gene regulation (Figure 7a).
Furthermore, mitochondria are known to move between adjacent cells via tunneling nanotubes not only in normal cells but also in cancer cells [46,47]. One possible cause of cancer erosion is the transfer of mitochondria from normal cells to cancer cells, resulting in decreased mitochondria in normal cells, which may lead to decreased metabolism and rotation of primary cilia (Figure 7b).
Considering that age-related changes in mitochondrial function and hypothermia due to low metabolism affect proteins and enzymes, we are proposing a carcinogenic mechanism.
In the early stages of mammalian DNA replication, the double helix of DNA needs to be unwound, and DNA helicases use ATP to unwind it into single-stranded DNA. However, in the 5' to 3' direction of DNA synthesis, DNA is too tightly wound, resulting in a positive superhelix. Therefore, a longer positive superhelix indicates greater resistance to unwinding, and it stops the replication fork (Figure 8a). For ensuring that DNA replication proceeds smoothly, the positive superhelix must be neutralized by winding the DNA to introduce a superhelix in the opposite direction (negative superhelix) into the DNA [48]. This can be achieved using the DNA topoisomerase I, which is expressed evenly in the nucleoplasm during mitosis and is required to neutralize the positive superhelix by breaking one of the two DNA strands without the need for ATP, which allows the other strand to pass between the breaks and changes the linking number by one when the breaks rejoin [49,50] (Figure 8b).
The role of DNA topoisomerase I can be supplemented by DNA topoisomerase IIα [51,52]. This enzyme is highly expressed during mitosis and is required, in the presence of ATP, to simultaneously break and rotate two DNA strands. The DNA topoisomerase IIα changes the number of linkages by two when the breaks rejoin and neutralizes the positive superhelix (Figure 8c).
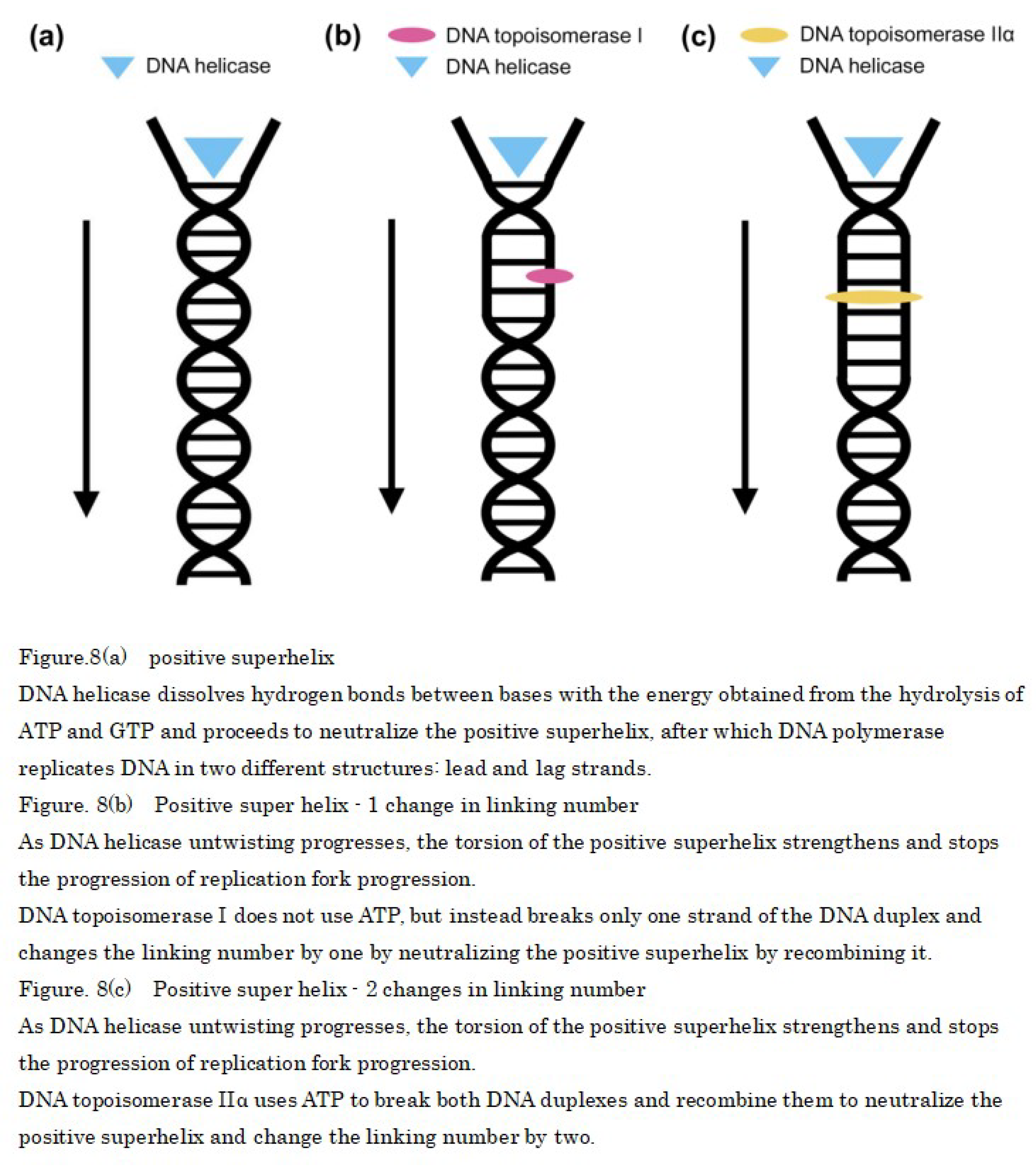
These observations indicated that after DNA helicase uses ATP to unwind the DNA double helix at the start of DNA replication, ATP is temporarily low; however, during the growth phase, DNA topoisomerase IIα is used owing to the abundance of ATP, and two DNA strands are cut simultaneously to achieve rapid replication. However, after the growth phase, DNA topoisomerase I is used because of the decrease in ATP, and it is possible that only one of the two DNA strands is cut, resulting in a decrease in the rate of DNA replication.
In addition, DNA topoisomerase IIα is expressed in dividing neural progenitor cells, and its expression ceases as they differentiate. Instead, DNA topoisomerase IIβ began to be expressed, and it is required to activate the transcription of several genes whose expression was induced in neurons.
Once single-stranded DNA is formed by DNA helicase, daughter strand synthesis occurs through the effects of DNA polymerase; however, after DNA replication, sister chromatids must be separated. DNA topoisomerase IIα is responsible for this and the activity of DNA topoisomerase IIα which simultaneously cleaves two DNA strands in the presence of ATP and allows the passage of another DNA strand between them is essential for the division and separation of sister chromatids in decatenanization, which dissolves the entanglement (catenan) that occurs between two daughter duplex DNA strands after DNA replication [53]. In cases when the DNA topoisomerase IIα is inhibition, the two replicated chromosomes cannot be separated, and if the genome that needs to be distributed via cell division cannot be separated, cell division fails and chromosome fusion occurs (Figure 9).
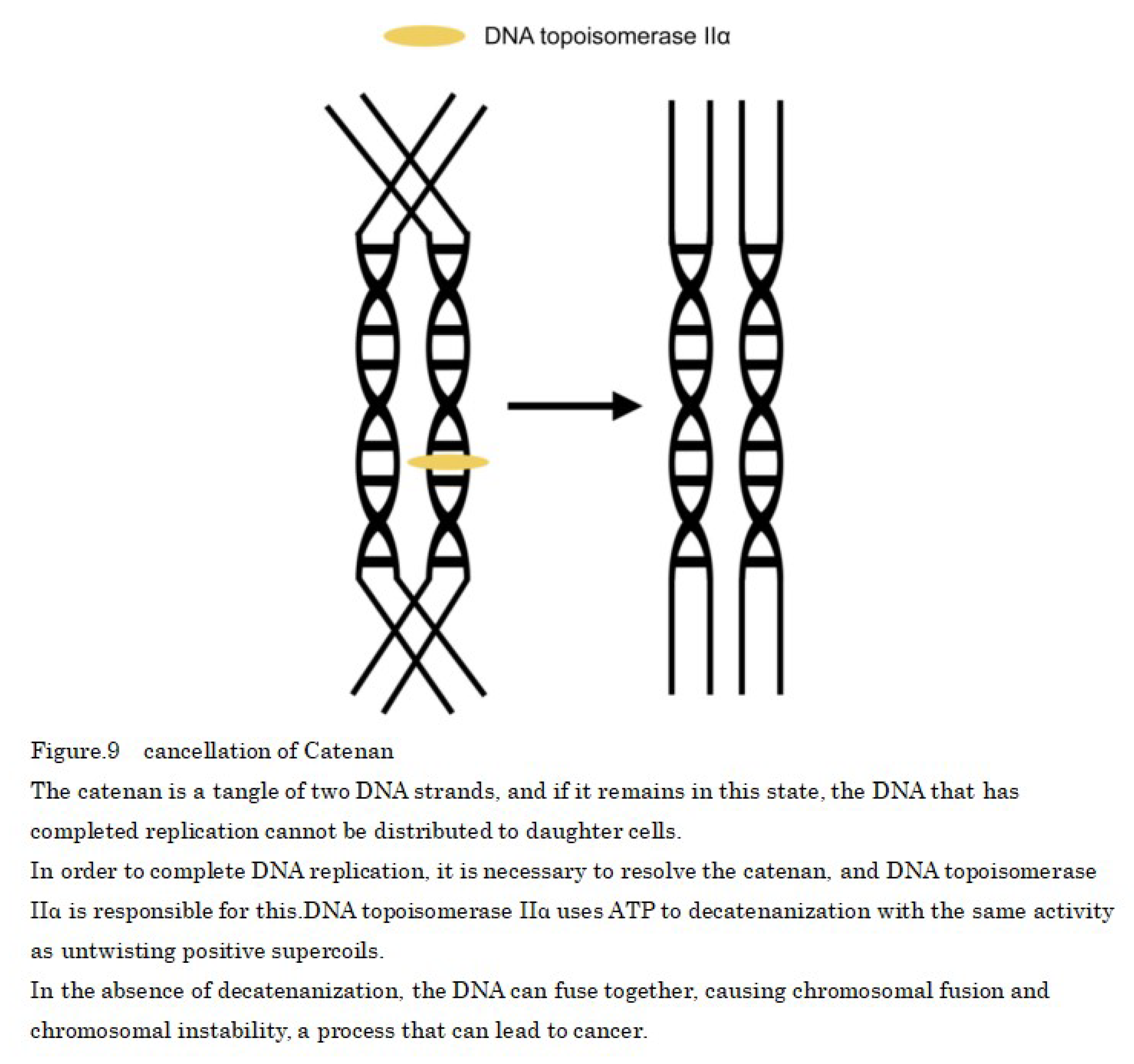
This is suggesting to be the result of delayed chromosomal segregation of DNA topoisomerase IIα due to decreased ATP production from age-related mitochondrial depletion and loss of normal function due to disruption or mutation of two alleles of DNA topoisomerase IIα. In addition, temperature sensitivity may be observed when mouse DNA topoisomerase IIα is introduced into a yeast DNA topoisomerase IIα disruptant strain by the plasmid shuffle method after mutagen treatment.
Regarding the temperature sensitivity of DNA topoisomerase IIα, it can growth at 28°C but cannot growth at 35°C and shows negligible growth at 37°C. However, this is not due to a decrease in the protein of DNA topoisomerase IIα, it instead occurs owing to a decrease in enzyme activity. Therefore, a temperature of 35°C, at which DNA topoisomerase IIα cannot growth at all, coincides with the temperature at which cancer cells proliferate most, suggesting that a decrease in body temperature due to a metabolic decline causes chromosome fusion because the DNA topoisomerase IIα cannot growth and chromosomes cannot be separated.
Conclusion
These facts suggested the possibility that etiology of cancer in mammals and other eukaryotes is involves the decrease in the number of mitochondria, which leads to a decrease in collagen and the products of the proto-oncogenes, which creates gaps between cell–cell attachments and allows the primary cilia to rotate, which consequently causes the primary cilia to rotate like node cilia that form during fetal development. These changes cause generate asymmetrical water flow, which induces the expression of Nodal and Lefty signaling factors that define the left side; however, in older adults, the body temperature can be close to 35°C [54,55], which affects growth of the temperature-sensitive DNA topoisomerase IIα. Furthermore, DNA topoisomerase IIα takes longer to separate chromosomes when ATP is low and cells divide with fused chromosomes without separation. As a result, chromosomes are destruction forming cells with properties different from their original nature, i.e., cancer, and the absence of apoptosis of cancer cells by mitochondria is suggested to be the cause of cancer growth.
This is consistent with the results of an experiment comparing the weekly averages of number of hamster wheel rotations, body weight, and urinary inorganic phosphate and pH in cancer and usually mice, which showed that the hamster wheel turnover was temporarily lower in cancer mice before skin cancer (Fig. 1(a)) and that the increase and decrease in weight after skin cancer was severe (Fig. 1(b)), indicating that protein production is slowed down in a state of low metabolism. However, there was no significant difference in inorganic phosphate in urine (Fig. 1(c)), suggesting that cancer mice may accumulate large amounts of lactic acid in their bodies because they synthesize ATP through anaerobic respiration, or that inorganic phosphate in the body easily binds to calcium ions, reducing their function, which is essential for muscle contraction.However, there is no significant difference in inorganic phosphate in urine (Fig 1(d)), indicating that cancer mice synthesize more ATP through anaerobic respiration, resulting in the production of more lactic acid in the body and fatigue accumulation, but when comparing urine pH (Fig 1(e)), the usually mice have a lower pH before death in life span, even considering kidney deterioration, indicating that the blood is more acidic, which is an aging phenomenon.
In addition, new blood vessels must be regenerated for benign tumors in mammals and other organisms to change into malignant tumors and metastasize [56]; however, no distant cancer cell metastasis has been reported in plants even though they have leaf veins corresponding to blood vessels, and because the primary cilia of plants have a structure without a central body, ions from outside the cell flow in highly likely do not rotate the primary cilia, preventing cancer metastasis [57,58].
In this study, by focusing on the primary cilia rotation and the properties of temperature-sensitive DNA topoisomerase IIα, which are the mechanisms of carcinogenesis, we investigated the previously reported chromosomal instability and loss of alleles of tumor suppressor genes, and were able to mark the steps involved in carcinogenesis in greater detail than previous evidence such as DNA methylation abnormalities and collagen phagocytosis. However, the effects of mitochondrial depletion alone are limited to the prevention of carcinogenesis the future, research on the avoidance of mitochondrial DNA damage associated with aging and treatment that promotes self-proliferation of mitochondria is warranted to be effective. The above findings indicate that promoting metabolism may lead to cancer prevention and treatment, and a result highlighting a decline in metabolism that causes cancer in addition to DNA replication errors is very important, which has been conventionally believed to be the cause of cancer.
Data availability statement for share upon reasonable request data sharing policy: Not applicable
Competing interests
This paper was prepared by an individual and has no competing interests.
Funding
This study was self-funded.
Acknowledgements
In this study, aimed to clarify the structure of cancer, and were able to obtain useful knowledge by investigating many papers. I would like to express my heartfelt gratitude to all the pioneers of cancer research for being able to present the results of this research.
References
- Nichols, C.A.; Gibson, W.J.; Brown, M.S.; Kosmicki, J.A.; Busanovich, J.P.; Wei, H.; Urbanski, L.M.; Curimjee, N.; Berger, A.C.; Gao, G.F.; et al. Loss of heterozygosity of essential genes represents a widespread class of potential cancer vulnerabilities. Nat. Commun. 2020, 11, 2517. [Google Scholar] [CrossRef]
- Engeland, K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022, 29, 946–960. [Google Scholar] [CrossRef]
- Nishiyama, A.; Nakanishi, M. Navigating the DNA methylation landscape of cancer. Trends Genet. 2021, 37, 1012–1027. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-H.; Wu, C.-F.; Rajasekaran, N.; Shin, Y.K. Loss of Tumor Suppressor Gene Function in Human Cancer: An Overview. Cell. Physiol. Biochem. 2018, 51, 2647–2693. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Zhang, H. Extracellular matrix: an important regulator of cell functions and skeletal muscle development. Cell Biosci. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Baart, V.M.; Houvast, R.D.; de Geus-Oei, L.F.; Quax, P.H.A.; Kuppen, P.J.K.; Vahrmeijer, A.L.; Sier, C.F.M. Molecular imaging of the urokinase plasminogen activator receptor: opportunities beyond cancer. EJNMMI Res. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Zhang, H.; Menzies, K.J.; Auwerx, J. The role of mitochondria in stem cell fate and aging. Development 2018, 145, dev143420. [Google Scholar] [CrossRef] [PubMed]
- van der Reest, J.; Cecchino, G.N.; Haigis, M.C.; Kordowitzki, P. Mitochondria: Their relevance during oocyte ageing. Ageing Res. Rev. 2021, 70, 101378. [Google Scholar] [CrossRef]
- Nissanka, N.; Moraes, C.T. Mitochondrial DNA damage and reactive oxygen species in neurodegenerative disease. FEBS Lett. 2017, 592, 728–742. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Tamrakar, P.; Rosenthal, R.E.; Fiskum, G. Calcium uptake and cytochrome c release from normal and ischemic brain mitochondria. Neurochem. Int. 2018, 117, 15–22. [Google Scholar] [CrossRef]
- Rossi, A.; Pizzo, P.; Filadi, R. Calcium, mitochondria and cell metabolism: A functional triangle in bioenergetics. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Bazylianska, V.; Kalpage, H.A.; Wan, J.; Vaishnav, A.; Mahapatra, G.; Turner, A.A.; Chowdhury, D.D.; Kim, K.; Morse, P.T.; Lee, I.; et al. Lysine 53 Acetylation of Cytochrome c in Prostate Cancer: Warburg Metabolism and Evasion of Apoptosis. Cells 2021, 10, 802. [Google Scholar] [CrossRef] [PubMed]
- Rong, N.; Mistriotis, P.; Wang, X.; Tseropoulos, G.; Rajabian, N.; Zhang, Y.; Wang, J.; Liu, S.; Andreadis, S.T. Restoring extracellular matrix synthesis in senescent stem cells. FASEB J. 2019, 33, 10954–10965. [Google Scholar] [CrossRef]
- Higgins, M.; Obaidi, I.; McMorrow, T. Primary cilia and their role in cancer (Review). Oncol. Lett. 2019, 17, 3041–3047. [Google Scholar] [CrossRef]
- Cao, M.; Zou, X.; Li, C.; Lin, Z.; Wang, N.; Zou, Z.; Ye, Y.; Seemann, J.; Levine, B.; Tang, Z.; et al. An actin filament branching surveillance system regulates cell cycle progression, cytokinesis and primary ciliogenesis. Nat. Commun. 2023, 14, 1–18. [Google Scholar] [CrossRef]
- Daggubati V, Vykunta A, Choudhury A, et al. Hedgehog target genes regulate lipid metabolism to drive basal cell carcinoma and medulloblastoma.Research Square. 2023;1.
- Kim, D.H.; Ahn, J.S.; Han, H.J.; Kim, H.-M.; Hwang, J.; Lee, K.H.; Cha-Molstad, H.; Ryoo, I.-J.; Jang, J.-H.; Ko, S.-K.; et al. Cep131 overexpression promotes centrosome amplification and colon cancer progression by regulating Plk4 stability. Cell Death Dis. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Hoffmann, I. Role of Polo-like Kinases Plk1 and Plk4 in the Initiation of Centriole Duplication—Impact on Cancer. Cells 2022, 11, 786. [Google Scholar] [CrossRef]
- Wu, Q.; Li, B.; Liu, L.; Sun, S.; Sun, S. Centrosome dysfunction: a link between senescence and tumor immunity. Signal Transduct. Target. Ther. 2020, 5, 1–9. [Google Scholar] [CrossRef]
- Sabourian, P.; Yazdani, G.; Ashraf, S.S.; Frounchi, M.; Mashayekhan, S.; Kiani, S.; Kakkar, A. Effect of Physico-Chemical Properties of Nanoparticles on Their Intracellular Uptake. Int. J. Mol. Sci. 2020, 21, 8019. [Google Scholar] [CrossRef]
- Adachi N. koutou sinkaku seibutu no ni gata DNA topoisomera-ze no kouzou to kinou[Structure and function of type II DNA topoisomerase from higher eukaryotes]. Seikagaku J Jpn Biochem Soc. 1995;67(1):36–40. Japanese.
- Madabhushi, R. The Roles of DNA Topoisomerase IIβ in Transcription. Int. J. Mol. Sci. 2018, 19, 1917. [Google Scholar] [CrossRef]
- Kumiko, T. Temperature-dependent gene expression in hyperthermia. Japanese journal of health & research. 2018;39:46-53. [CrossRef]
- Mielczarek-Lewandowska, A.; Hartman, M.L.; Czyz, M. Inhibitors of HSP90 in melanoma. Apoptosis 2019, 25, 12–28. [Google Scholar] [CrossRef]
- Somero, G.N. The cellular stress response and temperature: Function, regulation, and evolution. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2020, 333, 379–397. [Google Scholar] [CrossRef]
- Chen, W. Thermometry and interpretation of body temperature. Biomed. Eng. Lett. 2019, 9, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Ichise, H.; Tsukamoto, S.; Hirashima, T.; Konishi, Y.; Oki, C.; Tsukiji, S.; Iwano, S.; Miyawaki, A.; Sumiyama, K.; Terai, K.; et al. Functional visualization of NK cell-mediated killing of metastatic single tumor cells. eLife 2022, 11. [Google Scholar] [CrossRef]
- Glenn, A.; Armstrong, C.E. Physiology of red and white blood cells. Anaesth. Intensiv. Care Med. 2019, 20, 170–174. [Google Scholar] [CrossRef]
- Bowden Davies, K.A.; Pickles, S.; Sprung, V.S.; Kemp, G.J.; Alam, U.; Moore, D.R.; Tahrani, A.A.; Cuthbertson, D.J. Reduced physical activity in young and older adults: metabolic and musculoskeletal implications. Ther. Adv. Endocrinol. Metab. 2019, 10. [Google Scholar] [CrossRef]
- Park, Y.; Choi, J.-E.; Hwang, H.-S. Protein supplementation improves muscle mass and physical performance in undernourished prefrail and frail elderly subjects: a randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2018, 108, 1026–1033. [Google Scholar] [CrossRef]
- Rubio-Aliaga, I.; Krapf, R. Phosphate intake, hyperphosphatemia, and kidney function. Pfl?gers Arch. Eur. J. Physiol. 2022, 474, 935–947. [Google Scholar] [CrossRef]
- Rajkumar, P.; Pluznick, J.L. Acid-base regulation in the renal proximal tubules: using novel pH sensors to maintain homeostasis. Am. J. Physiol. Physiol. 2018, 315, F1187–F1190. [Google Scholar] [CrossRef]
- Matzke, N.J.; Lin, A.; Stone, M.; Baker, M.A.B. Flagellar export apparatus and ATP synthetase: Homology evidenced by synteny predating the Last Universal Common Ancestor. BioEssays 2021, 43, 2100004. [Google Scholar] [CrossRef]
- Guo, H.; Suzuki, T.; Rubinstein, J.L. ; Japan Structure of a bacterial ATP synthase. eLife 2019, 8. [Google Scholar] [CrossRef]
- Nakamura, S.; Minamino, T. Flagella-Driven Motility of Bacteria. Biomolecules 2019, 9, 279. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Shiozawa, K.; Katoh, T.A.; Minegishi, K.; Ide, T.; Ikawa, Y.; Nishimura, H.; Takaoka, K.; Itabashi, T.; Iwane, A.H.; et al. Role of Ca 2+ transients at the node of the mouse embryo in breaking of left-right symmetry. Sci. Adv. 2020, 6, eaba1195. [Google Scholar] [CrossRef]
- Little, R.B.; Norris, D.P. Right, left and cilia: How asymmetry is established. Semin. Cell Dev. Biol. 2021, 110, 11–18. [Google Scholar] [CrossRef]
- Szenker-Ravi, E.; Ott, T.; Khatoo, M.; de Bellaing, A.M.; Goh, W.X.; Chong, Y.L.; Beckers, A.; Kannesan, D.; Louvel, G.; Anujan, P.; et al. Discovery of a genetic module essential for assigning left–right asymmetry in humans and ancestral vertebrates. Nat. Genet. 2021, 54, 62–72. [Google Scholar] [CrossRef]
- Petri, N.D. Evolutionary Diversity of the Mechanisms Providing the Establishment of Left-Right Asymmetry in Metazoans. Russ. J. Dev. Biol. 2020, 51, 84–98. [Google Scholar] [CrossRef]
- Ide, T.; Twan, W.K.; Lu, H.; Ikawa, Y.; Lim, L.-X.; Henninger, N.; Nishimura, H.; Takaoka, K.; Narasimhan, V.; Yan, X.; et al. CFAP53 regulates mammalian cilia-type motility patterns through differential localization and recruitment of axonemal dynein components. PLOS Genet. 2020, 16, e1009232. [Google Scholar] [CrossRef]
- Xie, H.; Kang, Y.; Liu, J.; Huang, M.; Dai, Z.; Shi, J.; Wang, S.; Li, L.; Li, Y.; Zheng, P.; et al. Ependymal polarity defects coupled with disorganized ciliary beating drive abnormal cerebrospinal fluid flow and spine curvature in zebrafish. PLOS Biol. 2023, 21, e3002008. [Google Scholar] [CrossRef]
- Mehmet V. Cell-Extracellular Matrix Adhesion Assay. Epidermal Cells. 2019;2109:209-217. [CrossRef]
- Janiszewska, M.; Primi, M.C.; Izard, T. Cell adhesion in cancer: Beyond the migration of single cells. J. Biol. Chem. 2020, 295, 2495–2505. [Google Scholar] [CrossRef]
- Mei, J.; Yang, X.; Xia, D.; Zhou, W.; Gu, D.; Wang, H.; Liu, C. Systematic summarization of the expression profiles and prognostic roles of the dishevelled gene family in hepatocellular carcinoma. Mol. Genet. Genom. Med. 2020, 8, e1384. [Google Scholar] [CrossRef]
- Gambichler, T.; Ardabili, S.; Lang, K.; Dreißigacker, M.; Scheel, C.; Brand-Saberi, B.; Skrygan, M.; Stockfleth, E.; Käfferlein, H.; Brüning, T.; et al. Expression of Lefty predicts Merkel cell carcinoma-specific death. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 2016–2020. [Google Scholar] [CrossRef] [PubMed]
- Hekmatshoar, Y.; Nakhle, J.; Galloni, M.; Vignais, M.-L. The role of metabolism and tunneling nanotube-mediated intercellular mitochondria exchange in cancer drug resistance. Biochem. J. 2018, 475, 2305–2328. [Google Scholar] [CrossRef]
- Liu, D.; Gao, Y.; Liu, J.; Huang, Y.; Yin, J.; Feng, Y.; Shi, L.; Meloni, B.P.; Zhang, C.; Zheng, M.; et al. Intercellular mitochondrial transfer as a means of tissue revitalization. Signal Transduct. Target. Ther. 2021, 6, 1–18. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Dröge, P. Chromatin Architectural Factors as Safeguards against Excessive Supercoiling during DNA Replication. Int. J. Mol. Sci. 2020, 21, 4504. [Google Scholar] [CrossRef] [PubMed]
- Spakman, D.; Bakx, J.A.M.; Biebricher, A.S.; Peterman, E.J.G.; Wuite, G.J.L.; A King, G. Unravelling the mechanisms of Type 1A topoisomerases using single-molecule approaches. Nucleic Acids Res. 2021, 49, 5470–5492. [Google Scholar] [CrossRef]
- Soren, B.C.; Dasari, J.B.; Ottaviani, A.; Lacovelli, F.; Fiorani, P. Topoisomerase IB: a relaxing enzyme for stressed DNA. Cancer Drug Resist. 2019, 3, 18–25. [Google Scholar] [CrossRef]
- McKie, S.J.; Neuman, K.C.; Maxwell, A. DNA topoisomerases: Advances in understanding of cellular roles and multi-protein complexes via structure-function analysis. BioEssays 2021, 43, 2000286–e2000286. [Google Scholar] [CrossRef]
- Karol S, Arkadi M, Heather M R. Topoisomerase II contributes to DNA secondary structure-mediated double-stranded breaks. Nucleic Acids Research. 2020;48(12):6654–6671. [CrossRef]
- Broeck, A.V.; Lotz, C.; Drillien, R.; Haas, L.; Bedez, C.; Lamour, V. Structural basis for allosteric regulation of Human Topoisomerase IIα. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Miklós S, János G. Chapter 23 - Thermoregulation and age. Handbook of Clinical Neurology. 2018;156:377-395. [CrossRef]
- Geneva, I.I.; Cuzzo, B.; Fazili, T.; Javaid, W. Normal Body Temperature: A Systematic Review. Open Forum Infect. Dis. 2019, 6, ofz032. [Google Scholar] [CrossRef]
- Elebiyo, T.C.; Rotimi, D.; Evbuomwan, I.O.; Maimako, R.F.; Iyobhebhe, M.; Ojo, O.A.; Oluba, O.M.; Adeyemi, O.S. Reassessing vascular endothelial growth factor (VEGF) in anti-angiogenic cancer therapy. Cancer Treat. Res. Commun. 2022, 32, 100620. [Google Scholar] [CrossRef]
- Seyfried, T.N.; Huysentruyt, L.C. On the Origin of Cancer Metastasis. Crit. Rev. Oncog. 2013, 18, 43–73. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Liu, B. Microtubule nucleation for the assembly of acentrosomal microtubule arrays in plant cells. New Phytol. 2019, 222, 1705–1718. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



