
Submitted:
17 March 2024
Posted:
18 March 2024
You are already at the latest version
Abstract
The vomeronasal system serves as a pivotal element in mammalian sensory biology, allowing the perception of chemical signals essential for social communication via the activation of two main receptor families, V1R and V2R. The expression of V1Rs and V2Rs is linked with that of the G-protein alpha-subunits, Gαi2 and Gαo, respectively. The exploration of the evolutionary pathways of V1Rs and V2Rs across all mammalian species remains a main challenge, juxtaposing available genomic data against emerging immunohistochemical evidence. However, recent investigations, in contrast to what is predicted by the currently annotated genomic sequences, have revealed the expression of Gαo in the vomeronasal neuroepithelium of wild canids, including wolves and foxes. In the present study, the employment of a specific antibody raised against the mouse V2R2, member of the C-family of vomeronasal receptors, V2Rs, has confirmed the expression of this receptor in the fox and wolf, but it has revealed the lack of expression in the dog. This may reflect the impact of domestication on the regression of the VNS in this species, in contrast to their wild counterparts, and underscores the effects of artificial selection on sensory functions. Thus, these findings suggest a more refined chemical detection capability in wild species.
Keywords:
vomeronasal organ
; V2R
; G-proteins
; Canidae
; immunohistochemistry
1. Introduction
The ability of organisms to adapt to their environment is crucial for the survival of both individual members and the species as a whole. Evolutionary processes have endowed mammals with complex sensory systems, specialized organs crucial for sensing environmental cues and enabling continuous adaptation. Among these, the chemical senses, particularly olfactory subsystems, are key to managing essential survival functions, including communication among members [1], social interactions [2,3], and mating behaviors [4,5]. Chemical communication between individuals depends on chemical signals, called semiochemicals [6,7], produced by individuals, released into the environment through secretions [8,9] , and recognised by the olfactory systems of animals [10,11].
The principal mechanisms for odor and pheromone detection comprise the main olfactory system (MOS) and the vomeronasal system (VNS). The former operates through the main olfactory epithelium and the olfactory bulb and has a significant linkage to the limbic system, playing the main role in both memory retrieval and conscious perception [12]. On the other hand, the VNS, which comprises the vomeronasal organ (VNO) and the accessory olfactory bulb (AOB), is implicated in non-conscious influences in sexual behaviour [13,14], maternal identification [15,16], and the detection of predators [17,18], and, therefore, it is specifically tuned to sensing of pheromones [19], kairomones [10], steroids [20], and major histocompatibility complex (MHC) molecules [21,22].
Despite their proximity, these two systems maintain anatomical independence, exhibiting significant morphological and functional disparities, which suggest that they likely evolved separately [23,24,25,26]. However, functional dichotomy has been questioned, based on the presence of vomeronasal receptors (VR) in the olfactory mucosa and odorant receptors in the VNO [27,28,29].
Rodent species represent the primary model for studies concerning the VNS in mammals [30,31]. Shinohara and colleagues [32] first revealed the presence of Gαi2 and Gαo into non overlapping regions of the AOB, showing that the rostral part expressed Gαi2 whereas the caudal one, Gαo. Since the vomeronasal sensory neurons (VSNs) send their axonal projections to the AOB, it was possible to demonstrate that also the VNO is provided with a similar anatomical organization in several rodent species, in that, Gαi2 is specifically expressed in the apical neurons of the VNO whereas Gαo is expressed in the basal ones [33,34,35]. Unraveling the involvement of both G-proteins within the signal transduction sequence in VSNs proved essential to the discovery of the two main VR families. In fact, in the mouse VNO, Dulac and Axel [36] identified a set of receptors, named V1Rs, whose expression pattern specifically matched the Gαi2 expression profile.
The identification of a second VR family, type-2 V2R, in rat and mouse occurred in 1997 through simultaneous studies [37,38,39], cementing the notion of dual independent sensory pathways. In fact, V2Rs were shown to exclusively co-express with Gαo in the basal neurons of the VNO. Thus, the immunohistochemical profiling of Gαi2 and Gαo since has become a key method for indicating V1R and V2R receptor expression in the VNO, respectively. However, later studies, in the VNO of other mammals, such as goat [40], dog [41,42], horse and marmoset [41], sheep [43], hyrax [44], cat [45], hippopotami [46], meerkat [47], and cow and pig [48], demonstrated the expression of Gαi2 but not of Gαo. Unexpectedly, the wallaby (genus Notamacropus) was initially proposed as an alternative model in the organization of G-protein expression, only showing Gαo immunostaining, which was localized in the basal neurons of the VNO, but, unexpectedly, scattered throughout the AOB without any evident compartmentalization [49]; however, it has recently been demonstrated that this marsupial genus possesses canonical immunohistochemical labelling of Gαo and Gαi2 in both VNO and AOB [50].
The differentiation of two segregated pathways for pheromonal information processing, as demonstrated by immunohistochemistry, has received further support from genomic studies in different species [51,52,53,54]. However, more recent immunohistochemical observations have revealed the expression of Gαo in the VNO of species that, based on the available genomic data, were assumed to lack V2Rs, specifically in wild canids such as the fox and wolf [55,56], suggesting caution when dealing with incompletely annotated genomes, as is often the case of wild species.
In this context, a phenotypic characterization of the expression of vomeronasal receptors, especially of V1Rs and V2Rs in the VNO, would be desirable. For this reason, we have characterized the expression, through an immunohistochemical approach, of V2Rs in the VNO of wolf and fox, in which Gαo is abundantly expressed [55,56]. In this immunohistochemical study, we have also included, the VNO of the dog since, in this species, there is no definitive information to date [42,57].
2. Results
In our study, we have applied immunohistochemical techniques to investigate the expression pattern of V2R2 in the vomeronasal epithelium of three canid species: two wild, the wolf and the fox, and one domestic, the dog.
We have taken advantage of the availability of anti-V2R2, an antibody that recognizes all functional receptors of family-C V2Rs [58]. Anti-V2R2 is a polyclonal antibody that has been exhaustively characterized and validated in the mouse and rat VNO [59,60]. We used this antibody as family-C V2R are highly conserved throughout species.
The results of the immunolabeling with anti-V2R2 in the wolf VNO are presented in Figure. 1. The analysis revealed extensive labeling throughout the entire extent of the vomeronasal neuroepithelium, with pronounced intensity in its basal part. Moreover, the apical portion of the epithelium displayed a more diffuse labeling, in the dendritic processes of the VSNs. This pattern suggests a significant expression of V2R2 in the wolf VNO (Figure 1A).
Counterstaining with hematoxylin further confirmed the observed labeling pattern and facilitated the delineation of the structural characteristics of the cellular strata and the underlying lamina propria as well as the localization of V2R2 within specific cellular compartments (Figure 1B). Higher magnification of both non-counterstained (Figure 1C) and counterstained (Figure 1D,E) images provided a clearer insight into the cellular specificity of the V2R2 expression. The labeling corresponded predominantly to cells identified as VSNs, based on their spatial placement and distinctive morphological features. These cells exhibited a clean and sharply defined pattern of labeling, primarily concentrated in the soma. Additionally, we observed a diffuse labeling pattern in the apical region of the epithelium (Figure 1C). The counterstained slides showed that the labeling in the VSNs was concentrated in the apical portion of their somata (Figure 1D). Dendrites originating from these somata extended towards the apical surface, culminating in the formation of immunopositive dendritic knobs (Figure 1D).
Interestingly, not all the receptor cells displayed immunopositivity for V2R2, with some cellular somata in various locations remaining unstained (Figure 1D, E). Additionally, our observations confirmed the absence of immunopositivity in the diverse basal stem cells (Figure 1E). These findings underscore the specific localization and expression patterns of V2R2 within the wolf VNO, highlighting the receptor potential role in pheromonal signaling and its implications in the sensory functions of this species.
In the immunohistochemical study of V2R2 expression in the sensory epithelium of the fox VNO, we have as well obtained consistent labeling throughout the vomeronasal sensory neuroepithelium, especially pronounced in the basal area (Figure 2). Additionally, the apical zone shows a diffuse pattern of labeling (Figure 2A). Hematoxylin counterstaining of the neuroepithelium facilitates the visualization of the structural features of the cellular layers and the lamina propria (Figure 2B). No counterstained higher magnification images confirm the established labeling pattern. This labeling displays a distinct arrangement, mainly concentrated in the soma. Additionally, the apical part of the epithelium exhibits a more widespread staining pattern (Figure 2C). Counterstaining at higher magnification shows the labeling mostly concentrated at the somata apical end. The dendritic ends are also immunopositive. As expected, not every VSN cell is immunopositive for V2R2. The basal stem cells display no immunopositivity (Figure 2D,E). These findings highlight the expression of V2R2 within the VNO sensory epithelium of the fox.
The immunohistochemical labeling of the dog VNO using the anti-V2R2 antibody did not result in immunostaining of any structural components of the organ in all the examined sections (Figure 3A,E), which contrasts with the clear labeling obtained in the vomeronasal sensory epithelium of the wolf (Figure 3D). Counterstaining of the immunostained sections allowed for the clear distinction between the sensory epithelium and the non-sensory or respiratory epithelium, highlighting the structural organization of the dog VNO (Figure 3B,C). The sensory epithelium displayed the neuronal layers, the mucomicrovillar complex on its surface, and the underlying lamina propria. All of which were found to be immunonegative, confirming the absence of V2R2 immunoreactivity in these structures (Figure 3E).
Finally, given that the expression of V2Rs is tightly linked to that of Gao, we conducted an immunohistochemical study on VNO sections of the wolf and fox vomeronasal organ to assess whether this transduction protein was also detectable in these samples. The results are presented in Figure 4. In both species, immunopositivity for Gao was observed in the sensory epithelium of the VNO. The immunolabeling comprised a subpopulation of non-receptor cells and extended to both the cellular somas and the dendritic processes present on the luminal surface (Figure 4A,B).
3. Discussion
The vomeronasal system in mammals has undergone highly sophisticated and complex evolution, leading to significant diversity within vomeronasal receptor families that are characterized by extensive gene diversity and gene inactivation across species [25,61]. Understanding how this diversity arose requires a phylogenetic perspective, in which the system is characterized from a morphological, biochemical, and comparative genomic viewpoint, avoiding making extrapolations between groups, even those that appear to be very closely related, and taking into account the interaction between the main and accessory olfactory systems [26,62].
For decades, it has been widely accepted that there are two models of expression for the two main families of vomeronasal receptors, V1R and V2R: a segregated model (expressing both families) and a uniform model (expressing only V1R). The segregated model would include Rodents [24,63], Lagomorphs [64], Marsupials [33], and Strepsirrhine Primates [65,66], while the rest of the investigated mammals would fall under the uniform model [19,41]. The distinction between these models was based on information originally derived from the expression of the Gαi2 and Gαo, as they are considered relevant markers for the expression of V1Rs and V2Rs, respectively. The development of genomics in recent decades has helped to clarify the enormous disparity and diversity of morphological data available on the vomeronasal system. However, there are still some mismatches between genetic and morphological information in numerous mammalian groups [67,68]. Current genomic evidence sometimes clashes with histochemical data, and vice versa as immunological features challenge the information provided by genomics.
In our study of the vomeronasal system in wild canids, specifically wolves and foxes, we encountered the latter situation. While the current genomic information seems to exclude the presence of V2Rs in these species, thus assigning them to the uniform model, our immunohistochemical data demonstrate the expression of both Gαo and V2Rs at the protein level, thereby aligning the VNO of fox and wolf with the segregated model.
To our knowledge, this study represents the first demonstration of the expression of a V2R beyond a laboratory rodent model, namely rat and mouse. This has also allowed the confirmation of the likely role in pheromone transduction of Gαo positive cells found in both species, fox and wolf. As V2R2 belongs to family-C V2Rs, this finding has additional implications. Studies on the expression of the V2R2 have demonstrated that genes of family-C are expressed in all basal neurons of the VNO and that are highly conserved throughout species making its antibody useful for detecting Gαo pathways in many mammals.
The results obtained in the current study regarding immunostaining with V2R2 in the VNO of both wolf and fox are clearly consistent with the expression of this receptor in the vomeronasal neuroepithelium. Specifically, the staining is clear both in the somas and in the dendritic processes that reach the mucomicrovillar complex covering the luminal surface of the epithelium. It is noteworthy that there is no staining in the axonal terminals that leave the somas at their basal part, with staining concentrated on the apical part of the somas. This absence of axonal staining coincides with observations made by Martini and colleagues [58] in mouse and rat.
Although previous studies on the expression of Gαo in the VNO of fox and wolf were already available [55,56], for this study, we have confirmed these results using antibodies against Gαo from two different commercial brands. In both cases, the staining pattern is comparable to that observed with the V2R2 antibody. It is noteworthy that, while in the case of laboratory rodents there is a basal-apical zonation of the VNO neuroepithelium in terms of G-protein and receptor expression, in canids, the reduced size of the organ does not allow to highlight this differentiation.
A second notable aspect of our study is the absence of immunoreactivity for V2R2 in the VNO of the domestic dog. This finding, however, aligns with the accepted absence of Gαo expression found in the vomeronasal epithelium of this species [42]. The contrast in the expression patterns of Gαo and V2R2 between dogs and the closely related wild canids, like wolves and foxes, offers valuable evidence that complements existing morphological analyses in the VNS of these species [55,56,69,70], which show remarkable differences in the degree of differentiation of both the VNO and the AOB. These features may suggest a regression of the system due to the effects of domestication. This issue has also been postulated in the study of the main olfactory system of the dog through different approaches [71,72,73], therefore, our study may impact the understanding of some of the effects of domestication.
Finally, regarding the discrepancy between the available genomic information on the absence of V2R receptors in the genome of canids and the phenotypic results provided by G proteins, a probable explanation might lie in the absence of exhaustively annotated complete genomes in species such as the fox and the wolf. It is feasible that genomic studies may be overlooking the presence of V2R receptors. For example, in the recent attempt to sequence and assemble the red fox genome, intact V2R genes were not reported [74].
In conclusion, this study reinforces the existence of a segregated model in the development of the VNS in wild canids, with the expression of the two vomeronasal receptor families, V1R and V2R, in species previously thought to exhibit deterioration of the latter. Additionally, it supports the notion that domestication may have produced a dramatic effect on the capacity of detecting pheromonal information.
4. Materials and Methods
In this research, we employed samples from three adult male wolves (Canis lupus signatus), three adult male foxes (Vulpes vulpes), and three adult male mixed-breed dogs (Canis lupus familiaris). The wolves were sourced from wildlife rehabilitation centers located in the Galicia provinces and had succumbed to fatal accidents. We selected only those individuals that had recently passed away and exhibited no signs of external or internal injuries to the head for inclusion in our study. The foxes came from activities organized by the Galician Hunting Federation. They were obtained in the field, the same day of their shooting, with a maximum of two hours delay. The procurement of wolves and fox samples was conducted in compliance with the necessary authorizations from the Galician Environment, Territory, and Housing Department, under the approval codes EB-009/2020 and EB-007/2021. The dogs subjected to necropsy were adult mesocephalic canines, sourced solely from the Department of Clinical Sciences at our institution, where they had died for various clinical conditions. Their heads were intact, displaying neither clinical nor postmortem signs of neurological disorders.
Upon their arrival to the Unit of Anatomy of the Faculty of Veterinary in Lugo, all heads were promptly dissected. The VNOs were exposed by removing the nasal bones and the lateral walls of the nasal cavity. The bone tissue encasing the VNO on the ventral and medial sides was meticulously dissected away from each sample, allowing for processing without the necessity for decalcification. Subsequently, the samples were immersed in Bouin’s fluid (BF) for a 24-hour period, then transferred to 70% ethanol, embedded in paraffin, and sectioned using a microtome. They were systematically sectioned in a transverse plane along their entire length, from the caudal end to the rostral end, at a thickness of 6-7 µm.
Immunohistochemistry methodology
After deparaffinizing and rehydrating the samples, they were processed without performing antigen retrieval. This approach was chosen to avoid unmasking epitopes that could result in non-specific binding. The first step involved treating all samples with a 3% H2O2 solution to inhibit endogenous peroxidase. Subsequently, samples were immersed in a 2.5% horse serum solution, compatible with the ImmPRESS Anti- Rabbit IgG reagent kit (Vector Laboratories, Burlingame, CA, USA) to preclude non-specific binding. Samples were incubated overnight at 4ºC with the primary antibodies.
The next day, samples were incubated for 30 min with the ImmPRESS VR Polymer HRP Anti-Rabbit IgG. Prior to the visualization stage, all samples were rinsed for 10 minutes in 0.2 M Tris-HCl buffer at pH 7.61. DAB chromogen was used for visualizing. A 0.05% 3,3′-diaminobenzidine (DAB) chromogen solution and a 0.003% H2O2 solution, in 0.2 M Tris-HCl buffer were used. The DAB reagent develops into a brown precipitate in the presence of the hydrogen peroxide solution, which enables the visualization of the reaction.
Negative controls omitted the primary antibodies. As positive control we employed sections from tissues known to express the protein of interest.
To carry out the counterstaining procedure, after taking the necessary microphotographs for the immunohistochemical study, the slides were immersed in xylene for 24 hours to facilitate the removal of the cover slips. Once this was accomplished, they were cleared and rehydrated, to stain them with hematoxylin for 5 minutes. Finally, the samples were dehydrated, cleared, and mounted once more, and microphotographs were taken of them.
Primary Antibodies
The antibody against V2R2 was developed by Prof. Tirindelli (University of Parma, Italy). It is a polyclonal antibody raised in rabbit [60]. For the immunohistochemical study of the expression of the G protein alpha subunit Gαo, two commercial polyclonal antibodies were used. The reason for employing two different antibodies was to confirm the immunolabeling obtained with this protein, which in both cases was analogous. The antibodies used were from Santa Cruz Biotechnology (sc-387) and Medical & Biological Laboratories (MBL-551).
Image Capture
Images were digitally captured using a Karl Zeiss Axiocam MRc5 camera coupled with a Zeiss Axiophot microscope. Adobe Photoshop CS4 was employed for brightness, contrast and balance adjustment; however, no enhancements, additions, or relocations of the image features were made. Additionally, an image-stitching software (PTGuiPro) was used for low magnification images composed of several photographs.
Author Contributions
Conceptualization, I.O.L., T.S., R.T., and P.S.Q; Methodology, I.O.L., M.V.T., A.L.B., L.F., and P.S.Q; Investigation, I.O.L., M.V.T., A.L.B., L.F., T.S., R.T. and P.S.Q.; Resources, T.S., R.T., and P.S.Q; Writing – Original Draft Preparation, I.O.L., and P.S.Q; Writing – Review & Editing, I.O.L., M.V.T., T.S., R.T., and P.S.Q; Supervision, P.S.Q.; Project Administration, P.S.Q.; Funding Acquisition, P.S.Q.
Funding
This research was funded by CONSELLO SOCIAL DA UNIVERSIDADE DE SANTIAGO DE COMPOSTELA, grant number 2022-PU004.
Institutional Review Board Statement
Not applicable, as all the animals employed in this studydied by natural causes..
Informed Consent Statement
Not applicable, as this research did not involve any humans.
Data Availability Statement
All relevant data are within the manuscript, and are fully available without restriction.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Woszczyło, M.; Jezierski, T.; Szumny, A.; Niżański, W.; Dzięcioł, M. The Role of Urine in Semiochemical Communication between Females and Males of Domestic Dog (Canis Familiaris) during Estrus. Animals 2020, 10, 2112. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.A.; Kendrick, K.M. Mammalian Social Odours: Attraction and Individual Recognition. Phil. Trans. R. Soc. B 2006, 361, 2061–2078. [Google Scholar] [CrossRef]
- Navarro-Moreno, C.; Barneo-Muñoz, M.; Ibáñez-Gual, M.V.; Lanuza, E.; Agustín-Pavón, C.; Sánchez-Catalán, M.J.; Martínez-García, F. Becoming a Mother Shifts the Activity of the Social and Motivation Brain Networks in Mice. iScience 2022, 25, 104525. [Google Scholar] [CrossRef] [PubMed]
- Meredith, M. Vomeronasal Organ Removal before Sexual Experience Impairs Male Hamster Mating Behavior. Physiol. Behav. 1986, 36, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Maico, L.M.; Burrows, A.M.; Mooney, M.P.; Siegel, M.I.; Bhatnagar, K.P.; Smith, T.D. Size of the Vomeronasal Organ in Wild Microtus with Different Mating Strategies. Acta Biol. Hung. 2003, 54, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Hayes, R.A.; Richardson, B.J.; Wyllie, S.G. Semiochemicals and Social Signaling in the Wild European Rabbit in Australia: I. Scent Profiles of Chin Gland Secretion from the Field. J. Chem. Ecol. 2002, 28, 363–384. [Google Scholar] [CrossRef]
- Burger, B.V. Mammalian Semiochemicals. In The Chemistry of Pheromones and Other Semiochemicals II; Schulz, S., Ed.; Topics in Current Chemistry; Springer Berlin Heidelberg: Berlin, Heidelberg, 2004; ISBN 978-3-540-21308-6. [Google Scholar]
- Salazar, I.; Fdez de Troconiz, P.; Prieto, M.D.; Cifuentes, J.M.; Quinteiro, P.S. Anatomy and Cholinergic Innervation of the Sinus Paranalis in Dogs. Anat. Histol. Embryol. 1996, 25, 49–53. [Google Scholar] [CrossRef]
- Tirindelli, R.; Dibattista, M.; Pifferi, S.; Menini, A. From Pheromones to Behavior. Physiological Reviews 2009, 89, 921–956. [Google Scholar] [CrossRef]
- Fortes-Marco, L.; Lanuza, E.; Martinez-Garcia, F. Of Pheromones and Kairomones: What Receptors Mediate Innate Emotional Responses?: Pheromones and Kairomones. Anat. Rec. 2013, 296, 1346–1363. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, R.M.; Ghirardi, F.; Tirindelli, R. Lacrimal Gland Removal Impairs Sexual Behavior in Mice. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Reep, R.L.; Finlay, B.L.; Darlington, R.B. The Limbic System in Mammalian Brain Evolution. Brain Behav Evol 2007, 70, 57–70. [Google Scholar] [CrossRef]
- Muñiz-de Miguel, S.; Barreiro-Vázquez, J.D.; Sánchez-Quinteiro, P.; Ortiz-Leal, I.; González-Martínez, Á. Behavioural Disorder in a Dog with Congenital Agenesis of the Vomeronasal Organ and the Septum Pellucidum. Vet. Rec. Case Rep. 2023, 11, e571. [Google Scholar] [CrossRef]
- Zilkha, N.; Chuartzman, S.G.; Sofer, Y.; Pen, Y.; Cum, M.; Mayo, A.; Alon, U.; Kimchi, T. Sex-Dependent Control of Pheromones on Social Organization within Groups of Wild House Mice. Current Biology 2023, 33, 1407–1420. [Google Scholar] [CrossRef]
- Del Cerro, M. Role of the Vomeronasal Input in Maternal Behaviour. Psychoneuroendocrinology 1998, 23, 905–926. [Google Scholar] [CrossRef] [PubMed]
- Elwood, R.W.; Stolzenberg, D.S. Flipping the Parental Switch: From Killing to Caring in Male Mammals. Animal Behaviour 2020, 165, 133–142. [Google Scholar] [CrossRef]
- Apfelbach, R.; Blanchard, C.D.; Blanchard, R.J.; Hayes, R.A.; McGregor, I.S. The Effects of Predator Odors in Mammalian Prey Species: A Review of Field and Laboratory Studies. Neurosci. Biobehav. Rev. 2005, 29, 1123–1144. [Google Scholar] [CrossRef] [PubMed]
- Brechbuhl, J.; Moine, F.; Klaey, M.; Nenniger-Tosato, M.; Hurni, N.; Sporkert, F.; Giroud, C.; Broillet, M.-C. Mouse Alarm Pheromone Shares Structural Similarity with Predator Scents. Proceedings of the National Academy of Sciences 2013, 110, 4762–4767. [Google Scholar] [CrossRef]
- Torres, M.V.; Ortiz-Leal, I.; Sanchez-Quinteiro, P. Pheromone Sensing in Mammals: A Review of the Vomeronasal System. Anatomia 2023, 2, 346–413. [Google Scholar] [CrossRef]
- Xie, W.; Chen, M.; Shen, Y.; Liu, Y.; Zhang, H.; Weng, Q. Vomeronasal Receptors Associated with Circulating Estrogen Processing Chemosensory Cues in Semi-Aquatic Mammals. IJMS 2023, 24, 10724. [Google Scholar] [CrossRef] [PubMed]
- Leinders-Zufall, T.; Brennan, P.; Widmayer, P.; S. , P.C.; Maul-Pavicic, A.; Jäger, M.; Li, X.-H.; Breer, H.; Zufall, F.; Boehm, T. MHC Class I Peptides as Chemosensory Signals in the Vomeronasal Organ. Science 2004, 306, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Leinders-Zufall, T.; Ishii, T.; Chamero, P.; Hendrix, P.; Oboti, L.; Schmid, A.; Kircher, S.; Pyrski, M.; Akiyoshi, S.; Khan, M.; et al. A Family of Nonclassical Class I MHC Genes Contributes to Ultrasensitive Chemodetection by Mouse Vomeronasal Sensory Neurons. J. Neurosci. 2014, 34, 5121–5133. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, C.J. Neurobehavioral Evidence for the Involvement of the Vomeronasal System in Mammalian Reproduction. Neurosci. Biobehav. Rev. 1979, 3, 301–341. [Google Scholar] [CrossRef]
- Halpern, M.; Martínez-Marcos, A. Structure and Function of the Vomeronasal System: An Update. Prog. Neurobiol. 2003, 70, 245–318. [Google Scholar] [CrossRef] [PubMed]
- Swaney, W.T.; Keverne, E.B. The Evolution of Pheromonal Communication. Behav. Brain Res. 2009, 200, 239–247. [Google Scholar] [CrossRef]
- Salazar, I.; Barrios, A.W.; Sánchez-Quinteiro, P. Revisiting the Vomeronasal System From an Integrated Perspective. Anat. Rec. 2016, 299, 1488–1491. [Google Scholar] [CrossRef]
- Rodriguez, I.; Greer, C.A.; Mok, M.Y.; Mombaerts, P. A Putative Pheromone Receptor Gene Expressed in Human Olfactory Mucosa. Nat. Genet. 2000, 26, 18–19. [Google Scholar] [CrossRef]
- Trinh, K.; Storm, D.R. Vomeronasal Organ Detects Odorants in Absence of Signaling through Main Olfactory Epithelium. Nat. Neurosci. 2003, 6, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Hagino-Yamagishi, K. Diverse Systems for Pheromone Perception: Multiple Receptor Families in Two Olfactory Systems. Zoological Science 2008, 25, 1179–1189. [Google Scholar] [CrossRef]
- Barrios, A.W.; Núñez, G.; Sanchez Quinteiro, P.; Salazar, I. Anatomy, Histochemistry, and Immunohistochemistry of the Olfactory Subsystems in Mice. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef]
- Ibarra-Soria, X.; Levitin, M.O.; Saraiva, L.R.; Logan, D.W. The Olfactory Transcriptomes of Mice. PLoS Genet. 2014, 10, e1004593. [Google Scholar] [CrossRef]
- Shinohara, H.; Asano, T.; Kato, K. Differential Localization of G-Proteins Gi and Go in the Accessory Olfactory Bulb of the Rat. J. Neurosci. 1992, 12, 1275–1279. [Google Scholar] [CrossRef]
- Halpern, M.; Shapiro, L.S.; Jia, C. Differential Localization of G Proteins in the Opossum Vomeronasal System. Brain Res. 1995, 677, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Halpern, M. Subclasses of Vomeronasal Receptor Neurons: Differential Expression of G Proteins (Giα2 and Goα) and Segregated Projections to the Accessory Olfactory Bulb. Brain Research 1996, 719, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Berghard, A.; Buck, L. Sensory Transduction in Vomeronasal Neurons: Evidence for G Alpha o, G Alpha I2, and Adenylyl Cyclase II as Major Components of a Pheromone Signaling Cascade. J. Neurosci. 1996, 16, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Dulac, C.; Axel, R. A Novel Family of Genes Encoding Putative Pheromone Receptors in Mammals. Cell 1995, 83, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Herrada, G.; Dulac, C. A Novel Family of Putative Pheromone Receptors in Mammals with a Topographically Organized and Sexually Dimorphic Distribution. Cell 1997, 90, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Matsunami, H.; Buck, L.B. A Multigene Family Encoding a Diverse Array of Putative Pheromone Receptors in Mammals. Cell 1997, 90, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Ryba, N.J.P.; Tirindelli, R. A New Multigene Family of Putative Pheromone Receptors. Neuron 1997, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Takigami, S.; Mori, Y.; Ichikawa, M. Projection Pattern of Vomeronasal Neurons to the Accessory Olfactory Bulb in Goats. Chem. Senses 2000, 25, 387–393. [Google Scholar] [CrossRef]
- Takigami, S. Morphological Evidence for Two Types of Mammalian Vomeronasal System. Chem. Senses 2004, 29, 301–310. [Google Scholar] [CrossRef]
- Salazar, I.; Cifuentes, J.M.; Sánchez-Quinteiro, P. Morphological and Immunohistochemical Features of the Vomeronasal System in Dogs. Anat. Rec. 2013, 296, 146–155. [Google Scholar] [CrossRef]
- Salazar, I.; Quinteiro, P.S.; Alemañ, N.; Cifuentes, J.M.; Troconiz, P.F. Diversity of the Vomeronasal System in Mammals: The Singularities of the Sheep Model. Microsc. Res. Tech. 2007, 70, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Suárez, R.; Fernández-Aburto, P.; Manger, P.R.; Mpodozis, J. Deterioration of the Gαo Vomeronasal Pathway in Sexually Dimorphic Mammals. PLoS ONE 2011, 6, e26436. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sánchez-Quinteiro, P. A Detailed Morphological Study of the Vomeronasal Organ and the Accessory Olfactory Bulb of Cats. Microsc. Res. Tech. 2011, 74, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, D.; Watanabe, K.; Nishihara, K.; Ono, Y.S.; Nakamura, K.G.; Yuhara, K.; Tomikawa, S.; Sugimoto, M.; Kobayashi, S.; Horiuchi, N.; et al. Histological Properties of Main and Accessory Olfactory Bulbs in the Common Hippopotamus. Brain Behav. Evol. 2017, 90, 224–231. [Google Scholar] [CrossRef]
- Torres, M.V.; Ortiz-Leal, I.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Neuroanatomical and Immunohistological Study of the Main and Accessory Olfactory Bulbs of the Meerkat (Suricata Suricatta). Animals 2021, 12, 91. [Google Scholar] [CrossRef]
- Kondoh, D.; Kawai, Y.K.; Watanabe, K.; Muranishi, Y. Artiodactyl Livestock Species Have a Uniform Vomeronasal System with a Vomeronasal Type 1 Receptor (V1R) Pathway. Tissue Cell 2022, 77, 101863. [Google Scholar] [CrossRef]
- Schneider, N.Y.; Fletcher, T.P.; Shaw, G.; Renfree, M.B. The Vomeronasal Organ of the Tammar Wallaby. J. Anat. 2008, 213, 93–105. [Google Scholar] [CrossRef]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Does a Third Intermediate Model for the Vomeronasal Processing of Information Exist? Insights from the Macropodid Neuroanatomy. Brain. Struct. Funct. 2022, 227, 881–899. [Google Scholar] [CrossRef]
- Grus, W.E.; Shi, P.; Zhang, Y. -p.; Zhang, J. Dramatic Variation of the Vomeronasal Pheromone Receptor Gene Repertoire among Five Orders of Placental and Marsupial Mammals. Proceedings of the National Academy of Sciences 2005, 102, 5767–5772. [Google Scholar] [CrossRef]
- Young, J.M.; Trask, B.J. V2R Gene Families Degenerated in Primates, Dog and Cow, but Expanded in Opossum. Trends Genet. 2007, 23, 212–215. [Google Scholar] [CrossRef]
- Grus, W.E.; Zhang, J. Distinct Evolutionary Patterns between Chemoreceptors of 2 Vertebrate Olfactory Systems and the Differential Tuning Hypothesis. Molecular Biology and Evolution 2008, 25, 1593–1601. [Google Scholar] [CrossRef]
- Dong, D.; Jin, K.; Wu, X.; Zhong, Y. CRDB: Database of Chemosensory Receptor Gene Families in Vertebrate. PLoS ONE 2012, 7, e31540. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; López-Beceiro, A.; Sanchez-Quinteiro, P. The Vomeronasal Organ of Wild Canids: The Fox ( Vulpes Vulpes ) as a Model. Journal of Anatomy 2020, 237, 890–906. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; Barreiro-Vázquez, J.; López-Beceiro, A.; Fidalgo, L.; Shin, T.; Sanchez-Quinteiro, P. The Vomeronasal System of the Wolf ( Canis Lupus Signatus ): The Singularities of a Wild Canid. Journal of Anatomy 2024, 00, joa–14024. [Google Scholar] [CrossRef]
- Dennis, J.C.; Allgier, J.G.; Desouza, L.S.; Eward, W.C.; Morrison, E.E. Immunohistochemistry of the Canine Vomeronasal Organ. J. Anat. 2003, 202, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Silvotti, L.; Shirazi, A.; Ryba, N.J.P.; Tirindelli, R. Co-Expression of Putative Pheromone Receptors in the Sensory Neurons of the Vomeronasal Organ. J. Neurosci. 2001, 21, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Silvotti, L. The Vomeronasal Receptor V2R2 Does Not Require Escort Molecules for Expression in Heterologous Systems. Chemical Senses 2005, 30, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Silvotti, L.; Moiani, A.; Gatti, R.; Tirindelli, R. Combinatorial Co-expression of Pheromone Receptors, V2Rs. Journal of Neurochemistry 2007, 103, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.; Antunes, A. Vomeronasal Receptors in Vertebrates and the Evolution of Pheromone Detection. Annu. Rev. Anim. Biosci. 2017, 5, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Mier Quesada, Z.; Portillo, W.; Paredes, R.G. Behavioral Evidence of the Functional Interaction between the Main and Accessory Olfactory System Suggests a Large Olfactory System with a High Plastic Capability. Front. Neuroanat. 2023, 17, 1211644. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. The Vomeronasal System of the Newborn Capybara: A Morphological and Immunohistochemical Study. Sci. Rep. 2020, 10, 13304. [Google Scholar] [CrossRef] [PubMed]
- Villamayor, P.R.; Cifuentes, J.M.; Quintela, L.; Barcia, R.; Sanchez-Quinteiro, P. Structural, Morphometric and Immunohistochemical Study of the Rabbit Accessory Olfactory Bulb. Brain Struct. Funct. 2020, 225, 203–226. [Google Scholar] [CrossRef] [PubMed]
- Hohenbrink, P.; Mundy, N.I.; Zimmermann, E.; Radespiel, U. First Evidence for Functional Vomeronasal 2 Receptor Genes in Primates. Biol. Lett. 2013, 9, 20121006. [Google Scholar] [CrossRef]
- Hohenbrink, P.; Dempewolf, S.; Zimmermann, E.; Mundy, N.I.; Radespiel, U. Functional Promiscuity in a Mammalian Chemosensory System: Extensive Expression of Vomeronasal Receptors in the Main Olfactory Epithelium of Mouse Lemurs. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef]
- Zhang, Z.; Nikaido, M. Inactivation of ancV1R as a Predictive Signature for the Loss of Vomeronasal System in Mammals. Genome Biology and Evolution 2020, 12, 766–778. [Google Scholar] [CrossRef]
- Yohe, L.R.; Krell, N.T. An Updated Synthesis of and Outstanding Questions in the Olfactory and Vomeronasal Systems in Bats: Genetics Asks Questions Only Anatomy Can Answer. Anat. Rec. 2529. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; Fidalgo, L.E.; López-Beceiro, A.; Sanchez-Quinteiro, P. Can Domestication Shape Canidae Brain Morphology? The Accessory Olfactory Bulb of the Red Fox as a Case in Point. Ann. Anat. 2022, 240, 151881. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; Vargas-Barroso, V.; Fidalgo, L.E.; López-Beceiro, A.M.; Larriva-Sahd, J.A.; Sánchez-Quinteiro, P. The Olfactory Limbus of the Red Fox (Vulpes Vulpes). New Insights Regarding a Noncanonical Olfactory Bulb Pathway. Front. Neuroanat. 2023, 16, 1097467. [Google Scholar] [CrossRef]
- Jezierski; Ensminger; Papet CANINE OLFACTION SCIENCE AND LAW: Advances in Forensic Science, Medicine; CRC PRESS: S.l., 2016; ISBN 978-0-367-77811-8.
- Bird, D.J.; Jacquemetton, C.; Buelow, S.A.; Evans, A.W.; Van Valkenburgh, B. Domesticating Olfaction: Dog Breeds, Including Scent Hounds, Have Reduced Cribriform Plate Morphology Relative to Wolves. Anat. Rec. 2021, 304, 139–153. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; López-Callejo, L.N.; Fidalgo, L.E.; López-Beceiro, A.; Sanchez-Quinteiro, P. Comparative Neuroanatomical Study of the Main Olfactory Bulb in Domestic and Wild Canids: Dog, Wolf and Red Fox. Animals 2022, 12, 1079. [Google Scholar] [CrossRef]
- Kukekova, A.V.; Johnson, J.L.; Xiang, X.; Feng, S.; Liu, S.; Rando, H.M.; Kharlamova, A.V.; Herbeck, Y.; Serdyukova, N.A.; Xiong, Z.; et al. Red Fox Genome Assembly Identifies Genomic Regions Associated with Tame and Aggressive Behaviours. Nat Ecol Evol 2018, 2, 1479–1491. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Immunohistochemical labelling of the wolf VNO using anti-V2R2 antibody. (A) Labeling is observed throughout the entire strip of the vomeronasal sensory neuroepithelium (SE), particularly in its basal part. Additionally, the apical portion of the epithelium exhibits a more diffuse labeling. (B) Counterstaining with hematoxylin of the neuroepithelium confirms this labeling pattern and allows for the identification of the structural characteristics of the cellular strata and lamina propria (LP). (C) At higher magnification, the labeling is observed to correspond to cells that, because of their spatial placement and morphological characteristics, are identified as neuroreceptor cells (arrowheads). This labeling exhibits a clean and well-defined pattern, predominantly localized to the soma. In addition, a more diffuse pattern of labeling is evident in the apical region of the epithelium. (D-E) Higher magnification of the counterstained areas shown in 1B allows for the determination of the morphological features of the immunopositive structures. It is noticeable that in the immunopositive cells (arrowhead), the labeling is concentrated in the apical portion of the somas. From this portion, dendrites emerge and project towards the apical surface to form the immunopositive dendritic knobs (arrows). Not all receptor cells are immunopositive for V2R2, with cellular somas in different locations being immunonegative (open arrowhead). Finally, no immunopositivity is observed in the various basal cells (asterisk and open arrows). (1) Apical processes; (2) Sustentacular cells layer; (3) Neuroreceptor cells layer. MV, Mucomicrovillar complex; VD, Vomeronasal duct; Vv, Blood vessels. Scale bars: (A): 250 μm; (B-E): 100 μm.
Figure 1.
Immunohistochemical labelling of the wolf VNO using anti-V2R2 antibody. (A) Labeling is observed throughout the entire strip of the vomeronasal sensory neuroepithelium (SE), particularly in its basal part. Additionally, the apical portion of the epithelium exhibits a more diffuse labeling. (B) Counterstaining with hematoxylin of the neuroepithelium confirms this labeling pattern and allows for the identification of the structural characteristics of the cellular strata and lamina propria (LP). (C) At higher magnification, the labeling is observed to correspond to cells that, because of their spatial placement and morphological characteristics, are identified as neuroreceptor cells (arrowheads). This labeling exhibits a clean and well-defined pattern, predominantly localized to the soma. In addition, a more diffuse pattern of labeling is evident in the apical region of the epithelium. (D-E) Higher magnification of the counterstained areas shown in 1B allows for the determination of the morphological features of the immunopositive structures. It is noticeable that in the immunopositive cells (arrowhead), the labeling is concentrated in the apical portion of the somas. From this portion, dendrites emerge and project towards the apical surface to form the immunopositive dendritic knobs (arrows). Not all receptor cells are immunopositive for V2R2, with cellular somas in different locations being immunonegative (open arrowhead). Finally, no immunopositivity is observed in the various basal cells (asterisk and open arrows). (1) Apical processes; (2) Sustentacular cells layer; (3) Neuroreceptor cells layer. MV, Mucomicrovillar complex; VD, Vomeronasal duct; Vv, Blood vessels. Scale bars: (A): 250 μm; (B-E): 100 μm.
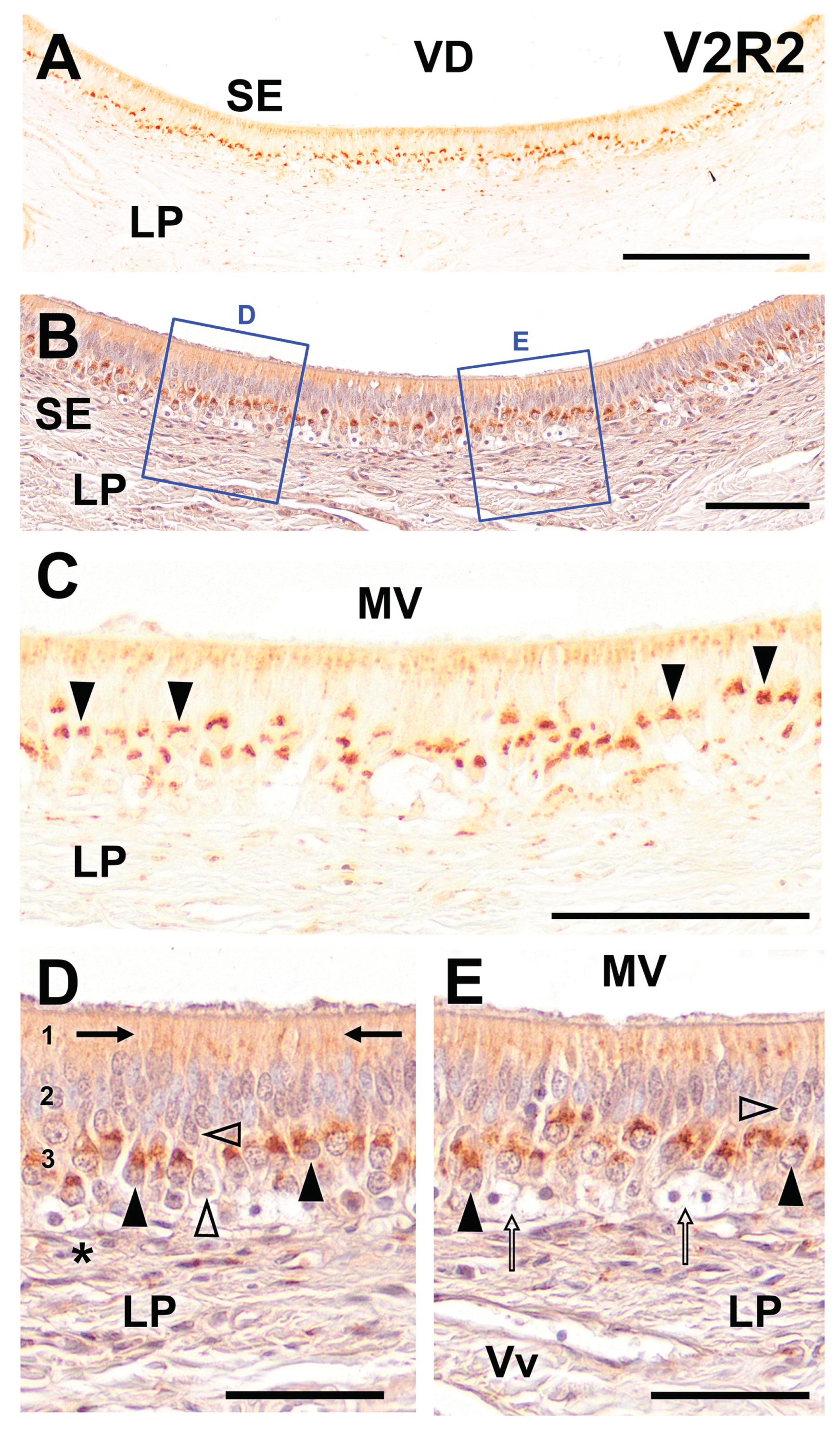
Figure 2.
Immunohistochemical labelling of the fox VNO using anti-V2R2 antibody. (A) Throughout the vomeronasal sensory neuroepithelium (SE), consistent labeling is evident, especially pronounced in the basal area. Additionally, the apical zone shows a diffuse pattern of labeling. (B) Hematoxylin counterstaining of the neuroepithelium facilitates the visualization of the structural features of the cellular layers and lamina propria (LP). (C) Higher magnification corroborates the established labeling pattern and also enables detailed recognition of the cellular strata and the underlying lamina propria. This labeling displays a distinct arrangement, mainly concentrated in the soma. Additionally, the apical part of the epithelium exhibits a more widespread staining pattern. (D-E) Detailed imaging of the counterstained SE, including the section shown in 2B, permits the assessment of the morphological features of the immunopositive structures. The immunopositive cells (arrowhead) have labeling concentrated at the somata apical end. The dendritic ends are as well immunopositive (arrows). Not every vomeronasal neuron is immunopositive for V2R2, with some cell bodies in different locations showing immunonegativity (open arrowhead). The basal cells display no immunopositivity (asterisk). Scale bars: (A): 500 μm; (B-E): 100 μm.
Figure 2.
Immunohistochemical labelling of the fox VNO using anti-V2R2 antibody. (A) Throughout the vomeronasal sensory neuroepithelium (SE), consistent labeling is evident, especially pronounced in the basal area. Additionally, the apical zone shows a diffuse pattern of labeling. (B) Hematoxylin counterstaining of the neuroepithelium facilitates the visualization of the structural features of the cellular layers and lamina propria (LP). (C) Higher magnification corroborates the established labeling pattern and also enables detailed recognition of the cellular strata and the underlying lamina propria. This labeling displays a distinct arrangement, mainly concentrated in the soma. Additionally, the apical part of the epithelium exhibits a more widespread staining pattern. (D-E) Detailed imaging of the counterstained SE, including the section shown in 2B, permits the assessment of the morphological features of the immunopositive structures. The immunopositive cells (arrowhead) have labeling concentrated at the somata apical end. The dendritic ends are as well immunopositive (arrows). Not every vomeronasal neuron is immunopositive for V2R2, with some cell bodies in different locations showing immunonegativity (open arrowhead). The basal cells display no immunopositivity (asterisk). Scale bars: (A): 500 μm; (B-E): 100 μm.
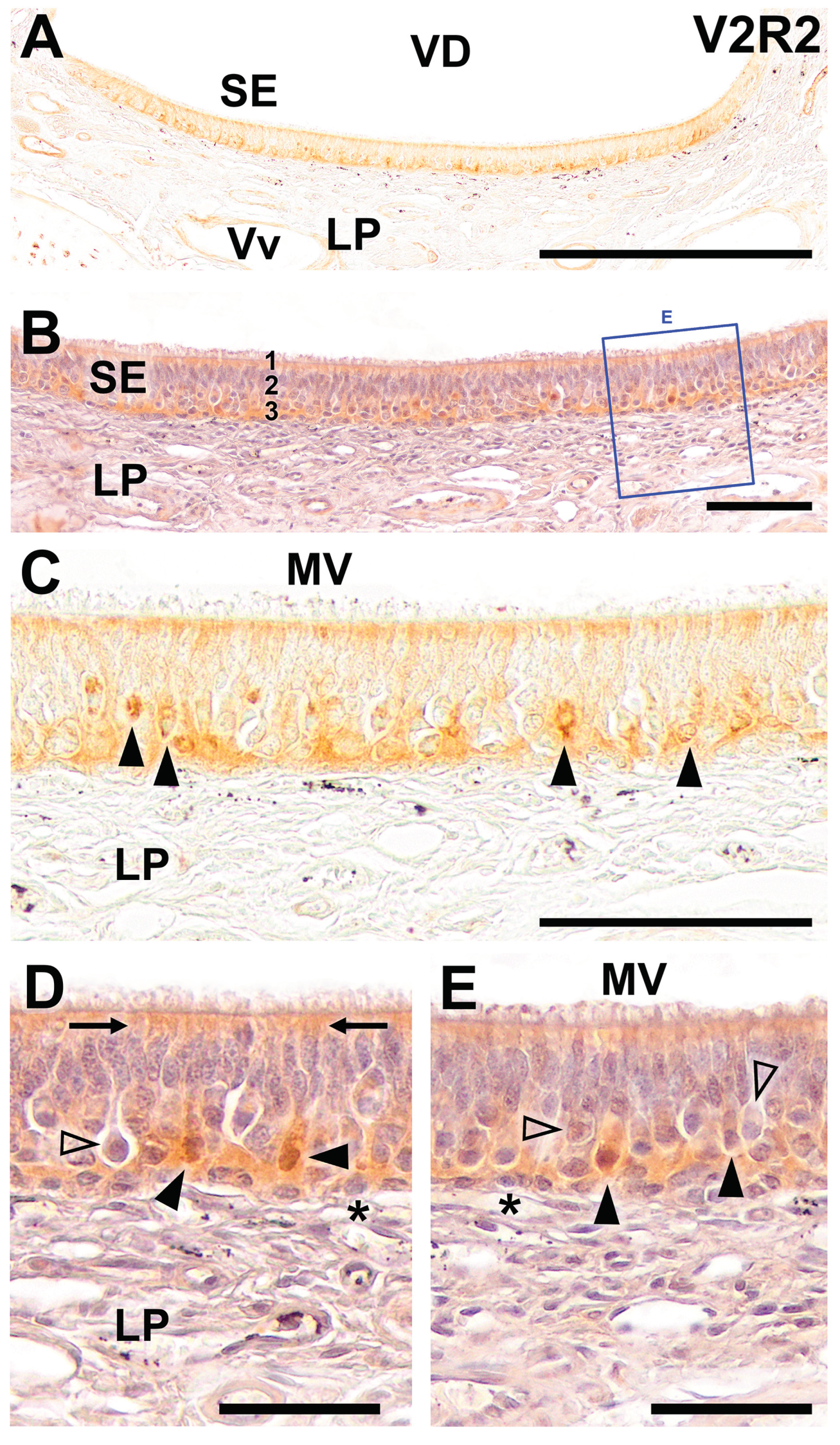
Figure 3.
Immunohistochemical labelling of the dog VNO using anti-V2R2 antibody. (A) The immunohistochemical study of the dog VNO using antibodies against V2R2 did not produce any immunostaining in any of the structural components that constitute it. An immunonegative section is shown. (B) The same section hematoxylin counterstained reveals the differentiation between the sensory epithelium (SE) and the non-sensory or respiratory epithelium (RE). (C) An image of the SE at higher magnifications, in which the neuroepithelium cellular strata, the mucomicrovillar complex, and the underlying lamina propria can be seen, all of them immunonegative. (D-E) High magnification images of the VNO sensory epithelium. Comparison of the wolf anti-V2R2 immunopositive labelling (D), with the dog immunolabelling (E), with produces a complete lack of immunostaining. Scale bars: (A): 500 μm; (B): 250 μm; (C): 100 μm; (D,E): 25 μm.
Figure 3.
Immunohistochemical labelling of the dog VNO using anti-V2R2 antibody. (A) The immunohistochemical study of the dog VNO using antibodies against V2R2 did not produce any immunostaining in any of the structural components that constitute it. An immunonegative section is shown. (B) The same section hematoxylin counterstained reveals the differentiation between the sensory epithelium (SE) and the non-sensory or respiratory epithelium (RE). (C) An image of the SE at higher magnifications, in which the neuroepithelium cellular strata, the mucomicrovillar complex, and the underlying lamina propria can be seen, all of them immunonegative. (D-E) High magnification images of the VNO sensory epithelium. Comparison of the wolf anti-V2R2 immunopositive labelling (D), with the dog immunolabelling (E), with produces a complete lack of immunostaining. Scale bars: (A): 500 μm; (B): 250 μm; (C): 100 μm; (D,E): 25 μm.
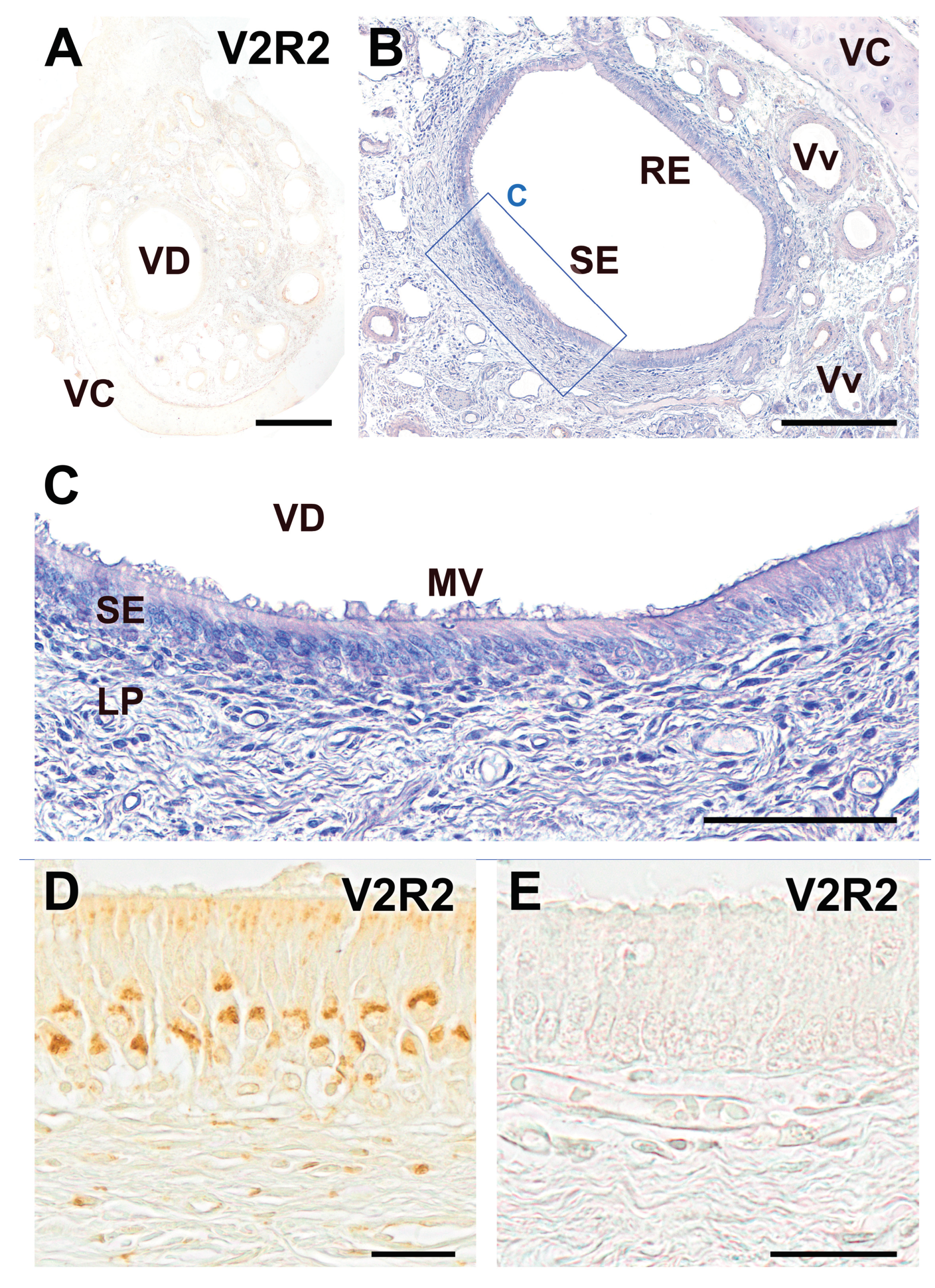
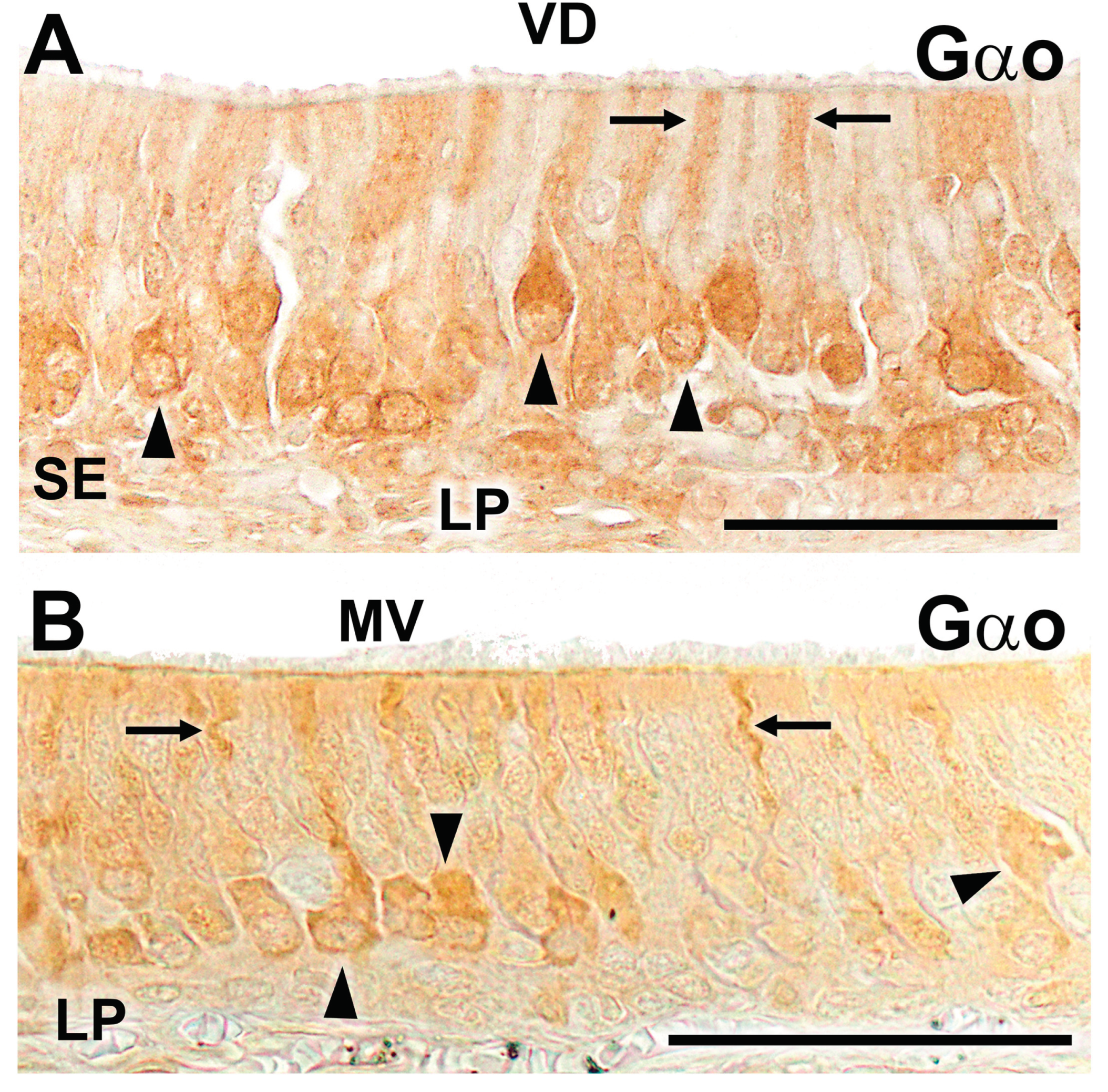
Figure 4.
Immunohistochemical labelling of the wolf and fox VNO using anti-Gαo antibody. (A) Immunohistochemical labeling against Gαo in the sensory epithelium of the wolf VNO results in positive staining in a subpopulation of sensory neurons, which are predominantly located in the basal position. The labeling concentrates on the cellular somata (arrowheads) and the dendritic processes (arrows). (B) The study with the same antibody in the fox VNO produces a similar staining pattern, with immunopositive cellular somata (arrowheads) and dendrites (arrows). Scale bars: (A-B): 50 μm.
Figure 4.
Immunohistochemical labelling of the wolf and fox VNO using anti-Gαo antibody. (A) Immunohistochemical labeling against Gαo in the sensory epithelium of the wolf VNO results in positive staining in a subpopulation of sensory neurons, which are predominantly located in the basal position. The labeling concentrates on the cellular somata (arrowheads) and the dendritic processes (arrows). (B) The study with the same antibody in the fox VNO produces a similar staining pattern, with immunopositive cellular somata (arrowheads) and dendrites (arrows). Scale bars: (A-B): 50 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



