
Submitted:
16 March 2024
Posted:
18 March 2024
You are already at the latest version
Abstract
Excess consumption of a high-fat diet has been identified as one of the contributing factors for obesity. Women are at higher risk for adult obesity than men, predisposing them to a different set of detrimental disease conditions. Furthermore, aging studies show that physiological decline also has a serious impact on changes in the endocrine properties of white adipose tissue. Howev-er, there is still relatively little known about the factors associated with obesity and aging and their compounding impact on women’s health. To investigate adipocytokine secretion profile changes, female C3H/HeJ mice were induced to obesity through long-term consumption of a high-fat diet. Weight gain measurements and Echo MRI analysis of fat composition showed in-creases were a due solely to the high fat content in the diet. Adipocytokine secretions were ana-lyzed in media conditioned from harvested visceral fat tissue that was organ cultured ex-vivo. Adipocytokine analysis across diets and age showed significant increases in Adiponectin and Leptin secretion in high-fat diets, accelerated increases in Resistin secretion with high-fat diets. Aging induced increased secretion of Lipcalin-2, Pentraxin-3 and Serpin E1, MCP-1 and ICAM-1, regardless of diet. Furthermore, comparisons of organoid cultured protein secretions and flash-frozen tissue samples differed greatly, suggesting the WAT organoid cultures may yield information more reflective of in situ conditions. Taken together, our results show that diet and aging significantly impact secretions from adipose tissue, that may contribute to women’s health issues.
Keywords:
Adipocytokines
; Organoid culture
; Ex-vivo white fat tissue culture
; Diet-induced obesity
; Aging
; Adipocyte-secreted factors
; Conditioned medium.
1. Introduction
White adipose tissue (WAT) is no longer considered just an insulating cushion for the body’s internal organs [1]. It is an endocrine organ secreting adipokines and adipocytokines that have paracrine and endocrine effects [2,3]. Their changes in expression, followed with the infiltration of pro-inflammatory immune cells under obese conditions, are responsible for the emergence of local and systemic pathogenic condition such as Type-2 diabetes mellitus, cancer (liver, colon, breast, ovary, prostate), fatty liver, hypertension, Alzheimer disease, depression, and asthma [4,5,6,7,8]. Obesity is reaching a pandemic proportion in the Western Hemisphere and the most common contributing factor behind this consequence is the excessive consumption of a calorie-induced and energy dense diet [9]. Epidemiological evidence suggests a positive correlation of high-fat diet with the development of obesity [10,11]. Due to physiological decline, aging is another critical variable that also has a serious impact on changes in the endocrine properties of white adipose tissue. Age-associated changes such as decline in fat mass and sex hormones, redistribution of lipids from subcutaneous to visceral deposits, decline in brown adipose tissue function, reduction in differentiation and proliferation potential of adipose progenitor stem cells, cellular senescence, or cell stemness and adipogenesis, give rise to multifactorial effects that alter the secretion pattern of WAT [12,13,14]. This ultimately hampers metabolic homeostasis through ectopic lipid deposition, inflammation, disruption in glucose metabolism and insulin sensitivity [12,13,14]. Excess adiposity under these circumstances accelerates the aging process [12]. A few studies have reported the correlation between the changes in adipocytokine secretion with aging and obesity, under different dietary intervention, mostly in male mice [15,16,17,18,19,20]. Furthermore, fewer studies explore these changes in female mice [21]. In the current study, a diet-driven induction of obesity (DIO) approach was used in a long-term study on CH3/HeJ female mice to investigate the effects of a high-fat diet and aging towards changing the expression of endocrine factors secreted from WAT.
To navigate the association between the obesity-mediated changes in WAT and the emergence of various metabolic disorders in organs at the molecular level, different in vitro, in vivo, and clinical study models of WAT are available [3]. However, significant variability between these study models can affect the relevance of experimental outcomes [1,22]. One of the most common strategies to study WAT-associated disease biology at the cellular and molecular level is an in vitro research approach, where adipocyte cell lines are used to mimic the in vivo effect of the WAT [3]. By definition, WAT is heterogeneous, containing preadipocytes, adipocytes, macrophages, stem cells, neutrophils, lymphocytes, endothelial cells, and other cell types [12]. Hence, the adipocyte cell model used for in vitro research differs significantly from the in vivo form of WAT, potentially impacting the interpretation of experimental outcomes [1,22]. Furthermore, according to the CDC, women are reported to have a higher prevalence for adult obesity than men [23] which in turn, is responsible for the emergence of different detrimental disease conditions in women [24,25,26]. To better understand how a high-fat diet and aging change adipocyte-secreted factors of WAT in the in vivo microenvironment, the current study generated conditioned medium derived from visceral fat of female mice cultured ex vivo, using an organ culture technique [1,22]. Organ culture techniques allow for maintaining organotypic cell-cell and cell-matrix interactions of WAT ex vivo [1,22] and more closely recapitulate traits of WAT from the in vivo microenvironment.
There are various strains of mice for which their responses to exposure to high-fat diets have been characterized [27,28]. The most used mouse strain for diet induction of obesity studies is C57BL/6 mice. However, this mouse strain has susceptibility for fat accumulation, weight gain, hyperglycemia and insulin resistance when fed on a high-fat diet [29,30]. To circumvent this predisposition, and to evaluate the impact of a high-fat diet alone in contributing to the changes in adipocytokine secretion in WAT under obese condition in vivo through a DIO approach, the C3H/HeJ mouse strain was used. C3H/HeJ is a general strain of mice used for cancer, immunology, inflammation, and cardiovascular related research studies in biomedical sciences. To date, no study has reported on their susceptibility to obesity, which made the model suitable for investigating the impact of diet-mediated induction of obesity and/or aging on changes in the secretion pattern of adipocytokines. This current study reports the changes in diet and age-induced adipocytokine secretion profiles from visceral fat in female C3H/HeJ mice using a DIO approach. The findings yield important information towards understanding the connection between the high-fat diet-based influence on the aberrant secretions of adipocytokines with the emergence of obesity-associated health disorders and aging in women. Thus, results from this study will help, not only in establishing this mouse strain as a study model for obesity and its associated metabolic disorders, but also yield important information on how adipocyte-secreted factors from WAT change with a high-fat diet and aging in females, leading to new areas of studying their impact on women’s health.
2. Materials and Methods
2.1. Mouse Study Design
The mice used in this specific study were a part of a much larger study under Texas Tech University IACUC protocol 19021-02. For this study specifically, a total of 32 C3H/HeJ female mice, at 4 weeks of age, were utilized. At all times, mouse care and handling were followed as per the protocol. This strain of mice was selected as a robust, generic strain, unmodified for disease susceptibility or resistance. After two weeks of acclimatization, female C3H/HeJ mice were randomized into two customized diets. Diets, prepared by Research Diets, Inc., (New Brunswick, NJ), were categorized based on dietary fat content (control fat: 11% fat kcals, or high-fat (HF): 46% fat kcals %). Total dietary components and formulations are listed in Table A1. Mice were housed in a ventilated cage (4 per cage) with a 12-h light/dark cycle at 22-23 ̊ C and 70% humidity. They also had access to their respective diets and water ad libitum. Their weight and food intake were measured weekly. At the end of the dietary intervention period, (12 months and 18 months for this study), mice were fasted for 2 hr before euthanasia. At collection, 16 females (8 control and 8 HF) from 4 cages (2 control and 2 HF) were euthanized per experimental conditions and visceral fat collected.
2.2. Whole Body Composition Analysis
To analyze the weight gain status based on the body composition of the control and experimental groups of mice, the Echo MRITM Body Composition Analyzer E26-292-BH (EchoMRI Inc., Houston, TX) was used. Echo MRI provides precise measurement of fat-mass, lean tissue, free water and total water composition of a live animal (i.e., mouse) using a nuclear magnetic resonance method [31,32,33,34,35,36,37,38]. The body composition of all mice was analyzed at baseline 0 and monthly until the termination of the experiment. For purposes of this study, only months 12 and 18 were analyzed for adipocytokine secretion changes.
2.3. Tissue Extraction
Mouse visceral fat, anatomically located around the branches of the superior and inferior mesenteric arteries, was used for the study [22,39]. The rationale for choosing visceral fat over other fat deposits is based on its active metabolic potential to produce hormones and cytokines over other fat deposits [16,39,40]. Following euthanasia, mice were first positioned anatomically and pinned in place. Using a scalpel, an incision was made along the midline extending caudally to the pubic symphysis. Forceps were used to grasp the skin, and the scalpel was used to carefully separate the skin from the peritoneum. This allowed access to the visceral white adipose tissue (WAT) deposits, of which a 5 mm3 portion was collected and placed into appropriately labeled 2 ml microcentrifuge tubes. Microcentrifuge tubes containing visceral WAT samples were then flash-frozen in liquid nitrogen and stored at -80°C. Approximately 5 mm3 of the remaining fat was used for the organ culture procedures described in the following section.
2.4. Protein Extraction
The protein extraction process was performed cautiously and under low temperatures (4°C) to prevent protein degradation. The reagents used were stored as per manufacturer’s instructions and kept on ice when used. Moreover, any additional microcentrifuge tubes to be used were stored at -80°C prior to use and immediately placed on ice after being removed from -80°C storage. Similar to the additional microcentrifuge tubes, when visceral WAT samples were removed for protein extraction, they were kept on ice. To begin the protein extraction process, ~ 400 μl of RIPA buffer (Lot# XG348655, Thermo Scientific) was added to a 2 ml microcentrifuge tubes. For every 100 μl of RIPA buffer, 1 μl of Halt Protease Inhibitor Cocktail 100X (Thermo Fisher Scientific, Product# 1862209) was added as outlined in the manufacturer recommendations. The broad spectrum of protease inhibitor cocktail contains AEBSF, aprotinin, E-64, bestatin, leupeptin, and pepstatin A. After thoroughly vortexing the solution, it was then transferred to sterile, pre-cooled 2 ml homogenization tubes containing 1.4 mm ceramic soft tissue homogenizing beads (VWR, Catalog # 10158-610). An average of ~ 50 mg of sample tissue could be added to the tube before inverting to ensure complete coating with the RIPA buffer/ Halt Protease Inhibitor cocktail mixture. Visceral WAT samples were then homogenized using the Bead Mill Homogenizer (Catalog # 75840-022, VWR), that has been preset to 6 m/s for 30 seconds. Samples were removed and immediately placed on the rotating apparatus to allow for continuous agitation for an hour. Following agitation, samples were placed into a mini-spin microcentrifuge (Eppendorf 5452 Minispin Centrifuge) and spun at 13,000 RPM for 15 minutes. Following centrifugation, the sample separates into a clear supernatant with a white lipid layer on the top. A micropipette was used to pierce the lipid layer and remove the protein extract before transferring it to a new, properly labeled 1.5 ml microcentrifuge tube. Protein concentration would then be determined prior to final storage at -80°C to reduce the number of freeze-thaw cycles occurring to preserve protein integrity.
2.5. Protein Quantification
The protein concentration for each sample was then quantified using the Pierce Bicinchoninic Acid (BCA) Protein Assay Kit (Thermo Fisher Scientific). The quantification assay was conducted following the manufacturer’s recommendation. Prior to quantification, a portion of each sample was diluted by a factor of ten with RIPA buffer to ensure it fell within the assay’s detectable limits for optimal accuracy. After quantification, the obtained value could be multiplied by the dilution factor to obtain the protein concentration for each sample.
2.6. Visceral Fat Tissue Harvest for Organ Culture
Visceral WAT deposits were surgically removed from female mice within 30 mins of euthanasia. The collected fat deposit was placed in a 50-cc tube containing a sterile mixture of Medium 199 supplemented with glutamine and 25 nM HEPES (Thermo Fisher Scientific, Catalog# 12340030), 50 μg/mL Gentamycin (Millipore Sigma, Lot# 058M4860V), 1 % Penicillin-Streptomycin (Thermo Fisher Scientific, Catalog # 15140122). Under aseptic conditions, the fat tissue was then coarsely minced using sterile scissors, to avoid hypoxia. Once minced, the sample can remain at room temperature for an hour. The tube was capped and transported to the lab within 30 minutes. Next, a nylon mesh (Mfr # 148147- item # EW-06631-15, Cole Palmer-Scientific Expert) ~240-300 μm, was made into a cone shape and placed on a 500 mL bottle opening. At least 300 mL sterile 1X PBS was poured over the minced fat to drain all the blood and other waste from the minced fat deposit. After cleaning, the fat tissues were recovered from the mesh and put into a petri dish using sterile forceps. The petri dishes were weighed beforehand and adjusted with the weight of fat tissue to ~0.5 gm of fat per dish. Fat tissue weight may be overestimated as it was not dry. Following this, ~15 mL of culture media containing a mixture of M199 media supplemented with 50 μg/mL gentamycin, 1% penicillin-streptomycin, 0.5 μg/mL amphotericin B (Thermo Fisher Scientific, Catalog # 15290018), 0.7 nM of insulin (Sigma-Aldrich; Catalog # I0516), and 250 nM of dexamethasone (Enzo Life Sciences; Catalog# BML-EI126-0001) was added to each dish (Table 1). The media supernatants from every petri dish were collected every 24 hours for one week and replenished with Medium 199 and freshly added hormones. These collected media supernatants are the conditioned media used for determining adipocytokine profiles.
2.7. Adipocytokine Analysis from Organoid Cultures
The adipose secretion profiling of the conditioned media collected from the organ cultured visceral fat was done using a Proteome Profiler Mouse Adipokine Array Kit (R and D System, Catalog# ARY013), following the manufacturer’s protocol. Chemiluminescence was detected with the Odyssey® Fc Imaging System (LI-COR Biosciences, Lincoln, NE), with 8 min exposure time. This profiler kit contained antibodies printed on nitrocellulose membranes against 38 commonly occurring adipocytokines secreted by mouse adipocytes (shown in Figure 1 and listed in Table 2). This approach was used to generate a broad overview of the adipocytokines secreted by mouse adipocytes rather than using an ELISA kit for a particular adipocytokine separately.

Table 2.
List of adipocytokine duplicates printed onto the nitrocellulose membranes.
| Coordinate | Adipokine | Coordinate | Adipokine |
|---|---|---|---|
| A1, A2 | Reference spots | C15, C16 | IL-10 |
| A23, A24 | Reference spots | C17, C18 | IL-11 |
| B1, B2 | Adiponectin | C19, C20 | Leptin |
| B3, B4 | AgRP | C21, C22 | LIF |
| B5, B6 | ANGPT-L1 | C23, C24 | Lipocalin-2 |
| B7, B8 | C-Reactive Protein | D1, D2 | MCP-1 |
| B9, B10 | DPPIV | D3, D4 | M-CSF |
| B11, B12 | Endocan | D5, D6 | Oncostatin M |
| B13, B14 | Fetuin A | D7, D8 | Pentraxin 2 |
| B15, B16 | FGF Acidic | D9, D10 | Pentraxin-3 |
| B17, B18 | FGF-21 | D11, D12 | Pref-1 |
| B19, B20 | HGF | D13, D14 | RAGE |
| B21, B22 | ICAM-1 | D15, D16 | RANTES |
| B23, B24 | IGF-I | D17, D18 | RBP4 |
| C1, C2 | IGF-II | D19, D20 | Resistin |
| C3, C4 | IGFBP-1 | D21, D22 | Serpin E1 |
| C5, C6 | IGFBP-2 | D23, D24 | TIMP-1 |
| C7, C8 | IGFBP-3 | E1, E2 | TNF-α |
| C9, C10 | IGFBP-5 | E3, E4 | VEGF-A |
| C11, C12 | IGFBP-6 | F1, F2 | Reference Spots |
| C13, C14 | IL-6 | F23, F24 | PBS (negative Control) |
2.8. Adipocytokine Protein Levels from Flash-Frozen Tissue Samples
Flash frozen samples of adipose tissue a different to work with than other tissue types due to the concentrations of lipids. Therefore, a modified protein extraction approach was used. Briefly, 0.5 ml RIPA buffer without Triton X-100 was added to ~100 mg adipose tissue in a 2.0-ml tube with protease inhibitor. Halt Protease Inhibitor Cocktail at10 µl/ml was used. The recommended Ripa Lysis Buffer contained 50 mmol/l Tris-HCl (pH 8.0); 0.25 mol/l NaCl; 5 mmol/l EDTA ;1% Triton X-100 (v/v). Tissue was homogenized using the TissueLyser II (Qiagen) at the highest frequency for 3-5 min with the addition of one stainless steel bead into the 2 ml tube. The sample was kept on ice. Once clear, the sample was centrifuged at 6,000 x g for 15 min at 4 °C (9500 RPM). The fat cake (refers to the white lipid layer on top of the aqueous layer) was carefully removed and the loose pellet resuspended. Note: Alternatively, use the pipette tip to penetrate the fat cake and transfer the solution and the pellet to a new 1.5-ml tube. Triton X-100 was added to a final concentration of 1% (v/v), mixed well, and sample kept on ice. The sample was incubated at 4 °C for 30-60 min, then centrifuged at 12,000 x g for 15 min at 4 °C (13500 RPM). The upper lipid layer was removed and the supernatant transferred to a new 1.5-ml tube. The samples were stored at -80 °C until analyzed.
The following ELISA kits were purchased for determining levels of adipocytokines from FF-WAT based on the prior results from the mouse adipokine array. The 6 adipokines/adipocytokines with the strongest signal were assayed using these ELISA kits. For each ELISA, 400 µg of tissue lysate was used to run on the Adipokine membrane. For this study, the manufacturer’s instructions were followed.
Invitrogen Mouse Adiponectin ELISA Kit - Cat# KMP0041
Invitrogen Mouse Leptin ELISA Kit - Cat# KMC2281
Invitrogen MCP-1 ELISA Kit - Cat# BMS6005
Invitrogen Mouse Resistin ELISA Kit - Cat# EMRETN
Invitrogen Mouse VEGF-A ELISA Kit - Cat# BMS619-2
Proteintech Mouse TIMP1 ELISA Kit - Cat# KE10039
2.9. Statistical Analysis
The statistical interpretation of the data was performed using GraphPad Prism 9 software (GraphPad Software Inc., San Diego, CA, USA). For mouse weight changes, Total % Fat (or Lean) Mass columns are the average mass type of all samples/average total mass of all samples * 100. Statistics were analyzed using the Welch’s t test assuming inequal variance between groups (cage effect). Raw statistical data is found in Table A2. Each experiment was conducted independently 3 times with at least 2 technical replicates and the resultant data was expressed as mean ± standard error. The statistical significance between the treatment group and control was determined using Two-way analysis of variance (ANOVA). Tukey’s post hoc test was employed for multiple comparison of means between groups. Overall, a p value <0.05 was considered statistically significant.
3. Results
3.1. Changes in Weight and Body Composition in Control Fat and High-Fat Fed Mice
To measure the changes in weight, fat and lean mass composition of female C3H/HeJ mice with respect to the given diet, their weight and body composition variables were noted by weighing once every week and performing Echo MRI every month, respectively. In Table 2, experimental samples represent the animals that were sacrificed at the given time point (Months 12 and 18). The progressive samples represent the animals that were not sacrificed on Month 12 and had progressed to Month 18. The sample size less than 8 represent the unexpected death of mice in their respective cages, randomly selected for the sacrifice on Month 12 and Month 18.
The changes in the cumulative weight, fat, and lean mass composition of female C3H/HeJ mice, grouped based on control diet and high-fat diet, on Month 12 and Month 18 were compared with that of control, baseline mice at time 0. The mice started the study with an average weight of 17.2 g at time 0. Female C3H/HeJ mice, fed a high-fat diet, were observed to have more than 100 % (~119%) increase in their body weight on Month 12 from the initial weight measured for mice at time 0. In contrast, measurements of changes in the weight of mice fed the control diet, on Month 12 was much less ~41%. The trend in weight gain of both the mice groups on Month 18 is similar to that of Month 12. However, on Month 18, both the control and experimental group of mice showed a reduced percentage of weight gain than they had on Month 12 due to aging.
The changes in the percentage of fat mass status of the control and experimental group of mice were in proportion to their weight gain status. Mice fed on high-fat diet showed a 1000-fold increase in their fat mass from the fat mass found for the mice at time 0. Whereas similar comparative measurements for fat mass of mice fed on control fat diet, is close to 100-fold - which is much less in comparison to high-fat diet fed mice at Month 12. A similar trend of reduction in fat mass occurred with reduction in the percentage of weight gain in the control and experimental group of mice on Month 18. Moreover, the trend in changes of the percentage of lean mass composition for both the control and experimental mice group, in comparison to the initial measure of lean mass composition found for control mice in time 0, paralleled the observed changes in the percentage of weight gain and fat mass composition at Month 12 and Month 18.
Table 2.
Weight gain and body composition details of control and high-fat diet fed female C3H/HeJ mice over time. Note: *Prog = Progressive (mice carried from Month 12 to Month 18). †Exper = Experimental (Euthanized on the given month).
Table 2.
Weight gain and body composition details of control and high-fat diet fed female C3H/HeJ mice over time. Note: *Prog = Progressive (mice carried from Month 12 to Month 18). †Exper = Experimental (Euthanized on the given month).
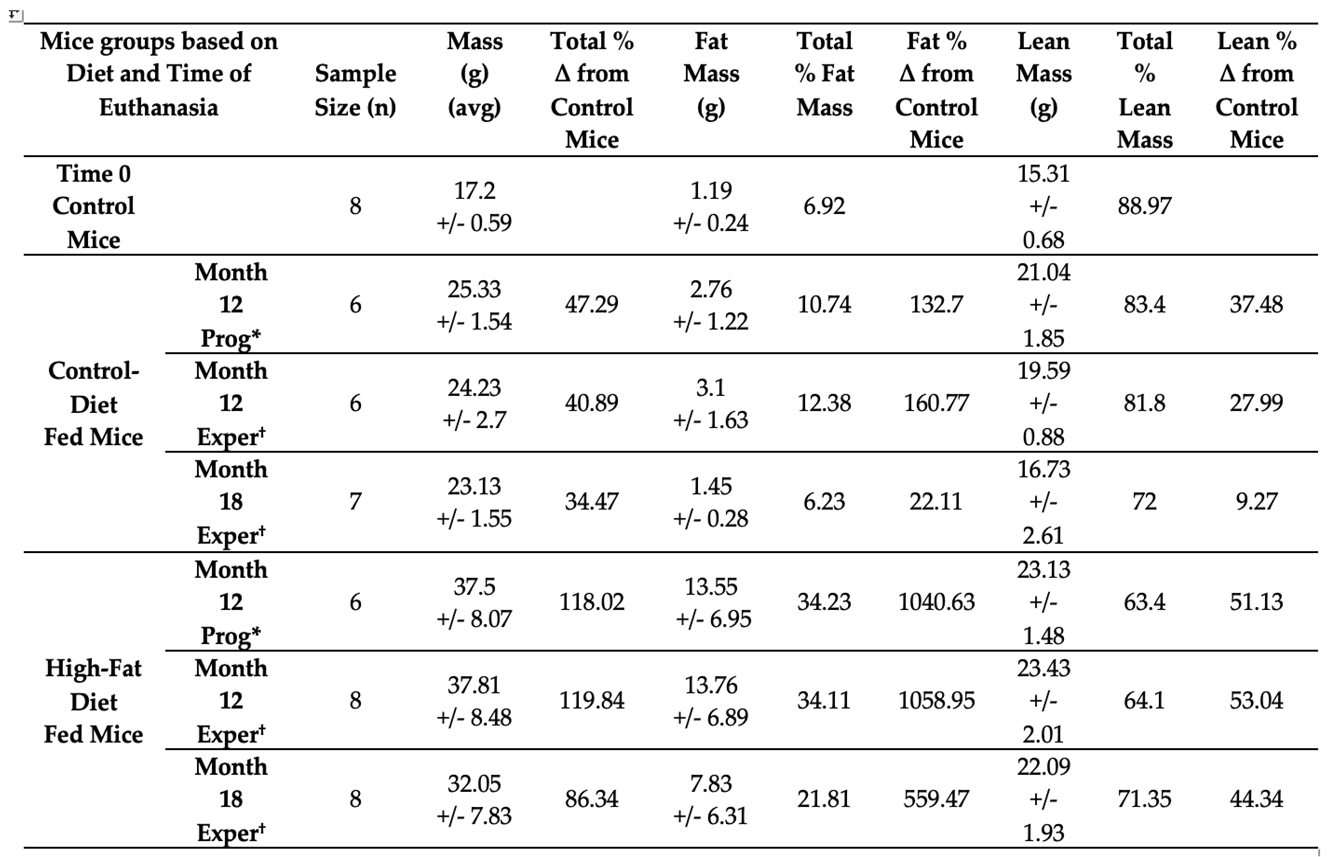
Note: *Prog = Progressive (mice carried from Month 12 to Month 18). †Exper = Experimental (Euthanized on the given month).
3.2. Secretome Profiles of Visceral Fat Harvested from High-Fat and Control Diet Fed C3H/HeJ Mice Differ Significantly with Diet and Aging
For reference, the conditioned medium collected from ex vivo organ cultured visceral fat will be heretofore represented as ExV-WAT-CM in this study. Explanation of the adipocytokine arrangement is described in the Methods section. The presence of different adipocytokines appear as dots. Their various intensities correlate directly to the changes in secretion patterns of the ex vivo cultured visceral fat harvested from control and high-fat diet fed female C3H/HeJ mice at Month 12 and Month 18 in ExV-WAT-CM.
Figure 1 shows the signal output for the adipocytokines from the array kit. In the dot blots, the reference spots represent the intensity of a reference antibody present on the nitrocellulose membrane against which the signal intensity of the secreted ExV-WAT-CM adipocytokines, at separate time points, was measured. The changes in adipocytokine secretions between the conditioned medium have been designated as arbitrary units on the Y-axis of the bar graph as per the instructions from the Proteome Profiler Mouse Adipokine Array Kit (R and D System, Catalog# ARY013). The calculated expression levels of these adipocytokines in Mean ± SEM are presented in Table 2. Figure 2 and Figure 3 show visual comparisons of the data included in Table 3. It is important to note the scale breaks in the y-axes for all the secreted adipocytokine results except Adiponectin and Resistin. The scale breaks were added because without them, there would be no meaningful visualization of the results representative of Table 2.
Table 3.
Adipocytokine secretions from visceral fat harvested from control and high-fat diet fed female C3H/HeJ mice at Month 12 and Month 18.
Table 3.
Adipocytokine secretions from visceral fat harvested from control and high-fat diet fed female C3H/HeJ mice at Month 12 and Month 18.
|
Adipocytokines |
Control Fat (Month 12) (Mean±SEM) |
High-Fat (Month 12) (Mean±SEM) |
Control Fat (Month 18) (Mean±SEM) |
High-Fat (Month 18) (Mean±SEM) |
|---|---|---|---|---|
| Ref Spot 1 | 0.039 ± 0.0023 | 0.043 ± 0.0021 | 0.056 ± 0.0004 | 0.050 ± 0.0020 |
| Ref Spot 2 | 0.043 ± 0.0023 | 0.042 ± 0.0016 | 0.049 ± 0.0004 | 0.047 ± 0.0019 |
| Adiponectin | 0.033 ± 0.0020 | 0.046 ± 0.0010 | 0.060 ± 0.0003 | 0.057 ± 0.0023 |
| ICAM-1 | 0.004 ± 0.0002 | 0.003 ± 0.0002 | 0.009 ± 0.0001 | 0.011 ± 0.0008 |
| IL-6 | 0 | 0 | 0.003 ± 6.36E-05 | 0.002 ± 0.0002 |
| Leptin | 0.005 ± 0.0002 | 0.003 ± 0.0002 | 0.002 ±7.10E-05 | 0.008 ± 0.0004 |
| Lipocalin-2 | 0.018 ± 0.0008 | 0.015 ± 0.0004 | 0.020 ± 0.0003 | 0.025 ± 0.0013 |
| MCP-1 | 0.009 ± 0.0008 | 0.011 ± 0.0008 | 0.019 ± 0.0001 | 0.028 ±0.0026 |
| MCS-F | 0.002 ± 0.0002 | 0.000 ± 9.95E-05 | 0.001 ± 4.11E-05 | 0.002 ± 0.0003 |
| Pentraxin-3 | 0.005 ± 0.0002 | 0.006 ± 0.0005 | 0.014 ± 0.0002 | 0.020 ± 0.0015 |
| RBP4 | 0.003 ± 0.0002 | 0.003 ± 0.0001 | 0.004 ± 0.0002 | 0.005 ± 0.0003 |
| Resistin | 0.032 ± 0.0015 | 0.050 ± 0.0015 | 0.051 ± 0.0005 | 0.048 ± 0.0025 |
| Serpin E1 | 0.015 ± 0.0007 | 0.018 ± 0.0004 | 0.024 ± 0.0003 | 0.023 ± 0.0011 |
| TIMP-1 | 0.017 ± 0.0008 | 0.009 ± 0.0008 | 0.003 ± 0.0003 | 0.009 ± 0.0014 |
| VEGF | 0.011 ± 0.0008 | 0.005 ± 0.0004 | 0.002 ± 3.86E-05 | 0.006 ± 0.0004 |
| Ref Spot 3 | 0.040 ± 0.0023 | 0.044 ± 0.0010 | 0.050 ± 0.0003 | 0.045 ± 0.0021 |
As shown in Figure 2 and Figure 3A and Table 3, Adiponectin analyzed ExV-WAT-CM, exhibited significant increases in secretion in both control and high-fat diet fed mice as a result of age. Furthermore, there were significant increases in Adiponectin secretion between control and HF diet fed mice at 12 months. However, this difference was not observed at 18 months. Interestingly, the difference in Adiponectin secretion at 12 months in the HF diet fed mice was comparable to the secretion levels in the control mice measured at 18 months (not significant). It is also important to note that of the ExV-WAT-CM samples measured, Adiponectin was secreted in the highest concentrations along with Resistin.
Results for Leptin showed mice fed a high-fat diet had significantly increased Leptin secretion with aging, whereas the control diet fed mice trended downward. Moreover, the WAT secretion levels of Leptin also significantly increased in mice with the increase of fat mass with aging (Figure 2 and Figure 3B and Table 3). Even with an increase in the level of Leptin, mice on the high-fat diet were obese, failing to respond to reduced feeding signals from Leptin, which is most likely due to the development of Leptin resistance or insufficient Leptin sensitivity in the brain receptors [41].
Adipocytes also secrete chemokines. Monocyte chemoattractant protein-1 (MCP-1) is a chemokine that attracts blood monocytes to the site of inflammation [42]. The elevation in the level of MCP-1 is thought to be correlated with age-related induction of inflammation (inflammaging) [43,44]. The data generated from the ExV-WAT-CM samples shows that HF diet fed mice exhibit significantly increased secretions of MCP-1 with aging, irrespective of diet (Figure 2 and Figure 3C and Table 3). Furthermore, while there was an upward trend in control diet fed mice, this difference was not significant with aging, suggesting that high-fat diet alone promotes inflammatory events in aging mice.
Another important secretory protein from adipocytes (and other cells) is Resistin. It is a hormone which impairs the action of insulin and induces glucose intolerance, leading to the development of Type 2 Diabetes Mellitus [45]. Figure 2 and Figure 3D and Table 3, showed a significant increase in Resistin secreted in ExV-WAT-CM samples in the mice fed a HF diet at Month 12 when compared to the control diet. Interestingly, a significant increase in the expression of Resistin was also observed in control diet fed mice between Month 12 to Month 18, while the difference in secreted concentrations of Resistin in the HF diet fed mice did not change much with aging. It is also important to note that of the ExV-WAT-CM samples measured, Resistin was secreted in the highest concentrations along with Adiponectin, as indicated by the y-axis scale break.
TIMP-1, a member of the family of tissue inhibitors of metalloproteinases, physiologically plays a role in the inhibition of matrix metalloproteinase, which in turn, causes degradation of the extracellular matrix (ECM) and tissue remodeling. Analysis of TIMP-1 in our study showed significantly decreased expression of TIMP-1 with aging in control diet fed ExV-WAT-CM mouse samples (Figure 2 and Figure 3E and Table 3). In contrast, the group fed a HF diet did not demonstrate significant changes in the expression of TIMP-1 with aging. Moreover, the expression level of TIMP-1 was decreased in mice fed a HF diet at Month 12. It is of note that the measurements from the 18-month samples varied greatly in their distribution between mice.
Vascular endothelial growth factor (VEGF) is a protein secreted by adipocytes and other cells that drives angiogenesis. Its overexpression in tumors is associated with increased angiogenesis, proliferation, and metastasis [46,47,48]. Similar to our TIMP-1 results, the ExV-WAT-CM analyzed secretions demonstrated significant decreases in the presence of VEGF for control diet fed mice between 12 and 18 months (with aging) (Figure 2 and Figure 3F and Table 3). Furthermore, the levels of VEGF between 12 and 18 months was basically unchanged, and while there was significantly less VEGF present in these samples versus the 12-month controls, the difference was not significant compared to 18-month controls samples.
ICAM-1 is a cell surface glycoprotein receptor, expressed by immune, endothelial and epithelial cells [32]. Its expression becomes upregulated in response to inflammatory stimulation, regulating the recruitment of leukocytes to the sites of inflammation [32]. Our results (Figure 2 and Figure 4 and Table 3). showed that, irrespective of diet, ICAM-1 secretion in ExV-WAT-CM was significantly increased in mice with aging, while there was no significant difference between diets at each respective timepoint. These results suggest ICAM-1′s effects were more affected by aging than diet (in this case HF versus control).
Lipocalin 2 is an adipocytokine produced from WAT that is responsible for transporting hydrophobic molecules like steroid hormones, fatty acids, etc., to their target site of action [49]. Results from the current study showed an increase in the secretion of Lipocalin-2 from ExV-WAT-CM in mice with aging, irrespective of the fat content in the diet (Figure 2 and Figure 4 and Table 3).
Adipocytes also secrete other proteins that directly or indirectly play a role in inflammation. IL-6 is a pro-inflammatory cytokine known to be secreted from adipocytes [50,51,52]. IL-6 expression in the ExV-WAT-CM samples at 12 months was undetectable regardless of diet (Figure 2 and Figure 4 and Table 3). With aging, female mice expressed IL-6 at Month 18 and both control and HF diet samples were significantly greater than 12 months. Surprisingly however, at 18 months, the control diet samples showed significantly greater IL-6 secretion than the HF diet fed mice.
Pentraxin-3 is an adipocytokine secreted under inflammatory conditions and reported to promote metastasis [53,54]. In our study, Pentraxin-3 was observed to be significantly upregulated with aging, irrespective of diet (Figure 2 and Figure 4 and Table 3). Furthermore, there were no significant differences in Pentraxin-3 secretion between diet groups at either time point.
Serpin E1 is a plasminogen activator inhibitor that functions to stabilize fibrinogen clot formation wound healing [55]. As shown in Figure 2 and Figure 4 and Table 3, Serpin E1 secretions from ExV-WAT-CM increased significantly with aging for the control diet mice. However, secretion profiles for HF diet fed mice, while trending upwards with aging, were not significant. Like the Pentraxin-3 results, the Serpin E1 results were not significant between groups at either timepoint.
M-CSF is a growth factor cytokine, mediating local survival, proliferation and differentiation signaling to mononuclear phagocytic cells at the site of inflammation [31]. We observed that there were no differences in secretion of M-CSF across time or diet. There was a trend towards increased expression of M-CSF when mice were fed a high-fat diet (Figure 2 and Figure 4 and Table 3).
3.3. Adipocytokine Profiles of Flash-Frozen Visceral Fat Tissue Harvested from High-Fat and Control Diet Fed C3H/HeJ Mice Differ Significantly from Corresponding Organoid Cultured Samples
One of the major goals of this study was to determine whether there is a difference from flash frozen WAT sample secretions versus fresh samples. This is important because it reveals whether frozen WAT tissue accurately represents changes in adipocytokine secretion compared to freshly harvested samples. Inconsistencies could have repercussions to data interpretation for in vitro studies. To distinguish between the results, and for ease of identification, the flash frozen samples are heretofore represented as FF-WAT to be compared to the ex vivo samples represented as ExV-WAT-CM.
Figure 5 is a graphical representation of the results analyzed by ELISA for the levels of Adiponectin, Leptin, Resistin, MCP-1, TIMP-1 and VEGF from FF tissue samples. These are also the 6 secreted adipocytokines analyzed from medium obtained from cultured organoids in Figure 3. It is important to note the y-axis scale breaks when comparing these results.
Interestingly, Adiponectin analyzed from FF-WAT reflected similar increases in protein levels as ExV-WAT-CM (Figure 3), for both control and high-fat diet fed mice as a result of age. However, the protein levels between groups at each time point was very similar. Interestingly, the levels of Adiponectin were markedly higher in the FF-WAT samples. It is also important to note that of the FF-WAT samples measured, Adiponectin exhibited the highest concentrations (Figure 5).
Results for Leptin showed mice fed a high-fat diet had significantly decreased Leptin secretion with aging. Moreover, the WAT secretion levels of Leptin also trended upward in control diet fed mice (Figure 5). Also important to note was the substantial increase in Leptin concentrations in FF-WAT compared to the ExV-WAT-CM. Although statistical comparisons were not performed, the amount of measurable Leptin was 5x greater in the FF-WAT samples. These results are contrary to the adipokine secretion results from ExV-WAT-CM (Figure 3), in which we observed an increase in Leptin in high-fat diet fed mice. Even with an increase in the level of Leptin, mice on the high-fat diet were obese, failing to respond to reduced feeding signals from Leptin, which is most likely due to the development of Leptin resistance or insufficient Leptin sensitivity in the brain receptors [41].
Monocyte chemoattractant protein-1 (MCP-1) is a chemokine that attracts blood monocytes to the site of inflammation [42]. The elevation in the level of MCP-1 is thought to be correlated with age-related induction of inflammation (inflammaging) [43,44]. Results for MCP-1 from FF-WAT showed mice fed a high-fat diet had significantly decreased concentrations with aging. Moreover, the WAT secretion levels of MCP-1 in control diet fed mice were not significantly different (Figure 5). Although statistical comparisons were not performed, the amount of measurable MCP-1 was ~2-3x greater in the ExV-WAT-CM samples (note the scale break in Figure 2 y-axis). These results are contrary to the secretion results from (Figure 5), in which we observed a decrease in MCP-1 in high-fat diet fed mice.
Resistin levels in the FF-WAT samples basically remained unchanged between diet and aging (Figure 5). Additionally, there were no significant changes in Resistin expression between diets for each time point. Furthermore, the FF-WAT samples of Resistin were ~5X lower than the levels observed in ExV-WAT-CM.
Analysis of TIMP-1 expression from FF-WAT (Figure 5), showed significant increase in expression in control diets with aging, but no significant changes in the high-fat diet fed mice. Furthermore, there was no significant difference between diet groups when compared at each time point. This contrasted with results for ExV-WAT-CM samples, where the control diet fed group were observed to decrease in expression significantly with aging. However, the expression level of TIMP-1 was comparable between ExV and FF samples in the high-fat diet fed mice, with no significant changes and approximately the same signal intensity measured over time.
Vascular endothelial growth factor (VEGF) results for FF-WAT expression by ELISA was not significant for either diet or aging at any level (Figure 5).
4. Discussion
C3H/HeJ is a generic strain of laboratory mice without any documented susceptibility for obesity or type 2 diabetes. Thus, choosing this mouse model for evaluating the changes in adipocytokine expression in the metabolically active visceral fat, based on the variability of dietary fat consumption for a long-term aging study, yielded important information about how a high-fat diet for a long term impacts physiological homeostasis without any existing predisposition. In this study, the visceral fat was organ cultured ex vivo, and the conditioned media was collected to determine the expression of adipocytokines. This approach allowed us to determine how the diet impacted the physiological condition of visceral adipose tissue in vivo and how that impact resulted in significant changes in the expression of adipocytokines from that visceral fat cultured ex vivo. As expected, the data presented on this study demonstrated that excessive consumption of a high-fat diet contributed to the weight gain in the mice. This was further supported by fat mass composition data, measured through Echo MRI. It showed that the 1000-fold increase of fat deposition in high-fat diet fed mice at 12 months. These data, in comparison to data from the control diet fed mice, provides further evidence that the increase in the weight and fat mass composition in the C3H/HeJ mouse model is solely due to the high-fat content included in the diet.
Data shown in Figure 2, Figure 3 and Figure 4 and Table 3 confirmed our hypothesis that significant changes exist in the expression of adipocytokines in ExV-WAT-CM, secreted from visceral fat harvested from high-fat and control fat diet fed female C3H/HeJ mice at Month 12 and Month 18. As there are very few studies reporting adipocytokine secretions from fat deposits in female mice or C3H/HeJ mice, most of the insights for the discussion of the current study have been drawn from previously reported observations for the corresponding adipocytokines secreted from other mouse strains.
One of the critical adipocytokines secreted by WAT is Adiponectin. Not only it is the most abundant adipocytokine secreted from the adipose tissue [56], but it also plays a protective role in minimizing the effects of inflammation, maintaining whole-body energy homeostasis by expediting lipid and carbohydrate metabolism, increasing insulin sensitivity for the glucose transport system, as well as inhibiting hepatic gluconeogenesis and lipogenesis [57].
In Figure 2, results suggest that the secretion of Adiponectin increased at the mid to advanced stage of life in female C3H/HeJ mice when they are on a high-fat diet. This result contradicts epidemiological and experimental evidence in obesity studies reporting that decreased expression of Adiponectin is correlated with the increase in fat mass [58,59,60,61,62]. One possible explanation for this discrepancy may be associated with the total intake of calories and not the changes in fat mass as a function of dietary fat or source of calories intake [63,64,65]. Furthermore, the total caloric intake may play a critical role at molecular level to control the expression of Adiponectin as suggested by in vivo studies using C57BL/6 mice [66,67]. Qiao et al., reported that 2 groups of C57BL/6 mice, receiving the same number of calories through the high-fat and low-fat diet regimens, did not exhibit any changes in WAT-based Adiponectin gene expression or blood Adiponectin levels, even though high-fat diet fed mice gained increased adiposity [67]. Subsequently, different dietary feeding regimens and treatments (low-fat control diet, 30% calorie restriction regimen, a treadmill exercise regimen with a low-fat control diet and continuation of high-fat diet) on the high-fat diet fed C57BL/6 mice, showed that only calorie restriction in the high-fat fed C57BL/6 mice increased the expression Adiponectin [67]. According to the dietary formulations for the C3H/HeJ mice (Appendix 1), both groups of mice (control and high-fat), were given similar calorie containing diets. Taken together, these reports coupled with our findings, suggest that increase in fat mass, with high-fat dietary intake in C3H/HeJ mice, does not significantly contribute to the changes in Adiponectin expression. Rather, the intake of total calories given similar calorie diets containing high-fat and control fat content, could be the reasons for the observed changes in Adiponectin expression with this current study. Moreover, most of the study findings discussed above, are based primarily on findings interpreted from male mice. Thus, when differences in physiology are considered, the sex of the mice could be a reason for the discrepancy in the Adiponectin secretion reported in this study from those of previous studies.
Leptin is another adipocytokine secreted profusely in ExV-WAT-CM [68]. Under normal physiological conditions, as a hormone, it regulates appetite and energy expenditure and helps to maintain normal weight [69]. However, the circulatory levels have been reported to increase in obese individuals with increased fat mass [41,68]. Even with increased plasma levels of Leptin, mice on high-fat diets keep progressing to obesity due to Leptin resistance [41]. Our results from the ExV-WAT-CM samples are in line with these findings. In our study, C3H/Hej female mice fed a high-fat diet were observed to have significantly increased Leptin secretion to the control fat diet with aging. These results are in line with studies demonstrating that Leptin signaling decreases with obesity [41].
The changes in Resistin secretion levels suggest that high-fat diet consumption increases Resistin expression sooner in the mouse lifespan, as increased expression in the control diet fed C3H/HeJ mice was observed at Month 18th. This suggests that continuous consumption of high-fat diet promotes the occurrence of age-associated metabolic comorbidities earlier in life by increasing the secretion of Resistin from visceral fat.
TIMP-1, a member of the family of tissue inhibitors of metalloproteinases, physiologically plays a role in the inhibition of matrix metalloproteinase, which in turn, causes degradation of the extracellular matrix (ECM) and tissue remodeling [70]. A number of clinical and in vivo studies reported in male mice have found increased circulatory and fat tissue-based expression of TIMP-1 under obese conditions [71,72]. Studies have proposed the role of TIMP-1 in adipogenesis through TIMP1-mediated extracellular matrix remodeling and adipocyte expansion [72,73,74]. Reduced expression of TIMP-1 with aging implies less space available for adipose tissue to expand, which can lead to “mechanical stuffing” of expanded adipose tissue with continuous high-fat diet consumption, followed by hypoxia and associated inflammatory condition. In contrast, an in vivo study in a TIMP-1 deficient female mice model reported a pronounced increased in body weight compared to wild type mice, with increased weight and size of white adipose tissue attributed to hyperphagia [75]. Taking these results into consideration, it is possible that there is a link between decreased expression of TIMP-1 in ExV-WAT-CM samples of female mice fed on high-fat diet and hyperphagia, explaining their increased weight gain and adipose tissue mass. More study observations are needed to address this possibility.
Based on the observations for TIMP-1 and VEGF, the data suggests that the secretion for both adipocytokines decreased with aging with control fat diet consumption. However, with consumption of the high-fat diet, their secretion remained stable with aging. The decreased expression of VEGF in aged control diet fed as well as young and aged high-fat fed female C3H/HeJ female mice in this current study suggests a possible impairment in angiogenic capacity in adipose tissue progressing to hypoxia and dysfunction by limiting the removal of fatty acids from the extracellular spaces, exhibited through enhanced fat mass as well as inflammatory events in adipose tissue (shown later) [76].
Lipocalin results parallel reports from previous studies demonstrating a link between the elevated expression of Lipocalin 2 with obesity and glucose intolerance [77,78,79,80]. However, these findings are controversial as other studies have demonstrated a protective effect of Lipocalin-2 against metabolic deterioration with aging [81,82]. Clarifying these studies and elucidating the reason needs further research.
Observations for the adipocytokines Pentraxin-3 and Serpin-1 parallel those reported in previous studies correlating their increased expression with obesity and age-related disease [83,84,85,86]. Pentraxin-3, an inflammatory biomarker, secreted under inflammatory conditions, was found to promote cell stemness [87], and metastasis [53,54]. Serpin E1, a plasminogen activator inhibitor encoded by the SERPIN E1 gene, primarily plays a role in the stabilization of fibrinogen clot formation and maintenance of wound healing [55]. However, studies have also reported Serpin E1 signaling to have a role in malignant progression and resistance [88,89,90,91], hepatic steatosis [92] and aging [93]. These same results were observed for ICAM and IL-6. Considering the influence of the above mentioned adipocytokines in pathogenic and metabolic disorder, it is possible that increased secretion of Pentraxin-3, Serpin E1 ICAM and IL-6 from ExV-WAT-CM samples of female mice fed on high-fat diet are involved in age-associated disease pathogenesis in this female C3H/HeJ mice model. This study’s findings concur with the current paradigm that age-associated pathogenesis is the direct outcome of inflammatory changes during aging [33,34,35].
The differences in flash frozen WAT and proteins secreted from organoid cultures into the medium are important to consider. The data for ExV-WAT secreted proteins compared to the FF-WAT samples have shown that there is a difference in the levels of adipocytokines from the same visceral fat sample processed differently. FF-WAT is a sample of the whole, heterogeneous visceral fat tissue that has been flash frozen before protein extraction. After protein extraction and quantification, the changes in the expression of embedded adipocytokines were evaluated. However, it is important to distinguish that apart from the cellular proteins analyzed by the Array Kit, adipocytokines are secretory proteins which are released systematically in vivo. When the visceral fat was organ cultured ex vivo, the conditioned media approach was used to evaluate the expression of those same adipocytokines. This approach allows us to determine how the diet impacted the visceral adipose tissue in vivo and how that impact resulted in significant changes in the expression of adipocytokines measured through this ex vivo process. Of the 3 signaling factors primarily secreted by WAT, a strong discrepancy was observed between Leptin and Resistin. Results for Adiponectin and Leptin were greatest in FF-WAT unlike ExV-WAT where Adiponectin and Resistin were highest. This difference in key adipocytokines between processing methods needs further study.
This is the first report of the secretion patterns of adipocytokines from visceral fat in C3H/HeJ mice after exposure to high-fat diet. The impact of high-fat diet and aging on the adipocytokine secretions from visceral fat has been summarized in Table 3. Collectively, with the current study, it appears that inflammatory adipocytokine secretions from ExV-WAT-CM samples of visceral fat in female C3H/HeJ mice, might promote their age-associated pathogenesis.
Table 3.
Summary of the Impact of Aging and High-Fat Diet on the Selected Adipocytokine Secretions.
| Aging | Adiponectin, Leptin, MCP-1, TIMP-1, VEGF, IL-6,ICAM-1, Lipocalin-2, Serpin E1, M-CSF |
| High Fat Diet | Adiponectin, Leptin, Resistin, TIMP-1, VEGF |
One limitation of the study is the possibility for C3H/HeJ mice to develop a spontaneous mutation in the lipopolysaccharide response locus. This was later found to be detected as a toll-like receptor-4 gene (Tlr4-gene) [94,95,96] resulting in a defective TLR-4, making those mice with the spontaneous mutation susceptible to endotoxins [94,95,96]. The relevance of TLR-4 in the current study may have implications for the obesity-induced inflammatory response [97,98,99]. Moreover, TLR4 is also involved in the crosstalk between adipocytes and immune cells, leading to an induction of proinflammatory environment in white adipose tissue [100,101]. However, the influence of TLR4 loss of function is controversial and other studies have shown that C3H/HeJ mice with this mutation are protected against the development of diet-induced obesity [102]. Future studies could address the frequency and status of this mutation to better understand the molecular signaling between adipocytes and immune cells in this model. While not considered a limitation, it should also be noted that, as this study was specific to female C3H/HeJ mice, results in males may not be comparable.
5. Conclusions
Because mouse physiology can vary greatly from one strain to another, it is difficult to determine which factor has the stronger impact on the altered secretions of adipocytokines from visceral fat – high-fat diet, or aging. However, with the current study findings, it can be surmised that aging plays a major role in the altered secretion of adipocytokines from visceral fat tissue from these female mice. The study findings discussed above successfully demonstrated that a high-fat-based diet alone changes the secretion pattern of adipocytokines from visceral fat adipose tissue and suggests that continuous high-fat diet consumption for long term (in this case the lifespan of the mice), increases the likelihood of age-related disease and morbidities occurring earlier. More research focusing on obesity, aging and women’s health is needed. The insights gained from the current study will be helpful in considering the C3H/HeJ mouse strain for the future in vivo obesity-mediated metabolic disorder studies. It is our hope that the findings from this study will add important information to the growing field of research into women’s health issues.
Author Contributions
Conceptualization, N.M. and L.G.; methodology, N.M., B.B. and C.B.; formal analysis, N.M., C.B., and B.B.; investigation, N.M.; resources, L.G.; data curation, N.M., L.G.; writing—original draft preparation, N.M.; writing—review and editing, B.B., C.B., and L.G.; supervision, L.G.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Empirical Foods Inc.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available from the corresponding author upon reasonable request.
Acknowledgments
We acknowledge the Texas Tech University Department of Biological Sciences, Texas Tech University Graduate School, Texas Tech University Association of Biologists for Grants-in-Aid, Dr. Cathy Wakeman and Dr. Naima Moustaid-Moussa for instrumentation access and advice.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Table A1.
Total dietary components and formulations.
|
D20013102 |
D20013106 |
|
| Product # |
11 kcal% Fat Control Diet |
46 kcal% Fat High-Fat Diet |
| Ingredient (g) | ||
| Casein | 200 | 200 |
| L-Cystine | 3 | 3 |
| Corn Starch | 452.2 | 72.8 |
| Maltodextrin 10 | 75 | 100 |
| Sucrose | 175.21 | 175.21 |
| Cellulose | 50 | 50 |
| Soybean Oil | 25 | 25 |
| Lard | 0 | 0 |
| Beef Fat, Bunge | 20 | 177.5 |
| Mineral Mix S10026 (No Ca, P, K) | 0 | 0 |
| Mineral Mix S10026A (No Ca, P, K, Na, Cl) | 5 | 5 |
| Dicalcium Phosphate | 13 | 13 |
| Calcium Carbonate | 5.5 | 5.5 |
| Potassium Citrate, 1 H2O | 16.5 | 16.5 |
| Sodium Chloride | 2.546 | 2.546 |
| Vitamin Mix V10001 | 10 | 10 |
| Choline Bitartrate | 2 | 2 |
| Cholesterol | 0.6 | 0.49 |
| Yellow Dye #5, FD&C | 0 | 0 |
| Red Dye #40, FD&C | 0.05 | 0 |
| Blue Dye #1, FD&C | 0 | 0.05 |
| Total | 1055.606 | 845.798 |
| gm | ||
| Protein | 179.0 | 179.0 |
| Carbohydrate | 712.4 | 358.0 |
| Fat | 47.4 | 204.9 |
| Fiber | 50.0 | 50.0 |
| Cholesterol | 0.66 | 0.66 |
| gm% | ||
| Protein | 17.0 | 20.8 |
| Carbohydrate | 67.5 | 41.7 |
| Fat | 4.5 | 23.9 |
| Fiber | 4.7 | 5.8 |
| Cholesterol | 0.062 | 0.08 |
| kcals | ||
| Protein | 716.0 | 716.0 |
| Carbohydrate | 2849.6 | 1432.0 |
| Fat | 426.6 | 1844.1 |
| Total | 3992.2 | 3992.1 |
| kcal% | ||
| Protein | 18 | 18 |
| Carbohydrate | 71 | 36 |
| Fat | 11 | 46 |
| Total | 100 | 100 |
| kcal/gm | 3.8 | 4.6 |
| Linoleic Acid, gm/kg Diet (NRC Requirement is 6.8) | 12.9 | 21.9 |
| Arginine, gm/kg Diet (NRC Requirement is 3.0) | 5.6 | 6.9 |
| Histidine, gm/kg Diet (NRC Requirement is 2.0) | 4.3 | 5.2 |
| Isoleucine, gm/kg Diet (NRC Requirement is 4.0) | 7.1 | 8.7 |
| Leucine, gm/kg Diet (NRC Requirement is 7.0) | 14.9 | 18.3 |
| Valine, gm/kg Diet (NRC Requirement is 5.0) | 8.7 | 10.7 |
| Threonine, gm/kg Diet (NRC Requirement is 4.0) | 6.7 | 8.3 |
| Lysine, gm/kg Diet (NRC Requirement is 4.0) | 12.3 | 15.1 |
| Methionine, gm/kg Diet (NRC Requirement is 5.0; half can be Cystine) | 4.7 | 5.8 |
| Cystine, gm/kg Diet (NRC Requirement is - see above) | 3.0 | 3.6 |
| Phenylalanine, gm/kg Diet (NRC Requirement is 7.6; half can be Tyrosine) | 7.9 | 9.7 |
| Tyrosine, gm/kg Diet (NRC Requirement is - see above) | 8.5 | 10.5 |
| Tryptophan, gm/kg Diet (NRC Requirement is 1.0) | 2.0 | 2.4 |
| Sodium (mg/kg diet) | 979 | 1203 |
| Sodium (mg/3982.5 kcal) | 1033 | 1033 |
References
- Carswell, K.A.; Lee, M.J.; Fried, S.K. Culture of isolated human adipocytes and isolated adipose tissue. Methods Mol Biol 2012, 806, 203–214. [Google Scholar] [CrossRef]
- Poddar, M.; Chetty, Y.; Chetty, V.T. How does obesity affect the endocrine system? A narrative review. Clin Obes 2017, 7, 136–144. [Google Scholar] [CrossRef]
- Dufau, J.; Shen, J.X.; Couchet, M.; De Castro Barbosa, T.; Mejhert, N.; Massier, L.; Griseti, E.; Mouisel, E.; Amri, E.Z.; Lauschke, V.M.; et al. In vitro and ex vivo models of adipocytes. Am J Physiol Cell Physiol 2021, 320, C822–c841. [Google Scholar] [CrossRef] [PubMed]
- Redinger, R.N. The pathophysiology of obesity and its clinical manifestations. Gastroenterol Hepatol (N Y) 2007, 3, 856–863. [Google Scholar]
- Blüher, M. Obesity: global epidemiology and pathogenesis. Nat Rev Endocrinol 2019, 15, 288–298. [Google Scholar] [CrossRef]
- Després, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Qatanani, M.; Lazar, M.A. Mechanisms of obesity-associated insulin resistance: many choices on the menu. Genes Dev 2007, 21, 1443–1455. [Google Scholar] [CrossRef]
- Mubtasim, N.; Moustaid-Moussa, N.; Gollahon, L. The Complex Biology of the Obesity-Induced, Metastasis-Promoting Tumor Microenvironment in Breast Cancer. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Wisse, B.E.; Kim, F.; Schwartz, M.W. Physiology. An integrative view of obesity. Science 2007, 318, 928–929. [Google Scholar] [CrossRef]
- Lissner, L.; Heitmann, B.L. Dietary fat and obesity: evidence from epidemiology. Eur J Clin Nutr 1995, 49, 79–90. [Google Scholar]
- Golay, A.; Bobbioni, E. The role of dietary fat in obesity. Int J Obes Relat Metab Disord 1997, 21 Suppl 3, S2–11. [Google Scholar]
- Mancuso, P.; Bouchard, B. The Impact of Aging on Adipose Function and Adipokine Synthesis. Front Endocrinol (Lausanne) 2019, 10, 137. [Google Scholar] [CrossRef]
- Varghese, M.; Song, J.; Singer, K. Age and Sex: Impact on adipose tissue metabolism and inflammation. Mech Ageing Dev 2021, 199, 111563. [Google Scholar] [CrossRef]
- Ou, M.Y.; Zhang, H.; Tan, P.C.; Zhou, S.B.; Li, Q.F. Adipose tissue aging: mechanisms and therapeutic implications. Cell Death Dis 2022, 13, 300. [Google Scholar] [CrossRef] [PubMed]
- Peek, V.; Neumann, E.; Inoue, T.; Koenig, S.; Pflieger, F.J.; Gerstberger, R.; Roth, J.; Matsumura, K.; Rummel, C. Age-Dependent Changes of Adipokine and Cytokine Secretion From Rat Adipose Tissue by Endogenous and Exogenous Toll-Like Receptor Agonists. Front Immunol 2020, 11, 1800. [Google Scholar] [CrossRef]
- Fontana, L.; Eagon, J.C.; Trujillo, M.E.; Scherer, P.E.; Klein, S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 2007, 56, 1010–1013. [Google Scholar] [CrossRef] [PubMed]
- Castro-Barquero, S.; Casas, R.; Rimm, E.B.; Tresserra-Rimbau, A.; Romaguera, D.; Martínez, J.A.; Salas-Salvadó, J.; Martínez-González, M.A.; Vidal, J.; Ruiz-Canela, M.; et al. Loss of Visceral Fat is Associated with a Reduction in Inflammatory Status in Patients with Metabolic Syndrome. Mol Nutr Food Res 2023, 67, e2200264. [Google Scholar] [CrossRef]
- Cobos-Palacios, L.; Ruiz-Moreno, M.I.; Vilches-Perez, A.; Vargas-Candela, A.; Muñoz-Úbeda, M.; Benítez Porres, J.; Navarro-Sanz, A.; Lopez-Carmona, M.D.; Sanz-Canovas, J.; Perez-Belmonte, L.M.; et al. Metabolically healthy obesity: Inflammatory biomarkers and adipokines in elderly population. PLoS One 2022, 17, e0265362. [Google Scholar] [CrossRef]
- Cobos-Palacios, L.; Muñoz-Úbeda, M.; Gallardo-Escribano, C.; Ruiz-Moreno, M.I.; Vilches-Pérez, A.; Vargas-Candela, A.; Leiva-Gea, I.; Tinahones, F.J.; Gómez-Huelgas, R.; Bernal-López, M.R. Adipokines Profile and Inflammation Biomarkers in Prepubertal Population with Obesity and Healthy Metabolic State. Children (Basel) 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Senkus, K.E.; Crowe-White, K.M.; Bolland, A.C.; Locher, J.L.; Ard, J.D. Changes in Adiponectin:Leptin ratio among older adults with obesity following a 12-month exercise and diet intervention. Nutr Diabetes 2022, 12, 30. [Google Scholar] [CrossRef]
- Oraha, J.; Enriquez, R.F.; Herzog, H.; Lee, N.J. Sex-specific changes in metabolism during the transition from chow to high-fat diet feeding are abolished in response to dieting in C57BL/6J mice. Int J Obes (Lond) 2022, 46, 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.X.; Couchet, M.; Dufau, J.; de Castro Barbosa, T.; Ulbrich, M.H.; Helmstädter, M.; Kemas, A.M.; Zandi Shafagh, R.; Marques, M.A.; Hansen, J.B.; et al. 3D Adipose Tissue Culture Links the Organotypic Microenvironment to Improved Adipogenesis. Adv Sci (Weinh) 2021, 8, e2100106. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity and Severe Obesity Among Adults: United States, 2017-2018. NCHS Data Brief 2020, 1–8. [Google Scholar]
- Neuhouser, M.L.; Aragaki, A.K.; Prentice, R.L.; Manson, J.E.; Chlebowski, R.; Carty, C.L.; Ochs-Balcom, H.M.; Thomson, C.A.; Caan, B.J.; Tinker, L.F.; et al. Overweight, Obesity, and Postmenopausal Invasive Breast Cancer Risk: A Secondary Analysis of the Women’s Health Initiative Randomized Clinical Trials. JAMA Oncol 2015, 1, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.L. Obesity-associated cardiovascular risk in women: hypertension and heart failure. Clin Sci (Lond) 2021, 135, 1523–1544. [Google Scholar] [CrossRef]
- Hruby, A.; Manson, J.E.; Qi, L.; Malik, V.S.; Rimm, E.B.; Sun, Q.; Willett, W.C.; Hu, F.B. Determinants and Consequences of Obesity. Am J Public Health 2016, 106, 1656–1662. [Google Scholar] [CrossRef]
- Sclafani, A. Animal models of obesity: classification and characterization. Int J Obes 1984, 8, 491–508. [Google Scholar]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr Res Rev 2010, 23, 270–299. [Google Scholar] [CrossRef]
- Surwit, R.S.; Feinglos, M.N.; Rodin, J.; Sutherland, A.; Petro, A.E.; Opara, E.C.; Kuhn, C.M.; Rebuffé-Scrive, M. Differential effects of fat and sucrose on the development of obesity and diabetes in C57BL/6J and A/J mice. Metabolism 1995, 44, 645–651. [Google Scholar] [CrossRef]
- Siersbæk, M.S.; Ditzel, N.; Hejbøl, E.K.; Præstholm, S.M.; Markussen, L.K.; Avolio, F.; Li, L.; Lehtonen, L.; Hansen, A.K.; Schrøder, H.D.; et al. C57BL/6J substrain differences in response to high-fat diet intervention. Sci Rep 2020, 10, 14052. [Google Scholar] [CrossRef] [PubMed]
- Fixe, P.; Praloran, V. M-CSF: haematopoietic growth factor or inflammatory cytokine? Cytokine 1998, 10, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.M.; Wiesolek, H.L.; Sumagin, R. ICAM-1: A master regulator of cellular responses in inflammation, injury resolution, and tumorigenesis. J Leukoc Biol 2020, 108, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Bruunsgaard, H.; Ladelund, S.; Pedersen, A.N.; Schroll, M.; Jørgensen, T.; Pedersen, B.K. Predicting death from tumour necrosis factor-alpha and interleukin-6 in 80-year-old people. Clin Exp Immunol 2003, 132, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Rowsey, P.J.; Bishop, B.L.; Ward, W.O.; Macphail, R.C. Serum biomarkers of aging in the Brown Norway rat. Exp Gerontol 2011, 46, 953–957. [Google Scholar] [CrossRef]
- Kwon, H.J.; Sung, B.K.; Kim, J.W.; Lee, J.H.; Kim, N.D.; Yoo, M.A.; Kang, H.S.; Baek, H.S.; Bae, S.J.; Choi, J.S.; et al. The effect of lipopolysaccharide on enhanced inflammatory process with age: Modulation of NF-κB. J Am Aging Assoc 2001, 24, 163–171. [Google Scholar] [CrossRef]
- Kovner, I.; Taicher, G.Z.; Mitchell, A.D. Calibration and validation of EchoMRI™ whole body composition analysis based on chemical analysis of piglets, in comparison with the same for DXA. Int J Body Compos Res 2010, 8, 17–29. [Google Scholar]
- Jones, A.S.; Johnson, M.S.; Nagy, T.R. Validation of quantitative magnetic resonance for the determination of body composition of mice. Int J Body Compos Res 2009, 7, 67–72. [Google Scholar]
- Tinsley, F.C.; Taicher, G.Z.; Heiman, M.L. Evaluation of a quantitative magnetic resonance method for mouse whole body composition analysis. Obes Res 2004, 12, 150–160. [Google Scholar] [CrossRef]
- Mandarim-de-Lacerda, C.A.; Del Sol, M.; Vásquez, B.; Aguila, M.B. Mice as an animal model for the study of adipose tissue and obesity. Int J Morphol 2021, 39, 1521–1528. [Google Scholar] [CrossRef]
- O'rourke, R.W.; Metcalf, M.D.; White, A.E.; Madala, A.; Winters, B.R.; Maizlin, I.I.; Jobe, B.A.; Roberts, C.T.; Slifka, M.K.; Marks, D.L. Depot-specific differences in inflammatory mediators and a role for NK cells and IFN-gamma in inflammation in human adipose tissue. Int J Obes (Lond) 2009, 33, 978–990. [Google Scholar] [CrossRef]
- Amitani, M.; Asakawa, A.; Amitani, H.; Inui, A. The role of Leptin in the control of insulin-glucose axis. Front Neurosci 2013, 7, 51. [Google Scholar] [CrossRef]
- Yoshimura, T. The chemokine MCP-1 (CCL2) in the host interaction with cancer: a foe or ally? Cell Mol Immunol 2018, 15, 335–345. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Schafer, M.J.; Noren Hooten, N.; Atkinson, E.J.; Evans, M.K.; Baker, D.J.; Quarles, E.K.; Robbins, P.D.; Ladiges, W.C.; LeBrasseur, N.K.; et al. Circulating levels of monocyte chemoattractant protein-1 as a potential measure of biological age in mice and frailty in humans. Aging Cell 2018, 17. [Google Scholar] [CrossRef]
- Kim, S.-H. Age-Associated Increasing of MCP-1 in Adults. J. Exp. Biomed. Sci. 2007, 13, 183–187. [Google Scholar]
- Tripathi, D.; Kant, S.; Pandey, S.; Ehtesham, N.Z. Resistin in metabolism, inflammation, and disease. Febs j 2020, 287, 3141–3149. [Google Scholar] [CrossRef]
- Takahashi, Y.; Kitadai, Y.; Bucana, C.D.; Cleary, K.R.; Ellis, L.M. Expression of vascular endothelial growth factor and its receptor, KDR, correlates with vascularity, metastasis, and proliferation of human colon cancer. Cancer Res 1995, 55, 3964–3968. [Google Scholar]
- Salven, P.; Ruotsalainen, T.; Mattson, K.; Joensuu, H. High pre-treatment serum level of vascular endothelial growth factor (VEGF) is associated with poor outcome in small-cell lung cancer. Int J Cancer 1998, 79, 144–146. [Google Scholar] [CrossRef]
- Niu, G.; Chen, X. Vascular endothelial growth factor as an anti-angiogenic target for cancer therapy. Curr Drug Targets 2010, 11, 1000–1017. [Google Scholar] [CrossRef]
- Jaberi, S.A.; Cohen, A.; D’Souza, C.; Abdulrazzaq, Y.M.; Ojha, S.; Bastaki, S.; Adeghate, E.A. Lipocalin-2: Structure, function, distribution and role in metabolic disorders. Biomed Pharmacother 2021, 142, 112002. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int Immunol 2021, 33, 127–148. [Google Scholar] [CrossRef]
- Yao, X.; Huang, J.; Zhong, H.; Shen, N.; Faggioni, R.; Fung, M.; Yao, Y. Targeting interleukin-6 in inflammatory autoimmune diseases and cancers. Pharmacol Ther 2014, 141, 125–139. [Google Scholar] [CrossRef]
- Uciechowski, P.; Dempke, W.C.M. Interleukin-6: A Masterplayer in the Cytokine Network. Oncology 2020, 98, 131–137. [Google Scholar] [CrossRef]
- Ying, T.H.; Lee, C.H.; Chiou, H.L.; Yang, S.F.; Lin, C.L.; Hung, C.H.; Tsai, J.P.; Hsieh, Y.H. Knockdown of Pentraxin 3 suppresses tumorigenicity and metastasis of human cervical cancer cells. Sci Rep 2016, 6, 29385. [Google Scholar] [CrossRef]
- Song, T.; Wang, C.; Guo, C.; Liu, Q.; Zheng, X. Pentraxin 3 overexpression accelerated tumor metastasis and indicated poor prognosis in hepatocellular carcinoma via driving epithelial-mesenchymal transition. J Cancer 2018, 9, 2650–2658. [Google Scholar] [CrossRef]
- Iwaki, T.; Urano, T.; Umemura, K. PAI-1, progress in understanding the clinical problem and its aetiology. Br J Haematol 2012, 157, 291–298. [Google Scholar] [CrossRef]
- Lubbers, E.R.; List, E.O.; Jara, A.; Sackman-Sala, L.; Cordoba-Chacon, J.; Gahete, M.D.; Kineman, R.D.; Boparai, R.; Bartke, A.; Kopchick, J.J.; et al. Adiponectin in mice with altered GH action: links to insulin sensitivity and longevity? J Endocrinol 2013, 216, 363–374. [Google Scholar] [CrossRef]
- Nguyen, T.M.D. Adiponectin: Role in Physiology and Pathophysiology. Int J Prev Med 2020, 11, 136. [Google Scholar] [CrossRef]
- Fruebis, J.; Tsao, T.S.; Javorschi, S.; Ebbets-Reed, D.; Erickson, M.R.; Yen, F.T.; Bihain, B.E.; Lodish, H.F. Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc Natl Acad Sci U S A 2001, 98, 2005–2010. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone Adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med 2001, 7, 941–946. [Google Scholar] [CrossRef]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat Med 2001, 7, 947–953. [Google Scholar] [CrossRef]
- Combs, T.P.; Berg, A.H.; Obici, S.; Scherer, P.E.; Rossetti, L. Endogenous glucose production is inhibited by the adipose-derived protein Acrp30. J Clin Invest 2001, 108, 1875–1881. [Google Scholar] [CrossRef]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K.; et al. Paradoxical decrease of an adipose-specific protein, Adiponectin, in obesity. Biochem Biophys Res Commun 1999, 257, 79–83. [Google Scholar] [CrossRef]
- Kern, P.A.; Di Gregorio, G.B.; Lu, T.; Rassouli, N.; Ranganathan, G. Adiponectin expression from human adipose tissue: relation to obesity, insulin resistance, and tumor necrosis factor-alpha expression. Diabetes 2003, 52, 1779–1785. [Google Scholar] [CrossRef]
- Barnea, M.; Shamay, A.; Stark, A.H.; Madar, Z. A high-fat diet has a tissue-specific effect on Adiponectin and related enzyme expression. Obesity (Silver Spring) 2006, 14, 2145–2153. [Google Scholar] [CrossRef]
- Bullen, J.W., Jr.; Bluher, S.; Kelesidis, T.; Mantzoros, C.S. Regulation of Adiponectin and its receptors in response to development of diet-induced obesity in mice. Am J Physiol Endocrinol Metab 2007, 292, E1079–E1086. [Google Scholar] [CrossRef]
- Lee, B.; Shao, J. Adiponectin and energy homeostasis. Rev Endocr Metab Disord 2014, 15, 149–156. [Google Scholar] [CrossRef]
- Qiao, L.; Lee, B.; Kinney, B.; Yoo, H.S.; Shao, J. Energy intake and Adiponectin gene expression. Am J Physiol Endocrinol Metab 2011, 300, E809–E816. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: an endocrine organ. Arch Med Sci 2013, 9, 191–200. [Google Scholar] [CrossRef]
- Morton, G.J.; Schwartz, M.W. Leptin and the central nervous system control of glucose metabolism. Physiol Rev 2011, 91, 389–411. [Google Scholar] [CrossRef]
- Brew, K.; Nagase, H. The tissue inhibitors of metalloproteinases (TIMPs): an ancient family with structural and functional diversity. Biochim Biophys Acta 2010, 1803, 55–71. [Google Scholar] [CrossRef]
- Kralisch, S.; Bluher, M.; Tonjes, A.; Lossner, U.; Paschke, R.; Stumvoll, M.; Fasshauer, M. Tissue inhibitor of metalloproteinase-1 predicts adiposity in humans. Eur J Endocrinol 2007, 156, 257–261. [Google Scholar] [CrossRef]
- Chavey, C.; Mari, B.; Monthouel, M.N.; Bonnafous, S.; Anglard, P.; Van Obberghen, E.; Tartare-Deckert, S. Matrix metalloproteinases are differentially expressed in adipose tissue during obesity and modulate adipocyte differentiation. J Biol Chem 2003, 278, 11888–11896. [Google Scholar] [CrossRef]
- Sharma, A.M.; Staels, B. Review: Peroxisome proliferator-activated receptor gamma and adipose tissue--understanding obesity-related changes in regulation of lipid and glucose metabolism. J Clin Endocrinol Metab 2007, 92, 386–395. [Google Scholar] [CrossRef]
- Meissburger, B.; Stachorski, L.; Röder, E.; Rudofsky, G.; Wolfrum, C. Tissue inhibitor of matrix metalloproteinase 1 (TIMP1) controls adipogenesis in obesity in mice and in humans. Diabetologia 2011, 54, 1468–1479. [Google Scholar] [CrossRef]
- Gerin, I.; Louis, G.W.; Zhang, X.; Prestwich, T.C.; Kumar, T.R.; Myers, M.G., Jr.; Macdougald, O.A.; Nothnick, W.B. Hyperphagia and obesity in female mice lacking tissue inhibitor of metalloproteinase-1. Endocrinology 2009, 150, 1697–1704. [Google Scholar] [CrossRef]
- Corvera, S.; Solivan-Rivera, J.; Yang Loureiro, Z. Angiogenesis in adipose tissue and obesity. Angiogenesis 2022, 25, 439–453. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, K.S.; Kraegen, E.W.; Sweeney, G.; Zhang, J.; Tso, A.W.; Chow, W.S.; Wat, N.M.; Xu, J.Y.; Hoo, R.L.; et al. Lipocalin-2 is an inflammatory marker closely associated with obesity, insulin resistance, and hyperglycemia in humans. Clin Chem 2007, 53, 34–41. [Google Scholar] [CrossRef]
- Yan, Q.W.; Yang, Q.; Mody, N.; Graham, T.E.; Hsu, C.H.; Xu, Z.; Houstis, N.E.; Kahn, B.B.; Rosen, E.D. The adipokine Lipocalin 2 is regulated by obesity and promotes insulin resistance. Diabetes 2007, 56, 2533–2540. [Google Scholar] [CrossRef]
- Zhang, Y.; Foncea, R.; Deis, J.A.; Guo, H.; Bernlohr, D.A.; Chen, X. Lipocalin 2 expression and secretion is highly regulated by metabolic stress, cytokines, and nutrients in adipocytes. PLoS One 2014, 9, e96997. [Google Scholar] [CrossRef]
- Guo, H.; Jin, D.; Zhang, Y.; Wright, W.; Bazuine, M.; Brockman, D.A.; Bernlohr, D.A.; Chen, X. Lipocalin-2 deficiency impairs thermogenesis and potentiates diet-induced insulin resistance in mice. Diabetes 2010, 59, 1376–1385. [Google Scholar] [CrossRef]
- Deis, J.A.; Guo, H.; Wu, Y.; Liu, C.; Bernlohr, D.A.; Chen, X. Adipose Lipocalin 2 overexpression protects against age-related decline in thermogenic function of adipose tissue and metabolic deterioration. Mol Metab 2019, 24, 18–29. [Google Scholar] [CrossRef]
- Law, I.K.; Xu, A.; Lam, K.S.; Berger, T.; Mak, T.W.; Vanhoutte, P.M.; Liu, J.T.; Sweeney, G.; Zhou, M.; Yang, B.; et al. Lipocalin-2 deficiency attenuates insulin resistance associated with aging and obesity. Diabetes 2010, 59, 872–882. [Google Scholar] [CrossRef]
- Alessi, M.C.; Juhan-Vague, I. PAI-1 and the metabolic syndrome: links, causes, and consequences. Arterioscler Thromb Vasc Biol 2006, 26, 2200–2207. [Google Scholar] [CrossRef]
- Samad, F.; Loskutoff, D.J. Tissue distribution and regulation of plasminogen activator inhibitor-1 in obese mice. Mol Med 1996, 2, 568–582. [Google Scholar] [CrossRef]
- Landin, K.; Stigendal, L.; Eriksson, E.; Krotkiewski, M.; Risberg, B.; Tengborn, L.; Smith, U. Abdominal obesity is associated with an impaired fibrinolytic activity and elevated plasminogen activator inhibitor-1. Metabolism 1990, 39, 1044–1048. [Google Scholar] [CrossRef]
- Guo, H.; Qiu, X.; Deis, J.; Lin, T.Y.; Chen, X. Pentraxin 3 deficiency exacerbates lipopolysaccharide-induced inflammation in adipose tissue. Int J Obes (Lond) 2020, 44, 525–538. [Google Scholar] [CrossRef]
- Thomas, C.; Henry, W.; Cuiffo, B.G.; Collmann, A.Y.; Marangoni, E.; Benhamo, V.; Bhasin, M.K.; Fan, C.; Fuhrmann, L.; Baldwin, A.S.; et al. Pentraxin-3 is a PI3K signaling target that promotes stem cell-like traits in basal-like breast cancers. Sci Signal 2017, 10. [Google Scholar] [CrossRef]
- Zhang, Q.; Lei, L.; Jing, D. Knockdown of SERPINE1 reverses resistance of triple-negative breast cancer to paclitaxel via suppression of VEGFA. Oncol Rep 2020, 44, 1875–1884. [Google Scholar] [CrossRef]
- Chen, S.; Li, Y.; Zhu, Y.; Fei, J.; Song, L.; Sun, G.; Guo, L.; Li, X. SERPINE1 Overexpression Promotes Malignant Progression and Poor Prognosis of Gastric Cancer. J Oncol 2022, 2022, 2647825. [Google Scholar] [CrossRef]
- Azimi, I.; Petersen, R.M.; Thompson, E.W.; Roberts-Thomson, S.J.; Monteith, G.R. Hypoxia-induced reactive oxygen species mediate N-cadherin and SERPINE1 expression, EGFR signalling and motility in MDA-MB-468 breast cancer cells. Sci Rep 2017, 7, 15140. [Google Scholar] [CrossRef]
- Wang, S.; Pang, L.; Liu, Z.; Meng, X. SERPINE1 associated with remodeling of the tumor microenvironment in colon cancer progression: a novel therapeutic target. BMC Cancer 2021, 21, 767. [Google Scholar] [CrossRef]
- Levine, J.A.; Oleaga, C.; Eren, M.; Amaral, A.P.; Shang, M.; Lux, E.; Khan, S.S.; Shah, S.J.; Omura, Y.; Pamir, N.; et al. Role of PAI-1 in hepatic steatosis and dyslipidemia. Sci Rep 2021, 11, 430. [Google Scholar] [CrossRef]
- Khan, S.S.; Shah, S.J.; Klyachko, E.; Baldridge, A.S.; Eren, M.; Place, A.T.; Aviv, A.; Puterman, E.; Lloyd-Jones, D.M.; Heiman, M.; et al. A null mutation in SERPINE1 protects against biological aging in humans. Sci Adv 2017, 3, eaao1617. [Google Scholar] [CrossRef]
- Sultzer, B.M. Endotoxin-induced resistance to a staphylococcal infection: cellular and humoral responses compared in two mouse strains. J Infect Dis 1968, 118, 340–348. [Google Scholar] [CrossRef]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps gene product. J Immunol 1999, 162, 3749–3752. [Google Scholar] [CrossRef]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef]
- Benomar, Y.; Taouis, M. Molecular Mechanisms Underlying Obesity-Induced Hypothalamic Inflammation and Insulin Resistance: Pivotal Role of Resistin/TLR4 Pathways. Front Endocrinol (Lausanne) 2019, 10, 140. [Google Scholar] [CrossRef]
- Jin, C.; Flavell, R.A. Innate sensors of pathogen and stress: linking inflammation to obesity. J Allergy Clin Immunol 2013, 132, 287–294. [Google Scholar] [CrossRef]
- Kuzmich, N.N.; Sivak, K.V.; Chubarev, V.N.; Porozov, Y.B.; Savateeva-Lyubimova, T.N.; Peri, F. TLR4 Signaling Pathway Modulators as Potential Therapeutics in Inflammation and Sepsis. Vaccines (Basel) 2017, 5. [Google Scholar] [CrossRef]
- Engin, A.B. Adipocyte-Macrophage Cross-Talk in Obesity. Adv Exp Med Biol 2017, 960, 327–343. [Google Scholar] [CrossRef]
- McKernan, K.; Varghese, M.; Patel, R.; Singer, K. Role of TLR4 in the induction of inflammatory changes in adipocytes and macrophages. Adipocyte 2020, 9, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Mieda, T.; Itoh, M.; Shimoda, Y.; Kamei, Y.; Ogawa, Y. Attenuation of obesity-induced adipose tissue inflammation in C3H/HeJ mice carrying a Toll-like receptor 4 mutation. Biochem Biophys Res Commun 2007, 354, 45–49. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of Mouse Adipokine Array and coordinates for antibodies against 38 commonly occurring adipocytokines.
Figure 1.
Schematic representation of Mouse Adipokine Array and coordinates for antibodies against 38 commonly occurring adipocytokines.

Figure 2.
Adipokine array showing dot blot results of different adipocytokines on nitrocellulose membrane. Dots are adipocytokine signals detected after treatment with visceral, fat-derived, conditioned medium harvested from C3H/HeJ female mice fed on control or high-fat diets, both for Month 12 and Month 18. A) The negative control is M199 media without FBS, used for conditioning cells. B) Organ cultured visceral fat harvested from control diet fed mice at Month 12. C) Organ cultured visceral fat harvested from high-fat diet fed mice at Month 12. D) Organ cultured visceral fat harvested from control fat diet fed mice at Month 18. E) Organ cultured visceral fat harvested from high-fat diet fed mice at Month 18.
Figure 2.
Adipokine array showing dot blot results of different adipocytokines on nitrocellulose membrane. Dots are adipocytokine signals detected after treatment with visceral, fat-derived, conditioned medium harvested from C3H/HeJ female mice fed on control or high-fat diets, both for Month 12 and Month 18. A) The negative control is M199 media without FBS, used for conditioning cells. B) Organ cultured visceral fat harvested from control diet fed mice at Month 12. C) Organ cultured visceral fat harvested from high-fat diet fed mice at Month 12. D) Organ cultured visceral fat harvested from control fat diet fed mice at Month 18. E) Organ cultured visceral fat harvested from high-fat diet fed mice at Month 18.
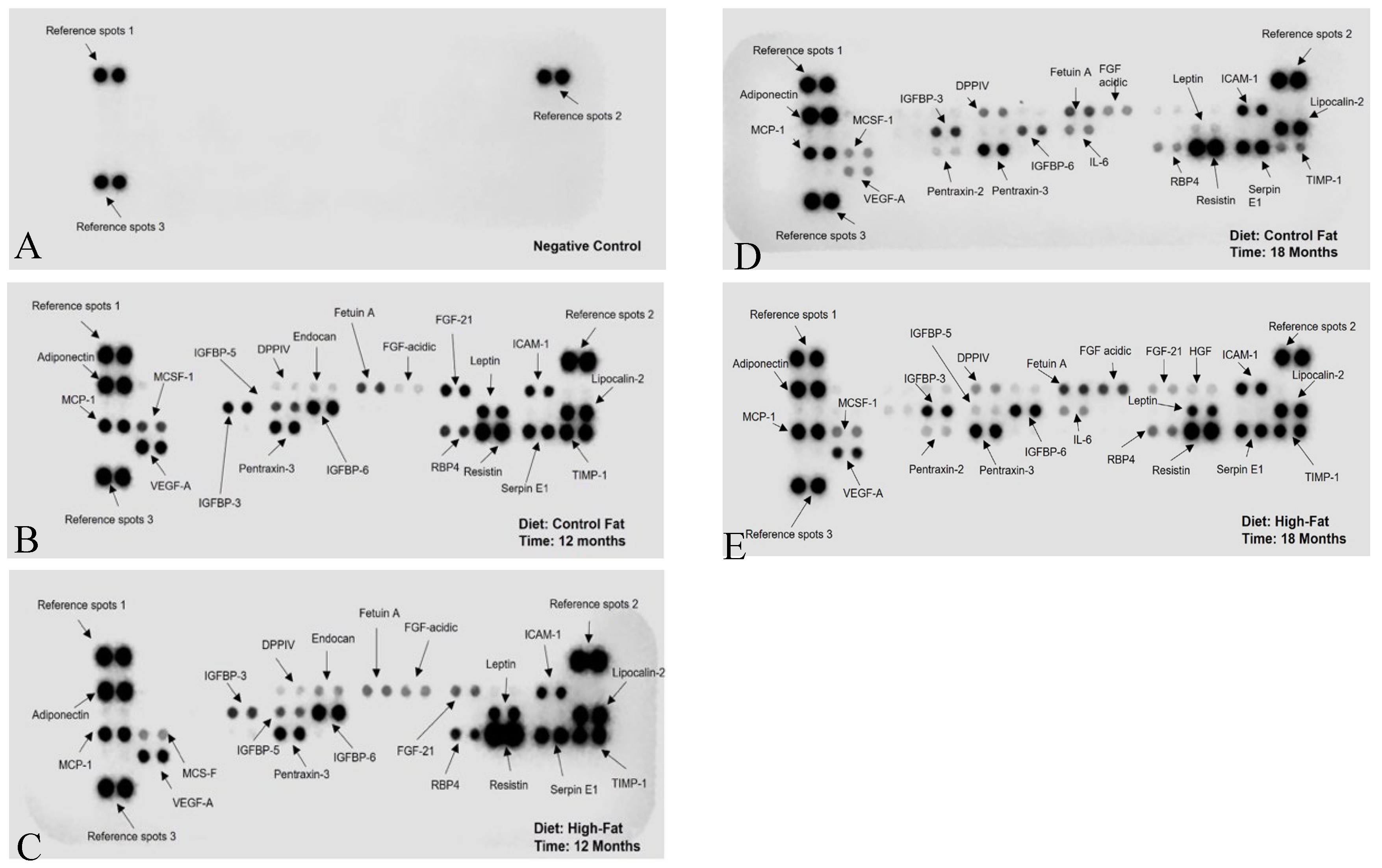
Figure 3.
Comparison of secretions of adipokines Leptin and Adiponectin and adipocytokines. A) Adiponectin, B) Leptin, (C) MCP-1, D) Resistin, E) TIMP-1, F) VEGF in the Ex-WAT-CM harvested from control and high-fat diet fed female C3H/HeJ mice at Month 12 and Month 18. Each data point represents the mean value of the secretome from 2 technical replicates for 3 independent experiments. Adipocytokine secretion changes over time, between control and high-fat diet, was determined using Two-way ANOVA followed by Tukey’s post hoc test for multiple comparison. A p value ≤ 0.05 was considered statistically significant and identified by one asterisk (*), two asterisks (**) indicates p ≤ 0.01, three asterisks (***) indicates p ≤ 0.001 and four asterisks (****) indicates p ≤ 0.0001. p-value > 0.05 was considered statistically nonsignificant and not marked on the graphs. Note: As ExV-WAT-CM samples are conditioned medium collected from Ex Vivo visceral adipose tissue, the changes in adipocytokine secretions between the conditioned medium have been designated as arbitrary units for the Y-axis following the manufacturer’s kit instruction.
Figure 3.
Comparison of secretions of adipokines Leptin and Adiponectin and adipocytokines. A) Adiponectin, B) Leptin, (C) MCP-1, D) Resistin, E) TIMP-1, F) VEGF in the Ex-WAT-CM harvested from control and high-fat diet fed female C3H/HeJ mice at Month 12 and Month 18. Each data point represents the mean value of the secretome from 2 technical replicates for 3 independent experiments. Adipocytokine secretion changes over time, between control and high-fat diet, was determined using Two-way ANOVA followed by Tukey’s post hoc test for multiple comparison. A p value ≤ 0.05 was considered statistically significant and identified by one asterisk (*), two asterisks (**) indicates p ≤ 0.01, three asterisks (***) indicates p ≤ 0.001 and four asterisks (****) indicates p ≤ 0.0001. p-value > 0.05 was considered statistically nonsignificant and not marked on the graphs. Note: As ExV-WAT-CM samples are conditioned medium collected from Ex Vivo visceral adipose tissue, the changes in adipocytokine secretions between the conditioned medium have been designated as arbitrary units for the Y-axis following the manufacturer’s kit instruction.
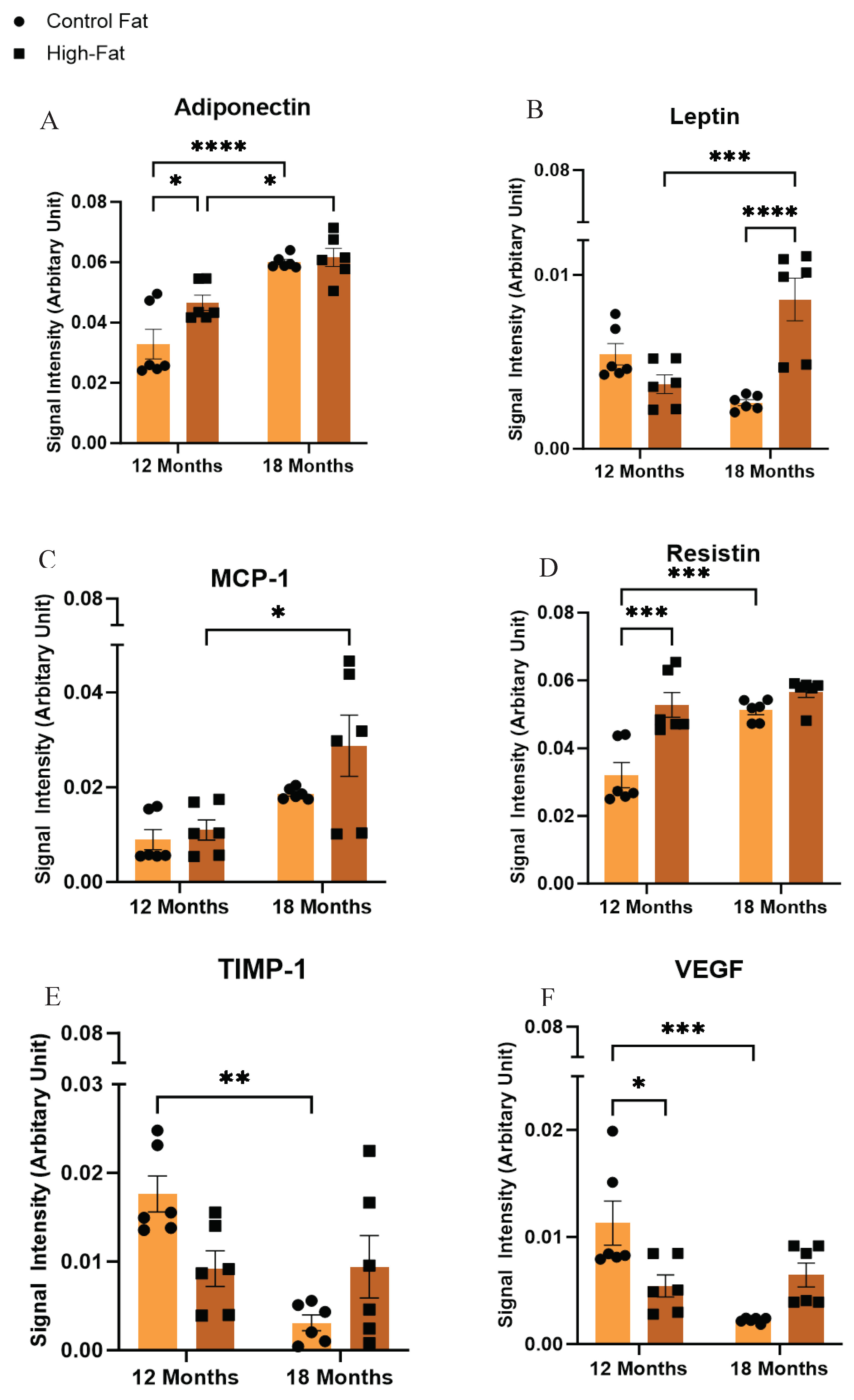
Figure 4.
Changes in adipocytokine secretions. A) IL-6, B) ICAM-1, C) Lipocalin-2, D) Serpin E1, E) Pentraxin-3, F) M-CSF in the ExV-WAT-CM harvested from control and high-fat diet fed mice female C3H/HeJ mice at Month 12 and Month 18. Each data point represents the mean value of the secretome from 2 technical replicates for 3 independent experiments. Adipocytokine secretion changes over time, between control and high-fat diets, was measured using two-way ANOVA followed by Tukey’s post hoc test for multiple comparison. A p value p ≤ 0.05 was considered statistically significant and identified by one asterisk (*), two asterisks (**) indicate p ≤ 0.01 and three asterisks (***) indicates p ≤ 0.001. Note: As ExV-WAT-CM samples are conditioned medium collected from Ex Vivo visceral adipose tissue, the changes in adipocytokine secretions between the conditioned medium have been designated as arbitrary units for the Y-axis following the manufacturer’s kit instruction.
Figure 4.
Changes in adipocytokine secretions. A) IL-6, B) ICAM-1, C) Lipocalin-2, D) Serpin E1, E) Pentraxin-3, F) M-CSF in the ExV-WAT-CM harvested from control and high-fat diet fed mice female C3H/HeJ mice at Month 12 and Month 18. Each data point represents the mean value of the secretome from 2 technical replicates for 3 independent experiments. Adipocytokine secretion changes over time, between control and high-fat diets, was measured using two-way ANOVA followed by Tukey’s post hoc test for multiple comparison. A p value p ≤ 0.05 was considered statistically significant and identified by one asterisk (*), two asterisks (**) indicate p ≤ 0.01 and three asterisks (***) indicates p ≤ 0.001. Note: As ExV-WAT-CM samples are conditioned medium collected from Ex Vivo visceral adipose tissue, the changes in adipocytokine secretions between the conditioned medium have been designated as arbitrary units for the Y-axis following the manufacturer’s kit instruction.
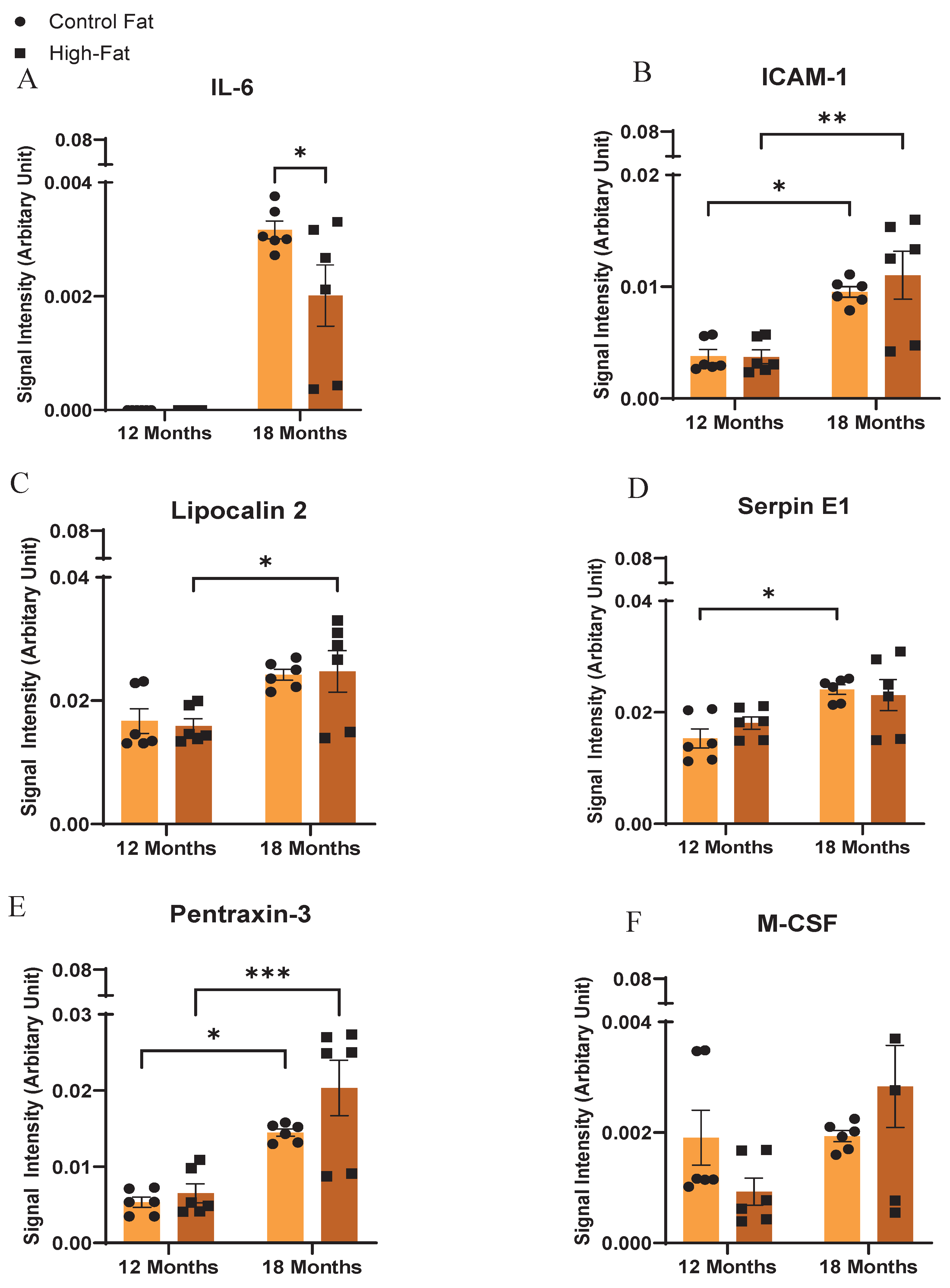
Figure 5.
Graphic representation of observable changes in the proteins extracted from visceral fat flash frozen tissue (FF-WAT), harvested from control and high fat diet fed mice at Month 12 and Month 18. A) Adiponectin, B) Leptin, (C) MCP-1, D) Resistin, E) TIMP-1, F) VEGF in the FF-WAT-tissue harvested from control and high-fat diet fed female C3H/HeJ mice at Month 12 and Month 18. Each data point represents the mean value of the protein analyzed from 3 technical replicates for 3 independent experiments. Adipocytokine protein changes over time, between control and high-fat diet, was determined using Two-way ANOVA followed by Tukey’s post hoc test for multiple comparison. A p value ≤ 0.05 was considered statistically significant and identified by one asterisk (*), two asterisks (**) indicates p ≤ 0.01. The changes in adipocytokine protein levels have been designated as arbitrary units for the Y-axis following the manufacturer’s kit instruction. Note the scale breaks in the y-axis units based on protein expression levels.
Figure 5.
Graphic representation of observable changes in the proteins extracted from visceral fat flash frozen tissue (FF-WAT), harvested from control and high fat diet fed mice at Month 12 and Month 18. A) Adiponectin, B) Leptin, (C) MCP-1, D) Resistin, E) TIMP-1, F) VEGF in the FF-WAT-tissue harvested from control and high-fat diet fed female C3H/HeJ mice at Month 12 and Month 18. Each data point represents the mean value of the protein analyzed from 3 technical replicates for 3 independent experiments. Adipocytokine protein changes over time, between control and high-fat diet, was determined using Two-way ANOVA followed by Tukey’s post hoc test for multiple comparison. A p value ≤ 0.05 was considered statistically significant and identified by one asterisk (*), two asterisks (**) indicates p ≤ 0.01. The changes in adipocytokine protein levels have been designated as arbitrary units for the Y-axis following the manufacturer’s kit instruction. Note the scale breaks in the y-axis units based on protein expression levels.
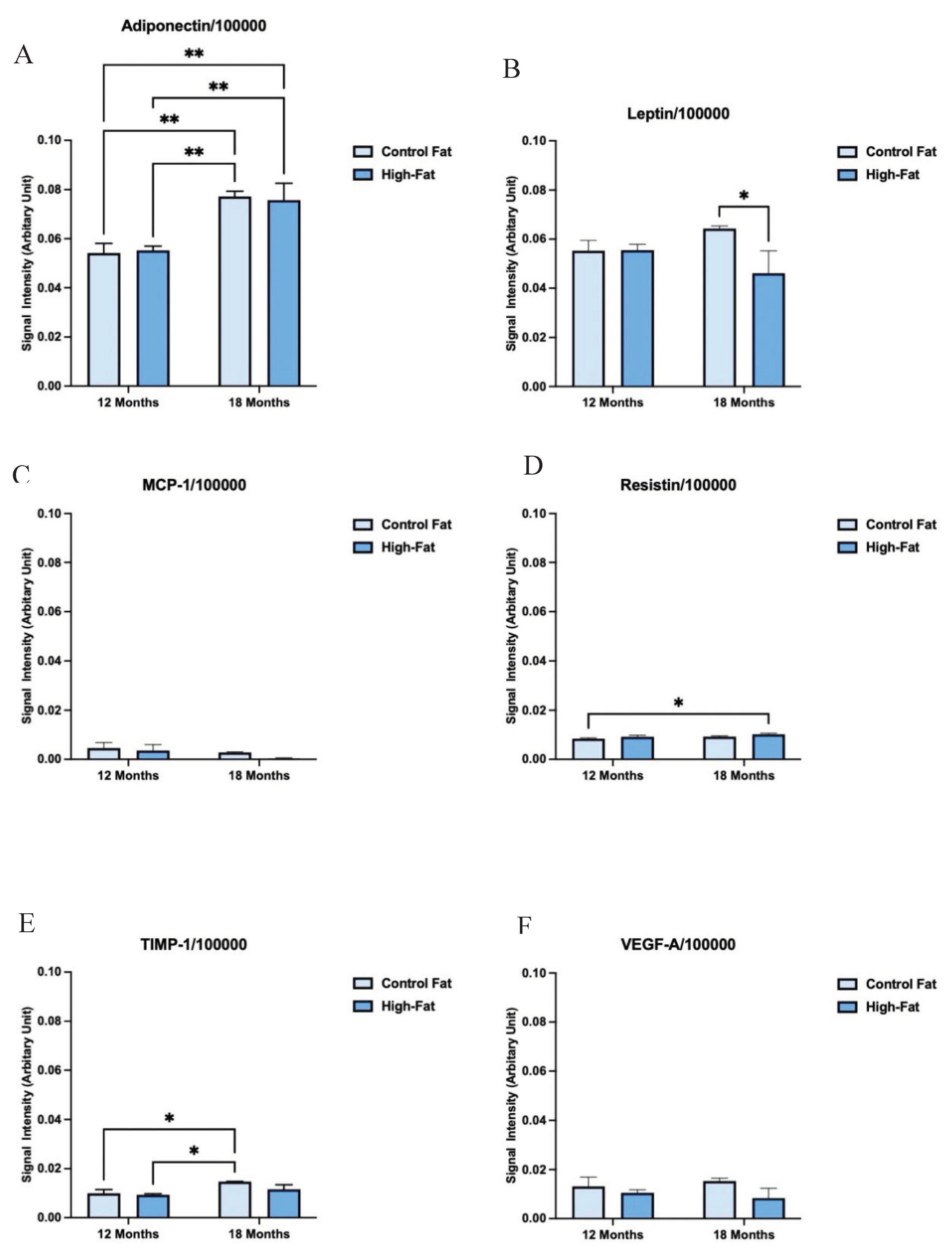
Table 1.
Media and Ingredients used for Organoid Culture.
| Ingredients | Amount | Function/Role |
|---|---|---|
| M199 media | 500 mL | Source of glucose, amino acid, vitamins, and inorganic salts |
| Gentamycin | 50 μg/mL | Antibiotic |
| Penicillin-Streptomycin | 1% or 5 mL | Antibiotics |
| Insulin | 0.7 nM | Aids in lipogenesis and fat accumulation |
| Dexamethasone | 250 nM | Helps in adipogenic gene expression. |
| Amphotericin B | 0.5 μg/mL | Antifungal agent |
| HEPES | 25 nM | Buffering agent |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



