
Submitted:
20 March 2024
Posted:
20 March 2024
You are already at the latest version
Abstract
Cell-culture meat is a novel concept to replace traditional meat produced from animals, and to satisfy rising food demands due to the world population growth. It is still a young field and requires to solve technical issues including cell sources and types, efficient culture for large scale production, taste and texture for customer satisfaction. The current review summarized traditional beef meat production and evaluation methods for customer satisfaction. In particular, serum biomarkers for beef meat evaluation are discussed, which can be applicated for cell-culture meat production.
Keywords:
cell-culture meat
; beef meat evaluation
; serum biomarkers
; cell culture media
1. Introduction:
Meat is an important food providing proteins to most people in the world. Global meat consumption is continuously increased, reached 286 million tonnes in 2013 and is expected to rise to 486 million tonnes by 2050 [1]. The global population is growing and the world population reached 7.5 billion in 2015, which may be expected to reach 10 billion by 2050, with current resources, producing enough meat for the increasing population appears to be a difficult task (Figure 1) [2].
Not only animal sources of meat, but also meat quality is a major determinant of consumer satisfaction, which differs country to country depending on culture and heritage [3]. Although meat quality is a complex concept, ISO (International Organization for Standardization) defined general meat quality, in particular, for beef meat, as the totality of features and the characteristics of a product or service that impact its ability to meet stated or implied requirements [4]. A number of characteristics, including colour, tenderness, water holding capacity, pH, appearance, smell, and taste, are used to evaluate beef meat quality. Animal species, sex, age, feeding, and pre- and post-slaughter conditions all have an impact on these parameters. Additionally, the definition of beef meat quality varies according to farmers, processors/companies, and customers. High yield was the most essential criterion for farmers' perceptions of meat quality, functional quality seemed significant for processors' perceptions of meat quality, and eating quality or palatability is significant for consumers' perceptions of meat quality [5].
It is unlikely that existing animal agriculture and meat production techniques will be able to meet the rising demand for meat. An alternative technique is required to replace traditional meat production. Cell-culture meat is a novel concept in meat industries, which can produce edible biomass by in vitro culture of stem cells harvested from the muscle of live animals [6]. Compared to conventional meat production, the in vitro technology ensures public safety, nutritional security, and an ethical manner of producing meat, while using less water and emitting fewer greenhouse gases [7]. There are still numerous challenges to be solved, including those related to cells (selection, collection, and maintenance), culture techniques for high yield, secure production, and appropriate quality [8]. Specifically, serum markers of evaluation are used to evaluate the yield and quality of traditional beef meat production. Additionally, the possible use of serum indicators as dietary supplements in culture media for meat production in cell cultures is considered. We aim to update knowledge on serum markers for assessing the quality of beef meat and explore their possible applications to the production of cultured meat.
2. Conventional evaluation of meat quality:
2.1. Color
Meat color plays a significant role in influencing consumer purchasing decisions; color is an indicator of freshness and quality, and also attractiveness at the point of sale [9]. Many factors affect meat color, which include animal breed, aging, dietary factors, type of muscle, animal management, pre and post-slaughter conditions, packaging, pre, and postmortem myoglobin chemistry [9,10]. Myoglobin, a heme containing protein, occurs in the high concentration in muscles of animals and is the principle protein responsible for meat color. The oxidation state of iron atom in the heme of myoglobin determines the meat color: purple, red, or brown for deoxymyoglobin, oxymyoglobin, or metmyoglobin, respectively. Oxidation of oxymyoglobin and deoxymyoglobin to metmyoglobin accounts for the discoloration of meat.
2.2. Tenderness
Meat tenderness is one of the major quality traits for consumer acceptability of fresh meat, repurchase decisions, and willingness to pay premium prices [11]. The tenderness of beef meat is a complex trait influenced by pre and post-harvest factors, and evaluated by instrumental and sensory analysis methods [12]. Different types of mechanisms are involved in the meat tenderization process. After slaughtering the animal/carcass tissue undergo an anoxic state, which will affect all metabolic pathways, leads to an adaptation of most metabolic process and prevents cell functions by an increase in the concentration of several enzymes [13].
2.3. Water Holding Capacity
Water Holding Capacity (WHC) is defined as the ability of meat to hold all or part of its own water, and a major quality assigned of fresh meat [14]. It influences consumer acceptance, nutritive value, and the final weight of the product. Carcass muscle contains the major portion of water (75%), protein (20%), and less quantity (5%) of remaining elements like carbohydrates, organic compounds, lipids, etc. [15]. WHC can be measured as drip loss dependent on the body temperature of the carcass and the velocity, humidity, and temperature of the air in the chilling room [16]. Many factors influence WHC, which include physiological (pH, breed, sex, age, muscle type, area of muscle), rearing conditions (feeding, administration, pre slaughter), slaughter conditions (killing method, handling, packing, storage, freezing, thawing, and aging) [17].
2.4. Beef Meat Grading Systems
Standard evaluation of meat quality and grading system are important for delivering consistent meat products to consumers. Evaluation criteria and methods differ depending on the types of meat, and most countries have their own grading systems. The grading system for beef meat is well established and generally based on the yield and the degree of marbling [18]. Marbling is visible distribution of intramuscular fat, and critically associated with beef meat flavor and juiciness, which highly influence a consumer’s consumption decisions and marketing demand. Representative grading systems for beef meat quality are summarized in Table 1.
3. Serum Biomarkers for Meat Quality:
Meat quality is evaluated after slaughtering animals. However complex pre-slaughtering factor, including genetic background, gender, nutrition, maturity and environmental conditions, determines meat quality. Muscle tissues and body fluids from live animals have been used to study pre-slaughtering factors for a high carcass grading and to develop a biomarker for meat quality evaluation. In this review, we focus on serum biomarkers related to intramuscular fat for the marbling score.
3.1. Vitamin A
Restricted vitamin A diet during fattening states of beef cattle has been shown to increase intramuscular fat with a higher carcass grade. Retinol is a fat-soluble vitamin belonging to the vitamin A family. A high concentration of serum retinol was negatively correlated with intramuscular fat and the marbling score of beef meat, while no significant correlation was obtained with the subcutaneous fat thickness [28]. Ruminants can absorb vitamin A which is converted into retinol or retinyl esters by many digestive enzymes [29]. Retinol is converted to retinoic acid through a two-step oxidation process catalyzed by alcohol dehydrogenase 1 and retinaldehyde dehydrogenases [30]. Retinoic acid was shown to regulate genes in adipocyte differentiation which in turn regulates cell surface receptors such as the retinoid X receptor (RXR) and the retinoic acid receptor (RAR) [31]. Retinoic acid suppressed adipocyte differentiation by activation of the CRABP-II/RAR pathway and decreases preadipocyte hyperplasia by activating related genes and inhibiting adipogenesis. Retinoic acid stimulates lipid oxidation and plays a crucial role in hyperplasia and hypertrophy [31,32]. This indicates controlling the blood retinol during the fattening phage gives high marbling quality meat.
3.2. Blood Urea-Nitrogen (BUN)
Nutrition plays an important role in meat quality, a sufficient amount of protein diet is critical for meat production, excess amount of protein diet leads to wastage and elevated blood urea nitrogen (BUN), Low amount of protein containing diet, failed to induce significantly lower blood urea nitrogen (BUN) production. Balanced diets maintain the intestinal flora which helps to increase the number of rumen microbes, which enhances the feed utilization and nutrient absorption in beef cattle and reduce wastage [33]. Rumen microbes breakdown the protein into small peptides, Non-protein nitrogen (NPN) such as urea also can be made into ruminal microbial protein following enzymatic conversion or breakdown of the NPN to ammonia in the rumen [34,35]. Unused ammonia is converted into urea from the deamination of amino acids; urea then circulates in the blood to the kidneys and is excreted with urine. Blood Urea Nitrogen concentrations were positively correlated with ruminal ammonia nitrogen concentrations [36,37,38]. Blood urea nitrogen concentration may be a useful marker for monitoring the protein energy status of cattle, and higher concentration decreased the back fat thickness and also increased the yield.
3.3. Non-Esterified Fatty Acid (NEFA)
Beef meat quality is related to intramuscular fat as well as its fatty acid composition. Intramuscular fat in beef meat consists of on average 50 % saturated fatty acids, 45 % monounsaturated fatty acids, and 5% polyunsaturated fatty acids. Fat deposition is an important prospect of meat quality; it is a complex of phospholipids, cholesterol, and triacylglycerides which is an important energy reserve in muscles. Meat is a significant source of long-chain omega-3 fatty acids, vitamins, proteins, and essential minerals. Fatty acid composition influences meat juiciness, aroma, tenderness, color, nutritive value, shelf-life, and market value [39,40,41].
The relationship between fatty acid percentages in meat and overall palatability is always correlated to intramuscular fat in beef. Fatty acid composition related to the flavor of meat, increases the amount of oleic acid in meat, simultaneously increasing the palatability of meat due to the fat softness associated with this fatty acid. Oleic acid reduces total plasma cholesterol and total LDL cholesterol. Reports suggest that oleic acid not only increased the palatability of meat but also supported functionally healthy food for humans [40]. Conjugated linoleic acid (CLA) amounts are rich in meat and it promotes anti-carcinogenic effects, reduction of atherosclerosis and alters the immune response. It is an important nutrient in human nutrition [41,42]. There is a strong positive correlation between stearic acid, myristic acid, and marbling score.
3.4. Total Cholesterol (TCH)
Cholesterol is necessary for the structure and function of the cells it is a pliable fat-like substance that is found in all cells of animal tissue. Cholesterol is an important component in the nutritional composition of meat. It acts as a precursor of hormonal regulation and serves as a source of energy to the organism and it can be stored as lipid droplets [43]. Carcass body weight positively correlated with total blood cholesterol concentration, and negatively correlated with yield index, marbling score, and rib-eye muscle. The cholesterol content of meat varies in different species of muscles, different muscles from the same species. Cholesterol content in processed meat is greatly varied from raw meat, because of the changes in the proportion of ingredients and formulation. Bone marrow, liver, kidney, and brain contain greater content of cholesterol up to several hundred mg/100g [42,44].
3.5. Paraoxonase 1 (PON1)
Paraoxonase1 (PON1) is a calcium-dependent serum antioxidative enzyme; it is associated with high-density lipoprotein (HDL) and plays a key role in inhibiting the oxidation of low-density lipoprotein (LDL). PON1 is synthesized in the liver and secreted into blood; it acts as a marker for early diagnosis of liver damage, like fatty liver diseases in cows [45,46]. PON1 during bovine in vitro oocyte maturation can increase the blastocyst development rate, indicating it plays an important role in the fertility of female cows. Previous reports suggest that PON1 levels are higher in females, especially during the parturition period [47,48,49,50]. PON1 levels were lower than females and castrated males indicated a gender-dependent difference. These differences correlate with the quality of meat. Analysis of three different grades of meat quality showed the PON1 concentration was lower in grade -3 than in grade-1&2. The marbling score was significantly higher in females and castrated males than in males. Certain polymorphic variants of PON1 (PON1/EcoRV and PON1/AluI loci) showed positive associations with carcass body weight, a genetic gain of tenderness, and meat color [51]. This evidence suggests that PON1 would be a useful serum marker for beef meat quality, fatty liver diagnosis, and parturition changes analysis.
3.6. Insulin:
Insulin is an important regulator for metabolic activities like glucose, lipid & protein metabolism, and it also affects growth, fertility, development, lifespan and maintains energy homeostasis in both animals and humans. Insulin can also influence glucose metabolism indirectly through changes in free fatty acids generated from hidden fat and stimulates fat synthesis, and inhibits lipid breakdown [52,53].
Glycolysis is one of the key determinants of meat quality. It is a key metabolic pathway in the conversion of muscle to meat and helps to generate ATP for cellular functions [53]. Insulin is associated with total protein and albumin and maintains a balance between synthesis and utilization. In some conditions, it increases the synthesis and decreases the degradation in favor of the anabolic process. Maintaining plasma amino acid concentrations is crucial [54,55]. Insulin regulates the lipoprotein lipase (LPL) in the clearance of plasma levels of triglycerides (TG) and LDL. TG and LPL are positively correlated to marbling [56]. Blood retinol is negatively correlated with insulin. Retinol influences insulin resistance. Unfortunately, serum retinol negatively correlated with meat color and marbling scores [57]. Insulin has negatively correlated with leptin. Leptin also increases insulin sensitivity, not only by decreasing adiposity and lipotoxicity but also through insulin-independent action [58]. Leptin is related to beef marbling, brightness, and firmness. These factors indicate insulin is a good marker of beef quality.
3.7. Leptin
Leptin is an adipocyte-derived hormone it plays a crucial role in food intake and energy metabolism, and body composition in mammals. Leptin concentrations are strongly associated with adipocyte mass. Adipocyte mass increases, and simultaneously concentrations of leptin also increase, because adipocyte size may influence leptin synthesis [59,60]. Serum concentrations of leptin were positively correlated with a marbling score, kidney, pelvic, and heart fat (KPH), fat depth, quality grade, and yield grade of meat [61]. Leptin and G6PDH (Glucose-6-phosphate dehydrogenase) are closely related to the deposition of Intramuscular fat adipose tissue in beef cattle, and insulin stimulates leptin secretion from adipose tissue [62,63]. This indicates serum leptin is a good marker for meat marbling, intramuscular fat, and meat quality.
3.8. Aspartic Acid Transaminase (AST) & Alanine Transaminase (ALT)
Serum AST & ALT levels are commonly measured clinically as biomarkers for liver health in animals and humans. AST and ALT are two important enzymes involved in the glucose metabolism amino acid metabolism pathways [64,65].
Aspartate aminotransferase (AST) is a transaminase enzyme that catalyzes the conversion of aspartate and α-ketoglutarate to oxaloacetate and glutamate and plays an important role in the synthesis of phosphoenolpyruvic acid (PEP), which is an intermediate in the production of glucose and alanine. AST and cytoplasmic free calcium are also associated with apoptosis and glycolysis. One of the important metabolic pathways involved in the transformation of muscle into meat is glycolysis. Lower AST levels can increase the amount of cytoplasmic free calcium by increasing mitochondrion depolarization and boosting calcium-mediated protease activity. This is crucial for improving the tenderness and quality of meat. AST regulates glucose levels by inhibiting the expression of glycolytic enzymes to maintain normal glucose homeostasis [66]. This indicates AST could be a useful serum marker for meat tenderness, color and quality grade.
Alanine aminotransferase (ALT) is the most important marker of liver dysfunction. ALT catalyzes the transfer of an amino group from L-alanine to α-ketoglutarate, the products of this reversible transamination reaction being pyruvate and L-glutamate, and plays an important role, as an enzyme involved in gluconeogenesis in the liver and glycolysis in the muscle, in the transport of glucose and alanine between the liver and muscle [67,68]. High levels of ALT indicate liver dysfunction, it may negatively affect meat quality, marbling, and yield index.
3.9. Total Protein (TP)
Serum total protein indicates the total amount of protein present in the blood (Albumin and Globulins); albumin helps to prevent fluid from leaking out of blood vessels. Globulins are an important part of the immune system. Serum total protein is an indicator that reflects protein metabolism in the organism, and a higher total protein concentration denotes a higher level of protein deposition in the organism, total protein levels in serum depend on animal age, breed, dietary supplementation, climatic conditions, etc. TP positively correlated with carcass body weight, springiness, gumminess, chewiness and drip loss [44]. Serum total protein is extremely correlated with lipid content which influences the meat quality [67]. The role of serum indicators on meat quality is seen in Table 2.
4. Cultured Meat and Limitations:
Conventional meat/ Traditional meat production methods are notorious for their significant depletion of natural resources and substantial contribution to global greenhouse gas emissions (GHGs), thereby posing considerable ethical and public health concerns [69,70].
In contrast, cultured meat production holds promise for addressing these issues, yet it encounters several limitations. These limitations include challenges associated with cell line screening, formulation of culture media/ serum free media formulations, selection of appropriate scaffold materials, constraints in scaling up manufacturing processes, and fostering consumer acceptance [71].
Despite its potential advantages, the scientific community must address these limitations to ensure the viability and widespread adoption of cultured meat as a sustainable alternative to conventional meat production.
4.1. Cell Line Selection for Cultured Meat Production
The selection of appropriate starter culture cells for cultured meat production is crucial, with ideal traits including self-renewal capacity, ability to differentiate, and adaptability to culture conditions. The quality and characteristics of the cultured meat primarily depends upon the properties of the cultured cells utilized. Primary cells sourced from live or deceased animal biopsies represent one avenue for cell acquisition. Stem cells include muscle cells (MC), satellite cells (SC), adipogenic stem cells (ASC), and induced pluripotent stem cells (iPSC), stand out as promising candidates owing to their undifferentiated nature, rendering them amenable to in vitro culture. Among these, adult stem cell types such as Myosatellite cells exhibit particular promise for cultured meat production, albeit necessitating additional support, such as co-culture, to facilitate the development of fat and connective tissues [71,72,73].
4.2. Serum-Free Media Formulation
One of the key cost drivers for cultured meat is the cell culture medium. Culture media comprises vitamins, amino acids, carbohydrates, organic and inorganic salts, and other nutrients that are required for cell proliferation and differentiation. The majority of media is made from animal sources, such as fetal bovine serum. It renders cultured meat generated from serum media inappropriate for vegan and specific religious consumption (Halal& Kosher) [73,74].
The precise composition of the cultured meat media has not yet been determined. Components originating from genetically modified plants and microbial fermentation, such as corn steep solids, yeast, or soy extracts, have a high concentration of critical nutrients for cell development and are prospective replacements for serum in cultured meat production [75,76,77]. Beefy-9; a serum-free media that efficiently supports long-term iPSC cells, Knockout TM Serum Replacement (KSR), is a serum-free formulation medium commonly utilized in the culture of ESCs and induced pluripotent stem cells (iPSCs). It effectively maintains stemness and promotes cell proliferation over time, as in an FBS-containing medium. There have been positive findings from research using a serum-free medium with the inclusion of extra proteins and the development of innovative media such as AIM-V, Sericin, and Ultroser-G. Amino acid-rich plant-based media derived from maitake mushroom extract was found to be optimal for fish explant development and surface area expansion [76,77,78]. Cyanobacteria can be employed as a potential food source for cell growth in meat culture [79,80].
4.3. Scaffold
It is very difficult to grow cells in media and get tissue structured without the appropriate provision of a scaffold, a solid surface is required for growing myoblast for attachment and maximizing their surface [81]. Scaffolds not only provide support for the cells but also allow for a continual flow of nutrients and oxygen, which aids in the maintenance of important metabolic functions in cultured meat cells. Scaffolds are porous 3D structures that provide particular support for cell growth, adhesion, differentiation, and maturation. The benefit of having an edible scaffold is that it can improve the texture of the product, add flavor, and aid in co-culture by avoiding the removal of cultured tissues from the scaffold and potential harm during this process [82,83].
Scaffold materials that are commonly mixed with synthetic polymers include starch, cellulose, and its derivatives chitin, alginate, and agarose. As scaffolding in cultured meat development, recombinant protein-based materials such as fibrin, collagen, gelatin, and keratin may be of interest [84]. The current state of research on animal-free biomaterial scaffolds is quite restricted. Aleph Farms uses a more advanced strategy to manufacture a fully formed steak by co-culturing numerous cell types including microsatellites, endothelial cells, and extracellular secreting cells within a three-dimensional (3D) porous scaffold [85,86]. BSA/Starch scaffold is a sophisticated type of scaffold composed of protein and carbohydrate components that is suitable for tissue growth [87].
4.4. Limitations of Cultured Meat Production
Cultured meat may be biologically equivalent to traditional meat, but there are still many technical difficulties to solve to produce fresh meat like cultured meat. Traditional meat consists mostly of skeletal muscle which grows in long, thin fibers (Intramuscular fat (IMF)), but the case of cultured meat may require intensive research on this [88].
Industrial Production of lab grown meat is associated with economic challenges as well. Cultured meat is closely related to forecast demand and the intensity of the production process. The industrial scale of culture meat production requires bioreactors in volumes up to several thousand liters [89]. Considerable optimization is needed to produce cultured meat efficiently at a huge scale with very low expenditures. In a conventional 5,000-liter bioreactor, it is predicted that 8 trillion muscle cells will be needed to create 1 kilogram of protein [90]. Many companies like Mosa meats and Memphis meats are building pilot plants but have not established large-scale manufacturing. To prepare chicken bits at the “1880” restaurant in Singapore, US startup “Eat Just” employed a bioreactor with a 10,000-liter capacity. The company proposed to have bioreactors of about 50,000 liter capacity for the commercialization of in-vitro meat around the globe [6]. Further innovations are needed to develop scaffolds and bioreactor conditions that enable the large-scale production of cultured meat.
4.5. Overcoming Solutions
Tissue engineering, gene therapy, transgene-free technology like microfluidics, and cell therapy, can provide potential solutions to the current cell line development for cultured meat production. iPSCs are generated from differentiated cells by inducing the expression of pluripotent transcription factors that can be induced by viral vector-mediated transduction. iPSCs can reach immortality through epigenetic changes and up-regulation of the enzyme telomerase, which are helpful for cell expansion in large-scale production [91,92].
Large-scale production of in-vitro meat culture requires a series of bioreactors, which provide a controlled environment for various parameters such as temperature, pH, dissolved oxygen, carbon dioxide, continuous supply of nutrients, and waste removal. Industrial bioreactors for meat culture should be designed in such a way that facilitates cell suspension in a near-continuous medium, maintain lower fluid shear force, proper growth of cells similar to native tissue, facilitates the separation of suspended tissue, and should provide an environment for appropriate growth and cell differentiation leading to increased efficiency. Efficient bioreactors should also be automated with real-time monitoring and support the continuous supply of nutrients and removal of waste products [73,89]. Media recycling also is an option to reduce production costs, but recycling technologies are in the budding stage and require further development [93,94,95].
Consumer acceptance is the major attribute of cultured meat production, they do not accept new technologies easily, such as genetically modified organisms, due to a lack of awareness and suspicious attitude towards science, its inaccessibility, and unfamiliarity could further intensify food neophobia. One of the major things is that “awareness” of cultured meat is the finest predictor of acceptance [96,97]. People are more willing to try cultured meat when presented with more information about possible benefits and an affordable price. Clarify the belief/religious and behavioral intention to culture meat, and provide tracking, tracing, and sensorial information, these factors may influence the consumer acceptability of cultured meat [98]. Figure 2 represents, the factors involved in the commercialization of cultured meat.
5. Potential Application of Serum Markers for Culture Meat Production:
Muscle tissue engineering can be applied to produce cultured meat for human consumption it is an alternative to conventional meat. The motive of cultured meat production is animal/serum-free cultured meat development. The serum contains lots of proteins, lipids, enzymes, electrolytes, hormones, and other elements which support cell growth. In-vitro meat might be biologically equivalent to conventional meat but there are still many technical difficulties to solve to produce fresh meat-like cultured meat [99]. Serum influences the cultured meat color, taste, aroma, etc. Due to a lack of serum or serum-free media utilization for cultured meat production, the end product may lead to quality and quantity differences from conventional meat [88]. However, cultured meat production media/serum-free media to date have had limited success, suggesting that a better understanding of optimization conditions is needed.
Cultured muscle tissues generally have a pale color due to the deficiency of myoglobin, which is a potential biomarker for meat color, and the amount of myoglobin content in meat may influence the color of meat (dark to pale). Myoglobin contains heme, which has iron in the center of its structure, and its expression is suppressed under ambient oxygen conditions. Several approaches have been suggested to increase the myoglobin content of cultured meat [100]. The addition of metmyoglobin (oxidized form of myoglobin) to the culture media was shown to increase the cell proliferation capacity and myoglobin content in the cultured cells [101]. However, myoglobin contents were still much lower compared to traditional meat, and the resulting color was brown because there might be a limit to the number of nutrients, the cells can incorporate in in-vitro conditions, to solve this issue needs to be optimizing the culture conditions and media. Retinol (RBP-4) and vitamin-E supplements, according to prior research, may improve the color of the meat.
The composition of amino acids in meat is important because they determine the quality and taste, cultured meat has fewer amino acids and nucleotide composition and when compared to traditional meat, it gives taste and tenderness differences compared to a normal steak, and it shows a negative effect on consumer acceptance and economy. Cultured meat of beef showed that the aspartic acid and glutamic acid range was lower than the conventional meat, aspartic acid is a potential marker related to the meat taste and palatability. Glutamic acid was most closely related to taste. It is necessary to find a method to increase the ratio of aspartic acid and glutamic acid when culturing satellite cells [102]. We suggest micro-supplements of amino acids and vitamin D3 may improve the amino acid composition, taste and tenderness of cultured meat.
Nucleotide-related compounds like adenosine-5 monophosphate, inosine-5 monophosphate; inosine, and hypoxanthine were responsible for the umami, mouth feel, and juiciness senses. In the case of cultured meat, inosine-5 mono phosphate content was significantly lower than traditional meat it may cause weaker umami intensity [103]. It is necessary to find a solution to increase the number of nucleotide derivatives that involve umami, and juiciness in culture meat production. The addition of vitamin E and protein-rich components in the medium may improve the juiciness and umami taste of cultured meat.
Insulin is a potential meat quality marker, deficiency of insulin in culture media, may significantly affect induced pluripotent stem cell (iPSCs) attachment, proliferation, morphology, expression, colony formation, cell cycle profile, and protein phosphorylation. In cultured meat production, the differentiation of pluripotent or adipose-derived stem cells into adipocytes can be stimulated by insulin and insulin growth factors [104]. The addition of natural substitutes instead of insulin cannot meet such criteria. It is necessary to solve this problem needs to be optimizing the culture media supplement with insulin.
Retinol is the alcohol form of vitamin A, and it is associated with the maintenance of self-renewal and differentiation of embryonic stem cells and pluripotent stem cells. Retinol executes its function via retinoic acid and regulates the function of genes involved in cell proliferation, differentiation, and development [105,106]. Retinol executes its function by activating the phosphatidylinositol 3 (PI3) kinase signaling pathway via the insulin-like growth factor 1 (IGF1) receptor [107]. Retinol deficiency in culture media may lead to associated immune deficiency causing the impaired function of lymphocytes and detrimental to blood cells [105]. So this information gives a clear idea of retinol deficiency in media may decelerate stem cell proliferation, differentiation, and development and may influence cultured meat production yield and quality.
Cholesterol is an essential component for mammalian cells, it plays a crucial role in cellular functions such as the biosynthesis of bile acids, production of steroid hormones, embryonic development, water salts, electrolyte balance, and cell proliferation [108]. The serum contains numerous cholesterols and lipids; it will lead to support cell survival, at the same time cholesterol deficient cells suffer from impaired osmatic resistance disrupting the cell membrane, apoptosis, and necrosis [109]. Serum-free media contains fewer nutrients and growth factors when compared to commercial media, it is necessary to optimize the serum-free media and keep the right amount of cholesterol-like compounds for enhancing cell growth and cultured meat quality.
Paraoxonase-1 (PON1) is a calcium-dependent serum protein, involved in lipid metabolism and plays a key role against oxidative stress. PON1 is an HDL-associated protein that can hydrolyze oxidized LDL-cholesterol, with potential atheroprotective effects [110]. PON1 shows a positive association with the fatty acid composition of the adipose tissue [111]. PON1 level was positively associated with meat quality, supplementation of PON1 may increase the fatty acid composition and adipose tissue formation which may enhance the quality of cultured meat.
Leptin is a hormone synthesized and secreted from adipocytes and plays a crucial role in biological functions in cattle. Leptin concentration is positively correlated with meat thickness, marbling score, and intramuscular fat, and is also negatively correlated with cholesterol content [112,113,114]. This indicates supplementation of leptin in meat culture media gives us promising results and improves the meat yield and quality.
Non- esterified fatty acids (NEFA) are involved in the improvement of the intramuscular fat content and fatty acid composition of meat. Cellular lipids and albumin associated lipids are involved in the regulation of stem cell differentiation. NEFA and meat composition are closely related because they influence flavor, texture, and caloric content [115,116]. These facts point to NEFA's significant contribution to the development of cultured meat. Blood urea nitrogen (BUN) was positively correlated with total protein, albumin, and NEFA; this indicates that BUN affects the protein content, flavor, and texture of meat. Micro supplementation of Blood Urea Nitrogen (BUN) and also other compounds may enhance cultured meat quality and yield [60]. In the development of stem cells, serum total protein (TP) in the media, supplies energy to increase the rate of cell division, regeneration, and proliferation. Total protein is necessary for metabolic processes and affects yield grade, yield score and quality of the meat [117].
Alanine aminotransferase (ALT) was positively connected with AST, NEFA, albumin, and BUN and negatively correlated with leptin, cholesterol, and glucose These connections could affect the color and quality of meat [58].
The enzyme serum aspartate aminotransferase (AST), which is present in the liver, heart, skeletal muscles, and brain, is widely distributed. It is necessary for cell survival because it promotes cell growth and prevents mitochondrial disintegration by controlling the release of calcium into the cytoplasm. The link between AST and cytoplasmic free calcium, which is robust, is essential for the death of cells. In order to maintain calcium homeostasis during the postmortem period, increased AST activity can prevent the release of apoptotic agents as well as the activation of calcium-mediated enzymes, whereas decreased AST activity may be associated with a rapid apoptotic process. Lower AST activity may lead to a faster pH decrease and generate inferior-quality meat in the post mortem muscles. Higher AST activity showed a slower pH decline and WBS values (Warner-Bratzler shear force), which led to less force being needed to break down the cooked meat [66,118]. AST levels can regulate apoptosis, glycolysis and rate of lactate generation. This highlights how crucial AST supplementation in culture media is.
6. Conclusion:
In contrast to conventional meat production, cultured meat production offers a promising answer to future meat shortages, global food security, and the preservation of the environment resources. Despite the fact that it ought to be economical, it is still at the budding stage and has not yet been developed for large-scale production. Additionally, important problems including social, religious, and legal difficulties have an impact on the production of cultured meat. The price of culture media will always be too expensive; hence culture conditions must be optimized. Researchers concentrated on better and more sustainable production inputs, including cell selection, culture conditions, and co-culturing, specialized bioreactors, appropriate scaffolding material, and formulation of serum-free media.
There is currently no optimal medium for cultured meat production, as scientists are concentrating on serum-free media implementations. The type of grown cells determines how the media should be optimized. Promising alternatives to serum media for the development of cultured meat include materials generated from plants and bacteria that have undergone genetic engineering. Beefy-9, AIM-V, Sericin, Ultroser-G and KnockoutTM Serum Replacement (KSR) are the serum-free culture media, yet still concentrating on production and yield concerns due to a media gap during media optimization, some essential elements that are necessary to raise meat grade and quality were lacking. In this study, we discussed significant concerns regarding conventional meat quality and yield, particularly serum markers of evaluation, and we also made some suggestions regarding the potential use of serum markers for the production of cell-cultured meat.
Therefore, to increase the output and quality of the cultured meat, productive research on the optimization of cultured meat production settings is needed. We propose that, adding serum markers to the culture media is a cutting-edge and secure method for increasing the production and quality of cultured meat. Consumer awareness is the only way to combat food phobia, clarify consumers' beliefs and behaviors regarding cultured meat, and provide comprehensive product tracking, tracing, sensory information, and an affordable price. These elements may attract consumers to cultured meat and have an impact on their acceptance of it.
Author Contributions
C.S.K investigation and writing original draft, J.K designed the study, provided the supervision and support. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2020R1A6A1A03044512 and 2020R1I1A3060716) and the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (IPET) through the High Value-added Food Technology Development Program, funded by the Ministry of Agriculture, Food, and Rural Affairs (MAFRA) (321026-05).
Conflicts of Interest
Authors declared that no conflict of interest.
References
- Valin, H.; Sands, R.D.; van der Mensbrugghe, D.; Nelson, G.C.; Ahammad, H.; Blanc, E.; Bodirsky, B.; Fujimori, S.; Hasegawa, T.; Havlik, P.; et al. The Future of Food Demand: Understanding Differences in Global Economic Models. Agric. Econ. 2014, 45, 51–67. [Google Scholar] [CrossRef]
- Sans, P.; Combris, P. World Meat Consumption Patterns: An Overview of the Last Fifty Years (1961-2011). Meat Sci. 2015, 109, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.T.; Kim, G.D.; Hwang, Y.H.; Ryu, Y.C. Control of Fresh Meat Quality through Manipulation of Muscle Fiber Characteristics. Meat Sci. 2013, 95, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Geletu, U.S.; Usmael, M.A.; Mummed, Y.Y.; Ibrahim, A.M. Quality of Cattle Meat and Its Compositional Constituents. Vet. Med. Int. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Grunert, K.G.; Bredahl, L.; Brunsø, K. Consumer Perception of Meat Quality and Implications for Product Development in the Meat Sector-a Review. Meat Sci. 2004, 66, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Sharma, N.; Sharma, S.; Mehta, N.; Verma, A.K.; Chemmalar, S.; Sazili, A.Q. In-Vitro Meat: A Promising Solution forsustainability of Meat Sector. J. Anim. Sci. Technol. 2021, 63, 693. [Google Scholar] [CrossRef]
- Tuomisto, H.L.; Teixeira De Mattos, M.J. Environmental Impacts of Cultured Meat Production. Environ. Sci. Technol. 2011, 45, 6117–6123. [Google Scholar] [CrossRef]
- Post, M.J. Cultured Beef: Medical Technology to Produce Food. J. Sci. Food Agric. 2014, 94, 1039–1041. [Google Scholar] [CrossRef]
- Smith, G.C.; Belk, K.E.; Sofos, J.N.; Tatum, J.D.; Williams, S.N. Economic Implications of Improved Color Stability in Beef. Antioxidants muscle foods Nutr. Strateg. to Improv. Qual. 2000, 397–426. [Google Scholar]
- Gagaoua, M.; Hughes, J.; Terlouw, E.M.C.; Warner, R.D.; Purslow, P.P.; Lorenzo, J.M.; Picard, B. Proteomic Biomarkers of Beef Colour. Trends Food Sci. Technol. 2020, 101, 234–252. [Google Scholar] [CrossRef]
- O’Quinn, T.G.; Legako, J.F.; Brooks, J.C.; Miller, M.F. Evaluation of the Contribution of Tenderness, Juiciness, and Flavor to the Overall Consumer Beef Eating Experience. Transl. Anim. Sci. 2018, 2, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Dunshea, F.; Ha, M.; Purslow, P.; Miller, R.; Warner, R.; Vaskoska, R.S.; Wheeler, T.L.; Li, X. Meat Tenderness: Underlying Mechanisms, Instrumental Measurement, and Sensory Assessment. Meat Muscle Biol. 2021, 4. [Google Scholar] [CrossRef]
- Ouali, A.; Gagaoua, M.; Boudida, Y.; Becila, S.; Boudjellal, A.; Herrera-Mendez, C.H.; Sentandreu, M.A. Biomarkers of Meat Tenderness: Present Knowledge and Perspectives in Regards to Our Current Understanding of the Mechanisms Involved. Meat Sci. 2013, 95, 854–870. [Google Scholar] [CrossRef]
- Warner, R.D. The Eating Quality of Meat—IV Water-Holding Capacity and Juiciness. Lawrie’s Meat Sci. Eighth Ed. 2017, 419–459. [Google Scholar] [CrossRef]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. ScientificWorldJournal. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Delele, M.A.; Kuffi, K.D.; Geeraerd, A.; De Smet, S.; Nicolai, B.M.; Verboven, P. Optimizing Precooling of Large Beef Carcasses Using a Comprehensive Computational Fluid Dynamics Model. J. Food Process Eng. 2019, 42. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Lonergan, S.M. Mechanisms of Water-Holding Capacity of Meat: The Role of Postmortem Biochemical and Structural Changes. Meat Sci. 2005, 71, 194–204. [Google Scholar] [CrossRef]
- Font-i-Furnols, M.; Tous, N.; Esteve-Garcia, E.; Gispert, M. Do All the Consumers Accept Marbling in the Same Way? The Relationship between Eating and Visual Acceptability of Pork with Different Intramuscular Fat Content. Meat Sci. 2012, 91, 448–453. [Google Scholar] [CrossRef]
- What’s Your Beef – Prime, Choice or Select? | USDA. Available online: https://www.usda.gov/media/blog/2013/01/28/whats-your-beef-prime-choice-or-select?page=1 (accessed on 30 April 2023).
- What’s Your Beef – Prime, Choice or Select? | USDA. Available online: https://www.usda.gov/media/blog/2013/01/28/whats-your-beef-prime-choice-or-select?page=1 (accessed on 17 October 2022).
- Grading - Lone Mountain Cattle. Available online: https://www.lonemountaincattle.com/about-wagyu/grading/ (accessed on 30 April 2023).
- Australian Wagyu Grading – The Wagyu Shop. Available online: https://wagyushop.com/pages/australian-wagyu-grading (accessed on 30 April 2023).
- Wagyu Grading Comparison – Wagyu Prime. Available online: https://wagyuprime.com/pages/beef-grading-comparison (accessed on 30 April 2023).
- What the Beef? Canadian vs American Beef Grading -. Available online: https://smellaque.com/2020/08/12/what-the-beef-canadian-vs-american-beef-grading/ (accessed on 30 April 2023).
- Beef Quality Grading in Europe: Shifting to a Consumer Centred Perception of Beef Quality | Farming Connect. Available online: https://businesswales.gov.wales/farmingconnect/news-and-events/technical-articles/beef-quality-grading-europe-shifting-consumer-centred-perception-beef-quality (accessed on 30 April 2023).
- Liu, J.; Chriki, S.; Ellies-Oury, M.P.; Legrand, I.; Pogorzelski, G.; Wierzbicki, J.; Farmer, L.; Troy, D.; Polkinghorne, R.; Hocquette, J.F. European Conformation and Fat Scores of Bovine Carcasses Are Not Good Indicators of Marbling. Meat Sci. 2020, 170, 108233. [Google Scholar] [CrossRef]
- 축산물품질평가원 홈페이지. Available online: https://www.ekape.or.kr/index.do (accessed on 30 April 2023).
- Knutson, E.E.; Menezes, A.C.B.; Sun, X.; Fontoura, A.B.P.; Liu, J.H.; Bauer, M.L.; Maddock-Carlin, K.R.; Swanson, K.C.; Ward, A.K. Effect of Feeding a Low-Vitamin A Diet on Carcass and Production Characteristics of Steers with a High or Low Propensity for Marbling. Animal 2020, 14, 2308–2314. [Google Scholar] [CrossRef]
- Chen, W.; Chen, G. The Roles of Vitamin A in the Regulation of Carbohydrate, Lipid, and Protein Metabolism. J. Clin. Med. 2014, 3, 453. [Google Scholar] [CrossRef] [PubMed]
- Reichert, B.; Yasmeen, R.; Jeyakumar, S.M.; Yang, F.; Thomou, T.; Alder, H.; Duester, G.; Maiseyeu, A.; Mihai, G.; Harrison, E.H.; et al. Concerted Action of Aldehyde Dehydrogenases Influences Depot-Specific Fat Formation. Mol. Endocrinol. 2011, 25, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Gorocica-Buenfil, M.A.; Fluharty, F.L.; Bohn, T.; Schwartz, S.J.; Loerch, S.C. Effect of Low Vitamin A Diets with High-Moisture or Dry Corn on Marbling and Adipose Tissue Fatty Acid Composition of Beef Steers. J. Anim. Sci. 2007, 85, 3355–3366. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.C.; DeSantis, D.; Soltanian, H.; Croniger, C.M.; Noy, N. Retinoic Acid Upregulates Preadipocyte Genes to Block Adipogenesis and Suppress Diet-Induced Obesity. Diabetes 2012, 61, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Tshuma, T.; Fosgate, G.T.; Hamman, R.; Holm, D.E. Effect of Different Levels of Dietary Nitrogen Supplementation on the Relative Blood Urea Nitrogen Concentration of Beef Cows. Trop. Anim. Health Prod. 2019, 51, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The Rumen Microbiome: A Crucial Consideration When Optimising Milk and Meat Production and Nitrogen Utilisation Efficiency. Gut Microbes 2019, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Xu, Z.; Shen, Z.; Tian, Y.; Shen, H. Dietary Energy Level Promotes Rumen Microbial Protein Synthesis by Improving the Energy Productivity of the Ruminal Microbiome. Front. Microbiol. 2019, 10, 847. [Google Scholar] [CrossRef] [PubMed]
- Spek, J.W.; Dijkstra, J.; Van Duinkerken, G.; Bannink, A. A Review of Factors Influencing Milk Urea Concentration and Its Relationship with Urinary Urea Excretion in Lactating Dairy Cattle. J. Agric. Sci. 2013, 151, 407–423. [Google Scholar] [CrossRef]
- Müller, C.B.M.; Görs, S.; Derno, M.; Tuchscherer, A.; Wimmers, K.; Zeyner, A.; Kuhla, B. Differences between Holstein Dairy Cows in Renal Clearance Rate of Urea Affect Milk Urea Concentration and the Relationship between Milk Urea and Urinary Nitrogen Excretion. Sci. Total Environ. 2021, 755. [Google Scholar] [CrossRef]
- Xia, C.; Rahman, M.A.U.; Yang, H.; Shao, T.; Qiu, Q.; Su, H.; Cao, B. Effect of Increased Dietary Crude Protein Levels on Production Performance, Nitrogen Utilisation, Blood Metabolites and Ruminal Fermentation of Holstein Bulls. Asian-Australasian J. Anim. Sci. 2018, 31, 1643–1653. [Google Scholar] [CrossRef]
- Schumacher, M.; Delcurto-Wyffels, H.; Thomson, J.; Boles, J. Fat Deposition and Fat Effects on Meat Quality-A Review. Anim. an open access J. from MDPI 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Kazala, E.C.; Lozeman, F.J.; Mir, P.S.; Laroche, A.; Bailey, D.R.C.; Weselake, R.J. Relationship of Fatty Acid Composition to Intramuscular Fat Content in Beef from Crossbred Wagyu Cattle. J. Anim. Sci. 1999, 77, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Ladeira, M.M.; Schoonmaker, J.P.; Swanson, K.C.; Duckett, S.K.; Gionbelli, M.P.; Rodrigues, L.M.; Teixeira, P.D. Review: Nutrigenomics of Marbling and Fatty Acid Profile in Ruminant Meat. Animal 2018, 12, s282–s294. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Nutritional Composition of Red Meat. Nutr. Diet. 2007, 64, S113–S119. [Google Scholar] [CrossRef]
- Muchenje, V.; Hugo, A.; Dzama, K.; Chimonyo, M.; Strydom, P.E.; Raats, J.G. Cholesterol Levels and Fatty Acid Profiles of Beef from Three Cattle Breeds Raised on Natural Pasture. J. Food Compos. Anal. 2009, 22, 354–358. [Google Scholar] [CrossRef]
- Dinh, T.T.N.; Thompson, L.D.; Galyean, M.L.; Brooks, J.C.; Patterson, K.Y.; Boylan, L.M. Cholesterol Content and Methods for Cholesterol Determination in Meat and Poultry. Compr. Rev. Food Sci. Food Saf. 2011, 10, 269–289. [Google Scholar] [CrossRef]
- Kota, S.K.; Meher, L.K.; Kota, S.K.; Jammula, S.; Krishna, S.V.S.; Modi, K.D. Implications of Serum Paraoxonase Activity in Obesity, Diabetes Mellitus, and Dyslipidemia. Indian J. Endocrinol. Metab. 2013, 17, 402. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, G.; Bacchetti, T.; Moroni, C.; Savino, S.; Liuzzi, A.; Balzola, F.; Bicchiega, V. Paraoxonase Activity in High-Density Lipoproteins: A Comparison between Healthy and Obese Females. J. Clin. Endocrinol. Metab. 2005, 90, 1728–1733. [Google Scholar] [CrossRef] [PubMed]
- Rincón, J.; Madeira, E.M.; Campos, F.T.; Mion, B.; Silva, J.F.; Absalón-Medina, V.A.; Butler, W.R.; Corrêa, M.N.; Pegoraro, L.; Schneider, A. Exogenous Paraoxonase-1 during Oocyte Maturation Improves Bovine Embryo Development in Vitro. Reprod. Domest. Anim. 2016, 51, 827–830. [Google Scholar] [CrossRef]
- Silveira, P.A.S.; Butler, W.R.; LaCount, S.E.; Overton, T.R.; Barros, C.C.; Schneider, A. Polymorphisms in the Anti-Oxidant Paraoxonase-1 (PON1) Gene Associated with Fertility of Postpartum Dairy Cows. Theriogenology 2019, 125, 302–309. [Google Scholar] [CrossRef]
- Rincón, J.; Madeira, E.M.; Campos, F.T.; Mion, B.; Silva, J.F.; Absalón-Medina, V.A.; Butler, W.R.; Corrêa, M.N.; Pegoraro, L.; Schneider, A. Exogenous Paraoxonase-1 during Oocyte Maturation Improves Bovine Embryo Development in Vitro. Reprod. Domest. Anim. 2016, 51, 827–830. [Google Scholar] [CrossRef]
- Silveira, P.A.S.; Butler, W.R.; LaCount, S.E.; Overton, T.R.; Barros, C.C.; Schneider, A. Polymorphisms in the Anti-Oxidant Paraoxonase-1 (PON1) Gene Associated with Fertility of Postpartum Dairy Cows. Theriogenology 2019, 125, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Ji, A.G.; Huai, Y.H.; Zhou, Z.K.; Li, J.Y.; Zhang, L.P.; Xu, S.Z.; Gao, X.; Ren, H.Y.; Chen, J.B. Association between PON1 Gene SNPs and Growth and Carcass Traits in Beef Cattle. Asian-Australasian J. Anim. Sci. 2008, 21, 1097–1102. [Google Scholar] [CrossRef]
- Stajkovic, S.; Vasilev, D.; Teodorovic, V.; Karabasil, N. Postmortem Glycolysis and Pork Quality. IOP Conf. Ser. Earth Environ. Sci. 2019, 333, 012032. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, F. Tissue-Specific Insulin Signaling in the Regulation of Metabolism and Aging. IUBMB Life 2014, 66, 485–495. [Google Scholar] [CrossRef]
- Newsholme, E.A.; Dimitriadis, G. Integration of Biochemical and Physiologic Effects of Insulin on Glucose Metabolism. Exp. Clin. Endocrinol. Diabetes, 2. [CrossRef]
- Goldberg, I.J.; Eckel, R.H.; Abumrad, N.A. Regulation of Fatty Acid Uptake into Tissues: Lipoprotein Lipase- and CD36-Mediated Pathways. J. Lipid Res. [CrossRef]
- Jindřichová, E.; Kratochvílová, S.; Kovář, J. Glucose Administration Downregulates Lipoprotein Lipase Activity in Vivo: A Study Using Repeated Intravenous Fat Tolerance Test. Physiol. Res. 2007, 56, 175–181. [Google Scholar] [CrossRef]
- Graham, T.E.; Yang, Q.; Blüher, M.; Hammarstedt, A.; Ciaraldi, T.P.; Henry, R.R.; Wason, C.J.; Oberbach, A.; Jansson, P.-A.; Smith, U.; et al. Retinol-Binding Protein 4 and Insulin Resistance in Lean, Obese, and Diabetic Subjects. N. Engl. J. Med. 2006, 354, 2552–2563. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.H.; Cho, W.K.; Lee, S.S. Investigation of Blood Biomarkers Related to Meat Quality and Quantity in Hanwoo Steers. Asian-Australasian J. Anim. Sci. 2018, 31, 1923. [Google Scholar] [CrossRef]
- Kuźnicka, E.; Gabryszuk, M.; Kunowska-Slósarz, M.; Gołębiewski, M.; Balcerak, M. Plasma Leptin as a Predictor for Carcass Composition in Growing Lambs. Can. J. Anim. Sci. 2017, 97, 193–198. [Google Scholar] [CrossRef]
- Geary, T.W.; McFadin, E.L.; MacNeil, M.D.; Grings, E.E.; Short, R.E.; Funston, R.N.; Keisler, D.H. Leptin as a Predictor of Carcass Composition in Beef Cattle. J. Anim. Sci. 2003, 81, 1–8. [Google Scholar] [CrossRef]
- Kononoff, P.J.; Deobald, H.M.; Stewart, E.L.; Laycock, A.D.; Marquess, F.L.S. The Effect of a Leptin Single Nucleotide Polymorphism on Quality Grade, Yield Grade, and Carcass Weight of Beef Cattle. J. Anim. Sci. 2005, 83, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Asakuma, S.; Morishita, H.; Sugino, T.; Kurose, Y.; Kobayashi, S.; Terashima, Y. Circulating Leptin Response to Feeding and Exogenous Infusion of Insulin in Sheep Exposed to Thermoneutral and Cold Environments. Comp. Biochem. Physiol. - A Mol. Integr. Physiol. 2003, 134, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, M.; Faulconnier, Y.; Leroux, C.; Jurie, C.; Cassar-Malek, I.; Bauchart, D.; Boulesteix, P.; Pethick, D.; Hocquette, J.F.; Chilliard, Y. Glucose-6-Phosphate Dehydrogenase and Leptin Are Related to Marbling Differences among Limousin and Angus or Japanese Black x Angus Steers. J. Anim. Sci. 2007, 85, 2882–2894. [Google Scholar] [CrossRef]
- McGill, M.R. The Past and Present of Serum Aminotransferases and the Future of Liver Injury Biomarkers. EXCLI J. 2016, 15, 817. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R. The Past and Present of Serum Aminotransferases and the Future of Liver Injury Biomarkers. EXCLI J. 2016, 15, 817–828. [Google Scholar] [CrossRef]
- Lee, B.; Choi, Y.M. Association of Serum Glucose, Serotonin, Aspartate Aminotransferase, and Calcium Levels with Meat Quality and Palatability Characteristics of Broiler Pectoralis Major Muscle. Anim. an Open Access J. from MDPI 2022, 12. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Dai, C.; Li, J.; Huang, P.; Li, Y.; Ding, X.; Huang, J.; Hussain, T.; Yang, H. Effects of Dietary Energy on Growth Performance, Carcass Characteristics, Serum Biochemical Index, and Meat Quality of Female Hu Lambs. Anim. Nutr. 2020, 6, 499. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.R.; Lott, J.A.; Nolte, F.S.; Gretch, D.R.; Koff, R.S.; Seeff, L.B. Diagnosis and Monitoring of Hepatic Injury. I. Performance Characteristics of Laboratory Tests. Clin. Chem. 2000, 46, 2027–2049. [Google Scholar] [CrossRef] [PubMed]
- Salter, A.M. The Effects of Meat Consumption on Global Health. Rev. Sci. Tech. 2018, 37, 47–55. [Google Scholar] [CrossRef]
- Greger, M. The Human/Animal Interface: Emergence and Resurgence of Zoonotic Infectious Diseases. Crit. Rev. Microbiol. 2007, 33, 243–299. [Google Scholar] [CrossRef]
- Balasubramanian, B.; Liu, W.; Pushparaj, K.; Park, S. The Epic of In Vitro Meat Production—A Fiction into Reality. Foods 2021, 10. [Google Scholar] [CrossRef]
- Shaikh, S.; Lee, E.J.; Ahmad, K.; Ahmad, S.S.; Chun, H.J.; Lim, J.H.; Lee, Y.H.; Choi, I. Cell Types Used for Cultured Meat Production and the Importance of Myokines. Foods 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Stephens, N.; Di Silvio, L.; Dunsford, I.; Ellis, M.; Glencross, A.; Sexton, A. Bringing Cultured Meat to Market: Technical, Socio-Political, and Regulatory Challenges in Cellular Agriculture. Trends Food Sci. Technol. 2018, 78, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Guttieres, D.; Koenigsberg, A.; Barone, P.W.; Sinskey, A.J.; Springs, S.L. Large-Scale Cultured Meat Production: Trends, Challenges and Promising Biomanufacturing Technologies. Biomaterials 2022, 280. [Google Scholar] [CrossRef]
- Karnieli, O.; Friedner, O.M.; Allickson, J.G.; Zhang, N.; Jung, S.; Fiorentini, D.; Abraham, E.; Eaker, S.S.; Yong, T.K.; Chan, A.; et al. A Consensus Introduction to Serum Replacements and Serum-Free Media for Cellular Therapies. Cytotherapy 2017, 19, 155–169. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.N.; Cosenza, Z.A.; Baar, K.; Block, D.E. Considerations for the Development of Cost-Effective Cell Culture Media for Cultivated Meat Production. Compr. Rev. food Sci. food Saf. 2021, 20, 686–709. [Google Scholar] [CrossRef]
- Fujita, H.; Endo, A.; Shimizu, K.; Nagamori, E. Evaluation of Serum-Free Differentiation Conditions for C2C12 Myoblast Cells Assessed as to Active Tension Generation Capability. Biotechnol. Bioeng. 2010, 107, 894–901. [Google Scholar] [CrossRef]
- Kuo, H.H.; Gao, X.; DeKeyser, J.M.; Fetterman, K.A.; Pinheiro, E.A.; Weddle, C.J.; Fonoudi, H.; Orman, M. V.; Romero-Tejeda, M.; Jouni, M.; et al. Negligible-Cost and Weekend-Free Chemically Defined Human IPSC Culture. Stem cell reports 2020, 14, 256–270. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.A.; Khan, S.; Murid, M.; Asif, Z.; Oboturova, N.P.; Nagdalian, A.A.; Blinov, A.V.; Ibrahim, S.A.; Jafari, S.M. Marketing Strategies for Cultured Meat: A Review. Appl. Sci. 2022, 12, 8795. [Google Scholar] [CrossRef]
- Benjaminson, M.A.; Gilchriest, J.A.; Lorenz, M. In Vitro Edible Muscle Protein Production System (MPPS): Stage 1, Fish. Acta Astronaut. 2002, 51, 879–889. [Google Scholar] [CrossRef]
- Ben-Arye, T.; Shandalov, Y.; Ben-Shaul, S.; Landau, S.; Zagury, Y.; Ianovici, I.; Lavon, N.; Levenberg, S. Textured Soy Protein Scaffolds Enable the Generation of Three-Dimensional Bovine Skeletal Muscle Tissue for Cell-Based Meat. Nat. Food 2020, 1, 210–220. [Google Scholar] [CrossRef]
- MacQueen, L.A.; Alver, C.G.; Chantre, C.O.; Ahn, S.; Cera, L.; Gonzalez, G.M.; O’Connor, B.B.; Drennan, D.J.; Peters, M.M.; Motta, S.E.; et al. Muscle Tissue Engineering in Fibrous Gelatin: Implications for Meat Analogs. npj Sci. Food 2019, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Orellana, N.; Sánchez, E.; Benavente, D.; Prieto, P.; Enrione, J.; Acevedo, C.A. A New Edible Film to Produce In Vitro Meat. Foods 2020, 9. [Google Scholar] [CrossRef]
- Wollschlaeger, J.O.; Maatz, R.; Albrecht, F.B.; Klatt, A.; Heine, S.; Blaeser, A.; Kluger, P.J. Scaffolds for Cultured Meat on the Basis of Polysaccharide Hydrogels Enriched with Plant-Based Proteins. Gels 2022, 8, 94. [Google Scholar] [CrossRef]
- Xiang, N.; Yuen, J.S.K.; Stout, A.J.; Rubio, N.R.; Chen, Y.; Kaplan, D.L. 3D Porous Scaffolds from Wheat Glutenin for Cultured Meat Applications. Biomaterials 2022, 285. [Google Scholar] [CrossRef]
- Bomkamp, C.; Skaalure, S.C.; Fernando, G.F.; Ben-Arye, T.; Swartz, E.W.; Specht, E.A. Scaffolding Biomaterials for 3D Cultivated Meat: Prospects and Challenges. Adv. Sci. 2022, 9, 2102908. [Google Scholar] [CrossRef] [PubMed]
- Prasopdee, T.; Sinthuvanich, C.; Chollakup, R.; Uttayarat, P.; Smitthipong, W. The Albumin/Starch Scaffold and Its Biocompatibility with Living Cells. Mater. Today Commun. 2021, 27, 102164. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Kumar, S.; Fayaz, H. In Vitro Meat Production: Challenges and Benefits over Conventional Meat Production. J. Integr. Agric. 2015, 14, 241–248. [Google Scholar] [CrossRef]
- Allan, S.J.; De Bank, P.A.; Ellis, M.J. Bioprocess Design Considerations for Cultured Meat Production With a Focus on the Expansion Bioreactor. Front. Sustain. Food Syst. 2019, 3, 44. [Google Scholar] [CrossRef]
- Faustman, C.; Hamernik, D.; Looper, M.; Zinn, S.A. Cell-Based Meat: The Need to Assess Holistically. J. Anim. Sci. 2020, 98, 1–7. [Google Scholar] [CrossRef]
- Zidarič, T.; Milojević, M.; Vajda, J.; Vihar, B.; Maver, U. Cultured Meat: Meat Industry Hand in Hand with Biomedical Production Methods. Food Eng. Rev. 2020, 12, 498–519. [Google Scholar] [CrossRef]
- Moritz, M.S.M.; Verbruggen, S.E.L.; Post, M.J. Alternatives for Large-Scale Production of Cultured Beef: A Review. J. Integr. Agric. 2015, 14, 208–216. [Google Scholar] [CrossRef]
- Pajčin, I.; Knežić, T.; Azoulay, I.S.; Vlajkov, V.; Djisalov, M.; Janjušević, L.; Grahovac, J.; Gadjanski, I. Bioengineering Outlook on Cultivated Meat Production. Micromachines 2022, 13. [Google Scholar] [CrossRef]
- Escobar, M.I.R.; Cadena, E.; Nhu, T.T.; Cooreman-Algoed, M.; De Smet, S.; Dewulf, J. Analysis of the Cultured Meat Production System in Function of Its Environmental Footprint: Current Status, Gaps and Recommendations. Foods 2021, 10, 2941. [Google Scholar] [CrossRef] [PubMed]
- Reiss, J.; Robertson, S.; Suzuki, M. Cell Sources for Cultivated Meat: Applications and Considerations throughout the Production Workflow. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Tomiyama, A.J.; Kawecki, N.S.; Rosenfeld, D.L.; Jay, J.A.; Rajagopal, D.; Rowat, A.C. Bridging the Gap between the Science of Cultured Meat and Public Perceptions. Trends Food Sci. Technol. 2020, 104, 144–152. [Google Scholar] [CrossRef]
- Teixeira, A.; Rodrigues, S. Consumer Perceptions towards Healthier Meat Products. Curr. Opin. Food Sci. 2021, 38, 147–154. [Google Scholar] [CrossRef]
- Chriki, S.; Hocquette, J.F. The Myth of Cultured Meat: A Review. Front. Nutr. 2020, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Post, M.J.; Hocquette, J.F. New Sources of Animal Proteins: Cultured Meat. New Asp. Meat Qual. From Genes to Ethics 2017, 425–441. [Google Scholar] [CrossRef]
- Simsa, R.; Yuen, J.; Stout, A.; Rubio, N.; Fogelstrand, P.; Kaplan, D.L. Extracellular Heme Proteins Influence Bovine Myosatellite Cell Proliferation and the Color of Cell-Based Meat. Foods 2019, 8. [Google Scholar] [CrossRef]
- Fraeye, I.; Kratka, M.; Vandenburgh, H.; Thorrez, L. Sensorial and Nutritional Aspects of Cultured Meat in Comparison to Traditional Meat: Much to Be Inferred. Front. Nutr. 2020, 7, 35. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.T.; Choi, J.S.; Hur, S.J.; Kim, G.D.; Kim, C.J.; Lee, E.Y.; Bakhsh, A.; Hwang, Y.H. A Comparative Study on the Taste Characteristics of Satellite Cell Cultured Meat Derived from Chicken and Cattle Muscles. Food Sci. Anim. Resour. 2022, 42, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Cui, J.; Liu, X.; Zhang, Y.; Qin, N.; Luo, Y. Application of Artificial Neural Network to Predict the Change of Inosine Monophosphate for Lightly Salted Silver Carp (Hypophthalmichthys Molitrix) during Thermal Treatment and Storage. J. Food Process. Preserv. 2017, 41. [Google Scholar] [CrossRef]
- Shahbazi, M.; Cundiff, P.; Zhou, W.; Lee, P.; Patel, A.; D’Souza, S.L.; Abbasi, F.; Quertermous, T.; Knowles, J.W. The Role of Insulin as a Key Regulator of Seeding, Proliferation, and MRNA Transcription of Human Pluripotent Stem Cells. Stem Cell Res. Ther. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Khillan, J.S. Vitamin A/Retinol and Maintenance of Pluripotency of Stem Cells. Nutrients 2014, 6, 1209–1222. [Google Scholar] [CrossRef]
- Rajala, K.; Vaajasaari, H.; Suuronen, R.; Hovatta, O.; Skottman, H. Effects of the Physiochemical Culture Environment on the Stemness and Pluripotency of Human Embryonic Stem Cells. Stem Cell Stud. 2011, 1, e3. [Google Scholar] [CrossRef]
- Kawaguchi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A Membrane Receptor for Retinol Binding Protein Mediates Cellular Uptake of Vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Bernecker, C.; Köfeler, H.; Pabst, G.; Trötzmüller, M.; Kolb, D.; Strohmayer, K.; Trajanoski, S.; Holzapfel, G.A.; Schlenke, P.; Dorn, I. Cholesterol Deficiency Causes Impaired Osmotic Stability of Cultured Red Blood Cells. Front. Physiol. 2019, 10, 1529. [Google Scholar] [CrossRef]
- Fernández, C.; Lobo, M.D.V.T.; Gómez-Coronado, D.; Lasunción, M.A. Cholesterol Is Essential for Mitosis Progression and Its Deficiency Induces Polyploid Cell Formation. Exp. Cell Res. 2004, 300, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Kulka, M.; Kołodziejska-Lesisz, J.; Kluciński, W. Serum Paraoxonase 1 (PON1) Activity and Lipid Metabolism Parameters Changes in Different Production Cycle Periods of Holstein-Friesian, Polish Red and Norwegian Breeds. Pol. J. Vet. Sci. 2016, 19, 165–173. [Google Scholar] [CrossRef]
- Kulka, M.; Bełtowski, J.; Kluciński, W.; Orłowska, M.; Kołodziejska, J.; Kleczkowski, M. Serum Paraoxonase-1 Activity of Dairy Holstein-Fresian Cows in Different Lactation Stages--Preliminary Study. Pol. J. Vet. Sci. 2014, 17, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Zhang, M.; Yang, L.; Jin, Y. Correlations between Circulating Leptin Concentrations and Growth Performance, Carcass Traits, and Meat Quality Indexes in Finishing Simmental × Luxi Bulls Fed High-Concentrate Diets. Anim. Sci. J. 2020, 91. [Google Scholar] [CrossRef] [PubMed]
- Berg, E.P.; McFadin, E.L.; Maddock, K.R.; Goodwin, R.N.; Baas, T.J.; Keisler, D.H. Serum Concentrations of Leptin in Six Genetic Lines of Swine and Relationship with Growth and Carcass Characteristics. J. Anim. Sci. 2003, 81, 167–171. [Google Scholar] [CrossRef]
- Luo, G.; Wang, L.; Hu, S.; Du, K.; Wang, J.; Lai, S. Association of Leptin MRNA Expression with Meat Quality Trait in Tianfu Black Rabbits. Anim. Biotechnol. 2022, 33, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Fish, K.D.; Rubio, N.R.; Stout, A.J.; Yuen, J.S.K.; Kaplan, D.L. Prospects and Challenges for Cell-Cultured Fat as a Novel Food Ingredient. Trends food Sci. Technol. 2020, 98, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Tor, M.; Vilaró, F.; Ros-Freixedes, R.; Álvarez-Rodríguez, J.; Bosch, L.; Gol, S.; Pena, R.N.; Reixach, J.; Estany, J. Circulating Non-Esterified Fatty Acids as Biomarkers for Fat Content and Composition in Pigs. Anim. an Open Access J. from MDPI 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Yeo, G.C.; Weiss, A.S. Soluble Matrix Protein Is a Potent Modulator of Mesenchymal Stem Cell Performance. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 2042–2051. [Google Scholar] [CrossRef]
- Wang, C.; Chen, H.; Ying, W. Cytosolic Aspartate Aminotransferase Mediates the Mitochondrial Membrane Potential and Cell Survival by Maintaining the Calcium Homeostasis of BV2 Microglia. Neuroreport 2018, 29, 99–105. [Google Scholar] [CrossRef]
Figure 1.
Worldwide meat consumption and the anticipated increase in population by 2050.
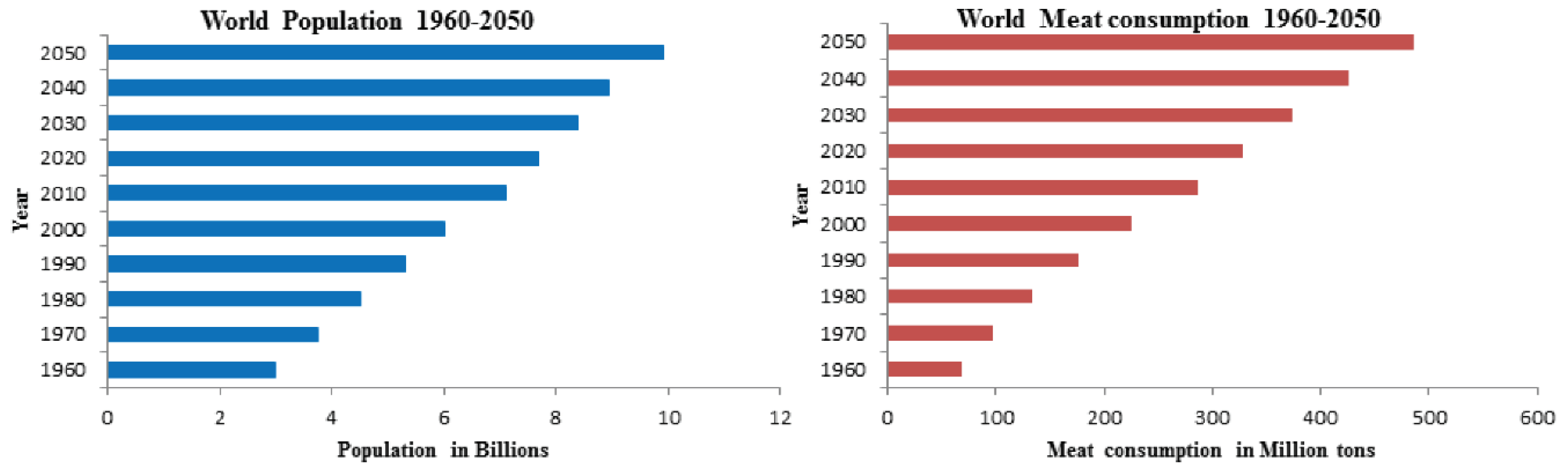
Figure 2.
Factors involved in the commercialization of cultured meat.
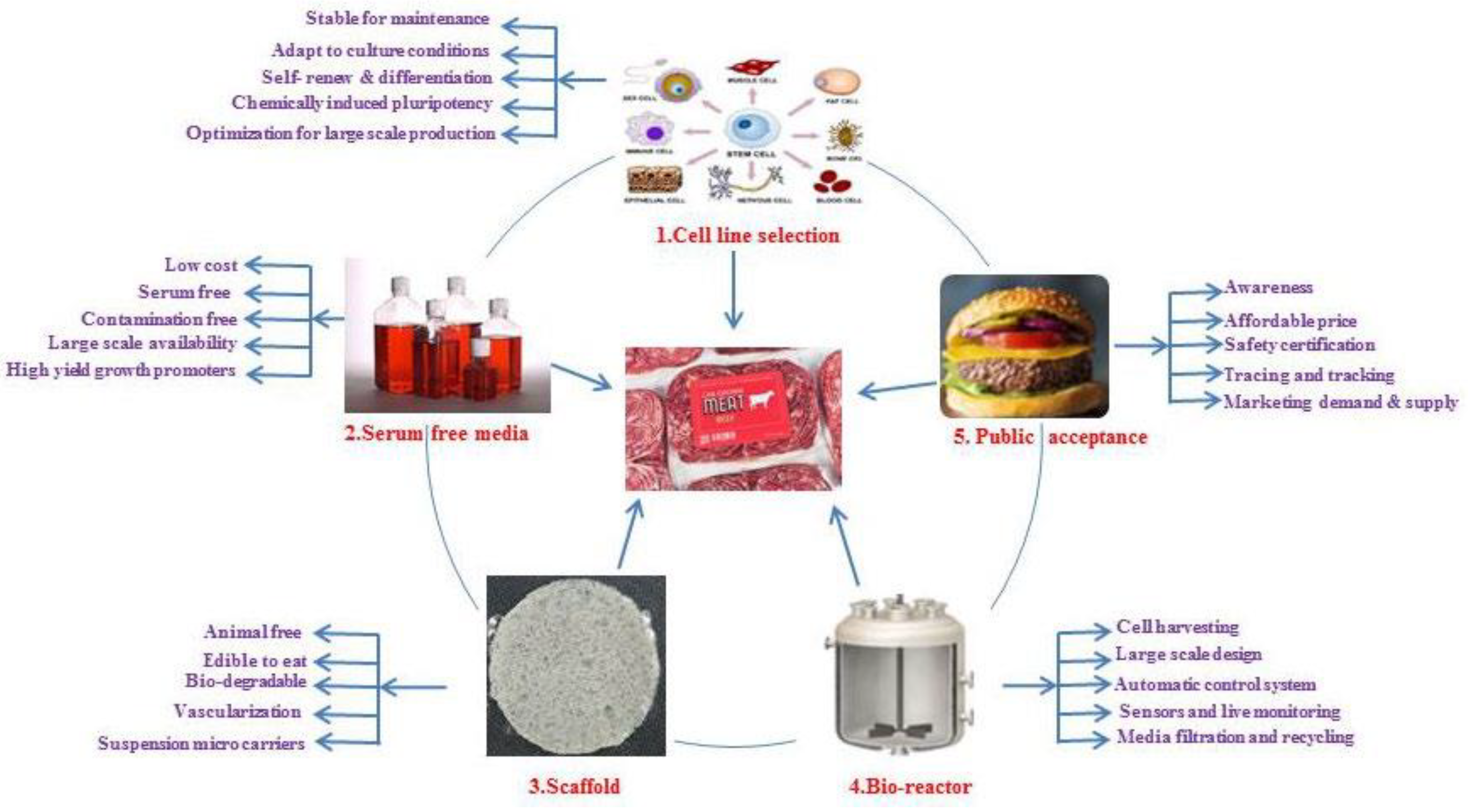

Table 1.
Top beef meat grading systems and quality selection used globally.
| Country | Board | Type of Grade | Classification | Quality (Intra muscular fat/ Beef marbling grade) | References |
|---|---|---|---|---|---|
| USA | USDA (United States Department of Agriculture) |
Prime Choice Select Standard Commercial Utility Cutter Canner |
USDA classified the beef grades based on marbling and maturity of the steak Marbling (Abundant to Practically Devoid) Maturity- (Grade A to E) Maturity divide into 5 groups based on carcass age A-9- 30 Months, B- 30–42 Months, C- 42–72 Months, D- 72-96 Months, E- > 96 Months Colour and tenderness of the steak based on maturity Grade-A- Red, porous and soft Grade-B- Slightly red and slightly soft Grade-C- Tinged with red, slightly hard Grade-D- Rather white, moderately hard Grade-E-White, nonporous, extremely hard |
Prime-Abundant &above (Grade A&B) Choice – Moderate(Grade A&B) Select - Slight (Grade A) Standard-Slight traces (Grade A&B) Commercial – Small (Grade C, D&E) Utility – Traces ( Grade C, D&E) Cutter - Practically Devoid (Grade E) Canner- Practically Devoid (Grade E) |
[19,20] |
| Japan | JMGA (Japanese Meat Grading Association) | A (A5, A4, A3, A2, A1) B C |
JMGA classified the meat grades based on yield, and quality Yield is determined by rib eye area, rib thickness, subcutaneous fat thickness, and chilled carcass weight. Yield grade divide into A,B,C (A: 72 and greater B: 69 -72 C: less than 69) Quality grade is classified by yield percentages estimated by an equation. Quality grades divide into five groups (5-1) based on marbling, meat color, texture, fat color and quality. Based on marbling, meat color, quality scores divide into 1-12 classes A5 - No.8~No.12 A4 - No.5~No.7 A3 - No.3~No.4 A2 - No.2 A1 - No.1 A- Represents yield grade. 5- 1- Represents quality grade. No 1-12 - Represents quality score. |
A5- Very abundant (No.8~No.12) A4- Abundant (No.5~No.7) A3- Standard (No.3~No.4) A2- Scarce (No.2) A1-Very scarce (No.1) |
[21] |
|
Australia |
Meat and Livestock Australia (MLA) |
AUS-MEAT - M9 to M0 MSA - 100-1100+ |
MSA classified the meat grades based on meat colour, marbling, fat depth, carcass weight, maturity and ultimate pH Based on intramuscular fat, meat grades divide into M9-M0 (AUS-MEAT) MSA grades on a scale of 100 (no intramuscular fat) to 1190 (extreme amounts of intramuscular fat) |
M9- Extreme (1100+) M8- Excellent (1000-1100) M7- Very Good (900-1000) M6- Good (800-900) M5- Good (700-800) M4- Average (600-700) M3- Fair (500-600) M2- Poor (400-500) M1- Poor (300-400) M0- Poor (100-200) |
[22,23] |
| Canada | CBGA (Canadian Beef Grading Agency) |
Prime A (AAA, AA, A) B (B1-B4) D (D1-D4) E |
CBGA classified the beef, based on maturity, sex, conformation (muscling), fat and meat (color, texture and marbling). Youthful cattle may receive a quality grade of Canada Prime, AAA, AA, A or B1-B4, Carcasses from mature cattle may receive a grade of D1-D4 or E. Yield grades 1, 2, 3 and 4 estimates the percentage of the carcass that is saleable at retail. |
Prime-Superior (11.5%) AAA- Best (4.62%) AA -Better (3.11%) A -Good (2.92%) B1 - Better B2 - Good B3 - Good B4 - Fair |
[24] |
| Europe | EUROP grid | E. U. R. O. P | EUROP grid is used to classify a carcass according to its conformation (shape) and fat level Conformation is assessed on an E to P basis, with E being a convex and shapely carcass, R being an average shape or straight profile, and P being a plainer carcass with a concave profile. Fat is assessed on a 1 to 5 basis, with 1 being very lean and 5 being very fat. U, O and P are subdivided to give a high (+) or low (–) classification, and fat classes 4 and 5 are subdivided to low (L) and high (H). |
E- excellent U- Very Good R- Good O- Fair P- Poor |
[25,26] |
|
Korea |
KAPE (Korea Institute for Animal Products Quality Evaluation) |
Grade 1++ Grade 1+ Grade 1 Grade 2 Grade 3 |
South Korea has its own meat grading system. Beef meat is classified by 5 grades that include 1++, 1+, 1, 2, and 3, depending on the degree of marbling, meat color, fat color, firmness of rib eye and maturity. |
Grade 1++ - Superior (15.6-17 %) Grade 1+ - Excellent (12.3-15.5%) Grade1 - Good (9-12.2%) Grade 2 - Fair (5-9%) Grade 3 - Poor (0-5 %) |
[27] |
* Note – Europe meat grade intramuscular fat information not available.
Table 2.
Serum markers for beef meat evaluation and their functions in muscle development.
| Serum markers | Serum levels | Correlation | References |
|---|---|---|---|
| Retinol | 1.10 ± 0.012 IU/ mL |
High retinol content (> 1.10) poorly/negatively associated with meat marbling score. Retinol concentration positively correlates with meat color. |
[28] |
| Blood Urea Nitrogen (BUN) | 6.02 ± 0.6 mg/10 mL |
An indication of the cattle's protein energy status may be found in the blood urea nitrogen concentration (BUN) level. |
[33] |
|
Non-esterified fatty acid (NEFA) |
2.237 ± 0.12 μEq/10mL |
Palatability and intramuscular fat positively correlate with non-esterified fatty acid content. |
[40] |
| Total cholesterol | 128.03 ± 6.00 mg/mL |
The amount of total blood cholesterol was strongly/positively connected with carcass body weight & negatively correlated with marbling score. |
[43] |
| Paraoxonase 1(PON1) | 72.3 ± 35.6 mU/mL |
PON1 positively correlated with Marbling score, it is higher in female and castrated male than in male, indicating a gender-dependent difference. |
[51] |
| Insulin | 35.35 ± 2.37 ng/mL |
Meat marbling score and meat color are both favorably and adversely linked with insulin. |
[59] |
| Leptin | 30.34 ± 1.37 ng/mL |
Leptin have a negatively correlation with meat marbling score |
[59] |
| Aspartic acid transaminase (AST) | 0.54 ± 0.0324 U/10mL |
AST is positively connected with meat color and adversely correlated with the yield index and meat marbling score. |
[69] |
| Alanine transaminase (ALT) | 0.51 ± 0.043 U/10mL |
ALT has a positive correlation with the yield index and the meat marbling score and a negative correlation with the color of the meat. |
[68] |
| Total Protein (TP) | 1.1 ± 0.04 g/10 mL |
TP has a positive correlation with the yield index and the meat marbling score |
[44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



