Submitted:
22 March 2024
Posted:
25 March 2024
You are already at the latest version
Abstract
The effects of soluble silicon fertilization on monocots and dicots have been widely studied. However, little is known regarding its effects on protecting epiphytes against insect and fungal pests. The efficacy of silicon fertilizer to reduce damage by thrip pest complexes, namely: Thrips palmi Karny, Frankliniella occidentalis Pergande, Chaetanaphothrips orchidii Moulton, and Chaetanaphothrips signipennis Bagnall (Thysanoptera: Thripidae), and fungal pathogens: Botrytis cinerea Persoon (Helotiales: Sclerotiniaceae) and Fusarium spp. Link (Hypocreales: Nectriaceae) was examined during a nine-month greenhouse trial in Hawaii. The trial assessed yield, quality, and pest damage across three common varieties of dendrobiums. All replicates received additional soluble silicon fertilizer applications alternating weekly between soil drench and foliar (50 mg Si/plant) applications. Yield, quality, and spray length, pest damage, plant vigor, SPAD, and leaf temperature were measured. Data were analyzed using a generalized linear model (glm) with repeated measures followed by post-hoc pair-wise comparisons in R, version 4.3.1. Treatment effects were significant at P
Keywords:
Thrips palmi
; melon thrips
; silicon
; Fertilizer
; Nutrient Management
; Western flower thrips
; Frankliniella occidentalis
; Dendrobium
; Orchid
; Fusarium spp.
; Integrated Pest Management
1. Introduction
1.1. Dendrobium Production in Hawaii
Dendrobium (Dendrobium spp., (Swartz), Asperagales: Orchidaceae) production has a rich historical and cultural significance in Hawaii. Dendrobiums are commonly used in lei for celebrations, ceremonies, and hula. The island of Hawaii has many diverse climates which have led to an explosion of varied orchid types, species, and hybrids; thus, the majority of orchid production occurs on Hawaii island, and it has been endearingly nicknamed “The Orchid Isle”. The first statistical record of dendrobium production in Hawaii was published in 1969 by the Hawaii Department of Agriculture (HDOA), with 228,000 cut sprays sold for a wholesale value of $50,000 (Hara and Hata 1999). Historically dendrobium production was one of the fastest growing agricultural industries in Hawaii (Hara and Hata 1999). In 2020, cut and potted orchids were valued at $12.2 million in Hawaii and dendrobium orchid sales alone were valued at $3.16 million (USDA NASS 2021). This is a slight decrease from 2018 production values ($14 million for cut and potted orchids) due in large part to the 2018 Kilauea lava eruption and the negative impacts of the COVID-19 pandemic (Loke 2018, Eng et al. 2021, Kirk et al. 2021). Damage to farmers from the Fissure 8 lava flow totaled $27.9 million in farm losses, with $13.3 million in losses from floriculture and nursery industry, namely orchid production (Loke 2018). Orchid production in Hawaii is in the process of rebuilding from these events. Nationally, the industry continues to grow from 137 producers in 2018 to 175 producers in 2019 (USDA NASS 2020). Total orchid wholesale value for 2019 was $305 million nationally, and among indoor houseplants potted orchids account for one-third of the total wholesale value of all potted flowering crops in the U.S. (USDA NASS 2020).
1.2. Pests of Dendrobiums:
Hawaii orchid growers battle a wide range of insect and pathogen pests. The year-round growing season makes pest management especially challenging. Thrips are the most common insect pest of orchid flowers in Hawaii. There are over ten common thrips species and four fungal pathogens of dendrobium alone (Table 1). Thrips damage terminal bud growth leaving it stunted and deformed. At lower density levels damage can occur as scarring on petals (Sakimura et al. 1986). In severe infestations, thrips can damage every bloom produced in infested greenhouses (Hata and Hara 1992), and destroy newly formed sprays, rendering yield to essentially zero. Without proper management of thrips, dendrobium production would not remain profitable. Surveys conducted by the University of Hawaii and HDOA have identified western flower thrips (Frankliniella occidentalis Pergande (Thysanoptera: Thripidae)) and melon thrips (Thrips palmi Karny (Thysanoptera: Thripidae)) as the two most prevalent species of thrips infesting dendrobium (Hata et al. 1993). These two invasive species were introduced to Hawaii in 1955 and 1982 respectively (Waterhouse and Norris 1989, Johnson 1986). Populations can grow rapidly as they have a relatively short life cycle and can reproduce both sexually and asexually. Hawaii ‘s tropical climates allow pest populations to remain at damaging thresholds (Gill et al. 2012) year-round as temperatures do not significantly drop during winter months (Tsai et, al. 1995), highlighting the importance of sustainable pest management strategies. Both species of thrips are polyphagous, and their host range includes at least 50 different plant species (CABI 2023, Wang and Chu 1986, Immaraju et al. 1992, Johnson 1986, Mau and Kessing 1993). Both species are vectors for several economically significant plant viruses, including at least six known tospoviruses specifically transmitted by T. palmi (Nagata et al. 2002). T. palmi is still classified as a federal quarantine action pest which means there is a zero-tolerance level for orchids exported from Hawaii to the continental United States and other un-infested areas around the world. Entire flower shipments are rejected by inspection officials as a result of T. palmi and F. occidentalis infestations (Nakahara 1985, Gardner 1991). Due to these strict export regulations dendrobium growers employ significant efforts to control pests. The international economic importance of T. palmi and F. occidentalis necessitates the development of sustainable management.
Other economically important pests include a range of fungal pathogens (Table 1). Major economically important pathogens of dendrobium orchids include gray mold (Botrytis cinerea (Persoon) Helotiales: Sclerotiniaceae) and fusarium (Fusarium spp. (Link) Hypocreales: Nectriaceae) (Ito and Aragaki 1977, Hara and Hata 1999, Srivastava et al. 2018). Gray mold causes dark necrotic flecks on flowers, although other fungal species also incite necrotic flecking that resemble gray mold infections (Ito and Aragaki 1977, Uchida and Aragaki, 1979). Fusarium has been reported as a pathogen in dendrobium in Hawaii and Asia, and causes necrotic leaf spots that can become severe blights. Fusarium also incites stem, pseudobulb, and root rot symptoms (Srivastava et al. 2018). Humidity and airflow play a large role in fungal pathogens in dendrobium production and is the greatest environmental factor in promoting fungal problems for both the potted and cut dendrobium industry in Hawaii.
1.3. Current Management Challenges:
Dendrobium growers face a multitude of issues on the farm including an ever-shorter list of chemical control options for pests and a growing demand in the consumer marketplace for reduced pesticide use. Growers understand that the sole use of chemicals to control insect pests is not a sustainable long-term solution (Seal et al. 2013). Currently, growers are battling issues with pesticide resistant populations that have the ability to decimate entire nurseries, this includes: T. palmi, F. occidentalis, Chaetanaphothrips orchidii (Moulton) (anthurium thrips), and Chaetanaphothrips signipennis (Bagnall) (banana rust thrips). Thrips are especially difficult to control due to their short life cycle, parthenogenetic reproduction, and their preference of hiding deep inside blossom and petals, making contact with pesticides challenging (Hata et al. 1991). Additionally, thrips pupae evade treatment as they are usually found in the soil and growing media, requiring carefully timed insecticide applications occurring in succession (Hollingsworth et al. 2001). Thus, growers have come to heavily rely on a short list of systemic insecticides.
Fusarium spp. can also be difficult to control if control measures are implemented too late in the disease cycle. Many available fungicides are prophylactic and there remain few therapeutic chemical options registered in Hawaii. Although growers often employ integrated tactics to control thrips and fungal pathogens, they rely heavily on the use of chemical pesticides.
In 2000, a growers’ perception survey, conducted by the College of Tropical Agriculture and Human Resources (CTAHR) at the University of Hawaii at Manoa, found 23.8% of growers reported that the average number of pesticide applications (excluding fungicides) made per year for growers of cut and potted orchids was 25.8 and 29.3 applications/year respectively (Hollingsworth et al. 2000). Organophosphates and pyrethroids were the most common types of pesticides applied (Hollingsworth et al. 2000). These high application rates reflect the industry’s reliance on costly chemical options.
The cost of agricultural inputs in Hawaii is further exacerbated by increases in shipping costs, most notably during the summer of 2020 due to the economic strain of the COVID-19 pandemic. The Public Utilities Commission approved a 46% rate increase for Hawaii’s only regulated inter-island cargo shipping company, Young Brothers (Ancheta 2020).
Grower dependency on chemical pest control in dendrobium production in Hawaii generated the need for sustainable and economical alternative management options. Soluble silicon (Si) fertilizer is one input that has received attention for its potential use in controlling both biotic and abiotic stressors. A wealth of scientific evidence demonstrating the benefits of Si against a range of various environmental stresses has been established over the past several decades (Reynolds et al. 2009), however, little research has been conducted on the effects of Si on epiphytic plants such as dendrobium.
1.4. Silicon-Mediated Resistance
Si is a non-essential nutrient that is observed in substantial concentrations in plant tissues, and is linked to plant defense against insects and pathogens (Fateux et al. 2005, Alhousari and Greger 2018). Silicon accumulation varies in plants and plants have previously been categorized by the percent of silicon in dry shoot weight of a plant (Jones and Handreck 1967). Si content can vary based on plant species and is also dependent on application method, soil properties, Si source and Si amount. Some orchid species were reported to absorb and show increased tissue Si concentrations in response to increased Si fertilization (Vendrame et al. 2010) and Phaelanopsis orchids are considered Si accumulators (Mantovani et al. 2018). With the discovery of Si transporters, it has been suggested that Si accumulation should be categorized by the presence or absence of aquaporins (Ma et al. 2006). No work was found on the presence of aquaporins in dendrobium.
The understanding of how Si confers beneficial effects against insect pests and fungal pathogens has evolved over the last several years. Si was originally thought to provide primarily a physical defense, called the mechanical barrier hypothesis (Wagner 1940). More recently, the mechanical barrier hypothesis has been challenged, and other mechanisms have been suggested, including defense priming (Samuels et al. 1991, Rodrigues et al. 2004, Liang et al. 2015, Kvedaras et al. 2010), effector-triggered immunity (Hogenhout and Bos 2011), and the apoplastic obstruction hypothesis (Coskun et al. 2019).
While there is limited available literature on Si effects in dendrobium (Mantovani et al. 2018), the effects on thrips and fungi have been studied in other plant species. For example, Si has been shown to induce defense related enzymes (Cai et al. 2008), induce photochemical efficiency and adjust mineral uptake (Gao et al. 2011) in rice (Oryza sativa (L.) Poales: Poaceae) that help resist rice blast disease (Magnaporthe oryzae). Silicon was also found to positively affect sorghum (Sorghum bicolor) infected with anthracnose (Colletotrichum sublineolum) (Resende et al. 2012). Additionally, Si has been proven to suppress Fusarium spp. in banana (Musa acuminata), tomato (Solanum lycopersicum), and potato (Solanum tuberosum) (Li et al. 2009, Huang et al. 2011, Fortunato et al. 2012). Not all studies showed efficacy against fungal pathogens in plants, especially necrotrophs (Coskun et al. 2019), such as Botrytis. Previous studies have also examined thrips management using Si treatments with varied results.
Several studies found that Si applications led to decreased thrips counts and lesions due to increased mortality of nymphs in tomato (Almeida et al. 2009) and eggplant (Solanum melogena) (Almeida et al. 2008). Similarly, reductions in thrips populations occurred after treatment with Si in groundnut (Arachis hypogea) (Parthiban et al. 2018). However, in pepper plants (Capsicum annuum), researchers found that Si applications had no significant effect on thrips populations (Dogramaci et al. 2013). Likewise, Si applications did not affect thrips populations in sugar cane (Saccharum spp.), but did affect stalk borer (Eldana saccharina), indicating that Si-mediated resistance may be more developed in different plant tissues (Keeping and Sewpersad 2014). Mantovani et al., 2018 found net positive benefit of Si in dendrobium growth, with concentrations of 27 and 16 mmol L-1 from potassium silicate and monosilicic acid sources respectively. To date, no studies have been published exploring the effects of Si against thrips pests and fungal pathogens in dendrobium production.
1.5. Objectives
The purpose of this study was to evaluate the effects of soluble silicon dioxide (SiO2) on naturally occurring thrips and fungus populations on three popular dendrobium cultivars (‘Uniwai Mist’, ‘Uniwai Supreme’, and ‘Uniwai Royale’) in a greenhouse production setting. The nature of the experimental design allowed for the evaluation of the short and long-term effects of SiO2 on the two most common Dendrobium pests (thrips and fungus) in Hawaii, and examine potential differences in response to Si in three popular varieties.
2. Materials and Methods
2.1. Experiment Design:
An experiment investigating the effects of soluble silicon fertilizer on three popular varieties of dendrobium orchids was conducted at Waiakea Research Station (19.644, -155.080), Hilo, Hawaii from August 1, 2020 to May 30, 2021 in a greenhouse (Conely’s Greenhouse Manufacturing and Sales, Montclair, CA), with 30% shade. Treatments were arranged in a split plot design with main plots arranged in an RCBD, consisting of a negative control versus soluble silicon dioxide fertilizer (Mainstay Si®, Redox (SiO2)) and three popular cultivars as sub plots, totaling 6 blocks. Subplots consisted of three varieties of dendrobiums: white (Dendrobium cv ‘Uniwai Mist’), lavender (Dendrobium cv ‘Uniwai Supreme’), and purple (Dendrobium cv ‘Uniwai Royale’). Dendrobium orchids were 3 years mature at the onset of the experiment. Si was applied at a rate slightly higher than the recommended drench label rate of 3.27 gal/acre weekly, switching between foliar and drench applications. This rate was calculated to deliver 50 mg Si to each plant at each application (Dr. Wendy Zellner, 2020, Personal Communication). We applied the fertilizer solution using a one-liter hand pump sprayer with TeeJet® flat fan nozzle, size 8004, for foliar applications, and applying 200 ml of solution to the root zone for drench applications via 250 ml beaker vessel by hand. Overhead sprinklers irrigated plants for 3 minutes twice daily, using municipal water, and RainBird® Micro-Quick 0.038 inch diameter sprinkler nozzle.
Dendrobiums were grown in blue rock medium in plastic bags. All experimental units received Nutricote® 14-14-14 (American Horticultural Supply, Inc ©, Oxnard, CA), controlled-release fertilizer at a rate of 300lb/acre/year at the onset of the trial and 6 months into the trial, in keeping with conventional cultivation practices of dendrobium in Hawaii (Hara and Hata 1999). We relied on naturally occurring insect and fungal populations in the field. No pesticides were applied throughout the duration of the trial.
2.2. Data Collection:
Data on several dependent variables were collected throughout the trial and can be categorized into the three categories outlined below.
Harvest and quality: Sprays that had reached 67-75% maturity (meaning two thirds to three fourths of their lower blooms had fully opened and the top remaining 25-30% buds were unopened) were harvested every other week and yield and quality were assessed during the time of harvest (Hara and Hata, 1999). Quality was characterized by: (i) percent damage to sprays, stems, blooms and buds, and (ii) length of spray (cm). Sprays with less than 80% damage to blooms and stem were deemed salable. The remaining sprays were considered unmarketable and were considered “throw aways”. Harvest and quality data was collected for nine months between September 9, 2020, to May 15, 2021. Market prices are higher for longer sprays and an important consideration for grower profits.
Insect and Disease Monitoring: Incidence of pest and disease was monitored weekly for five months between September 25, 2020 to February 25, 2021. Proportional thrips damage to blooms, sprays, buds, and stems was assessed. Scarring on flower sepals, petals, buds, and stems, as well as deformed and arrested spray development due to thrips damage was quantified on a percent damage scale from 0-100. No attempt to attribute damage to specific species was made as thrips damage on dendrobium presents similarly across all species. Additionally, pest counts were collected at the same time, however, thrips count is a less precise measure of thrips populations in the field, as many are small and hide in tight places of the orchid making them difficult to observe. To avoid disturbing the natural thrips populations in the greenhouse, we minimized the sampling through thrips removal. However, thrips were identified to species from representative field samples every 2 months and the populations consisted of T. palmi, F. occidentalis, C. signipennis, and C. orchidii.
Additionally, other insects were recorded through direct visual observation of both beneficial and pest species. Sampling was standardized by allowing observation to occur over a 30 second interval per plant and only one person collected this data for the duration of the trial.
Fungal pathogens were characterized by percent area of leaf affected by fungal legions, necrotic or blackened leaf spots, or percent area affected dark necrotic flecks on the flowers. Fusarium sp. and Botrytis spp. were the only two fungal pathogens observed throughout the trial. Their symptoms were very distinct from each other and easily identified in the field. In addition, samples were collected, and pathogens identified in lab via microplate method.
From these measurements we were able to analyze total disease, total thrips damage, and total beneficials in the analysis.
Plant health and growth: Data were collected weekly on number of new canes, number of new sprays for nearly seven months, between July 1, 2020, and January 22, 2021. Data were collection began one month prior to treatment applications to determine baseline plant vigor measurements. Soil Plant Analysis Development (SPAD) chlorophyll meter, and leaf temp data was collected every 14 days between August 30, 2020, to October 2, 2020. SPAD index was measured (portable chlorophyll meter; SPAD-502, Minolta, Japan) as it is one of the most commonly used diagnostic tools to measure crop nitrogen status in the field (Chubachi et al. 1986). SPAD measurements were taken at a position two thirds from leaf base on the newest mature leaf of the tallest cane. A total of three SPAD measurements per leaf were recorded and averaged (Paris-Felipo et al. 2020). Leaf temperature was measured using a Klein® dual laser infrared thermometer at an optimal distance of 12 inches from the newest mature leaf. Three measurements per leaf were recorded and averaged. Leaf temperature was considered because it has been recognized as important for plant function and influences photosynthesis, respiration, and transpiration (He et al. 1998).
Vase Life: Harvested flowers were transported to Komohana Research and Extension Center (19.696, -155.090), Hilo, Hawaii, where the stems were cut, and flowers were transferred to vases which were filled with room temperature distilled water. The number of individual flowers on each sample inflorescence was counted. Flowers were kept in vases at an ambient temperature of 22.2 degrees Celsius. Every five days vase water was replaced, and number of individual flowers was counted on each sample inflorescence. Once the number of flowers reached half or less than half of the original number of flowers, that sample was discarded. Days to discard following harvest was recorded as the vase life measurement. Vase life data were collected later in the trial from December 1, 2020, to June 1, 2021.
Silicon Accumulation: Six months into the experiment on December 16, 2020, we collected five mature leaves from each experimental unit, and dried leaves at 60 degrees Celsius for 72 hours in a Despatch® model LBB2-27-1, 4800-watt oven, after which we sent them to the Agricultural Diagnostic Service Center (ADSC) at University of Hawaii for analysis. Silicon content was determined gravimetrically in the plant tissue as the residue after acid digestion (Snyder 2001).
Analysis: All data were analyzed using a combination of Analysis of Variance (ANOVA) for normally distributed data sets and Generalized Linear Model (GLM) specifying Poisson distribution for count data using the glmm package in R, version 4.3.1. Post-hoc pairwise comparisons were performed using Tukey’s HSD following ANOVA analysis and estimated marginal means (emmeans + contrast) following GLM in R.
3. Results
Results indicated significant benefit of soluble silicon fertilizer (P<0.001) among insect and disease, spray length, and marketable yield.
3.1. Harvest
Spray length at harvest: This experiment demonstrated that soluble silicon fertilizer had a significant influence on spray length at harvest (F value = 69.208; df = 1, 189, P<0.001) (Figure 1). Variety factor was also significant (F=7.272; df = 2, 189, P<0.001) with silicon treated white having the longest sprays (M = 53.4 cm ± 2.8), and was significantly longer than the silicon treated purple variety, which had the shortest sprays (M = 42.3 cm ± 2.1) among the treated varieties, neither being significantly different from the treated lavender in terms of spray length (Figure 1). However, the lavender variety demonstrated the largest benefit of Si in terms of spray length, with an average increase spray length of of 22.9 cm ± 2.9 and purple benefiting the least in spray length from silicon fertilizer, with an average increase spray length of 16.9 cm ± 2.3 (Figure 1).
Thrip damage on harvested sprays: The amount of thrips damage observed at harvest was statistically significant between silicon treated and untreated control plants irrespective of variety (F = 64.5; df = 1, 189, P< 0.001). We observed the greatest amount of thrips damage in the control lavender variety and the least amount of thrips damage to the Si treated lavender variety. Again, demonstrating that the lavender variety benefited the greatest from the silicon treatment, while the purple variety benefited the least. The average difference in percent thrip damage in Si versus control treated plants was 54.1% ± 8.3, and 40.6% ± 5.8, lavender and purple varieties respectively (Figure 2).
Marketable yield: The degree of thrip damage observed ocularly translated to marketable yield, and the same trend held true. Silicon effects on marketable yield were significant (F = 63.977; df = 1, 189, P<0.001) while the variety term was not significant at α = 0.05. Lavender again benefited the greatest with 55.3% increase in marketable yield, compared with white which had an increase of 43.1% in marketable sprays, and lastly purple with the 42.7% increase in marketable sprays (Figure 3).
3.2. Insect and Disease Monitoring
Thrip damage: Silicon fertilizer significantly lowered thrips damage (F = 147.223; df = 1, 175, P<0.001) across all varieties. Variety was not a significant factor. However, date played a significant role (F = 31.677; df = 16, 175, P<0.001) (Figure 4, Figure 5 and Figure 6). Differences among treatments were observed between six and eight weeks, becoming significant (α = 0.05) between eight and twelve weeks post initial silicon applications.
Fungal Damage: The degree of infestation by B. cinerea. was minor over the length of the study. For this reason, we combined all fungal data into one data set. Significant differences were evidenced among total disease observed throughout the life of the trial among treatment and variety factors (F = 88.341, df = 1, 188, P<0.001; F = 3.469, df = 2, 188, P=0.0322 respectively). We observed the greatest fungal infections on the untreated control lavender variety (M= 9.6% ± 1.5), which was significantly different from the untreated white variety, which exhibited the least amount of fungal pathogen damage; neither significantly different from the untreated purple variety (Figure 7). The silicon treated white variety had the lowest infestation rate (M= 0.367% ± 0.13). Varieties with the greatest fungal damage benefited the most from the use of silicon (Figure 7).
3.3. Plant Growth and Health
Plant Vigor: Plant vigor was determined by the sum of total sprays after seven months of collecting data weekly. There were no significant differences (α = 0.05) between silicon treated and untreated plants in terms of the number of sprays produced, although the total number of sprays produced was significant among varieties (F=22.977, df = 2, 189, P<0.001). Purple was the highest yielding variety, followed by white and lavender (Table 2). The purple variety was the greatest producing cultivar but overall benefited the least from Si applications, while the lavender variety was the lowest producing variety and overall benefited the most from Si applications (Figure 1, Figure 2, Figure 3 and Figure 7).
SPAD and Temp measurements: There was no significant difference (α = 0.05) between treatment effects on SPAD readings (Figure 8), however, there were significant differences among the varieties in SPAD readings (F = 14.797, df = 2, 174, P<0.001). The white variety consistently had the higher SPAD measurements, and the lavender variety consistently had the lowest reading. Leaf temperature was not significant (α = 0.05) between treatments, nor among varieties. It is important to note, that leaf temperature between treated and control plants was marginally significant at (α = 0.10, F = 2.91, df = 1, 174, P = 0.0897), and the silicon treated plants had consistently lower leaf temperature for the majority of the five weeks we collected data, however, that difference ranged from 0.5-1 C◦ depending on variety (Figure 9). We see greater temperature differences among treatment groups at higher ambient temperatures and little to no difference among treatments below 23.5 degrees Celsius.
3.4. Vase Life
Formal ANOVA could not be conducted due to the unbalanced nature of vase life data. When we began collecting vase life data four months into the trial, there were already high thrip densities in the greenhouse and untreated control varieties had very low yield, thus there were over 6.5 times the yield of silicon treated sprays versus control. On average silicon treated sprays exhibited a longer vase life, days to discard, (M = 25 days ± 7.2) than their control counterparts (M = 13 days ± 8.8), with no differences observed among varieties. Further research is needed to explore the potential benefit of Si on improving vase life.
3.5. Silicon Accumulation
All silicon treated plants had slightly higher silicon accumulation (ug/g) but none significantly higher than their untreated counterparts (Figure 10).
4. Discussion
The benefits of soluble silicon fertilization in dendrobium production in Hawaii are evident throughout the results of this trial. On average, irrespective of variety, Si treated dendrobium exhibited 53.4% less damage from thrips, and a 91% reduction in pathogen infection, which translated to a varietal yield increases of 43.1-55.3%. These dramatic benefits were observed even though there was no significant increase in Si accumulation in treated versus untreated plants. The varieties that received the greatest benefit from Si (Figure 1, Figure 2, Figure 3 and Figure 7) exhibited the least Si accumulation within its plant tissues (lavender variety, Figure 10). Thus, challenging our conventional understanding of the mechanisms of Si regarding crop protection.
The mechanisms by which Si provides protection against arthropod pests and disease is still largely unknown and varies by system. Historically, Si was believed to offer plant protection mainly by acting as a physical barrier to reduce insect feeding and deter disease infections. Silicon has been demonstrated to enhance phytoliths within plant tissues and change the shape and size of trichome bases on leaves (Frantz et al. 2008). Both aid in deterring arthropod feeding by herbivorous pests. One of the original theories developed from evidence that Si thickened the plant cell walls making it more difficult for insects with piercing-sucking mouthparts to penetrate plant tissue and/or wearing down the mandibles of chewing insects, such as caterpillars. However, our Si accumulation data indicate that Si may work more as a cofactor or inducing factor that has a cascading effect and potentially changes gene expression and/or plant cell architecture (among other things) to provide plant protection, rather than on accumulation alone. Additionally, the dynamics of Si accumulation or sensitivity may vary in different tissues (Dr. Jon Suzuki 2023 personal communication).
Recent evidence suggests that Si plays a crucial role in the Systemic Acquired Resistance (SAR) of plants. SAR is understood to confer long-lasting protection against a broad spectrum of microorganisms. There is research to support the theory that indirect defenses by Si are mediated by herbivore-induced plant volatiles (HIPVs) released in response to insect feeding (Alhousari and Greger 2018). These HIPVs, modulated by the Jasmonic acid (JA) pathway, promote biological control by attracting predators and parasitoids of insect pests. Therefore, understanding the role of Si as part of both a physical barrier and as part of an induced-defense system is vital to knowing how to incorporate it as part of a larger IPM program.
Our results show the promise of incorporating Si as part of an IPM program for dendrobium production in Hawaii and possibly other epiphytic cropping systems such as Anthuriums and Bromeliads. Incorporation of Si can be considered a “Bottom-Up” tactic, defined as part of ecologically based methods that impact the tri-trophic interactions among crop, pest, and predator within agroecosystems to optimize IPM. Bottom-up management tactics often rely on strengthening natural plant defenses through nutrient management. Slow growing plants, such as dendrobiums, require adequate time for the uptake and metabolism of fertilizers and other plant protection inputs. Understanding the effects of Si on invasive thrip pests is critical to the development and understanding of its complete benefits, offering a greater understanding of the biological trade-offs and practical insights into the relationships between plant nutrition, plant stress responses, pests, and diseases.
4.1. Role of Si in an IPM Program
There are many possibilities for successfully incorporating Si into an existing IPM program. These possibilities include: adding Si to media/soil as a supplement to growers’ current fertilizer regime, adding soluble Si to tank mixes during pesticide applications, incorporating soluble Si in irrigation or fertigation systems during times of high pest pressure or likely abiotic stress. There is also the potential to incorporate Si applications into a pesticide resistance management program since it may offer plant protection with little to no selection pressure. To potentially utilize Si as part of a pesticide resistance management program there are many questions that require further examination. The difficulty being the answers to these questions may differ by cropping system.
For Si to function as an effective part of a pesticide resistance management program we would need to understand the following basic questions: (1) Does Si reduce pest population densities? (2) Does Si alter pest host selection? (3) Does Si alter plant tissues to make it a less nutritious host for herbivore pests? (4) Does Si make it more difficult for pests to penetrate plant tissue? (5) Does Si change the chemical ecology, specifically HIPVs, of the plant? (6) Are none of these factors altered (population density, host selection, nutrition, rigidity, and cell wall thickness) and damage is simply less visible on plants treated with Si?
Applied questions regarding application rates, methods (foliar versus drench), timing of application, and use in conjunction with pesticides also need to be considered when developing Si as a practical IPM tool. However, our results indicate that Si has the potential to positively impact the production of dendrobium, and possibly other epiphytic crops, in Hawaii.
4.2. Where and When Si Shines
Throughout the trial we observed that some varieties benefited more from Si than others. Although this difference was not statistically significant, this observed variation could be due to various factors. In our trial the lavender variety experienced the most damage by thrips and fungal diseases, and also the greatest yield benefit from Si against both pests (Figure 2, Figure 3 and Figure 7). We believe the degree of benefit of Si is correlated to the degree of the stressor. It could also be that plant protection provided by Si is variety dependent and depends on the presence of certain Si transporter genes, along with other factors to allow for the uptake and transport of Si. The fact that Si has been shown to help with a variety of abiotic and biotic stressors in the field, including: insect pests, pathogens, heat and water stress etc., (Mantovani et al. 2018, Fortunato et al. 2012, Resende et al. 2012, Gao et al. 2011, Kvedaras et al. 2010, Alemeida et al. 2009, Li et al. 2009, and Fateux et al. 2005) and that the benefits of Si have been variable across pest species, pest complexes, and cultivation systems (Dogramaci et al. 2013, and Keeping and Sewpersad 2014) muddies not only our understanding of Si, but also how to best incorporate it into any IPM system. Within floriculture and nursery industries, the diversity of plant species and cultivars within species is vast and it is common, due to traditional breeding, for some cultivars to be highly resistant to certain pests, disease, and other abiotic factors, and susceptible to other like factors. It may be that Si will most benefit each cultivar in the area in which it is most susceptible. In this scenario, researchers must evaluate each cultivar’s susceptibility to any given pest, and collect data on the proper parameters to capture the potential benefit of Si.
Additionally, timing and length of application played an important role in the success of this trial. We didn’t begin to see beneficial effects of Si until weeks 10-12. If we had concluded the trail after 2 months, and were only interested in short-term effects, we wouldn’t have necessarily seen positive results and missed the delayed benefits of Si. In the case of this trial, Si had a cumulative effect over time providing plant protection against four common thrip pests and two classes of common fungal pathogens. However, we observed a diminishing return on investment after 5 months of weekly applications depending on the variety (Figure 4, Figure 5 and Figure 6). No pesticides were applied during this trial and future research should examine the use of Si in conjunction with pesticides and its possible value as supplement to pesticide programs.
5. Summary
While it is clear dendrobium orchids benefit from Si applications, the timing, rate, application method, frequency of application and context in which it is utilized will require further development by researchers. The variability of Si to confer benefits against various pest and disease across different cultivars further compounds the difficulty in determining proper application factors described above. Understanding the short-term and long-term effects of Si and its ability to be used prophylactically and/or therapeutically will give us a better understanding of how to sustainably utilize this element in various stages of plant growth and cropping systems.
Author Contributions
Conceptualization, J.B. Bloese, Methodology, J.B. Bloese & R. Galanti, Validation, J. B. Bloese, R. Galanti, T. Kow, and R. Porter, Formal Analysis, J.B. Bloese, Investigation, J.B. Bloese, Resources, J.B. Bloese, R. Galanti, and R. Porter, Data curation, J.B. Bloese, R. Porter, and T. Kow, Writing- Original Draft Preparation, J.B. Bloese, and R. Galanti, Writing- Review and Editing, J.B.Bloese and R. Galanti, Visualization, J.B.Bloese, Supervision, J.B.Bloese, Project Administration, J.B. Bloese, Funding Acquisition, J.B. Bloese and R. Galanti.
Funding
This research was funded by USDA Animal and Plant Health Inspection Services (APHIS), Plant Protection and Quarantine (PPQ) grant project: Best Management Practices for Plant Quarantine Security of Hawaii’s Tropical Flowers and Foliage, agreement number AP19PPQFO000C484.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors thank Dr. Wendy Zellner, research faculty at the University of Toronto, for her valuable assistance in determining application rate and method. Dr. Jon Suzuki biologist at United States Department of Agriculture, Agricultural Research Services (USDA ARS) is acknowledged for his insights in the discussion regarding silicon accumulation and gene expression. The authors would also like to thank Cindy McCarty for her help with data entry and Paul Bloese for reviewing the manuscript for organization, flow, and clarity.
Conflicts of Interest
The authors declare that this research was performed in the absence of any financial or commercial relationships that could be construed as a potential conflict of interest.
References
- Alhousari, F.; Greger, M. Silicon and Mechanisms of Plant Resistance to Insect Pests. Plants 2018, 7, 33. [Google Scholar] [CrossRef]
- Almeida, G.D.; Pratissoli, D.; Zanuncio, J.C.; Vicentini, V.B.; Holtz, A.M.; Serrão, J.C. Calcium silicate and organic mineral fertilizer applications reduce phytophagy by Thrips palmi Karny (Thysanoptera: Thripidae) on eggplants (Solanum melongena L. ). Interciencia 2008, 33, 835–838. [Google Scholar]
- Almeida, G.D.; Pratissoli, D.; Zanuncio, J.C.; Vicentini, V.B.; Holtz, A.M.; Serrão, J.E. Calcium silicate and organic mineral fertilizer increase the resistance of tomato plants to Frankliniella schultzei. Phytoparasitica 2009, 37, 225–230. [Google Scholar] [CrossRef]
- Ancheta D. 2020. Neighbor Islanders brace for higher prices after Young Brothers gets 46% rate increase. Hawaii News Now. Published: Aug. 18, 2020 at 10:07 AM HST. Updated: Aug. 18, 2020 at 10:09 AM HST. Retrieved June 2022, from https://www.hawaiinewsnow.com/2020/08/17/puc-approved-emergency-percent-rate-increase-young-brothers/.
- CABI. 2023. Thrips palmi. In: Invasive Species Compendium. Wallingford, UK: CAB International. Retrieved from https://www.cabi.org/isc/datasheet/53745#todistribution.
- Cai, K.; Gao, D.; Luo, S.; Zeng, R.; Yang, J.; Zhu, X. Physiological and cytological mechanisms of silicon-induced resistance in rice against blast disease. Physiol. Plant 2008, 134, 324–333. [Google Scholar] [CrossRef]
- Chubachi, T.; Asano, I.; Oikawa, T. The diagnosis of nitrogen nutrition of rice plants using chlorophyll meter. Soil Sci. Plant Nutr. 1986, 57, 190–193. [Google Scholar]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Belanger, R.R. The Controversies of silicon’s role in plant biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef]
- Dogramaci, M.; Arthurs, S.P.; Chen, J.; Osborne, L. Silicon Applications Have Minimal Effects on Scirtothrips dorsalis (Thysanoptera: Thripidae) Populations on Pepper Plant, Capsicum annum L. Fla. Entomol. 2013, 96, 48–54. [Google Scholar] [CrossRef]
- Eng, S.; Khun, T.; Esquivel, M.; Ooki, N.; Bloese, J.; Sand, S.; Lincoln, N. Farmers’ Perceived Needs of Extension’ Support During Covid-19 in Hawai’i. Journal of Extension 2021, 59, 15. [Google Scholar] [CrossRef]
- Fateux, F.; Remus-Borel, W.; Menzies, J.G.; Belanger, R.R. Silicon and plant disease resistance against pathogenic fungi. FEMS Microbiol. Lett. 2005, 249, 1–6. [Google Scholar] [CrossRef]
- Fortunato, A.A.; Rodrigues, F.A.; Baroni, J.C.P.; Soares, G.C.B.; Rodriguez, M.A.D.; Pereira, O.L. Silicon suppresses Fusarium wilt development in banana plants. J. Phytopathol. 2012, 160, 674–679. [Google Scholar] [CrossRef]
- Frantz, J.M.; Locke, J.C.; Datnoff, L.; Omer, M.; Widrig, A.; Strutz, D.; Horst, L.; Krause, C.R. Detection, distribution, and quantification of silicon in floriculture crops utilizing three distinct analytical methods. Communications in Soil Science and Plant Analysis 2008, 39, 2734–2751. [Google Scholar] [CrossRef]
- Gao, D.; Cai, K.; Chen, J.; Luo, S.; Zeng, R.; Yang, J.; Zhu, X. Silicon enhances photochemical efficiency and adjusts mineral nutrient absorption in Magnaporthe oryzae infected rice plants. Acta Physiol. Plant. 2011, 33, 675–682. [Google Scholar] [CrossRef]
- Gardener, W.D. 1991. Pest-related flower shipment rejections, pp. 49–51. In K. W. Leonhardt, D.O. Evans and J.M. Halloran [eds.], The Hawaii Tropical Cut Flower Industry Conference, 29-31 March 1990, Hilo, Hawaii, College of Tropical Agriculture and Human Resources, Honolulu.
- Gill, S.; Dutky, E.; Raupp, M.; Davidson, J.; Nakahara, S. 2012. Thrips Management in Greenhouses. University of Maryland Extension, FS-762-2012.
- Hara, A.H.; Hata, T.Y. 1999. Pests and pest management. Pg. 29-45. In: K. Leonhardt, and K. Sewake [eds.], Growing dendrobium orchids in Hawaii: Production and management guide. University of Hawaii, Honolulu, HI.
- Hata, T.Y.; Hara, A.H.; Hansen, J.D. Feeding preference of melon thrips on orchids in Hawaii. Hortsci. 1991, 26, 1294–1295. [Google Scholar] [CrossRef]
- Hata, T.Y.; Hara, H.A. Anthurium thrips, Chaetanaphothrips orchidii (Moulton): Biology and insecticidal control on Hawaiian anthuriums. Trop. Pest Manag. 1992, 38, 230–233. [Google Scholar] [CrossRef]
- Hata, T.Y.; Hara, A.H.; Hu, B.K.S.; Kaneko, R.T.; Tenbrink, V.L. Field sprays and insecticidal dips after harvest for pest management of Frankliniella occidentalis and Thrips palmi (Thysanoptera: Thripidae) on orchids. Journal of Economic Entomology 1993, 86, 1483–1789. [Google Scholar] [CrossRef]
- He, J.; Khoo, G.H.; Hew, C.S. Susceptibility of CAM Dendrobium leaves and flowers to high light and high temperature under natural tropical conditions. Environmental and Experimental Botany 1998, 40, 255–264. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Bos, J.I.B. Effector proteins that modulate plant–insect interactions. Curr. Opin. in Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Hollingsworth, R.G.; Hara, A.H.; Sewake, K.T. Pesticide use and grower perceptions of pest problems on ornamental crops in Hawaii. J. Ext. 2000, 38. [Google Scholar]
- Hollingsworth, R.G.; Hara, A.H.; Sewake, K.T. Scouting for thrips in orchid plants. CTAHR Publication IP-3, College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa. 2001.
- Huang, C.H.; Roberts, P.D.; Datnoff, L.E. Silicon suppresses Fusarium crown and root rot of tomato. J. Phytopathol. 2011, 159, 546–554. [Google Scholar] [CrossRef]
- Immaraju, J.; Paine, T.D.; Bethke, J.A. Western Flower Thrips (Thysanoptera: Thripidae) Resistance to Insecticides in Coastal California Greenhouses. J. Econ. Entomology. 1992, 85, 9–14. [Google Scholar] [CrossRef]
- Ito, J.S.; Aragaki, M. Botrytis blossom blight of Dendrobium. Phytopathology 1977, 67, 820–824. [Google Scholar] [CrossRef]
- Jana, S.; Jeong, B.R. Silicon: The most under-appreciated element in horticulture crops. Trends Hortic. Res. 2014, 4, 1–19. [Google Scholar]
- Johnson, M. Population trends of a newly introduced species, Thrips palmi (Thysanoptera: Thripsidae), on commercial watermelon plantings in Hawai’i. J. Econ. Entomology. 1986, 79, 718–720. [Google Scholar] [CrossRef]
- Jones, L.H.P.; Handreck, K.A. Silica in soils, plants, and animals. Adv. Agron. 1967, 19, 107–149. [Google Scholar]
- Keeping, M.G.; Miles, N.; Sewpersad, C. Silicon reduces impact of plant nitrogen in promoting stalk borer (Eldana saccharina) but not sugarcane thrips (Fulmekiola serrata) infestations in sugarcane. Front. Plant. Sci. 2014, 4, 289. [Google Scholar] [CrossRef] [PubMed]
- Kirk, E.; Sand, S.; Bloese, J.; Gutierrez-Coarite, R.; Keach, J.; Eng, S. 2021. COVID-19 Hawaii agriculture survey: Initial and on-going impacts. Cooperative Extension. College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa.
- Kvedaras, O.L.; An, M.; Choi, Y.S.; Gurr, G.M. Silicon enhances natural enemy attraction and biological control through induced plant defences. Bull. Entomol. Res. 2010, 100, 367–371. [Google Scholar] [CrossRef]
- Li, Y.; Bi, Y.; Ge, Y.; Sun, X.; Wang, Y. Antifungal activity of sodium silicate on Fusarium sulphureum and its effect on dry rot of potato tubers. J. Food Sci. 2009, 74, 213–218. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Belanger, R.R.; Gong, H.; Song, A. 2015. Silicon in agriculture: From theory to practice. Dordrecht, the Netherlands: Springer.
- Loke, M. 2018. Farm disaster survey results Kilauea East rift zone eruptions, 2018 report. Cooperative Extension Service. College of Tropical Agriculture and Natural Resources. University of Hawaii at Manoa.
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Mantovani, C.; Prado, R.M.; Pivetta, K.F.L. Silicon foliar application on nutrition and growth of Phalaenopsis and Dendrobium orchids. Sci. Hortic. 2018, 241, 83–92. [Google Scholar] [CrossRef]
- Mau, R.F.; Kessing, J.L. 1993. Frankliniella occidentalis (Pergande). Retrieved from http://www.extento.hawaii.edu/Kbase/crop/Type/f_occide.htm.
- Nagata, T.; Almeida, A.C.L.; Resende, R.O.; Avila, A.C. 2002. The transmission specificity and efficiency of tospoviruses. Pg. 45-52. In: R. Marullo, and L. Mound [eds.], Thrips and Tospoviruses: Proc. 7th Intl. Symp. Thysanoptera. Australian Nat. Insect Collection, Canberra, Australia.
- Nakahara, L.M. 1985. Thrips palmi on dendrobium, pp 29-31. In Proc. 1985 Hawaii Commercial Dendrobium Growers Conf., 11-12 Oct., Cooperative Extension Service. University of Hawaii, Honolulu.
- Parthiban, P.; Chinniah, C.; Murali Baskaran, R.K.; Suresh, K.; Karthick, S. Influence of calcium silicate application on the population of sucking pests of groundnut (Arachis hypogaea L. ). Silicon. 2018, 11, 1687–1692. [Google Scholar] [CrossRef]
- Peris-Felipo, F.J.; Benavent-Gil, Y.; Hernandez-Apaolaza, L. Silicon beneficial effects on yield, fruit quality, and shelf-life of strawberries grown in different culture substrates under different iron status. Plant Physiol. Biochem. 2020, 152, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 2021, 283. [Google Scholar] [CrossRef]
- Resende, R.S.; Rodrigues, F.A.; Cavatte, P.C.; Martins, S.C.V.; Moreira, W.R.; Chaves, A.R.M.; DaMatta, F.M. Leaf gas exchange and oxidative stress in sorghum plants supplied with silicon and infected by Colletotrichum sublineolum. Phytopathology 2012, 102, 892–898. [Google Scholar] [CrossRef]
- Reynolds, M.G.; Keeping, J.H.; Meyer, E.H. Silicon-augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; McNally, D.J.; Datnoff, L.E.; Jones, J.B.; Labb, E.C.; Benhamou, N.; Menzies, J.G.; Belanger, R.R. Silicon enhances the accumulation of diterpenoid phytoalexins in rice: A potential mechanism for blast resistance. Phytopathology 2004, 94, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Sakimura, K.; Nakahara, L.M.; Denmark, H.A. A thrips, Thrips palmi Karny (Thysanoptera: Thripsidae) Fla. Dept. Agri. Consumer Serv., Division of Plant Ind., Entomol. Circ. 1986, 280.
- Samuels, A.; Glass, A.; Ehret, D.; Menzies, J. Distribution of silicon in cucumber leaves during infection by powdery mildew fungus (Sphaerotheca fuliginea). Canad. J. of Bot. 1991, 69, 140–146. [Google Scholar] [CrossRef]
- Seal, D.R.; Kumar, V.; Kakkar, G.; Mello, S.C. Abundance of Adventive Thrips palmi (Thysanoptera: Thripidae) Populations in Florida During the First Sixteen Years. Fla. Entomol. 2013, 96, 789–796. [Google Scholar] [CrossRef]
- Srivastava, S.; Kadooka, C.; Uchida, J.Y. Fusarium species as pathogen on orchids. Microbiol. Res. 2018, 207, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Snyder, G.H. Chapter 11 Methods for silicon analysis in plants, soil, and fertilizers. Studies in Plant Science. Elsevier 2001, 8, 185–196. [Google Scholar] [CrossRef]
- Tsai, J.H.; Yue, B.; Webb, S.E.; Funderburk, J.E.; Hsu, H.T. Effects of Host Plant and Temperature on Growth and Reproduction of Thrips palmi (Thysanoptera: Thripidae). Environ. Entomol. 1995, 24, 1598–1603. [Google Scholar] [CrossRef]
- Uchida, J.Y.; Aragaki, M. Etiology of necrotic flecks on Dendrobium blossoms. Phytopathology 1979, 69, 1115–1117. [Google Scholar] [CrossRef]
- USDA NASS. 2020. Floriculture Crops 2019 Summary (December 2020). USDA, National Agricultural Statistics Service. Retrieved from chrome- https://www.nass.usda.gov/Publications/Todays_Reports/reports/floran20.pdf.
- USDA NASS. 2021. Pacific Region – Hawaii, Hawaii Horticulture and Nursery Products Annual Summary 2020. Retrieved from https://www.nass.usda.gov/Statistics_by_State/Hawaii/Publications/Flowers_and_Nursery_Products/2021 Hawaii Whole Flower.pdf.
- Vendrame, W.A.; Palmateer, A.J.; Pinares, A.; Moore, K.A.; Datnoff, L.E. Silicon fertilization affects growth of hybrid Phalaenopsis orchid liners. Horttechnology. 2010, 20, 603–607. [Google Scholar] [CrossRef]
- Wagner, F. The importance of silicic acid for the growth of some cultivated plants, their metabolism, and their susceptibility to true mildews. Phytopathol. Zeitschrift. 1940, 12, 427–479. [Google Scholar]
- Waterhouse, D.F.; Norris, K.R. 1989. Chapter 4 Frankliniella occidentalis (Pergande). pp. 24–35. In: Biological Control Pacific Prospects - Supplement 1. Australian Centre for International Agriculture Research: Canberra. 123 pages.
- Wang, C.L.; Chu, Y.I. Rearing method of southern yellow thrips, Thrips palmi Karny, in the laboratory. Plant Prot. Bull. (Taiwan, R.O.C) 1986, 28, 411. [Google Scholar]
Figure 1.
Spray length (cm) at harvest (67-75% mature blooms) by dendrobium cultivar and treatment (Si versus untreated control). Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
Figure 1.
Spray length (cm) at harvest (67-75% mature blooms) by dendrobium cultivar and treatment (Si versus untreated control). Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
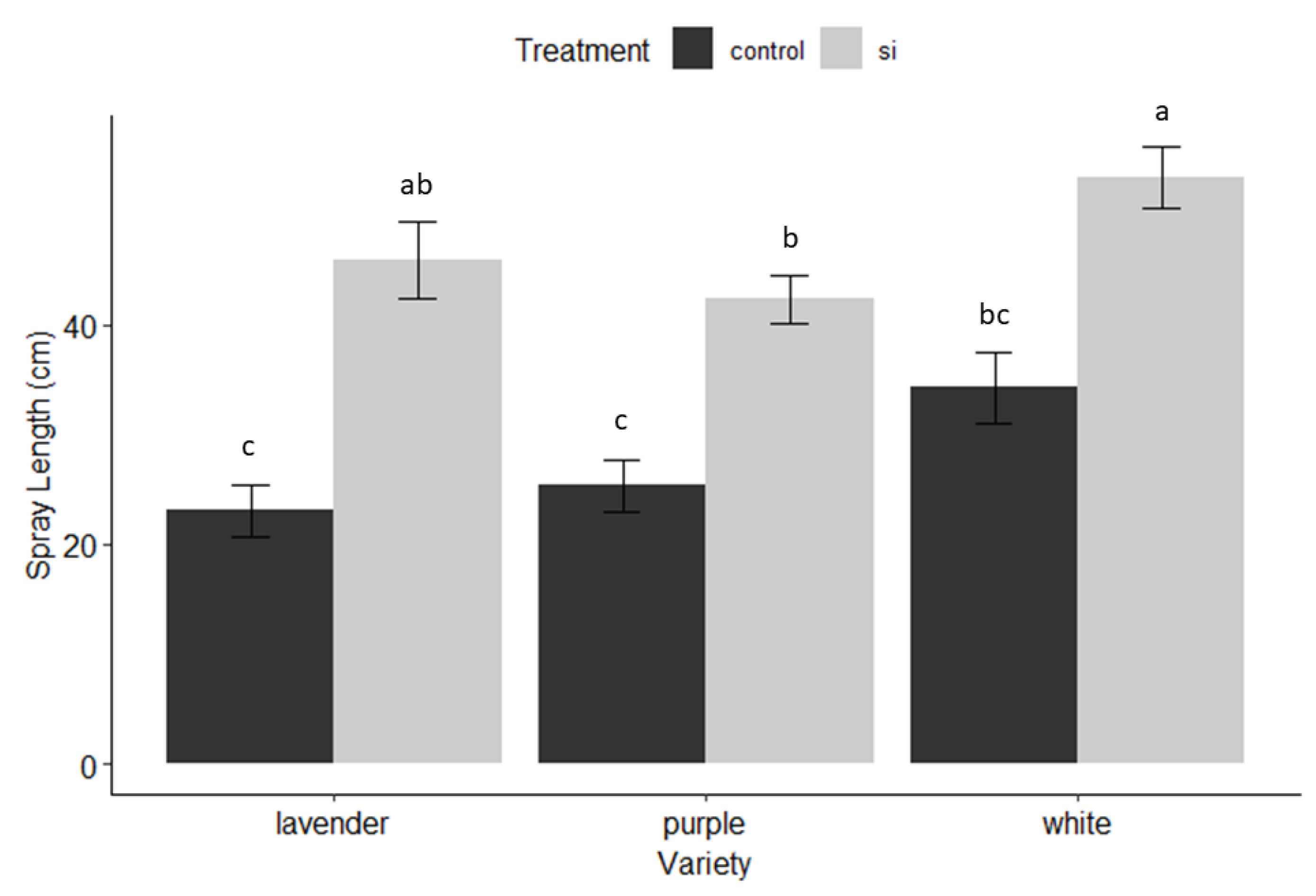
Figure 2.
Proportion of thrips damage to blooms, sprays, buds, and stems, as well as deformed and arrested spray development due to thrips damage on a percent damage scale from 0-100 by dendrobium cultivar and treatment (Si versus untreated control). Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
Figure 2.
Proportion of thrips damage to blooms, sprays, buds, and stems, as well as deformed and arrested spray development due to thrips damage on a percent damage scale from 0-100 by dendrobium cultivar and treatment (Si versus untreated control). Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
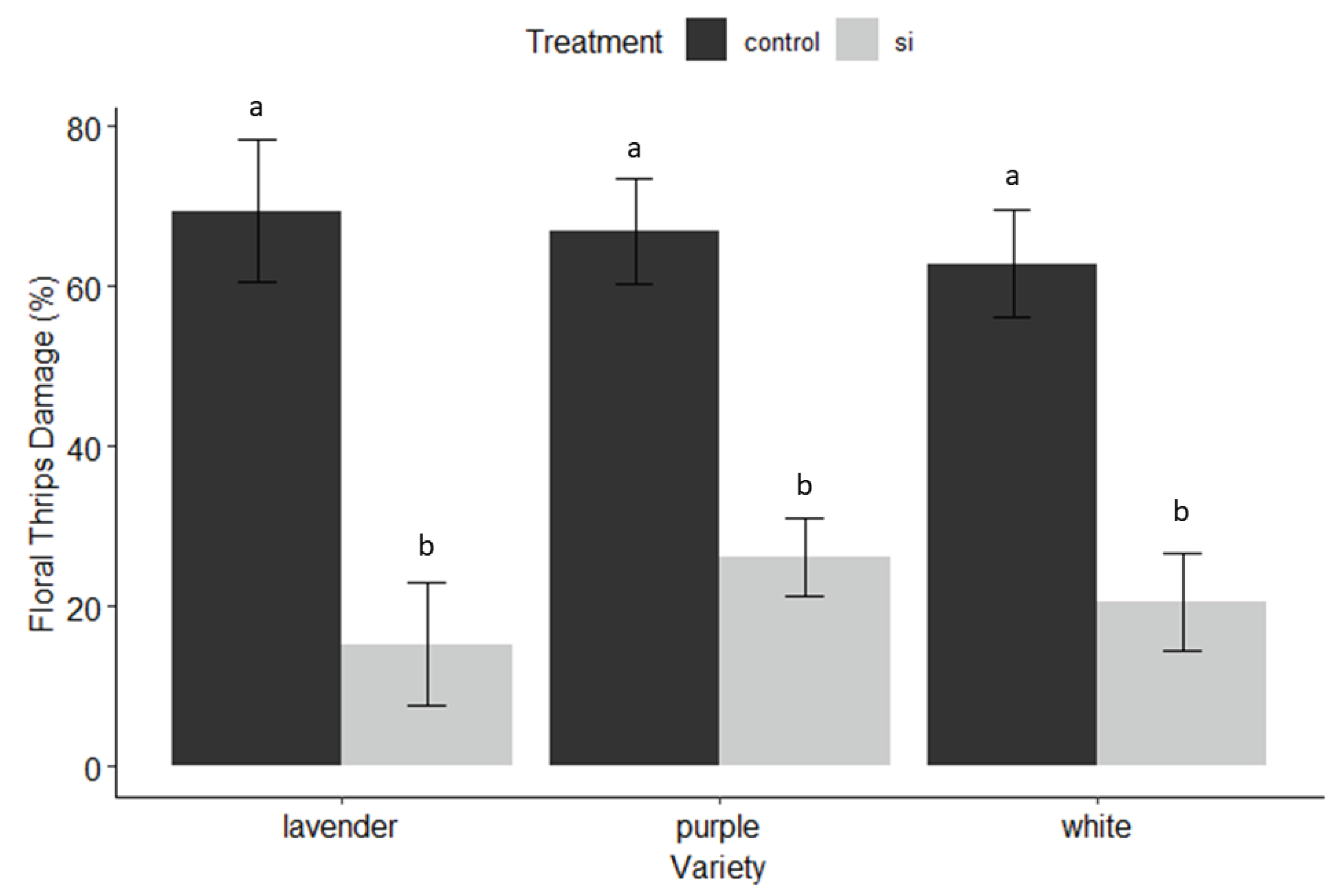
Figure 3.
Percent marketable yield by dendrobium cultivar and treatment (Si versus untreated control). Marketable yield determined as percent of total sprays with less than 80% damage to blooms and stem and thus deemed salable. Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
Figure 3.
Percent marketable yield by dendrobium cultivar and treatment (Si versus untreated control). Marketable yield determined as percent of total sprays with less than 80% damage to blooms and stem and thus deemed salable. Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
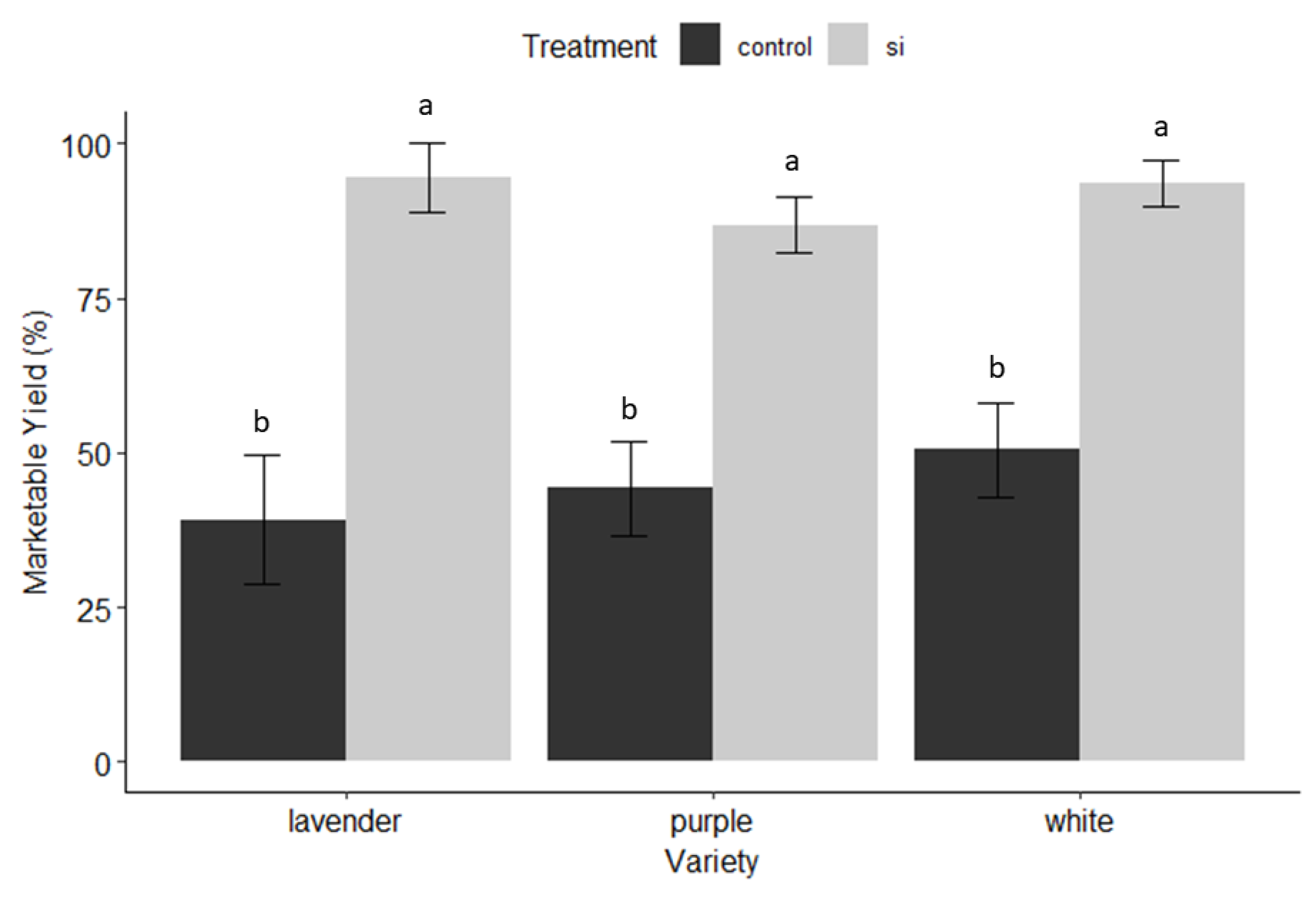
Figure 4.
The percentage of thrips damage to blooms, sprays, buds, stems, and canes on the purple dendrobium variety (‘Uniwai Royale’) over the length of the trail from September 1, 2020, to March 1, 2021.
Figure 4.
The percentage of thrips damage to blooms, sprays, buds, stems, and canes on the purple dendrobium variety (‘Uniwai Royale’) over the length of the trail from September 1, 2020, to March 1, 2021.
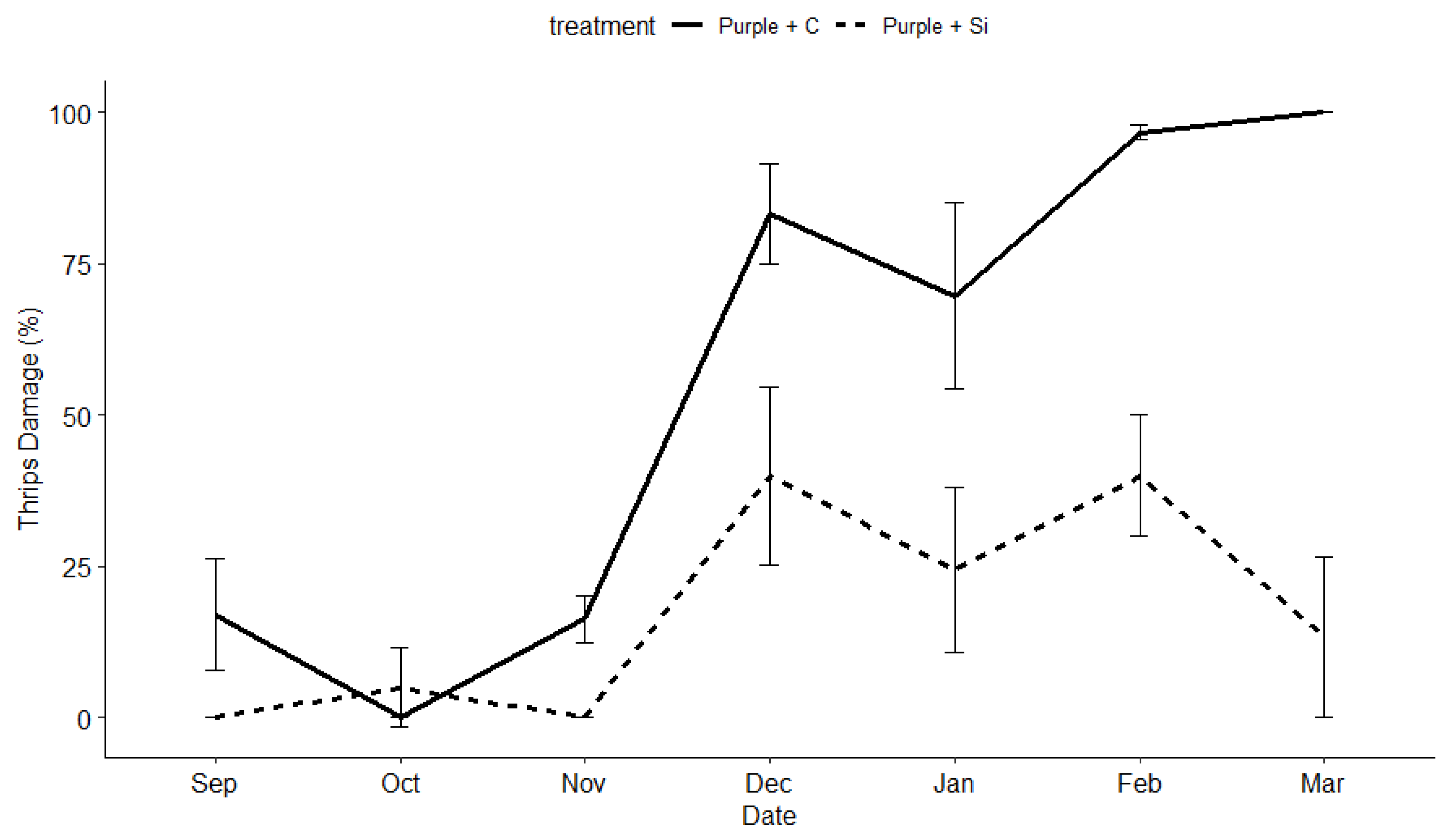
Figure 5.
The percentage of thrips damage to blooms, sprays, buds, stems, and canes on the white dendrobium variety (‘Uniwai Mist’) over the length of the trail from September 1, 2020, to March 1, 2021.
Figure 5.
The percentage of thrips damage to blooms, sprays, buds, stems, and canes on the white dendrobium variety (‘Uniwai Mist’) over the length of the trail from September 1, 2020, to March 1, 2021.
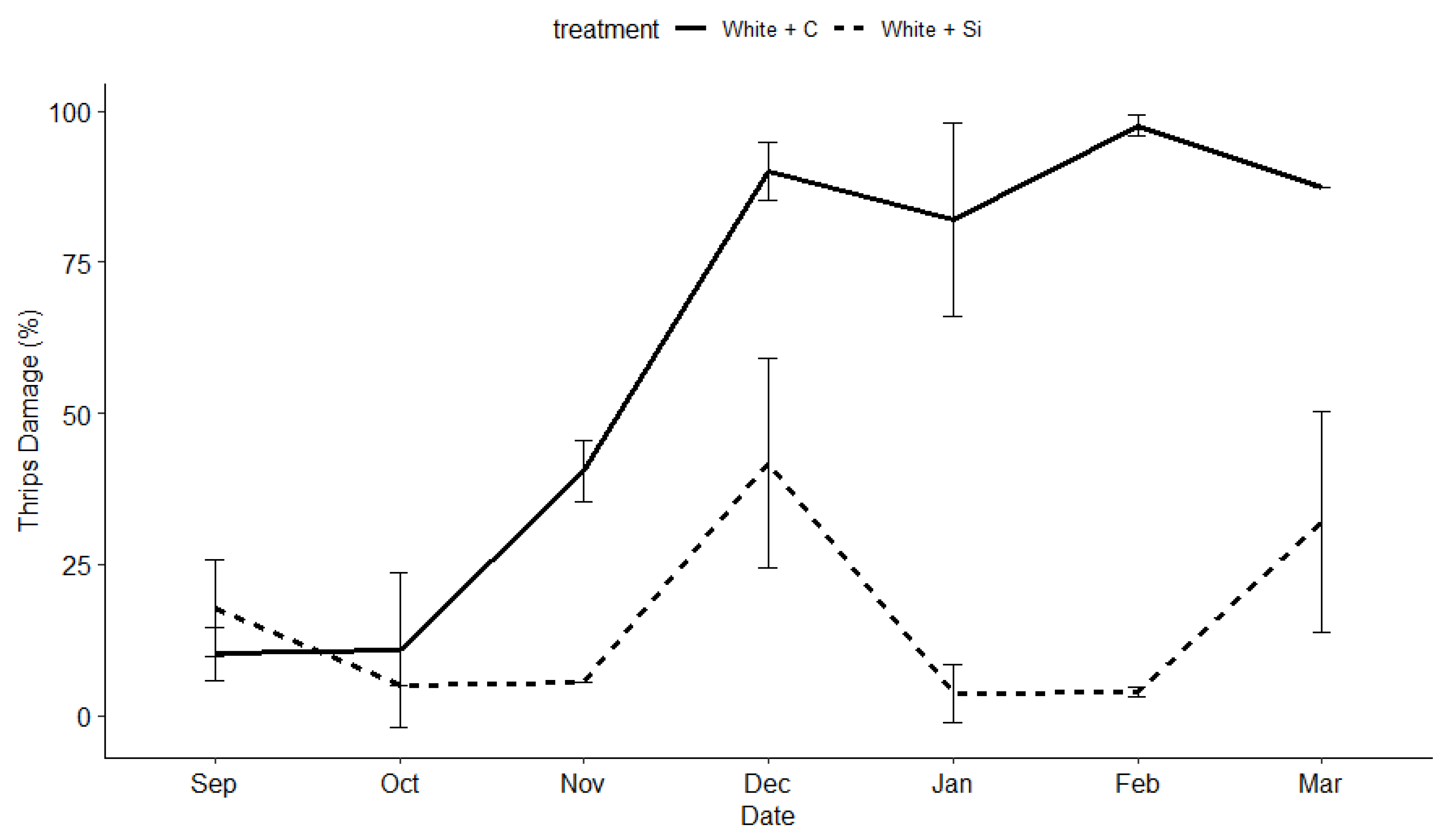
Figure 6.
The percentage of thrips damage to blooms, sprays, buds, stems, and canes on the lavender dendrobium variety (‘Uniwai Supreme’) over the length of the trail from September 1, 2020, to March 1, 2021.
Figure 6.
The percentage of thrips damage to blooms, sprays, buds, stems, and canes on the lavender dendrobium variety (‘Uniwai Supreme’) over the length of the trail from September 1, 2020, to March 1, 2021.
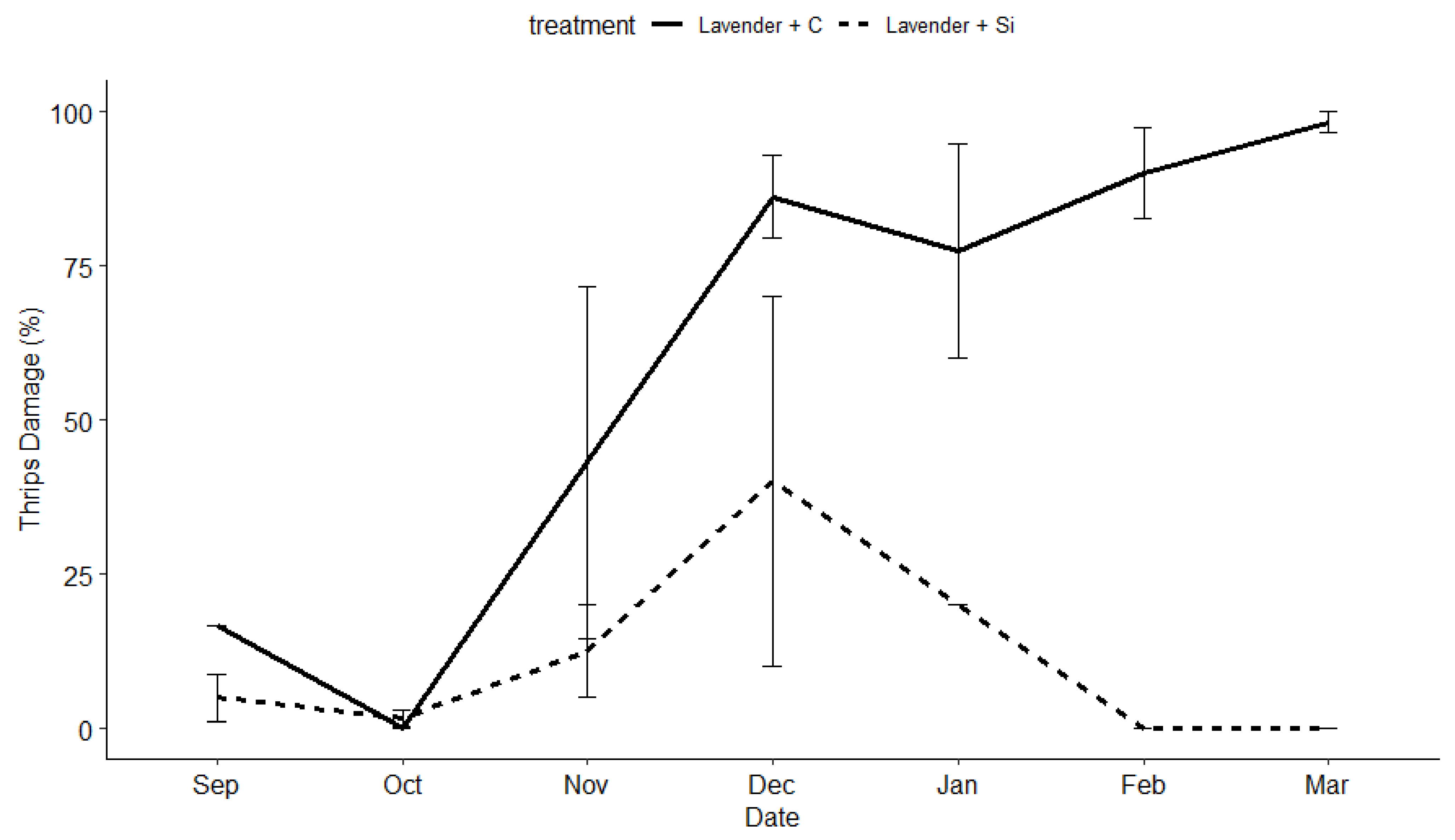
Figure 7.
Percentage of leaf and flower area exhibiting disease symptoms (Botrytis cinerea and Fusarium spp.) by dendrobium cultivar and treatment (Si versus untreated control). Disease symptoms were characterized by percent area of leaf affected by fungal legions, necrotic or blackened leaf spots, and/or percent area showing dark necrotic flecks on the flowers per plant. Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
Figure 7.
Percentage of leaf and flower area exhibiting disease symptoms (Botrytis cinerea and Fusarium spp.) by dendrobium cultivar and treatment (Si versus untreated control). Disease symptoms were characterized by percent area of leaf affected by fungal legions, necrotic or blackened leaf spots, and/or percent area showing dark necrotic flecks on the flowers per plant. Letters indicate significant differences between treatments according to pairwise comparisons between groups using R’s emmeans pairs() function.
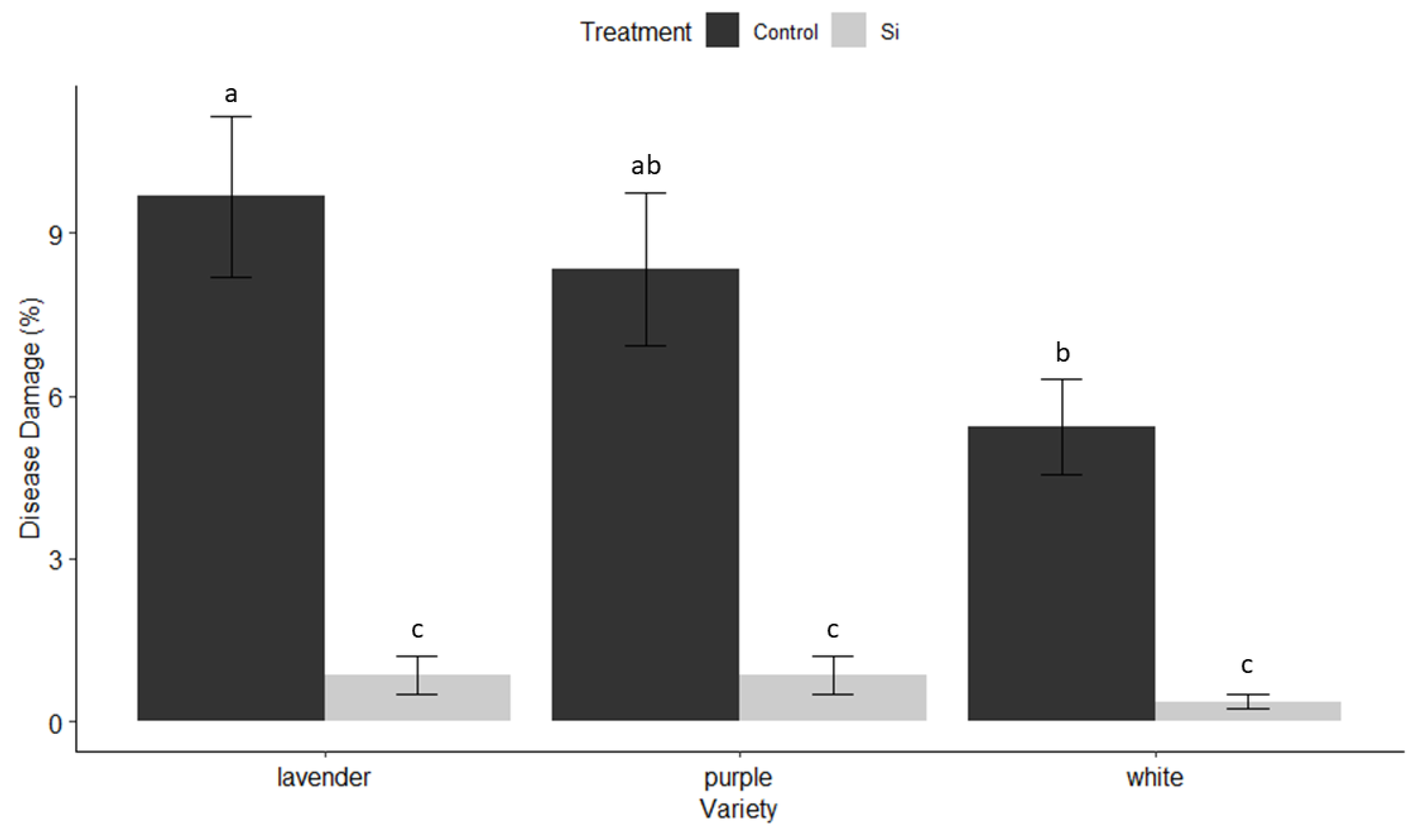
Figure 8.
SPAD index (portable chlorophyll meter) measurements taken every 14 days between August 30, 2020 to October 2, 2020 across all varieties and treatment factors. (A) Mean SPAD readings (± SE) over time for Si treated and untreated control lavender variety plants. (B) Mean SPAD readings (± SE) over time for Si treated and untreated control white variety plants. (C) Mean SPAD readings (± SE) over time for Si treated and untreated control purple variety plants. No significant differences were observed among treatments.
Figure 8.
SPAD index (portable chlorophyll meter) measurements taken every 14 days between August 30, 2020 to October 2, 2020 across all varieties and treatment factors. (A) Mean SPAD readings (± SE) over time for Si treated and untreated control lavender variety plants. (B) Mean SPAD readings (± SE) over time for Si treated and untreated control white variety plants. (C) Mean SPAD readings (± SE) over time for Si treated and untreated control purple variety plants. No significant differences were observed among treatments.
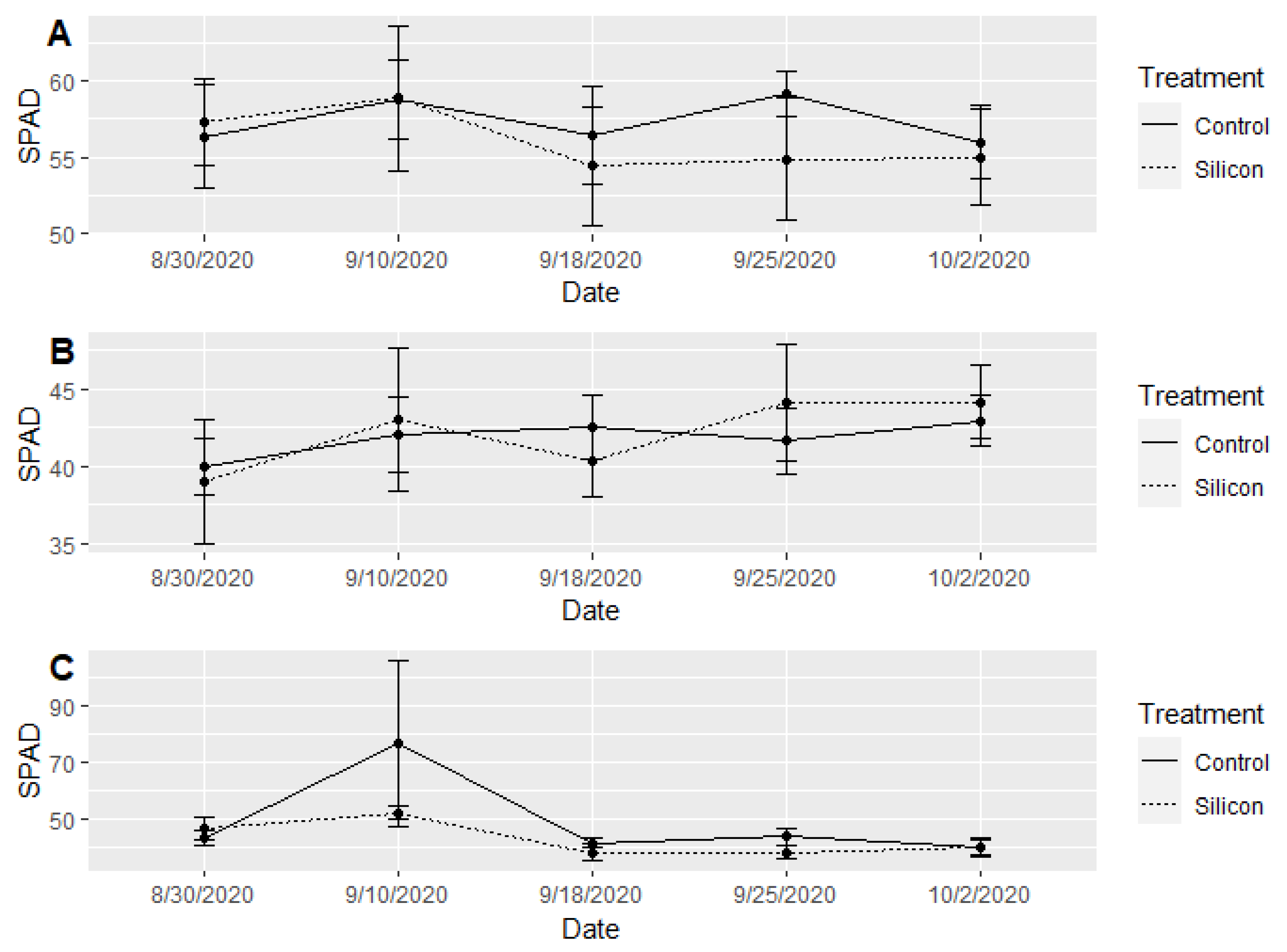
Figure 9.
Mean leaf temperatures (degrees Celsius) collected every 14 days between August 30, 2020 to October 2, 2020 across all varieties and treatment factors. (A) Mean leaf temperatures (± SE) for Si treated and untreated control lavender variety plants. (B) Mean leaf temperatures (± SE) for Si treated and untreated control white variety plants. (C) Mean leaf temperatures (± SE) for Si treated and untreated control purple variety plants.
Figure 9.
Mean leaf temperatures (degrees Celsius) collected every 14 days between August 30, 2020 to October 2, 2020 across all varieties and treatment factors. (A) Mean leaf temperatures (± SE) for Si treated and untreated control lavender variety plants. (B) Mean leaf temperatures (± SE) for Si treated and untreated control white variety plants. (C) Mean leaf temperatures (± SE) for Si treated and untreated control purple variety plants.
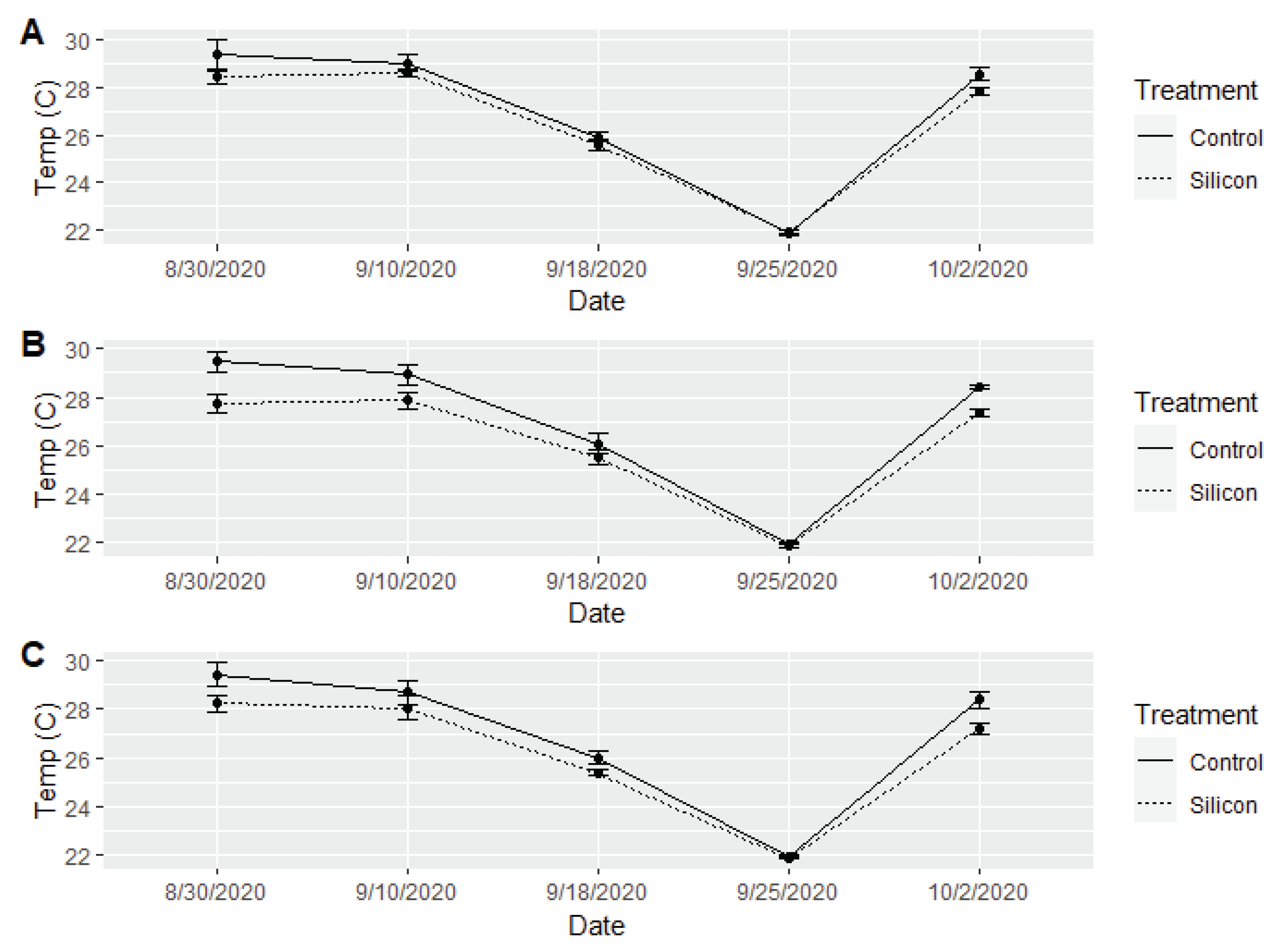
Figure 10.
Silicon accumulation (ug/g) across all variety and treatment levels determined gravimetrically in the leaf plant tissue as the residue after acid digestion. Tukey’s HSD found no significant differences between silicon treated and untreated controls for any of the varieties.
Figure 10.
Silicon accumulation (ug/g) across all variety and treatment levels determined gravimetrically in the leaf plant tissue as the residue after acid digestion. Tukey’s HSD found no significant differences between silicon treated and untreated controls for any of the varieties.


Table 1.
List of the most common thrips and fungal pests of dendrobium orchids in Hawaii.
| Scientific Name | Common Name | Categorization |
|---|---|---|
| Chaetanaphothrips orchidii* | Anthurium thrips | Thrips |
| Frankliniella occidentalis* | western flower thrips | Thrips |
| Frankliniella shultzei | yellow flower thrips | Thrips |
| Haplothrips gowdeyi | black flower thrips | Thrips |
| Chaetanaphothrips signipennis* | banana rust thrips | Thrips |
| Heliothrips haemorrhoidalis | greenhouse thrips | Thrips |
| Hercinothrips femoralis | banded greenhouse thrips | Thrips |
| Thrips hawaiiensis | Hawaiian flower thrips | Thrips |
| Thrips palmi* | melon thrips | Thrips |
| Thrips tabaci | onion thrips | Thrips |
| Botrytis cinerea* | gray mold | Fungus |
| Fusarium proliferatum* | Fusarium leaf spot | Fungus |
| Fusarium solani* | Fusarium leaf spot | Fungus |
| Phyllosticta capitalensis | Phyllosticta leaf spot | Fungus |
* Observed in our trial.
Table 2.
Values are the total numbers of sprays produced (N) by each experimental unit for the length of the trial by variety and treatment.
Table 2.
Values are the total numbers of sprays produced (N) by each experimental unit for the length of the trial by variety and treatment.
| TOTAL SPRAYS (N) | ||
| VARIETY | sILICON | cONTROL |
|
PURPLE DENDROBIUM CV ‘UNIWAI ROYALE’ |
155 ± 2.00a | 156 ± 2.40a |
|
WHITE DENDROBIUM CV ‘UNIWAI MIST’ |
99 ± 1.46a | 108 ± 1.97 a |
|
LAVENDER DENDROBIUM CV ‘UNIWAI SUPREME’ |
49 ± 1.86 a | 51 ± 1.20 a |
Letters in the same row indicate no significant differences between treatments among varieties according to pairwise comparisons between groups using R’s emmeans pairs() function.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



