
Submitted:
26 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
Pseudomonas aeruginosa is an opportunistic pathogen, especially dominant in people with cystic fibrosis; the drug resistance expressed by this pathogen and its capacity for adaptation generate a great challenge for its treatment and control, increasing morbidity and mortality rates globally. In this sense, the search for new treatment alternatives is imminent today, with products of plant origin being an excellent alternative to be used. The objective of this research was to evaluate the antibacterial and antibiofilm potential and to explore the possible effect of ethanolic extracts of wood and bark of Duguetia vallicola on the cell membrane. Microdilution assays showed inhibition of bacterial growth by more than 50%, with the lowest concentration evaluated (62.5μg/mL) of both extracts. Furthermore, we report the ability of both extracts to inhibit mature biofilms, with inhibition percentages between 48.4% and 93.7%. Intracellular material leakage experiments (260/280 nm), extracellular pH measurement, and fluorescence microscopy with acridine orange (AO) and ethidium bromide (EB) showed cell membrane damage. This indicates that the anti-bacterial action of ethanolic extracts of D. vallicola is associated with damage to the integrity of the cell membrane and consequent death of these pathogens. These results serve as a reference for future studies in establishing the mechanisms of action of these extracts.
Keywords:
Ethanolic extracts
; Duguetia vallicola
; antibacterial activity
; antibiofilm
; Pseudomonas aeruginosa.
1. Introduction
Pseudomonas aeruginosa is a gram-negative pathogen that causes illness and death in people with immunosuppressive and chronic conditions [1,2]. It is dominant in people with cystic fibrosis [3,4,5], burns or who have acquired this pathogen at the surgical site, causing coinfections and generalized sepsis [6]; its high intrinsic resistance to antiseptics and antibiotics [7,8], make it a pathogen of great clinical relevance due to the high rates of morbidity and mortality, especially in immunocompromised people [9]. According to the World Health Organization (WHO), these pathogens represent between 7 and 12% of isolates in healthcare-associated infections (HAIs) [10]. The pathogenicity, adaptability and flexibility of P. aeruginosa is due to the expression of various virulence factors [1,3,7], highlighting the production of powerful biofilms [4,11], that facilitate their multi-drug resistance and evasion of the immune system [12]. Thus, P. aeruginosa infections represent a serious public health problem at a global level [13]. Therefore, the search for therapeutic alternatives that help mitigate the problem generated by this pathogen is important today.
In this context, compounds of natural origin, especially plants, are an excellent alternative to be used, since they are an invaluable source of metabolites with recognized medicinal properties [14,15]. Compounds obtained from members of the Annonaceae family are well known for their outstanding biological activity [16,17,18,19,20,21]. The genus Duguetia is one of the most abundant of this family, from which various compounds have been obtained that include: terpenes, lignans, flavonoids, aromatic compounds and mainly it has been documented with a high content of alkaloids [22]. Alkaloids have been widely reported for their important biological activities such as: antitumor [23,24], antimicrobials [25,26,27,28], trypanocidal, leishmanicidal [23], antiplasmodial [17], antivirals [29], antioxidant, anti-inflammatory and antinociceptive, among other [30,31,32,33].
Particularly, alkaloids such as N-methylaurotetanine and its O-methylated and O-acetylated derivatives have been isolated and characterized from Duguetia vallicola, which have been documented with antioxidant activity [30]; glaziovine, with neuropharmacological properties [34] and isoquinoline alkaloids (duguevalline, cleistopholine, O-methylmoschatoline, (-)-oliveroline and (-)-oliveridine) with antiplasmodial activity against Plasmodium falciparum [17]. However, there are no reports on the antibacterial potential of D. vallicola extracts against clinical isolates of P. aeruginosa. We hypothesize that its bark and wood extracts have an antibacterial effect against P. aeruginosa. This research aimed to evaluate the antibacterial and antibiofilm potential of ethanolic extracts of wood and bark of D. vallicola, as well as to explore its mechanism of action, targeting the cell membrane. The results of this research contribute to the search for molecules and compounds of plant origin with antimicrobial potential against P. aeruginosa.
2. Results
2.1. Identification of Alkaloids in the Wood Extract.
Two alkaloids with an oxoaporphin nucleus known as liriodenine (1) and O-methylmoschatoline (2) were obtained from the D. vallicola wood extract, along with an aporphinic alkaloid known as nornuciferine (3). Alkaloids 1 and 3 are reported for the first time in D. vallicola. Their structures (Figure 1), were established by comparison of their physical and spectral data with those published in the literature. The 1H-NMR, 13C-NMR, HMQC and HMBC data for compounds 1, 2 and 3 are presented in Tables S1, S2 and S3, respectively. The 1H-NMR spectra are presented in Figures S1, S2, S3, S4 and S5.
2.2. Antibacterial Susceptibility Testing
The ethanolic extracts of wood and bark of D. vallicola showed antibacterial activity against all the clinical isolates of P. aeruginosa studied; we observed a reduction in the percentage of growth of bacteria treated with the extracts, compared to the untreated isolates used as a control. Figure 2 and Figure 3 show the similar tendency among isolates to increase the percentage of growth reduction as the concentration of wood and bark extracts increases, respectively. The wood extract, at the lowest concentration evaluated (62.5μg/mL), achieved more than 50% growth inhibition of the CLI15, CLI50 and CLI52 isolates. With the bark extract (62.5μg/mL), more than 50% and 100% growth reduction was achieved in the isolates CLI52, CLI54, CLI56, CLI57, CLI97, CLI212.
Table 1 shows the MIC90 values of the extracts for each of the clinical isolates evaluated. The effect of the extracts on the inhibition of P. aeruginosa varied between each isolate. The lowest MIC90 of the wood extracts (180 μg/mL) and bark (62.5 μg/mL) were observed with the isolates CLI15 and CLI57 respectively, while the highest MIC90 of the wood extracts (3800 μg/mL) and bark (6300 μg/mL), were observed with the CLI12 isolate.
2.3. Biofilm Reduction
All P. aeruginosa isolates produced biofilms in different proportions. The isolates CLI12, CLI51, CLI52, CLI55, and CLI57 were strong producers of biofilms in polystyrene microplates, while the rest of the isolates were weak and moderate producers of biofilms (Figure 4a). The effect of bark and wood extracts against P. aeruginosa biofilms varied between each isolate. Table 2 shows the inhibition percentages when the MIC90 of the extracts was added to the biofilms formed in each isolate; a percentage of biofilm biomass reduction was obtained between 11.5% and 93.7% with the wood extract and between 16.0% and 48.4% with the bark extract (Figure 4b), after 1 h of exposure; while the percentage of biofilm biomass reduction in cells treated with ciprofloxacin (CIP) ranged between 0.00% and 77.19%. The Kruskal Wallis test with a value of p < 0.64 and the Dunnett test (0.99 ˃ P < 0.70) for the wood and bark extracts respectively, indicated that there are no statistically significant differences between the effect of CIP and the extracts evaluated. -two in the inhibition of mature biofilms in P. aeruginosa.
2.4. Effect of Extracts on Cell Membrane Integrity
2.4.1. Leakage of Nucleic Acids and Proteins Through the Cell Membrane
The action of wood and bark extracts on the integrity of P. aeruginosa membranes was evaluated by release assays of intracellular constituents that absorb at 260/280 nm, such as nucleic acids and proteins. These tests were carried out at 0, 30, 60, 120 and 240 minutes after treatment with the MIC of the extracts for each isolate. As seen in Figure 5, the effect of the extracts on the integrity of the cell membrane varied between the isolates, but in all cases leakage of intracellular material was observed. In the CLI15 isolate, the OD260/OD280 values in the groups treated with the extracts are significantly higher from time zero, compared to the groups treated with CIP; a greater effect of the bark extract is highlighted, while for the CLI57 isolate a greater effect is observed with the wood extract. These results suggest alteration in the integrity of the cell membrane of P. aeruginosa caused by the extracts.
2.4.2. Measurement of Extracellular pH
The extracellular pH measurements of P. aeruginosa treated with the extracts, CIP and untreated cells are shown in Figure 6. As seen in the four isolates, the cells treated with the MIC of the bark and wood extracts showed a significant decrease in extracellular pH, compared to untreated cells (INO) and cells treated with CIP (2 µg/mL); this behavior was similar in all P. aeruginosa isolates. These results support the alteration in the integrity of the cell membrane caused by treatment with D. vallicola extracts.
2.4.3. LIVE/DEAD Assays
Isolates of P. aeruginosa treated with the extracts of D. vallicola, CIP and without treatment were observed under fluorescence microscopy. Acridine orange (AO) diffuses through intact cytoplasmic membranes in living cells where it interacts with DNA, emitting bright green fluorescence; in contrast, ethidium bromide (EB) penetrates only cells with damaged membranes and cell walls in dead cells, intercalates with DNA and emits orange-red fluorescence [35]. As seen in Figure 7, untreated cells (control), with completely green fluorescence, grew well after 24 h; while dead cells, with red fluorescence, were observed massively in the groups treated with the extracts; similar results to the group treated with CIP.
3. Discussion
Pseudomonas aeruginosa is a common cause of healthcare-associated infections, particularly pneumonia and in patients with structural lung disease such as cystic fibrosis. The increasing trend of resistance to antimicrobials (including multidrug-resistant (MDR) isolates) in recent years, and the presence of various virulence mechanisms that increase their ability to cause serious infections [36,37,38,39], represent a great challenge in the treatment of this pathogen. This awakens our interest in the search for compounds and new therapeutic options for the control of P. aeruginosa, with products of plant origin being an excellent alternative.
In this research, we report for the first time the antibacterial activity of ethanolic extracts of bark and wood of D. vallicola, against intrahospital isolates of P. aeruginosa. The effect of the extracts was different between the isolates and in all cases it was concentration dependent. These results coincide with those reported by Sousa [40], who found differences in the efficiency of essential oils extracted from the bark of D. lanceolata against the strains Staphylococcus aureus, Streptococcus pyogenes, Escherichia coli, P. aeruginosa and Candida albicans; since the antimicrobial effect varies depending on the concentration and the microorganism evaluated. Studies carried out with extracts from other species of the genus, D. gardneriana, D. moricandiana and D. quitensis have also reported antibacterial activity against S. aureus, S. pyogenes and E. coli, attributing this effect to the content of terpenes present in the essential oils evaluated [41].
The extracts of D. vallicola have been little studied; the presence of various types of isoquinoline alkaloids has been demonstrated, including pseudopalmatine, isoboldine, isocorydine, N-methylaurotetanine, oliveridine, oliveroline, among others [22], however, studies of antimicrobial activity of its extracts or compounds have not been reported. Alkaloids are a large and structurally diverse group of compounds that have served as scaffolds for important antibacterial drugs such as metronidazole and quinolones. They have been widely studied for their potential, not only antibacterial, but also antibiotic enhancers; furthermore, the effects of these compounds on virulence gene regulatory systems, such as quorum sensing and virulence factors, have also been reported [42,43,44]. Isoquinoline alkaloids of plant origin comprise a broad source of multimodal agents with unique structural diversity and a varied range of pharmacological activities [45], they are of great interest due to their promising biological activity [46], highlighting their antibacterial, antifungal, antiviral and antiparasitic activity, among others [47]. Likewise, studies reported by [48,49], have documented the antibacterial structure-activity relationship of isoquinoline alkaloids of the type protoberberine, protopine, benzophenanthridine, aporphine and bisbenzylisoquinoline, evidencing damage to the integrity of the membrane and cell wall, inhibition of efflux pumps and related enzymes, damage in bacterial DNA and protein synthesis, as mechanisms of antibacterial action of these compounds. Taking into account the great antibacterial potential reported for isoquinoline alkaloids and the presence of a diverse group of these alkaloids in D. vallicola, we could suggest their contribution to the inhibitory activity of ethanolic extracts of wood and bark against P. aeruginosa.
The notorious persistence of P. aeruginosa in clinical environments is attributed to its ability to form antibiotic-resistant biofilms; these function as a framework to enclose bacteria on surfaces and protect them from environmental stress, preventing phagocytosis and therefore, giving them the capacity for colonization, persistence and contributing to their multi-drug resistance [50,51,52]. We report the ability of ethanolic extracts of bark and wood to inhibit mature biofilms in P. aeruginosa (48.4% up to 93.7%), this effect being similar to that observed in cells treated with CIP in some cases. The antibiofilm activity of species of the genus Duguetia had not been reported.
Inhibition of biofilm formation by berberine in P. aeruginosa [53] and S. epidermidis [54] has been reported. Likewise, the action of sanguinarine, a benzophenanthridine alkaloid of plant origin, to inhibit bacterial biofilms has been documented [55] and the action against fungal biofilms of the chelerythrine-sanguinarine combination has also been demonstrated [56]. Taking into account the content of isoquinolinic alkaloids present in D. vallicola extracts [22], these could, at least in part, be responsible for the antibiofilm action evidenced in this research. These results are the first report of the antibiofilm action of D. vallicola extracts on P. aeruginosa.
On the other hand, the experiments of leakage of intracellular material (260/280), measurement of extracellular pH and fluorescent staining with AO and EB, showed alteration in the integrity of the cell membrane, indicating that the ethanolic extracts of D. vallicola, alter the integrity of the membrane of these pathogens, as an antibacterial mechanism, causing cell death. Our results provide new and important insights into the antibacterial and antibiofilm potential of D. vallicola extracts against P. aeruginosa; besides, we show the cell membrane as a possible target of its action; however, more studies are required to elucidate the mechanisms involved in the antimicrobial activity of this plant species. This research provides a reference for future research on the mechanisms of antimicrobial action against pathogenic bacteria of intrahospital origin.
Materials and Methods
4.1. Reagents
Mueller-Hinton broth (MHB) (Sigma, Mendota Heights, MN, USA) was used for the determination of MIC and cultures of bacterial isolates. Tryptic Soy Agar (TSA) and Tryptic Soy Broth (TSB) (Becton, Dickinson and Company, San Diego, CA, USA), Mueller-Hinton agar (MHA) (Sigma, Mendota Heights, MN, USA), and Brain Heart Infusion (BHI) broth (Sigma-Aldrich, St. Louis, MO, USA) were also used for bacterial cultures. Dimethyl
sulfoxide (DMSO), phosphate-buffered saline (PBS), crystal violet (CV), acridine orange (AO), ethidium bromide, (EB) and antibiotic ciprofloxacin (CIP), used in this study were obtained from Sigma-Aldrich, St. Louis, MO, USA; ammonium hydroxide (NH4OH), dichloromethane (DCM), hydrochloric acid (HCl), methanol (MeOH) and glacial acetic acid were obtained from Carlo Erba Reagents, Milano, Italy.
4.2. Obtaining the Extracts
The wood and bark of Duguetia vallicola were collected from a specimen located in the El Corozo village (8° 33' 39"N, 75°45'30"W) municipality of Montería Córdoba, a specimen rests in the Herbarium of the Botanical Garden Joaquín Antonio Uribe, from the city of Medellín under collection number JAUM 037841. The plant material, free of impurities, was dried at room temperature and pulverized in a knife mill. The dried and ground material (250 g of bark and 300 g of wood) were extracted separately by percolation with 96% ethanol until exhaustion. Each extract was concentrated in a rotary evaporator (Hei-VAP Core, Wood Dale IL, USA), until obtaining 32g of ethanolic bark extract and 11.4g of ethanolic wood extract.
4.3. Obtaining alkaloids from wood extract.
An 8 g fraction of the ethanolic wood extract was alkalinized to pH 8-9 with 100 mL of 5% ammonium hydroxide (NH4OH) and extracted twice with 100 mL portions of dichloromethane (DCM). The organic phase was partitioned with 150 mL of 3% hydrochloric acid (HCl) and the aqueous phase obtained was alkalinized with NH4OH until pH 8-9. Finally, it was extracted twice with 100 mL portions of DCM, obtaining a total alkaloid fraction of 235 mg. The fractionation of the total alkaloids was carried out by chromatography in successive columns (silica gel 60 (0.063-0.200mm) Merck®), eluting with DCM/MeOH mixtures from 99:1 to 90:10. The chromatographic monitoring was done with aluminum thin layer chromatography TLC plate, silica gel coated with flourescent indicator F254 (Merck®). 11.4 mg of liriodenine (1), 16.1 mg of O-methylmoschatoline (2) and 7.8 mg of nornuciferin (3) were obtained. Their structures were established by comparison of their physical and spectral data with those published in the literature, using 1H-NMR, 13C-NMR, DEPT, COSY 1H-1H, HMQC and HMBC spectra, performed on a 400 MHz Bruker Advance DRX spectrometer, in deuterated chloroform (CDCl3). The mass spectrum was obtained in electron impact ionization mode at 70 eV.
4.4. Strains
Twelve clinical isolates of P. aeruginosa (CLI12, CLI15, CLI50, CLI51, CLI52, CLI54, CLI55, CLI56, CLI57, CLI58, CLI97, CLI212) were used in this study. The isolates were cultured from blood and urine culture samples from patients hospitalized at the Social Health Service S.A.S. from the city of Sincelejo, Colombia. All microorganisms were identified using standard methods: Vitek® 2 Compact. Biomerieux SA. BHI medium and cetrimide agar were used to maintain cultures until testing was performed.
4.5. Antibacterial Susceptibility Testing
The minimum inhibitory concentration (MIC) of the ethanolic extracts of wood and bark of D. vallicola against clinical isolates was defined as the lowest concentration at which 90% (MIC90) of bacterial growth was inhibited, compared to the control (untreated cells). MIC90 was determined by performing broth microdilution assays, using 96-well microtiter plates (Nunclon Delta, Thermo Fisher Scientific, Waltham, MA, USA), as described in Clinical Laboratory Standards Institute method M07-A9 (CLSI) [57], with minor modifications. Serial dilutions were performed in MHB to obtain final concentrations of 2000, 1000, 500, 250, 125, and 62.5 μg/mL of the extracts in each reaction well. To carry out the experiments, a stock solution of each extract was prepared at 25000 μg/mL in 10% DMSO. Cells without treatment and cells treated with CIP 2 μg/mL were used as negative control and positive control respectively. The assays were developed at a final volume of 200 µL per well as follows: 100 µL of bacterial inoculum at a concentration of 108 CFU/mL and 100 µL of the extracts, adjusted to reach the previously described concentrations in the reaction wells final. Wells with bacterial inoculum, without extracts and with CIP (2 μg/mL) were used as growth controls and positive controls, respectively. The plates were incubated at 37 °C for 24 h. The experiments were performed in triplicate. The inhibition of bacterial growth by the extracts was determined by changes in optical density using a SYNERGY LX microplate reader (Biotek), at 600 nm, from the beginning of incubation to the end of incubation (24 h). Finally, the percentage of inhibition of bacterial growth [58] was calculated using the following equation:
where ODt24: optical density of the test well at 24 h post-inoculation; ODt0: optical density of the test well at 0 h post-inoculation; ODgc24: optical density of the growth control well at 24 h post-inoculation; ODgc0: optical density of the growth control well at 0 h post-inoculation.
%Inhibition = (1 - (ODt24 - ODt0/ODgc24 - ODgc0)) × 100
4.6. Quantitative Assessment of Biofilm Formation
The effect of D. vallicola wood and bark extracts on mature P. aeruginosa biofilms was evaluated following the protocol described by [59,60], with minor modifications. For biofilm formation, bacterial colonies from 24 h of incubation in TSA were used, standardizing the bacterial inoculum at 108 cells/mL. Then, in 96-well polystyrene microplates, 200 µL of the bacterial inoculum was discharged into each well and incubated at 37 °C for 24 h. Subsequently, the broth was removed from the microplates and 200 µL of the wood and bark extracts (evaluated separately), at the MIC concentration of each isolate, were added to TSB broth and incubated at 37 °C for 1 hour. Subsequently, the floating cells were removed and the biofilms from the bottom of the wells were washed with deionized water. Excess moisture was then removed by tapping the microplates on sterile napkins and the plates were dried for 5 minutes. Six replicates of each experiment were performed. Cultures without extracts were used as a negative control and cultures with CIP as a positive control. Biofilm reductions were quantified by staining the wells with 200 µl of 0.1% CV for 20 min. The samples were washed with deionized water until excess dye was removed; the excess water was carefully dried and then the CV was solubilized in 250 µL of 30% glacial acetic acid. Absorbance values were measured at 590 nm (OD590), using a SYNERGY LX microplate reader (Biotek). Biofilm production was grouped into the following categories: OD590 < 0.1: non-producers (NP), OD590 0.1–1.0: weak producers (WP), OD590 1.1–3.0: moderate producers (MP) and OD590 > 3.0: producers strong (SP). Biofilm reduction was calculated [61] using the following equation:
where AbsCO: absorbance of the control sample and AbsExt: absorbance of the sample treated with the extracts.
% Biofilm reduction: AbsCO - AbsExt/AbsCO × 100
4.7. Effect of Extracts on Cell Membrane Integrity
The evaluation of the action of ethanolic extracts of wood and bark of D. vallicola on the cell membrane of P. aeruginosa was carried out through experiments of intracellular material leakage, measurement of extracellular pH and cell fluorescence microscopy treated with each of the evaluated extracts.
4.7.1. Leakage of Nucleic Acids and Proteins Through the Cell Membrane
The release of intracellular material was measured according to the methodology proposed by [62], with some modifications. Cells cultured in nutrient broth were centrifuged at 3000 g for 20 minutes, washed three times and resuspended in 20 ml of PBS (pH 7.0). The cell suspension was then treated with the wood and bark extracts (MIC for each isolate; separate experiments) and incubated at 37 °C for 0, 30, 60, and 120 min. Subsequently, 2 mL of the samples were collected and centrifuged at 3000 g for 20 min. Then, to determine the concentration of the released constituents, 2 mL of supernatant was used to measure the absorbance at 260/280 nm with a Spectroquant® Prove 300 UV/Vis spectrophotometer. Samples without extracts and samples with CIP were used as controls. All assays were performed in triplicate.
4.7.2. Measurement of Extracellular pH
The measurement of the extracellular pH of P. aeruginosa after treatment with D. vallicola wood and bark extracts was determined according to [61], with some modifications. 20 mL of the bacterial suspension (1x107 CFU/mL) in nutrient broth was incubated at 37 °C for 24 h. The samples were then centrifuged at 3000 g for 20 min; the sediment was collected, resuspended and washed three times with double-distilled water, resuspending again in 20 ml of sterile double-distilled water. After the addition of the extracts (MIC of each isolate), the extracellular pH of P. aeruginosa was determined at 0, 30, 60 and 120 min, using a Schott® Instruments Handylab pH 11 pH meter. They were used as controls samples without extracts and samples with CIP.
4.7.3. LIVE/DEAD Assays
The LIVE/DEAD assays were developed following the methodology proposed by [35]. A suspension of P. aeruginosa (107 CFU/mL) was placed on sterile slides and incubated for 24 h. The cells were then washed three times with PBS. Subsequently, the MIC of the extracts of each isolate and CIP were added to the experimental groups and the bacterial inoculum in nutrient broth was used as a control. The prepared slides were incubated at 37 °C for 24 h and then washed three times with PBS. Together, AO (5 µL, 100 mg/L) and EB (5 µL, 100 mg/L) were mixed under dark conditions and added to the slides under dark conditions for 30 sec. The samples were then observed on an Olympus BX43 fluorescence microscope and photographed with a DP72 camera.
4.8. Data Analysis
The results were analyzed using GraphPad Prism version 8.0 software and the Microsoft Excel program. Initially, the Shapiro Wilk test was used to determine the distribution of the data. Subsequently, Pearson's correlation coefficient was used to measure the degree of linearity, the correlation between the concentration of the extracts and the percentage reduction in bacterial growth. To compare the effect of the extracts and CIP on biofilm reduction, the Kruskal Wallis test and the Dunnett test were used; this test was also used to compare the effects of the extracts and CIP on the leakage of intracellular material through the membrane (260/280 nm) and to compare the effects of the treatments on the extracellular pH of P. aeruginosa.
5. Conclusions
In this study we investigated the antibacterial potential of ethanolic extracts of Duguetia vallicola wood and bark against clinical isolates of Pseudomonas aeruginosa, as well as their role in biofilm inhibition. Furthermore, we explored the action against the cell membrane of these pathogens. We demonstrate the antibacterial action of the extracts under study against P. aeruginosa, this effect being associated with damage to the integrity of the cell membrane, in addition to its action against bacterial biofilms. Additionally, we report the presence of three isoquinoline alkaloids in the wood of D. vallicola. It is necessary to continue these studies, with a view to elucidating the mechanisms of antibacterial action and the composition of the extracts of this species, which promises as an alternative tool for the treatment and control of intrahospital pathogens such as P. aeruginosa.
Supplementary Materials
The following supporting information can be downloaded at: The following supporting information can be downloaded at the website of this paper posted on Preprints.org.. Table S1. NMR Spectroscopic Data (1H 400 MHz, 13C 100 MHz) of 1 in CDCl3. Table S2. NMR Spectroscopic Data (1H 400 MHz, 13C 100 MHz) of 2 in CDCl3. Table S3. NMR Spectroscopic Data (1H 400 MHz, 13C 100 MHz) of 3 in CDCl3. Figure S1. 1H-NMR spectrum (amplification 6.2 – 9.1 ppm) of 1, (CDCl3, 400 MHz). Figure S2. 1H-NMR spectrum (amplification 4.0 – 9.2 ppm) of 2, (CDCl3, 400 MHz). Figure S3. 1H-NMR spectrum (amplification 7.5 – 9.2 ppm) of 2, (CDCl3, 400 MHz). Figure S4. 1H-NMR spectrum of 3, (CDCl3, 400 MHz). Figure S5. 1H-NMR spectrum (amplifications) of 3, (CDCl3, 400 MHz).
Author Contributions
Conceptualization, O.I.C.M., D.S.Q. and A.A.O.; methodology, O.I.C.M., D.S.Q. and A.A.O.; validation, O.I.C.M., D.S.Q. and A.A.O.; formal analysis, O.I.C.M. and A.A.O.; writing—original draft preparation, O.I.C.M. and A.A.O.; writing—review and editing, O.I.C.M., D.S.Q. and A.A.O.; supervision, O.I.C.M. and A.A.O.; project administration, A.A.O.; funding acquisition, A.A.O. and O.I.C.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded with resources from the SI-FCB-01-22 project (Call to strengthen Research Seedbeds, 2022), of the University of Córdoba, Montería, Colombia.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
The authors thank the University of Córdoba for funding the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Diggle, S. P.; Whiteley, M. Microbe Profile: Pseudomonas aeruginosa: Opportunistic Pathogen and Lab Rat. Microbiol. (United Kingdom) 2020, 166, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Callejas-Díaz, A.; Fernández-Pérez, C.; Ramos-Martínez, A.; Múñez-Rubio, E.; Sánchez-Romero, I.; Vargas Núñez, J. A. Impacto de La Bacteriemia Por Pseudomonas aeruginosa En Un Hospital de Tercer Nivel: Mortalidad y Factores Pronósticos. Med. Clin. (Barc). 2019, 152, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Martín, I.; Sainz-Mejías, M.; McClean, S. Pseudomonas aeruginosa: An Audacious Pathogen with an Adaptable Arsenal of Virulence Factors. Int. J. Mol. Sci. 2021, 22, 3128. [Google Scholar] [CrossRef] [PubMed]
- Mielko, K. A.; Jabłoński, S. J.; Milczewska, J.; Sands, D.; Łukaszewicz, M.; Młynarz, P. Metabolomic Studies of Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2019, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Camus, L.; Vandenesch, F.; Moreau, K. From Genotype to Phenotype: Adaptations of Pseudomonas aeruginosa to the Cystic Fibrosis Environment. Microb. Genomics 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Paz-Zarza, VM, Mangwani-Mordani, S. , Martínez-Maldonado, A., Álvarez-Hernández, D., Solano-Gálvez, SG, & Vázquez-López, R. Pseudomonas aeruginosa: Patogenicidad y Resistencia Antimicrobiana En La Infección Urinaria. Patogenia 2019, 36, 180–189. [Google Scholar]
- Chevalier, S.; Bouffartigues, E.; Bodilis, J.; Maillot, O.; Lesouhaitier, O.; Feuilloley, M. G. J.; Orange, N.; Dufour, A.; Cornelis, P. Structure, Function and Regulation of Pseudomonas aeruginosa Porins. FEMS Microbiol. Rev. 2017, 41, 698–722. [Google Scholar] [CrossRef] [PubMed]
- Huszczynski, S. M.; Lam, J. S.; Khursigara, C. M. The Role of Pseudomonas aeruginosa Lipopolysaccharide in Bacterial Pathogenesis and Physiology. Pathogens 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Pang, Z.; Li, H.; Yang, T.; Guo, L.; Liu, L.; Mei, J.; Song, X.; Xie, T.; Zhang, Y.; He, X.; Lin, T. J.; Xie, Z. CXCL4 Contributes to Host Defense against Acute Pseudomonas aeruginosa Lung Infection. PLoS One 2018, 13, 1–19. [Google Scholar] [CrossRef]
- Aman, S.; Mittal, D.; Shriwastav, S.; Tuli, H. S.; Chauhan, S.; Singh, P.; Sharma, S.; Saini, R. V.; Kaur, N.; Saini, A. K. Prevalence of Multidrug-Resistant Strains in Device Associated Nosocomial Infection and Their in Vitro Killing by Nanocomposites. Ann. Med. Surg. 2022, 78. [Google Scholar] [CrossRef]
- Sharma, G.; Rao, S.; Bansal, A.; Dang, S.; Gupta, S.; Gabrani, R. Pseudomonas aeruginosa Biofilm: Potential Therapeutic Targets. Biologicals 2014, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B. R.; Lin, T. J.; Cheng, Z. Antibiotic Resistance in Pseudomonas aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Y.; Dickter, J. K. Nosocomial Infections: A History of Hospital-Acquired Infections. Gastrointest. Endosc. Clin. N. Am. 2020, 30, 637–652. [Google Scholar] [CrossRef]
- Avato, P. Editorial to the Special Issue –“Natural Products and Drug Discovery". Molecules 2020, 25, 1128. [Google Scholar] [CrossRef]
- Aylate, A.; Agize, M.; Ekero, D.; Kiros, A.; Ayledo, G.; Gendiche, K. In-Vitro and In-Vivo Antibacterial Activities of Croton macrostachyus Methanol Extract against E. coli and S. aureus. Adv. Anim. Vet. Sci. 2017, 5, 107–114. [Google Scholar] [CrossRef]
- Lúcio, A. S. S. C.; Almeida, J. R. G. da S.; da-Cunha, E. V. L.; Tavares, J. F.; Barbosa Filho, J. M. Alkaloids of the Annonaceae: Occurrence and a Compilation of Their Biological Activities; 2015; Vol. 74. [CrossRef]
- Pérez, E.; Saez, J.; Blair, S.; Franck, X.; Figadere, B. Isoquinoline Alkaloids from Duguetia vallicola Stem Bark with Antiplasmodial Activity. Lett. Org. Chem. 2004, 1, 102–104. [Google Scholar] [CrossRef]
- Menezes, L.; D’Sousa Costa, C.; Rodrigues, A. C.; Santo, F. R.; Nepel, A.; Dutra, L. M.; Silva, F.; Soares, M. B.; Barison, A.; Costa, E. V.; Bezerra, D. P. Cytotoxic Alkaloids from the Stem of Xylopia laevigata. Molecules 2016, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alves, D. S.; Machado, A. R. T.; Campos, V. A. C.; Oliveira, D. F.; Carvalho, G. A. Selection of Annonaceae Species for the Control of Spodoptera frugiperda (Lepidoptera: Noctuidae) and Metabolic Profiling of Duguetia lanceolata Using Nuclear Magnetic Resonance Spectroscopy. J. Econ. Entomol. 2016, 109, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Maia, D. S.; Lopes, C. F.; Saldanha, A. A.; Silva, N. L.; Sartori, Â. L. B.; Carollo, C. A.; Sobral, M. G.; Alves, S. N.; Silva, D. B.; de Siqueira, J. M. Larvicidal Effect from Different Annonaceae Species on Culex quinquefasciatus. Environ. Sci. Pollut. Res. 2020, 27, 36983–36993. [Google Scholar] [CrossRef]
- Fuentes, L.; Contreras, O. I.; Angulo, A. A. Evaluation of in Vitro Antioxidant Activity of Extracts from Duguetia vallicola J. F. Macbr. – Annonaceae. Rev. Fac. Ciencias Básicas 2021, 1, 63–63. [Google Scholar] [CrossRef]
- Pérez, E. G.; Cassels, B. K. Alkaloids from the Genus Duguetia. Alkaloids Chem. Biol. 2010, 68, 83–156. [Google Scholar] [CrossRef] [PubMed]
- 23 Da Silva, D. B.; Tulli, E. C.; Militão, G. C.; Costa-Lotufo, L. V.; Pessoa, C.; de Moraes, M. O.; Albuquerque, S.; de Siqueira, J. M. The Antitumoral, Trypanocidal and Antileishmanial Activities of Extract and Alkaloids Isolated from Duguetia furfuracea. Phytomedicine 2009, 16, 1059–1063. [Google Scholar] [CrossRef]
- Jin, Q.; Yang, D.; Dai, Z.; Khan, A.; Wang, B.; Wei, X.; Sun, Y.; Zhao, Y. L.; Wang, Y. F.; Liu, Y. P.; Zhao, X. D.; Luo, X. D. Antitumor Aporphine Alkaloids from Thalictrum wangii. Fitoterapia 2018, 128, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.; Dunbar, D. C.; Takamatsu, S.; Walker, L. A.; Clark, A. M. Antimalarial, Cytotoxic, and Antifungal Alkaloids from Duguetia hadrantha. J. Nat. Prod. 2001, 64, 559–562. [Google Scholar] [CrossRef]
- Hu, J.; Shi, X.; Chen, J.; Mao, X.; Zhu, L.; Yu, L.; Shi, J. Alkaloids from Toddalia asiatica and Their Cytotoxic, Antimicrobial and Antifungal Activities. Food Chem. 2014, 148, 437–444. [Google Scholar] [CrossRef]
- Almeida, M. C.; Resende, D. I.; da Costa, P. M.; Pinto, M. M.; Sousa, E. Tryptophan Derived Natural Marine Alkaloids and Synthetic Derivatives as Promising Antimicrobial Agents. Eur. J. Med. Chem. 2021, 209, 112945. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y. Y.; Zhou, H.; Peng-Deng; Zhang, B. Q.; Zhang, Z. J.; Wang, G. H.; Zhang, S. Y.; Wu, Z. R.; Wang, Y. R.; Liu, Y. Q. Antimicrobial Activity of Natural and Semi-Synthetic Carbazole Alkaloids. Eur. J. Med. Chem. 2023, 259. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Z.; Mukhamadiev, A.; Feng, J.; Gao, Y.; Zhuansun, X.; Han, R.; Chong, Y.; Jafari, S. M. Formulation Optimization and Characterization of Carvacrol-Loaded Nanoemulsions: In Vitro Antibacterial Activity/Mechanism and Safety Evaluation. Ind. Crops Prod. 2022, 181. [Google Scholar] [CrossRef]
- Arango, O.; Pérez, E.; Granados, H.; Rojano, B.; Sáez, J. Inhibition of Lipid Peroxidation and Free Radical Scavenging Capacity of Alkaloids Isolated from Two Annonaceae, Xylopia amazonica Cf. And Duguetia vallicola. Actual Biol 2004, 26, 105–110. [Google Scholar] [CrossRef]
- Vieira Lemos, D. E. C.; Cavalcante-Silva, L. H. A.; Almeida Lima, É. De; Alves, A. F.; Carneiro Lúcio, A. S.; Barbosa-Filho, J. M.; Mascarenhas, S. R. Anti-Inflammatory Effect of Discretamine, a Protoberberine Alkaloid Isolated from Duguetia moricandiana. Nat. Prod. Commun. 2017, 12, 1595–1597. [Google Scholar] [CrossRef]
- Do Santos, R. C.; de Souza, A. V.; Andrade-Silva, M.; Vera Cruz, Karoliny C. ; Leite Kassuya, C. A.; Lima Cardoso, C. A.; Vieira, M. do C.; Nazari Formagio, A. S. Antioxidant, Anti-Rheumatic and Anti-Inflammatory Investigation of Extract and Dicentrinone from Duguetia furfuracea (A. St.-Hil.) Benth. & Hook. F. J. Ethnopharmacol. 2018, 211, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, A. A.; Vieira, L.; Ribeiro, R. I. de A.; Thomé, R. G.; Santos, H. B. dos; Silva, D. B.; Carollo, C. A.; Oliveira, F. M. de; Lopes, D. de O.; Siqueira, J. M. de; Soares, A. C. Chemical Composition and Evaluation of the Anti-Inflammatory and Antinociceptive Activities of Duguetia furfuracea Essential Oil: Effect on Edema, Leukocyte Recruitment, Tumor Necrosis Factor Alpha Production, INOS Expression, and Adenosinergic and Opioid. J. Ethnopharmacol. 2019, 231, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Pérez, E.; Saez, J.; Cassels, B. A Convenient, Renewable Source of the Anxiolytic Proaporphine Alkaloid Glaziovine: Duguetia vallicola Leaves. J. Chil. Chem. Soc. 2005, 50, 553–557. [Google Scholar] [CrossRef]
- Contreras, O.; Angulo, A.; Santafé, G. Mechanism of Antifungal Action of Monoterpene Isoespintanol against Clinical Isolates of Candida tropicalis. Molecules 2022, 27, 5808. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.; Kollef, M. The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef] [PubMed]
- Jangra, V.; Sharma, N.; Chhillar, A. K. Therapeutic Approaches for Combating Pseudomonas aeruginosa Infections. Microbes Infect. 2022, 24, 104950. [Google Scholar] [CrossRef] [PubMed]
- Horcajada, J. P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef]
- Aman, S.; Mittal, D.; Shriwastav, S.; Tuli, H. S.; Chauhan, S.; Singh, P.; Sharma, S.; Saini, R. V.; Kaur, N.; Saini, A. K. Prevalence of Multidrug-Resistant Strains in Device Associated Nosocomial Infection and Their in Vitro Killing by Nanocomposites. Ann. Med. Surg. 2022, 78. [Google Scholar] [CrossRef] [PubMed]
- Sousa, O. V.; Del-Vechio-Vieira, G.; Alves, M. S.; Araújo, A. A.; Pinto, M. A.; Amaral, M. P.; Rodarte, M. P.; Kaplan, M. A. Chemical Composition and Biological Activities of the Essential Oils from Duguetia lanceolata St. Hil. Barks. Molecules 2012, 17, 11056–11066. [Google Scholar] [CrossRef]
- Dos Santos, A. C.; Nogueira, M. L.; de Oliveira, F. P.; Costa, E. V.; Bezerra, D. P. Essential Oils of Duguetia Species A. St. Hill (Annonaceae): Chemical Diversity and Pharmacological Potential. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- Cushnie, T. P.; Cushnie, B.; Lamb, A. J. Alkaloids: An Overview of Their Antibacterial, Antibiotic-Enhancing and Antivirulence Activities. Int. J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Dahmer, J.; do Carmo, G.; Mostardeiro, M. A.; Neto, A. T.; da Silva, U. F.; Dalcol, I. I.; Morel, A. F. Antibacterial Activity of Discaria americana Gillies Ex Hook (Rhamnaceae). J. Ethnopharmacol. 2019, 239. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; He, Y.; Li, F.; Chai, C.; Zhang, J.; Guo, J.; Chen, C.; Wang, J.; Zhu, H.; Hu, Z.; Zhang, Y. Antibacterial Activity against Drug-Resistant Microbial Pathogens of Cytochalasan Alkaloids from the Arthropod-Associated Fungus Chaetomium globosum TW1-1. Bioorg. Chem. 2019, 83, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Plazas, E.; Avila M, M.; Muñoz, D. R.; Cuca S, L. Natural Isoquinoline Alkaloids: Pharmacological Features and Multi-Target Potential for Complex Diseases. Pharmacol. Res. 2022, 177, 106126. [Google Scholar] [CrossRef] [PubMed]
- Pässler, U.; Knölker, H. J. The Pyrrolo[2,1-a]Isoquinoline Alkaloids; 2011; Vol. 70. [CrossRef]
- Tuzimski, T.; Petruczynik, A. New Trends in the Practical Use of Isoquinoline Alkaloids as Potential Drugs Applicated in Infectious and Non-Infectious Diseases. Biomed. Pharmacother. 2023, 168. [Google Scholar] [CrossRef] [PubMed]
- Nair, J. J.; Wilhelm, A.; Bonnet, S. L.; van Staden, J. Antibacterial Constituents of the Plant Family Amaryllidaceae. Bioorganic Med. Chem. Lett. 2017, 27, 4943–4951. [Google Scholar] [CrossRef]
- Yan, Y.; Li, X.; Zhang, C.; Lv, L.; Gao, B.; Li, M. Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review. Antibiotics 2021, 10. [Google Scholar] [CrossRef]
- Thi, M. T. T.; Wibowo, D.; Rehm, B. H. A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P. K.; Yeung, A. T. Y.; Hancock, R. E. W. Antibiotic Resistance in Pseudomonas aeruginosa Biofilms: Towards the Development of Novel Anti-Biofilm Therapies. J. Biotechnol. 2014, 191, 121–130. [Google Scholar] [CrossRef]
- Parsek, M. R.; Tolker-Nielsen, T. Pattern Formation in Pseudomonas aeruginosa Biofilms. Curr. Opin. Microbiol. 2008, 11, 560–566. [Google Scholar] [CrossRef]
- Aswathanarayan, J. B.; Vittal, R. R. Inhibition of Biofilm Formation and Quorum Sensing Mediated Phenotypes by Berberine in: Pseudomonas aeruginosa and Salmonella typhimurium. RSC Adv. 2018, 8, 36133–36141. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yao, X.; Zhu, Z.; Tang, T.; Dai, K.; Sadovskaya, I.; Flahaut, S.; Jabbouri, S. Effect of Berberine on Staphylococcus epidermidis Biofilm Formation. Int. J. Antimicrob. Agents 2009, 34, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, W.; Liu, M.; Zhang, J.; Yang, M.; Wang, T.; Qian, W. In Vitro Anti-Biofilm Efficacy of Sanguinarine against Carbapenem-Resistant Serratia marcescens. Biofouling 2021, 37, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Yang, M.; Li, X.; Sun, Z.; Li, Y.; Wang, X.; Wang, T. Anti-Microbial and Anti-Biofilm Activities of Combined Chelerythrine-Sanguinarine and Mode of Action against Candida albicans and Cryptococcus neoformans in Vitro. Colloids Surfaces B Biointerfaces 2020, 191, 111003. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard — Ninth Edition; 2012; Vol. 32.
- Quave, C. L.; Plano, L. R. W.; Pantuso, T.; Bennett, B. C. Effects of Extracts from Italian Medicinal Plants on Planktonic Growth, Biofilm Formation and Adherence of Methicillin-Resistant Staphylococcus aureus. J. Ethnopharmacol. 2008, 118, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Serio, A.; Chaves-López, C.; Anniballi, F.; Auricchio, B.; Goffredo, E.; Cenci-Goga, B. T.; Lista, F.; Fillo, S.; Paparella, A. Biofilm Formation, Pigment Production and Motility in Pseudomonas Spp. Isolated from the Dairy Industry. Food Control 2018, 86, 241–248. [Google Scholar] [CrossRef]
- Yousefpour, Z.; Davarzani, F.; Owlia, P. Evaluating of the Effects of Sub-MIC Concentrations of Gentamicin on Biofilm Formation in Clinical Isolates of Pseudomonas aeruginosa. Iran. J. Pathol. 2021, 16, 403–410. [Google Scholar] [CrossRef]
- Donadu, M. G.; Peralta-ruiz, Y.; Usai, D.; Maggio, F.; Molina-hernandez, J. B.; Rizzo, D.; Bussu, F.; Rubino, S.; Zanetti, S.; Paparella, A.; Chaves-López, C. Colombian Essential Oil of Ruta graveolens against Nosocomial Antifungal Resistant Candida Strains. J. Fungi 2021, 7, 383. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cai, J.; Chen, H.; Zhong, Q.; Hou, Y.; Chen, W.; Chen, W. Antibacterial Activity and Mechanism of Linalool against Pseudomonas aeruginosa. Microb. Pathog. 2020, 141, 103980. [Google Scholar] [CrossRef]
Figure 1.
Structures of liriodenine (1), O-methylmoschatoline (2) and nornuciferine (3), isolated from Duguetia vallicola wood extract.
Figure 1.
Structures of liriodenine (1), O-methylmoschatoline (2) and nornuciferine (3), isolated from Duguetia vallicola wood extract.
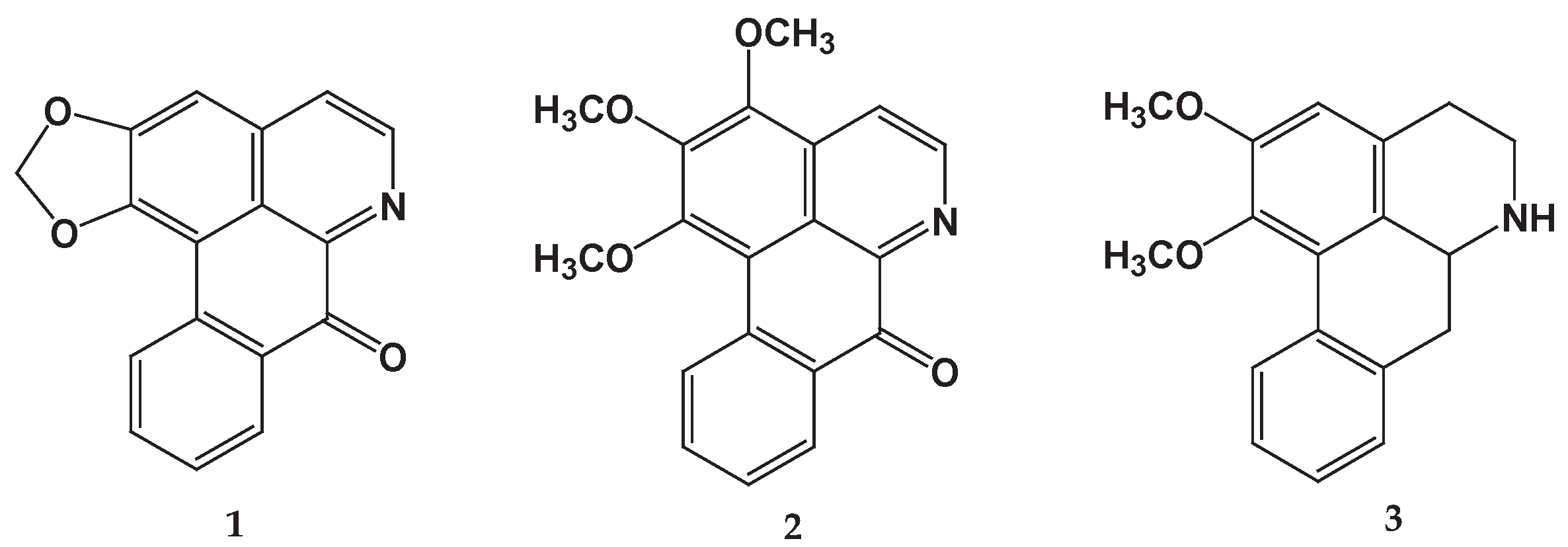
Figure 2.
Reduction in the growth of P. aeruginosa isolates exposed to ethanolic wood extract. A positive linear relationship was observed between the concentration of the extract and the percentage of inhibition of bacterial growth. The higher the concentration of the extract, the greater the percentage reduction in bacterial growth; this is consistent with the Pearson correlation coefficient (0.84 < r < 0.98) for all isolates. Furthermore, the hypothesis test on the correlation coefficient yields a p value < 0.05, which indicates that, with 95% confidence, there is a significant linear relationship.
Figure 2.
Reduction in the growth of P. aeruginosa isolates exposed to ethanolic wood extract. A positive linear relationship was observed between the concentration of the extract and the percentage of inhibition of bacterial growth. The higher the concentration of the extract, the greater the percentage reduction in bacterial growth; this is consistent with the Pearson correlation coefficient (0.84 < r < 0.98) for all isolates. Furthermore, the hypothesis test on the correlation coefficient yields a p value < 0.05, which indicates that, with 95% confidence, there is a significant linear relationship.
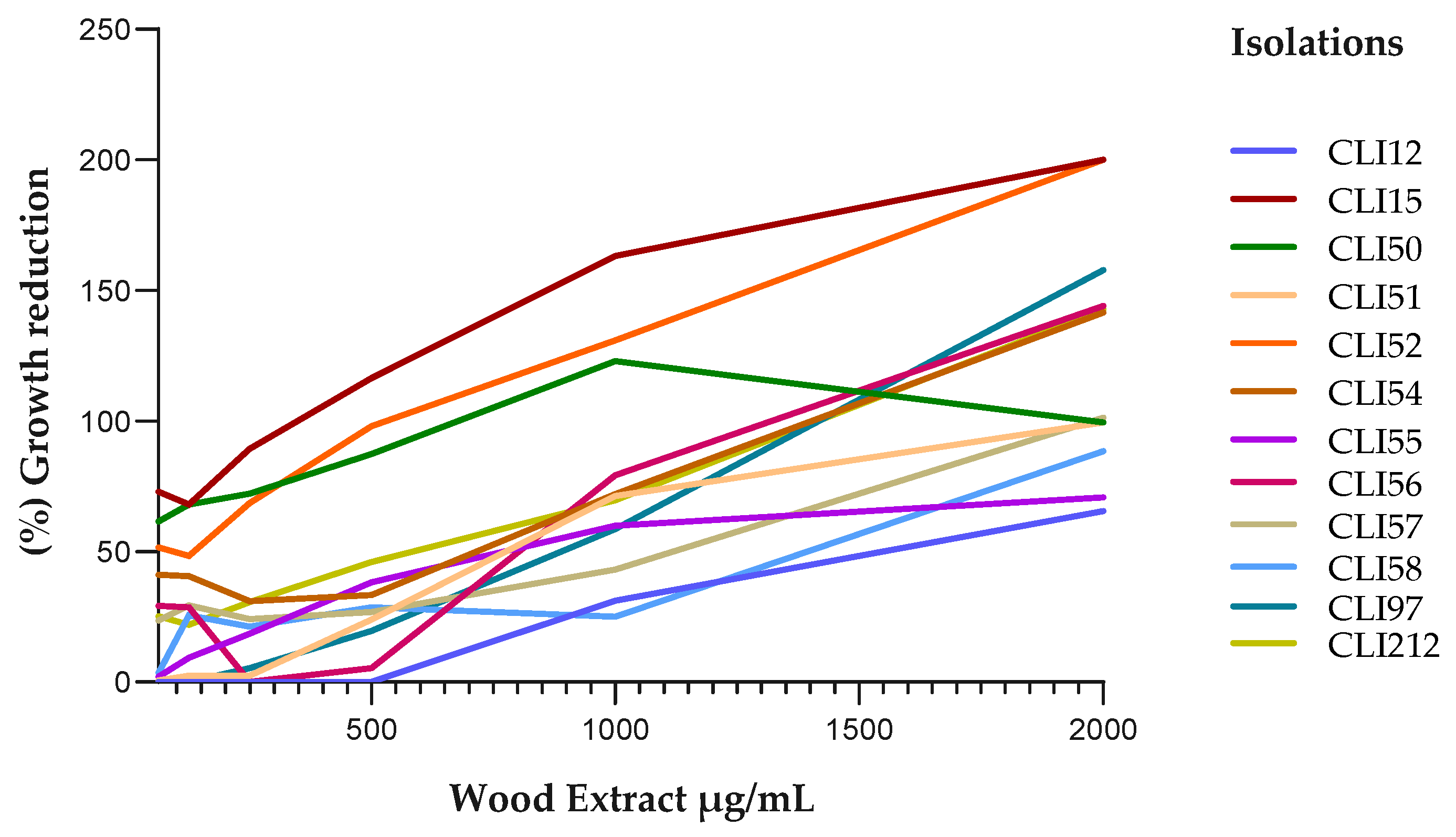
Figure 3.
Reduction in growth of P. aeruginosa isolates exposed to ethanolic bark extract. A positive linear relationship was observed between the concentration of the extract and the percentage of inhibition of bacterial growth.
Figure 3.
Reduction in growth of P. aeruginosa isolates exposed to ethanolic bark extract. A positive linear relationship was observed between the concentration of the extract and the percentage of inhibition of bacterial growth.
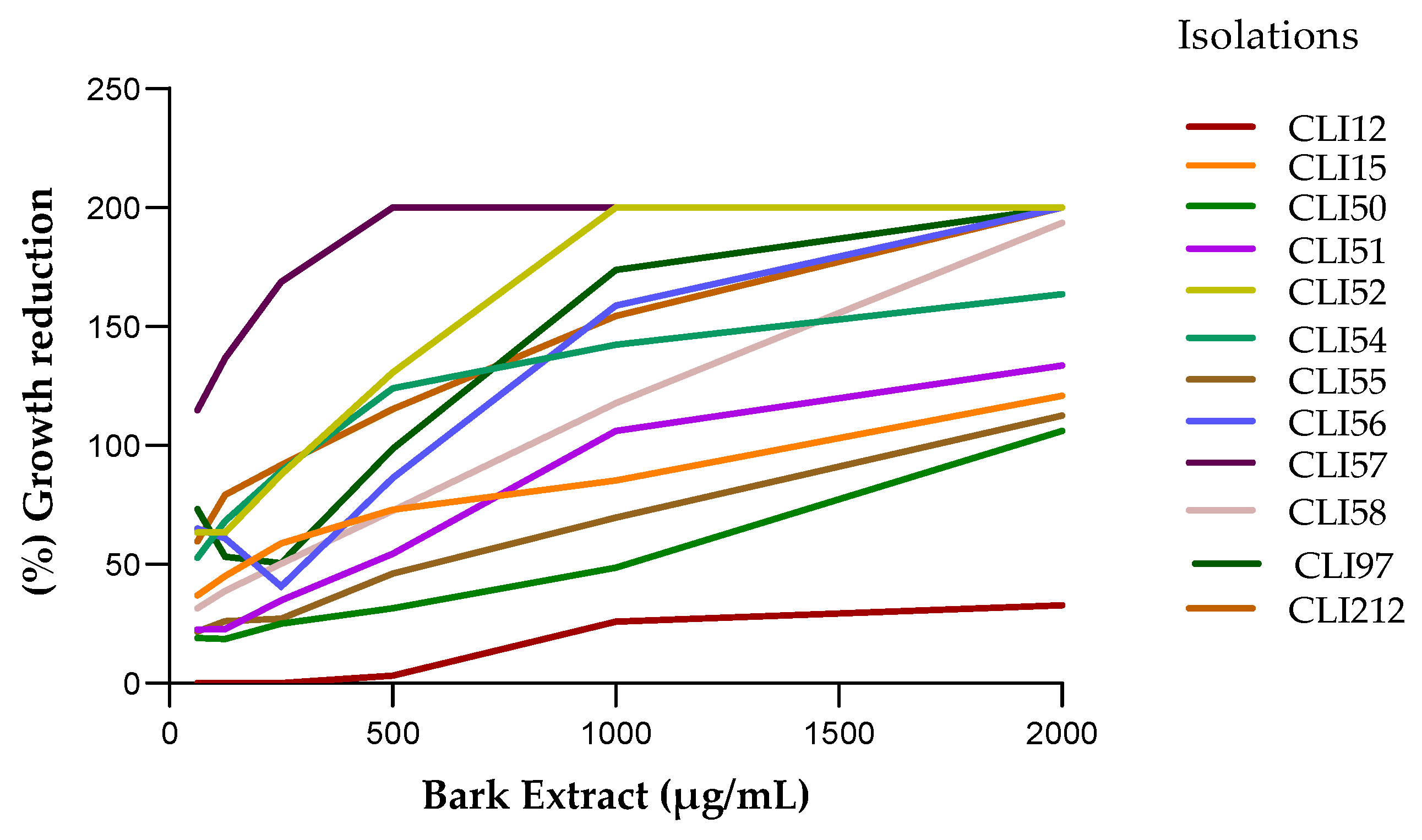
Figure 4.
Effect of bark and wood extracts on P. aeruginosa. (a) Biofilm formation at 37 °C for 24 hours. (b) Percentage reduction of mature biofilms, after 1 h of treatment with the MIC90 of each extract and CIP (2μg/mL).
Figure 4.
Effect of bark and wood extracts on P. aeruginosa. (a) Biofilm formation at 37 °C for 24 hours. (b) Percentage reduction of mature biofilms, after 1 h of treatment with the MIC90 of each extract and CIP (2μg/mL).
Figure 5.
Release of intracellular content at 260/280 nm versus time of P. aeruginosa treated with wood and bark extracts (MIC μg/mL) and CIP (2 μg/mL). The figure shows the OD260/OD280 values of the CLI15 and CLI57 isolates treated with the extracts and CIP at different times. The results are expressed as the absorbance of the sample (treated with extracts) minus the absorbance of the control (samples without extracts).
Figure 5.
Release of intracellular content at 260/280 nm versus time of P. aeruginosa treated with wood and bark extracts (MIC μg/mL) and CIP (2 μg/mL). The figure shows the OD260/OD280 values of the CLI15 and CLI57 isolates treated with the extracts and CIP at different times. The results are expressed as the absorbance of the sample (treated with extracts) minus the absorbance of the control (samples without extracts).
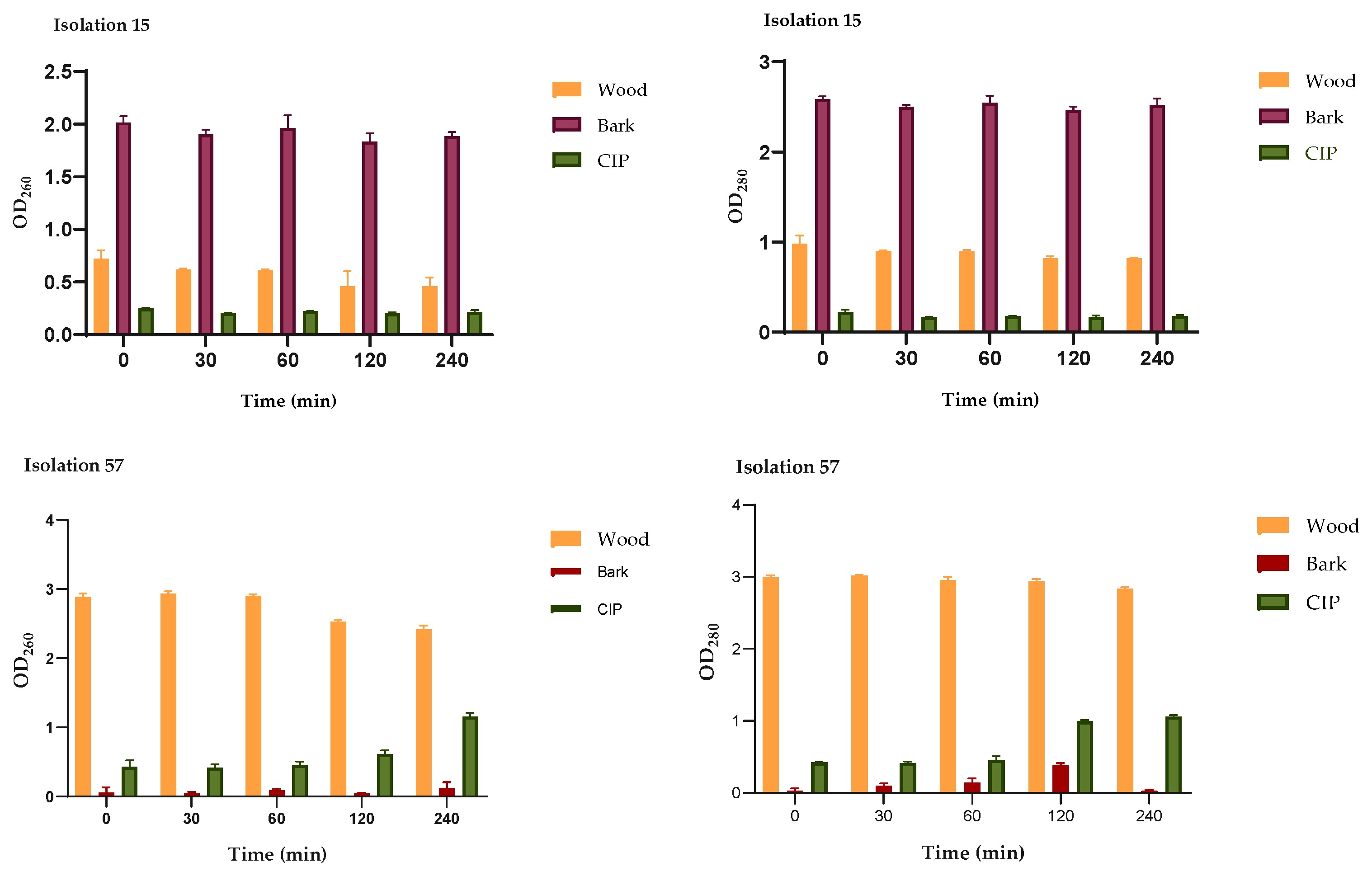
Figure 6.
Measurement of extracellular pH of P. aeruginosa treated with D. vallicola extracts, untreated cells (INO) and CIP (2 μg/mL). The results of the ANOVA indicate that there are significant differences between the extracts compared to the INO control (p < 0.0001) for all isolates. In cells treated with CIP, there are no significant differences with respect to cells without treatment (control). The Dunnett test with a confidence level of 95% indicates that there are significant differences between the effect of the extracts on extracellular pH values compared to the effect of CIP.
Figure 6.
Measurement of extracellular pH of P. aeruginosa treated with D. vallicola extracts, untreated cells (INO) and CIP (2 μg/mL). The results of the ANOVA indicate that there are significant differences between the extracts compared to the INO control (p < 0.0001) for all isolates. In cells treated with CIP, there are no significant differences with respect to cells without treatment (control). The Dunnett test with a confidence level of 95% indicates that there are significant differences between the effect of the extracts on extracellular pH values compared to the effect of CIP.
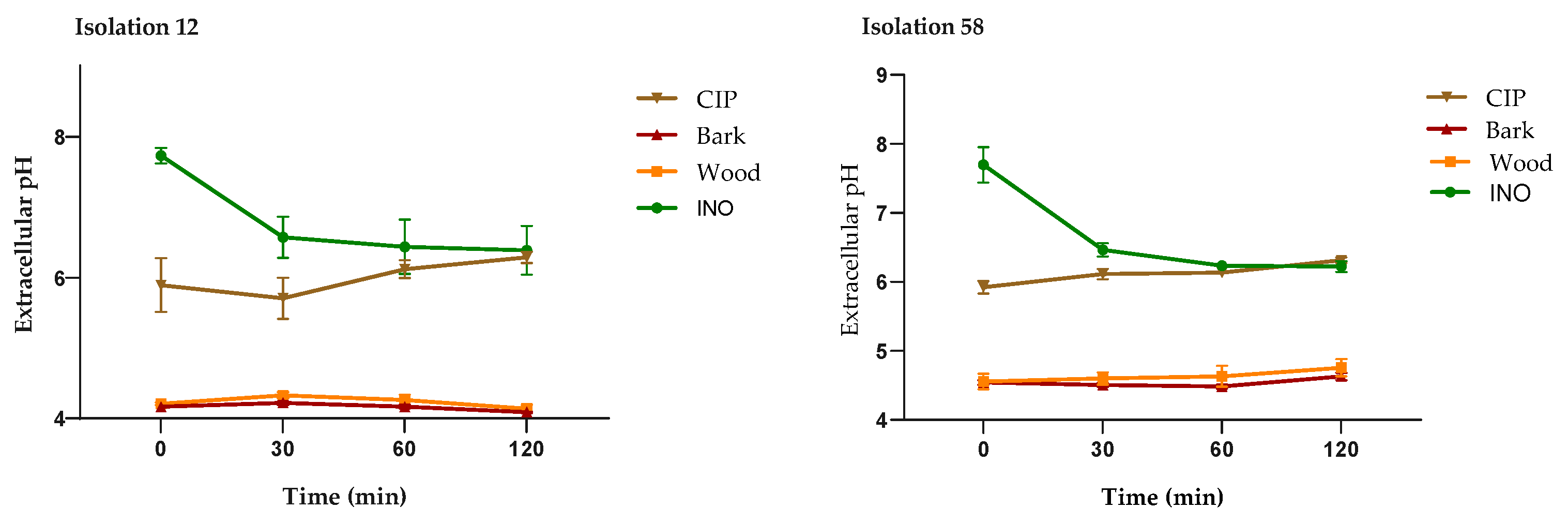
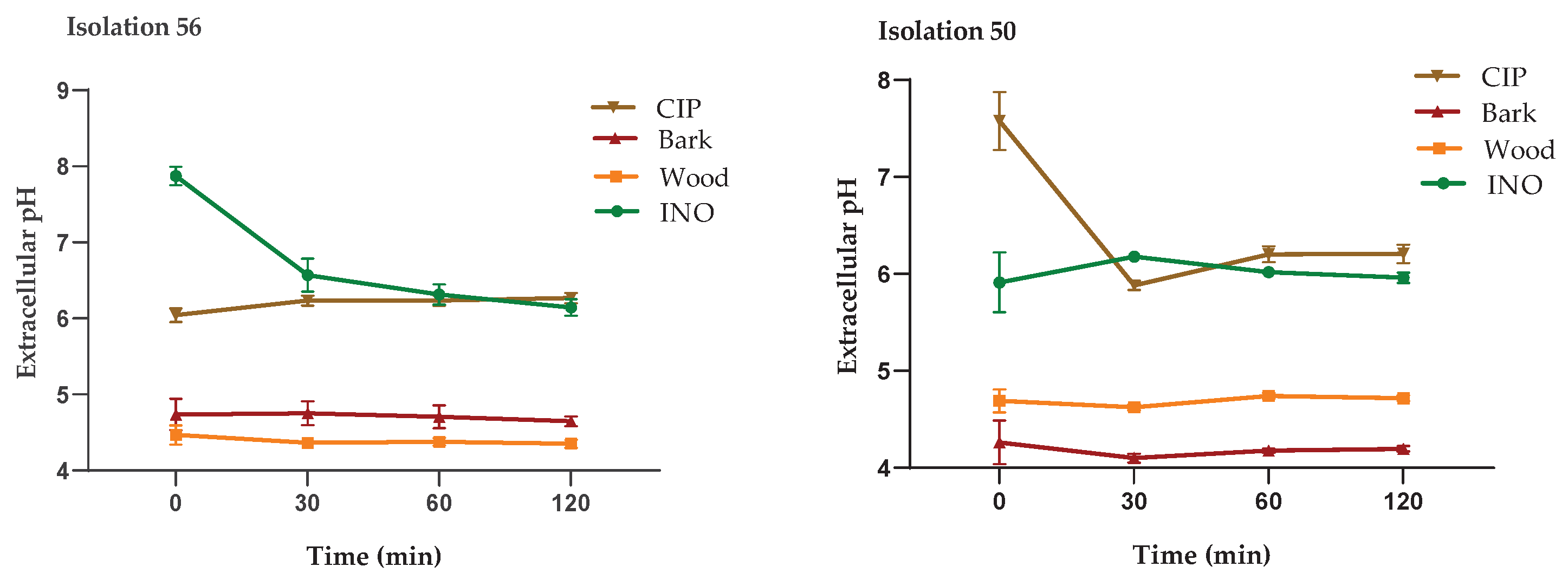
Figure 7.
Fluorescence microscopy of P. aeruginosa without treatment (control), treated with the bark and wood extracts, and treated with CIP after 24 h. Live cells with intact membranes appear green, while dead cells with damaged membranes appear red and orange.
Figure 7.
Fluorescence microscopy of P. aeruginosa without treatment (control), treated with the bark and wood extracts, and treated with CIP after 24 h. Live cells with intact membranes appear green, while dead cells with damaged membranes appear red and orange.
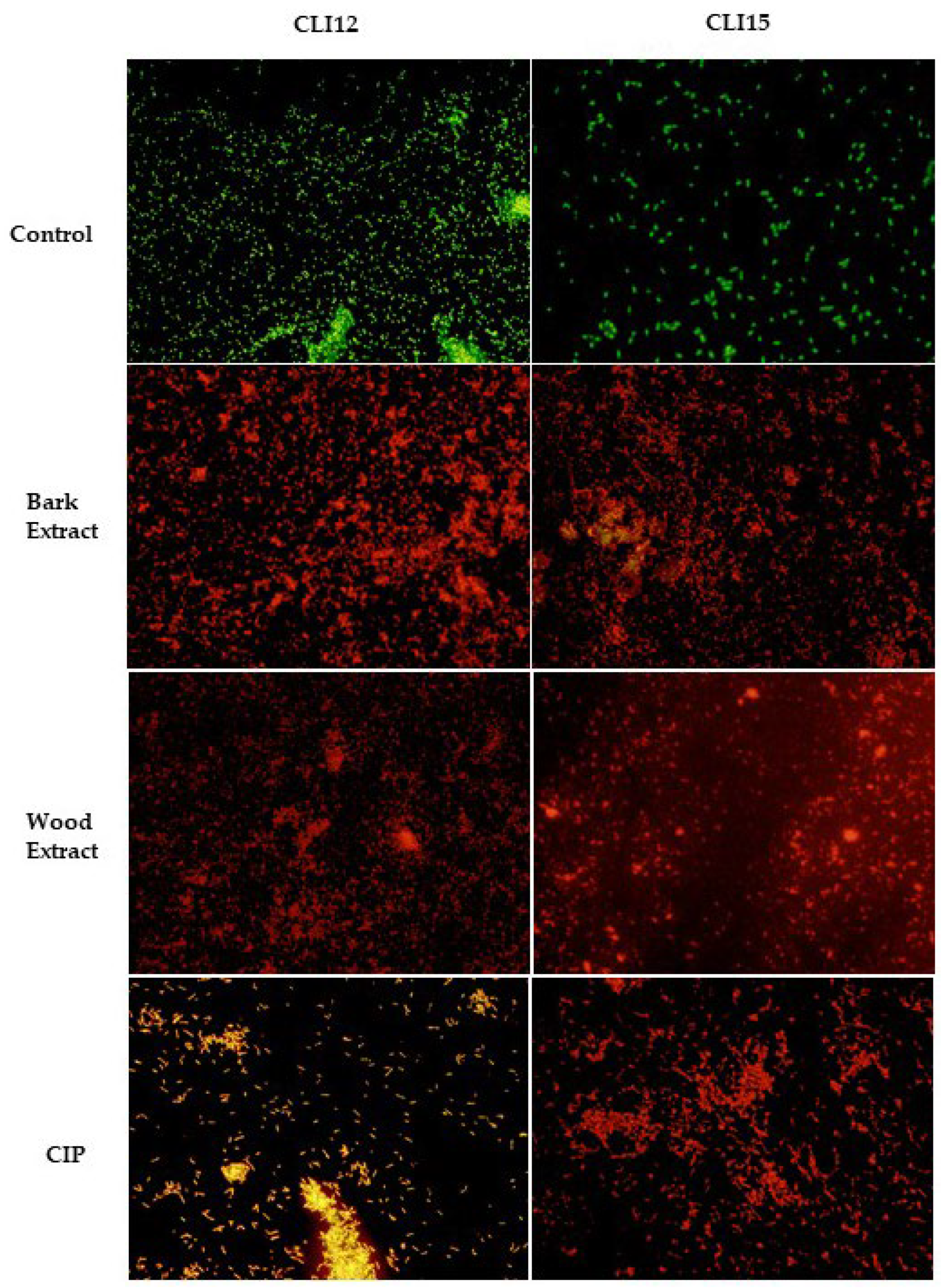

Table 1.
Minimum inhibitory concentration (MIC90) values of ethanolic extracts of wood and bark (μg/mL), against clinical isolates of P. aeruginosa..
Table 1.
Minimum inhibitory concentration (MIC90) values of ethanolic extracts of wood and bark (μg/mL), against clinical isolates of P. aeruginosa..
| P. aeruginosa | Wood Extract | Bark Extract |
| MIC90 | MIC90 | |
| CLI 12 | 3800 | 6300 |
| CLI 15 | 180 | 1460 |
| CLI 50 | 500 | 2200 |
| CLI 51 | 3000 | 1600 |
| CLI 52 | 490 | 73 |
| CLI 54 | 1270 | 300 |
| CLI 55 | 3120 | 2000 |
| CLI 56 | 1480 | 392 |
| CLI 57 | 2600 | 62.5 |
| CLI 58 | 2790 | 794 |
| CLI 97 | 1560 | 250 |
| CLI 212 | 1430 | 100 |
Table 2.
Percentages of inhibition of mature biofilms of ethanolic extracts of wood and bark vs. CIP in P. aeruginosa.
Table 2.
Percentages of inhibition of mature biofilms of ethanolic extracts of wood and bark vs. CIP in P. aeruginosa.
| P. aeruginosa | Wood | Bark | CIP |
| CLI12 | 11.51 | 44.79 | 33.03 |
| CLI15 | 0.00 | 0.00 | 0.00 |
| CLI50 | 0.00 | 38.97 | 64.09 |
| CLI51 | 37.09 | 38.85 | 15.49 |
| CLI52 | 20.91 | 25.51 | 77.19 |
| CLI54 | 0.00 | 0.00 | 0.00 |
| CLI55 | 15.20 | 16.07 | 11.28 |
| CLI56 | 0.00 | 0.00 | 31.41 |
| CLI57 | 56.79 | 0.00 | 64.19 |
| CLI58 | 57.58 | 31.80 | 39.07 |
| CLI97 | 54.31 | 48.46 | 0.00 |
| CLI212 | 93.74 | 0.00 | 26.63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



