
Submitted:
28 March 2024
Posted:
28 March 2024
You are already at the latest version
Abstract
Extensive green roofs provide for many ecosystem services in urban environments. The efficacy of these services is influenced by the substrate characteristics and vegetational composition. Despite their key role in plant performance and productivity, or their contribution to nitrogen fixation or carbon sequestration, green roof microbial communities have received little attention so far. Here, we investigated whether microbial communities in extensive green roof substrates contain a core community, as well as whether they are affected by green roof plant diversity. Eleven green roofs, planted either with a mixture of grasses, wildflowers and succulents (Sedum spp.), or solely species of Sedum, were seasonally sampled to investigate prokaryotic and fungal communities via metabarcoding. Although Sedum-herbs-grasses roofs contained more plant species, we observed no discriminant microbial communities between both roof types, likely due to sharing the same substrate textures and the vegetational layers that became more similar throughout time. Examining key microbial taxa revealed that most taxa are also dominant phylotypes in soils across different biomes. Many bacterial core taxa are capable of nitrogen fixation and most fungal key taxa are stress-tolerant saprotrophs, endophytes, or both. Microbial community composition further revealed high similarities with aerial samples taken in urban environments. Considering the extreme edaphic conditions that periodically arise in green roof substrates, further investigation is warranted to determine the extent to which the dynamic and intriguing microbial communities are influenced by the aeromicrobiome.
Keywords:
bacteria
; fungi
; microbes
; urban
; green roof
; Aeromicrobiome
1. Introduction
Green roofs, or vegetated roofs, are increasingly recognized as valuable urban green infrastructures due to their contribution to a number of ecosystem services in built-up environments. For example, green roofs retain stormwater runoff [1,2,3], remove air pollution [4], cool down building interiors [5,6], mitigate heat island effects [7,8], and conserve urban biodiversity [9,10,11]. The extent to which green roofs contribute to the above-mentioned ecosystem services depends on the characteristics of the substrate layer and the vegetational composition.
Traditionally, green roofs are classified based on the thickness and composition of the substrate layer, the type of vegetation, and their primary function [12]. Most of the installed green roofs are “extensive” green roofs, which comprise thin layers (<15 cm) of porous mineral substrate low in organic matter (<10%). They are primarily installed to retain stormwater runoff, and their lightweight substrates offer the advantage that the roofs do not require costly structural reinforcements to the buildings. The composition of such substrates, however, results in a vegetational layer that is adapted to the xerothermic conditions that periodically arise [13,14]. As a result, extensive green roofs are typically planted with either monocultures of succulents (species of Sedum) or succulents in combination with xerotolerant herbaceous species and grasses [15].
Green roof plant communities are not only shaped by the physio-chemical parameters of the substrate [16,17,18], but they will also be affected by the associated microbes in and around their roots. Soil microbial communities are known to vary in response to plant composition. Plant life forms, determined by plant history traits, play a significant role in shaping microbial community composition, which can be attribute to factors such as microbial-host specificity [19,20,21] or plant root structural differences [22]. Thus, it is expected that a greater plant diversity on extensive green roofs will result in more diverse and enriched microbial communities. Furthermore, soil bacteria and fungi are known to affect plant communities. For example, they shape plant diversity [23,24,25], contribute to overall plant performance and productivity [26,27,28], and influence plant functional traits [29,30]. Consequently, in green roofs, microbes can improve plant resilience to stress induced by the adverse edaphic conditions [31,32], resulting in improved ecosystem services. In addition to supporting the vegetational layer, microbes are integral to many other ecosystem services such as nitrogen-fixation [33,34], carbon sequestration [35,36], or decomposition [37], underscoring the importance of examining microbial diversity in green roof substrates.
While numerous studies have examined the application of inoculants in extensive green roof substrates [38,39,40,41,42,43,44], there has been comparatively limited research into identifying the trends that shape microbial communities within these novel urban habitats. The few studies that investigated green roof microbial assemblages via metabarcoding revealed that the communities are diverse and compositionally distinct from other urban green infrastructures, such as parcs or bioswales [45,46,47,48]. Community assembly is believed to be primarily driven by environmental filtering rather than stochastic processes [46] although the surrounding environment has also been found to play an integral role in structuring green roof microbial communities [49]. Microbial communities have been linked to the roof vegetation in some studies [50], but not in all [45]. This is not surprising, considering the variation in green roof construction. More research is warranted to elucidate the main drivers behind microbial community assemblages in the substrates of green roofs. Furthermore, what remains lacking is an overall picture of the core microbiome, i.e., a set of taxa that consistently occur within green roof substrates, especially since revealing the core microbiome and its ecological roles is crucial for understanding ecosystem services.
Here, we add to the limited body of knowledge about microbial communities in green roof substrates by employing 16S ribosomal RNA (Prokaryota) and internal transcribed spacer 2 (ITS2; Fungi) metabarcoding. Eleven extensive green roofs were selected across three cities in Flanders (Belgium) and sampled once throughout all four succeeding seasons to identify the core microbial taxa residing in the substrates. Furthermore, we selected green roofs that were either planted with species of Sedum or a combination of Sedum, wildflowers and grasses to examine whether plant diversity affect microbial community composition.
2. Materials and Methods
2.1. Study Area and Green Roof Characteristics
This study was performed in the Flemish Region of Belgium, Western Europe. The area has a temperate maritime climate. Throughout the duration of the study, a mean annual temperature of 11.5 °C was recorded, and the average rainfall amounted to 798.6 mm/m2 [51]. We selected eleven extensive green roofs (Table 1) in three cities that are up to 100 km away from each other: Antwerp (51.2194°N, 4.4024°E), Ghent (51.0543°N, 3.7174°E) and Hasselt (50.9307°N, 5.3325°E). They are constructed on flat roofs using commercial extensive green roof substrate, i.e., mineral substrate (mainly pumice and crushed brick) low in organic matter content (<10%) (Figure S1). The roofs differ in the vegetation that was initially planted: either only species of Sedum (Sedum-moss roofs) or species of Sedum combined with wildflowers and grasses (Sedum-herbs-grasses roofs) [12]. All roofs differ in the size of their vegetated areas (25 - 777 m2), their year of construction (4 - 15 years before the start of our study), and the height at which they are located (3.2 - 22.3 m above ground level). However, the average values of these variables do not differ between both roof types.
2.2. Data Collection and DNA Extraction
Plant species richness on all green roofs was assessed by identifying the growing forbs and grasses to the species level in May, June, August and September 2019. Samples for microbial analyses were collected in three consecutive days (one day per city) during four occasions (once per season): in April, July and October 2019, and in January 2020. Three samples were collected from every roof in April, except for roof 11 (R11; not accessible at that moment). In the remaining months four samples were taken per roof, resulting in a total of 162 samples.
The samples were collected by randomly placing plastic squares of 30 x 30 cm on every roof (Figure 1). Photographs were taken to assess the percentage cover of i) Sedum, ii) wildflowers and grasses and iii) bare substrate. Prior to sampling the substrate, the vegetational layer was removed. For each sample, the top 5 cm of substrate was collected and transferred to sterile plastic bags. Subsequently, the total substrate depth was measured within the plots. The substrate samples were kept on ice during transportation and stored at 4 °C for one week until further processing.
Substrate samples were sieved in the lab at 2 mm. Quadruplicate aliquots of 250 mg were weighted and transferred to Eppendorf tubes, resulting in 648 samples. These samples were stored at -21 °C until DNA extraction, which was performed using DNeasy PowerSoil Pro kits (Qiagen, Hilden, Germany) following the manufacturer’s protocol. One empty disruption tube was included to serve as a negative control, while another tube containing the ZymoBIOMICS Microbial Community DNA Standard (Zymo Research, Irvine, CA, USA) was included as a positive control.
2.3. Metabarcoding Substrate Microbial Communities
Prokaryotic communities were investigated by amplifying the hypervariable V4 region of the 16S rRNA gene using the primers 515F (5`- GTGYCAGCMGCCGCGGTAA) and 806R (5’-GGACTACHVGGGTWTCTAAT) [52]. Fungal communities were examined via the hypervariable internal transcribed spacer 2 (ITS2) region between the 5.8S and 28S rRNA genes using the primers gITS86F (5’- GTGARTCATCGARTCTTTGAA) and ITS4R (5’- TCCTCCGCTTATTGATATGC) [53]. All primers had Nextera transposase adapters (Illumina, San Diego, CA, USA) attached to them. PCR reactions were conducted in 25 μl containing 1× Q5 reaction buffer (New England Biolabs, Ipswich, MA, USA), 200 μM dNTPs, 0.25 μM of both primers, 0.02 U/μL Q5 Hot-Start High Fidelity DNA polymerase, and 1 μL of DNA template. The thermocycling conditions consisted of an initial denaturation at 98°C for 150 s – 28 cycles of 10 s at 98 °C, 30 s at 55/57 °C (Prok/Fun), 30 s at 72 °C – a final extension for 7 min at 72 °C. Downstream detection of the PCR products was carried out by agarose gel electrophoresis, and quadruplicate samples were pooled based on the intensity of the bands. Pooled PCR products were purified using AMPure XP beads (Beckman Coulter, Brea, CA, USA) and subsequently indexed using Nextera XT indices (Illumina, San Diego, CA, USA). The 25 μL PCR mixtures contained 1× Q5 reaction buffer, 200 μM dNTPs, 0.02 U/μL Q5 Hot-Start High Fidelity DNA polymerase, 2.5 μL of both indices, and 1 μL of DNA template. Amplification conditions were as follows: 98 ̊C for 3 min – 11x (98 ̊C for 10 s / 55 ̊C for 30 s / 72 ̊C for 30 s) – 72 ̊C for 7 min. Indexed samples were purified (as described above) and quantified with a Qubit dsDNA HS assay kit on a Qubit 2.0 fluorometer (Thermo Fisher Scientific, Waltham, MA, USA). Purified indexed samples were brought to equimolar concentration, pooled and sequenced on a MiSeq system at Hasselt University (Belgium) using a MiSeq Reagent Kit v3 (Illumina, San Diego, CA, USA), which enables a maximum read length of 2x300 bp.
2.4. Data Analysis
R v4.1.2 [54] was used to perform the analyses. First, primers were removed from the raw amplicon sequencing data via the R-package cutadapt v2.9 [55]. Next, reads were processed into exact amplicon sequence variants (ASVs) via dada2 v1.22.0 [56]. Based on the quality profiles of the reads, we truncated the forward reads at position 230/230 (Prokaryota/Fungi) and the reverse reads at position 120/160. The maximum number of expected errors was set to 1 and the number of ambiguous nucleotides to 0. We used standard parameters in all subsequent steps (error rate-learning, sample inference, merging and chimera removal). Unique ASVs were aligned against the UNITE v9.0 reference database for Fungi [57] or SILVA v138.1 for Bacteria and Archaea [58]. The R-package phyloseq v1.38.0 [59] was used to merge the ASV-feature table, taxonomy table and table containing the metadata.
Prior to performing the analyses we pre-processed the data. Control samples (negative controls and mock communities) were investigated and discarded from the dataset. ASVs that were not assigned at the level of phylum were discarded, since many were found to belong to plant species when blasted against GenBank [60]. Bacterial ASVs identified as chloroplasts or mitochondria were also discarded. Samples were rarefied to account for differences in library size, following the investigation of sequencing depth via rarefaction curves through the R-package vegan v2.5.7 [61].
We compared the microbial communities of both roof types by examining alpha- and beta-diversity metrics. Regarding alpha-diversity, ASV-richness and Shannon index were calculated via vegan, and Faith’s phylogenetic diversity (PD) via picante v.1.8.2 [62]. Significant differences in all three metrics between both roof types were measured via Wilcoxon rank-sum tests. Principal coordinate analysis (PCoA) was performed on Weighted UniFrac distance matrices [63] to compare the microbial community compositions (beta-diversity) of both roof types. The effect of roof type on community composition was tested using permutational analysis of variance (PERMANOVA, R-package vegan) after validating homogeneity of dispersion. Figures were made via ggplot2 v3.3.6 [64].
To further characterize the microbial communities we identified key (core and discriminant) microbial taxa. Core taxa were considered to be ASVs that are present in at least 75% of all samples taken, having a relative abundance of at least 0.1%. Discriminant taxa were investigated via linear discriminant analysis (LDA) effect size (Lefse) via the R-package microbiomeMarker v1.3.3 [65]. Prior to running the lefse LDA-analyses, we transformed the rarefied table to total sum scaling (relative abundance) and normalized the sum of the values to 1M. We also added an individual identity to each ASV to prevent the script from merging all ASVs belonging to the same genus. Wilcoxon rank-sum tests (cutoff at 0.01) were used to identify differentially abundant features between both roof types, and linear discriminant analysis (LDA) estimated the effect size for the significant features (threshold score of 4.0).
3. Results
3.1. Sedum-Moss Roofs vs Sedum-Herbs-Grasses Roofs
We identified all growing plant species on the eleven extensive green roofs during four occasions in the growth season (May, June, August and September 2019) (Table S1). When summed up, Sedum-herbs-grasses roofs have significantly more herbs and grasses than Sedum-moss roofs (p-value 0.014; Figure 2a). The vegetational layers were further characterized by measuring the plant coverage (excluding mosses) on top of the plots that were sampled for the microbial analyses. On average, total plant coverage between both roof types does not differ (p-value 0.202; Figure 2b), but herbs and grasses account for a higher percentage on Sedum-herbs-grasses roofs (p-value < 0.001; Figure 2c). Simultaneously we measured the substrate depth of the plots. The substrate layer is, overall, significantly thicker in Sedum-herbs-grasses roofs compared to Sedum-moss roofs (p-value <0.001, Figure 2d).
3.2. Prokaryotic Diversity
Samples were rarefied to 4,352 reads to account for uneven sampling depths (Figure S2a). Eight samples were excluded from the dataset due to an insufficient number of reads. Across the remaining 154 samples (670,208 reads), 8,308 prokaryotic ASVs (of which 78 archaeal) are discovered (average amplicon length 253 ± 2 bp). The taxonomic classification of the inferred ASVs via the SILVA database reveals three dominant phyla across all investigated green roofs, i.e., Proteobacteria (31.4%; 1,738 ASVs), Actinobacteriota (23.8%; 1,167 ASVs) and Acidobacteriota (14.5%; 774 ASVs) (Figure 3a). Extending the results to the ten most prevalent prokaryotic phyla revealed six more bacterial phyla [Bacteroidota (5.8%; 1,119 ASVs), Chloroflexi (5.2%; 654 ASVs), Verrucomicrobiota (4.0%; 403 ASVs), Gemmatimonadota (4.0%; 398 ASVs), Myxococcota (3.8%; 377 ASVs), Planctomycetota (3.4%; 797 ASVs)] and one archaeal phylum: Crenarchaeota (1.3%; 47 ASVs).
None of the calculated alpha-diversity metrics differ statistically significantly between Sedum-moss roofs (n=72) and Sedum-herbs-grasses roofs (n=82), i.e., ASV richness (p-value 0.73), Shannon-index (p-value 0.99) and Faith’s phylogenetic diversity (p-value 0.30) (Figure 4a-c). The prokaryotic communities of both roof types differ compositionally significantly (p-value 0.001, R2 0.062) and samples taken from Sedum-moss roofs have more variation in their community compositions than samples from Sedum-herbs-grasses roofs (p-value 0.003) (Figure 4d).
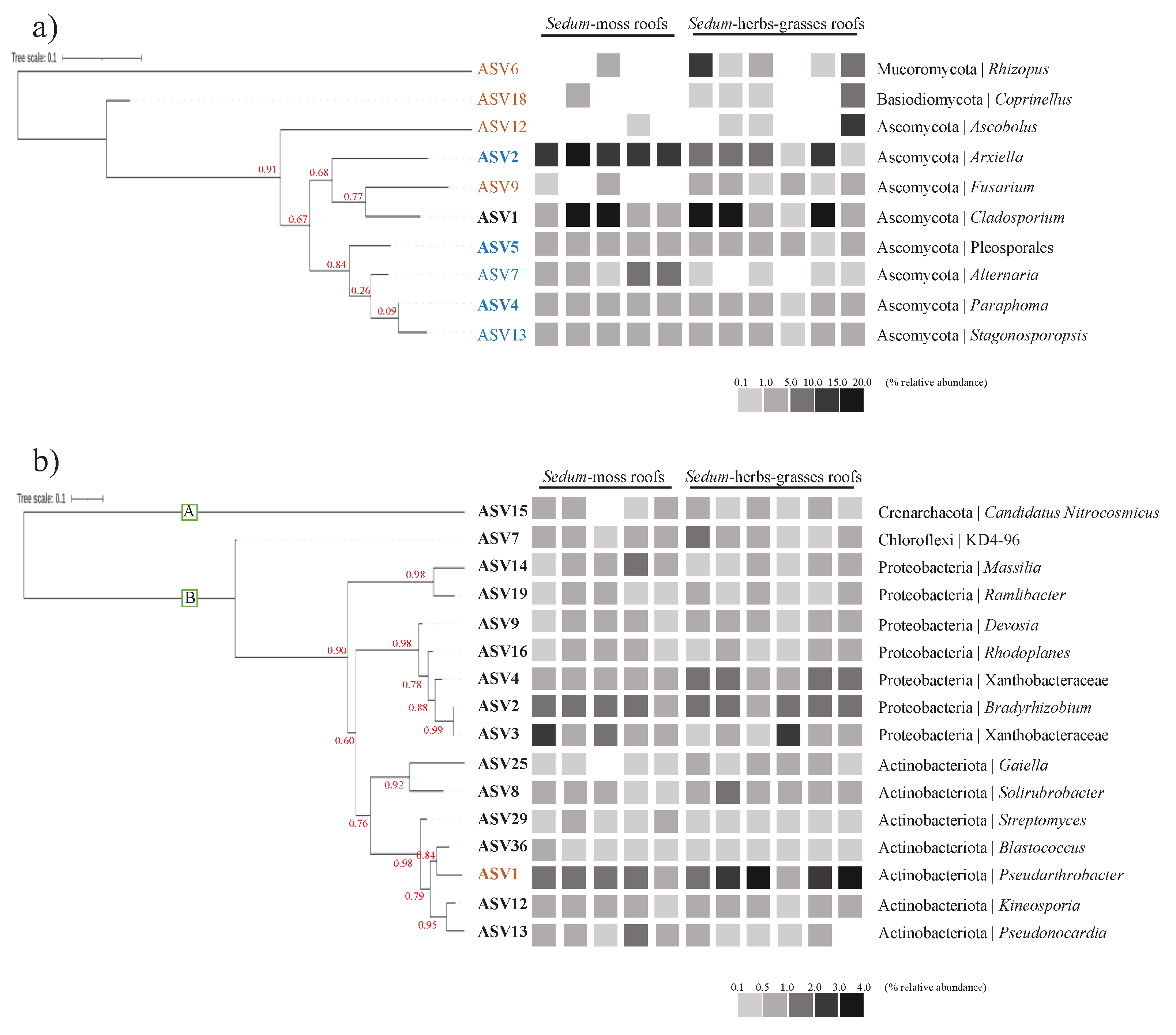
Regarding the key prokaryotic taxa, 16 taxa are found to be present in at least 75% of all samples, having a relative abundance ≥ 0.1%: 15 bacterial core taxa and 1 archaeal core taxon (Figure 5a). Pseudarthrobacter sp. is the only ASV found to be discriminant between both roof types, being more enriched in Sedum-herbs-grasses roofs.
Figure 3.
Relative abundances of the main prokaryotic (a) and fungal (b) phyla.
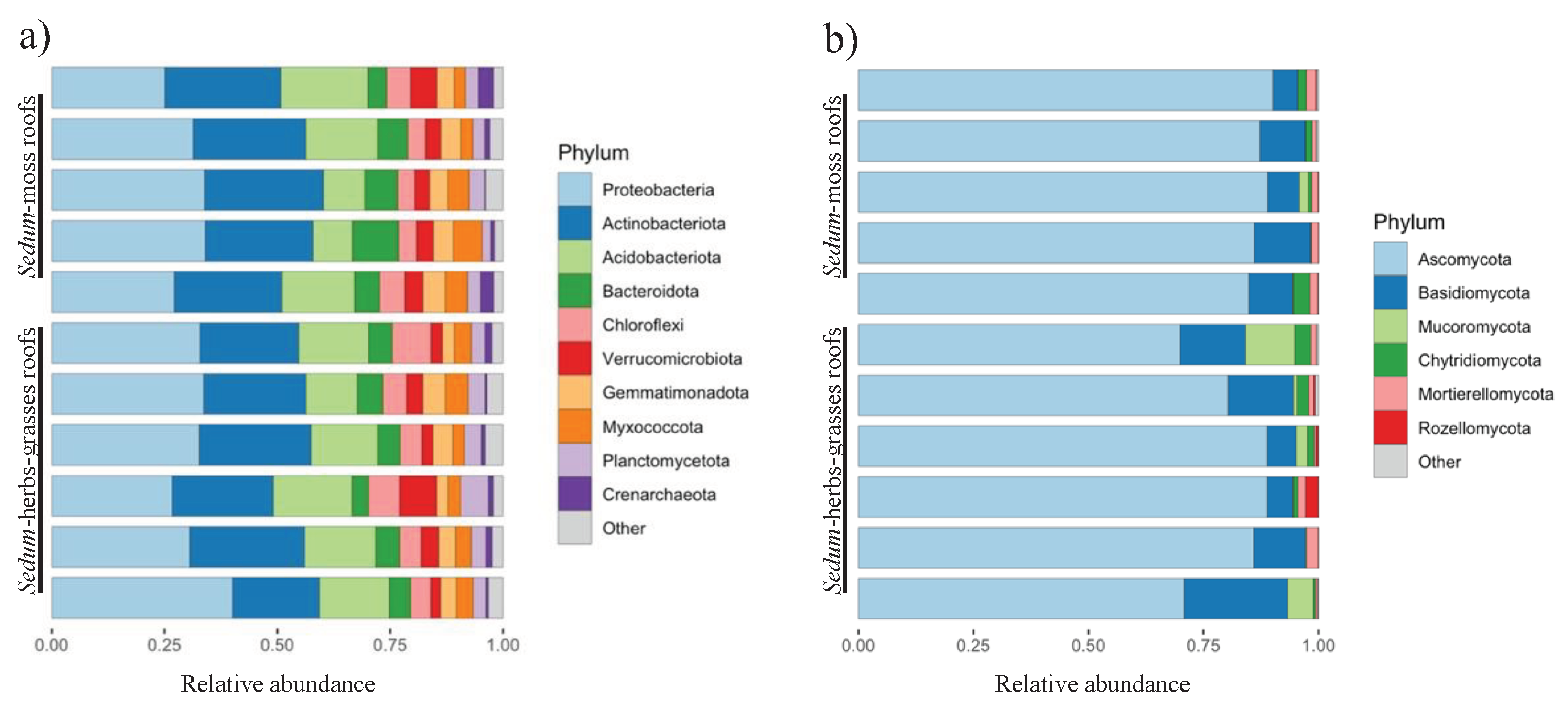
Figure 4.
Diversity metrics for prokaryotic (a-d) and fungal (e-h) communities in Sedum-moss roofs (blue) and Sedum-herbs-grasses roofs (orange). Boxplots span the interquartile range (i.e., the range between the 25th to 75th percentile), lines within boxes denote the median. Circles denote 95% confidence intervals. No significant differences are found between the alpha-diversity metrics of both roof types (p-value > 0.05; Wilcoxon rank-sum test).
Figure 4.
Diversity metrics for prokaryotic (a-d) and fungal (e-h) communities in Sedum-moss roofs (blue) and Sedum-herbs-grasses roofs (orange). Boxplots span the interquartile range (i.e., the range between the 25th to 75th percentile), lines within boxes denote the median. Circles denote 95% confidence intervals. No significant differences are found between the alpha-diversity metrics of both roof types (p-value > 0.05; Wilcoxon rank-sum test).
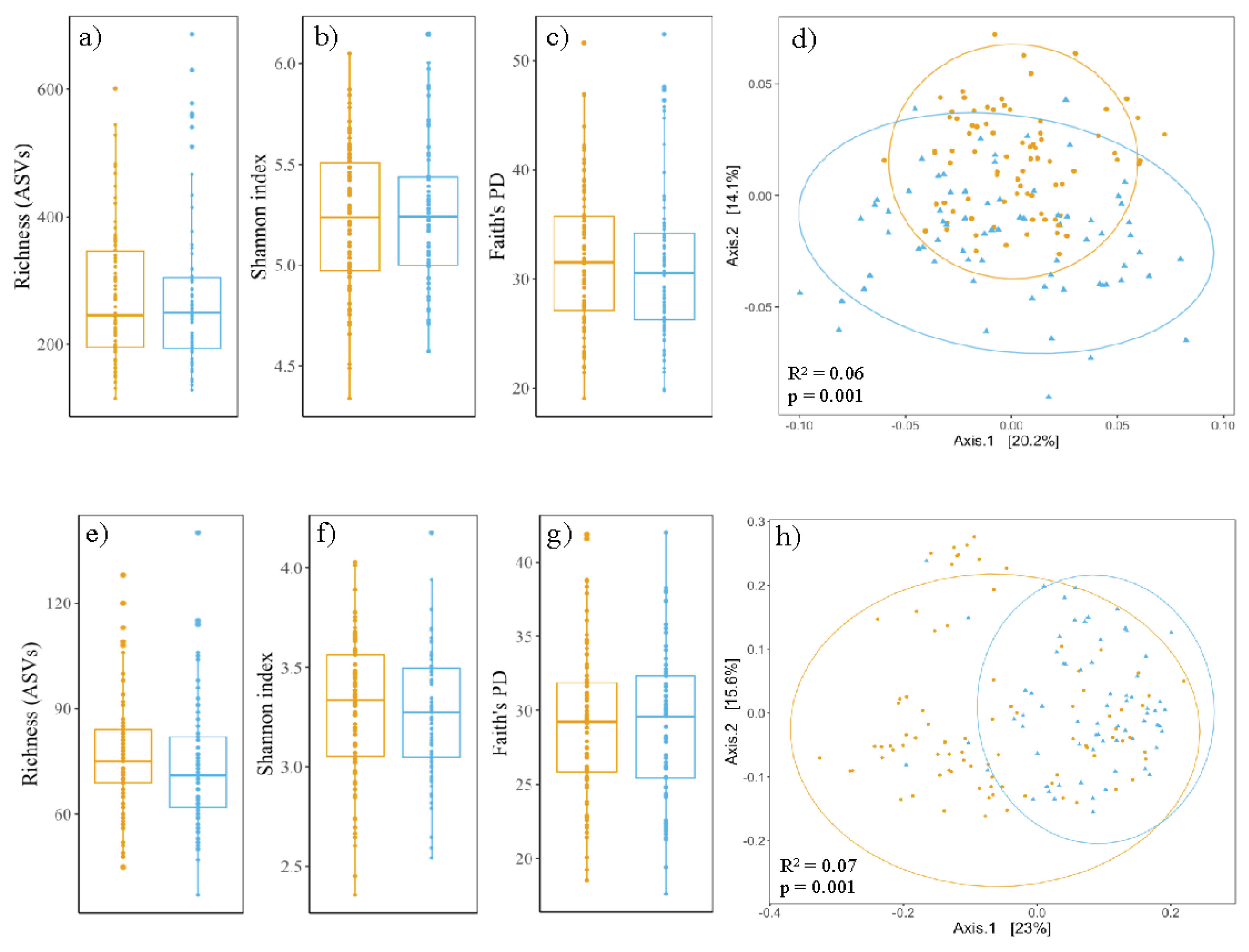
Figure 5.
Key fungal (a) and prokaryotic (b) taxa (ASVs). Core taxa, having a relative abundance of >0.1% in at least 75% of all samples, are displayed in bold. Microbial ASVs that are more abundantly present, as investigated via LEfSe (p < 0.01, LDA log10 scores ≥4.0), in Sedum-moss roofs are displayed in blue, those in Sedum-herbs-grasses roofs in orange. The numbers of the ASVs correspond to their relative abundances in the full dataset, with 1 being the most abundantly present ASV. The heatmaps show the average relative abundance of each key ASV per roof. Support values for the nodes are displayed in red.
Figure 5.
Key fungal (a) and prokaryotic (b) taxa (ASVs). Core taxa, having a relative abundance of >0.1% in at least 75% of all samples, are displayed in bold. Microbial ASVs that are more abundantly present, as investigated via LEfSe (p < 0.01, LDA log10 scores ≥4.0), in Sedum-moss roofs are displayed in blue, those in Sedum-herbs-grasses roofs in orange. The numbers of the ASVs correspond to their relative abundances in the full dataset, with 1 being the most abundantly present ASV. The heatmaps show the average relative abundance of each key ASV per roof. Support values for the nodes are displayed in red.
3.3. Fungal Diversity
Samples were rarefied to 2,331 reads (Figure S2b) to account for uneven sampling depths. Eight samples were excluded from the dataset due to an insufficient number of reads. Across the remaining 154 samples (358,974 reads), 2,671 fungal ASVs are discovered (average amplicon length 273 ± 33 bp). The taxonomic classification of the inferred ASVs via the UNITE database reveals one prevalent fungal phylum across all investigated green roofs, i.e., Ascomycota (84.0% of all reads; 1,683 ASVs), followed by Basidiomycota (10.4%; 587 ASVs), Mucoromycota (2.0%; 11 ASVs), Chytridiomycota (1.6%; 188 ASVs), Mortierellomycota (1.4%; 54 ASVs) and Rozellomycota (0.4%; 44 ASVs) (Figure 3b).
Fungal diversity analyses follow similar patterns as those observed for the prokaryotic communities. None of the calculated alpha-diversity metrics differ statistical significantly between Sedum-moss roofs (n=69) and Sedum-herbs-grasses roofs (n=85), i.e., ASV richness (p-value 0.06), Shannon-index (p-value 0.37) and Faith’s phylogenetic diversity (p-value 0.80) (Figures 4e-g). The fungal communities of both roof types differ compositionally significant (p-value 0.001, R2 0.070), but in contrast to the prokaryotic communities, fungal communities of Sedum-moss roofs have more variation in their compositions than samples from Sedum-herbs-grasses roofs (p-value <0.001) (Figure 4h).
Regarding the key fungal taxa (Figure 5b), four fungal ASVs are recovered in at least 75% of all samples, having a relative abundance ≥ 0.1%. Two ASVs are prominently abundant, i.e., Cladosporium sp. (10.4 ± 7.9 %) and Arxiella sp. (9.8 ± 6.3 %). Nine taxa were found to be discriminant: ASVs within Rhizopus, Coprinellus, Ascobolus and Fusarium are more enriched in Sedum-herbs-grasses roofs, while ASVs withing Arxiella, Alternaria, Paraphoma, Stagonosporosis and an ASV belonging to Pleosporales are more abundant in Sedum-moss roofs.
4. Discussion
Our study aimed to characterize the microbial communities residing in the substrates of extensive green roofs by investigating the presence of a core microbiome (ASVs). Furthermore, since species of Sedum do not root deeply into the substrate [66] and many plant species have host-specific microbes in their rhizobiomes [67,68], we investigated whether extensive green roofs planted with a combination of succulents (species of Sedum), flowers and grasses (i.e., Sedum-herbs-grasses roofs) harbor microbial communities that are more enriched and compositionally different from those in extensive green roofs solely planted with species of Sedum (i.e., Sedum-moss roofs).
Overall, prokaryotic communities in extensive green roof substrates are dominated by three phyla, i.e., Proteobacteria, Actinobacteriota and Acidobacteriota. This corresponds to the results obtained in previous green roof studies [46,49]. Among the 8,308 ASVs, we identified 15 prokaryotic core taxa, comprising one archaeal and 14 bacterial ASVs. Considering that few bacterial taxa are shared between any pair of unique soil samples [69,70], the low number of core taxa retrieved is not unexpected. Within the bacterial core community, a significant portion is composed of ASVs withing the orders Rhizobiales and Burkholderiales, taxa that have also been found in a previous green roof study [49]. Furthermore, that study recovered abundant nitrogenase (nifH) genes affiliated with Rhizobiales, which suggest active nitrogen fixation by these strains. Also, our results show similarities with a study that recently examined bacterial communities in soils from nine distinct biomes across the world [71]. While the amount of variation in the relative abundances of major bacterial phyla across the biomes they examined is substantial, their overall distributions align with our findings. Our results differ only in an increased relative contribution of Myxococcota and Crenarchaeota (Archaea). However, they excluded reads assigned to Archaea prior to the analyses. Another study exploring soils from a variety of terrestrial ecosystems did yield comparable findings related to Archaea [72]. As for Myxococcota, it is worth nothing that at the time of their study, this phylum was still affiliated with the class Deltaproteobacteria [73], potentially explaining why it did not emerge as a dominant phylum. Further comparing their results to ours revealed that, expect for the ASVs assigned to KD4-96 (Chloroflexi), Gaiella, and Pseudoarthrobacter, all bacterial core taxa identified in our study were also found to be dominant in soils globally.
Regarding the effect of the vegetational layer on substrate microbial community composition, Sedum-moss roofs and Sedum-herbs-grasses roofs do not differ in any of the investigated alpha-diversity metrics, neither for the prokaryotic nor for the fungal communities. The composition of prokaryotic and fungal communities is, however, affected by the type of roof but the amount of variation that could be explained by this variable is relatively low (6.2% and 7.0%, respectively). Consequently, we assume that the two types of extensive green roof studied by us do not differ enough from each other to host significantly different substrate microbiomes. Indeed, across all roofs, the composition of the substrates used is nearly the same and, although we observed significantly thicker substrate layers in Sedum-herbs-grasses roofs than in Sedum-moss roofs, the absolute difference is probably too small to exert any substantial effect. Regarding the vegetational layer, the Sedum-herbs-grasses roofs included in this study have indeed more plant species than Sedum-moss roofs, which is also reflected in higher cover percentages of herbs and grasses. However, a significant number of plant species on Sedum-herbs-grasses roofs are species that are also found to have colonized Sedum-moss roofs (Table S1). Although we do not have a list of plant species that were initially planted on the roofs, we suspect that many species disappeared due to the extreme edaphic conditions that periodically arise on these roofs. Our results contradict those from Hoch et al. [50] who, having a similar setup, did observe different fungal communities in extensive green roofs with wildflowers, grasses and Sedum compared to those in roofs planted solely with species of Sedum. However, their study design differs from ours. While we sampled randomly, they sampled either next to Sedum or next to species within Asteraceae upon comparing both roof types. Consequently, many of the indicator taxa they retrieved from extensive green roofs planted with wildflowers fall within Glomeromycota, phylotypes of which are known to occur in high relative abundance with Asteraceae plants [74].
In our study, the number of reads belonging to Glomeromycota is negligible. At phylum level, fungal communities are dominated by Ascomycota, which corresponds to previous green roof papers [45,46,50]. Furthermore, Egidi et al. [75] examined dominant fungal phylotypes in the same soil samples from the study that investigated bacterial communities in soils globally [71], taken across 9 different biomes, and found Ascomycota to be the most phylotype-rich and abundant lineage. Through comparing whole genomes from dominant Ascomycota with less dominant, generalist fungi they uncovered a higher number of genes associated with stress-tolerance and resource uptake, indicating that dominant Ascomycota might be better in colonising a wide range of environments. This would also explain why all identified fungal core taxa in our study are Ascomycota, i.e., Cladosporium, Arxiella, Paraphoma and an ASV within the order Pleosporales. Nine fungal phylotypes are found to be discriminant between both roof types under investigation. Rhizopus, Coprinellus, Ascobolus and Fusarium are found in most Sedum-herbs-grasses roofs but in almost none of the Sedum-moss roofs. However, these phylotypes do not show similar trends in occurrence across the Sedum-moss roofs (e.g., Rhizopus occurs only on roof 3, while Coprinellus only on roof 2). Therefore, we remain reluctant in stating that roof type is the main variable responsible for their distributions, and further research is needed to clarify this. Five fungal taxa are discriminant for Sedum-moss roofs, i.e., Alternaria, Paraphoma, Stagonosporosis, Arxiella, and an ASV in Pleosporales. Although they are found in higher relative abundances on these roofs, the absolute differences between Sedum-herbs-grasses roofs are negligible. Nevertheless, all discriminant fungal taxa contribute substantially to the overall fungal communities, judged by their total relative abundances in our dataset, warranting their inclusion in the investigation of their ecological characteristics.
Reviewing the lifestyles of the fungal key taxa via FungalTraits [76] revealed that all taxa are saprobic or plant-pathogenic (endophytic), or both. Sharing multiple lifestiles can be explained by the limited size of the amplicons obtained in this study, which prevents us from identifying the taxa to the species level. For example, the most abundant fungal ASV in our study is assigned to Cladosporium and it contributes to 9.24% of all reads. However, our rarefied dataset includes 11 ASVs belonging to this genus (although their summed contribution was little more, i.e., 9.87% of all reads). Some species within Cladosporium are known endophytes, whether or not parasitic, but many more are saprophytes. Blasting the ASV on GenBank [60] was inconclusive as the amplicon showed 100% identity with multiple species within this genus. Therefore, we recommend that future studies employ long-read technologies, such as Oxford Nanopore Technologies, which greatly improves taxonomic assignment.
Furthermore, it is known that endophytic fungi can switch to a pathogenic lifestyle [77] due to, e.g., a host shift, an imbalance in nutrient exchange or microbial interactions [78]. This is interesting, given the fact that the edaphic conditions in the substrate of extensive green roofs can be extreme at moments. Frequent periods of heat and drought in summer, but also floodings during heavy rainfall, most likely induce stress to all members of the biotic component. We suspect that many key fungal taxa respond quickly to the different environmental conditions by altering their lifestyle. For example, Marttinen et al. [79] identified fungal isolates from brown, necrotic parts of mosses on green roofs and inoculated them to spreading earthmoss (Physcomitrella patens) to investigate pathogenicity. Amongst the most pathogenic fungal strains were species within Trichoderma. However, Rumble et al. [40] inoculated green roofs with Trichoderma and while bryophyte coverage was indeed found to be lower in March and July, it increased in January. Consequently, switching lifestyles depending on the specific environmental conditions would be a plausible explanation for the dominance of the key taxa, making extensive green roofs fascinating habitats to study.
Many key taxa from this study are also frequently found in aerial samples from urban environments, contributing significantly to the aeromicrobiome. For example, Cladosporium is known for its high sporulation rate [80] and has been found to be amongst the most common fungal taxa, if not the most dominant one, in aerial samples as investigated via high-throughput DNA sequencing [81,82,83,84]. The same studies also found many of the other dominant phylotypes in our study, e.g., Alternaria or Paraphoma. Furthermore, conidia of Arxiella and Fusarium, also key fungal taxa, are previously found in rainwater collected from gutters [85]. The same patterns, although to a lesser extent, are also observed for our prokaryotic key taxa. Members within the orders Rhizobiales and Burkholderiales have been identified as some of the most representative taxa in the air [86,87,88]. This raises the question to what extent the aeromicrobiome influences green roof microbial communities, as has also been posed by McGuire et al. [45]. It could partially explain the fact that many green roofs show distinct prokaryotic and fungal communities, an observation that has previously also been made [45].
Considering i) the similarities with aerial samples taken from urban environments, ii) the shallow substrate layers of extensive green roofs and, consequently, frequent periods of heat, drought but also flooding, and iii) that most fungal key taxa are stress-tolerant phylotypes, plausibly capable to switch between lifestyles, we conclude that green roof substrates harbor dynamic and intriguing microbial communities, meriting further investigation. This warrants the application of a wider array of molecular techniques. In order to elucidate the main drivers behind microbial community assembly in green roof substrates, we recommend that future studies i) employ long-read technologies to improve taxonomic resolution, ii) consider sequencing the aeromicrobiome surrounding the green roofs under investigation to investigate the impact of airborne microbes on their metabarcoding results, and iii) explore qPCR transcriptomics to identify the metabolically active component of the microbiome present.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: An overview of the substrate textures from every investigated green roof; Figure S2: Rarefaction curves; Table S1: A list of the plant species that are identified on every green roof.
Author Contributions
T.V.D., L.S., S.T., T.A. and F.R. designed the study. T.V.D. and L.S. conducted soil sampling; C.V.M. identified the plant species and calculated cover percentages. T.V.D. processed the microbial samples. T.V.D. and V.S. performed the analyses. T.V.D., V.S., T.A. and F.R. analyzed the data. T.V.D. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The authors T.V.D. and L.S. and the research were financed by Fonds Wetenschappelijk Onderzoek (FWO) via a Strategic Basic Research project (S002818N: “EcoCities: Green roofs and walls as a source for ecosystem services in future cities”).
Data Availability Statement
The prokaryotic and fungal metabarcoding samples are uploaded in the Short Read Archive (SRA) of the National Center for Biotechnology Information (NCBI) and currently processed (temporary submissionID: SUB14339010). They will be publicly available at the time of publishing.
Acknowledgments
We would like to express our sincere gratitude to Julie Claes for her technical assistance during sequencing, and to all the green roof owners for granting us permission to take samples.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- DeNardo, J.C.; Jarrett, A.R.; Manbeck, H.; Beattie, D.J.; Berghage, R. Stormwater mitigation and surface temperature reduction by green roofs. Trans. ASAE 2005, 48, 1491-1496. [CrossRef]
- VanWoert, N.D.; Rowe, D.B.; Andresen, J.A.; Rugh, C.L.; Fernandez, R.T.; Xiao, L. Green roof stormwater retention: Effects of roof surface, slope, and media depth. J. Environ. Qual. 2005, 34, 1036-1044. [CrossRef]
- Mentens, J.; Raes, D.; Hermy, M. Green roofs as a tool for solving the rainwater runoff problem in the urbanized 21st century? Landscape and Urban Planning 2006, 77, 217-226. [CrossRef]
- Yang, J.; Yu, Q.; Gong, P. Quantifying air pollution removal by green roofs in Chicago. Atmos. Environ. 2008, 42, 7266-7273. [CrossRef]
- Del Barrio, E.P. Analysis of the green roofs cooling potential in buildings. Energy and Buildings 1998, 27, 179-193.
- Jaffal, I.; Ouldboukhitine, S.-E.; Belarbi, R. A comprehensive study of the impact of green roofs on building energy performance. Renewable Energy 2012, 43, 157-164. [CrossRef]
- Alexandri, E.; Jones, P. Temperature decreases in an urban canyon due to green walls and green roofs in diverse climates. Building and Environment 2008, 43, 480-493. [CrossRef]
- Santamouris, M. Cooling the cities - A review of reflective and green roof mitigation technologies to fight heat island and improve comfort in urban environments. Solar Energy 2014, 103, 682-703. [CrossRef]
- Williams, N.; Lundholm, J.; MacIvor, J.S. FORUM: Do green roofs help urban biodiversity conservation? Journal of Applied Ecology 2014, 51. [CrossRef]
- Jacobs, J.; Beenaerts, N.; Artois, T. Green roofs and pollinators, useful green spots for some wild bee species (Hymenoptera: Anthophila), but not so much for hoverflies (Diptera: Syrphidae). Sci. Rep. 2023, 13, 1449. [CrossRef]
- Van Dijck, T.; Klerkx, H.; Thijs, S.; Rineau, F.; Van Mechelen, C.; Artois, T. Sedum as host plants for caterpillars? Introducing gut content metabarcoding to green roof research. Urban Ecosystems 2023, 26, 955-965. [CrossRef]
- FLL (Landscape Development and Landscaping Research Society e.V). Green roof guidelines: guidelines for the planning. Construction and Maintenance of Green Roofs 2018.
- Dunnett, N.; Kingsbury, N. Planting Green Roofs and Living Walls; Timber Press: Portland, Oregon, USA, 2004.
- Lundholm, J.; MacIvor, J.S.; MacDougall, Z.; Ranalli, M. Plant Species and Functional Group Combinations Affect Green Roof Ecosystem Functions. PLoS One 2010, 5, e9677. [CrossRef]
- Van Mechelen, C.; Dutoit, T.; Kattge, J.; Hermy, M. Plant trait analysis delivers an extensive list of potential green roof species for Mediterranean France. Ecological Engineering 2014, 67, 48-59. [CrossRef]
- Emilsson, T. Vegetation development on extensive vegetated green roofs: Influence of substrate composition, establishment method and species mix. Ecological Engineering 2008, 33, 265-277. [CrossRef]
- Olszewski, M.; Holmes, M.; Young, C. Assessment of Physical Properties and Stonecrop Growth in Green Roof Substrates Amended with Compost and Hydrogel. HortTechnology 2010, 20. [CrossRef]
- Ondoño, S.; Martínez-Sánchez, J.J.; Moreno, J. The inorganic component of green roof substrates impacts the growth of Mediterranean plant species as well as the C and N sequestration potential. Ecol. Indic. 2015, 61. [CrossRef]
- Kowalchuk, G.A.; Buma, D.S.; de Boer, W.; Klinkhamer, P.G.L.; van Veen, J.A. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie van Leeuwenhoek 2002, 81, 509-520. [CrossRef]
- Santos-González, J.C.; Finlay, R.D.; Tehler, A. Seasonal dynamics of arbuscular mycorrhizal fungal communities in roots in a seminatural grassland. Appl. Environ. Microbiol. 2007, 73, 5613-5623. [CrossRef]
- Bever, J.; Platt, T.; Morton, E. Microbial Population and Community Dynamics on Plant Roots and Their Feedbacks on Plant Communities. Annu. Rev. Microbiol. 2012, 66, 265-283. [CrossRef]
- Haichar, F.Z.; Marol, C.; Berge, O.; Rangel-Castro, J.I.; Prosser, J.I.; Balesdent, J.; Heulin, T.; Achouak, W. Plant host habitat and root exudates shape soil bacterial community structure. ISME J 2008, 2, 1221-1230. [CrossRef]
- Reynolds, H.; Packer, A.; Bever, J.; Clay, K. Grassroots ecology: Plant-microbe-soil interactions as drivers of plant community structure and dynamics. Special Feature Ecology 2003, 84, 2281-2291. [CrossRef]
- van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69-72. [CrossRef]
- Van der Heijden, M.; Bardgett, R.; van Straalen, N.M. Van Der Heijden MGA, Bardgett RD, Van Straalen NM.. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology letters 2008, 11, 296-310. [CrossRef]
- Klironomos, J.; McCune, J.; Hart, M.; Neville, J. Klironomos JN, McCune J, Hart M, Neville J.. The influence of arbuscular mycorrhizae on the relationship between plant diversity and productivity. Ecology Letters 2000, 3, 137-141. [CrossRef]
- Rodriguez, R.J.; Redman, R.S.; Henson, J.M. The Role of Fungal Symbioses in the Adaptation of Plants to High Stress Environments. Mitigation and Adaptation Strategies for Global Change 2004, 9, 261-272. [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1-4. [CrossRef]
- Friesen, M.L.; Porter, S.S.; Stark, S.C.; Wettberg, E.J.v.; Sachs, J.L.; Martinez-Romero, E. Microbially Mediated Plant Functional Traits. Annual Review of Ecology, Evolution, and Systematics 2011, 42, 23-46. [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: the microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789-799. [CrossRef]
- McGuire, K.L.; Payne, S.G.; Orazi, G.; Palmer, M.I. In Green Roof Ecosystems. Springer International Publishing AG, 2015; Chapter 7, p 175-191.
- Fulthorpe, R.; MacIvor, J.S.; Jia, P.; Yasui, S.-L.E. The Green Roof Microbiome: Improving Plant Survival for Ecosystem Service Delivery. Frontiers in Ecology and Evolution 2018, 6. [CrossRef]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621-631. [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429-448. [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and Fungal Contributions to Carbon Sequestration in Agroecosystems. Soil Sci. Am. J. 2006, 70, 555-569. [CrossRef]
- Trivedi, P.; Anderson, I.C.; Singh, B.K. Microbial modulators of soil carbon storage: integrating genomic and metabolic knowledge for global prediction. Trends Microbiol. 2013, 21, 641-651. [CrossRef]
- Schimel, J.P.; Schaeffer, S.M. Microbial control over carbon cycling in soil. Frontiers in Microbiology 2012, 3, 348. [CrossRef]
- Molineux, C.; Connop, S.; Gange, A. Manipulating soil microbial communities in extensive green roof substrates. Sci. Total Environ.2014, 493, 632-638. [CrossRef]
- Molineux, C.; Gange, A.; Newport, D. Using soil microbial inoculations to enhance substrate performance on extensive green roofs. Sci. Total Environ.2016, 580. [CrossRef]
- Rumble, H.; Gange, A. Microbial inoculants as a soil remediation tool for extensive green roofs. Ecological Engineering 2017, 102, 188-198. [CrossRef]
- Rumble, H.; Finch, P.; Gange, A.C. Can microbial inoculants boost soil food webs and vegetation development on newly constructed extensive green roofs? Urban Forestry & Urban Greening 2022, 75, 127684. [CrossRef]
- Xie, L.; Lehvävirta, S.; Timonen, S.; Kasurinen, J.; Niemikapee, J.; Valkonen, J.P.T. Species-specific synergistic effects of two plant growth—promoting microbes on green roof plant biomass and photosynthetic efficiency. PLoS One 2019, 13, e0209432. [CrossRef]
- Xie, L.; Lehvävirta, S.; Valkonen, J.P.T. Case study: Planting methods and beneficial substrate microbes effect on the growth of vegetated roof plants in Finland. Urban Forestry & Urban Greening 2020, 53, 126722. [CrossRef]
- Young, T.; Cameron, D.D.; Phoenix, G.K. Using AMF inoculum to improve the nutritional status of Prunella vulgaris plants in green roof substrate during establishment. Urban Forestry & Urban Greening 2015, 14, 959-967. [CrossRef]
- McGuire, K.L.; Payne, S.G.; Palmer, M.I.; Gillikin, C.M.; Keefe, D.; Kim, S.J.; Gedallovich, S.M.; Discenza, J.; Rangamannar, R.; Koshner, J.A.; et al. Digging the New York City Skyline: Soil Fungal Communities in Green Roofs and City Parks. PLoS One 2013, 8, e58020. [CrossRef]
- Gill, A.S.; Purnell, K.; Palmer, M.I.; Stein, J.; McGuire, K.L. Microbial Composition and Functional Diversity Differ Across Urban Green Infrastructure Types. Frontiers in Microbiology 2020, 11, 912. [CrossRef]
- Droz, A.G.; Coffman, R.R.; Eagar, A.C.; Blackwood, C.B. Drivers of fungal diversity and community biogeography differ between green roofs and adjacent ground-level green space. Environ. Microbiol. 2022, 24, 5809-5824. [CrossRef]
- Hénault, A.; Heim, A.; Brisson, J.; Dagenais, D.; De Bellis, T.; Chagnon, P.L. Stressful, isolated, yet diverse: Green roofs have rich microbiomes that are not dominated by oligotrophic taxa. Environ. Microbiol. 2022, 14, 766-774. [CrossRef]
- Mitchell, M.E.; Hamilton, T.L.; Uebel-Niemeier, C.; Hopfensperger, K.N.; Buffam, I. Nitrogen cycling players and processes in green roof ecosystems. Applied Soil Ecology 2018, 132, 114-125. [CrossRef]
- Hoch, J.M.; Rhodes, M.E.; Shek, K.L.; Dinwiddie, D.; Hiebert, T.C.; Gill, A.S.; Salazar Estrada, A.E.; Griffin, K.L.; Palmer, M.I.; McGuire, K.L. Soil microbial assemblages are linked to plant community composition and contribute to ecosystem services on urban green roofs. Frontiers in Ecology and Evolution 2019, 7, 198. [CrossRef]
- KMI (Koninklijk Metereologisch Instituut van België). Analyse van het jaar 2019. Available online: https://www.meteobelgie.be/klimatologie/waarnemingen-en-analyses/jaar-2019/2254-analyse-van-het-jaar-2019. (accessed on 29 December 2023).
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; et al. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2016, 1. [CrossRef]
- Vancov, T.; Keen, B. Amplification of soil fungal community DNA using the ITS86F and ITS4 primers. FEMS Microbiol. Lett. 2009, 296, 91-96. [CrossRef]
- R Core Team. R: A language and environment for statistical computing. Available online: https://www.R-project.org/ (accessed on 20 March 2024).
- Martin, M. CUTADAPT removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17. [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581-583. [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.-H.; Taylor, Andy F.S.; May, Tom W.; Frøslev, T.G.; Pawlowska, J.; Lindahl, B.; Põldmaa, K.; Truong, C.; et al. The UNITE database for molecular identification and taxonomic communication of fungi and other eukaryotes: sequences, taxa and classifications reconsidered. Nucleic Acids Res. 2023, 52, D791-D797. [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590-596. [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 2013, 8, e61217. [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36-42. [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O'Hara, B.; Simpson, G.; Solymos, P.; Stevens, H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.2-1 2015, 2, 1-2.
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463-1464. [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228-8235. [CrossRef]
- Wickham, H. (2009). ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag New York. [CrossRef]
- Cao, Y.; Dong, Q.; Wang, D.; Zhang, P.; Liu, Y.; Niu, C. microbiomeMarker: an R/Bioconductor package for microbiome marker identification and visualization. Bioinformatics 2022, 38, 4027-4029. [CrossRef]
- Ji, P.; Sæbø, A.; Stovin, V.; Hanslin, H.M. Sedum root foraging in layered green roof substrates. Plant and Soil 2018, 430, 1-14. [CrossRef]
- Smalla, K.; Wieland, G.; Buchner, A.; Zock, A.; Parzy, J.; Kaiser, S.; Roskot, N.; Heuer, H.; Berg, G. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: plant-dependent enrichment and seasonal shifts revealed. Appl. Environ. Microbiol. 2001, 67, 4742-4751. [CrossRef]
- Tan, F.; Wang, J.; Chen, Z.; Feng, Y.; Chi, G.; Rehman, S.U. Assessment of the arbuscular mycorrhizal fungal community in roots and rhizosphere soils of Bt corn and their non-Bt isolines. Soil Biol. Biochem. 2011, 43, 2473-2479.
- Fierer, N.; Leff, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. U. S. A. 2012, 109, 21390-21395. [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 15684-15689. [CrossRef]
- Delgado-Baquerizo, M.; Oliverio, A.; Brewer, T.; Benavent-González, A.; Eldridge, D.; Bardgett, R.; Maestre, F.; Singh, B.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320-325. [CrossRef]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343-351. [CrossRef]
- Waite, D.W.; Chuvochina, M.; Pelikan, C.; Parks, D.H.; Yilmaz, P.; Wagner, M.; Loy, A.; Naganuma, T.; Nakai, R.; Whitman, W.B.; et al. Proposal to reclassify the proteobacterial classes Deltaproteobacteria and Oligoflexia, and the phylum Thermodesulfobacteria into four phyla reflecting major functional capabilities. Int. J. Syst. Evol. Microbiol. 2020, 70, 5972-6016. [CrossRef]
- Wehner, J.; Powell, J.R.; Muller, L.A.H.; Caruso, T.; Veresoglou, S.D.; Hempel, S.; Rillig, M.C. Determinants of root-associated fungal communities within Asteraceae in a semi-arid grassland. J. Ecol. 2014, 102, 425-436. [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.; Clemmensen, K.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.; Baldrian, P.; et al. FungalTraits: a user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Diversity 2020, 105, 1-16. [CrossRef]
- Boyce, K.; Andrianopoulos, A. Fungal dimorphism: The switch from hyphae to yeast is a specialized morphogenetic adaptation allowing colonization of a host. FEMS Microbio. Rev. 2015, 39. [CrossRef]
- Rai, M.; Agarkar, G. Plant-fungal interactions: What triggers the fungi to switch among lifestyles? Crit. Rev. Microbiol. 2016, 42, 428-438. [CrossRef]
- Marttinen, E.M.; Niemi-Kapee, J.; Laaka-Lindberg, S.; Valkonen, J.P.T. Fungal pathogens infecting moss green roofs in Finland. Urban Forestry & Urban Greening 2020, 55, 126812. [CrossRef]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P. The genus Cladosporium. Stud. Mycol. 2012, 72, 1-401. [CrossRef]
- Fröhlich-Nowoisky, J.; Pickersgill, D.; Despres, V.; Pöschl, U. High diversity of fungi in air particulate matter. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 12814-12819. [CrossRef]
- Núñez, A.; García, A.M.; Moreno, D.A.; Guantes, R. Seasonal changes dominate long-term variability of the urban air microbiome across space and time. Environment International 2021, 150, 106423. [CrossRef]
- Tordoni, E.; Ametrano, C.G.; Banchi, E.; Ongaro, S.; Pallavicini, A.; Bacaro, G.; Muggia, L. Integrated eDNA metabarcoding and morphological analyses assess spatio-temporal patterns of airborne fungal spores. Ecol. Indic. 2021, 121. [CrossRef]
- Marčiulynas, A.; Lynikienė, J.; Marčiulynienė, D.; Gedminas, A.; Menkis, A. Seasonal and Site-Specific Patterns of Airborne Fungal Diversity Revealed Using Passive Spore Traps and High-Throughput DNA Sequencing. Diversity 2023, 15, 539. [CrossRef]
- Gönczöl, J.; Révay, Á. Fungal spores in rainwater: Stemflow, throughfall and gutter conidial assemblages. Fungal Diversity 2004, 16.
- Bowers, R.; Clements, N.; Emerson, J.; Wiedinmyer, C.; Hannigan, M.; Fierer, N. Seasonal Variability in Bacterial and Fungal Diversity of the Near-Surface Atmosphere. Environ. Sci. Technol. 2013, 47. [CrossRef]
- Gandolfi, I.; Bertolini, V.; Ambrosini, R.; Bestetti, G.; Franzetti, A. Unravelling the bacterial diversity in the atmosphere. Appl. Microbiol. and Biotechnol. 2013, 97, 4727-4736. [CrossRef]
- Zhong, S.; Zhang, L.; Jiang, X.; Gao, P. Comparison of chemical composition and airborne bacterial community structure in PM2.5 during haze and non-haze days in the winter in Guilin, China. Sci. Total Environ. 2019, 655, 202-210. [CrossRef]
Figure 1.
Samples taken during the study. The inner blue squares measure 30 x 30 cm.

Figure 2.
Vegetational characteristics (a-c) and substrate depth (d) of Sedum-moss roofs (blue) vs Sedum-herbs-grasses roofs (orange). Boxplots span the interquartile range (i.e., the range between the 25th to 75th percentile), lines within boxes mark the median. Asterisks denote statistically significant differences between both roof types (p-value < 0.05; Wilcoxon rank-sum tests).
Figure 2.
Vegetational characteristics (a-c) and substrate depth (d) of Sedum-moss roofs (blue) vs Sedum-herbs-grasses roofs (orange). Boxplots span the interquartile range (i.e., the range between the 25th to 75th percentile), lines within boxes mark the median. Asterisks denote statistically significant differences between both roof types (p-value < 0.05; Wilcoxon rank-sum tests).
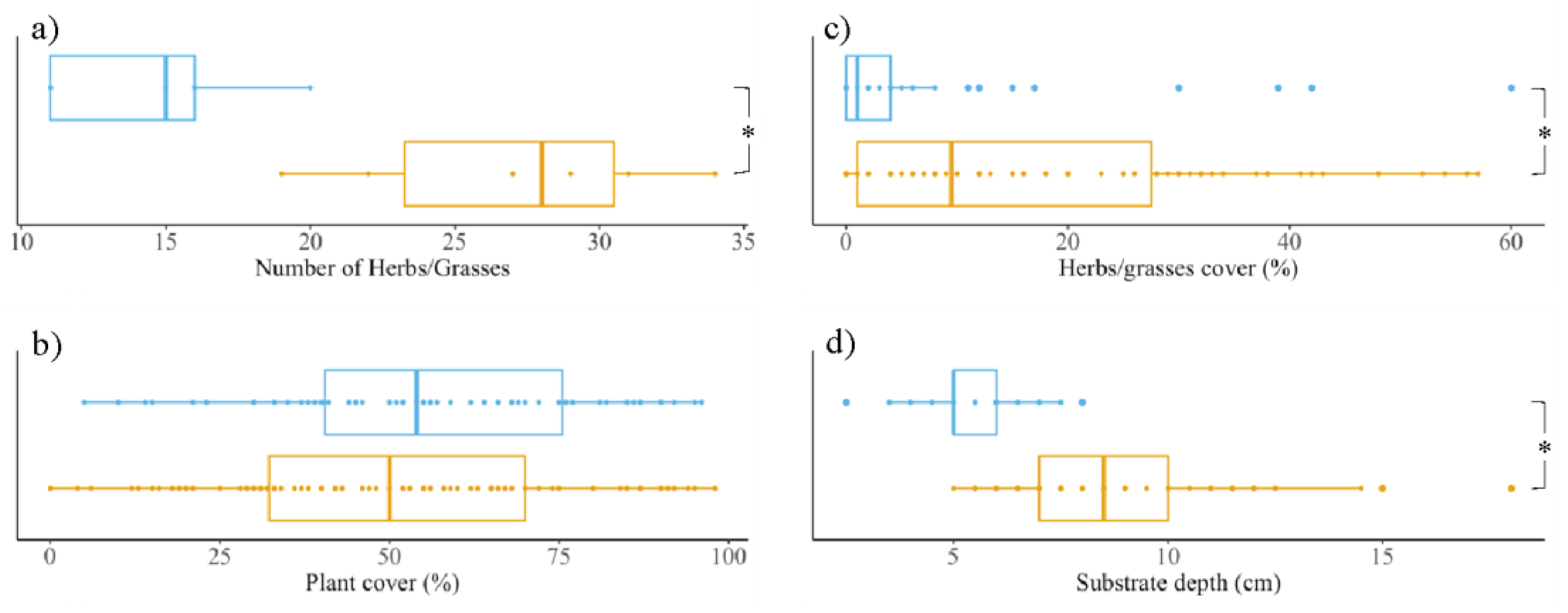

Table 1.
Green roof coordinates and descriptive characteristics.
| ID | City | Coordinates | Year | Height (m) | Area (m2) | Type |
|---|---|---|---|---|---|---|
| R1 | Ghent | 51.0239N 3.7665E | 2014 | 3.2 | 25 | Sedum-herbs-grasses |
| R2 | Ghent | 51.0479N 3.7419E | 2015 | 3.4 | 76 | Sedum-herbs-grasses |
| R3 | Ghent | 51.0457N 3.7509E | 2005 | 10.5 | 110 | Sedum-moss |
| R4 | Ghent | 51.0766N 3.7211E | 2013 | 8.4 | 588 | Sedum-moss |
| R5 | Hasselt | 50.9285N 5.3430E | 2015 | 7.0 | 225 | Sedum-herbs-grasses |
| R6 | Hasselt | 50.9338N 5.3419E | 2012 | 6.0 | 81 | Sedum-moss |
| R7 | Hasselt | 50.9263N 5.3410E | 2004 | 3.0 | 175 | Sedum-herbs-grasses |
| R8 | Antwerp | 51.1927N 4.4163E | 2014 | 22.3 | 708 | Sedum-moss |
| R9 | Antwerp | 51.1693N 4.3941E | 2008 | 9?0 | 280 | Sedum-moss |
| R10 | Antwerp | 51.2507N 4.4190E | 2009 | 17.4 | 777 | Sedum-herbs-grasses |
| R11 | Antwerp | 51.2302N 4.4165E | 2015 | 9.0 | 312 | Sedum-herbs-grasses |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



