Submitted:
28 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
Due to bacterial contamination, natural medicinal preservatives may be an alternative to enhance the quality of milk. Besides, thorough biocompatibility testing is deemed essential before integrating natural extracts into food products. To address this matter, our research focused on evaluating the antimicrobial potential of Plectranthus amboinicus, Flueggea leucopyrus, Cymbopogon citratus, and Bacopa monnieri against prevalent foodborne pathogens, including Staphylococcus aureus, Escherichia coli, Salmonella spp, and Pseudomonas aeruginosa and investigating the efficacy of plant extracts as natural preservatives along with biocompatibility and toxicity assessments. The methanolic extracts of plant materials were used for measuring antimicrobial activity using the agar well diffusion method, Minimum Inhibitory Concentration (MIC) and Minimum bactericidal concentration (MBC) assays. The preservative activity was tested by isolating pathogenic organisms in a sterilized milk medium and toxicity and biocompatibility was assessed using zebrafish embryo and brine shrimp lethality assays. At a concentration of 400 mg/ml, all plant extracts demonstrated notable activity against bacterial strains. B. monnieri at 400 mg/ml exhibited particularly significant effects on S. aureus (30.3 mm ± 0.6), surpassing the positive control, gentamicin (28.3 mm ± 0.6), while also displaying pronounced activity against E. coli. F. leucopyrus, at the same concentration, showed antimicrobial activity comparable to the positive control against both S. aureus (28.0 mm ± 1.0) and E. coli (27.0 mm ± 1.0). However, as there’s weak-mild toxicity in these herbs, prior to consumption, careful consideration of their toxicity and biocompatibility is imperative for ensuring their safe use.
Keywords:
Antimicrobial activity
; preservative activity
; biocompatibility
; Plectranthus amboinicus
; Bacopa monnieri
; Flueggea leucopyrus
; Cymbopogon citratus
1. Introduction
Milk is a major source of essential nutrients such as protein, calcium, and vitamin D for human beings. It has consistently played a crucial role in the dietary habits of people worldwide, forming an integral part of their consumption patterns [1]. The unique composition and properties make milk an excellent medium for bacterial growth, making it prone to microbial contamination. Microbial contamination of milk on dairy farms can be caused by factors such as herd hygiene, mastitis, production environment, chemicals, equipment control, storage facilities, and chemical hazards [2]. Spoilage of milk is a significant issue caused by the growth of microorganisms, leading to undesirable flavors, odors, and changes in texture and appearance. This deterioration can result in the loss of nutritional value, a decline in quality, and a reduced shelf life for milk [3]. The main microbiological hazards associated with raw milk consumption are Staphylococcus aureus, Salmonella spp., Listeria monocytogenes, Escherichia coli O157:H7, and Campylobacter jejuni. These pathogenic bacteria, transmitted through milk, pose a significant threat to human health, constituting approximately 90% of all diseases related to dairy consumption [4].
The dairy industry aims to sustain productivity and competitiveness in the expanding commercial market of milk by offering a diverse range of products and addressing consumer needs [5]. So, they are attentive to using chemical preservatives widely to prevent the spoilage of milk and extend its shelf life. Chemical preservatives play a major role in the preservation of milk, aiming to extend its shelf life and inhibit spoilage. Commonly employed preservatives include sodium benzoate, which combats yeast and mold growth, and sodium propionate, effective against mold and certain bacteria. Nisin, a natural antimicrobial peptide, serves as an additional defense against various bacteria. Moreover, Sorbic acid is utilized to inhibit the growth of mold and yeast, while natamycin acts as an antifungal agent to prevent mold proliferation. Hydrogen peroxide, functioning as a disinfectant, is employed to eliminate bacteria [6]. Nonetheless, their use has been shown to have negative consequences, including human health hazards, chemical residues in food and food chains, and the development of microbial resistance to the chemicals used [7]. Some common concerns include the possibility of allergic reactions, particularly in individuals sensitive to specific preservatives, leading to symptoms such as skin rashes, itching, or respiratory issues. Additionally, certain preservatives may have metabolic effects or interact with other substances in the body, potentially impacting health. Sensitivity to asthma can be a concern with certain preservatives, like sulfites, triggering symptoms in susceptible individuals. There is also apprehension about the development of resistant bacterial strains due to the use of antimicrobial agents, including preservatives, and the potential carcinogenicity of specific chemical preservatives is under investigation in various studies [8]. While regulatory agencies establish acceptable daily intake levels to ensure safe use, individuals are advised to be mindful of their dietary choices and, when possible, opt for foods with minimal or no preservatives or choose natural alternatives. Despite their efficacy, there is growing interest in exploring natural alternatives, such as plant extracts, due to concerns regarding potential health effects associated with some chemical preservatives. Thus, consumers have significant concerns regarding the risks posed by synthetic additives to human health; therefore, the use of these chemicals in food preservation is decreasing [9]. Therefore, there is a need to replace chemical preservatives with new eco-friendly, potentially effective, safe, and natural alternative preservatives. This shift aims to reduce the growth of pathogenic bacteria and extend the shelf life of milk and milk products [10].
Recently, many researchers have explored the potential utilization of plant extracts as natural preservatives. They have identified numerous herbal plants containing various constituents and biologically active molecules with antimicrobial properties. These studies have revealed that the mechanism of antimicrobial action in medicinal plant extracts can be harnessed as natural antimicrobial agents to extend the shelf life and maintain the quality of food, including milk [11]. These plant extracts present a wide variety of antimicrobial components that may possess crucial characteristics for inhibiting a broad spectrum of pathogens [12]. Many plants exhibit antimicrobial properties attributed to the essential oil constituents present in their extracts [13]. Some major antimicrobial components in plant extracts include aldehydes, ketones, polyphenols, ethers, alcohols, and hydrocarbons. Additionally, plants contain a diverse array of bioactive compounds such as flavonoids, terpenoids (including essential oils), glycosides, saponins, tannins, lignans, alkaloids, carotenoids, and lectins, each contributing to the plant’s defense mechanisms and offering various biological activities [7]. These components are abundant in the flowers, leaves, seeds, and bulbs of herbaceous plants [14].
Kapparawalliya (Plectranthus amboinicus), commonly known as Cuban oregano or Mexican mint, is a perennial herb recognized for its aromatic leaves used in culinary applications and traditional medicine. Lunuwila (Bacopa monnieri) is an herbaceous plant with succulent leaves, also known as Brahmi, traditionally acclaimed for its cognitive-enhancing properties in Ayurvedic medicine. Katupila (Flueggea leucopyrus), a deciduous shrub or small tree, is notable for its use in traditional medicine and as a source of bioactive compounds. Sera (Cymbopogon citratus), or lemongrass, is a tropical plant celebrated for its culinary uses and therapeutic properties, particularly its aromatic leaves infused for teas and essential oils. Each of these plants holds cultural significance and is being studied for potential applications in antimicrobial and preservative activities.
Herbal plants are generally considered safe and are entirely free or many have minimal side effects and low toxicity. However, it’s essential to be aware of potential adverse effects, especially when consumed in excess or by individuals with specific health conditions [15]. Recent studies, including one by [16], have evaluated the potential toxicity from both acute and chronic exposure, even with extracts considered to have low toxicity. Therefore, it is deemed necessary to assess the biocompatibility of herbs and their extracts before consumption, ensuring safety and minimizing the risk of potential side effects. Additionally, individual variations in response to herbal remedies underscore the importance of considering factors such as age, underlying health conditions, and concurrent medication use. Collaborative efforts between traditional knowledge and modern scientific research play a crucial role in establishing comprehensive guidelines for the safe and effective use of herbal remedies. Regular monitoring and updates based on emerging research contribute to the ongoing evaluation of herbal safety profiles, enhancing consumer awareness and promoting informed choices in herbal medicine utilization [17].
The present study aims to assess the antimicrobial and preservative activities of herbal plant extracts, including Kapparawalliya (Plectranthus amboinicus (Lour.) Spreng), Lunuwila (Bacopa monnieri (L.) Wettst), Katupila (Flueggea leucopyrus Willd.), and Sera (Cymbopogon citratus (DC.) Stapf). The evaluation specifically targets common milk pathogens such as Staphylococcus aureus, Salmonella spp., Escherichia coli, and Pseudomonas aeruginosa. Additionally, the study aims to investigate the biocompatibility and toxicity of these plant extracts, providing a comprehensive understanding of their potential applications and ensuring safety in usage.
2. Results and Discussion
2.1. Antimicrobial Susceptibility Testing
In the current study, various extracts of Plectranthus amboinicus, Bacopa monnieri, Flueggea leucopyrus, and Cymbopogon citratus were assessed for their antimicrobial activity against selected gram-negative and gram-positive bacteria, recognized as human pathogenic microorganisms. The susceptibility of each plant extract was examined using both agar-well diffusion and macro broth dilution assays.
2.1.1. Agar Well Diffusion Assay
The agar well diffusion assay technique, commonly employed to evaluate the antibacterial activity of plant or microbial extracts, was utilized in this investigation, following established methodologies [18]. The diameter of the zone of inhibition (ZOI) was measured to the nearest millimeter, and the mean ZOI was subsequently calculated. The results, including the mean and standard deviation (SD), of triplicate well diffusion assays for all plant extracts are presented in (Table 01).

Table 01.
Triplicate results of antimicrobial susceptibility test of selected herbal plant extracts. The results are expressed as the mean ZOI and standard deviation (SD) of triplicated well diffusion assays for all plant extracts.
Table 01.
Triplicate results of antimicrobial susceptibility test of selected herbal plant extracts. The results are expressed as the mean ZOI and standard deviation (SD) of triplicated well diffusion assays for all plant extracts.
| Selected herbal plant extracts with different concentrations | Inhibition zones diameter against tested organisms(mm) | |||
| S. aureus | P. aueroginosa | Salmonella spp. | E-coli | |
| 1.Katupila (Flueggea leucopyrus) | ||||
| 400 mg/dl | 0.6 | 0.6 | 2.0 | |
| 200 mg/dl | 1.0 | 0.6 | - | 0.6 |
| 100 mg/dl | 1.0 | - | 2.0 | |
| 50 mg/dl | 1.0 | 0.6 | - | |
| 25 mg/dl | 0.6 | - | - | 1.5 |
| 12.5 mg/dl | 0.0 | - | - | 0.6 |
| Positive control | 0.6 | 0.6 | 2.1 | 0.6 |
| Negative control | - | - | - | - |
| 2.Kapparawalliya (Plectranthus amboinicus) | ||||
| 400 mg/dl | 0.6 | 0.6 | 1.2 | 4.2 |
| 200 mg/dl | 2.5 | 0.6 | - | - |
| 100 mg/dl | 4.4 | - | - | - |
| 50 mg/dl | - | - | - | - |
| 25 mg/dl | - | - | - | - |
| 12.5 mg/dl | - | - | - | - |
| Positive control | 0.6 | 0.6 | 0.6 | |
| Negative control | - | - | - | - |
| 3.Lunuwila (Bacopa monnieri) | ||||
| 400 mg/dl | 0.6 | 0.6 | 0.6 | |
| 200 mg/dl | 1.0 | 0.6 | 1.2 | 0.6 |
| 100 mg/dl | 0.6 | 0.6 | 1.0 | 0.0 |
| 50 mg/dl | 0.6 | - | - | |
| 25 mg/dl | 0.6 | - | - | - |
| 12.5 mg/dl | - | - | - | - |
| Positive control | 0.6 | 0.6 | 2.1 | 0.6 |
| Negative control | - | - | - | - |
| 4.Sera (Cymbopogan citratus) | ||||
| 400 mg/dl | 1.0 | 0.6 | 0.6 | 2.0 |
| 200 mg/dl | 0.6 | 0.6 | - | 0.6 |
| 100 mg/dl | 1.0 | 0.0 | - | 2.0 |
| 50 mg/dl | - | - | - | 1.0 |
| 25 mg/dl | - | - | - | 1.5 |
| 12.5 mg/dl | - | - | - | 0.6 |
| Positive control | 0.6 | 2.1 | 2.1 | 0.6 |
| Negative control | - | - | - | - |
(-): inactive, no zone detected.
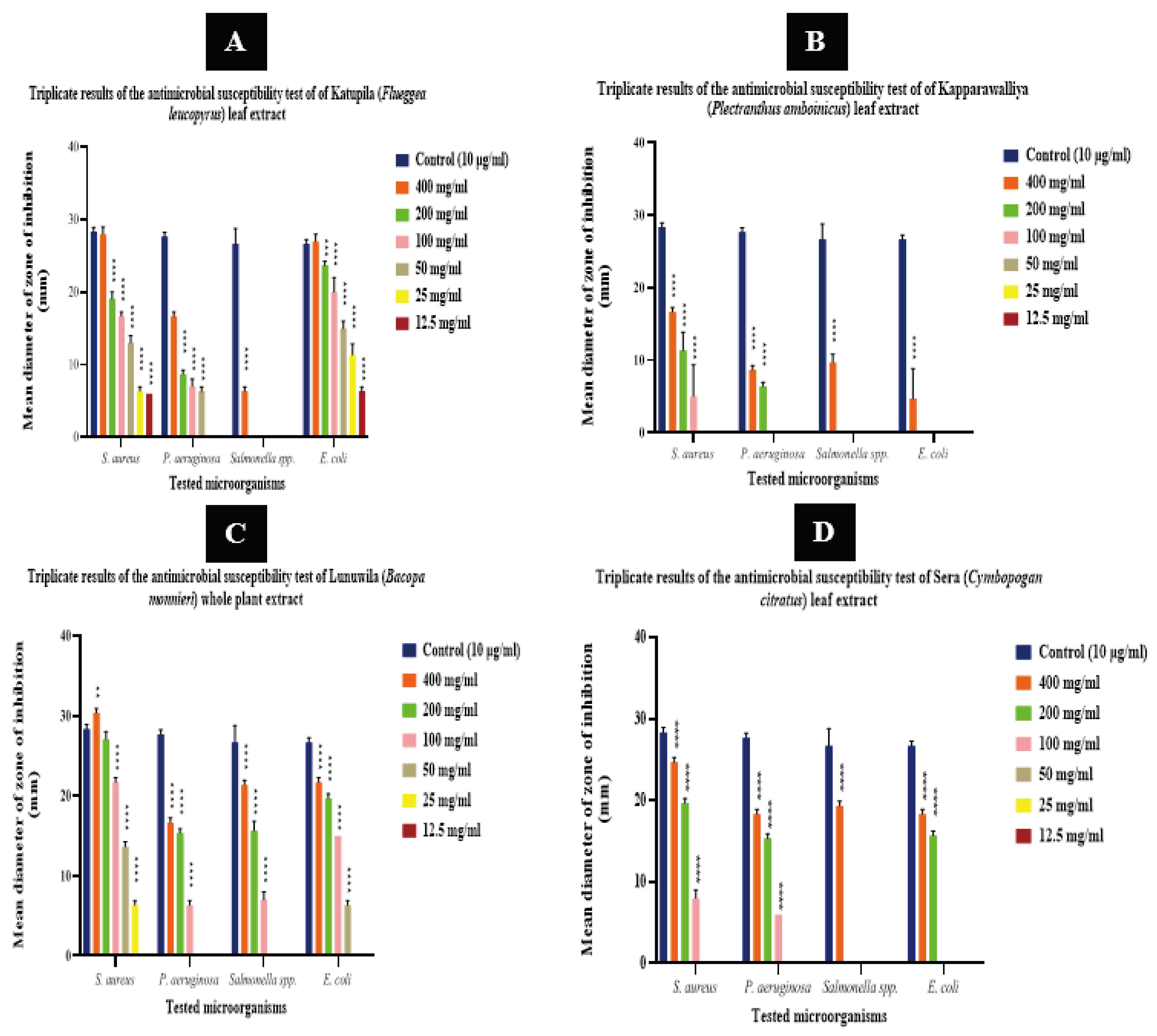
Notably, all tested herbal extracts at a concentration of 400 mg/ml demonstrated inhibitory effects against all bacterial strains examined. Particularly at 400 mg/ml, a prominent inhibitory activity was observed with B. monnieri, P. amboinicus, F. leucopyrus, and C. citratus against S. aureus, with zones of inhibitions (ZOI) values of 30.0 mm ± 0.6, 28.0 mm ± 1.0, 25.0 mm ± 1.0, and 16.7 mm ± 0.6, respectively. At that same concentration, F. leucopyrus, B. monnieri, and C. citratus exhibited a prominent antimicrobial activity against E. coli with zones of inhibitions of 27.0 mm ± 1.0, 21.7 mm ± 0.6, and 18.3 mm ± 0.6, respectively. All the plant extracts exhibited a lower activity against P. aeruginosa, while Salmonella spp. was the least susceptible among all the tested bacteria.
The whole plant extract of B. monnieri exhibited the highest antimicrobial activity against S. aureus and Salmonella spp., and the second-highest activity against E. coli and P. aeruginosa among all the tested methanolic extracts. At a concentration of 400 mg/ml, the whole plant extract of B. monnieri exhibited a significantly increased growth suppression potency against S. aureus (30.3 ± 0.6) compared to that of the standard reference antibiotic (28.3 ± 0.6) with a p-value ≤ 0.01. Even at a concentration of 200 mg/ml, B. monnieri demonstrated a zone of inhibition against S. aureus (27.0 mm ± 1.0) very close to the mean ZOI of gentamicin, and there was no significant difference from the controls. However, at concentrations of 100 mg/ml, 50 mg/ml, and 25 mg/ml, B. monnieri exhibited zones of 21.7 mm ± 0.6, 13.7 mm ± 0.6, and 6.3 mm ± 0.6, respectively, which were significantly lower. The mean diameters of the zone of inhibition for E. coli with B. monnieri at concentrations of 400 mg/ml, 200 mg/ml, 100 mg/ml, and 50 mg/ml were 21.7 mm ± 0.6, 19.7 mm ± 0.6, 15.0 mm ± 0.0, and 6.3 mm ± 0.6, respectively (P ≤ 0.0001). Even as B. monnieri displayed the highest activity against Salmonella spp., the mean diameters of the zone of inhibition at concentrations of 400 mg/ml, 200 mg/ml, and 100 mg/ml were 21.3 mm ± 0.6, 15.7 mm ± 1.2, and 7.0 mm ± 1.0, respectively, all significantly lower (P ≤ 0.0001). P. aeruginosa proved to be the least susceptible microorganism to B. monnieri extract. At a concentration of 400 mg/ml, the mean diameter of the zone of inhibition for P. aeruginosa with B. monnieri (16.7 mm ± 0.6) was identical to that of F. leucopyrus at 400 mg/ml. However, at 200 mg/ml, the zone of inhibition for P. aeruginosa (15.3 mm ± 0.6) surpassed that of F. leucopyrus (8.7 ± 0.6) at that concentration, while it decreased to 6.3 mm ± 0.6 at 100 mg/ml. No detectable inhibition zones were observed at the remaining concentrations. In a review by [19], the antimicrobial potential of B. monnieri against uropathogens and its bioactive compounds was explored. The review emphasized the antibacterial activity of B. monnieri against various bacteria, including Klebsiella pneumoniae and Proteus mirabilis. It also delved into the mechanisms of action, proposing that the antimicrobial activity of B. monnieri is linked to the presence of bioactive compounds such as alkaloids and saponins. According to [20], the diethyl ether extracts of B. monnieri exhibited antibacterial potency against S. aureus (gram-positive), while the ethyl acetate extract demonstrated effects on E. coli (gram-negative) at higher concentrations of 300 µg/mL. The ethanolic extract displayed potent antifungal activity against fungi, specifically Aspergillus flavus and Candida albicans, in comparison to diethyl ether and ethyl acetate-ether extracts. Both diethyl ether and ethyl acetate extracts demonstrated minimal antifungal effects, with more pronounced inhibitory effects observed against the tested bacteria. No inhibitory effects of the aqueous extract were observed at all concentrations (100 µg/mL, 200 µg/mL, and 300 µg/mL) against the examined bacterial and fungal species.
F. leucopyrus leaf extracts, at a concentration of 400 mg/ml, exhibited the highest antibacterial activity against E. coli, the second-highest antimicrobial activity against S. aureus, and the lowest activity against Salmonella spp. among all the tested methanolic plant extracts. The growth suppression potency of the F. leucopyrus leaf extract at this concentration against S. aureus (28.0 mm ± 1.0) and E. coli (27.0 mm ± 1.0) was closely comparable to that of the standard reference antibiotic, gentamicin (S. aureus - 28.3 mm ± 0.6 and E. coli - 26.7 mm ± 0.6), showing no significant deviation (P>0.05). At other tested concentration (200 mg/ml, 100 mg/ml, 50 mg/ml, 25 mg/ml, and 6.25 mg/ml), F. leucopyrus leaf extracts exhibited a zone of inhibition against both S. aureus and E. coli, with all ZOIs significantly lower than the controls. While S. aureus demonstrated the highest susceptibility to F. leucopyrus leaf extracts at 400 mg/ml, E. coli proved to be the most susceptible organism at all other tested concentrations. At 400 mg/ml, F. leucopyrus displayed a zone of inhibition of only 6.3 mm ± 0.6, while no discernible zone was observed for Salmonella spp. at the other concentrations of the leaf extracts. A study conducted by [21]investigated the antimicrobial activity of Flueggea leucopyrus leaf extracts. The findings of the study revealed that the extracts demonstrated inhibitory effects against various bacteria, including S. aureus, P. aeruginosa, and E. coli.
Among all the tested methanolic plant extracts, C. citratus leaf extracts exhibited the highest antimicrobial activity against P. aeruginosa. The mean diameters of the zone of inhibition for P. aeruginosa at concentrations of 400 mg/ml, 200 mg/ml, and 100 mg/ml were 18.3 mm ± 0.6, 15.3 mm ± 0.6, and 6.0 mm ± 0.0, respectively. The mean ZOIs at all concentrations against all tested bacteria were significantly lower than the controls. According to [22], the methanolic extract of C. citratus exhibited an intermediate level of antibacterial activity, with a smaller zone of inhibition compared to that obtained with the essential oil. This difference in activity could be attributed to the lower concentration of phytochemical constituents, such as flavonoids and tannins, in the methanolic extract. The essential oil, being highly concentrated, may contain a higher quantity of these phytochemicals, contributing to its greater antibacterial activity.
Among the tested plant extracts, the methanolic extracts of P. amboinicus exhibited the lowest antibacterial activity compared to the other plant extracts. But in a recent study investigating the antimicrobial activity of P. amboinicus solvent extracts against human pathogenic bacteria and fungi, it was found that the antimicrobial activity of P. amboinicus was more pronounced at a concentration of 100 mg/ml compared to 50 mg/ml. Specifically, the methanol extract of exhibited the highest antimicrobial activity when compared to the chloroform extract. The results indicated that P. amboinicus demonstrated significant antimicrobial activity against various bacteria and only one fungal yeast, Candida albicans [23].
The nature of cell wall structures makes gram-positive bacteria more susceptible to various chemical compounds compared to gram-negative bacteria [24]. This susceptibility difference arises because gram-negative bacteria possess an outer membrane with a lipopolysaccharide layer that acts as a barrier, making them impermeable to certain antimicrobial agents. In the current study, S. aureus demonstrated greater susceptibility to herbal plant extracts compared to other tested microorganisms. All the results obtained for the agar well diffusion assay is presented in (Figure 01).
Figure 01.
Results of the agar well diffusion assay for antimicrobial activity of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus against S. aureus, P. aeruginosa, Salmonella spp., and E. coli. The Y-axis represents mean ZOIs in mm, while the X-axis represents the microorganisms tested. The error bars represent the variability or uncertainty associated with the mean ZOI values. (A) Antimicrobial activity of F. leucopyrus leaf extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). (B) Antimicrobial activity of P. amboinicus leaf extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). (C) Antimicrobial activity of B. monnieri whole plant extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). (D) Antimicrobial activity of C. citratus leaf extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). Asterisks denote the concentration at which antimicrobial activity is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001).
Figure 01.
Results of the agar well diffusion assay for antimicrobial activity of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus against S. aureus, P. aeruginosa, Salmonella spp., and E. coli. The Y-axis represents mean ZOIs in mm, while the X-axis represents the microorganisms tested. The error bars represent the variability or uncertainty associated with the mean ZOI values. (A) Antimicrobial activity of F. leucopyrus leaf extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). (B) Antimicrobial activity of P. amboinicus leaf extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). (C) Antimicrobial activity of B. monnieri whole plant extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). (D) Antimicrobial activity of C. citratus leaf extract, presented as the mean diameter of the zone of inhibition (mm) with standard deviation (SD). Asterisks denote the concentration at which antimicrobial activity is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001).
2.1.2. Minimum Inhibitory Concentration (MIC)
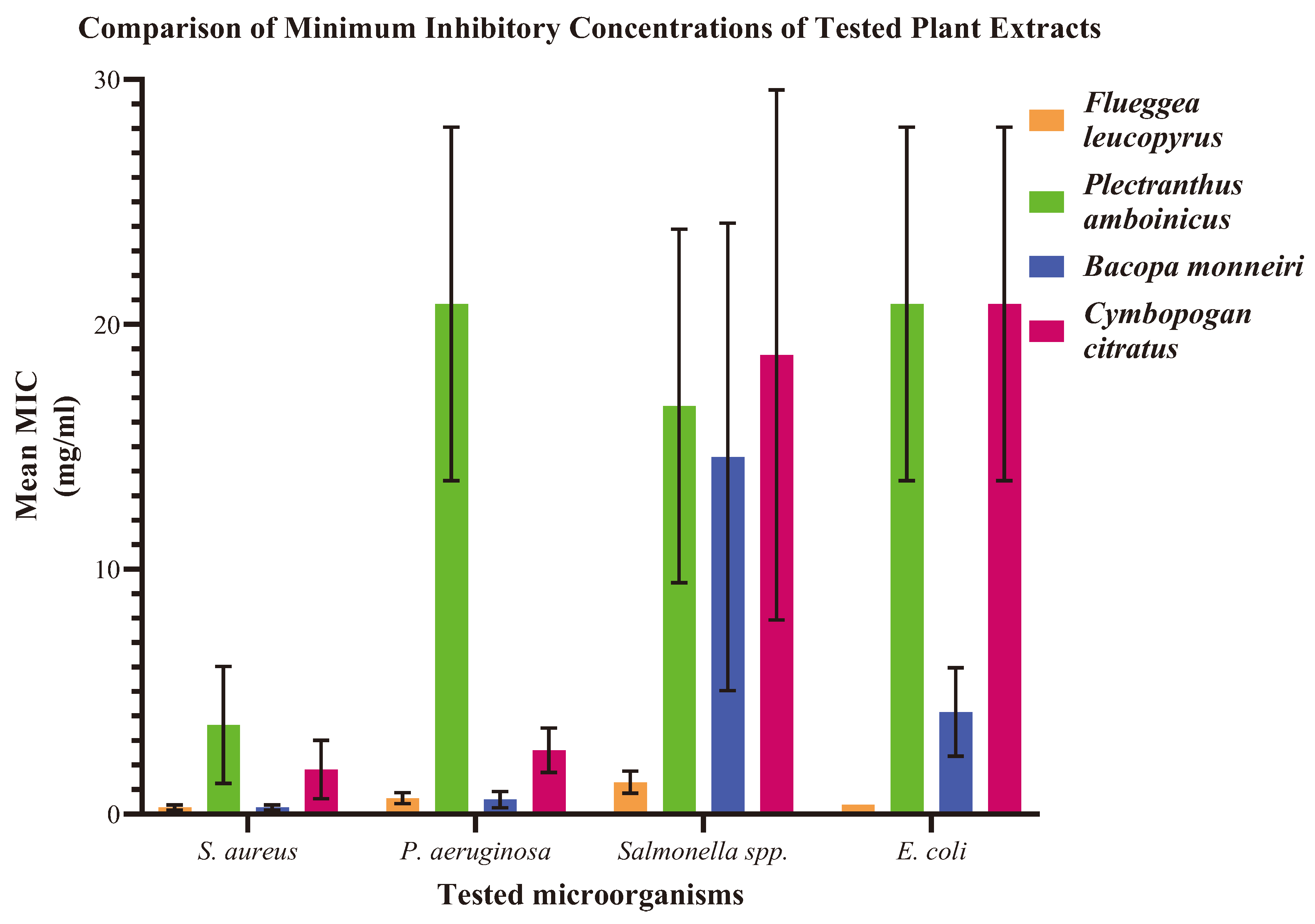
The MIC values for each plant extract against all tested microorganisms ranged from 0.3 mg/ml ± 0.1 to 20.8 mg/ml ± 7.2. F. leucopyrus and B. monnieri exhibited the lowest MIC against S. aureus, while P. amboinicus showed the highest MIC for S. aureus (Table 02).
Table 02.
MIC results of herbal plant extracts. The table presents the mean MIC and standard deviation (SD) values obtained from triplicate macro broth dilution assays for herbal plant extracts.
Table 02.
MIC results of herbal plant extracts. The table presents the mean MIC and standard deviation (SD) values obtained from triplicate macro broth dilution assays for herbal plant extracts.
| Observation of the growth results of the microorganisms at different concentrations | ||||
| S. aureus | P. aeruginosa | E. coli | Salmonella spp. | |
| Katupila (Flueggea leucopyrus) | ||||
| Mean MIC (mg/ml) | 0.3 | 0.7 | 1.3 | 0.4 |
| SD | 0.1 | 0.2 | 0.5 | 0.0 |
| Kapparawalliya (Plectranthus amboinicus) | ||||
| Mean MIC (mg/ml) | 3.7 | 20.8 | 16.7 | 20.8 |
| SD | 2.4 | 7.2 | 10.2 | 7.2 |
| Lunuwila (Bacopa monnieri) | ||||
| Mean MIC (mg/ml) | 0.3 | 0.6 | 14.6 | 4.2 |
| SD | 0.1 | 0.3 | 9.5 | 1.8 |
| Sera (Cymbopogon citratus) | ||||
| Mean MIC (mg/ml) | 1.8 | 2.6 | 18.75 | 20.8 |
| SD | 1.2 | 0.9 | 10.8 | 7.2 |
B. monnieri had the lowest MIC against P. aeruginosa, whereas P. amboinicus had the highest. F. leucopyrus demonstrated the lowest MIC against Salmonella spp., while C. citratus displayed the highest MIC against Salmonella spp. The lowest MIC against E. coli was recorded for F. leucopyrus, while C. citratus and P. amboinicus exhibited the highest MIC. Among the tested plant extracts, F. leucopyrus and B. monnieri demonstrated the highest potency of antimicrobial activity compared to others, whereas C. citratus exhibited a notable antimicrobial potency. P. amboinicus leaf extract displayed the least potential for antimicrobial activity compared to other plant extracts, as illustrated in (Figure 02).
Figure 02.
Comparison of Minimum Inhibitory Concentrations (MICs) of tested plant extracts against S. aureus, P. aeruginosa, Salmonella spp., and E. coli. The Y-axis represents mean MIC values in mg/ml, while the X-axis represents the microorganisms tested. The error bars represent the variability or uncertainty associated with the mean MIC values.
Figure 02.
Comparison of Minimum Inhibitory Concentrations (MICs) of tested plant extracts against S. aureus, P. aeruginosa, Salmonella spp., and E. coli. The Y-axis represents mean MIC values in mg/ml, while the X-axis represents the microorganisms tested. The error bars represent the variability or uncertainty associated with the mean MIC values.
2.1.3. Minimum Bactericidal Concentration (MBC)
Methanolic extracts of selected medicinal plants underwent further examination for Minimum Bactericidal Concentration (MBC) against the aforementioned microorganisms. The majority of MBC results were consistent with the observed zone of inhibition for antimicrobial activity. F. leucopyrus and B. monnieri plant extracts exhibited the highest antimicrobial activity, with MBC values ranging from 16.7 mg/ml to 400.0 mg/ml (Table 03).
Table 03.
MBC results of herbal plant extracts. The table presents the mean MBC and standard deviation (SD) values.
Table 03.
MBC results of herbal plant extracts. The table presents the mean MBC and standard deviation (SD) values.
| Observation of the growth results of the microorganisms at different concentrations | ||||
| S. aureus | P. aeruginosa | E. coli | Salmonella spp. | |
| Katupila (Flueggea leucopyrus) | ||||
| Mean MBC (mg/ml) | 16.7 | 66.7 | 400 | 20.8 |
| SD | 7.2 | 28.9 | 0.0 | 7.2 |
| Kapparawalliya (Plectranthus amboinicus) | ||||
| Mean MBC (mg/ml) | 200 | 266.7 | 400 | 400 |
| SD | 0.0 | 115.5 | 0.0 | 0.0 |
| Lunuwila (Bacopa monnieri) | ||||
| Mean MBC (mg/ml) | 41.7 | 116.7 | 83.3 | 83.3 |
| SD | 14.4 | 76.4 | 28.9 | 28.9 |
| Sera (Cymbopogon citratus) | ||||
| Mean MBC (mg/ml) | 83.3 | 133.3 | 400 | 266.7 |
| SD | 28.7 | 57.7 | 0.0 | 115.5 |
F. leucopyrus recorded the lowest MBC against S. aureus (16.7 mg/ml ± 7.2), while P. amboinicus exhibited the highest MBC against S. aureus (200 mg/ml ± 0.0). For P. aeruginosa, F. leucopyrus and B. monnieri demonstrated the lowest MBCs at 66.7 mg/ml ± 28.9 and 116.7 mg/ml ± 76.4, respectively, whereas P. amboinicus presented the highest (266.7 mg/ml ± 115.5). The lowest MBC against E. coli was found in F. leucopyrus (20.8 mg/ml ± 7.2), while P. amboinicus displayed the highest (400 mg/ml ± 0.0). B. monnieri also exhibited the lowest MBC against Salmonella spp. (83.3 mg/ml ± 28.9), while P. amboinicus and F. leucopyrus showed the highest (400 mg/ml ± 0.0). F. leucopyrus and B. monnieri demonstrated the highest potency of antimicrobial activity among the tested plant extracts, with C. citratus exhibiting notable potency. B. monnieri whole plant extract inhibited the growth of S. aureus, Salmonella spp., and E. coli at low concentrations, while F. leucopyrus leaf extract inhibited the growth of S. aureus, P. aeruginosa, and E. coli at low concentrations. In contrast, C. citratus inhibited the growth of S. aureus at a higher concentration. In addition, C. citratus leaf extract inhibited the growth of P. aeruginosa, Salmonella spp., and E. coli at concentrations of 133.3 mg/ml ± 57.7, 400 mg/ml ± 0.0, and 266.7 mg/ml ± 115.5, respectively. Meanwhile, P. amboinicus leaf extract exhibited the least potential for antimicrobial activity compared to other plant extracts.
Plant extracts exhibit a variety of phytochemical structures, and the antimicrobial properties of these extracts are contingent on the chemical composition of different plant parts. Hence, variations in antibacterial effects among plant species are attributed to phytochemical characteristics and species differences [25]. Plant-based antimicrobials hold significant therapeutic promise due to their ability to fulfill similar functions with fewer adverse effects compared to synthetic antimicrobials [26].
In the present study, achieving high antimicrobial activity close to the positive control required a much higher concentration compared to previous research. The observed concentration differences among F. leucopyrus, B. monnieri, C. citratus, and P. amboinicus can be attributed to variations in bioactive compound composition, extraction methods, testing conditions, microbial strains, and potential synergistic effects. Each plant may contain a distinct profile of antimicrobial compounds, influencing effectiveness at varying concentrations. Differences in extraction methods and testing conditions can impact bioactive compound yield and activity. Moreover, variations in microbial strain susceptibility and resistance mechanisms can necessitate higher concentrations for effective antimicrobial effects. Additionally, synergistic effects among compounds in the plant extracts may contribute to concentration requirements. Considering these factors is crucial when interpreting concentration differences in antimicrobial activity across studies [12]. Lastly, evaluating the antimicrobial and preservative activity of plant extracts is essential for assessing their potential as natural alternatives to synthetic antimicrobial agents, especially in industries such as food and pharmaceuticals.
2.2. Preservative Activity in Sterilized Milk Medium
Previous research has substantiated that natural antimicrobial compounds found in plants possess antimicrobial activity. The utilization of plant antimicrobial properties for milk and food preservation has garnered considerable attention due to the growing interest in natural and sustainable approaches [7]. Plant extracts containing bioactive compounds have been investigated for their potential as natural preservatives, and there is evidence suggesting their effectiveness. However, it’s crucial to evaluate their benefits in the context of the modern world and their potential to replace contemporary preservation techniques. Natural plant antimicrobials offer several advantages. Firstly, they are derived from renewable sources, rendering them environmentally friendly compared to synthetic chemicals. Additionally, plant antimicrobials are generally considered safe for consumption and do not leave behind harmful residues, addressing concerns related to chemical residues in food products [27]. Testing the preservative activity of plant extracts against milk pathogens using isolated milk pathogenic strains and a sterilized milk medium is a valuable approach for assessing the antimicrobial potential of these extracts.
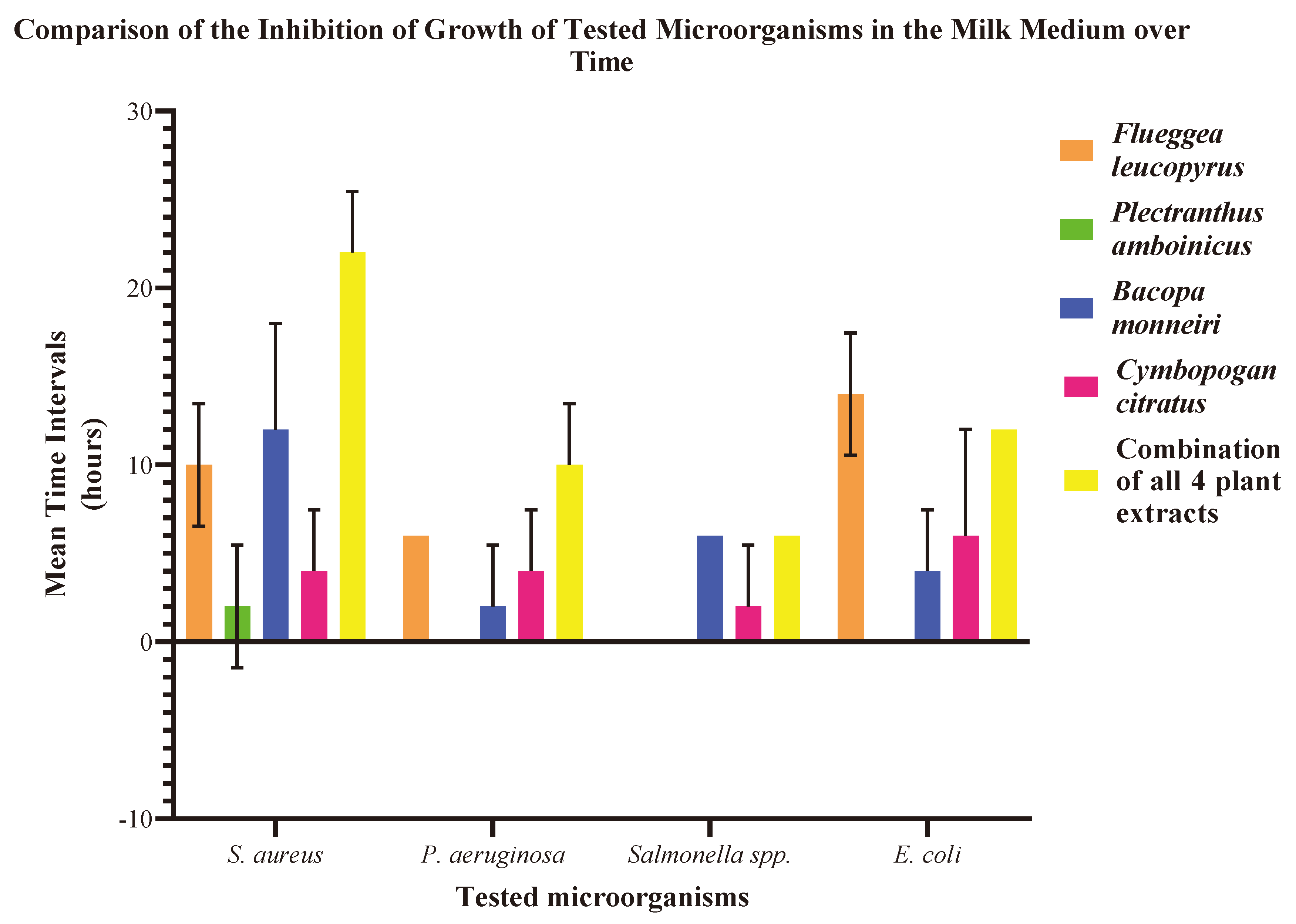
The inhibition time period for S. aureus was most pronounced with B. monnieri-treated milk medium, lasting 12.0 hours ± 6.0. F. leucopyrus demonstrated a mean inhibition duration of 10.0 hours ± 3.5 against S. aureus in milk, while C. citratus exhibited a comparatively shorter inhibition period of 4.0 hours ± 3.5. The growth of S. aureus was evident in the milk sample treated with P. amboinicus even after 6 hours of plant treatment, exhibiting a limited inhibitory effect with a mean time duration of 2.0 hours ± 3.5. The most prolonged period of growth inhibition for E. coli (14 hours ± 3.5) was noted in milk treated with F. leucopyrus, whereas B. monnieri and C. citratus demonstrated a more limited inhibitory effect on the growth of E. coli, with mean time periods of 4.0 hours ± 3.5 and 6.0 hours ± 6.0, respectively (Table 04).
Table 04.
Mean time durations with standard deviation up to which the growth of microorganisms is inhibited.
Table 04.
Mean time durations with standard deviation up to which the growth of microorganisms is inhibited.
| Plant Extract | Mean time durations with standard deviation up to which the growth of microorganisms is inhibited (hours) | |||
| S. aureus | P. aeruginosa | Salmonella spp. | E. coli | |
| Flueggea leucopyrus | 10.0 ± 3.5 | 6.0 ± 0.0 | 0.0 ± 0.0 | 14 ± 3.5 |
| Plectranthus amboinicus | 2.0 ± 3.5 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Bacopa monnieri | 12.0 ± 6.0 | 2.0 ± 3.5 | 6.0 ± 0.0 | 4.0 ± 3.5 |
| Cymbopogan citratus | 4.0 ± 3.5 | 4.0 ± 3.5 | 2 ± 3.5 | 6.0 ± 6.0 |
| Combination of all 4 plant extracts | 22 ± 3.5 | 10.0 ± 3.5 | 6.0 ± 0.0 | 12.0 ± 0.0 |
In milk medium treated with P. amboinicus, E. coli continued to grow even after 6 hours of plant treatment. Furthermore, P. aeruginosa growth remained unaffected by P. amboinicus-treated milk samples. F. leucopyrus, B. monnieri, and C. citratus inhibited P. aeruginosa growth for mean periods of 6.0 hours ± 0.0, 2.0 hours ± 3.5, and 4.0 hours ± 3.5, respectively. While F. leucopyrus and P. amboinicus did not inhibit the growth of Salmonella spp., B. monnieri and C. citratus successfully suppressed Salmonella spp. growth in milk medium for an average time period of 6.0 hours ± 0.0 and 2 hours ± 3.5, respectively. The most potent inhibition was witnessed when employing the combined extracts of all four plants. This synergistic blend effectively suppressed the growth of S. aureus, P. aeruginosa, Salmonella spp., and E. coli for 22 hours ± 3.5, 10.0 hours ± 3.5, 6.0 hours ± 0.0, and 12.0 hours ± 0.0, respectively (Figure 03).
Figure 03.
Mean time durations with standard deviation up to which the growth of microorganisms is inhibited. The Y-axis represents mean time duration in hours, while the X-axis represents the microorganisms tested. The error bars represent the variability or uncertainty associated with the mean MIC values.
Figure 03.
Mean time durations with standard deviation up to which the growth of microorganisms is inhibited. The Y-axis represents mean time duration in hours, while the X-axis represents the microorganisms tested. The error bars represent the variability or uncertainty associated with the mean MIC values.
According to [7], all tested herbal extracts demonstrated an inhibitory effect against pathogens. Gram-negative bacteria E. coli and P. aeruginosa exhibited lower susceptibility to the inhibitory activity of all herbal extracts used in the experiment compared to Gram-positive bacteria such as S. aureus, L. monocytogenes, and B. subtilis in both nutrient agar and sterilized milk mediums.
Therefore, these findings suggest the potential use of these herbal extracts in milk and milk products as natural antimicrobials.
2.3. Assessment of Biocompatibility and Toxicity
Previous studies have indicated that many herbal plants exhibit both antimicrobial and preservative properties. However, it is crucial to acknowledge that some of these plants may also possess toxicological properties. Assessing biocompatibility is essential to ensure the safety of plant extracts for use The zebrafish embryo assay and brine shrimp lethality assay are recognized as animal models suitable for evaluating the toxicological assessment of herbal plants and estimating the level of damage caused by compounds to both biological and non-biological materials [28]. While the results of these assays may not directly translate to human toxicity, they offer initial insights into the safety profile of the plant extracts.
2.3.1. Brine Shrimp Lethality Assay
- 1.
- Evaluation of the Toxicity of Methanolic Plant Extracts Dissolved in 10% DMSO
Plant crudes obtained through methanolic plant extracts, dissolved in 10% DMSO, were utilized for both antimicrobial susceptibility testing and assessing the preservative activity of these extracts against selected common milk pathogens. The brine shrimp lethality assay was employed to assess the toxicity of both 10% DMSO and the combined toxicity of methanolic plant extracts dissolved in 10% DMSO. The mortality rate of brine shrimp at different concentrations of plant extracts dissolved in 10% DMSO is given in (Table 05).
Table 05.
Mortality rate of Brine Shrimp after 24 hours of plant treatment dissolved in 10% DMSO.
| Concentration(mg/ml) | Mortality rate of Brine Shrimp after 24 hours of plant treatment (Mean% ± SD) | |||
| Flueggea leucopyrus | Plectranthus amboinicus | Bacopa monnieri | Cymbopogan citratus | |
| 400 | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 |
| 40 | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 |
| 4 | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 |
| 0.4 | 73.3 ± 5.8 | 33.3 ±5.8 | 63.3 ± 5.8 | 40.0 ± 0.0 |
| 0.04 | 46.7 ± 5.8 | 23.3 ± 5.8 | 50.0 ± 10 | 26.7 ± 5.8 |
| 10% DMSO | 23.3 ± 5.8 | 20.0 ± 0.0 | 26.7 ± 5.8 | 16.7 ± 5.8 |
| Sea water | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
No mortality among brine shrimp larvae was observed during the 24-hour experiment in seawater. Consequently, the survival rate was 100%, and the mortality rate was 0%. The mortality of brine shrimp in 10% DMSO ranged from 16.7% ± 5.8 to 26.7% ± 5.8, which was significantly higher (P ≤ 0.05) than that in seawater. Additionally, the mortality rate at different concentrations of 400, 40, 4, 0.4, and 0.04 mg/ml of each plant extract dissolved in 10% DMSO significantly increased (P ≤ 0.0001) compared to the mortality rate in seawater. Exposure to high concentrations of 400, 40, and 4 mg/ml of each plant extract dissolved in 10% DMSO resulted in 100% mortality in brine shrimp larvae. At a concentration of 0.4 mg/ml, mortality rates of 73.3% ± 5.8, 33.3% ± 5.8, 63.3% ± 5.8, and 40.0% ± 0.0 were observed for F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus, respectively. Furthermore, the mortality rate of the aforementioned plant extracts dissolved in 10% DMSO at 0.04 mg/ml was 23.3% ± 5.8, 20.0% ± 0.0, 26.7% ± 5.8, and 16.7% ± 5.8, respectively (Figure 04).
Figure 04.
(A) Mortality rate of Brine Shrimp after 24 hours of plant treatment dissolved in 10% DMSO. (B) Mortality rate (%) of brine shrimp larvae after 24 h exposure to increasing concentrations of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus methanolic extract dissolved in 0.1% DMSO (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). Asterisks denote the concentration at which brine shrimp mortality rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
Figure 04.
(A) Mortality rate of Brine Shrimp after 24 hours of plant treatment dissolved in 10% DMSO. (B) Mortality rate (%) of brine shrimp larvae after 24 h exposure to increasing concentrations of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus methanolic extract dissolved in 0.1% DMSO (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). Asterisks denote the concentration at which brine shrimp mortality rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
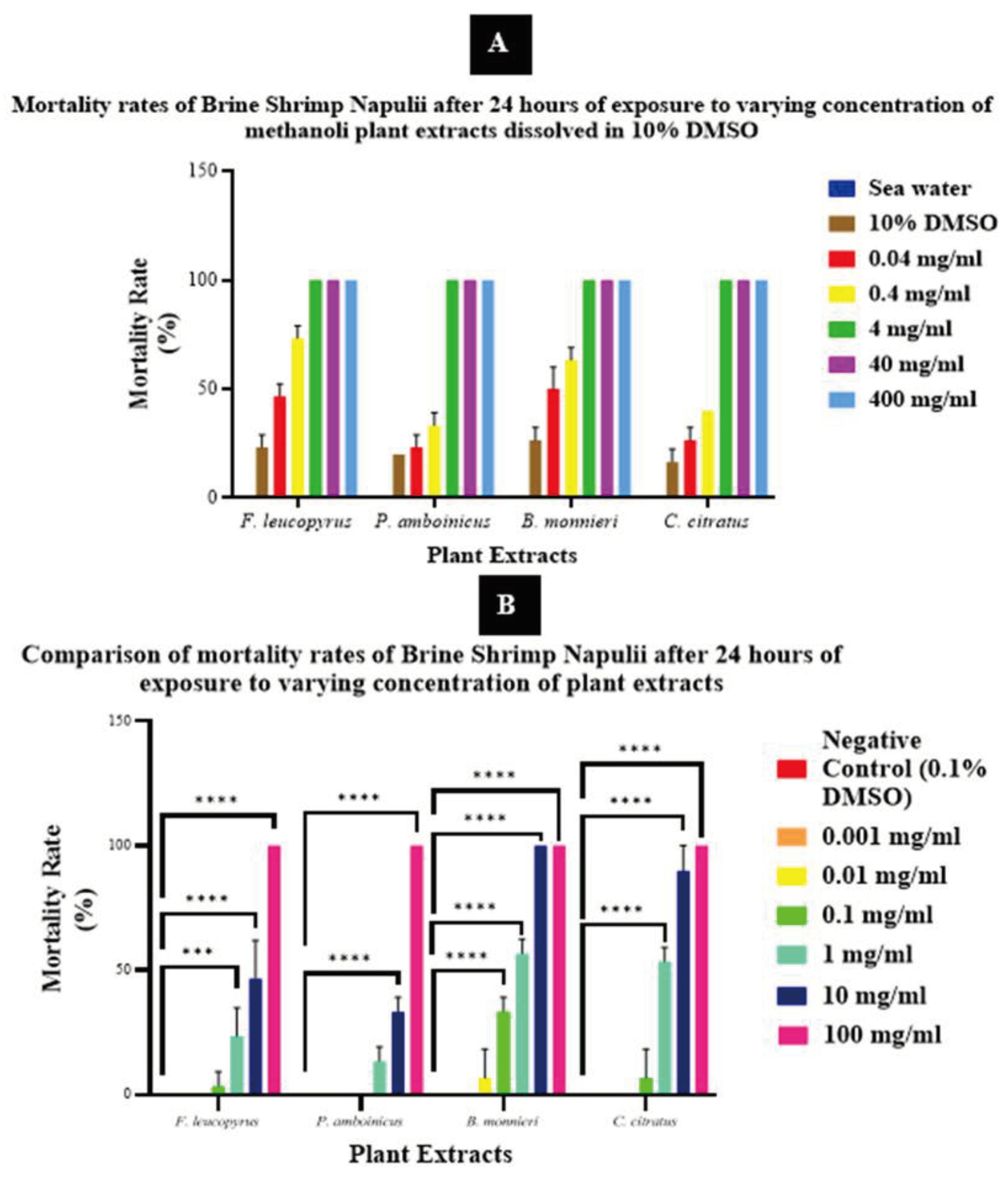
The heightened toxic effect observed may be attributed to the combined toxicity of both 10% DMSO and the plant extract in the brine shrimp lethality assay. In the brine shrimp lethality assay, the selection of a solvent for dissolving test compounds is critical, as the solvent can influence the observed toxicity [29]. Dimethyl sulfoxide (DMSO) is a frequently employed solvent due to its capacity to dissolve a diverse array of compounds. Nevertheless, it is essential to take into account the potential toxicity of DMSO itself, especially at higher concentrations [30]. In the case of 10% DMSO, it is generally considered a relatively high concentration for use in the brine shrimp lethality assay. Including solvent control groups is recommended due to potential solvent-related effects. DMSO toxicity varies with species, and brine shrimp are sensitive, showing adverse effects at higher DMSO concentrations. To accurately assess test compound toxicity, it’s advisable to use lower DMSO concentrations and conduct dose-response experiments [31]. In the present study, 10% DMSO resulted in a comparatively high mortality rate of 16.7% - 23.3% in brine shrimp nauplii.
- 2.
- Evaluation of the Toxicity of Methanolic Plant Extracts Dissolved in 0.1% DMSO
The brine shrimp lethality assay for methanolic plant extracts dissolved in 0.1% DMSO was conducted following the method proposed by [32] and [33] with minor modifications. The mortality rate of brine shrimp at different concentrations of plant extracts dissolved in 0.1% DMSO is given in (Table 06).
Table 06.
Mean mortality rates of brine shrimp nauplii after 24 hours of exposure to varying concentrations of methanolic plant extracts dissolved in 0.1% DMSO.
Table 06.
Mean mortality rates of brine shrimp nauplii after 24 hours of exposure to varying concentrations of methanolic plant extracts dissolved in 0.1% DMSO.
| Concentration(mg/ml) | Mortality rate of Brine Shrimp after 24 hours of plant treatment (Mean% ± SD) | |||
| Flueggea leucopyrus | Plectranthus amboinicus | Bacopa monnieri | Cymbopogan citratus | |
| 100 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 |
| 10 | 46.7 ± 15.3 | 33.3 ± 5.8 | 100.0 ± 0.0 | 90.0 ± 10.0 |
| 1 | 23.3 ± 11.5 | 13.3 ± 5.8 | 56.7 ± 5.8 | 53.3 ± 5.8 |
| 0.1 | 3.3 ± 5.8 | 0.0 ± 0.0 | 33.3 ± 5.8 | 6.7 ± 11.5 |
| 0.01 | 0.0 ± 0.0 | 0.0 ± 0.0 | 6.7 ± 11.5 | 0.0 ± 0.0 |
| 0.001 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Negative Control (0.1% DMSO) | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
According to [31], the recommended maximum working concentration (v/v) for DMSO, methanol, and ethanol is 1.25%, while for Tween 20, it is 0.16%. In the brine shrimp lethality assay, it has been established that a concentration of 0.1% DMSO is generally considered safe for brine shrimp. At this concentration, DMSO does not exhibit significant toxicity toward the brine shrimp, enabling an accurate assessment of the toxicity of test compounds. Brine shrimps are highly sensitive organisms, and elevated concentrations of DMSO can adversely affect their survival and development. Hence, it is advisable to use a lower concentration of DMSO, such as 0.1%, to minimize potential solvent-related toxicity. This ensures that the observed effects primarily stem from the test compounds under evaluation. For instance, a study by [29] assessed the toxicity of various solvents, including DMSO, in the brine shrimp lethality assay. Their findings indicated that concentrations up to 0.1% DMSO did not significantly impact the survival of brine shrimp larvae, suggesting that, at this concentration, DMSO itself did not exhibit substantial toxicity towards the brine shrimp.
According to the present study, no mortality of brine shrimp larvae was observed at the 24-hour time point with the 0.1% DMSO negative control. Thus, the survival rate was observed to be 100%. and the mortality rate was 0%. Further, 0.001 mg/ml dose of each plant extract dissolved in 0.1% DMSO exhibited a similar pattern to the negative control, with no significant deviation from the controls. Consequently, the mortality rate at this concentration remained at 0%. Similarly, exposure to 0.01 mg/ml doses of F. leucopyrus, P. amboinicus, and C. citratus leaf extracts resulted in a 0% mortality rate for brine shrimp larvae. At a dose of 0.01 mg/ml, a mortality rate of 6.7% ± 11.5 was observed for B. monnieri, which did not significantly differ from the control. Additionally, no mortality was observed at a concentration of 0.1 mg/ml of P. amboinicus leaf extract dissolved in 0.1% DMSO. At a concentration of 0.01 mg/ml, the mortality rates of F. leucopyrus and C. citratus were 3.3% ± 5.8 and 6.7% ± 11.5, respectively, without significant deviation from the controls. However, in the case of B. monnieri treatment at 0.1 mg/ml, the mortality rate (33.3% ± 5.8) was significantly affected (P ≤ 0.0001). Mortality rates of brine shrimp larvae exposed to F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus at a concentration of 1 mg/ml were 23.3% ± 11.5 (P ≤ 0.001), 13.3% ± 11.5 (P > 0.05), 5.5% ± 11.5 (P ≤ 0.001), and 53.3% ± 5.8 (P ≤ 0.001), respectively. The brine shrimp mortality in P. amboinicus was not significantly affected at that concentration, whereas the mortality in F. leucopyrus, B. monnieri, and C. citratus was significantly different at the same concentration. At a higher concentration of 10 mg/ml, the mortality rates of brine shrimp larvae exposed to F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus were 46.7% ± 15.3 (P ≤ 0.0001), 33.3% ± 5.8 (P ≤ 0.0001), 100.0% ± 0.0 (P ≤ 0.0001), and 90.0% ± 10.0 (P ≤ 0.0001), respectively. In high-dose treatment experiments involving all the mentioned plant extracts dissolved in 0.1% DMSO, mortality rates of brine shrimp larvae were significantly increased at 10 mg/ml and 100 mg/ml treatments, with a mortality rate of 100% observed at 100 mg/ml for each plant extract.
The LC50 (Lethal concentration) for each plant extract, dissolved in 0.1% DMSO, was calculated using GraphPad Prism 9.5.1. The resulting LC50 values for each plant extract are provided in (Table 07).
Table 07.
LC50 values for each plant extract dissolved in 0.1% DMSO.
| Plant Extract | Mean LC50 (mg/ml) ± SD |
| Flueggea leucopyrus | 12.7 ± 9.1 |
| Plectranthus amboinicus | 26.9 ±15.6 |
| Bacopa monnieri | 0.8 ± 0.1 |
| Cymbopogan citratus | 0.9 ± 0.2 |
The toxicity of herbal plants, expressed as LC50 values, is commonly assessed by comparison to either Meyer’s or Clarkson’s toxicity index (if LC50 < 1000 µg/ml is considered toxic, if LC50 > 1000 µg/ml considered as non-toxic) [34,35]. According to their toxicity index, [36] proposed the toxicity ranges of plant extracts based on LC50 results of the tested plant species. Therefore, toxicity assessment was conducted following the criteria outlined by [36], where LC50 values ranging between 0-0.1 mg/mL indicated high toxicity, LC50 between 0.1-0.5 mg/mL indicated moderate toxicity, LC50 between 0.5-1 mg/mL indicated weak-mild toxicity, and LC50 over 1 mg/mL indicated a non-toxic nature of the extract. The LC50 values of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus methanolic plant extracts dissolved in 0.1% DMSO ranged from 0.8 mg/ml ± 0.1 to 26.9 mg/ml ± 15.6. The LC50 values of F. leucopyrus (12.7 mg/ml ± 9.1) and P. amboinicus (26.9 ± 15.6) indicated the non-toxic nature of the extracts. In contrast, the LC50 of B. monnieri (0.8 mg/ml ± 0.1) and C. citratus (0.9 mg/ml ± 0.2) suggested a weakly-mild toxic nature. Consequently, the methanolic extract of B. monnieri dissolved in 0.1% DMSO exhibited the highest toxicity, while P. amboinicus demonstrated the least toxicity. According to [37], the brine shrimp lethality assay for the ethyl acetate extract of F. leucopyrus revealed LC50 values of 2779.63 μg/mL, suggesting no toxicity of the plant extract towards brine shrimp. This implies that LC50 values can vary with differences in plant extraction methods. The use of different extraction methods can result in the extraction of distinct sets and concentrations of bioactive compounds, influencing the toxicity of the plant extract [38]. Consequently, these variations can result in changes in the LC50 values, indicating different toxicological profiles of the extracted compounds. In a recent study conducted by [39], it was found that the methanolic plant extracts of P. amboinicus showed no toxicity in both zebrafish embryo and brine shrimp assays. Using the brine shrimp lethality bioassay, [40] tested the toxic effect of the aqueous extract of B. monnieri leaves and found it to exhibit low toxicity. In another study, [41] demonstrated through the A. salina lethality assay that C. citratus fractions were moderately toxic, with LC50 values ranging from 146.12 to 433.15 μg/ml.
2.3.2. Zebrafish FET293 Bioassay
The zebrafish embryo toxicity assay has gained significant attention for assessing the toxicity of natural products and herbal medicine [42]. It is particularly suitable for isolated compounds from herbal plants due to its ability to perform tests in small quantities. The transparent nature of zebrafish embryos allows for direct observation of developmental stages and endpoint toxicity assessment [43]. Additionally, this assay enables the evaluation of compound effects on various organs, such as the heart, brain, intestine, pancreas, cartilage, liver, and kidney, without requiring complicated processing [44].
Freeze-dried aqueous plant material was utilized in the zebrafish embryo assay (a modified version of OECD-FET assay) to mitigate the solvent effect of methanol, which could potentially impact the zebrafish embryo assay. Hatch rate and survival rate were observed at 24, 48, 54, 72, 80, and 96 hpf. Heart rate was recorded at 72 hpf, and developmental abnormalities were assessed at 96 hpf.
- Hatch Rate
Hatching or leaving the chorion, a crucial milestone in the early developmental cycle of zebrafish, is triggered biochemically when the embryo’s oxygen energy demands surpass the diffusion capabilities of O2 through the egg envelopes and perivitelline fluid [45]. This process involves the proteolytic hatching enzyme ZHE1 (zebrafish hatching enzyme), pivotal for “chorion softening” [46]. According to [47], exposure to heavy metals during this period can disrupt these enzymes, potentially altering or inhibiting the typical zebrafish hatching mechanism. In this study, we hypothesized that exposure to plant extracts may modulate the activities of hatching enzymes, potentially leading to a delay in the hatching process. However, further investigation is needed to unravel the exact mechanism of plant extracts on zebrafish hatching.
Throughout the treatments, control embryos were observed to hatch normally at each time point, eventually hatching all the examined embryos by the 72-hour time point. Furthermore, the 0.001 mg/ml dose of F. leucopyrus, P. amboinicus, and C. citratus leaf extracts also exhibited a consistent hatching pattern, indicating normalcy without significant deviation from the controls. However, only embryos exposed to the 0.001 mg/ml dose of B. monnieri completed hatching at 80 hours.
Embryos exposed to 0.01 and 0.1 mg/ml doses of P. amboinicus and C. citratus leaf extracts were observed hatching in an increasing pattern. Embryos exposed to 0.01 mg/ml concentrations of P. amboinicus and C. citratus completed hatching at the 72-hour time point without any significant alteration at any observed time point. In contrast, embryos exposed to the 0.1 mg/ml dose of both plant extracts completed hatching at the 80-hour time point. Although embryos exposed to the 0.1 mg/ml dose of C. citratus did not show any deviation from the controls, those exposed to the same concentration of P. amboinicus exhibited significant deviation at 54 and 72 hpf (P ≤ 0.0001).
At a dose of 0.01 mg/ml F. leucopyrus, embryos completed hatching at 80 hours, deviating significantly at the 54 (P ≤ 0.01) and 72-hour time points. However, those exposed to 0.1 mg/ml failed to hatch completely, leaving some embryos unhatched. This group exhibited significant deviations at 48, 54, and 72-hour (P ≤ 0.0001) time points and the hatching was not significantly affected compared to the control at 84 and 96-hour time points. Embryos exposed to 0.01 mg/ml and 0.1 mg/ml doses of B. monnieri whole plant extract were observed hatching in an increasing pattern, showing significant alterations from the control at the 72-hour time point. Eventually, all embryos hatched at the 80-hour time point.
According to [48], there was no significant difference in hatching rates between the control group and low concentrations (0.1% and 0.5%) for B. monnieri leaf extract. However, hatching rates decreased in the higher concentration groups (1%, 5%, and 10%) for the leaf extract. In contrast, embryonic hatching rates were significantly reduced in the silver nanoparticles (AgNPs) exposure groups, with a hatching rate of 53% in the 0.5% group, and no hatching observed in further concentrations. These findings suggest a notable dose-dependent decrease in hatching rates in both the leaf extract and biosynthesized AgNPs treated groups compared to the control.
As the hatch rates were examined in wild-type embryos at higher doses of 1, and 10 mg/ml, F. leucopyrus, B. monnieri, and P. amboinicus extracts demonstrated a significant effect on hatching, while the hatching pattern was significantly altered in general. The majority of embryos did not hatch at the higher doses (1 mg/ml, and 10 mg/ml) at any given time point. Although the hatching pattern of wildtype zebrafish embryos exposed to a 1 mg/ml concentration of C. citratus was significantly affected at 72 hpf (P ≤ 0.0001), it did not show a significant impact compared to the control at 48 hpf, 54 hpf, 84 hpf, and 96 hpf. When hatch rates were examined in wild-type embryos at higher doses of 10 mg/ml, C. citratus leaf extract exhibited a significant effect on hatching, with the overall hatching pattern significantly altered (Figure 05).
Figure 05.
Hatching rate (%) of wildtype zebrafish embryos at various time points (24, 48, 54, 72, 80, and 96 hpf) following exposure to different concentrations of aqueous extracts from F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean hatching rate (%), while the X-axis represents the zebrafish embryo age or observation time points. (A) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of F. leucopyrus leaf extract at various time points. (B) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of P. amboinicus leaf extract at various time points. (C) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of B. monnieri whole plant extract at various time points. (D) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of C. citratus leaf extract at various time points. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
Figure 05.
Hatching rate (%) of wildtype zebrafish embryos at various time points (24, 48, 54, 72, 80, and 96 hpf) following exposure to different concentrations of aqueous extracts from F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean hatching rate (%), while the X-axis represents the zebrafish embryo age or observation time points. (A) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of F. leucopyrus leaf extract at various time points. (B) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of P. amboinicus leaf extract at various time points. (C) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of B. monnieri whole plant extract at various time points. (D) Hatching rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of C. citratus leaf extract at various time points. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
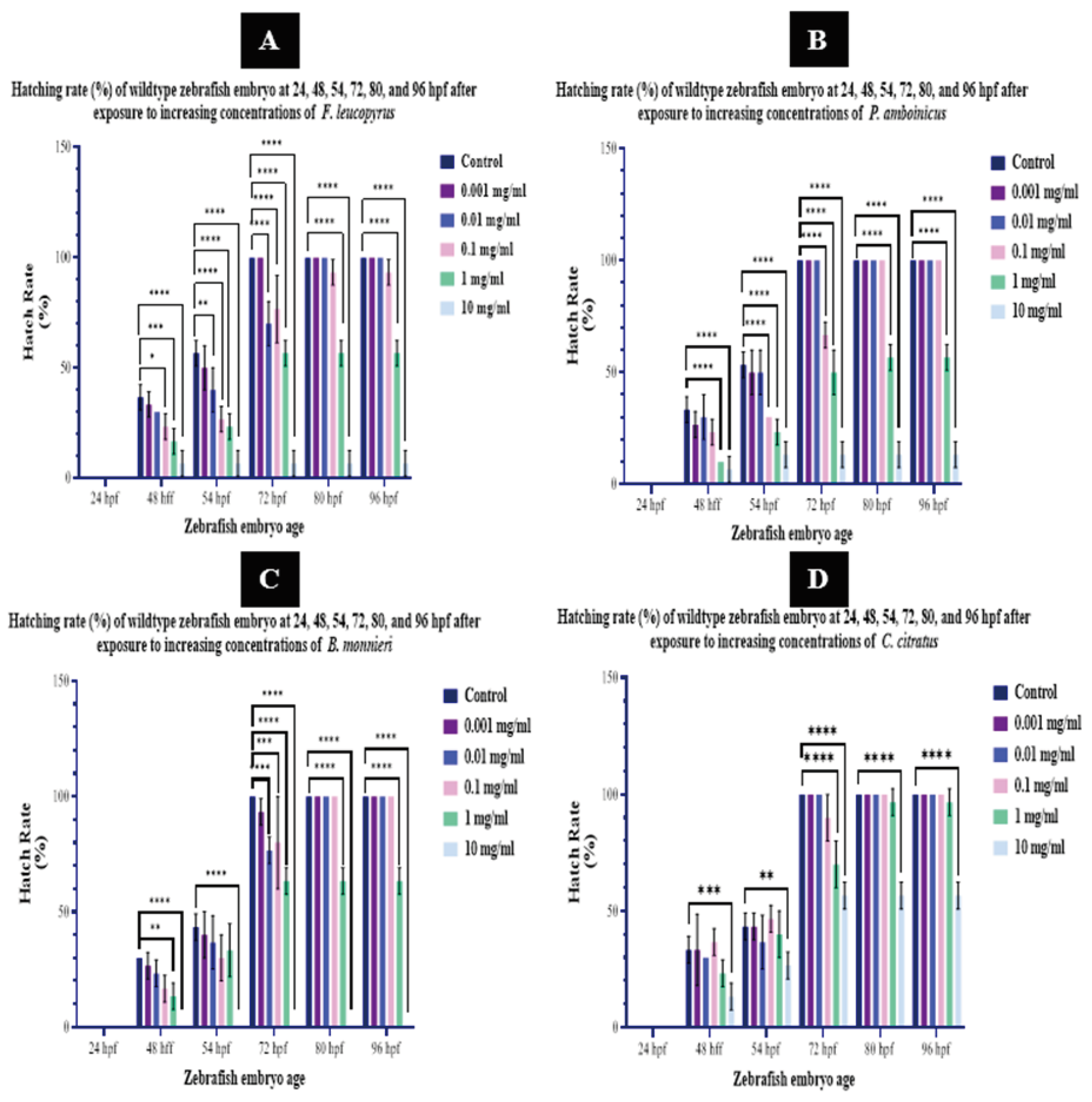
Assuming the hatching mechanism remains unaffected by exogenic factors, zebrafish embryos are anticipated to hatch within the typical timeframe of 48-96 hpf, with the majority hatching by 72 hpf [49]. Accordingly, in our study, the hatch rates of zebrafish embryos treated with F. leucopyrus, P. amboinicus, and C. citratus were normally distributed across time points from 24 hpf to 96 hpf. At the lowest dose (0.001 mg/ml), embryos in each treatment, except for B. monnieri, fully hatched at 72 hpf. B. monnieri-exposed embryos completed hatching at 80 hpf. Additionally, embryos exposed to 0.01 and 0.1 mg/ml concentrations of P. amboinicus, B. monnieri, and C. citratus completed hatching between 72-80 hpf. F. leucopyrus at 0.01 mg/ml resulted in hatching at 80 hpf, but at 0.1 mg/ml, some embryos remained unhatched. At higher doses (1 and 10 mg/ml), all plant extracts, except C. citratus at 1 mg/ml, significantly affected the hatching pattern, even starting from 48 hpf. Overall, our findings indicate that at the lowest doses, there is no substantial disruption to the typical hatching pattern of zebrafish embryos with any of the examined plant extracts. However, significant alterations in hatching patterns were observed at higher doses.
- 2.
- Survival Rate
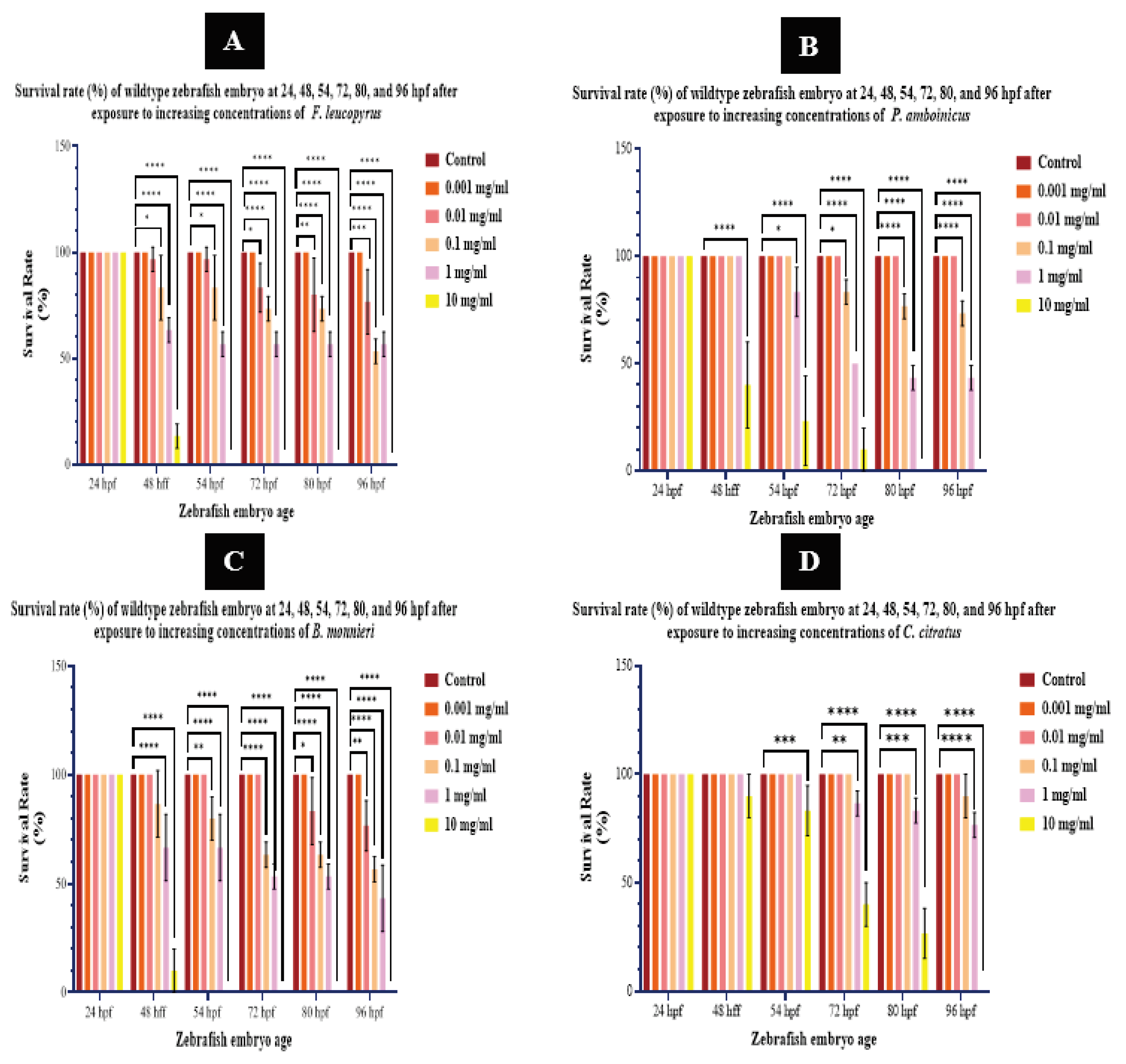
Mortality during early development can be attributed to the ability of toxicants to disrupt the functionality of multiple organs, ultimately leading to death from acute toxicity [50].
None of the zebrafish larvae in the group of wildtypes were subjected to death upon the treatment of any dose of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus aqueous extracts at 24-hour time point. Consequently, the survival rates were consistently 100%. Throughout the examined time points at 24, 48, 54, 72, 80, and 96 hpf after exposure to 0.001 mg/ml doses of all four extracts, zebrafish embryos were observed to have survived similarly to non-treated controls, with no recorded deaths at any given time point. Furthermore, at the 0.01 mg/ml dose of P. amboinicus and C. citratus leaf extracts, zebrafish embryos were observed to survive similarly to non-treated controls, which recorded no deaths at any given time point. But the survival of embryos was significantly affected by the 0.01 mg/ml dose of F. leucopyrus and B. monnieri from 72 hpf onwards. In high-dose treatment experiments with P. amboinicus leaf extract, the survival of wild-type zebrafish embryos was significantly reduced at 0.1, 1, and 10 mg/ml treatments at the 72, 80, and 96 hpf time points. The survival of embryos exposed to 0.1 mg/ml of C. citratus remained at 100% up to the 80-hour point and declined insignificantly at 96 hpf. In high-dose treatment experiments with C. citratus, the survival of wild-type zebrafish embryos was significantly reduced at 1 mg/ml and 10 mg/ml treatments at the 72, 80, and 96 hpf time points. According to [51], embryos exposed to solutions of essential oils of C. citratus at concentrations ranging from 0.04 to 0.46 mg/mL survived up to day 3At 0.1, 1, and 10 mg/ml doses of F. leucopyrus and 1 and 10 mg/ml doses of B. monnieri, the survival of embryos was significantly affected at 48, 54, 72, 80, and 96 hpf time points. Additionally, the survival of embryos exposed to the 0.1 mg/ml dose of B. monnieri was significantly affected at 54, 72, 80, and 96 hpf time points. No survival was observed for embryos exposed to a dose of 10 mg/ml at 96 hpf onward with C. citratus and 80 hpf onward with P. amboinicus, while no survival was observed from 54 hpf onward with 10 mg/ml doses of B. monnieri and F. leucopyrus (Figure 06).
Figure 06.
Survival rate (%) of wildtype zebrafish embryos at various time points (24, 48, 54, 72, 80, and 96 hpf) following exposure to different concentrations of aqueous extracts from F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean survival rate (%), while the X-axis represents the zebrafish embryo age or observation time points. (A) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of F. leucopyrus leaf extract at various time points. (B) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of P. amboinicus leaf extract at various time points. (C) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of B. monnieri whole plant extract at various time points. (D) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of C. citratus leaf extract at various time points. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
Figure 06.
Survival rate (%) of wildtype zebrafish embryos at various time points (24, 48, 54, 72, 80, and 96 hpf) following exposure to different concentrations of aqueous extracts from F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean survival rate (%), while the X-axis represents the zebrafish embryo age or observation time points. (A) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of F. leucopyrus leaf extract at various time points. (B) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of P. amboinicus leaf extract at various time points. (C) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of B. monnieri whole plant extract at various time points. (D) Survival rate (%) of wildtype zebrafish embryos exposed to increasing concentrations of C. citratus leaf extract at various time points. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
- 3.
- Heart Rate
The heart and vascular system are among the first organs to undergo development in vertebrate embryogenesis [52]. In fish, the formation of the cardiovascular system is a complex process involving intricate biochemical and molecular mechanisms [53]. This vital biological process encompasses a diverse array of genes, proteins, and signaling pathways [54], rendering it sensitive to the effects of chemicals, toxins, and environmental pollutants. Exposure to these substances can lead to the occurrence of cardiovascular abnormalities during early development [55]. Consequently, in this study, we measured the heart rates (beats/min.) of zebrafish embryos treated with plant extracts as a parameter to assess cardiovascular toxicity resulting from specific exposures to those plant extracts. The normal heartbeat of larvae is around 155 times/min [56]
The heart rates of embryos exposed to both low and high doses of P. amboinicus and C. citratus leaf extracts were not observed to be significantly altered from the controls. The mean heart rate of embryos within the control group and those exposed to 0.001 mg/ml, 0.01 mg/ml, 0.1 mg/ml, and 1 mg/ml doses of P. amboinicus was 143.5 ± 2.1 beats/min, 142.7 ± 1.5 beats/min, 144.3 ± 3.1 beats/min, 146.7 ± 3.0 beats/min, and 145.6 ± 3.9 beats/min, respectively. These values were all within the normal range of zebrafish heart rates. Similarly, the heart rates of embryos within the control group and those exposed to various concentrations of C. citratus were also within the normal range. The mean heart rate of embryos within the control group and those exposed to 0.001 mg/ml, 0.01 mg/ml, 0.1 mg/ml, and 1 mg/ml doses of C. citratus was 142.4 ± 0.9 beats/min, 146.4 ± 2.7 beats/min, 148.0 ± 4.0 beats/min, 150.5 ± 1.6 beats/min, and 150.5 ± 4.4 beats/min, respectively.
It was observed that the heart rate of embryos in the wild-type groups exposed to low doses of F. leucopyrus and B. monnieri extracts, including 0.001 and 0.01 mg/ml, was not significantly different from the control. The mean heart rate of embryos within the control, 0.001 mg/ml, and 0.01 mg/ml doses of F. leucopyrus were 144.5 ± 3.3 beats/min, 142.0 ± 1.8 beats/min, and 139.8 ± 2.5 beats/min, respectively—all within the normal range of zebrafish heart rates. Similarly, for B. monnieri, the mean heart rate within the control, 0.001 mg/ml, and 0.01 mg/ml doses was 144.7 ± 0.6 beats/min, 145.0 ± 2.0 beats/min, and 150.3 ± 0.6 beats/min, respectively, also falling within the normal range of zebrafish heart rates (Figure 07).
Figure 07.
Heart rate (beats/min) of wildtype zebrafish embryos at 72 hpf after exposure to increasing concentrations of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus aqueous extracts (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean heart rate (beats/min), and the X-axis represents the concentrations of plant extracts to which zebrafish embryos were exposed. (A) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of F. leucopyrus leaf extract. (B) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of P. amboinicus leaf extract. (C) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of B. monnieri whole extract. (D) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of C. citratus leaf extract. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
Figure 07.
Heart rate (beats/min) of wildtype zebrafish embryos at 72 hpf after exposure to increasing concentrations of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus aqueous extracts (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean heart rate (beats/min), and the X-axis represents the concentrations of plant extracts to which zebrafish embryos were exposed. (A) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of F. leucopyrus leaf extract. (B) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of P. amboinicus leaf extract. (C) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of B. monnieri whole extract. (D) Heart rate (beats/min) of 72 hpf wildtype zebrafish embryos after exposure to increasing concentration of C. citratus leaf extract. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
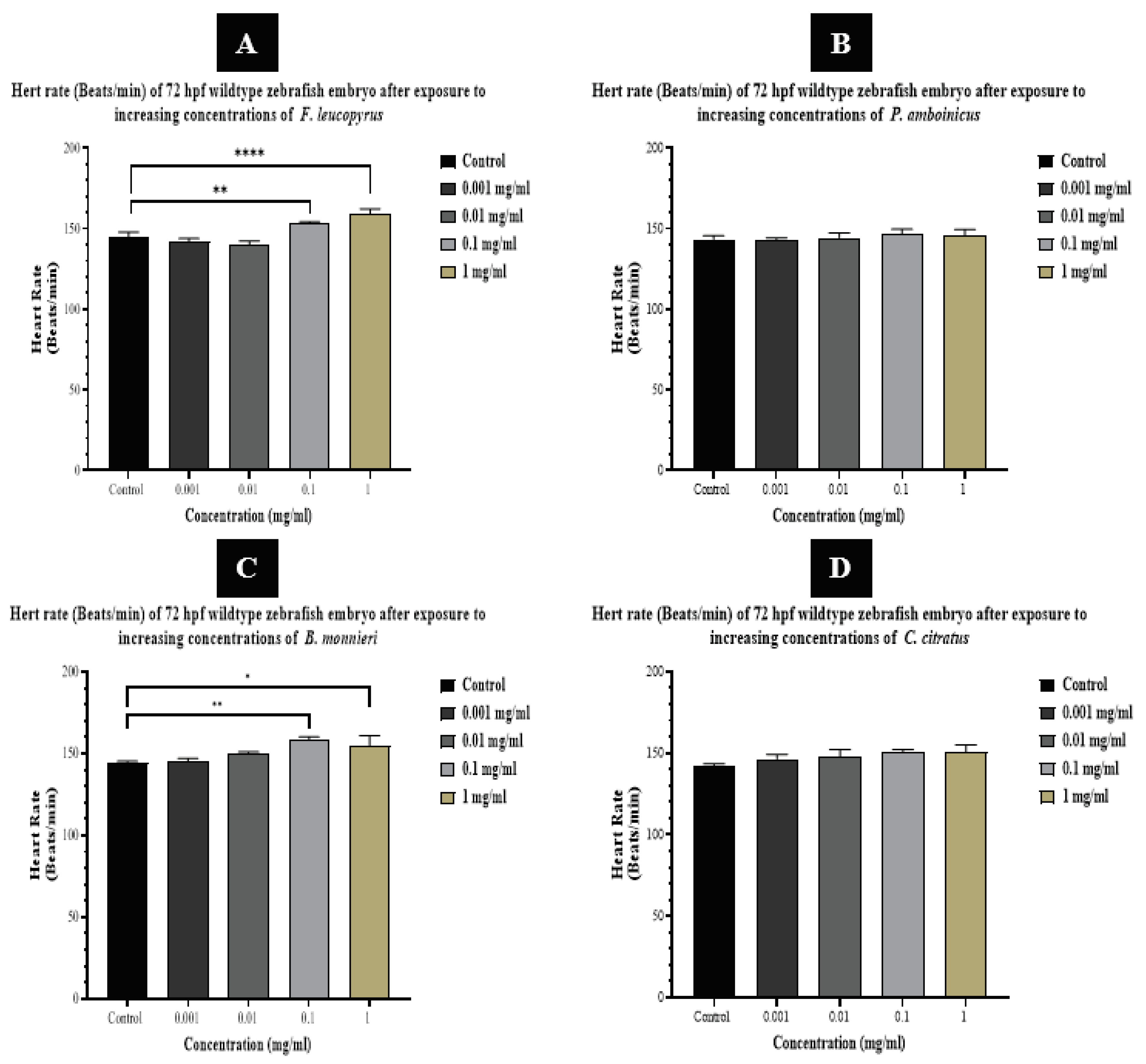
In high-dose treatment experiments with F. leucopyrus leaf extract, the heart rate of wild-type zebrafish embryos significantly increased at 0.1 mg/ml (P ≤ 0.01) and 1 mg/ml (P ≤ 0.0001) treatments at the 72-hour time point, with mean heart rates of 153.2 ± 1.1 beats/min and 159.2 ± 3.0 beats/min, respectively. Consequently, the heart rate of embryos treated with different concentrations of F. leucopyrus leaf extract varied from 139.8 ± 2.5 beats/min to 159.2 ± 3.0 beats/min. Similarly, in high-dose treatment experiments with B. monnieri whole plant extract, the heart rate of wild-type zebrafish embryos significantly increased at 0.1 mg/ml (P ≤ 0.01) and 1 mg/ml (P ≤ 0.05) treatments at the 72-hour time point, with mean heart rates of 158.2 ± 1.9 beats/min and 155.0 ± 6.1 beats/min, respectively. Thus, the heart rate of embryos treated with different concentrations of B. monnieri whole plant extract varied from 145.0 ± 2.0 beats/min to 158.2 ± 2.0 beats/min. Overall, none of the embryo heart rates exceeded the normal heart rate of zebrafish after exposure to each plant extract.
According to [48], changes in heart rate were observed in a dose-dependent manner with exposure to B. monnieri leaf extract and biosynthesized AgNPs, in comparison to the control group. The results have indicated a decrease in heartbeat rate as the concentration of the extract increases. But in the present study there was an increasement of the heart rates with the dose increasing.
- 4.
- Developmental Deformities
The structural deformities of wild-type zebrafish were observed to assess the toxicity of each plant extract on zebrafish in early developmental stages within the 0.001–10 mg/ml dose ranges at the 96-hour point. Three morphological differences were examined: Pericardial edema (PE), Yolk Sac edema (YSE), and Spinal Cord curvature (SCC) (Figure 08).
Figure 08.
Structural deformities (%) of wild-type zebrafish embryos at 96 hpf after exposure to increasing concentrations of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus aqueous extracts (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean percentage of structural deformities, and the X-axis represents the type of deformity observed in zebrafish at 96 hpf. (A) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of F. leucopyrus leaf extract. (B) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of P. amboinicus leaf extract. (C) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of B. monnieri whole extract. (D) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of C. citratus leaf extract. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
Figure 08.
Structural deformities (%) of wild-type zebrafish embryos at 96 hpf after exposure to increasing concentrations of F. leucopyrus, P. amboinicus, B. monnieri, and C. citratus aqueous extracts (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). The Y-axis represents mean percentage of structural deformities, and the X-axis represents the type of deformity observed in zebrafish at 96 hpf. (A) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of F. leucopyrus leaf extract. (B) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of P. amboinicus leaf extract. (C) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of B. monnieri whole extract. (D) Structural deformities (%) of 96 hpf wildtype zebrafish embryos after exposure to increasing concentration of C. citratus leaf extract. Asterisks denote the concentration at which zebrafish hatch rate is significantly different to controls (* = P ≤ 0.05, **= P ≤ 0.01, ***= P ≤ 0.001, ****= P ≤ 0.0001). Values represent mean ± SD.
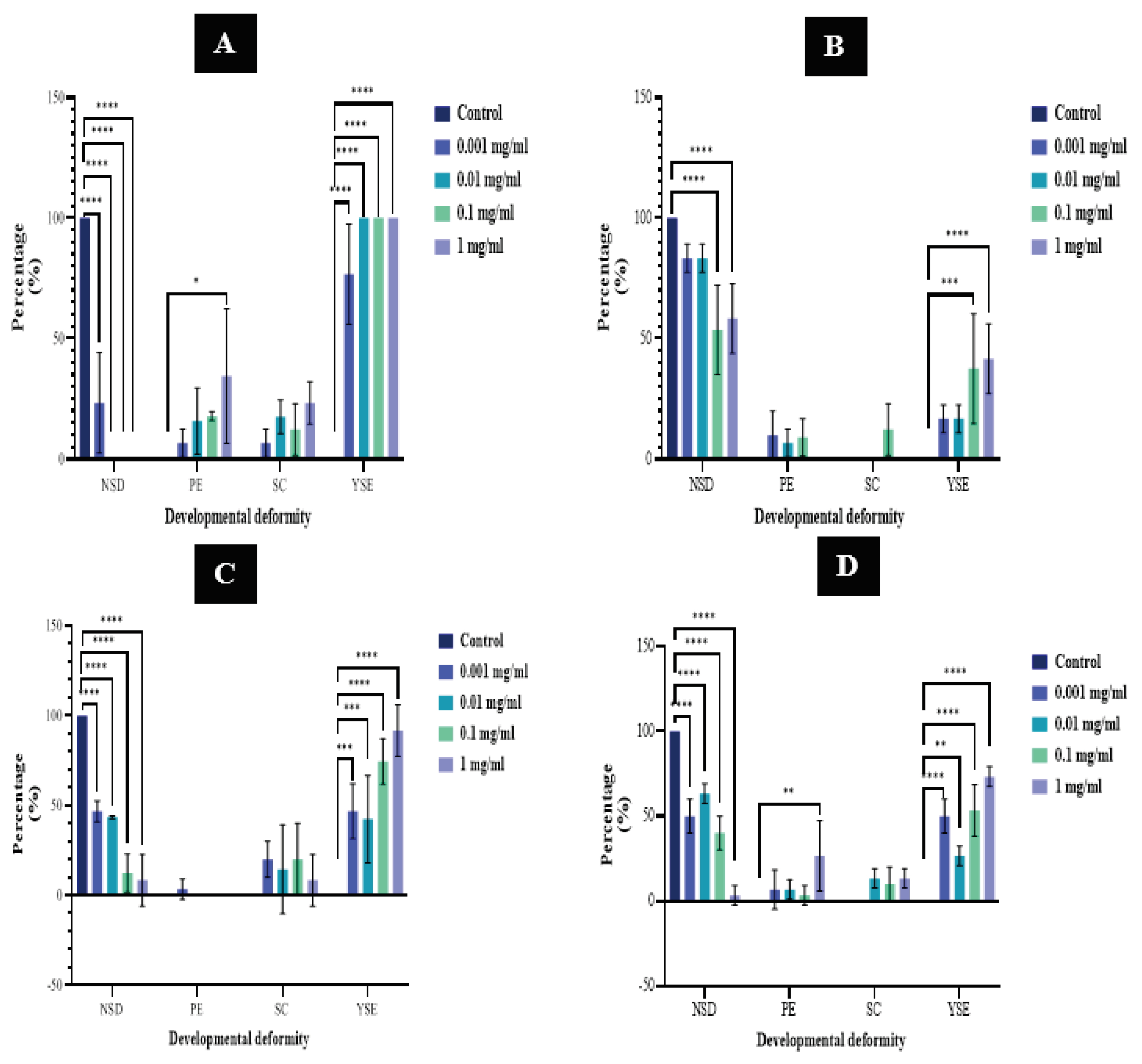
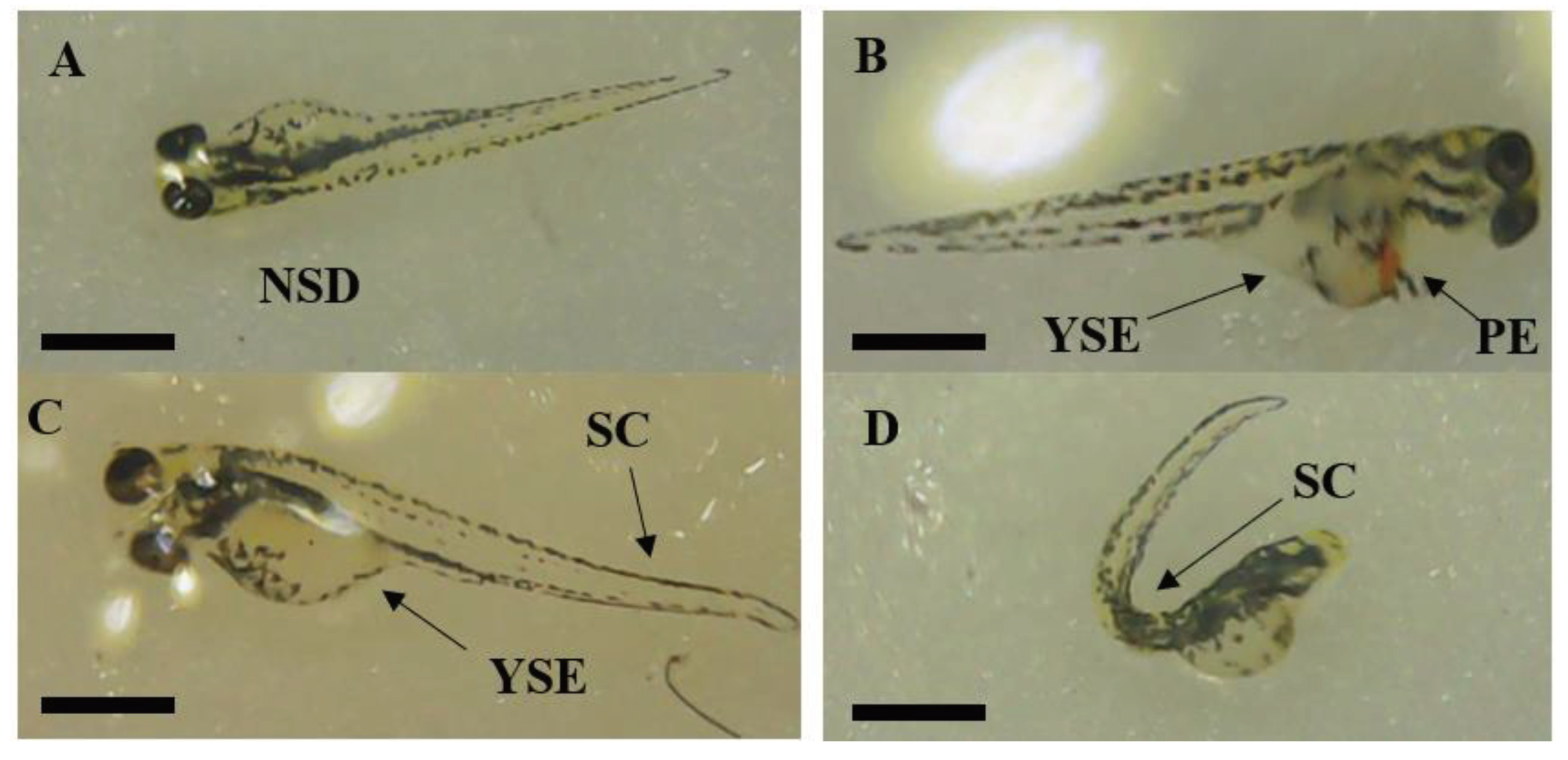
No abnormalities were observed in control embryos. Among wild-type zebrafish embryos exposed to F. leucopyrus leaf extract at various concentrations, only 23.3% ± 20.8 embryos at 0.001 mg/ml did not exhibit any structural deformities. All other surviving zebrafish exposed to F. leucopyrus displayed one of the three developmental abnormalities mentioned earlier (Figure 09).
Figure 09.
Structural deformities of 96 hpf zebrafish embryos after exposure to increasing concentration of F. leucopyrus leaf extract. (A) No Structural Deformities (NSD) in Control embryos. (B) Developmental abnormalities in zebrafish embryos exposed to 0.01 mg/ml of F. leucopyrus leaf extract showing pericardial edema (PE), and yolk sac edema (YSE). (C) 0.001 mg/ml of F. leucopyrus leaf extract showing yolk Sac edema (YSE). (D) 0.1 mg/ml of F. leucopyrus leaf extract showing spinal curvature (SC), and yolk sac edema (YSE). (E) 1 mg/ml of F. leucopyrus leaf extract showing spinal curvature (SC), and yolk sac edema (YSE), spinal curvature (SC). Scale bar = 3 mm in each image.
Figure 09.
Structural deformities of 96 hpf zebrafish embryos after exposure to increasing concentration of F. leucopyrus leaf extract. (A) No Structural Deformities (NSD) in Control embryos. (B) Developmental abnormalities in zebrafish embryos exposed to 0.01 mg/ml of F. leucopyrus leaf extract showing pericardial edema (PE), and yolk sac edema (YSE). (C) 0.001 mg/ml of F. leucopyrus leaf extract showing yolk Sac edema (YSE). (D) 0.1 mg/ml of F. leucopyrus leaf extract showing spinal curvature (SC), and yolk sac edema (YSE). (E) 1 mg/ml of F. leucopyrus leaf extract showing spinal curvature (SC), and yolk sac edema (YSE), spinal curvature (SC). Scale bar = 3 mm in each image.
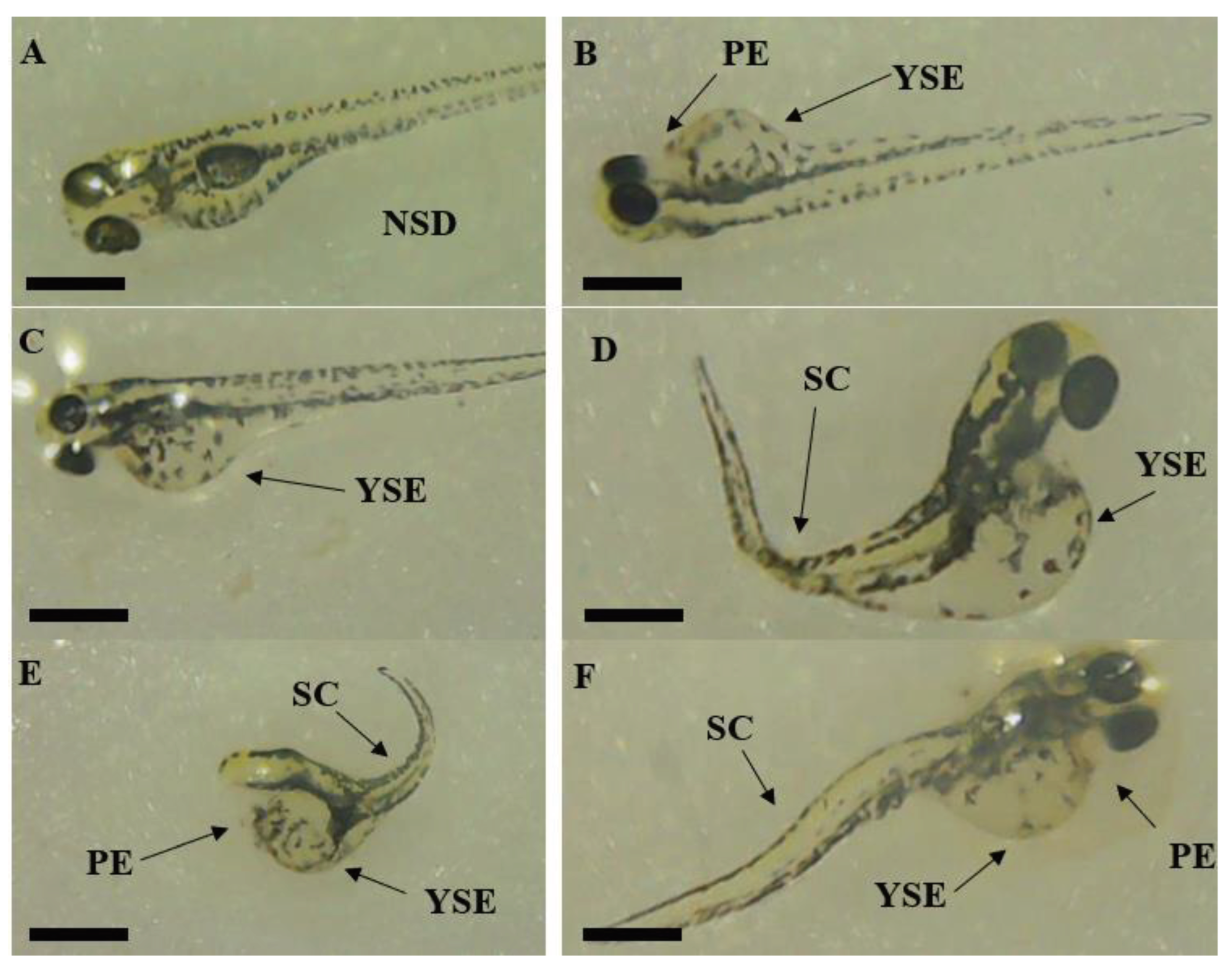
In B. monnieri treatment, the number of zebrafish larvae with NSDs decreased with increasing doses, and NSDs were significantly lower compared to controls at any concentration exposed. Among wild-type zebrafish embryos exposed to B. monnieri whole plant extract at different concentrations of 0.001, 0.01, 0.1, and 1 mg/ml, only 46.7% ± 5.8, 43.4% ± 0.9, 12.2% ± 10.7, and 8.3% ± 14.4 did not exhibit any structural deformities, respectively (Figure 10).
Figure 10.
Structural deformities of 96 hpf zebrafish embryos after exposure to increasing concentration of B. monnieri whole plant extract. (A) No Structural Deformities (NSD) in Control embryos. (B) Developmental abnormalities in zebrafish embryos exposed to 0.001 mg/ml of B. monnieri whole plant extract showing pericardial edema (PE), and yolk sac edema (YSE). (C) 0.01 mg/ml of B. monnieri whole plant extract showing yolk sac edema (YSE) and spinal curvature (SC). (D) 0.01 mg/ml of B. monnieri whole plant extract showing spinal curvature (SC). Scale bar = 3 mm in each image.
Figure 10.
Structural deformities of 96 hpf zebrafish embryos after exposure to increasing concentration of B. monnieri whole plant extract. (A) No Structural Deformities (NSD) in Control embryos. (B) Developmental abnormalities in zebrafish embryos exposed to 0.001 mg/ml of B. monnieri whole plant extract showing pericardial edema (PE), and yolk sac edema (YSE). (C) 0.01 mg/ml of B. monnieri whole plant extract showing yolk sac edema (YSE) and spinal curvature (SC). (D) 0.01 mg/ml of B. monnieri whole plant extract showing spinal curvature (SC). Scale bar = 3 mm in each image.
All other surviving zebrafish exposed to B. monnieri displayed one of the three developmental abnormalities mentioned earlier. Therefore, NSDs were observed to be significantly lower at doses of both F. leucopyrus and B. monnieri ranging from 0.001 to 10 mg/ml compared to the control.
Developmental abnormalities were comparatively fewer in embryos exposed to concentrations of 0.001 mg/ml and 0.01 mg/ml of P. amboinicus. Similarly, developmental abnormalities were comparatively fewer in embryos exposed to concentrations of 0.001 mg/ml, 0.01 mg/ml, and 0.1 mg/ml of C. citratus. A high number of NSDs were observed at all concentrations of P. amboinicus but significantly decreased at concentrations of 0.1 mg/ml and 1 mg/ml. Conversely, NSDs were significantly lower at all tested concentrations of C. citratus compared to control embryos.
The incidences of Spinal curvatures (SCs) and Pericardial edemas (PEs) were comparatively lower but detectable in all treatments of F. leucopyrus. Notably, PEs were significantly observed with F. leucopyrus at a dose of 1 mg/ml (P ≤ 0.05). Similarly, in the case of B. monnieri, SCs and PEs were comparatively lower, with SCs detectable at all tested concentrations and PEs observed only at a concentration of 0.001 mg/ml. Yolk Sac edemas (YSEs) were comparatively high and significantly observed in all treatments of F. leucopyrus and B. monnieri plant extracts. YSEs were present in 100% of F. leucopyrus leaf extracts at concentrations of 0.01, 0.1, and 1 mg/ml. YSEs were present in embryos exposed to B. monnieri whole plant extract at concentrations of 0.001, 0.01, 0.1, and 1 mg/ml, with averages of 46.7% ± 15.3, 42.3% ± 24.3, 74.4% ± 12.6, and 91.7% ± 14.4, respectively.
The wild-type zebrafish embryos exposed to doses of 0.001 and 0.01 mg/ml of P. amboinicus and 0.001 mg/ml dose of C. citratus, exhibited only PEs and YSEs. Among the wild-type zebrafish embryos exposed to P. amboinicus leaf extract at various concentrations, PEs were observed only at 10.0% ± 10.0 and 6.7% ± 5.8 for 0.001 mg/ml and 0.01 mg/ml, respectively, while YSEs were exhibited only at 16.7% ± 5.8 for both concentrations. From the wild-type zebrafish embryos exposed to C. citratus leaf extract at various concentrations, PEs were exhibited only in 6.7% ± 11.5 at 0.001 mg/ml, while YSEs were exhibited only in 50.0% ± 10.0. PEs were also comparatively less present at all tested concentrations but significant at 1 mg/ml. YSEs were comparatively higher compared to the other two abnormalities and were observed to be significant at all tested concentrations of C. citratus leaf extract. Furthermore, YSEs were significant only at 0.1 mg/ml (P ≤ 0.001) and 1 mg/ml (P ≤ 0.0001) concentrations of P. amboinicus leaf extract. With P. amboinicus, SCs were only observed at the 0.1 mg/ml dose, while they were comparatively low but present at all tested concentrations of C. citratus except at 0.001 mg/ml.
The most frequently observed developmental abnormality was yolk sac edema with all four plant extracts. Additionally, multiple abnormalities were observed in some of the survivors after 96 hpf at certain concentrations of the plant extracts tested. The results indicate that zebrafish embryos exposed to F. leucopyrus, B. monnieri, and C. citratus at concentrations of 0.001, 0.01, 0.1, and 1 mg/ml exhibited significantly higher YSEs among the survivors. Notably, YSEs were observed significantly only at doses of 0.1 and 1 mg/ml for P. amboinicus. Both SCs and PEs were significantly reduced with all plant extracts, although significant numbers of PEs were detected in F. leucopyrus and C. citratus only at a dose of 1 mg/ml. F. leucopyrus resulted in higher zebrafish survival compared to B. monnieri, but it also led to significantly more developmental abnormalities. Among the tested plants, C. citratus exhibited the highest embryo survival at the end of 96 hpf, with P. amboinicus ranking second. However, P. amboinicus showed significant developmental abnormalities only at 0.1 and 1 mg/ml, while C. citratus exhibited abnormalities at all concentrations.
In general, various studies have reported cases of edema, loss of blood circulation, and tail fin malformation observed in zebrafish embryos [57]. The occurrence of anomalies during embryonic development has been extensively studied in zebrafish. Osmotic balance plays a crucial role in maintaining physiological and metabolic mechanisms, particularly during early developmental stages. Fish embryos, in comparison, are more susceptible to exogenous toxicants, but the chorion layer serves a dual function by both shielding embryos from toxicants and acting as a membrane for osmotic balance in fish embryos [58]. A low osmotic environment is crucial for the precise development of embryos during the early stages. Studies conducted by [59] have highlighted the significance of maintaining osmotic balance, as an imbalance can disrupt metabolic and physiological mechanisms during the initial stages of embryonic development, leading to diminished embryogenesis and key developmental defects. Another study by [60] illustrated that the disruption of the circulatory system and osmoregulation are key phenomena that contribute to the development of mechanical forces, ultimately resulting in the collapse of the typical body structure and causing deformities in fish embryos. In a prior study, [61] reported bleeding tendency, yolk malformation, enlarged yolk, and shortened body length as abnormalities observed in zebrafish. The absence of embryo deaths, abnormalities, and teratogenic effects from some tested plant extracts suggests the safety of these plants.
The observed variations in toxicity and developmental effects between the brine shrimp lethality assay and the zebrafish embryo assay can be attributed to several factors. A significant factor is the divergence in extraction methods and solvents used in each assay, leading to the extraction of different sets of compounds with varying levels of toxicity and developmental effects. The brine shrimp lethality assay primarily focuses on acute toxicity, emphasizing immediate mortality. Results indicated that P. amboinicus and F. leucopyrus were non-toxic, suggesting that compounds extracted using the methanolic solvent diluted in 0.1% DMSO did not cause significant mortality in brine shrimp.
In contrast, the zebrafish embryo assay offers a more comprehensive assessment, evaluating both acute toxicity and developmental effects over time. The use of freeze-dried aqueous extracts in this assay introduces a different set of compounds compared to the methanolic extracts, contributing to variations in toxicity profiles and developmental impacts on zebrafish embryos. The higher survival of zebrafish embryos treated with F. leucopyrus, despite the presence of deformities in the survivors, suggests that certain compounds in F. leucopyrus may have protective effects on embryo survival, potentially enhancing survival rates despite deformities. On the other hand, B. monnieri exhibited low survival in the zebrafish assay, indicating potential toxicity towards zebrafish embryos. Interestingly, C. citratus, which showed weak toxicity in the brine shrimp assay, demonstrated the highest survival among the tested plants in the zebrafish assay. This suggests that compounds present in C. citratus may have protective or adaptive effects on zebrafish embryos, resulting in higher survival rates despite weak toxicity observed in the brine shrimp assay.
In summary, these differences underscore the significance of employing multiple assay systems to comprehensively evaluate the toxicity and developmental effects of plant extracts. Varied extraction methods, solvents, and the choice of assay organisms contribute to observed variations. Through the use of multiple assays, researchers can attain a more complete understanding of the potential toxic and developmental impacts of plant extracts.
So, these herbs possess potential as natural bio-preservatives for extending the shelf life of milk and milk products. Further investigation into various plant extract combinations is recommended, as their synergistic actions may enhance bacterial impact. To advance in this direction, it is recommended to conduct studies aimed at isolating and identifying the active compounds responsible for the observed antimicrobial and developmental effects of each plant extract. Such research would provide a solid basis for potential drug development and therapeutic applications. Moreover, to validate the safety and efficacy beyond zebrafish embryos, further in vivo studies using other animal models are warranted. Additionally, the potential environmental impact of using these plant extracts as bio-preservatives or pharmaceutical agents should not be overlooked. Exploring their effects on non-target organisms and the ecosystem is crucial to ensure their responsible use and minimize unintended consequences. Lastly, it is essential to underscore the significance of our findings in the context of the global challenges posed by antibiotic-resistant bacteria. With the rise of antimicrobial resistance, there is an urgent need for alternative and effective antimicrobial agents. The potent antimicrobial activity demonstrated by these plant extracts against a range of pathogenic bacteria highlights their potential as valuable candidates in the fight against antibiotic resistance.
3. Materials and Methods
3.1. Chemicals and Reagents
In the context of this research, a variety of essential chemicals and reagents were meticulously selected and sourced from reputable suppliers. Methanol was procured from Athula Enterprises, Kothalawala, Kaduwela, Sri Lanka. The laboratory essentials, including Muller Hinton agar (MHA), Muller Hinton broth (MHB), Dimethyl sulfoxide (DMSO), Nutrient agar (NA), and bacteriological agar, were obtained from Glorchem Enterprise, Colombo 11, Sri Lanka. Additionally, Gentamicin, was acquired from Rajya Osusala in Ragama, Sri Lanka.
3.2. Plant Extraction
Herbal plants, including Plectranthus amboinicus (Lour.) Spreng, Bacopa monnieri (L.) Wettst, Cymbopogon citratus (DC.) Stapf, and Flueggea leucopyrus Willd., were chosen based on a systematic literature review following PRISMA guidelines. The selection criteria took into account their demonstrated antimicrobial activity in prior studies, traditional Ayurvedic uses, and the feasibility of collection in Sri Lanka. Based on information and guidelines from Ayurveda doctors and traditional medical practitioners, all plants were collected in accordance with their natural habitats. Infection-free, fresh, and healthy plants were gathered from two distinct locations: Mawathagama, Kurunegala, and Katuwana, Hambantota, situated in the Northwestern and Southern provinces of Sri Lanka, respectively. All selected plants were authenticated by the Botany Division of the Bandaranaike Memorial Ayurvedic Research Institute, Nawinna, Maharagama where the voucher specimens have been deposited under accession number (Table 08)
Table 08.
Selected plant species for the investigations.
| Family | Species |
Local name1 English name2 |
Voucher Specimena | Part (S) of the plant used | Traditional uses |
| Lamiaceae | Plectranthus amboinicus (Lour.) Spreng | Kapparawalliya1 Country borage2 |
4103 | Leaves | Treatment of skin infections, mouth ulcers, hiccups, indigestion, and colic asthma. Often eaten raw or used as flavoring agents or incorporated as ingredients in the preparation of traditional food [62] |
| Plantaginaceae | Bacopa monnieri (L.) Wettst. | Lunuwila1 Water hyssop2 |
4104 | Whole plant | As a neuronal booster and cognitive enhancer [20]. As a curry [63]. |
| Phyllanthaceae | Flueggea leucopyrus Willd. | Katupila1 Spinous fluggea2 |
4105 | Leaves | Healing of chronic and non-healing wounds [64]. Treatment for cancers [37]. |
| Poaceae | Cymbopogan citratus (DC.) Stapf | Sera1 Lemon Grass2 |
4106 | Natural flavoring agents for food [65] |
a Plants were identified and authenticated at Bandaranaike Memorial Ayurveda Research institute, Nawinna, Maharagama. 1 Local name. 2 English name.
The collected plants were thoroughly washed in tap water to remove dust and muddy particles, followed by successive washings in distilled water for decontamination. The testing parts were then isolated. The washed plants were cut into small pieces and air-dried at room temperature (25±2 ∘C) in the shade until a constant mass was achieved [66]. Finally, the dried material was ground to a coarse powder in an electric food blender and stored in plastic containers for further analysis [67]. Leaves of F. leucopyrus, P. amboinicus, and C. citratus were used, while the whole plant was used in the case of B. monnieri.
3.2.1. Methanolic Plant Extract Preparation
Herbal plant extracts were prepared with minor modifications to the methodology proposed by [68]. The dry maceration procedure was followed, involving the mixing of a 1:5 ratio of plant material to methanol. Briefly, 100 g of dried plant material were soaked in 500 mL of methanol for 48 hours at room temperature (25 ± 2 ∘C), with occasional shaking. The plant material underwent a 48-hour maceration period. After two days, the plant extracts obtained were filtered through 4 layers of muslin cloth. Subsequently, they were concentrated to dryness and allowed to evaporate in the fume hood at room temperature. Dried extracts were then collected and stored at 4∘C (freezer) in the dark for further experiments [66]. Stock solutions of each plant extract were prepared in Falcon tubes with a concentration of 400 mg/ml in 10% dimethyl sulfoxide (DMSO) [69].
3.2.2. Freeze-dried Aqueous Extract Preparation
Each 100 g of powdered plant material was separately mixed with 500 ml of distilled water in a 1:5 ratio and soaked for 48 hours at room temperature (25 ± 2°C), with occasional shaking. After 2 days of maceration, the obtained extracts were filtered through 4 layers of muslin cloth. The filtrates were then transferred into 50 ml Falcon tubes and stored in a -20°C freezer in the laboratory until freeze-drying [70]. The filtrates were sent for freeze-drying at the University of Colombo using the LABCONCO freeze dryer. The process continued until a fine powder was obtained. The resulting yield was stored in sterilized airtight Falcon tubes in a -20 °C freezer for future use [71, 72]. Stock solutions of 400 mg/ml were prepared for each plant extract by dissolving 400 mg of crude plant material in 1 ml of distilled water.
3.3. Microbial Inoculum Preparation
Strains of bacteria, including Staphylococcus aureus (ATCC 25923), Salmonella spp., and Pseudomonas aeruginosa (ATCC 27853), were obtained from the culture collection of laboratories of Microbiology, Department of Biomedical Science, KIU, and Escherichia coli (ATCC 25922) from the Department of Microbiology, University of Colombo. Fresh bacterial inoculums were cultured and used throughout the research period. Pure bacterial cultures were maintained in nutrient agar at 4 ℃. Subcultures were maintained throughout the experiment using the streak plate method and incubated at 37 ℃ for 24 hours. Well-isolated colonies from overnight incubated subcultures were selected and transferred into a sterile Bijou bottle containing 5ml of sterile normal saline solution. The bacterial suspensions in Bijou bottles were uniformly mixed by vortexing. A 0.5 McFarland turbidity standard was prepared, and the bacterial suspension was adjusted to meet the 0.5 McFarland turbidity standards [73]. The 0.5 McFarland turbidity standard provides optical density comparable to a bacterial suspension with 1.5 × 108 colony-forming units (CFU/ml).
3.4. Antimicrobial Susceptibility Test
3.4.1. Agar Well Diffusion Assay
The antimicrobial activity of plant extracts was assessed using the agar well diffusion method. Muller Hinton agar (MHA) plates were prepared and labeled. Bacterial suspensions of 1-2 × 108 CFU/mL were obtained from fresh cultures and compared with 0.5 McFarland turbidity standards. The bacterial suspension was evenly seeded and streaked onto MHA plates using sterile cotton swabs [68]. Each MHA plate was inoculated using a sterile cotton swab, carefully swabbing from the top and moving from side to side down to the bottom of the plate. This process was repeated three times, rotating the plate approximately 60° each time to ensure an even distribution of inoculum, and finally, swabbing the rim of the agar. Inoculated plates were then allowed to dry at room temperature for 5 minutes. After drying, five holes/wells of 6 mm were punched in each plate using sterile micropipette tips (blue-colored). The bottom of the wells was sealed with molten MHA agar.
A stock solution of plant extract was prepared with a concentration of 400 mg/ml in 10% dimethyl sulfoxide (DMSO), and a dilution series was created for each plant extract using a twofold dilution (400 mg/ml, 200 mg/ml, 100 mg/ml, 50 mg/ml, 25 mg/ml, 12.5 mg/ml). Approximately 100 μl of different concentrations of methanolic extract dissolved in 10% DMSO was dispensed into labeled wells separately and allowed to diffuse at room temperature for 2 hours. Two plates were used for each plant extract against each organism: one plate for concentrations of 400 mg/ml, 200 mg/ml, 100 mg/ml, Negative control, and Positive control; and another plate for concentrations of 50 mg/ml, 25 mg/ml, 12.5 mg/ml, Negative control, and Positive control. For each organism, 10% DMSO served as the negative control, and Gentamicin (10 μg/ml) was used as the positive control. The plates were then incubated upright at 37°C for 18-24 hours [73].
After overnight incubation, the zones of inhibition (ZOI) were measured in millimeters and recorded separately. Antibacterial activity was confirmed by the presence of clear inhibition zones around each well. The diameter of the inhibition zone (mm) was measured. The experiment was conducted in triplicates, and the entire procedure was repeated thrice for each plant extract against each organism. For each replication, readings were taken in three fixed directions, and the average values were recorded [68].
3.4.2. Minimum Inhibitory Concentration (MIC)
The Minimum Inhibitory Concentration (MIC) was determined as the lowest concentration of extracts that inhibits visible growth (turbidity) of the test organisms after 24 hours of incubation. The MIC values were determined using the macro broth dilution method with some modifications. This test was conducted in sterile Khan tubes. The inoculum was prepared from 24-hour nutrient broth cultures, and suspensions were adjusted to 0.5 McFarland standard turbidity. Fifteen sterile Khan tubes were taken, and 1 ml of Muller Hinton broth (MHB) was added to each Khan tube, except the first one. To the first tube, 2 ml from the stock solution (400 mg/ml) of each plant extract dissolved in 10% DMSO was added. Subsequently, the plant extracts underwent a serial twofold dilution technique in sterile Khan tubes to prepare different concentrations for MIC determination, ranging from 400 mg/ml to 0.025 mg/ml. This was achieved by transferring 1 ml of the plant extract from the first tube to the second, and so on, until the last tube, where the last 1 ml was discarded. For each tube, 10 µl of microbial suspensions was added [74].
Tubes were sealed and incubated at 37 °C overnight for 24 hours. The experiments were performed in triplicate. Three control test tubes were included: one with MHB only (CB), another with the broth medium and sample/plant extract (CB+S), and the third with the medium and inoculums (CB+I), to serve as control tests. Gentamicin at 10 µg/ml was used as the positive control (Cp). After the overnight incubation, the growth of the test organisms was observed to determine the Minimum Inhibitory Concentration (MIC).
3.4.3. Minimum Bactericidal Concentration (MBC)
MBC was used to determine the minimum concentration of the plant extracts at which bacterial growth was completely inhibited, providing insights into their bactericidal activity. Bacterial cells from the MIC tubes were sub-cultured on freshly prepared solid nutrient agar by making streaks on the surface of the agar. The plates were divided into 8, 12, or 16 sections, depending on the tube number indicated in the MIC. Using a sterile straight wire, the inoculums were transferred from the lowest concentration that showed no visible microbial growth (MIC) to the highest concentration (stock solution) and cultured on nutrient agar [75]. The plates were incubated at 37 °C overnight for 24 hours. The section of the plate that exhibited no growth was considered the MBC for the plant extract used. The experiment was conducted in triplicate.
3.5. Testing on Isolated Milk Pathogenic Strains in Sterilized Milk Medium
The experimental procedures were conducted in accordance with the methodology suggested by [7], with minor adjustments. “Ambewela” sterilized fresh milk served as the growth medium to evaluate the antibacterial behavior and preservation activity of plant extracts against selected pathogens. Bacterial suspensions were prepared according to the 0.5 McFarland standard. The sterilized milk medium was inoculated with standard inocula of S. aureus, P. aeruginosa, Salmonella spp., and E. coli, supplemented with extracts from P. amboinicus, B. monnieri, C. citratus, and F. leucopyrus, each at a concentration of 5% (v/v). Microbial growth in the milk medium was observed at 37 °C for 24 hours at equal time intervals (6 hours, 12 hours, 18 hours & 24 hours). This involved transferring an inoculum from the tested milk medium supplemented with the respective plant extract and microorganism onto a nutrient agar plate using a straight wire at specific time intervals, followed by incubation of the plates at 37 °C for 24 hours.
3.6. Assessment of Biocompatibility and Toxicity
3.6.1. Brine Shrimp Lethality Assay
The brine shrimp lethality assay serves as a crucial tool for the preliminary cytotoxicity assessment of plant extracts and other substances, gauged by their ability to impact the survival of laboratory-cultured larvae (nauplii). The nauplii were exposed to varying concentrations of plant extracts for 24 hours, and the number of motile nauplii was calculated to determine the extract’s effectiveness. The experiment followed the methodology proposed by [33].
Three liters of seawater were measured and poured into a rectangular jar. The tip of an airline from an air pump was placed at the bottom of the water jar to maintain proper aeration. Fifteen grams of brine shrimp eggs were added at the top water level of the jar and mixed with the water. A light source (60–100-watt bulb) was positioned a few inches away from the jar and switched on. After 20-24 hours, the nauplii hatched. The eggs and nauplii were observed, and the nauplii were collected after an additional 24 hours. The hatched nauplii were separated from the empty eggs, and 10 nauplii were transferred into clean Khan tubes using a Pasteur pipette. This was achieved by turning off the air and switching off the lamp, causing the empty eggs to float while the brine shrimp concentrated in the water column. They were exposed to two different ten-fold diluted concentration series of plant extracts dissolved in 10% DMSO and 0.1% DMSO.
One set of tubes contained the actual concentrations of methanolic plant extracts dissolved in 10% DMSO, utilized for detecting antimicrobial and preservative activities against common milk pathogens. A tenfold dilution was prepared, ranging from 400 mg/ml to 0.004 mg/ml, in clean Khan tubes, each containing 10 nauplii in 1ml of sea water. This setup assessed the combined toxicity of 10% DMSO and plant extract. To evaluate the effect of 10% DMSO alone, a separate tube was dedicated, with 10 nauplii added to detect their survival and lethality.
For the other series, seven clean Khan tubes were taken and labeled. Plant extract weighing 100mg was measured using an analytical balance. A stock solution was then prepared by dissolving 100mg of plant extracts in 0.1% DMSO [32]. Concentrations of 100 mg/ml, 10 mg/ml, 1 mg/ml, 0.1 mg/ml, 0.01 mg/ml, and 0.001 mg/ml were prepared by serial 10-fold dilution. Subsequently, 1 ml of the prepared plant solutions was taken into six Khan tubes, each containing 10 nauplii and 1 ml of seawater. A negative control was established by adding 0.1% diluted DMSO with 10 nauplii in 1 ml of seawater to another tube. The number of survivors was counted, and the percentage of death/lethality (LC50) was calculated after 24 hours [33].
The mortality endpoint of this bioassay is defined as the absence of controlled forward motion during a 30-second observation. The percentage of lethality of the nauplii for each concentration and control was calculated. For each tube, the number of dead and live nauplii was counted, and the percentage of mortality was determined.
3.6.2. Zebrafish FET293 Embryo Assay
- Fish Fertilization Preparation
The zebrafish (Danio rerio) embryo toxicity experiments were conducted with wildtype zebrafish using a modified version of the OECD Fish Embryo Toxicity (FET) test [76]. For egg production, a breeding stock of unexposed, wild-type zebrafish with a well-documented fertilization rate of eggs was utilized. The fish were ensured to be free of macroscopic signs of infection and disease and had not received any pharmaceutical (acute or prophylactic) treatment for at least two months prior to spawning. Breeding fish were housed in aquaria with a recommended loading capacity of at least 1 L of water per fish and a fixed photoperiod of 12 - 16 hours.
In this study, wildtype adult zebrafish were housed in individual 1L tanks at a temperature of 27.0 °C, with a continuous water flow and a photoperiod of 12 hours light and 12 hours darkness (12L:12D). The aquarium water’s chemical parameters, including pH (7 ± 0.5) and electric conductivity (470 – 530 μs/m), were monitored daily. Additionally, nitrates (< 0.009 gm/l), nitrite (8-12 mg/l), and ammonia levels (<0.05 mg/l) were monitored weekly.
- 2.
- Eggs Production and Differentiation
Zebrafish spawning groups, housed in individual spawning tanks, were utilized to generate eggs. A 2:1 ratio of females to males was introduced into spawning tanks a few hours before the onset of darkness on the day before the test. Spawn traps were strategically placed in the tanks for efficient egg collection.
For the experiment, two wildtype adult zebrafish females and one male were moved to 3 L breeding tanks equipped with a 2 mm mesh insert and a transparent barrier separating males and females. The next morning, once the lights were on, the barrier was removed from each breeding tank, granting the zebrafish an hour for mating, spawning, and fertilization of the eggs. The petri dishes with the eggs were brought to the laboratory, where they underwent two rinses with aquatic water to eliminate adult debris. Subsequently, the eggs were transferred to new petri dishes, ensuring they were spread out without touching. Fertilized eggs exhibiting a transparent nature, undergoing cleavage, and displaying no apparent irregularities such as asymmetry, vesicle formation, or injuries to the chorion, were chosen through either digital microscopy or visual inspection. Coagulated embryos that exhibited a milk-white color were excluded from the selection. The chosen embryos were then incubated at 28°C in an incubator for 24 hours.
- 3.
- Start of Exposure and Embryo Treatment
The test commences immediately upon fertilization of the eggs and concludes after 96 hours of exposure. According to OECD guidelines, embryos should be immersed in the test solutions before blastodisc cleavage begins or, at the latest, by the 16-cell stage. In summary, ten zebrafish embryos at 24 hours post-fertilization (hpf) were introduced to each cell culture dish containing aquatic test media, zebrafish aquatics water (maintaining pH ± 0.5 and electric conductivity between 470 – 530 μs/ml), or E3 media (5mM NaCl, 0.17mM KCl, 0.33mM CaCl2, 0.33mM MgSO4, pH 7.2).
For this experiment, freeze-dried aqueous extracts were utilized. A tenfold dilution series with five concentrations was prepared for each plant extract, ranging from 10 mg/ml to 0.001 mg/ml in aquatic test media. Six sterile cell culture dishes were employed for one concentration series of each plant extract, including the negative control, with triplicates conducted for each plant extract. Eggs were allocated to well plates with 10 eggs on one plate for each test concentration of each plant extract and 10 eggs in dilution water (aquatic water) as a negative control on one plate. Zebrafish embryos in cell culture dishes were incubated in a controlled environment at 28 °C, 80% relative humidity, and a photoperiod of 12L:12D throughout the experiment. Zebrafish embryo hatching and mortality were assessed and recorded at 24, 48, 54, 72 and 80-hours post-fertilization (hpf) intervals.
- 4.
- Embryo Observation
Observations were conducted every 24 hours, extending up to 96 hours, utilizing digital microscopes for the examination of embryos. The percentage hatch rate, indicating the number of embryos hatched versus those not hatched at each time point, was calculated. Similarly, the percentage survival rate, representing the total number of living embryos versus dead embryos for each exposure dose at each time point, was reported [77].
The heart rate of zebrafish embryos (beats per minute) at 72 hpf was measured using a digital microscope connected to the Digital Viewer software and a stopwatch for both the control and each plant dose.
Photographs of zebrafish embryos exposed to control and varying plant extract doses were captured at 96 hpf using a digital microscope connected to the Digital Viewer software. These images were analyzed for developmental abnormalities, including no structural deformity (NSD), yolk sac edema (YSE), pericardial edema (PE), and spinal cord curvature (SSC). The percentage of each abnormality per well was then calculated.
3.7. Data Analysis
The experiments were conducted in triplicates, and the results were expressed as Mean ± SD. Data were compiled in an Excel database using GraphPad Prism 9.5.1 as analytical software. Mean ± SD and significant differences between the observations of control and exposed groups were analyzed using row statistics and two-way ANOVA, followed by Tukey’s multiple tests, respectively. A difference with a P-value < 0.05 was considered significant.
4. Conclusions
The results of the present study confirmed that B. monnieri and F. leucopyrus exhibited the highest antimicrobial activity against S. aureus and E. coli. B. monneiri showed superior activity against Salmonella spp., while C. citratus displayed the highest activity against P. aeruginosa. P. amboinicus exhibited relatively lower antimicrobial activity against all tested microorganisms. In the presence of specific microorganisms in sterilized milk, the addition of herbal plant extracts resulted in varying periods of viability reduction for milk pathogens. B. monnieri demonstrated the longest inhibition period for S. aureus, while F. leucopyrus showed the longest inhibition period for E. coli. Growth inhibition for Salmonella spp. and P. aeruginosa was minimal with individual plant extracts, but the combination of all four extracts exhibited the highest inhibition period for each organism. These herbs possess potential as natural bio-preservatives for extending the shelf life of milk and milk products. Further investigation into various plant extract combinations is recommended, as their synergistic actions may enhance bacterial impact.
The methanolic plant extracts, when diluted in 10% DMSO, resulted in significantly higher nauplii mortality due to the combined toxicity of DMSO and the plant extract. However, when dissolved in 0.1% DMSO, F. leucoyrys and P. amboinicus were found to be non-toxic, while B. monnieri and C. citratus exhibited weak toxicity. In the zebrafish embryo assay, early developmental stages demonstrated sensitivity to plant extract exposures (0, 0.001, 0.01, 0.1, 1, and 10 mg/ml). At 0.001 mg/ml, embryos hatched normally by the 72-hour time point, but hatch rates and survival decreased significantly with higher doses and time points. No survival was observed at the 10 mg/ml dose. Heart rates ranging from 139.8 to 159.2 beats/min were observed across all four plant extracts, falling within the normal range. P. amboinicus exhibited minimal developmental deformities, coupled with a high survival rate. Conversely, embryos exposed to B. monnieri and C. citratus showed significantly more developmental deformities than NSDs, despite high survival rates. While F. leucopyrus demonstrated high survival rates, the survivors displayed significantly more developmental abnormalities compared to the other three plant extracts. In conclusion, these plants demonstrated safety at low concentrations and the zebrafish embryo assay provided insights into the developmental effects of the plant extracts and this study opens promising avenues for future research and underscores the importance of a thorough investigation of these plant extracts. Understanding their active compounds, safety profiles, and environmental impact will be key in harnessing their full potential as effective antimicrobial agents and bio-preservatives in various applications.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. File S1: Supplementary materials.
Author Contributions
Conceptualization, K.J. and S.D.; methodology, K.J. and U.R.; software, K.J.; validation, K.J., S.D. and U.R.; formal analysis, K.J. and T.S.; investigation, K.J. and U.R.; resources, L.H. and J.D.; data curation, K.J.; writing—original draft preparation, K.J., S.D. and U.R.; writing—review and editing, K.J., S.D. and T.S.; visualization, K.J.; supervision, L.H. and J.D.; project administration, L.H.; funding acquisition, K.J. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
The animal study protocol was approved by the Institute of Biology (IOB), Sri Lanka (ERC IOBSL 19907 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors would like to express their gratitude to the Bandaranaike Memorial Ayurveda Research Institute, Nawinna, Maharagama, for the identification of plant materials, and to KIU Campus, Sri Lanka, for providing laboratory facilities.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| MIC | Minimum inhibitory concentration |
| MBC | Minimum bactericidal concentration |
| LC50 | Lethal concentration 50 |
| DMSO | Dimethyl sulfoxide |
| MHA | Mueller Hinton agar |
| MHB | Mueller Hinton broth |
| ZOI | Zone of inhibition |
| FET | Fish embryo toxicity |
| hpf | hours post fertilization |
| NSD | No structural deformity |
| YSE | Yolk sac edema |
| PE | Pericardial edema |
| SCC | Spinal cord curvature |
| SD | Standard deviation |
| AgNPs | Silver nano particles |
| OECD | Organization for Economic Co-operation and Development |
References
- Bezie, A. The Effect of Different Heat Treatment on the Nutritional Value of Milk and Milk Products and Shelf-Life of Milk Products. A Review. J. Dairy Vet. Sci. 2019, 11. [Google Scholar] [CrossRef]
- Fischer, W. J.; Schilter, B.; Tritscher, A. M.; Stadler, R. H. Contaminants of Milk and Dairy Products: Contamination Resulting from Farm and Dairy Practices. Encycl. Dairy Sci. Second Ed. 2011, 887–897. [Google Scholar] [CrossRef]
- R. Singh. Principles and Applications of Environmental Biotechnology for a Sustainable Future. Environ. Sci. 2017. [Google Scholar] [CrossRef]
- Berhe, G.; Wasihun, A. G.; Kassaye, E.; Gebreselasie, K. Milk-Borne Bacterial Health Hazards in Milk Produced for Commercial Purpose in Tigray, Northern Ethiopia. BMC Public Health 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Kongo, J. M.; Gomes, A. P.; Malcata, F. X. Monitoring and Identification of Bacteria Associated with Safety Concerns in the Manufacture of São Jorge, a Portuguese Traditional Cheese from Raw Cow’s Milk. J. Food Prot. 2008, 71, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Martin, N. H.; Torres-Frenzel, P.; Wiedmann, M. Invited Review: Controlling Dairy Product Spoilage to Reduce Food Loss and Waste. J. Dairy Sci. 2021, 104, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Siddique, F.; Latif, A.; Arshad, M.; Lashari, M. H. Antimicrobial Activity of Certain Herbal Plant Extracts against Pathogenic Microbes and Their Application in Sterilized Milk Medium. Pure Appl. Biol. 2021, 10, 378–387. [Google Scholar] [CrossRef]
- Velázquez-Sámano, G.; Collado-Chagoya, R.; Cruz-Pantoja, R. A.; Velasco-Medina, A. A.; Rosales-Guevara, J. Hypersensitivity Reactions to Food Additives. Rev. Alerg. Mex. 2019, 66, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Negi, P. S. Plant Extracts for the Control of Bacterial Growth: Efficacy, Stability and Safety Issues for Food Application. Int. J. Food Microbiol. 2012, 156, 7–17. [Google Scholar] [CrossRef]
- El-Sayed, S. M.; Youssef, A. M. Potential Application of Herbs and Spices and Their Effects in Functional Dairy Products. Heliyon 2019, 5. [Google Scholar] [CrossRef]
- Atanu, J. Application of Herbs in Functional Dairy Products – A Review. J. Dairy, Vet. Anim. Res. 2017, 5, 109–115. [Google Scholar] [CrossRef]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards Advances in Medicinal Plant Antimicrobial Activity: A Review Study on Challenges and Future Perspectives. Microorganisms 2021, 9, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Martínez, F. J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial Plant Compounds, Extracts and Essential Oils: An Updated Review on Their Effects and Putative Mechanisms of Action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef] [PubMed]
- Marjorie Murphy Cowan. Plant Products as Antimicrobial Agents. Am. Soc. Microbiol. 2007, 14, 128–130. [Google Scholar] [CrossRef]
- Ardalan, M.-R.; Rafieian-Kopaei, M. Is the Safety of Herbal Medicines for Kidneys under Question? J. nephropharmacology 2013, 2, 11–12. [Google Scholar]
- Oloya, B.; Namukobe, J.; Ssengooba, W.; Afayoa, M.; Byamukama, R. Phytochemical Screening, Antimycobacterial Activity and Acute Toxicity of Crude Extracts of Selected Medicinal Plant Species Used Locally in the Treatment of Tuberculosis in Uganda. Trop. Med. Health 2022, 50. [Google Scholar] [CrossRef] [PubMed]
- Ekor, M. The Growing Use of Herbal Medicines: Issues Relating to Adverse Reactions and Challenges in Monitoring Safety. Front. Neurol. 2014, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A. Evaluation of Antimicrobial Activity of Different Solvent Extracts of Saussurea Lappa. World J. Pharm. Pharm. Sci. 2017, 4, 12–18. [Google Scholar] [CrossRef]
- Mehta, J.; Utkarsh, K.; Fuloria, S.; Singh, T.; Sekar, M. Bioactive Molecules against Uropathogens — An In Silico Study to Identify Potential Lead Molecule ( s ) for the Development of New Drugs to Treat Urinary Tract Infections. 2022, No. Md.
- Fazlul, M. K. K.; Deepthi, S. P.; Irfan, M.; Farzana, Y.; Munira, B.; Nazmul, M. H. M. Antibacterial and Antifungal Activity of Various Extracts of Bacopa Monnieri. Int. J. Pharm. Res. 2019, 11, 1698–1702. [Google Scholar] [CrossRef]
- Karunarathnne, E. D. C.; Lokuwalpola, D. V; Sandaruwan, K. P. A. M.; Dabarera, M. D.; Wanigasekara, D. N. Antibacterial Activity of Flueggea Leucopyrus Willd. Int. Conf. Appl. Pure Sci. 2021, 31. [Google Scholar]
- Subramaniam, G.; Yew, X. Y.; Sivasamugham, L. A. Antibacterial Activity of Cymbopogon Citratus against Clinically Important Bacteria. South African J. Chem. Eng. 2020, 34, 26–30. [Google Scholar] [CrossRef]
- Sivaranjani, D.; Saranraj, P.; Manigandan, M.; Amala, K. Antimicrobial Activity of Plectranthus Amboinicus Solvent Extracts against Human Pathogenic Bacteria and Fungi. J. Drug Deliv. Ther. 2019, 9, 36–39. [Google Scholar] [CrossRef]
- Rowell, R. M.; Pettersen, R.; Han, J. S.; Rowell, J. S.; Tshabalala, M. A.; Service, F. - Cell Wall Chemistry; 2012. [CrossRef]
- Oskay, M.; Sari, D. Antimicrobial Screening of Some Turkish Medicinal Plants. Pharm. Biol. 2007, 45, 176–181. [Google Scholar] [CrossRef]
- Ginovyan, M.; Petrosyan, M.; Trchounian, A. Antimicrobial Activity of Some Plant Materials Used in Armenian Traditional Medicine. BMC Complement. Altern. Med. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.; Tapia-Rodríguez, M. R.; Baruzzi, F.; Ayala-Zavala, J. F. Plant Antimicrobials for Food Quality and Safety: Recent Views and Future Challenges. Foods 2023, 12. [Google Scholar] [CrossRef]
- Falya, Y.; Sumiwi, S. A.; Levita, J. Mini Review: Toxicity Study Of Plant Extracts. J. Pharm. Biol. Sci. 2020, 15, 25–32. [Google Scholar] [CrossRef]
- Chao, Wu. An Important Player in Brine Shrimp Lethality Bioassay: The Solvent. J. Adv. Pharm. Technol. Res. 2014, 5, 57–58. [Google Scholar]
- Sousa, M. I.; Correia, B.; Branco, A. F.; Rodrigues, A. S.; Ramalho-Santos, J. Effects of DMSO on the Pluripotency of Cultured Mouse Embryonic Stem Cells (MESCs). Stem Cells Int. 2020, 2020. [Google Scholar] [CrossRef]
- Geethaa, S.; Thavamany, P. J.; Chiew, S. P.; Thong, O. M. Interference from Ordinarily Used Solvents in the Outcomes of Artemia Salina Lethality Test. J. Adv. Pharm. Technol. Res. 2013, 4, 179–182. [Google Scholar] [CrossRef]
- George, M.; Josekumar, V. S. In Vitro Cytotoxicity Screening , Phytochemical Profile and Heavy Metal Analysis of Different Extracts of Acrostichum Heterophyllum L . 2016, 7 (March), 19–24.
- Sarah, Q. S.; Anny, F. C.; Misbahuddin, M. Brine Shrimp Lethality Assay. Bangladesh J. Pharmacol. 2017, 12, 186–189. [Google Scholar] [CrossRef]
- Meyer, B. N.; Ferrigni, N. R.; Putnam, J. E.; Jacobsen, L. B.; Nichols, D. E.; McLaughlin, J. L. Brine Shrimp: A Convenient General Bioassay for Active Plant Constituents. Planta Med. 1982, 45, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, C.; Maharaj, V. J.; Crouch, N. R.; Grace, O. M.; Pillay, P.; Matsabisa, M. G.; Bhagwandin, N.; Smith, P. J.; Folb, P. I. In Vitro Antiplasmodial Activity of Medicinal Plants Native to or Naturalised in South Africa. J. Ethnopharmacol. 2004, 92, 177–191. [Google Scholar] [CrossRef]
- Chalo, D. M.; Lukhoba, C.; Fidahuessein, D. S.; Nguta, J. M. Antimicrobial Activity, Toxicity and Phytochemical Screening of Selected Medicinal Plants of Losho, Narok County, Kenya. Biofarmasi J. Nat. Prod. Biochem. 2017, 15, 29–43. [Google Scholar] [CrossRef]
- Bulugahapitiya, V. P.; Munasinghe, M. M. A. B.; Hettihewa, L. M.; Kihara, N. Anti-Cancer Activity of <em>Fluggea Leucopyrus</Em> Willd (Katupila) against Human Ovarian Carcinoma and Characterization of Active Compounds. J. Sci. 2020, 11. [Google Scholar] [CrossRef]
- Sasidharan, S.; Chen, Y.; Saravanan, D.; Sundram, K. M.; Latha, L. Y. EXTRACTION, ISOLATION AND CHARACTERIZATION OF BIOACTIVE COMPOUNDS FROM PLANTS’ EXTRACTS. African J. Tradit. Complement. Altern. Med. 2011, 8, 93–130. [Google Scholar] [CrossRef]
- Paramasivam, D.; Balasubramanian, B.; Park, S.; Alagappan, P.; Kaul, T.; Liu, W.; Pachiappan, P. Phytochemical Profiling and Biological Activity of Plectranthus Amboinicus (Lour.) Mediated by Various Solvent Extracts against Aedes Aegypti Larvae and Toxicity Evaluation. Asian Pac. J. Trop. Med. 2020, 13, 494–502. [Google Scholar] [CrossRef]
- Charoenphon, N.; Kangwanrangsan, N.; Jiraungkoorskul, W. Artemia Salina Lethality and Histopathological Studies on Bacopa Monnieri Leaf Extract. Indian J. Anim. Res. 2018, 52, 610–614. [Google Scholar] [CrossRef]
- Mendes Hacke, A. C.; D’Avila da Silva, F.; Lima, D.; Rebuglio Vellosa, J. C.; Teixeira Rocha, J. B.; Marques, J. A.; Pereira, R. P. Cytotoxicity of Cymbopogon Citratus (DC) Stapf Fractions, Essential Oil, Citral, and Geraniol in Human Leukocytes and Erythrocytes. J. Ethnopharmacol. 2022, 291, 115147. [Google Scholar] [CrossRef]
- Chahardehi, A. M.; Arsad, H.; Lim, V. Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants. Plants 2020, 9, 1–35. [Google Scholar] [CrossRef]
- McGrath, P.; Li, C. Q. Zebrafish: A Predictive Model for Assessing Drug-Induced Toxicity. Drug Discov. Today 2008, 13, (9–10). [Google Scholar] [CrossRef]
- Parng, C.; Seng, W. L.; Semino, C.; McGrath, P. Zebrafish: A Preclinical Model for Drug Screening. Assay Drug Dev. Technol. 2002, 1 Pt 1, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Muller, E. B.; Lin, S.; Nisbet, R. M. Quantitative Adverse Outcome Pathway Analysis of Hatching in Zebrafish with CuO Nanoparticles. Environ. Sci. Technol. 2015, 49, 11817–11824. [Google Scholar] [CrossRef]
- Schoots, A. F. M.; Meijer, R. C.; Denucé, J. M. Dopaminergic Regulation of Hatching in Fish Embryos. Dev. Biol. 1983, 100, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Korwin-Kossakowski, M. Fish Hatching Strategies: A Review. Rev. Fish Biol. Fish. 2012, 22, 225–240. [Google Scholar] [CrossRef]
- Jansi, P. C. J.; Padua, J.; Freeda, R. In Vivo Toxic and Teratogenic Effects of Biologically Synthesized Silver Nanoparticles in The Embryos of the Zebrafish, Danio Rerio. Egypt. Acad. J. Biol. Sci. B. Zool. 2020, 12, 81–92. [Google Scholar] [CrossRef]
- Kent, M. L.; Buchner, C.; Barton, C.; Tanguay, R. L. Toxicity of Chlorine to Zebrafish Embryos. Dis. Aquat. Organ. 2014, 107, 235–240. [Google Scholar] [CrossRef]
- Wu, S. M.; Tsai, P. J.; Chou, M. Y.; Wang, W. Der. Effects of Maternal Cadmium Exposure on Female Reproductive Functions, Gamete Quality, and Offspring Development in Zebrafish (Danio Rerio). Arch. Environ. Contam. Toxicol. 2013, 65, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Piasecki, B.; Biernasiuk, A.; Skiba, A.; Skalicka-Woźniak, K.; Ludwiczuk, A. Composition, Anti-Mrsa Activity and Toxicity of Essential Oils from Cymbopogon Species. Molecules 2021, 26, 1–16. [Google Scholar] [CrossRef]
- Lawson, N. D.; Weinstein, B. M. In Vivo Imaging of Embryonic Vascular Development Using Transgenic Zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef]
- Herbert, S. P.; Stainier, D. Y. R. Molecular Control of Endothelial Cell Behaviour during Blood Vessel Morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564. [Google Scholar] [CrossRef]
- Rossant, J.; Howard, L. Signaling Pathways in Vascular Development. Annu. Rev. Cell Dev. Biol. 2002, 18, 541–573. [Google Scholar] [CrossRef] [PubMed]
- Massarsky, A.; Jayasundara, N.; Bailey, J. M.; Oliveri, A. N.; Levin, E. D.; Prasad, G. L.; Di Giulio, R. T. Teratogenic, Bioenergetic, and Behavioral Effects of Exposure to Total Particulate Matter on Early Development of Zebrafish (Danio Rerio) Are Not Mimicked by Nicotine. Neurotoxicol. Teratol. 2015, 51, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Li, H.; He, Q.; Yuan, W.; Chen, Z.; Yang, H. Developmental Toxicity of Diethylnitrosamine in Zebrafish Embryos/Juveniles Related to Excessive Oxidative Stress. Water. Air. Soil Pollut. 2018, 229. [Google Scholar] [CrossRef] [PubMed]
- Kiener, T. K.; Selptsova-Friedrich, I.; Hunziker, W. Tjp3/Zo-3 Is Critical for Epidermal Barrier Function in Zebrafish Embryos. Dev. Biol. 2008, 316, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Zhang, Y.; Yu, J.; Yu, Y.; Zhang, F.; Zhang, Z.; Wu, A.; Yan, X.; Zhou, Y.; Wang, F. Slc39a7/Zip7 Plays a Critical Role in Development and Zinc Homeostasis in Zebrafish. PLoS One 2012, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hill, A. J.; Bello, S. M.; Prasch, A. L.; Peterson, R. E.; Heideman, W. Water Permeability and TCDD-Induced Edema in Zebrafish Early-Life Stages. Toxicol. Sci. 2004, 78, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G. J.; Currie, P. D. Animal Models of Human Disease: Zebrafish Swim into View. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Joshua Garces, J. C.; Picardal, J. P.; Raymond, C. D.; Raymond Consuegra, C. D.; Michelle Custodio, F. B.; Paolo Pananganan, J. L.; Jean Ramirez, W. D. Teratogenicity and Acute Toxicity of Selected Philippine Indigenous Spices Using Brine Shrimp Lethality Assay and Zebrafish Assay. Pharmacophore 2020, 11, 88–105. [Google Scholar]
- Wickramaarachchi, D. L. C. K. F. W. W. U. I. Plectranthus Ambonicus and Plectranthus Zeylanicus: As Promising Medicinal Plants. Int. J. Sci. Res. 2018, 7, 1343–1347. [Google Scholar] [CrossRef]
- Barberyn Ayurveda Resorts. Ayurvedic Medicinal Plants of Sri Lanka Compendium. Barberyn Ayurveda Resorts and The Univhttps://herbssrilanka.com/rosemary/https://herbssrilanka.com/rosemary/ersity of Ruhuna. http://www.instituteofayurveda.org/plants/plants_detail.php?i=772&s=English_name.
- Sambhaji, D. T.; Shahan, A. A.; Kumar, V. In-Vitro Antimicrobial and Antioxidant Activity of Securinega Leucopyrus ( Willd ) Muell. 2020, 13 (September), 1251–1260.
- Cino Ceylon. Sera-Leaves-Lemon-Grass-Cymbopogon-Citratus. 2022. https://www.cinoceylon.com/cinoceylon-products/spices/sera-leaves-lemon-grass-cymbopogon-citratus.html.
- Nandapala, J. H. Y. P.; Napagoda, M. T.; Weerasinghe, N. P. Anticandidal Activity of Ten Selected Medicinal Plants from Southern and North Central Provinces of Sri Lanka. Int. J. KIU 2022, 3, 32–40. [Google Scholar] [CrossRef]
- Begashaw, B.; Mishra, B.; Tsegaw, A.; Shewamene, Z. Methanol Leaves Extract Hibiscus Micranthus Linn Exhibited Antibacterial and Wound Healing Activities. BMC Complement. Altern. Med. 2017, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Rivas Morales, C.; Castillo, S.; Leos-Rivas, C.; García-Becerra, L.; Ortiz Martínez, D. M. Antibacterial and Antibiofilm Activity of Methanolic Plant Extracts against Nosocomial Microorganisms. Evidence-based Complement. Altern. Med. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Alipoor, J.; Dabirzadeh, M.; Marofi, Y.; Sefidgar, S. A. A. Effects of Methanolic Extract of Ficus Carica Leaves on Cystic Echinococcosis. Med. Lab. J. 2017, 11, 15–20. [Google Scholar]
- Handa, S. S.; Khanuja, S. P. S.; Longo, G.; Dutt, D. Extraction Technologies for Medicinal and Aromatic Plants. United Nations Ind. Dev. Organ. Int. Cent. Sci. High Technol. 2008.
- Randima, K. A. D. K.; Perera, P. K.; Arawwawala, L. D. A. M. Comparative Analysis of Phytochemical and Antioxidant Activities of the Nishatipal Decoction and It ’ s Freeze Dried Powder. SLJIM 2020, 05, 329–339. [Google Scholar]
- How, Y. K.; Siow, L. F. Effects of Convection-, Vacuum-and Freeze-Drying on Antioxidant, Physicochemical Properties, Functional Properties and Storage Stability of Stink Bean (Parkia Speciosa) Powder. J. Food Sci. Technol. 2020, 57, 4637–4648. [Google Scholar] [CrossRef] [PubMed]
- Debalke, D.; Birhan, M.; Kinubeh, A.; Yayeh, M. Assessments of Antibacterial Effects of Aqueous-Ethanolic Extracts of Sida Rhombifolia’s Aerial Part. Sci. World J. 2018, 2018. [Google Scholar] [CrossRef]
- Rakholiya, K. D.; Kaneria, M. J.; Chanda, S. V. In Vitro Assessment of Novel Antimicrobial from Methanol Extracts of Matured Seed Kernel and Leaf of Mangifera Indica L. (Kesar Mango) for Inhibition of Pseudomonas Spp. and Their Synergistic Potential. Am. J. Drug Discov. Dev. 2015, 5, 13–23. [Google Scholar] [CrossRef]
- Parvekar, P.; Palaskar, J.; Metgud, S.; Maria, R.; Dutta, S. The Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Silver Nanoparticles against Staphylococcus Aureus. Biomater. Investig. Dent. 2020, 7, 105–109. [Google Scholar] [CrossRef]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test. OECD Guidel. Test. Chem. Sect. 2, OECD Publ. 2013, No. July, 1–22.
- Almeida, A. R.; Salimian, M.; Ferro, M.; Marques, P. A.; Goncalves, G.; Titus, E.; Domingues, I. Biochemical and Behavioral Responses of Zebrafish Embryos to Magnetic Graphene/Nickel Nanocomposites. Ecotoxicol. Environ. Saf. 2019, 186, 109760. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



