
Submitted:
28 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
We conducted a pilot study to analyze the differential methylation status of 20 primary acinar adenocarcinomas of the lungs. These adenocarcinomas had to be wild type in mutation analysis and had either high (TPS>50%; n=10) or low (TPS<1%; n=10) PD-L1 status to be integrated into our study. To examine the methylation of 866,895 specific sites, we utilized the Illumina Infinium EPIC bead chip array. Both hypermethylation and hypomethylation play significant roles in tumor development, progression, and metastasis. They also impact the formation of the tumor microenvironment, which plays a decisive role in tumor differentiation, epigenetics, dissemination, and immune evasion. The gained methylation patterns were correlated with PD-L1 expression. Our analysis has identified distinct methylation patterns in lung adenocarcinomas with high and low PD-L1 expression. After analyzing the correlation between methylation results of genes and promoters with their pathobiology, we found that tumors with high expression of PD-L1 tend to exhibit oncogenic effects through hypermethylation. On the other hand, tumors with low PD-L1 expression show loss of their suppressor functions through hypomethylation. The suppressor functions of hypermethylated genes and promoters are ineffective compared to simultaneously activated dominant oncogenic mechanisms. The tumor microenvironment supports tumor growth in both groups.
Keywords:
Lung cancer
; Epigenetic profiling
; Methylome
; Methylation analysis
; Programmed cell death ligand 1
; Precision medicine
1. Introduction
Lung cancer is a widespread form of cancer. The chances of surviving NSCLC vary from 19%-63% depending on the stage and available treatment options [1,2]. In real-world data analysis, the median overall survival was 351 days for all patients and 571 days for those with pathophysiologically relevant molecular aberrations.[3] Recent advances in precision medicine have improved outcomes for select patients with NSCLC who have activating mutations and/or activating translocations, particularly those with pulmonary adenocarcinomas.[4] Moreover, immune therapy with checkpoint inhibitors, particularly PD-1 and PD-L1 inhibitors, has shown promising outcomes with partial or even complete tumor regression in the case of pulmonary adenocarcinomas [5,6]. Biomarkers have been developed to identify molecular targets and select the most effective medications for individual patients. To treat lung cancer accurately, it is necessary to perform a precise diagnosis of the underlying tumor type, followed by a molecular panel that includes immunohistochemistry and next-generation sequencing (NGS) with RNA and DNA analyses in the case of NSCLC.[7]
In our routine investigation, we identify molecular targets relevant to therapy. These include mutations in EGFR, BRAF, KRAS, ERBB2, STK11, and rearrangements such as ALK, RET, ROS1, NTRK, and cMET Exon 14 splice mutation. Tumors such as lung cancer alter their microenvironment to decrease the effectiveness of physiological anti-tumor mechanisms[8]. Cancer cells upregulate PD-L1 to evade immune-regulated destruction [9]. PD-1 and PD-L1 checkpoint inhibitors block PD-L1, enabling them to destroy tumor cells again. Therefore, the expression of PD-L1 measured with the tumor proportion score (TPS) is used as a biomarker to predict the probability of immune-therapy response in lung cancer [10,11]. TPS is the percentage of stained viable tumor cells with complete or partial membranous staining at any intensity. The higher the TPS, the better the individual’s response to checkpoint inhibitor therapy is assumed[12]. Immune or target monotherapies can be combined with chemo- and radiotherapy in later therapy lines to improve and prolong therapy success[13]. Some patients become long-term survivors with a chronification of lung cancer.[14] Changes in the immune status result in changes in the methylation status in large tumor regions[15]. We conducted a comprehensive epigenomic methylation analysis to identify such molecular mechanisms in lung cancer with different PD-L1 statuses (i.e., PD-L1 high, TPS>50% and PD.L1 low, TPS<1%) and correlated the results with PD-L1 expression. Database searches such as GeneCards, National Institutes of Health, Alliance of Genome Resources, and The Cancer Genome Atlas (TCGA) outlined the pathobiology functions of genes and promotors significantly differentially methylated.
2. Materials and Methods
2.1. Tissue Collection and Immunohistochemical Analysis
Our study used formalin-fixed and paraffin-embedded (FFPE) archived samples from the University Institute of Pathology, University Hospital Salzburg, and Paracelsus Medical University. Salzburg, Austria. All tumors were primary pulmonary acinar adenocarcinomas with either high PD-L1 expression (TPS > 50%; n=10) or low PD-L1 expression (TPS < 1%; n=10). Standard molecular analyses were performed during routine workup, including DNA and RNA panels (described below). Only cases without molecular events in target genes or gene regions were included in the present study. The PD-L1 status of tumor cells was also determined during routine utilizing the DAKO 22C3pharm DX kit from Agilent, USA. Immunostainings were performed on a DAKO-Omnis System(Agilent, USA), and the tumor proportion score (TPS) was calculated according to the rules determined by the US Food and Drug Administration (FDA) and the National Comprehensive Cancer Network (NCCN).[11,16] For our differential methylation analysis, we selected 10 cases with high PD-L1 expression (TPS >50%, mean 82%) and ten patients with low PD-L1 expression (TPS <1%, mean 0,5%), as defined in Figure 1 and Example Images 1-4.
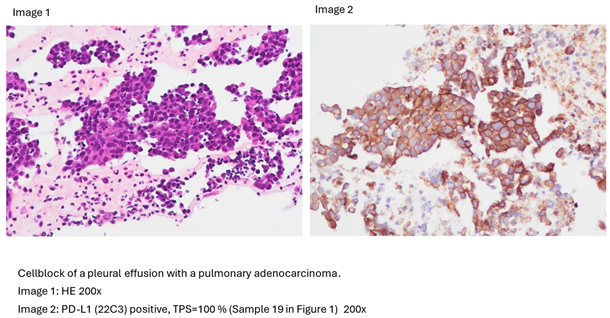
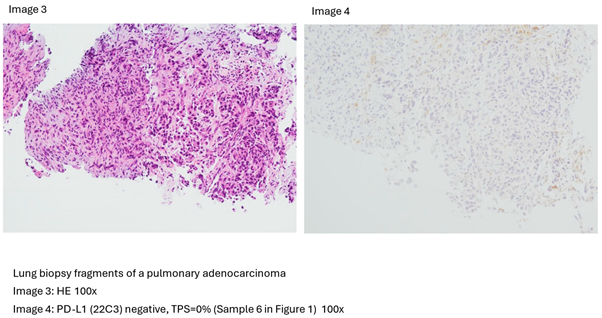
2.2. Molecular and Genetic Analysis
Tumor specimens with a minimum of 60% tumor content were used to ensure accurate mutation analysis. In some cases, we increased the tumor content by microdissecting the area of interest in tissue slides. We followed the manufacturer’s protocol for DNA extraction using a Maxwell system from Promega in Fitchburg, WI, USA. For mutation analysis of lung adenocarcinomas, we used either the AmpliSeq for Illumina Cancer Hotspot Panel v2 or the AmpliSeq for Illumina Focus Panel (Illumina, San Diego, California, USA). Next-generation sequencing was performed on an Illumina MiniSeq device following the manufacturer’s protocol. We used an Infinium Methylation EPIC Bead Chip and the Illumina protocol to identify methylation patterns for genome-wide screening[17].
2.3. Computational Data Analysis
We adhered to the three customary procedures for computational analysis of DNA methylation data, which included (I) data processing and quality control, (II) data visualization and statistical analysis, and (III) validation and interpretation.
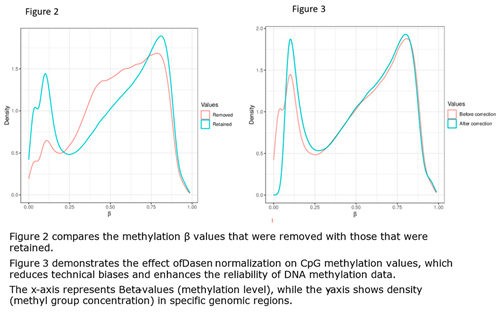
We analyzed methylation data using the Illumina Genome Studio Methylation Module and RnBeads on R statistical software[18]. Our mapping was based on the human genome reference builds (GRCh38.p14 patch release of the hg38 assembly). DNA methylation beta values were used, indicating the ratio of the intensity of the methylated bead type in variables between 0 and 1, depending on the combined locus intensity (ranging from 0% to 100% methylation). To ensure the reliability of our results, we removed probes enriched with single nucleotide polymorphisms (SNPs) and used Greedycut filtering to eliminate unreliable measurements with p>0.05 in an iterative algorithm. We also removed context-specific probes and those on sex chromosomes. The Dasen method was utilized to normalize beta values.[19] (please see Figures 2 and 3). Probes were marked with four genomic areas, including tiling regions (which were 5000 nucleotides long), genes (version Ensemble genes 75), promoters (version Ensemble genes 75), and CpG islands (CpG island track of the UCSC Genome Browser)[20]. We used the Limma method (linear model for microarray data) for on-site level p-value computation, a hierarchical linear model using an empirical Bayes approach.[21] To identify sites exhibiting differential variability between two sample groups, we used the diffVar method integrated into the missMethyl package and nominal p-values (nominal significances) as performed in other Epigenome-Wide Association Studies (EWAS)[22,23].
3. Results
Our analysis focused on identifying significant differences in the methylation status between PD-L1 high adenocarcinomas (TPS > 50%) and PD-L1 low cases (TPS < 1%) without any known relevant molecular aberration, with a particular interest in gene and promoter methylation status. The results showed a significantly different methylation pattern in both groups. (please see Figures 4 and 5)
We analyzed 20 samples with 866,895 methylation sites but had to exclude 17,371 sites due to overlapping with SNPs and 7,532 sites after applying the Greedycut algorithm. Additionally, we removed 18,597 probes located on sex chromosomes and 2,915 context-specific probes. We kept all samples but removed 46,415 probes, thus retaining 820,480 probes for final analyses. (Figure 3)
Our analysis identified 252,729 annotations in the tiling regions (length 5,000), 34,988 annotations in genes, 44,852 annotations in promoters, and 26,540 annotations in CpG islands. We only report the highest-ranking results previously linked to lung cancer in GeneCards and Alliance of Genome Resources. The detailed outcomes of the highest-ranking results can be found in Table 1.
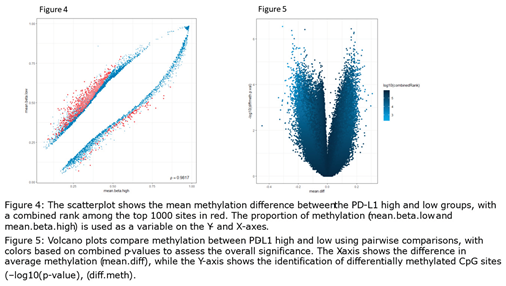
We classified genes and promoters based on the percentage of hypermethylation and hypomethylation and the most significant differences (delta values) in methylation in cases with high and low PD-L1 expression.
- Hypermethylated genes and promotors in PD-L1 high-expressing cases:
In our study, hypermethylation in the PD-L1 high group ranged from 73% to 90% methylation with p-values from 0.001 to 0.04. (Please refer to TABLE 1 for precise values). Hypermethylation was observed in the following locations in this group: SNORD114-14 (C/D Box 114-14, Small Nucleolar RNAs, snoRNAs), DCAF4L2 (DDB1 Associated Factor 4 Like 2 ), CELF2-AS1 (CELF2 Antisense RNA 1), LINCMD1 (Long Intergenic Non-Protein Coding RNA, Muscle Differentiation, MIR133BHG), LINC00528 (Long Intergenic Non-Protein Coding RNA 528). The most significant difference in methylation (delta value) between PD-L1 high and low expression groups was found in S100A7L2 (S100 Calcium Binding Protein A7 Like 2) (delta value = 17%, p-value = 0.002) and SOD1P3 (Superoxide Dismutase 1 Pseudogene 3) (delta value =14% and p-value = 0,02).
- Hypomethylated genes and promotors in PD-L1 high-expressing cases:
Hypomethylation was observed in genes and promoters of PD.L1 high-expressing adenocarcinomas at rates ranging from 14% to 21%, with p-values between 0.007 and 0.03. This was seen in the following locations: CAPS2 (cyclase-associated protein 2, calcyphosine 2), GLIPR1L2 (GLI Pathogenesis Related 1 Like 2), and IFITM3 (Interferon Induced Transmembrane Protein 3).
- Hypermethylated genes and promotors in PD-L1 low-expressing cases:
Our study found that tumors with low PD-L1 expression had hypermethylation in SNORD114-14 (methylation 90 % and p-value 0,04), while all other top results were pseudogenes or irrelevant to humans.
- Hypomethylated genes and promotors in PD-L1 low-expressing cases:
Locations with the lowest methylation in PD-L1 low expressing cases were found in miR124-3 (MicroRNA124-3), TRIM71 (Tripartite Motif Containing 71, LIN41), CAPS2 (Calcyphosine 2), UBE2QL1 (Ubiquitin Conjugating Enzyme E2 Q Family Like 1), and GLIPR1L2 (GLIPR1 Like 2). Methylation rates ranged from 15% to 30%, with p-values between 0.007 and 0. 01. The most significant methylation delta values between PD-L1 low/high were found for the NUMB gene (NUMB Endocytic Adaptor Protein) and LINC00528, with delta values of 19% / 20% and p-values of 0.001 and 0,0005.
4. Discussion
The expression of genes can differ due to various epigenetic mechanisms cells use to regulate DNA functions. One of these mechanisms is DNA methylation, which alters gene expression without changing its sequence. This modification can affect gene expression during cell differentiation. Methylation of a promoter region can regulate nearby gene expression, and excessive methylation can cause silencing mainly of DNA repair genes[24]. This is believed to be an early step towards cancer progression, as hypermethylation of DNA upstream blocks access to transcription factors and enzymes, ultimately inhibiting downstream gene activity[25]. Conversely, many tumors exhibit hypomethylated carcinogenic genes when compared to normal tissue[26,27]. Activating typically silenced genes can contribute to developing malignant neoplasias as individuals age. Both hypermethylation and hypomethylation are leading causes of oncogenesis, the former being more frequent and occurring at the CpG islands in the promoter region of the genes. In contrast, the latter occurs globally in various genomic sequences[28].
Several genes exhibit significant methylation status, directly influencing angiogenesis, active oxygen, calcium, and vessels closely related to the tumor microenvironment. [29] Additionally, the methylation status of several genes and promoters is associated with modifying the status of immune cells such as macrophages, lymphocytes, or neutrophils.[30] These genes and promoters help create an optimized microenvironment for tumor growth and development.[24,31]
Our research revealed significant differences in the methylation levels of genes and promoters between pulmonary adenocarcinomas with high and low expression of PD-L1. A few genes and promoters had similar methylation status simultaneously in both groups.
Pathobiological Mechanisms of Hypermethylated Genes and Promotors in the PD-L1 High-Expressing Tumors
SNORD114-14 and other small nucleolar RNAs (snoRNAs) are crucial in developing and spreading lung cancer. They control the balance between cell growth and death and promote the adaptability of cancer cells. SnoRNAs display both oncogenic and tumor-suppressive activities that are vital in the formation and progression of lung cancer. Dysregulation of snoRNAs is a contributing factor to lung cancer tumorigenesis and progression.[32]
DCAF4L2 (DDB1 Associated Factor 4 Like 2 ) is a protein-coding gene. Studies have revealed that the levels of DCAF4L2 are higher in patients with lung cancer and colorectal cancer, which can lead to more advanced stages of the disease and the spread of cancer cells to other parts of the body.[33]
CELF2-AS1 (CELF2 Antisense RNA 1) suppresses non-small cell lung carcinoma growth by inhibiting the PREX2-PTEN interaction, which regulates cell proliferation. Additionally, CELF2 protein expression is downregulated in tumor tissues as lung cancer and is associated with poor prognosis.[34]
LINCMD1 (Long Intergenic Non-Protein Coding RNA, Muscle Differentiation, MIR133BHG) inhibits cell proliferation, migration, and invasion of lung adenocarcinoma.[35,36]
LINC00528 (Long Intergenic Non-Protein Coding RNA 528) was identified as a long non-coding RNA found in computational analyses to be associated with neutrophils, lymphocytes, and macrophages involved in the immune reaction of the tumor microenvironment and strongly correlated with immunotherapy prognosis in lung cancer. The exact mechanism of how LINC00528 affects the immune system is not fully understood, but it is believed to regulate genes involved in the immune response.[37,38]
Despite being classified as a pseudogene, S100A7L2 (S100 Calcium Binding Protein A7 Like 2) has been found to play a significant role in inducing transdifferentiation from lung adenocarcinoma to squamous carcinoma. Moreover, it has been observed that the expression of this gene is regulated differently by the Hippo-YAP pathway in lung cancer cells, which contributes to tumor cell growth[39]. S100A7L2 is also involved in cell migration and invasion, creating a proinflammatory and proangiogenic environment that promotes tumor progression and metastasis.[40]
The gene SOD1P3 (Superoxide Dismutase 1 Pseudogene 3) substantially influences cancer development. SOD3 and SOD1P3 regulate active oxygen in the microenvironment and are down-regulated in lung cancer.[41] Furthermore, SOD1P3 regulates the processes of angiogenesis, metastasis, and invasion in lung cancer by controlling the levels of Interleukin6 and VEGF[42].
Pathobiological Mechanisms of Hypomethylated Genes and Promotors in the PD-L1 High-Expression Group
Our study found hypomethylation of specific genes and promoters (CAPS2, GLIPR1L2, and IFITM3) in the PD-L1 high-expression group.
CAPS2 (Calcyphosine 2) is overexpressed in lung cancer, where it promotes cell proliferation, migration, invasion, and metastasis by activating the ERK/MAPK and AKT signaling pathways. [43] The expression of CAPS2 is higher in lung cancer tissues when compared to normal lung tissues, and it is linked to the stage of the tumor, lymph node metastasis, and poor survival. As such, CAPS2 can be used as a biomarker to monitor lung cancer progression following therapy.[44]
GLIPR1L2 plays a significant role in cancer and immune defense. GLI pathogenesis-related 1 functions as a tumor suppressor in lung cancer, and during lung tumorigenesis, the expression of GLIPR1L2 is down-regulated. [45]
IFITM3 (Interferon-Induced Transmembrane Protein 3) regulates the growth and invasion of human lung adenocarcinoma and is believed to be a crucial factor in promoting carcinogenesis. According to recent research, IFITM3 expression levels are directly associated with tumor differentiation, lymph node, distant metastasis, and tumor node metastasis stages. Knockdown of IFITM3 is effective in suppressing lung cancer cell proliferation, invasion, and migration while inducing cell cycle arrest and apoptosis.[46]
Our database search results indicate that hypermethylated genes and promotors in PD-L1 high-expressing tumors have strong oncogenic effects and, additionally, promote the creation of a proinflammatory and proangiogenic milieu in the tumor microenvironment, associated with an increased number of immune cells such as neutrophils, macrophages and lymphocytes, and regulation of active oxygen and calcium [47]. On the other hand, hypermethylated genes and promotors in the PD-L1 high-expression group have suppressor effects that appear ineffective in reducing tumor progression compared to this group’s simultaneously activated dominant oncogenic mechanisms. Hypomethylation of genes and promoters in PD-L1 high-expressing tumors reduces oncogenic and tumor suppressive effects but ultimately supports lung cancer development.
Pathobiological Mechanisms of Hypermethylated Genes and Promotors in PD-L1 Low-Expressing Cases
Our study found that tumors with low PD-L1 expression had hypermethylation in a single gene and promotor, SNORD114-14, a potent oncogene. The pathobiological mechanisms are already listed above.
Pathobiological Mechanisms of Hypomethylated Genes and Promotors in the PD-L1 Low-Expressing Group
Our methylation analyses found that MIR124-3, TRIM71, CAPS2, UBE2QL1, and GLIPR1L2 were hypomethylated genes and promoters in pulmonary adenocarcinomas with low expression of PD-L1.
MIR124-3 (MicroRNA124-3) is a microRNA that affects mRNA stability and translation. It plays a role in both breast and lung cancer [48] and was shown to suppress metastasis of lung cancer.[49]
TRIM71 (Tripartite Motif Containing 71, LIN41) is a gene that encodes an E3 ubiquitin-protein ligase, which plays a role in the G1-S phase transition of the cell cycle. TRIM7 negatively regulates the NF-kappa B signaling pathway in lung cancer by degrading p65. [50,51] The expression of TRIM7 is diminished in tumor tissues compared to adjacent normal tissues, and its level is negatively correlated with the clinical stage of lung cancer. In vitro, TRIM7 substantially inhibits the proliferation and migration of tumor cells and promotes cell apoptosis, but its effects are less effective in vivo.[52]
CAPS2 and GLIPR1L2 were previously discussed in the group due to hypomethylation in PD-L1 high-expressing carcinomas.
The UBE2QL1 gene (Ubiquitin Conjugating Enzyme E2 Q Family Like 1) encodes a ubiquitin-conjugating enzyme in lung cancer cells. It plays a crucial role in regulating the integrity of lysosomes and controlling the selective macroautophagy or autophagy of the entire organelle, also known as lysophagy.[53] Lysosomal membrane permeabilization or complete rupture of lysosomes can lead to stress conditions relevant to degenerative diseases, infections, and cancer. [54]
LINC0528, please refer to the top results for PD-L1 high expression.
Based on several studies, the NUMB gene (NUMB Endocytic Adaptor Protein) acts as a tumor suppressor in lung carcinoma.[55] In lung adenocarcinoma, high levels of NUMB can inhibit tumor growth, invasion, the Notch pathway, and epithelial-mesenchymal transition. However, in lung squamous cell carcinoma, NUMB may promote cell proliferation. The loss or mutation of NUMB is associated with poor prognosis, tumor progression, and resistance to chemotherapy in NSCLC patients.[56]
In tumors with low PD-L1 expression, hypomethylation of genes and promoters mainly reduces suppressor function, while one potent oncogene is present. Based on this observation, we concluded that the process of hypomethylation leads to the development of tumors by reducing the effectiveness of the mechanisms that suppress tumor growth.[57] This suggests that tumors with high levels of PD-L1 expression in the lungs and other primary tumor sites increase oncogenic mechanisms, making them more aggressive than tumors with low PD-L1 expression, which lose tumor-suppressing mechanisms.[47]
Hypermethylated Genes and Promotors with the Same Methylation Status Simultaneously in PD-L1 High and Low-Expressing Carcinomas
The methylation status of the following genes and promoters is similar in both PD-L1 groups and, therefore, does not correlate with PD-L1 expression.
SNORD114-14 showed the highest levels of methylation in both PD-L1 high and PD-L1 low-expressing lung cancers, with methylation of 75% and 90 % and p values of 0,04.
CAPS2 and GLIPR1L2 were hypomethylated in both PD-L1 high and low expression groups with methylation of 14% and 26% and p-values of 0,005 for CAPS2 and 19%, 21%, and 30% and p-values of 0,008 for GLIPR1L2.
5. Conclusions
PD-L1 high-expressing tumors show hypermethylated genes and promotors with strong oncogenic effects (SNORD114-14, DCAF4L2, S100A7L2, and SOD1P3) and hypermethylated genes and promotors with tumor suppressor effects (CELF2-AS1, LINCMD1). Especially LINC00528 helps to create a tumor microenvironment that supports cancer development, growth, and progression. Hypomethylated genes and promotors in this group lose their tumor-suppressing effects (IFITM3 and GLIPR1L2) and also have a reduction of oncogene activity (CAPS2).
PD-L1 low-expressing tumors show a single potent hypermethylated oncogene (SNORD114-4) and hypomethylated genes and promotors which lose their suppressor function (MIR124-3, TRIM71, GLIPR1L2, and NUMB) and additionally reduce the functionality of CAPS2, an oncogene. Besides, UBE2QL1 and LINC0528 form a microenvironment that supports cancer growth.
PD-L1 independent genes and promotors with similar results in both tumor groups with low and high levels of PD-L1 show hypermethylation of SNORD114-14 and hypomethylation of CAPS2 and GLIPR1L2.
Lung carcinomas exhibiting high and low PD-L1 expression demonstrate distinct methylation patterns, indicating different paths through various mechanisms by which PD-L1 high-expressing and PD-L1 low-expressing lung cancers develop. It appears that the development of oncogenic effects primarily drives tumors with high expression of PD-L1, and carcinomas with low expression of PD-L1 tend to develop tumors mainly by reducing suppressor mechanisms. If genes and promoters are hypermethylated, leading to simultaneous upregulation of suppressor and oncogenic effects, suppressors seem less effective than dominant oncogenic mechanisms. We concluded that activation of oncogenes is equivalent to the more aggressive behavior of tumors in the PD-L1 high group. Remarkably, the tumor microenvironment supports tumor growth in both groups.
Author Contributions
Conceptualization, G.H., T.F.J.K.; KS. methodology, G.H., T.F.J.K., B.A.S., software, G.H., T.F.J.K.; validation, G.H., T.F.J.K.,B.A.S., KS.; formal analysis, G.H., T.F.J.K. investigation, G.H., T.F.J.K.; resources, K.S.; data curation, G.H., T.F.J.K.; writing—original draft preparation, G.H.; writing—review and editing, G.H., K.S.; visualization, G.H., T.F.J.K.; supervision, G.H., and K.S.; project administration, K.S.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
All procedures performed in this study were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards and approved by the Institutional Review Board and Ethics Committee of the district of Salzburg (415-E/2509/2- 2019, 24 April 2019), all samples were anonymized before study inclusion (non-identifiable samples).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available from the corresponding author on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chi, S.A.; Yu, H.; Choi, Y.L.; Park, S.; Sun, J.M.; Lee, S.H.; Ahn, J.S.; Ahn, M.J.; Choi, D.H.; Kim, K.; et al. Trends in Survival Rates of Non-Small Cell Lung Cancer With Use of Molecular Testing and Targeted Therapy in Korea, 2010-2020. JAMA Netw Open 2023, 6, e232002. [CrossRef]
- Paci, E.; Puliti, D.; Lopes Pegna, A.; Carrozzi, L.; Picozzi, G.; Falaschi, F.; Pistelli, F.; Aquilini, F.; Ocello, C.; Zappa, M.; et al. Mortality, survival and incidence rates in the ITALUNG randomised lung cancer screening trial. Thorax 2017, 72, 825-831. [CrossRef]
- Hardtstock, F.; Myers, D.; Li, T.; Cizova, D.; Maywald, U.; Wilke, T.; Griesinger, F. Real-world treatment and survival of patients with advanced non-small cell lung Cancer: a German retrospective data analysis. BMC Cancer 2020, 20, 260. [CrossRef]
- Godoy, L.A.; Chen, J.; Ma, W.; Lally, J.; Toomey, K.A.; Rajappa, P.; Sheridan, R.; Mahajan, S.; Stollenwerk, N.; Phan, C.T.; et al. Emerging precision neoadjuvant systemic therapy for patients with resectable non-small cell lung cancer: current status and perspectives. Biomark Res 2023, 11, 7. [CrossRef]
- Tang, Q.; Chen, Y.; Li, X.; Long, S.; Shi, Y.; Yu, Y.; Wu, W.; Han, L.; Wang, S. The role of PD-1/PD-L1 and application of immune-checkpoint inhibitors in human cancers. Front Immunol 2022, 13, 964442. [CrossRef]
- Wojtukiewicz, M.Z.; Rek, M.M.; Karpowicz, K.; Gorska, M.; Politynska, B.; Wojtukiewicz, A.M.; Moniuszko, M.; Radziwon, P.; Tucker, S.C.; Honn, K.V. Inhibitors of immune checkpoints-PD-1, PD-L1, CTLA-4-new opportunities for cancer patients and a new challenge for internists and general practitioners. Cancer Metastasis Rev 2021, 40, 949-982. [CrossRef]
- Bruno, R.; Fontanini, G. Next Generation Sequencing for Gene Fusion Analysis in Lung Cancer: A Literature Review. Diagnostics (Basel) 2020, 10. [CrossRef]
- Czajka-Francuz, P.; Prendes, M.J.; Mankan, A.; Quintana, A.; Pabla, S.; Ramkissoon, S.; Jensen, T.J.; Peiro, S.; Severson, E.A.; Achyut, B.R.; et al. Mechanisms of immune modulation in the tumor microenvironment and implications for targeted therapy. Front Oncol 2023, 13, 1200646. [CrossRef]
- Rizvi, N.A.; Hellmann, M.D.; Snyder, A.; Kvistborg, P.; Makarov, V.; Havel, J.J.; Lee, W.; Yuan, J.; Wong, P.; Ho, T.S.; et al. Cancer immunology. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science 2015, 348, 124-128. [CrossRef]
- Ruschoff, J.; Schildhaus, H.U.; Ruschoff, J.H.; Johrens, K.; Bocker-Edmonston, T.; Dietmaier, W.; Blaker, H.; Baretton, G.; Horst, D.; Dietel, M.; et al. [Not Available]. Pathologie (Heidelb) 2023. [CrossRef]
- Schildhaus, H.U. [Predictive value of PD-L1 diagnostics]. Pathologe 2018, 39, 498-519. [CrossRef]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. AAPS J 2021, 23, 39. [CrossRef]
- Turchan, W.T.; Pitroda, S.P.; Weichselbaum, R.R. Radiotherapy and Immunotherapy Combinations in the Treatment of Patients with Metastatic Disease: Current Status and Future Focus. Clin Cancer Res 2021, 27, 5188-5194. [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J Clin 2019, 69, 7-34. [CrossRef]
- Romero-Garcia, S.; Prado-Garcia, H.; Carlos-Reyes, A. Role of DNA Methylation in the Resistance to Therapy in Solid Tumors. Front Oncol 2020, 10, 1152. [CrossRef]
- De Marchi, P.; Leal, L.F.; Duval da Silva, V.; da Silva, E.C.A.; Cordeiro de Lima, V.C.; Reis, R.M. PD-L1 expression by Tumor Proportion Score (TPS) and Combined Positive Score (CPS) are similar in non-small cell lung cancer (NSCLC). J Clin Pathol 2021, 74, 735-740. [CrossRef]
- Kaur, D.; Lee, S.M.; Goldberg, D.; Spix, N.J.; Hinoue, T.; Li, H.-T.; Dwaraka, V.B.; Smith, R.; Shen, H.; Liang, G.; et al. Comprehensive evaluation of the Infinium human MethylationEPIC v2 BeadChip. Epigenetics Communications 2023, 3, 6. [CrossRef]
- Bock, C. Analysing and interpreting DNA methylation data. Nat Rev Genet 2012, 13, 705-719. [CrossRef]
- Wang, Y.; Gorrie-Stone, T.J.; Grant, O.A.; Andrayas, A.D.; Zhai, X.; McDonald-Maier, K.D.; Schalkwyk, L.C. InterpolatedXY: a two-step strategy to normalize DNA methylation microarray data avoiding sex bias. Bioinformatics 2022, 38, 3950-3957. [CrossRef]
- Gardiner-Garden, M.; Frommer, M. CpG islands in vertebrate genomes. J Mol Biol 1987, 196, 261-282. [CrossRef]
- Smyth, G.K. limma: Linear Models for Microarray Data. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor, Gentleman, R., Carey, V.J., Huber, W., Irizarry, R.A., Dudoit, S., Eds.; Springer New York: New York, NY, 2005; pp. 397-420.
- Flanagan, J.M. Epigenome-wide association studies (EWAS): past, present, and future. Methods Mol Biol 2015, 1238, 51-63. [CrossRef]
- Campagna, M.P.; Xavier, A.; Lechner-Scott, J.; Maltby, V.; Scott, R.J.; Butzkueven, H.; Jokubaitis, V.G.; Lea, R.A. Epigenome-wide association studies: current knowledge, strategies and recommendations. Clin Epigenetics 2021, 13, 214. [CrossRef]
- Bhootra, S.; Jill, N.; Shanmugam, G.; Rakshit, S.; Sarkar, K. DNA methylation and cancer: transcriptional regulation, prognostic, and therapeutic perspective. Med Oncol 2023, 40, 71. [CrossRef]
- Lee, C.J.; Ahn, H.; Jeong, D.; Pak, M.; Moon, J.H.; Kim, S. Impact of mutations in DNA methylation modification genes on genome-wide methylation landscapes and downstream gene activations in pan-cancer. BMC Med Genomics 2020, 13, 27. [CrossRef]
- Frost, H.R. Analyzing cancer gene expression data through the lens of normal tissue-specificity. PLoS Comput Biol 2021, 17, e1009085. [CrossRef]
- Chen, Y.; Breeze, C.E.; Zhen, S.; Beck, S.; Teschendorff, A.E. Tissue-independent and tissue-specific patterns of DNA methylation alteration in cancer. Epigenetics Chromatin 2016, 9, 10. [CrossRef]
- Stahl, M.; Kohrman, N.; Gore, S.D.; Kim, T.K.; Zeidan, A.M.; Prebet, T. Epigenetics in Cancer: A Hematological Perspective. PLoS Genet 2016, 12, e1006193. [CrossRef]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Zhang, S.; Gong, Z.; Li, X.; Cao, K.; Deng, H.; He, Y.; et al. The role of microenvironment in tumor angiogenesis. J Exp Clin Cancer Res 2020, 39, 204. [CrossRef]
- Surace, A.E.A.; Hedrich, C.M. The Role of Epigenetics in Autoimmune/Inflammatory Disease. Front Immunol 2019, 10, 1525. [CrossRef]
- de Mendoza, A.; Nguyen, T.V.; Ford, E.; Poppe, D.; Buckberry, S.; Pflueger, J.; Grimmer, M.R.; Stolzenburg, S.; Bogdanovic, O.; Oshlack, A.; et al. Large-scale manipulation of promoter DNA methylation reveals context-specific transcriptional responses and stability. Genome Biol 2022, 23, 163. [CrossRef]
- Mourksi, N.E.; Morin, C.; Fenouil, T.; Diaz, J.J.; Marcel, V. snoRNAs Offer Novel Insight and Promising Perspectives for Lung Cancer Understanding and Management. Cells 2020, 9. [CrossRef]
- Han, P.; Liu, Q.; Xiang, J. Monitoring methylation-driven genes as prognostic biomarkers in patients with lung squamous cell cancer. Oncol Lett 2020, 19, 707-716. [CrossRef]
- Yeung, Y.T.; Fan, S.; Lu, B.; Yin, S.; Yang, S.; Nie, W.; Wang, M.; Zhou, L.; Li, T.; Li, X.; et al. CELF2 suppresses non-small cell lung carcinoma growth by inhibiting the PREX2-PTEN interaction. Carcinogenesis 2020, 41, 377-389. [CrossRef]
- Cuttano, R.; Colangelo, T.; Guarize, J.; Dama, E.; Cocomazzi, M.P.; Mazzarelli, F.; Melocchi, V.; Palumbo, O.; Marino, E.; Belloni, E.; et al. miRNome profiling of lung cancer metastases revealed a key role for miRNA-PD-L1 axis in the modulation of chemotherapy response. J Hematol Oncol 2022, 15, 178. [CrossRef]
- Guz, M.; Rivero-Muller, A.; Okon, E.; Stenzel-Bembenek, A.; Polberg, K.; Slomka, M.; Stepulak, A. MicroRNAs-role in lung cancer. Dis Markers 2014, 2014, 218169. [CrossRef]
- Jiang, J.; Lu, Y.; Zhang, F.; Huang, J.; Ren, X.L.; Zhang, R. The Emerging Roles of Long Noncoding RNAs as Hallmarks of Lung Cancer. Front Oncol 2021, 11, 761582. [CrossRef]
- Gong, S.; Xu, M.; Zhang, Y.; Shan, Y.; Zhang, H. The Prognostic Signature and Potential Target Genes of Six Long Non-coding RNA in Laryngeal Squamous Cell Carcinoma. Front Genet 2020, 11, 413. [CrossRef]
- Liu, X.L.; Zuo, R.; Ou, W.B. The hippo pathway provides novel insights into lung cancer and mesothelioma treatment. J Cancer Res Clin Oncol 2018, 144, 2097-2106. [CrossRef]
- Padilla, L.; Dakhel, S.; Adan, J.; Masa, M.; Martinez, J.M.; Roque, L.; Coll, T.; Hervas, R.; Calvis, C.; Llinas, L.; et al. S100A7: from mechanism to cancer therapy. Oncogene 2017, 36, 6749-6761. [CrossRef]
- Zhang, Y.; Lu, X.; Zhang, Y.; Zhao, D.; Gong, H.; Du, Y.; Sun, H. The Effect of Extracellular Superoxide Dismutase (SOD3) Gene in Lung Cancer. Front Oncol 2022, 12, 722646. [CrossRef]
- Xiao-Jie, L.; Ai-Mei, G.; Li-Juan, J.; Jiang, X. Pseudogene in cancer: real functions and promising signature. J Med Genet 2015, 52, 17-24. [CrossRef]
- Pang, B.; Wu, N.; Guan, R.; Pang, L.; Li, X.; Li, S.; Tang, L.; Guo, Y.; Chen, J.; Sun, D.; et al. Overexpression of RCC2 Enhances Cell Motility and Promotes Tumor Metastasis in Lung Adenocarcinoma by Inducing Epithelial-Mesenchymal Transition. Clin Cancer Res 2017, 23, 5598-5610. [CrossRef]
- Herath, S.; Sadeghi Rad, H.; Radfar, P.; Ladwa, R.; Warkiani, M.; O’Byrne, K.; Kulasinghe, A. The Role of Circulating Biomarkers in Lung Cancer. Front Oncol 2021, 11, 801269. [CrossRef]
- Sheng, X.; Bowen, N.; Wang, Z. GLI pathogenesis-related 1 functions as a tumor-suppressor in lung cancer. Mol Cancer 2016, 15, 25. [CrossRef]
- Zhang, D.; Wang, H.; He, H.; Niu, H.; Li, Y. Interferon induced transmembrane protein 3 regulates the growth and invasion of human lung adenocarcinoma. Thorac Cancer 2017, 8, 337-343. [CrossRef]
- Dong, P.; Xiong, Y.; Yue, J.; Hanley, S.J.B.; Watari, H. Tumor-Intrinsic PD-L1 Signaling in Cancer Initiation, Development and Treatment: Beyond Immune Evasion. Front Oncol 2018, 8, 386. [CrossRef]
- Wu, K.L.; Tsai, Y.M.; Lien, C.T.; Kuo, P.L.; Hung, A.J. The Roles of MicroRNA in Lung Cancer. Int J Mol Sci 2019, 20. [CrossRef]
- Zhu, Q.; Zhang, Y.; Li, M.; Zhang, Y.; Zhang, H.; Chen, J.; Liu, Z.; Yuan, P.; Yang, Z.; Wang, X. MiR-124-3p impedes the metastasis of non-small cell lung cancer via extracellular exosome transport and intracellular PI3K/AKT signaling. Biomark Res 2023, 11, 1. [CrossRef]
- Loedige, I.; Gaidatzis, D.; Sack, R.; Meister, G.; Filipowicz, W. The mammalian TRIM-NHL protein TRIM71/LIN-41 is a repressor of mRNA function. Nucleic Acids Res 2013, 41, 518-532. [CrossRef]
- Treiber, T.; Treiber, N.; Plessmann, U.; Harlander, S.; Daiss, J.L.; Eichner, N.; Lehmann, G.; Schall, K.; Urlaub, H.; Meister, G. A Compendium of RNA-Binding Proteins that Regulate MicroRNA Biogenesis. Mol Cell 2017, 66, 270-284 e213. [CrossRef]
- Jin, J.; Lu, Z.; Wang, X.; Liu, Y.; Han, T.; Wang, Y.; Wang, T.; Gan, M.; Xie, C.; Wang, J.; Yu, B. E3 ubiquitin ligase TRIM7 negatively regulates NF-kappa B signaling pathway by degrading p65 in lung cancer. Cell Signal 2020, 69, 109543. [CrossRef]
- Schuck, S. Microautophagy - distinct molecular mechanisms handle cargoes of many sizes. J Cell Sci 2020, 133. [CrossRef]
- Kravic, B.; Behrends, C.; Meyer, H. Regulation of lysosome integrity and lysophagy by the ubiquitin-conjugating enzyme UBE2QL1. Autophagy 2020, 16, 179-180. [CrossRef]
- Garcia-Heredia, J.M.; Verdugo Sivianes, E.M.; Lucena-Cacace, A.; Molina-Pinelo, S.; Carnero, A. Numb-like (NumbL) downregulation increases tumorigenicity, cancer stem cell-like properties and resistance to chemotherapy. Oncotarget 2016, 7, 63611-63628. [CrossRef]
- Haider, T.; Pandey, V.; Banjare, N.; Gupta, P.N.; Soni, V. Drug resistance in cancer: mechanisms and tackling strategies. Pharmacol Rep 2020, 72, 1125-1151. [CrossRef]
- Shen, X.; He, Z.; Li, H.; Yao, C.; Zhang, Y.; He, L.; Li, S.; Huang, J.; Guo, Z. Distinct functional patterns of gene promoter hypomethylation and hypermethylation in cancer genomes. PLoS One 2012, 7, e44822. [CrossRef]
Figure 1.
shows the TPS values of all probes.
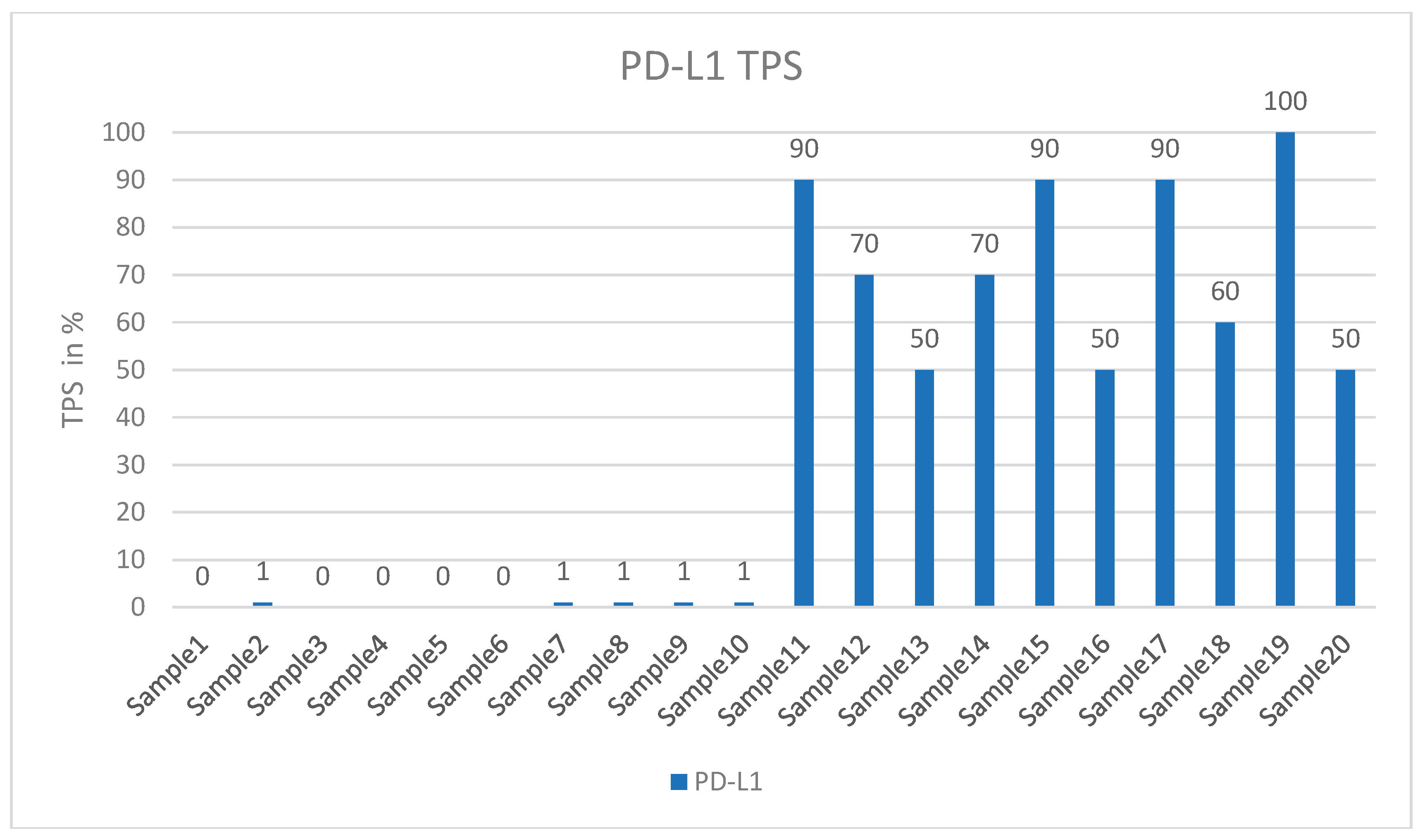

Table 1.
| PD-L1 Expression | Methylation Status | Gene | Methylation | P value | Chr |
| PD-L1 high | Highest methylation in gene | SNORD114-14 (, C/D Box 114-14). Small Nucleolar RNAs (SnoRNAs) | 0,9 | 0,04 | 14 |
| Highest methylation in promotor | DCAF4L2 (DDB1 Associated Factor 4 Like 2 ) | 0,82 | 0,02 | 8 | |
| CELF2-AS1 (CELF2 Antisense RNA 1) | 0,8 | 0.02 | 10 | ||
| LINCMD1 (Long Intergenic Non-Protein Coding RNA, Muscle Differentiation1, MIR133BHG) | 0,8 | 0,01 | 6 | ||
| Long Intergenic Non-Protein Coding RNA 528 (LINC00528) | 0,73 | 0,001 | 22 | ||
| Lowest methylation in gene | GLIPR1L2 (GLIPR1 Like 2) | 0,21 | 0,007 | 12 | |
| Lowest methylation in promotor | CAPS2 (Calcyphosine 2). | 0,14 | 0,005 | 12 | |
| GLIPR1L2 (GLI Pathogenesis Related 1 Like 2) | 0,19 | 0,008 | 12 | ||
| IFITM3 (Interferon Induced Transmembrane Protein 3 ) | 0,21 | 0,03 | 11 | ||
| PD-L1 low | Highest methylation in gene | SNORD114-14, (C/D Box 114-14) Small Nucleolar RNAs (SnoRNAs) |
0,9 | 0,04 | 14 |
| Highest methylation in promotor | SNORD114-14, (C/D Box 114-14) Small Nucleolar RNAs (SnoRNAs) |
0,75 | 0,04 | 14 | |
| Lowest methylation in gene | MIR124-3 (MicroRNA124-3) | 0,21 | 0,01 | 20 | |
| Lowest methylation in promotor | TRIM71 (Tripartite Motif Containing 71, LIN41) | 0,15 | 0,02 | 3 | |
| CAPS2 ( Calcyphosine 2). | 0,26 | 0;005 | 12 | ||
| UBE2QL1 (Ubiquitin Conjugating Enzyme E2 Q Family Like 1) | 0,3 | 0,007 | 5 | ||
| GLIPR1L2 (GLIPR1 Like 2) | 0,3 | 0,008 | 12 | ||
| PDL1 high/low | Difference* in gene methylation | S100A7L2 gene, also known as S100 Calcium Binding Protein A7 Like 2 | 0,74/0,57 Delta 17% |
0,002 | 1 |
| Difference* in promotor methylation | SOD1P3 (Superoxide Dismutase 1 Pseudogene 3), | 0,68/0,54 Delta 14% |
0,02 | 3 | |
| PDL1 low/high | Difference* in gene methylation | Long Intergenic Non-Protein Coding RNA 528 (LINC00528) | 0,54/0,73 Delta 19 % |
0,001 | 22 |
| Difference* in promotor methylation | NUMB gene, (NUMB Endocytic Adaptor Protein) | 0,31/0,51 Delta 20 % |
0,0005 | 14 |
* Average values of methylation (mean.mean.diff).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



