
Submitted:
29 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
The sexual reproduction of ascomycetes is controlled by the mating-type(MAT) locus. (Pseudo)homothallic reproduction has been hypothesized based on genetic data from Hirsutella sinensis (Genotype #1 of Ophiocordyceps sinensis). In this study, the differential occurrence of the mating-type genes in the MAT1-1 and MAT1-2 idiomorphs, which constitute the genetic control of O. sinensis reproduction, was analyzed in 237 strains and 5 genomes of H. sinensis. The mating-type genes were differentially transcribed in H. sinensis Strain L0106, and the MAT1-2-1 transcript in H. sinensis Strain 1229 was alternatively spliced, with an unspliced intron I containing stop codons; these findings constitute O. sinensis reproduction controls at the transcriptional and coupled transcriptional-translational levels. The occurrence of the mating-type genes and a-/α-pheromone receptor genes differed among the 5 H. sinensis genomes. The differential occurrence and transcription of these genes are inconsistent with self-fertilization under (pseudo)homothallism but suggest self-sterility of H. sinensis, which therefore needs a mating partner to achieve sexual outcrossing under heterothallism or hybridization. Consistent occurrence and transcription of the mating-type genes of both the MAT1-1 and MAT1-2 idiomorphs have been reported in natural and cultivated Cordyceps sinensis, but these findings are insufficient to determine the reproduction mode(s) because of the co-occurrence of >90 fungal species and multiple genomically independent genotypes of O. sinensis. The mutant MAT1-1-1 and α-pheromone receptor transcripts in natural C. sinensis result in the N-terminus or middle-truncated proteins, with changes in hydrophobicity and secondary structures of the proteins, suggesting heterogeneous fungal source(s) of the proteins and hybridization reproduction.
Keywords:
Differential transcription mating-type genes
; mutant MAT1-1-1 protein
; mutant α-pheromone receptor protein
; genotypes of Ophiocordyceps sinensis
; natural Cordyceps sinensis
; transcriptional regulation
; homothallism
; pseudohomothallism
; physiological heterothallism
; hybridization
Introduction
The sexual reproductive behavior of ascomycetes is controlled by transcription factors encoded at the mating-type (MAT) locus [Debuchy & Turgeon 2006; Jones & Bennett 2011; Zheng & Wang 2013; Wilson et al. 2015]. Hu et al. [2013] detected the MAT1-1-1 and MAT1-2-1 genes of the MAT1-1 and MAT1-2 idiomorphs in the genome assembly ANOV00000000 of Hirsutella sinensis Strain Co18 and hypothesized that H. sinensis (GC-biased Genotype #1 of Ophiocordyceps sinensis) experiences self-fertilization under homothallism. The MAT1-1-1 gene encodes a protein possessing a conserved α-box domain, and the MAT1-2-1 gene encodes a protein possessing a high-mobility-group (HMG) box domain [Zheng & Wang 2013; Wilson et al. 2015]. Bushley et al. [2013] described multicellular heterokaryotic structures of natural Cordyceps sinensis hyphae and ascospores containing mononucleated, binucleated, trinucleated, and tetranucleated cells; in addition, they detected the MAT1-1-1, MAT1-1-2 and MAT1-1-3 genes of the MAT1-1 idiomorph and the MAT1-2-1 gene of the MAT1-2 idiomorph, which are located >4 kb apart in the genome of H. sinensis Strain 1229. Based on these results, they hypothesized that H. sinensis underwent pseudohomothallism. However, Zhang et al. [2011] and Zhang and Zhang [2015] reported differential occurrence of the MAT1-1-1 and MAT1-2-1 genes in various H. sinensis strains or isolates and hypothesized that O. sinensis underwent facultative hybridization. These hypotheses regarding the sexual reproductive behavior of O. sinensis were proposed based on the genetic information of H. sinensis without considering the entire expression process of the H. sinensis mating-type genes, including genetic, epigenetic, transcriptional, posttranscriptional, translational and posttranslational modifications, as well as the activation and degradation of mating-type proteins.
Natural C. sinensis is one of most valued therapeutic agents in traditional Chinese medicine and has a rich history of clinical use in health maintenance, disease amelioration, postdisease and postsurgery recovery and antiaging therapy [Zhu et al. 1998a, 1998b, 2011]. The Chinese Pharmacopoeia defines natural C. sinensis as an insect-fungal complex containing the O. sinensis fruiting body and the remains of a Hepialidae moth larva (an intact, thick larval body wall with numerous bristles, intact larval intestine and head tissues, and fragments of other larval tissues) [Ren et al. 2013; Li et al. 2016b, 2022b; Zhu & Li 2017]. Studies of natural C. sinensis conducted using various histological, mycological and molecular approaches have demonstrated its multicellular heterokaryotic structure and genetic heterogeneity, as revealed by the detection of at least 17 genomically independent genotypes of O. sinensis [Kinjo & Zang 2001; Stensrud et al. 2007; Xiao et al. 2009; Zhang et al. 2009; Zhu et al. 2010, 2012; Chen et al. 2011; Li et al. 2013, 2016b, 2022b, 2023a, 2023b; Mao et al. 2013] and >90 fungal species spanning at least 37 genera and larval genes [Qiu et al. 1987; Li et al. 1988, 2016b, 2019, 2022b; Dai et al. 1989; Engh 1999; Jiang & Yao 2003; Chen et al. 2004; Stensrud et al. 2005; Leung et al. 2006; Zhu et al. 2007, 2023b; Yang et al. 2008; Zhang et al. 2010; Barseghyan et al. 2011; Xia et al. 2015; Wang et al. 2020]. The expression of mating-type genes during the sexual life of natural C. sinensis is much more complex than that in pure fungal cultures and is mutually and/or antagonistically disrupted by the expression of the metagenomes of multiple cocolonized fungi and by larval host innate immunity and acquired immunological responses during O. sinensis fungal infection and proliferation [Meng et al. 2015; Lu et al. 2016; Wang et al. 2016, 2018; Li et al. 2019, 2022b]. It has been suggested that larval tissues are not just culture medium that passively provide nutrients for fungal growth, similar to the medium used in in vitro culture or fermentation [Lu et al. 2016; Wang et al. 2018; Li et al. 2022b]. Notably, the Latin name Cordyceps sinensis has been indiscriminately used since the 1840s for both the teleomorph/holomorph of C. sinensis fungus and the wild insect-fungal complex, and the fungus was renamed O. sinensis in 2007 [Sung et al. 2007; Ren et al. 2013; Zhang et al. 2012; Zhu & Wu 2015; Li et al. 2016b, 2022b; Zhu & Li 2017]. In this paper, we refer to the fungus/fungi as O. sinensis and continue the customary use of the name C. sinensis to refer to the wild or cultivated insect-fungi complex, although this practice will likely be replaced by the discriminative use of exclusive Latin names in the future.
Many studies on natural C. sinensis and O. sinensis fungi have focused primarily on H. sinensis, which has been postulated to be the sole anamorph of O. sinensis [Wei et al. 2006]. Following this sole anamorph hypothesis, H. sinensis Strain EFCC7287, Genotype #1 of O. sinensis, was selected for use as a reference species for the renaming of Cordyceps sinensis to Ophiocordyceps sinensis [Sung et al. 2007]. However, this renaming did not involve the natural insect-fungal complex and did not cover the remaining 16 genotypes of O. sinensis fungi, for which pure cultures have not been available for taxonomy and nomenclature multigene studies or for genomics, transcriptomics, proteomics, natural chemistry and pharmacology studies [Engh 1999; Kinjo & Zang 2001; Stensrud et al. 2005, 2007; Xiao et al. 2009; Zhang et al. 2009, 2012; Zhu et al. 2010, 2012, 2018; Chen et al. 2011; Gao et al. 2011, 2012; Yao et al. 2011; Li et al. 2013, 2016b, 2016c, 2020, 2022b, 2023a, 2023b]. Numerous studies have reported that the sequences of O. sinensis Genotypes #2‒17, regardless of whether they are GC or AT biased, are not found in the genome assemblies ANOV00000000, JAAVMX000000000, LKHE00000000, LWBQ00000000, and NGJJ00000000 of the GC-biased Genotype #1 H. sinensis Strains Co18, IOZ07, 1229, ZJB12195, and CC1406-203, respectively [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020], providing evidence of the genomic independence of the 17 O. sinensis genotypes, which are interindividual O. sinensis fungi [Stensrud et al. 2007; Xiao et al. 2009; Zhang et al. 2009; Zhu et al. 2010, 2012; Chen et al. 2011; Li et al. 2016b, 2016c, 2020, 2021, 2022a, 2022b, 2023a, 2023b]. In this paper, we continue using the anamorphic name H. sinensis for Genotype #1 of O. sinensis, although a group of mycologists [Zhang et al. 2013] have improperly implemented the “One Fungus=One Name” nomenclature rule of the International Mycological Association while disregarding the presence of multiple genomically independent genotypes of O. sinensis fungi and inappropriately replacing the anamorphic name H. sinensis with the teleomorphic name O. sinensis [Ren et al. 2013; Zhu & Wu 2015; Zhu & Li 2017; Zhu et al. 2018; Li et al. 2022b].
The lifecycle of C. sinensis includes asexual and sexual growth stages during its development and maturation. O. sinensis infection of the larvae of the Hepialidae family initiates the lifecycle of natural C. sinensis, which includes several developmental phases: (1) formation of a stromal primordium; (2) immature C. sinensis with a short stroma (1‒2 cm) (O. sinensis grows asexually in the first two phases); (3) maturing C. sinensis in the transition from asexual to sexual growth with an approximately 3‒5 cm stroma without the formation of an expanded fertile portion close to the tip of the stroma; and (4) mature C. sinensis that grows sexually with a long stroma (usually >5 cm) with an expanded fertile portion close to the stromal tip, densely covering numerous ascocarps before and after ascospore ejection (Figure 1).
C. sinensis maturation is associated with dramatic changes in the mycobiota, metagenome, metatranscriptomic and proteomic expression, chemical constituent fingerprint and pharmacological functions [Zhu et al. 2007, 2010, 2012; Yang et al. 2008; Liang et al. 2010; Gao et al. 2011, 2012; Dong et al. 2014; Yao et al. 2014; Zhu & Wu 2015; Li et al. 2016b, 2016c, 2020, 2022b, 2023b; Zhu & LI 2017]. Li et al. [2023a, 2023b] reported naturally and semiejected C. sinensis ascospores associated with differential co-occurrence of several GC- and AT-biased genotypes of O. sinensis. The biomasses/abundances of GC- and AT-biased genotypes of O. sinensis change dynamically in an asynchronous, disproportional manner in the caterpillar body, stroma, stromal fertile portion (with ascocarps) and ascospores of natural C. sinensis during maturation [Xiao et al. 2009; Zhu et al. 2010, 2012; Gao et al. 2011, 2012; Yao et al. 2011; Zhu & Wu 2015; Li et al 2016b, 2023a, 2023b; Zhu & Li 2017]. Thus, it is essential to identify the maturation stages of natural C. sinensis specimens during the examination of mating-type gene expression.
Multiple genetic, genomic and transcriptomic sequences of numerous H. sinensis strains and natural C. sinensis specimens are available in the GenBank database, which enables further examination of the hypotheses regarding the sexual reproduction strategy of O. sinensis at the genome and transcriptome levels. This paper discusses the genetic occurrence of mating-type genes in 237 H. sinensis strains, the transcription of mating-type genes and pheromone receptor genes involved in O. sinensis sexual reproduction in H. sinensis and natural C. sinensis, and other features related to the potential mating process of O. sinensis.
Materials and Methods
Gene and Genome Sequences of H. sinensis Strains and Natural C. sinensis
Table 1 lists 237 H. sinensis strains that were used to obtain mating-type gene sequences, which are available in GenBank and the original papers [Zhang et al. 2009, 2011; Bushley et al. 2013; Hu et al. 2013; Liu et al. 2015, 2020; Zhang & Zhang 2015; Li et al. 2016a, 2021, 2022a; Jin et al. 2020; Shu et al. 2020]. Five sets of genome assemblies, ANOV00000000, JAAVMX000000000, LKHE00000000, LWBQ00000000, and NGJJ00000000, of H. sinensis Strains Co18, IOZ07, 1229, ZJB12195 and CC1406-20395, respectively, are available in GenBank [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020].
Genomic DNA from H. sinensis Strain Co18 was sequenced with the Roche 454 GS FLX system (Illumina HiSeq: 454), and the shotgun sequences were assembled using SOAPdenovo v.1.05 and Newbler v.2.3 [Hu et al. 2013]. Genomic DNA from Strain 1229 was sequenced with Illumina HiSeq sequencing technology, and the shotgun sequences were assembled using ABySS v.1.2.3 [Li et al. 2016a]. Genomic DNA from Strain ZJB12195 was sequenced with Illumina sequencing technology (HiSeq 2000 Sequencing System), and the shotgun sequences were assembled using SOAPdenovo v.2.0 [Jin et al. 2020]. Genomic DNA from Strain CC1406-203 was sequenced with continuous long reads (CLRs) following the Hierarchical Genome Assembly Process (HGAP) workflow (PacBioDevNet; Pacific Biosciences) and assembled with CA software (v.7.0) and the PacBio Rs_PreAssembler.1 module [Liu et al. 2020]. Genomic DNA from Strain IOZ07 was sequenced with PacBio Sequel sequencing technology, and the shotgun sequences were assembled using Canu v.1.7 [Shu et al. 2020].
Transcriptome and Metatranscriptome Assemblies and Transcripts of Mating-Type Genes of H. sinensis Strains and Natural C. sinensis
The transcriptome assembly GCQL00000000 is accessible in GenBank and was derived from fermented mycelia of the H. sinensis Strain L0106 [Liu et al. 2015]. The mycelia were collected for total RNA extraction from cultures grown for 3, 6 and 9 days. Total RNA (20 mg per sample) was subjected to mRNA purification, and total mRNA was used to construct a cDNA library and sequenced using Illumina HiSeq sequencing technology. The shotgun nucleotide sequences were assembled using SOAPdenovo v.2.0.
Natural C. sinensis specimens (unknown maturational status) were collected from Kangding County, Sichuan Province, China [Xiang et al. 2015]. Total RNA from this specimen was sequenced using 454 technology. Sequences longer than 50 bp from the 454 reads were assembled using GS De Novo Assembler software v 2.6 or Newbler 2.6 (454 Life Sciences Corporation, USA). The shotgun sequences were assembled as a metatranscriptome assembly under GenBank accession number GAGW00000000 using Newbler v.2.3 and 2.6.
Another metatranscriptome assembly was derived from specimens of mature natural C. sinensis that were collected from Deqin, Yunnan Province, China (cf. the Appendix of [Xia et al. 2017]). Total RNA was extracted from the fruiting bodies to construct a cDNA library, which was sequenced using the Illumina HiSeq2000 platform. The raw reads were assembled using Trinity (version r20140717) [Grabherr et al. 2011]. The sequencing reads were deposited in GenBank under BioProject #PRJNA382001. According to the Appendix of [Xia et al. 2017], the metatranscriptome assembly sequences were uploaded to the depository database, www.plantkingdomgdb.com/Ophiocordyceps_sinensis/data/cds/Ophiocordyceps_sinensis_CDS.fas, which is currently inaccessible, but a previously downloaded cDNA file was used for the metatranscriptomic analysis.
Sequence Alignment Analysis
All gene, genome, transcript, transcriptome and metatranscriptome sequences and other PCR-amplified DNA sequences of the mating-type and pheromone receptor genes of H. sinensis and natural C. sinensis were reanalyzed using the MegaBlast, discontinuous MegaBlast, Blastp, Tblastn, or blastn suite-SRA programs provided by GenBank (https://blast.ncbi.nlm.nih.gov/) to compare and align nucleotide and amino acid sequences.
Amino Acid Property and Scale Analysis
The hydrophobicity/hydrophilicity of the mating-type and pheromone receptor proteins of H. sinensis and natural C. sinensis were characterized based on the chemical-physical properties of the component amino acids using Peptide 2.0 (https://www.peptide2.com/N_peptide_hydrophobicity_hydrophilicity.php). The component amino acids were scaled (Table 2) and plotted at a window size of 21 amino acid residues for the α-Helix, β-Sheet, β-Turn and Coil of the proteins using the linear weight variation model of the ExPASy PlotScale algorithm (https://web.expasy.org/protscale/) provided by the SIB Swiss Institute of Bioinformatics [Chou & Fasman 1978; Deleage & Roux 1987; Gasteiger et al. 2005]. The amino acid hydrophobicity properties and the waveforms of PlotScale plots for the proteins were compared to explore potential alterations in the folding and secondary structures of the mating-type and pheromone receptor proteins.
Results
Differential Occurrence of Mating-Type Genes in H. sinensis Strains
A search of the GenBank database and the literature revealed intraspecific variations in 237 H. sinensis strains that reportedly contain the MAT1-1-1 and MAT1-2-1 genes (Table 1): 22 strains contain only the MAT1-1-1 gene, 66 strains harbor only the MAT1-2-1 gene, and 149 strains harbor both genes [Zhang et al. 2011, 2013; Bushley et al. 2013; Hu et al. 2013; Zhang & Zhang 2015; Li et al. 2016a, 2021, 2022a; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020]. These findings provide evidence of intraspecific variations in H. sinensis strains [Li et al. 2022a].
Bushley et al. [2013] identified 3 mating-type genes of the MAT1-1 idiomorph in the genome sequence KC437356 of H. sinensis Strain 1229: the MAT1-1-1 gene (6,530→7,748), MAT1-1-2 gene (4,683→6,183) and MAT1-1-3 gene (3,730→4,432). The sequences of these mating-type genes from KC437356 show 99.9–100% homology with the segment sequences of the genome assemblies LKHE01001116 (3,691←4,909; 5,374←6,874; 7,125←7,827) of Strain 1229; ANOV01017390 (302←1,519) and ANOV01017391 (276←1,776; 2,027←2,729) of Strain Co18; and JAAVMX010000001 (6,698,911→6,700,129; 6,696,939→6,698,439; 6,695,986→6,696,688) of Strain IOZ07 (Table 3) [Bushley et al. 2013; Hu et al. 2013; Li et al. 2016a; Shu et al. 2020]. Note that the arrows “→” and “←” indicate sequences in the sense and antisense chains of the genomes, respectively. However, the mating-type genes of the MAT1-1 idiomorph are absent from the genome assemblies NGJJ00000000 and LWBQ00000000 of H. sinensis Strains CC1406-203 and ZJB12195, respectively [Jin et al. 2020; Liu et al. 2020]. A BLAST search of the GenBank database identified the MAT1-1-1 gene in 127 H. sinensis strains with 98.0‒100% homology among the sequences but did not find any sequences of the MAT1-1-2 or MAT1-1-3 genes in these strains [Zhang et al. 2009, 2013; Bushley et al. 2013; Hu et al. 2013; Li et al. 2021, 2022b].
The MAT1-2-1 sequence JQ325153 of H. sinensis Strain GS09_121 [Zhang et al. 2014] exhibits 99.7–99.9% homology with the segment sequences of the genome assemblies LKHE01001605 (13,851←14,723), ANOV01000063 (9,319→10,191), NGJJ01000619 (23,021←23,893) and LWBQ01000021 (238,864←239,736) of H. sinensis Strains 1229, Co18, CC1406-203 and ZJB12195, respectively (Table 3) [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Liu et al. 2020]. However, the MAT1-2-1 gene is absent in the genome assembly JAAVMX000000000 of Strain IOZ07 [Shu et al. 2020]. Further searches of the GenBank database identified the MAT1-2-1 gene sequence in 183 H. sinensis strains with 97.3–100% homology [Zhang et al. 2009, 2011, 2013, 2014; Bushley et al. 2013; Hu et al. 2013; Zhang & Zhang 2015; Li et al. 2021].
Bushley et al. [2013] found the presence of monokaryotic, bikaryotic, trikaryotic and tetrakaryotic structures in multicellular hyphae and ascospores and showed that the MAT1-1 and MAT1-2 idiomorphs are not closely linked because the MAT1-1-1 and MAT1-2-1 genes are located more than 4 kb apart in the H. sinensis genome. They then proposed that H. sinensis undergoes pseudohomothallism. The MAT1-1-1 and MAT1-2-1 genes are located on contig_17390 (302←1,519) of ANOV01017390 and contig_63 (9,319→10,191) of ANOV01000063 in the genome of Strain Co18, and scaffold_1116 (3,691←4,909) of LKHE01001116 and scaffold_1605 (13,851←14,723) of LKHE01001605 in the genome of Strain 1229 [Hu et al. 2013; Li et al. 2016a]. The MAT1-1-1 gene is present in the genome assembly JAAVMX010000001 of Strain IOZ07, but the MAT1-2-1 gene is absent [Shu et al. 2020]. The MAT1-2-1 gene is present in the genome assemblies NGJJ01000619 and LWBQ01000021 of Strains CC1406-203 and ZJB12195, respectively, but the MAT1-1-1 gene is absent [Jin et al. 2020; Liu et al. 2020].
Zhang et al. [2011] cloned the MAT1-2-1 gene but not the mating-type genes within the MAT1-1 idiomorph in H. sinensis Strains CS2 and SCK05-4-3. The differential occurrence of mating-type genes was confirmed by genomic evidence in the H. sinensis strains CC1406-203, IOZ07 and ZJB12195 (Table 3) [Jin et al. 2020; Liu et al. 2020; Shu et al. 2020]. It is believed that the homothallic reproduction of ascomycetes requires the combined expression of the mating-type genes of both MAT1-1 and MAT1-2 idiomorphs within a single fungal cell and that the expression of only one of the MAT1-1-1 and MAT1-2-1 genes might reciprocally require the expression of complementary mating-type genes from another fungus of the same species for heterothallic outcrossing or from a different species for hybridization [Bushley et al. 2013; Zheng & Wang 2013; Wilson et al. 2015; Zhang et al. 2018]. Although the homothallism and pseudohomothallism hypotheses were proposed for H. sinensis based on the occurrence of the mating-type genes of both MAT1-1 and MAT1-2 idiomorphs in H. sinensis Strains Co18 and 1229 [Bushley et al. 2013; Hu et al. 2013], Zhang and Zhang [2015] reported the differential occurrence of mating-type genes of the MAT1-1 and/or MAT1-2 idiomorphs in 125 wild-type isolates of natural C. sinensis collected from different production areas in geographically remote locations and hypothesized that H. sinensis undergoes facultative hybridization.
Notably, Zhang and Zhang [2015] reported 4.9% and 6.1% allelic variation (93.9% and 95.1% similarity) in the full-length sequences of the MAT1-1-1 and MAT1-2-1 genes, respectively, and 4.7% and 5.7% variation in the exon sequences of numerous C. sinensis isolates. These variations might disrupt the translation of the coding sequences of the genes. However, a BLAST search of the GenBank database revealed much less variation (97.3–100% similarity) among 127 sequences of the MAT1-1-1 gene and 183 sequences of the MAT1-2-1 gene of H. sinensis strains and C. sinensis isolates, suggesting that Zhang and Zhang [2015] may not have uploaded all analyzed mating-type gene sequences to GenBank.
Differential Transcription of Mating-Type Genes in the Transcriptome of H. sinensis Strains
In addition to the mating-type genes in H. sinensis, transcription of the mating-type genes of both the MAT1-1 and MAT1-2 idiomorphs and the production and full activation of the encoded proteins within a single cell are needed to accomplish the homothallic mating process and to control the development and maturation of fruiting bodies, ascocarps and ascospores [Turgeon & Yoder 2000; Debuchy & Turgeo 2006; Jones & Bennett 2011; Zheng & Wang 2013; Wilson et al. 2015]. The sequences of the MAT1-1-1, MAT1-1-2 and MAT1-1-3 genes of H. sinensis are absent in the transcriptome assembly GCQL00000000 of H. sinensis Strain L0106 (Table 3) [Liu et al. 2015]. Zhang et al. [2011] found no transcripts of the mating-type genes within the MAT1-1 idiomorph in H. sinensis strains and wild-type C. sinensis isolates. However, Bushley et al. [2013] obtained the transcript cDNA of the MAT1-1-1 gene with the splicing of two introns in the mycelia of Strain 1229. Therefore, it is apparent that the genes of the MAT1-1 idiomorph are differentially transcribed in H. sinensis Strains 1229, CS2, L0106, and SCK05-4-3 and many other strains in which the genes of the MAT1-1 idiomorph are absent in their genomes (cf. Table 1).
The MAT1-2-1 gene JQ325153 of H. sinensis GS09_121 contains two introns: “Intron Phase-Two” for intron I (52 nucleotides [nt]; 187→238 of JQ325153) and “Intron Phase-One” for intron II (55 nt; 535→589 of JQ325153). Both intron phases disrupt the open reading frame and codons (Figure 2) [Zhang et al. 2011, 2014; Bushley et al. 2013; Zhang & Zhang 2015; Li et al. 2021]. Figure 2 also shows 3 other MAT1-2-1 genes (FJ654187, JQ325237, and KM197536) from H. sinensis Strains CS2, XZ12_16, and XZ12_16, respectively, which were 97.7–99.5% similar to the sequence of JQ325153. A single T-to-C transition downstream of the third stop codon was present in intron I in the sequences of JQ325237 and KM197536 and apparently had no impact on the three stop codons in intron I.
The coding sequences of 3 exons (388←671; 672←967; 968←1,153) of the MAT1-2-1 gene JQ325153 exhibit 99.7‒100% homology with the transcriptome assembly GCQL01020543 of H. sinensis Strain L0106 [Zhang et al. 2014; Liu et al. 2015]. Xia et al. [2017] confirmed that the MAT1-2-1 transcript sequence (OSIN7649) with the splicing of both introns I and II in the metatranscriptome assembly of natural C. sinensis (further discussion below). Bushley et al. [2013] reported that “comparison of DNA and cDNA sequences of MAT1-2-1 revealed only one spliced intron of 55 bp” in H. sinensis Strain 1229, which “did not result from contamination of genomic DNA” and illustrated the alternatively spliced MAT1-2-1 cDNA with unspliced intron I and spliced intron II. Figure 2 herein shows the MAT1-2-1 sequences, which possess 3 stop codons, “TAA” or “TGA”, within intron I (the first, third and 13th triplets), leading to translational arrest of the MAT1-2-1 transcript with unspliced intron I.
To rule out the possibility that gene mutations within MAT1-2-1 intron I occur to avoid translational interruption due to the presence of stop codons within intron I, a BLAST search of the GenBank database was performed. This search revealed that the intron I sequence of the MAT1-2-1 gene is 100% identical to the MAT1-2-1 gene segments of the genome assemblies LKHE01001605, NGJJ01000619, ANOV01000063 and LWBQ01000021 of Strains 1229, CC1406-203, Co18 and ZJB12195, respectively, and to the DNA sequences of more than 100 H. sinensis strains. A five-base insertion occurs in intron I of the MAT1-2-1 genes FJ654204 and FJ654205 in two H. sinensis isolates, XZ-LZ07-H1 and XZ-LZ07-H2, obtained from the Nyingchi District in Tibet, which was postulated to be the center of origin of the species H. sinensis [Zhang et al. 2009]. The insertion is located downstream of the first stop codon “TAA”. Thus, the alternatively spliced MAT1-2-1 gene transcript with unspliced intron I might represent an additional mechanism of the coupled transcriptional-translational regulation of the mating process of H. sinensis Strain 1229 [Bushley et al. 2013], distinct from the gene silencing used for transcriptional control (Table 3), although the full expression process (genetic, epigenetic, transcriptional, posttranscriptional, translational, and posttranslational modifications; protein activation-degradation; etc.) of the H. sinensis MAT1-2-1 gene needs to be thoroughly examined in future reproductive physiology studies.
The above transcriptome analysis of H. sinensis Strains 1229 and L0106 [Bushley et al. 2013; Liu et al. 2015] revealed the transcriptional and coupled transcriptional-translational controls of mating-type gene expression in H. sinensis. These results are inconsistent with the hypotheses of self-fertilization under homothallism and pseudohomothallism for H. sinensis proposed by Bushley et al. [2013] and Hu et al. [2013]. Instead, these findings suggest that O. sinensis requires sexual partners from monoecious or dioecious H. sinensis for physiological heterothallic reproduction or heterospecific fungal species to ensure hybridization if the species are able to overcome interspecific reproductive isolation.
The available evidence regarding mating-type gene transcription might explain why previous efforts to cultivate pure H. sinensis in research-oriented academic settings to induce the production of fruiting bodies and ascospores have consistently failed [Holliday & Cleaver 2008; Stone 2010; Hu et al. 2013; Zhang et al. 2013] and why a successful inoculation-cultivation project in a product-oriented industrial setting presented a species contradiction between the GC-biased Genotype #1 H. sinensis strains used as anamorphic inoculants and the teleomorphic AT-biased Genotype #4 of O. sinensis in the fruiting body of cultivated C. sinensis, as reported by Wei et al. [2016].
Differential Occurrence of Mating-Type Genes in Natural and Cultivated C. sinensis Insect-Fungi Complexes
The above sections discuss the differential occurrence and transcription of mating-type genes in pure H. sinensis strains (GC-biased Genotype #1 of O. sinensis) and wild-type C. sinensis isolates. Mycobiota studies have demonstrated the differential coexistence of >90 cocolonized fungi belonging to at least 37 fungal genera in the stroma and caterpillar body of natural C. sinensis [Zhang et al. 2010; Xia et al. 2015], which has been confirmed by metagenomic and metatranscriptomic studies and by the identification of >100 mitogenomic transcript repeats in natural and cultivated C. sinensis [Xiang et al. 2014; Meng et al. 2015; Xia et al. 2017; Wang et al. 2018; Zhang et al. 2018; Zhong et al. 2018; Li et al. 2019; Zhao et al. 2020; Yang et al. 2021]. Many studies have reported differential co-occurrence of several GC- and AT-biased genotypes of O. sinensis in different compartments (the stroma, caterpillar body, stromal fertile portion densely covered with numerous ascocarps, and ascospores) of natural C. sinensis, the sequences of which reside in the independent fungal genomes of different O. sinensis fungi [Engh 1999; Kinjo & Zang 2001; Chen et al. 2011; Stensrud et al. 2005, 2007; Xiao et al. 2009; Zhang et al. 2009; Zhu et al. 2010, 2012, 2018, 2019; Li et al. 2013; 2016b, 2016c, 2020, 2022b, 2023a, 2023b; Mao et al. 2013; Wei et al. 2016; Zhu & Li 2017]. Notably, the indiscriminate use of the Latin name Ophiocordyceps sinensis for multiple genotypes of O. sinensis fungi and natural and cultivated insect-fungal complexes has prompted scientists to carefully distinguish the study materials when composing and reading research papers/reports to ensure correct wording and understanding [Li et al. 2016b, 2020, 2022a, 2022b, 2023a, 2023b; Zhu & Li 2017].
The mating-type genes of both the MAT1-1 and MAT1-2 idiomorphs reportedly exist in natural and cultivated C. sinensis, for instance, (1) in the metagenome of mature natural C. sinensis [Xia et al. 2017]; (2) in the compartments of natural C. sinensis at different developmental stages [Zhong et al. 2018]; (3) in cultivated C. sinensis at different developmental phases [Li et al. 2019]; and (4) in the early-developed stroma and caterpillar body of natural C. sinensis with very low read count values and in 31 other C. sinensis specimens [Zhao et al. 2020]. However, the detection of the mating-type genes of both the MAT1-1 and MAT1-2 idiomorphs reported in these studies might not correctly represent the genetic-based capability for self-fertilization of a single H. sinensis cell due to the differential coexistence of multiple cocolonized fungi and multiple genotypes of O. sinensis in the compartments of natural and cultivated C. sinensis [Engh 1999; Kinjo & Zang 2001; Xiao et al. 2009; Zhang et al. 2009; Zhu et al. 2010, 2012; Gao et al. 2011, 2012; Chen et al. 2011; Yao et al. 2011; Li et al. 2013, 2016b, 2020, 2022, 2023a, 2023b; Mao et al. 2013; Zhang & Zhang 2015; Wei et al. 2016; Zhu & Li 2017; Zhong et al. 2018; Zhao et al. 2020; Yang et al. 2021]. Hence, the possibility of physiological heterothallic outcrossing or hybridization should not be neglected, particularly because of the species contradiction between the anamorphic inoculants of GC-biased H. sinensis strains used in industrial cultivation projects and the sole teleomorph of AT-biased Genotype #4 in cultivated C. sinensis (Wei et al. 2016).
Differential Transcription of Mating-Type Genes in Natural and Cultivated C. sinensis Insect-Fungi Complexes
Transcription of Mating-Type Genes in the Metatranscriptome Assemblies of Natural C. sinensis
Xiang et al. [2014] reported a metatranscriptome project with natural C. sinensis specimens (unknown maturation stage) collected from Kangding County of Sichuan Province, China. The assembled metatranscriptome segment 297←1,129 of GAGW01008880 aligns to the MAT1-1-1 gene sequence KC437356 but is shorter due to the absence of exon I, intron I and 48 bp of exon II of the MAT1-1-1 gene (Figure 3). Its intron II (48 bp) is spliced between transcript nucleotides 358 and 359. Excluding the missing segments, GAGW01008880 is 100% identical to the remaining coding sequences (771 bp of exon II and the full length of exon III) of the MAT1-1-1 gene.
Xia et al. [2017] reported a metagenome-metatranscriptome project with fully mature C. sinensis specimens collected from Deqin County of Yunnan Province, China. The metatranscriptome assembly OSIN7648 in the Appendix [Xia et al. 2017] exhibits 87.3% similarity to the MAT1-1-1 gene sequence KC437356 and possesses 3 spliced/deleted segments, introns I and II (52 bp and 48 bp in pink in Figure 3, respectively), and an additional deleted segment of 54 bp (in blue in Figure 3) within exon II. Excluding the 3 spliced/deleted segments, OSIN7648 shares 100% identity with the coding sequences of exons I and III and the remaining exon II.
MAT1-1-1 transcripts in both metatranscriptome assemblies of natural C. sinensis are short compared with the coding sequence (1,119 bp) of the MAT1-1-1 gene of KC437356 [Bushley et al. 2013; Xiang et al. 2014; Xia et al. 2017]. As described above, OSIN7648 (1,065 bp) contains an additional deleted segment (54 bp in blue in Figure 3) within exon II of the MAT1-1-1 gene [Xia et al. 2017], which may produce a middle-truncated MAT1-1-1 protein that is missing 18 aa residues. Compared with the MAT1-1-1 gene, GAGW01008880 is 286 bp shorter due to the absence of a 238-bp segment of exon I and a 48-bp segment of exon II [Xiang et al. 2014] and may produce an N-terminal-truncated MAT1-1-1 protein. Figure 3 also shows a 296-bp noncoding segment of GAGW01008880 flanking downstream of the stop codon “TAG”.
The transcript sequences of the MAT1-1-2 and MAT1-1-3 genes were not identified in the metatranscriptome assemblies of the natural C. sinensis specimens [Xiang et al. 2014; Xia et al. 2017].
The MAT1-2-1 gene sequence was not detected in the metatranscriptome assembly GAGW00000000 of natural C. sinensis collected from Kangding County of Sichuan Province (cf. Figure 2) [Xiang et al. 2014]. However, the MAT1-2-1 gene sequence was found in the metatranscriptome assembly OSIN7649 of natural C. sinensis collected from Deqin County of Yunnan Province, which exhibits 100% homology with the transcriptome assembly GCQL01020543 of H. sinensis Strain L0106 [Liu et al. 2015; Xia et al. 2017].
Transcription of Mating-Type Genes in Unassembled Metatranscriptome Sequencing Read Archives of Natural and Cultivated C. sinensis
Zhang et al. [2018] reported differential transcription of the mating-type genes of natural C. sinensis at the early maturing stage. The MAT1-1-1 gene (OCS_06642) was reportedly transcribed at a similar level in all five specimens collected from the counties of Qumalai (specimen #1), Gonghe (specimen #2) and Zaduo (specimen #3) of Qinghai Province and Naqu (specimen #4) and Nyingchi (specimen #5) in Tibet. However, the heatmap shown in Figure 3B of [Zhang et al. 2018] demonstrated very different transcription in all five specimens. The MAT1-1-3 gene (OCS_06644) was transcribed at a low level only in specimen #4 but was silent in the other specimens, whereas the MAT1-2-1 gene (OCS_00196) was transcribed at low levels in specimens #4 and #5 but was silent in specimens #1‒3.
Zhong et al. [2018] observed differential transcription of mating-type genes in the early-developed stroma and caterpillar body of natural C. sinensis collected from Yushu, Qinghai Province, China. They reported that “the MAT1-1-1 gene could be detected in all three groups of samples” but did not report whether the transcript or cDNA of the MAT1-1-1 gene was detected in parallel. The authors also reported that “MAT1-1-2 was not detected in all samples”. They identified the transcripts of the MAT1-1-3 and MAT1-2-1 genes at “a very low read count value” but were unsure whether they “might participate in fruiting body development” in natural C. sinensis. Thus, they encouraged further studies aiming to verify the transcription of mating-type genes and the functions of mating proteins in natural C. sinensis at different developmental stages.
Li et al. [2019] observed the differential transcription of the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs at very low levels (with read counts of 0‒40 and fragments per kilobase of exon model per million mapped fragments [FPKM] values of 0‒8.4) in cultivated C. sinensis at all developmental phases (an FPKM value less than 10 indicates very low levels of transcription). The mating-type genes of the MAT1-1 idiomorph were not transcribed in the hyphae (read count=0) but were differentially transcribed at very low levels in cultivated C. sinensis at other developmental stages. The MAT1-2-1 gene was transcribed at very low levels (FPKM 0.16–0.93) in cultivated C. sinensis at all developmental stages.
Zhao et al. [2020] reported nearly no transcription of the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs (transcripts per million reads [TPM] of 0–2.27). The authors concluded that these genes might not play any role in the initiation of fruiting body development in cultivated C. sinensis.
We conducted a BLAST search of the GenBank database against the unassembled transcript reads as part of (1) the metatranscriptome BioProject PRJNA325365 and the Sequence Read Archive (SRA) SRP076425 of natural C. sinensis [Zhang et al. 2018]; (2) the SRA SRP103894 of natural C. sinensis at an early developmental stage [Zhong et al. 2018]; (3) the BioProject GSE123085 of cultivated C. sinensis at all developmental phases [Li et al. 2019]; and (4) the BioProject PRJNA600609 of cultivated C. sinensis at the initiation stage [Zhao et al. 2020]. We identified hundreds to thousands of unassembled transcript reads of MAT1-1-1, MAT1-1-2 and MAT1-2-1 cDNA. These unassembled transcript reads share 73.8–100% similarity to the coding sequences of the H. sinensis MAT1-1-1, MAT1-1-2 and MAT1-2-1 genes. No match was found between sequences of the H. sinensis MAT1-1-3 gene and the unassembled transcript reads of natural and cultivated C. sinensis. This range of variation between the sequences of the H. sinensis mating-type genes and the unassembled transcript reads indicates multiple fungal sources of the transcripts in natural and cultivated C. sinensis.
This section discusses the coexistence of multiple mating-type genes at the metatranscriptome level in natural and cultivated C. sinensis [Xiang et al. 2014; Xia et al. 2017; Zhang et al. 2018; Zhong et al. 2018; Li et al. 2019; Zhao et al. 2020]. The differential transcription of the mating-type genes of both the MAT1-1 and MAT1-2 idiomorphs in natural and cultivated C. sinensis, which contain >90 fungal species spanning at least 37 genera and multiple GC- and AT-biased genotypes of O. sinensis, provides insufficient evidence to prove that H. sinensis is the source of the transcripts and therefore does not sufficiently support the hypothesis that H. sinensis (Genotype #1 of O. sinensis) uses homothallic/pseudohomothallic mating strategies [Zhang et al. 2011, 2013, 2014, 2018; Bushley et al. 2013; Hu et al. 2013; Xiang et al. 2014; Zhang & Zhang 2015; Xia et al. 2017; Zhong et al. 2018; Li et al. 2019, 2022b, 2023c, 2024; Zhao et al. 2020].
Variations in Mating Proteins
The above sections explored the differential occurrence and transcription of the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs at the genome/metagenome and transcriptome/metatranscriptome levels and discussed the genetic, transcriptional, and coupled transcriptional-translational controls of the mating processes of H. sinensis in natural and cultivated C. sinensis. Allelic mutations in mating-type genes and their transcripts might influence protein translation and posttranslational processes.
The MAT1-1-1 protein sequences derived from the gene sequences of 127 H. sinensis strains show 98.1‒100% nucleotide homology [Zhang et al. 2009, 2011, 2013, 2014; Bushley et al. 2013; Hu et al. 2013; Zhang & Zhang 2015]. Among these, 29 sequences contained one to seven conserved and/or nonconserved mutated residues of a total of 372 aa residues at single-variant sites (Figure 4), sharing 98.1‒99.7% similarity with the MAT1-1-1 protein sequence AGW27560 of Strain CS68-2-1229. The MAT1-1-1 protein sequence (354 aa) translated from the metatranscriptome assembly OSIN7648 obtained from natural C. sinensis specimens collected from Deqin County, Yunnan Province, China, shows 92.5‒94.9% similarity to the MAT1-1-1 protein sequence (372 aa) of H. sinensis, with a substitution of leucine (L)→phenylalanine (F) and a deletion of 18 aa (SMQREYQAPREFYDYSVS) (Figure 3 and Figure 4) [Zhang et al. 2011, 2014; Bushley et al. 2013; Hu et al. 2013; Zhang & Zhang 2015; Xia et al. 2017]. This mutation likely produces a middle-truncated MAT1-1-1 protein. The MAT1-1-1 protein (276 aa) translated from the metatranscriptome assembly GAGW01008880 obtained from natural C. sinensis specimens collected from Kangding County of Sichuan Province, China, is 96 aa shorter (MTTRNEVMQRLSSVRADVLLNFLTDDAIFQLASRY-HESTTEADVLTPVSTAAASRATRQTKEASCDRAKRPLNAFMAFRSYYLKLFPDVQQQKTASG) at its N-terminus (Figure 3 and Figure 4) [Zhang et al. 2011, 2014; Bushley et al. 2013; Hu et al. 2013; Xiang et al. 2014; Zhang & Zhang 2015]. This mutation likely produces an N-terminal-truncated MAT1-1-1 protein.
The absence of the MAT1-1-1 transcript in the transcriptome of H. sinensis Strain L0106 and the remarkable variations in the MAT1-1-1 protein conformation in natural C. sinensis as translated from the MAT1-1-1 transcripts in the metatranscriptome assemblies may suggest heterogeneous fungal sources of the protein in natural insect-fungal complexes (Table 3) [Xiang et al. 2014; Liu et al. 2015; Xia et al. 2017].
Table 4 lists the 4 types of amino acids in MAT1-1-1 proteins based on their chemical-physical and hydrophobic properties determined using Peptide 2.0 [Peters & Elofsson 2014; Simm et al. 2016]. Compared with the percentage of amino acids of the MAT1-1-1 protein AGW27560 of H. sinensis Strain CS68-2-1229, changes in the amino acids of the MAT1-1-1 protein sequences translated from the metatranscriptome assemblies GAGW01008880 and OSIN7648 of natural C. sinensis indicated altered overall hydrophobicity-hydrophilicity of the proteins [Bushley et al. 2013; Xiang et al. 2014; Xia et al. 2017].
Figure 5 shows ExPASy ProtScale plots for the α-Helix, β-Sheet, β-Turn and Coil of the following MAT1-1-1 proteins: the authentic protein AGW27560 of H. sinensis Strain CS68-2-1229 and the middle and N-terminal-truncated proteins (OSIN7648 and GAGW01008880, respectively) of natural C. sinensis. The variable segment region of the MAT1-1-1 protein is outlined with open boxes in green in all panels of Figure 5. Substantial waveform changes are present in all OSIN7648 panels. The N-terminal-truncated region of the MAT1-1-1 protein GAGW01008880 is marked with open boxes in blue in the AGW27560 and OSIN7648 panels. The alterations in hydrophobicity (Table 4) and the α-Helix, β-Sheet, β-Turn and Coil plots (Figure 5) of the MAT1-1-1 proteins indicate variable folding and secondary structures, indicating disturbed functionality in the fungal mating process and, in turn, the sexual reproduction of O. sinensis.
The MAT1-2-1 protein sequences derived from 183 H. sinensis strains show 98.0‒100% homology; 64 strains contain one to five conserved and/or nonconserved mutated residues out of 249 aa residues (Figure 2 and Figure 6) [Zhang et al. 2009, 2011, 2014; Bushley et al. 2013; Hu et al. 2013; Zhang & Zhang 2015]. The MAT1-2-1 protein sequence translated from the metatranscriptome of OSIN7649 obtained from natural C. sinensis collected from Deqin County in Yunnan Province shows 100% identity to the MAT1-2-1 protein sequence AFX66389 derived from the genome sequence KC429550 of H. sinensis Strain GS09_121 [Zhang et al. 2014; Xia et al. 2017]. However, the MAT1-2-1 transcript was absent in the metatranscriptome assembly GAGW00000000 of natural C. sinensis collected from Kangding County in Sichuan Province (cf. Table 3) [Xiang et al. 2014]. Table 4 shows no change in the component amino acids of the MAT1-2-1 proteins, AFX66389 of H. sinensis Strain GS09_121 and OSIN7649 of natural C. sinensis, indicating unchanged hydrophobicity of the proteins [Zhang et al. 2014; Xia et al. 2017].
In addition to the above protein sequence analyses, Zhang and Zhang [2015] reported 4.7% and 5.7% allelic variations in the coding sequences of the MAT1-1-1 and MAT1-2-1 genes, respectively, resulting in 5.9% and 5.6% variations in the MAT1-1-1 and MAT1-2-1 protein sequences, respectively. The allelic and amino acid variations are much greater than what we observed through GenBank database analysis, suggesting that Zhang and Zhang [2015] may not have uploaded all gene and protein sequences that they analyzed to GenBank. It is conceivable that the mutant MAT1-1-1 and MAT1-2-1 proteins of natural C. sinensis might not necessarily be produced by Genotype #1 H. sinensis of O. sinensis but might be produced by one of multiple genotypes of O. sinensis fungi and cocolonized fungi when considering the failure to detect mating-type genes and transcripts within the MAT1-1 and MAT1-2 idiomorphs in numerous H. sinensis strains or wild-type isolates from natural C. sinensis (cf. Table 1 and Table 3) and “to induce development of the C. sinensis fruiting bodies” during 40 years of experience using pure H. sinensis cultures in research-oriented academic settings, as summarized by Zhang et al. [2013].
Occurrence and Transcription of Pheromone Receptor Genes and Variations in Pheromone Receptor Proteins in H. sinensis and Natural C. sinensis
Heterothallic sexual development in morphologically indistinguishable fungal haploid cells is controlled by the growth of complementary mating mycelia that produce complementary mating proteins followed by gametangial contact and the fusion of hyphal or ascosporic cells to induce plasmogamy and karyogamy in fungi [Jones & Bennett 2011; Zheng & Wang 2013; Wilson et al. 2015]. These mating processes for ascomycetes occur between two types of fungal cells, a and α cells, and the mating of these fungal cells is regulated by secreted pheromones. Pheromones play essential roles in choreographing the interactions between mating partners by regulating intercellular mating communication and conveying information from one type of cell to another cell type of the same species for physiological heterothallism or from a different species for hybridization. However, the biochemical processes for a- and α-pheromone precursors in ascomycetes appear to be complex and are not fully understood for O. sinensis. To date, scientists have not identified the genes encoding a- and α-pheromones in the genome assemblies of H. sinensis Strains 1229, CC1406-203, Co18, IOZ07 and ZJB12195 [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020].
An α-pheromone is recognized by a-type pheromone receptors (class 4 G-protein-coupled receptors) on the surface of an a-cell and vice versa [Jones & Bennett 2011; Zheng & Wang 2013; Wilson et al. 2015]. The activated receptor plays crucial roles in the reciprocal interaction of mate recognition and the initiation of signaling and mating processes.
a-Factor-Like Pheromone Receptor in H. sinensis and Natural C. sinensis
Hu et al. [2013] reported the occurrence of an a-pheromone receptor gene (PRE-1; 3→1,386 of KE659055) in H. sinensis Strain Co18. It contains 3 exons separated by introns I and II (437→490 and 1,018→1,075 of KE659055, respectively) (Figure 7). The PRE-1 gene sequences of the genome assemblies LKHE01002330 (1,592→2,975), NGJJ01000446 (84,292←85,675), ANOV01020521 (3→1,386) and JAAVMX010000007 (3,515,715→3,517,197) of H. sinensis Strains 1229, CC1406-203, Co18 and IOZ07, respectively, show 100% homology with each other [Hu et al. 2013; Li et al. 2016a; Liu et al. 2020; Shu et al. 2020]. However, the sequence of the PRE-1 gene exhibited 95.3% similarity with the genome assembly LWBQ01000004 (451,200→452,640) of H. sinensis Strain ZJB12195 due to multiple insertions and several transversion alleles in exon I (Table 5, Figure 7) [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020]. The coding sequence of PRE-1 is absent in the transcriptome assembly GCQL00000000 of H. sinensis Strain L0106 [Liu et al. 2015], indicating either silent transcription or the complete absence of the gene in a-cells in H. sinensis Strain L0106, resulting in impaired mate recognition upon interaction with the α-factor pheromone.
The PRE-1 gene sequence is present in the metatranscriptome assemblies OSIN6252 and GAGW01004735/GAGW01004736 of natural C. sinensis specimens collected from Deqin County of Yunnan Province and Kangding County of Sichuan Province in China, respectively, showing 98.5‒100% homology to the coding sequence of the PRE-1 gene KE659055 of H. sinensis Strain Co18 (Table 5) [Hu et al. 2013; Xiang et al. 2014; Xia et al. 2017].
The metatranscript OSIN6252 (220→1461) is 100% identical to the a-pheromone receptor gene sequence KE659055 of H. sinensis Strain Co18 (Figure 7). The metatranscript GAGW01004735 (1→764) is missing a 232-bp segment in exon I at its 5’ end and a 257-bp segment in exon III at its 3’ end, while the metatranscript GAGW01004736 (1029←1165) covers the missing portion of exon III at the 3’ end of the a-pheromone receptor gene sequence (Figure 7). Although the fragmentation of GAGW01004735 and GAGW01004736 may represent an assembly error if they are assumed to be derived from the same fungal genome [Xiang et al. 2015], the integration of the two fragments is still 232 bp short at the 5’ end of exon I of the PRE-1 gene.
In addition to the complete absence of the PRE-1 transcript and protein in H. sinensis Strain L0106 [Liu et al. 2015], the protein sequences translated from the metatranscriptome assembly OSIN6252 of natural C. sinensis are 100% identical to the protein sequence EQK97482 of the a-pheromone receptor that was directly translated from the PRE-1 gene KE659055 of H. sinensis Strain Co18 (Table 5; Figure 7 and Figure 8) [Hu et al. 2013; Xia et al. 2017]. The metatranscriptome GAGW01004735/GAGW01004736 of natural C. sinensis is 99.4% homologous to the protein sequence EQK97482 of the a-pheromone receptor (Table 5) [Hu et al. 2013; Xiang et al. 2014]. The translated PRE-1 protein sequence of GAGW01004735 is missing 75 aa and 85 aa at its N- and C-termini, respectively, whereas the other metatranscript, GAGW01004736, covers the C-terminal portion of the PRE-1 protein with two nonconserved mutated aa residues (Figure 8).
Table 6 lists the 4 types of amino acids in α-pheromone receptor proteins based on their chemical-physical and hydrophobicity properties using Peptide 2.0 [Peters & Elofsson 2014; Simm et al. 2016]. The segment deletion and nonconserved aa mutations in the PRE-1 protein sequence produce a truncated a-pheromone receptor protein without significant changes in hydrophobicity (50.4–50.7% hydrophobic amino acids) (Table 6) or in α-Helix, β-Sheet, β-Turn and Coil plots of the a-pheromone receptor proteins (GAGW01004735-GAGW01004736 and OSIN6252) in natural C. sinensis (data not shown), which raises the question of whether the mutations cause altered conformation of the membrane proteins and their functionality in interindividual mating signal communication between mating partners.
α-Factor-like Pheromone Receptor in H. sinensis and Natural C. sinensis
Hu et al. [2013] reported the occurrence of the α-pheromone receptor gene (2,275→3,724 of KE653642) in H. sinensis Strain Co18. This receptor gene exhibits 97.5‒100% homology with the genome assemblies LKHE01001069 (13,389→13,603)/LKHE01000580 (23,846←24,749; 23,657←23,794), NGJJ01001310 (128,759←130,010), ANOV01006352 (1→138)/ANOV01006351 (2,275→3,528), JAAVMX010000012 (406,964→408,405) and LWBQ01000044 (488,856→490,264) in H. sinensis Strains 1229, CC1406-203, Co18, IOZ07 and ZJB12195, respectively (Figure 9, Table 4) [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020]. Introns I and II of the α-pheromone receptor gene are located at sites 3,091→3,240 and 3,528→3,586 of the sequence KE653642 of Strain Co18 [Hu et al. 2013]. A 133-bp segment deletion in exon I of the gene occurs between the genome assembly segments of LKHE01001069 and LKHE01000580 of H. sinensis Strain 1229 (Figure 9), which encodes a 45-aa deleted segment (ALVVAFAVLALTPAAKLRRPSSLLHLAGLAMCLARVGSLAVPA-LS) in the protein sequence.
The α-pheromone receptor gene contains 3 exons and 2 introns (Figure 9). The coding sequence of the gene of H. sinensis Strain Co18 shows 99.3‒99.7% homology to the transcriptome assemblies GCQL01007648/GCQL01017756/GCQL01015779 of H. sinensis Strain L0106 and the metatranscriptome assembly OSIN6424 of natural C. sinensis collected from Deqin County of Yunnan Province, China (Table 4) [Hu et al. 2013; Liu et al. 2015; Xia et al. 2017]. GCQL01007648 (1←235) covers only 235 bp at the 5’ end of the 720 bp exon I. GCQL01015779 (1→908) and GCQL01017756 (1,274←2,180) cover 444 bp and 436 bp, respectively, at the 3’ end of exon I, and the full-length sequences of exons II and III. There are two gaps (138 bp and 145 bp) between the sequences of GCQL01007648 and GCQL01017756 and between those of GCQL01007648 and GCQL01015779 [Liu et al. 2015].
The presence of the α-pheromone receptor transcript but not the a-pheromone receptor transcript in H. sinensis Strain L0106 suggests that the strain can only receive a mating signal from a sexual partner that contains a-cells and produces a-pheromone. The differential transcription of pheromone receptor genes in H. sinensis Strain L0106 (Table 4), together with the differential transcription of mating-type genes of the MAT1-1 and MAT1-2 idiomorphs (cf. Table 3), indicates that H. sinensis Strain L0106 needs a sexual partner that produces a-pheromone and protein(s) of the MAT1-1 idiomorph for sexual reproduction. The sexual partner may be the same species for physiological heterothallism or a different species for hybridization when homothallic or pseudohomothallic reproduction is impossible, as described above.
The metatranscriptome assembly OSIN6424 of natural C. sinensis contains the sequences of all 3 exons of the α-pheromone receptor gene with a 58-bp deletion (GTCGTCATCATCCTCCCCCTGGGCA-CCCTCGCCGCCCAGCGCATGACAGCCCGTCCCA) between nucleotides 885 and 886 in exon II of the cDNA (cf. Figure 9) [Xia et al. 2017]. Notably, the deletion of 58 bp is not a multiple of 3 and may represent a frameshift mutation that involves a peptide segment of 52 aa encoded by the 3’ end region of exon II with no stop codon, while intron II of the gene is likely in “Intron Phase 0”, and the frameshift mutation may not involve exon III of the gene.
The α-pheromone receptor gene is absent in the metatranscriptome assembly GAGW00000000 of natural C. sinensis collected from Kangding County of Sichuan Province [Xiang et al. 2014].
The translated protein sequence of the metatranscriptome assembly OSIN6424 (1→1,224) from natural C. sinensis shows 82.9% similarity to the α-pheromone receptor protein sequence EQK99119, which was translated directly from the segment sequence of KE653642 of H. sinensis Strain Co18. OSIN6424 exhibits an insertion (38 aa), two segment deletions (9 and 11 aa) and 15-aa residues of nonconserved mutations (Figure 10, Table 4) [Hu et al. 2013; Xia et al. 2017].
The translated protein sequence of GCQL01007648 (38←235) of H. sinensis Strain L0106 is 100% identical to the small N-terminal portion (66 aa) of EQK99119 [Hu et al. 2013; Liu et al. 2015]. The translated protein sequences of GCQL01017756 and GCQL01015779 overlap and cover the majority of the protein sequence through the C-terminus. These sequences show 86.4% and 87.0% similarity to EQK99119 with a deletion of the 59‒61 aa residues AQLGAALVVAFAVLALTPAAKLRRPSSLLHLAGLAMCLAR-VGSLAVPALSPAGYFYVLW(AG) and several conserved and/or nonconserved mutated residues.
Compared with the percentages of amino acids of the α-pheromone receptor protein EQK99119 of H. sinensis Strain Co18 as shown in Table 6, changes in the amino acids of α-pheromone receptor protein sequences translated from the transcriptome assemblies GCQL01017756 and GCQL01015779 of H. sinensis Strain L0106 and the metatranscriptome assembly OSIN6424 of natural C. sinensis indicated dramatic decreases in hydrophobic amino acids and complementary increases in acidic, basic and neutral amino acids [Hu et al. 2013; Liu et al. 2015; Xia et al. 2017].
Figure 11 shows the ExPASy ProtScale plots for the α-Helix, β-Sheet, β-Turn and Coil of the α-pheromone receptor proteins: the authentic protein EQK99119 of H. sinensis Strain Co18 and the variable α-pheromone receptor sequences translated from the transcriptome assemblies GCQL01017756 and GCQL01015779 of H. sinensis Strain L0106 and the metatranscriptome assembly OSIN6424 of natural C. sinensis. Substantial waveform changes were found in all the GCQL01017756, GCQL01015779 and OSIN6424 panels and are outlined with open boxes in green. The N-terminal-truncated region of the proteins is marked with open boxes in blue in the EQK99119 and OSIN6424 panels.
Zheng & Wang [2013] reported that the α-pheromone receptor is a G protein-coupled receptor possessing 7 transmembrane domains. The membrane protein of the α-pheromone receptor EQK99119 of H. sinensis Strain Co18 contains 55.1% hydrophobic amino acids (cf. Table 6), which is significantly greater than the approximately 41% hydrophobic amino acids in MAT1-1-1 proteins (cf. Table 4). The mutant α-pheromone receptor proteins in H. sinensis Strain L0106 and natural C. sinensis contain fewer hydrophobic amino acids (50.8–52.7%). The alterations in the hydrophobicity (cf. Table 6) and the α-Helix, β-Sheet, β-Turn and Coil plots (cf. Figure 11) of the α-pheromone receptor proteins of H. sinensis Strain L0106 and natural C. sinensis indicate variable folding and secondary structures of the receptor proteins, indicating altered functionality of the sexual signal reception from mating partners and, in turn, the sexual reproduction of O. sinensis, probably favoring hybridization.
Other Pheromone-Related Genes in H. sinensis and Natural C. sinensis
Hu et al. [2013] also identified genes encoding a pheromone-regulated membrane protein (65,257→66,887 of KE652182), the a-pheromone processing metallopeptidase Ste23 (10361→13752 of KE652396), and other related genes in H. sinensis Strain Co18. The proteins encoded by these genes participate in the biological processes of mating protein activation and signal transduction. The segment sequences of KE652182 and KE652396 exhibit 99.9‒100% homology to the genome assemblies LKHE01001238 (91,967→93,597)/LKHE01000585 (20,198←23,589), NGJJ01000102 (63,761→65,391)/NGJJ01001093 (42,301→45,691), ANOV01000086 (17,505→19,135)/ANOV01001222 (10,361→13,752) and JAAVMX010000009 (1,427,620→1,429,250)/JAAVMX010000008 (1,724,511→1,727,903) of H. sinensis Strains 1229, CC1406-203, Co18 and IOZ07, respectively, but 95.8‒98.4% similarity to the genome assemblies LWBQ01000047 (64,210→65,902)/LWBQ01000028 (337,499←339,681) of Strain ZJB12195 due to multiple insertion/deletion mutations and other transition and transversion mutations in alleles [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020].
The KE652182 gene encoding a pheromone-regulated membrane protein shows 95.2‒96.3% similarity to the transcriptome assembly GCQL01011718 of H. sinensis Strain L0106 and the metatranscriptome assemblies GAGW01006658 and OSIN1278 of natural C. sinensis collected from Kangding County of Sichuan Province and Deqin County of Yunnan Province, respectively, with five to six deletion gaps [Hu et al. 2013; Xiang et al. 2014; Liu et al. 2015; Xia et al. 2017].
The KE652396 gene encoding the a-pheromone processing metallopeptidase Ste23 exhibits 99.2‒100% homology to the transcriptome assembly GCQL010123128 of H. sinensis Strain L0106 and the metatranscriptome assemblies GAGW01006419/GAGW01005794 and OSIN0988 of natural C. sinensis collected from Kangding County of Sichuan Province and Deqin County of Yunnan Province, respectively [Hu et al. 2013; Xiang et al. 2014; Liu et al. 2015; Xia et al. 2017].
Discussion
Reproductive Behavior of H. sinensis, Genotype #1 of O. sinensis
Three reproduction hypotheses have been previously proposed for H. sinensis, the postulated sole anamorph of O. sinensis [Zhu & Li 2017; Li et al. 2022a, 2022b]: homothallism [Hu et al. 2013], pseudohomothallism [Bushley et al. 2013] and facultative hybridization [Zhang & Zhang 2015]. In theory, self-fertilization in ascomycetes becomes a reality when the mating-type genes of both the MAT1-1 and MAT1-2 idiomorphs are successfully transcribed and translated, and the mating proteins are synthesized and fully activated within a single fungal cell [Turgeon & Yoder 2000; Debuchy & Turgeo 2006; Jones & Bennett 2011; Zhang et al. 2011; Bushley et al. 2013; Hu et al. 2013; Zheng & Wang 2013; Wilson et al. 2015; Zhang & Zhang 2015]. According to this study, the differential occurrence of the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs in 237 H. sinensis strains (cf. Table 1) fails to support the genetic-based capability of self-fertilization. The transcriptome assembly GCQL00000000 of H. sinensis Strain L0106 contains a transcript of the MAT1-2-1 gene but no transcripts of the MAT1-1 idiomorph [Liu et al. 2015]. The differential occurrence and transcription of mating-type genes at the genomic and transcriptomic levels are not consistent with the self-fertilization hypothesis for H. sinensis under homothallic or pseudohomothallic reproduction; instead, they support a reproductive strategy of either physiological heterothallism or hybridization [Turgeon & Yoder 2000; Pfennig 2007; Zheng & Wang 2013; Wilson et al. 2015; Du et al. 2020; Hėnault et al. 2020; Samarasinghe et al. 2020; Steensels et al. 2021].
Bushley et al. [2013] detected the full sequences of the MAT1-1-1, MAT1-1-2, MAT1-1-3 and MAT1-2-1 genes in single-ascospore isolates using a genome walking/tail PCR strategy. They also observed multicellular heterokaryotic hyphae and ascospores of natural C. sinensis with mononucleated, binucleated, trinucleated and tetranucleated structures (cf. Figure 3 of [Bushley et al. 2013]). However, they detected the alternatively spliced transcript of the MAT1-2-1 gene of H. sinensis Strain 1229 with an unspliced intron I that contains 3 stop codons (cf. Figure 2); possible technical errors of DNA contamination were ruled out because intron II was normally spliced. This transcription phenomenon indicates translational interruption of the MAT1-2-1 transcript and the production of a largely truncated MAT1-2-1 protein encoded only by exon I of the gene without the majority of the protein, which is encoded by exons II and III. This inability to produce a full-length and functional MAT1-2-1 protein resulting in dysfunctional MAT1-2-1 mating might constitute a mechanism of coupled transcriptional-translational regulation in the control of H. sinensis reproduction resulting in the induction of self-sterility.
In addition to demonstrating the multicellular heterokaryotic structures of C. sinensis hyphae and ascospores by Bushley et al. [2013], their coauthors Li et al. [2013] obtained 15 cultures from a mono-ascospore of natural C. sinensis: seven homogeneous clones contained only GC-biased Genotype #1 H. sinensis, and eight other clones heterogeneously contained both GC-biased Genotype #1 and AT-biased Genotype #5 of O. sinensis. The sequences of the GC- and AT-biased genotypes of O. sinensis reside in independent genomes and belong to different fungi [Xiao et al. 2009; Zhu et al. 2010, 2012, 2018, 2019; Li et al. 2016b, 2022b; Zhu & Li 2017]. Li et al. [2023a, 2023b] observed two types of ascospores (fully and semiejected) of natural C. sinensis and found the coexistence of GC-biased Genotypes #1 and #13‒14 of O. sinensis, AT-biased Genotypes #5‒6 and #16 of O. sinensis, Samsoniella hepiali and an AB067719-type fungus. Zhang and Zhang [2015] hypothesized that the nuclei of binucleated hyphal and ascosporic cells (as well as mononucleated, trinucleated and tetranucleated cells) of natural C. sinensis likely contain different genetic material. Thus, these multicellular hyphal and ascosporic cells of natural C. sinensis might contain two or more sets of fungal genomes of independent species, which might be responsible for producing complementary mating-type proteins for sexual reproductive outcrossing.
In addition to the translated protein sequences derived from the genome assembly sequences of five H. sinensis strains, GenBank also lists 183 MAT1-2-1 protein sequences of various H. sinensis strains, which were not obtained directly through protein purification and amino acid sequencing but instead derived from the nucleotide sequences of the MAT1-2-1 gene or transcript of H. sinensis [Zhang et al. 2009, 2011, 2014; Bushley et al. 2013; Hu et al. 2013; Zhang & Zhang 2015]. These protein sequences show 98.4‒100% similarity to the translated protein sequence of the transcriptome assembly GCQL01020543 of Strain L0106 [Liu et al. 2015]. Zhang and Zhang [2015] reported 4.7% and 5.7% allelic variations in the coding sequences of the H. sinensis MAT1-1-1 and MAT1-2-1 genes, respectively, and predicted that these high rates of genetic variation might cause translation disturbance of the coding sequences or 5.9% and 5.6% variations in the amino acid sequences.
As mentioned above, the MAT1-1-1 gene is not expressed in H. sinensis Strains CS2, L0106 and SCK05-4-3 and is even absent in the genomes of many other H. sinensis strains (cf. Table 1 and Table 3) [Zhang et al. 2009, 2011, 2014; Liu et al. 2015; Zhang & Zhang 2015]. In addition to the absence of the MAT1-2-1 gene in many H. sinensis strains (genetic regulation of the mating process) (cf. Table 1 and Table 3), the alternatively spliced transcript of the MAT1-2-1 gene with unspliced intron I and spliced intron II in H. sinensis Strain 1229 provides a translation template for the production of a largely truncated and dysfunctional MAT1-2-1 protein encoded by exon I but not by exons II and III, which constitutes an example of coupled transcriptional-translational regulation of the mating process [Bushley et al. 2013]. To date, there have been no experimental reports on the parallel production and direct amino acid sequencing of the mating proteins of both the MAT1-1 and MAT1-2 idiomorphs within a pure culture of H. sinensis, although many papers have used the word “expression” to describe the transcription of mating-type genes without considering other aspects of gene expression, such as epigenetic, posttranscriptional, translational and posttranslational modifications and protein activation/degradation processes.
Based on the differential occurrence and transcription of the mating-type genes, regardless of whether H. sinensis (Genotype #1), the postulated sole anamorph of O. sinensis, might be monoecious or dioecious, there may be two or more H. sinensis populations capable of producing either of the mating proteins of the MAT1-1 and MAT1-2 idiomorphs and functioning reciprocally as sexual partners for successful physiological heterothallism crossing. If this assumption is correct, the sexual partners might possess indistinguishable H. sinensis-like morphological and growth characteristics [Engh 1999; Kinjo & Zang 2001; Chen et al. 2004, 2011; Zhang et al. 2009; Li et al. 2013, 2016b; Mao et al. 2013]. For instance, the indistinguishable H. sinensis Strains 1229 and L0106 produce complementary transcripts of the mating-type genes and mating proteins of the MAT1-1 and MAT1-2 idiomorphs, as well as the a- and α-pheromone receptor genes, which are differentially transcribed and produce variable receptor proteins in the indistinguishable H. sinensis Strains Co18 and L0106. If the physiological heterothallism hypothesis is incorrect for O. sinensis, one of the mating proteins might be produced by heterospecific fungal species, which would result in plasmogamy and the formation of heterokaryotic cells (cf. Figure 3 of [Bushley et al. 2013]) to ensure a successful hybridization process if the heterospecific species are able to break interspecific reproduction isolation, similar to many cases of fungal hybridization that probably facilitate adaptation to the extremely adverse ecological environment on the Tibet-Qinghai Plateau [Pfennig 2007; Du et al. 2020; Hėnault et al. 2020; Samarasinghe et al. 2020; Steensels et al. 2021]. Alternatively, to complete physiological heterothallism or hybridization reproduction, mating partners might exist in three-dimensionally adjacent hyphal cells, which might make their mating choices and communicate with each other through a mating signal-based transduction system of pheromones and pheromone receptors and form “H”-shaped crossings of multicellular hyphae, as observed by Hu et al. [2013], Bushley et al. [2013] and Mao et al. [2013]. In particular, Mao et al. [2013] reported the observation of “H”-shaped morphology in C. sinensis hyphae that contained either AT-biased Genotype #4 or #5 of O. sinensis without the co-occurrence of GC-biased Genotype #1 H. sinensis, and the AT-biased O. sinensis genotypes shared indistinguishable H. sinensis-like morphological and growth characteristics.
Sexual Reproduction Strategy during the Lifecycle of Natural C. sinensis
The differential occurrence and transcription of the mating-type genes of both the MAT1-1 and MAT1-2 idiomorphs have also been observed in natural and cultivated C. sinensis, which contain multiple genotypes of O. sinensis and numerous fungal species [Engh 1999; Kinjo & Zang 2001; Chen et al. 2004, 2011; Xiao et al. 2009; Zhang et al. 2009, 2018; Zhu et al. 2010, 2012; Gao et al. 2011, 2012; Li et al. 2013, 2016b, 2016b, 2019, 2020, 2022b, 2023a, 2023b; Mao et al. 2013; Wei et al. 2016; Zhu & Li 2017; Zhong et al. 2018; Zhao et al. 2020; Yang et al. 2021]. As described above, transcriptome and metatranscriptome studies have shown differential transcription of mating-type genes in different maturation stages of natural and cultivated C. sinensis (cf. Figure 1), with a wide range of similarities compared with the sequences of the H. sinensis genes and transcripts and the unassembled metatranscriptome sequence reads, possibly indicating heterogeneous fungal sources of the transcripts. The middle and N-terminal truncated MAT1-1-1 proteins observed in natural C. sinensis exhibit variable hydrophobicity and alterations in the α-Helix, β-Sheet, β-Turn and Coil (cf. Figure 3, Figure 4 and Figure 5, Table 4), suggesting heteromorphic folding and altered primary and secondary structures of the MAT1-1-1 proteins, which could result in dysfunctional or anomalous fungal mating processes and may indicate the heterospecific fungal sources of the proteins needed for accomplishing hybridization in natural C. sinensis. In addition, the variable α-pheromone receptor proteins observed in natural C. sinensis exhibit changes in hydrophobicity and in the α-Helix, β-Sheet, β-Turn and Coil of the proteins (cf. Figure 9, Figure 10 and Figure 11, Table 6), indicating altered primary and secondary structures of the α-pheromone receptor proteins that could result in altered functionality in the sexual signal reception from mating partners in natural C. sinensis. Although the coexistence of larval tissues and fungal mycelia in the caterpillar body of natural C. sinensis indicates that larval tissues are not just culture medium that passively provide nutrients for fungal growth [Li et al. 2022b], the impact of host immunological reactions on the mating-type gene transcription of O. sinensis has not been explored, which represents the key process in the production of O. sinensis fruiting bodies and in the sexual life of natural and cultivated C. sinensis.
Regardless of whether H. sinensis (Genotype #1 of O. sinensis) is monoecious or dioecious, the sexual reproductive process of O. sinensis might require mating partners with the same or a different genotype of O. sinensis or even another fungal species to produce complementary mating proteins for physiological heterothallism or hybridization outcrossing. Thus, O. sinensis might have more than one monoecious or dioecious anamorph to accomplish the sexual reproduction of O. sinensis and the lifecycle of natural C. sinensis. These fungal partners might stay within a single heterokaryotic hyphal and ascosporic cell with mononucleated, binucleated, trinucleated or tetranucleated structures [cf. Figure 3 of Bushley et al. 2013], which would suggest that the following scientific observations from prior studies need to be reassessed:
(1) Li et al. [2013] detected GC-biased H. sinensis (Genotype #1) and AT-biased Genotype #5 of O. sinensis in eight of 15 cultures from mono-ascospores of natural C. sinensis. The authors misinterpreted all AT-biased genotypes as the “ITS pseudogene” components of the H. sinensis genome, while AT-biased Genotypes #4, #6 and #15‒17 were not detected by Li et al. [2013] in the cultures of the mono-ascospores and the sequences of all AT-biased genotypes residing not in the genomes of GC-biased H. sinensis Strains 1229, CC1406-203, Co18, IOZ07 and ZJB12195 but instead in the genomes of independent O. sinensis fungi [Xiao et al. 2009; Hu et al. 2013; Li et al. 2016a, 2016b, 2022a, 2022b, 2023a, 2023b, 2024; Zhu & Li 2017; Zhu et al. 2018, 2019; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020].
(2) Zhu et al. [2010, 2012] reported the cooccurrence of GC-biased Genotypes #1 and #2 of O. sinensis in the stromata of natural C. sinensis. The ITS sequences of these two genotypes share 94.7% sequence similarity [Li et al. 2016b, 2022b, 2023b; Zhu & Li 2017]. The Genotype #2 sequences were located outside the phylogenetic clade of Genotype #1 in the Bayesian trees and did not reside in the genome of Genotype #1 H. sinensis (Li et al. 2022b, 2023b, 2024). The abundances of the two genotypes undergo dynamic alterations in an asynchronous, disproportional manner in the stroma of natural C. sinensis during maturation (Figure 12) [Zhu et al. 2012; Li et al. 2022b], indicating the genomic independence of the two GC-biased genotypes as evidence of independent O. sinensis fungi.
(3) Chen et al. [2011] reported the detection of the Genotype #1 H. sinensis sequence AJ488255 from the caterpillar body of a natural C. sinensis specimen (#H1023) collected from Qinghai Province in China and the Genotype #7 sequence AJ488254 with multiple transversions and transition point mutations from the stroma of the same specimen [Li et al. 2016b, 2022b, 2023b; Zhu & Li 2017]. The GC-biased Genotype #7 sequence is located within the phylogenetic clade of GC-biased Genotype #1 in the Bayesian trees but does not reside in the genome of GC-biased Genotype #1 H. sinensis (Li et al. 2022b, 2023b, 2024).
(4) The cooccurrence of multiple AT-biased genotypes of O. sinensis and GC-biased Genotype #1 H. sinensis of O. sinensis in different combinations has been observed in the stroma, caterpillar body, ascocarps and ascospores of natural C. sinensis [Xiao et al. 2009; Zhu et al. 2010, 2012; Gao et al. 2011, 2012; Yao et al. 2011; Zhu & Li 2017; Li et al. 2022b, 2023a, 2023b]. The sequences of the mutant O. sinensis genotypes do not reside in the genome of GC-biased H. sinensis but instead belong to the genomes of independent fungi [Xiao et al. 2009; Zhu et al. 2010, 2012, 2019; Gao et al. 2011, 2012; Yao et al. 2011; Hu et al. 2013; Li et al. 2016a, 2016b, 2022b, 2023a, 2023b, 2024; Zhu & Li 2017; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020]. The abundances of the AT- and GC-biased genotypes of O. sinensis undergo dynamic alterations in an asynchronous, disproportional manner in the caterpillar body and stroma of C. sinensis during maturation, with a consistent predominance of the AT-biased genotypes, not GC-biased Genotype #1 H. sinensis, in the stromata [Zhu et al. 2010, 2012; Gao et al. 2011, 2012; Li et al. 2016b, 2022b; Zhu & Li 2017].
(5) Mao et al. [2013] identified AT-biased Genotype #4 or #5 of O. sinensis fungus without the co-occurrence of GC-biased H. sinensis in natural C. sinensis specimens collected from production areas in geographically remote locations. They also observed that the AT-biased mutant genotypes shared indistinguishable H. sinensis-like morphologic and growth characteristics and were able to form “H”-shaped hyphal crossings and anastomoses during germination, which are related to the sexual reproduction of O. sinensis. Similarly, Kinjo and Zang [2001] reported the detection of AT-biased Genotype #4 or #5 of O. sinensis in several natural C. sinensis specimens collected from remote production areas and GC-biased Genotype #1 H. sinensis in other C. sinensis specimens collected from different production areas.
(6) Hu et al. [2013] reported that H. sinensis has hermaphroditic and homothallic characteristics. Forty larvae of Hepialidae sp. were inoculated with a mixture of two pure H. sinensis strains, Co18 and QH195-2. Fungal inoculation induced death and mummification of the larvae but failed to induce the development of fruiting bodies and ascospores, indicating biological separation of the larval death/mummification process and fungal fruiting body development process. The authors cited two other studies [Holliday & Cleaver 2008; Stone 2010] and commented that inoculation of ghost moth larvae of the Hepialidae family with pure H. sinensis consistently failed to produce fruiting bodies and ascospores. Zhang et al. [2013] (coauthors of [Hu et al. 2013]) summarized 40 years of experience in artificial cultivation of C. sinensis and concluded that “it is very difficult in our laboratory to induce development of the C. sinensis fruiting bodies, either on culture medium or on insects.”
(7) Wei et al. [2016] reported a species contradiction between anamorphic inoculants (3 strains of GC-biased Genotype #1 H. sinensis: 130508-KD-2B, 20110514 and H01-20140924-03) and the sole teleomorph of AT-biased Genotype #4 of O. sinensis in the fruiting body of cultivated C. sinensis. In addition, Figure 6 of [Wei et al. 2016] shows two phylogenetically distinct teleomorphs of O. sinensis: AT-biased Genotype #4 of O. sinensis in cultivated C. sinensis and GC-biased Genotype #1 in the natural C. sinensis specimen G3, which was used as the teleomorphic reference in the phylogenetic analysis. Because the sequences of AT- and GC-biased genotypes of O. sinensis reside in independent genomes of different fungi [Xiao et al. 2009; Hu et al. 2013; Zhu et al. 2010, 2012, 2018, 2019; Li et al. 2016a, 2016b, 2020a, 2022b, 2024; Jin et al. 2020; Liu et al. 2020; Shu et al. 2020], Wei et al. [2016] demonstrated two distinct teleomorphs of O. sinensis and questioned the true causal fungus/fungi and anamorph-teleomorph connections of O. sinensis according to Koch’s postulates and the sole anamorph and sole teleomorph hypotheses proposed 10 years ago by the same group of key authors [Wei et al. 2006].
(8) Tolypocladium sinense in natural C. sinensis was first identified and published by Li [1988]. It was subsequently isolated from natural C. sinensis and characterized morphologically and genetically [Jiang & Yao 2003; Chen et al. 2004; Leung et al. 2006]. Engh [1999] reported the molecular identification of the Cordyceps-Tolypocladium complex in natural C. sinensis. The “Cordyceps” sequence AJ786590 obtained by Engh [1999] was published and uploaded to GenBank by Stensrud et al. [2005] and phylogenetically clustered into AT-biased Group B (Genotype #4) of O. sinensis, along with other C. sinensis sequences, by Stensrud et al. [2007]. Barseghyan et al. [2011] performed a macro/micromycology study and concluded that H. sinensis, which is presumed to be psychrophilic, and T. sinensis, which is presumed to be mesophilic, are dual anamorphs of O. sinensis. Notably, the O. sinensis fungus, which has H. sinensis-like morphology and growth characteristics, was not genotyped molecularly in that study.
(9) Genotypes #13 (KT339190) and #14 (KT339178) of O. sinensis have been identified in either semiejected or fully ejected multicellular heterokaryotic ascospores collected from the same specimen of natural C. sinensis [Zhu & Li 2017; Li et al. 2022b, 2023b]. The two genotypes feature precise reciprocal substitutions of large DNA segments due to chromosomal intertwining interactions and genetic material recombination between two parental fungi, Genotype #1 H. sinensis (Group A by Stensrud et al. [2007]) and an AB067719-type Group E fungus [Zhu & Li 2017; Li et al. 2022b, 2023b]. A pure culture of the AB067719-type fungus has not been obtained, and its taxonomic position is unclear. More than 900 sequences highly homologous to AB067719, including Alternaria sp., Ascomycota sp., Aspergillus sp., Avena sp., Berberis sp., Colletotrichum sp., Cordyceps sp., Cyanonectria sp., Dikarya sp., Fusarium sp., Gibberella sp., Hypocreales sp., Juglans sp., Lachnum sp., Nectria sp., Nectriaceae sp., Neonectria sp., and Penicillium sp., have been uploaded to GenBank [Li et al. 2023b]. Chromosomal intertwining and genetic material recombination may occur after plasmogamy and karyogamy of heterospecific parental fungi under sexual reproduction hybridization or parasexuality, which is characterized by the prevalence of heterokaryosis and results in concerted chromosome loss for transferring/substituting genetic materials without conventional meiosis [Bennett & Johnson 2003; Sherwood & Bennett 2009; Bushley et al. 2013; Seervai et al. 2013; Nakamura et al. 2019]. The phenomena of precise vertical transfer and reciprocal substitution of genetic materials between the chromosomes of heterospecific parental fungi that occurred differentially between the two types of ascospores collected from the same specimen of natural C. sinensis are distinct from the randomness and arbitrariness of horizontal environmental gene drift.

Table 7.
Percent similarities between the sequences of the ITS1, 5.8S and ITS2 segments of the parental fungi (Genotype #1 H. sinensis AB067721 and the AB067719-type Group E fungus) and the segment sequences of the offspring variant Genotypes #13 (KT339190, in the semiejected ascospores) and #14 (KT339178, in the fully ejected ascospores) of O. sinensis (adapted from [Zhu & LI 2017; Li et al. 2022b, 2023b]).
Table 7.
Percent similarities between the sequences of the ITS1, 5.8S and ITS2 segments of the parental fungi (Genotype #1 H. sinensis AB067721 and the AB067719-type Group E fungus) and the segment sequences of the offspring variant Genotypes #13 (KT339190, in the semiejected ascospores) and #14 (KT339178, in the fully ejected ascospores) of O. sinensis (adapted from [Zhu & LI 2017; Li et al. 2022b, 2023b]).
| Percent similarity | ||||
|---|---|---|---|---|
| ITS1-5.8S-ITS2 | ITS1 | 5.8S | ITS2 | |
| vs. Genotype #13 of O. sinensis (KT339190) | ||||
| Genotype #1 AB067721 O. sinensis | 86.3% | 100% | 94.8% | 64.2% |
| Group E AB067719 fungus | 88.2% | 71.5% | 100% | 99% |
| vs. Genotype #14 of O. sinensis (KT339178) | ||||
| Genotype #1 AB067721 O. sinensis | 87.7% | 67.9% | 94.9% | 100% |
| Group E AB067719 fungus | 89.2% | 100% | 100% | 71.5% |
(10) Paecilomyces hepiali was first isolated from natural C. sinensis by Dai et al. [Qiu et al. 1987; Dai et al. 1989; Jiang & Yao 2003] and renamed Samsoniella hepiali by Wang et al. [2020]. A close association of psychrophilic H. sinensis and mesophilic S. hepiali has been found in the caterpillar body, stroma, stromal fertile portion densely covered with ascocarps, and ascospores of natural C. sinensis and even in the formation of a fungal complex in “pure” H. sinensis strains that were isolated from natural C. sinensis and provided as gifts by a well-known mycology taxonomist [Jiang & Yao 2003; Chen et al. 2004; Zhu et al. 2007, 2010, 2012; Yang et al. 2008; Yao et al. 2011; Zhu & Wu 2015; Li et al. 2016b, 2016c, 2023b; Zhu & Li 2017]. Whether certain strains of these two fungal species would select each other as sexual partners will depend on their mating choices for hybridization and their abilities to break interspecific isolation barriers to adapt to extremely harsh ecological environments on the Qinghai‒Tibet Plateau and the seasonal change from the extremely cold winter when C. sinensis is in its asexual growth phase and to the spring and early summer when C. sinensis switches to the sexual reproduction phase [Pfennig 2007; Du et al. 2020; Hėnault et al. 2020; Samarasinghe et al. 2020; Steensels & Gallone 2021].
Conclusions
This paper analyses the differential occurrence and transcription of the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs and pheromone receptor genes of H. sinensis. The genomic and transcriptomic evidence is inconsistent with the self-fertilization hypothesis under homothallism and pseudohomothallism for H. sinensis but instead suggests self-sterility in O. sinensis, which utilizes physiological heterothallism or fungal hybridization strategies for sexual reproduction in natural and cultivated C. sinensis. H. sinensis Strains L0106 and 1229, either monoecious or dioecious, differentially transcribed the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs and might become sexual partners for physiological heterothallic outcrossing. H. sinensis Strain L0106 possesses the transcript of an α-pheromone receptor gene but not an a-pheromone receptor gene, indicating its ability to receive a mating signal from an a-pheromone secreted by a-cells of another fungus, either the same or different fungal species, to accomplish heterothallism or hybridization outcrossing for the development and maturation of the fruiting body, ascocarps and ascospores of natural C. sinensis. The mutations of MAT1-1-1 and α-pheromone receptor proteins observed in natural C. sinensis result in dramatically altered hydrophobicity properties and secondary protein structures, probably suggesting the heterogeneity of fungal source(s) of the proteins in the natural C. sinensis insect-fungal complex and sexual reproduction of O. sinensis under heterothallism or hybridization.
Author Contributions
Conceptualization, XZL, YLL and JSZ; methodology, JSZ; formal analysis, JSZ; data curation, XZL and JSZ; writing–original draft preparation, JSZ; writing—review and editing, XZL., YLL and JSZ; project administration, YLL; funding acquisition, YLL. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a grant from the Science and Technology Department of Qinghai Province, China, grant number 2021-SF-A4, “Study on key technologies of conservation of natural resource and industrial upgrading of Cordyceps sinensis”, the major science and technology projects in Qinghai Province.
Institutional Review Board Statement
Not applicable because this paper is public data reanalysis.
Informed Consent Statement
Not applicable because this paper is public data reanalysis.
Data Availability Statement
All sequence data are available in the GenBank database, except for the metatranscriptome sequences of natural C. sinensis that were submitted to the depository database www.plantkingdomgdb.com/Ophiocordyceps_sinensis/data/cds/Ophiocordyceps_sinensis_CDS.fas by Xia et al. [2017], which is currently inaccessible, but a previously downloaded cDNA file was used for the metatranscriptomic analysis.
Acknowledgments
The authors are grateful to Prof. Mu Zang, Prof. Ru-Qin Dai, Prof. Ping Zhu, Prof. Zong-Qi Liang for consultations and assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Barseghyan, G.S.; Holliday, J.C.; Price, T.C.; Madison, L.M.; Wasser, S.P. Growth and cultural-morphological characteristics of vegetative mycelia of medicinal caterpillar fungus Ophiocordyceps sinensis G.H. Sung et al. (Ascomycetes) Isolates from Tibetan Plateau (P.R.China). Intl. J. Med. Mushrooms. 2011, 13, 565–581. [Google Scholar] [CrossRef]
- Bennett, R.J.; Johnson, A.D. Completion of a parasexual cycle in Candida albicans by induced chromosome loss in tetraploid strains. EMBO J. 2003, 22, 2505–2515. [Google Scholar] [CrossRef] [PubMed]
- Bushley, K.E.; Li, Y.; Wang, W.-J.; Wang, X.-L.; Jiao, L.; Spatafora, J.W.; Yao, Y.-J. Isolation of the MAT1-1 mating type idiomorph and evidence for selfing in the Chinese medicinal fungus Ophiocordyceps sinensis. Fungal Biol. 2013, 117, 599–610. [Google Scholar] [CrossRef]
- Chen, C.-S.; Hseu, R.-S.; Huang, C.-T. Quality control of Cordyceps sinensis teleomorph, anamorph, and Its products. Chapter 12, in (Shoyama, Y., Ed.) Quality Control of Herbal Medicines and Related Areas. InTech, Rijeka, Croatia (2011). pp. 223–238. www.intechopen.com.
- Chen, Y.-Q.; Hu, B.; Xu, F.; Zhang, W.-M.; Zhou, H.; Qu, L.-H. Genetic variation of Cordyceps sinensis, a fruit-body-producing entomopathegenic species from different geographical regions in China. FEMS Microbiol. Lett. 2004, 230, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Chou, P.Y.; Fasman, G.D. Prediction of the secondary structure of proteins from their amino acid sequence. Adv. Enzymol. Relat. Areas. Mol. Biol. 1978, 47, 45–148. [Google Scholar] [CrossRef]
- Dai, R.-Q.; Lan, J.-L.; Chen, W.-H.; Li, X.-M.; Chen, Q.-T.; Shen, C.-Y. Discovery of a new fungus Paecilomyces hepiali Chen & Dai. Acta Agricult. Univ. Pekin. 1989, 15, 221–224. [Google Scholar]
- Debuchy R, Turgeo BG. Mating-Type Structure, Evolution, and Function in Euascomycetes. In (eds. Kües U. & Fischer, R.) Growth, Differentiation and Sexuality. Springer (2006). pp. 293–323.
- Deleage, G.; Roux, B. An algorithm for protein secondary structure prediction based on class prediction. Protein Engineering, Design and Selection 1987, 1, 289–294. [Google Scholar] [CrossRef]
- Dong, Y.-Z.; Zhang, L.-J.; Wu, Z.-M.; Gao, L.; Yao, Y.-S.; Tan, N.-Z.; Wu, J.-Y.; Ni, L.-Q.; Zhu, J.-S. Altered proteomic polymorphism in the caterpillar body and stroma of natural Cordyceps sinensis during maturation. PLoS ONE. 2014, 9, e109083. [Google Scholar] [CrossRef]
- Du, X.-H.; Wu, D.-M.; Kang, H.; Wang, H.-C.; Xu, N.; Li, T.-T.; Chen, K.-L. Heterothallism and potential hybridization events inferred for twenty-two yellow morel species. IMA Fungus. 2020, 11, 4. [Google Scholar] [CrossRef]
- Engh, I.B. Molecular phylogeny of the Cordyceps-Tolypocladium complex. Candidate scientific thesis, Department of Biology, University of Oslo, 1999.
- Gao, L.; Li, X.-H.; Zhao, J.-Q.; Lu, J.-H.; Zhao, J.-G.; Zhu, J.-S. Maturation of Cordyceps sinensis associates with alterations of fungal expressions of multiple Ophiocordyceps sinensis mutants with transition and transversion point mutations in stroma of Cordyceps sinensis. J. Peking Univ. (Health Sci.). 2012, 44, 454–463. [Google Scholar]
- Gao, L.; Li, X.-H.; Zhao, J.-Q.; Lu, J.-H.; Zhu, J.-S. Detection of multiple Ophiocordyceps sinensis mutants in premature stroma of Cordyceps sinensis by MassARRAY SNP MALDI-TOF mass spectrum genotyping. J. Peking Univ. (Health Sci.). 2011, 43, 259–266, https://www.ncbi.nlm.nih.gov/pubmed/21503123. [Google Scholar]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the Expasy Server. Chapter 52 (In) John M. Walker (ed): The Proteomics Protocols Handbook, Humana Press (2005). pp. 571–607.
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; Chen, Z.-H.; Mauceli, E.; Hacohen, N.; Gnirke, A.; Rhind, N.; di Palma, F.; Birren, B.W.; Nusbaum, C.; Lindblad-Toh, K.; Friedman, N.; Regev, A. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–130. [Google Scholar] [CrossRef] [PubMed]
- Hėnault, M.; Marsit, S.; Charron, G.; Landry, C.R. The effect of hybridization on transposable element accumulation in an undomesticated fungal species. eLife. 2020, 9, e60474. [Google Scholar] [CrossRef] [PubMed]
- Holliday, J.; Cleaver, M. Medicinal value of the caterpillar fungi species of the genus Cordyceps (Fr. ) Link (Ascomycetes). A review. Int. J. Med. Mushrooms. 2008, 10, 219–234. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Y.-J.; Xiao, G.-H.; Zheng, P.; Xia, Y.-L.; Zhang, X.-Y.; St Leger, R.J.; Liu, X.-Z.; Wang, C.-S. Genome survey uncovers the secrets of sex and lifestyle in caterpillar fungus. Chin. Sci. Bull. 2013, 58, 2846–2854. [Google Scholar] [CrossRef]
- Jiang, Y.; Yao, Y.-J. A review for the debating studies on the anamorph of Cordyceps sinensis. Mycosistema. 2003, 22, 161–176. [Google Scholar]
- Jin, L. -.Q.; Xu, Z.-.W.; Zhang, B.; Yi, M.; Weng, C.-.Y.; Lin, S.; Wu, H.; Qin, X.-T.; Xu, F.; Teng, Y.; Yuan, S.-J.; Liu, Z.-Q.; Zheng, Y.-G. Genome sequencing and analysis of fungus Hirsutella sinensis isolated from Ophiocordyceps sinensis. AMB Expr. 2020, 10, 105. [Google Scholar] [CrossRef]
- Jones, S.K.; Bennett, R.J. Fungal mating pheromones: choreographing the dating game. Fungal Genet. Biol. 2011, 48, 668–676. [Google Scholar] [CrossRef]
- Kinjo, N.; Zang, M. Morphological and phylogenetic studies on Cordyceps sinensis distributed in southwestern China. Mycoscience. 2001, 42, 567–574. [Google Scholar] [CrossRef]
- Leff, S.E.; Rosenfeld, M.G.; Evans, R.M. Complex transcriptional units: diversity in gene expression by alternative RNA processing. Ann. Rev. Biochem. 1986, 55, 1091–117. [Google Scholar] [CrossRef]
- Leung, P.-H.; Zhang, Q.-X.; Wu, J.-Y. Mycelium cultivation, chemical composition and antitumour activity of a Tolypocladium sp. fungus isolated from wild Cordyceps sinensis. J. Appl. Microbiol. 2006, 101, 275–283. [Google Scholar] [CrossRef]
- Li, C.-L. A study of Tolypocladium sinense C.L. Li. sp. nov. and cyclosporin production. Acta Mycol. Sinica. 1988, 7, 93–98. [Google Scholar]
- Li, X.; Wang, F.; Liu, Q.; Li, Q.-P.; Qian, Z.-M.; Zhang, X.-L.; Li, K.; Li, W.-J.; Dong, C.-H. Developmental transcriptomics of Chinese cordyceps reveals gene regulatory network and expression profiles of sexual development-related genes. BMC Genomics. 2019, 20, 337. [Google Scholar] [CrossRef]
- Li, X.-Z.; Li, Y.-L.; Wang, Y.-N.; Zhu, J.-S. Genetic and phylogenetic features of the repetitive ITS copies in the Hirsutella sinensis genome and the genomically independent AT-biased Ophiocordyceps sinensis genotypes. BioRxiv (the preprint server for biology) February 03, 2024. https://www.biorxiv.org/content/10.1101/2022.12.09. 03 February 5198. [Google Scholar]
- Li, X.-Z.; Li, Y.-L.; Yao, Y.-S.; Xie, W.-D.; Zhu, J.-S. Further discussion with Li et al. (2013, 2019) regarding the “ITS pseudogene hypothesis” for Ophiocordyceps sinensis. Mol. Phylogenet. Evol. 2020, 146, 106728. [Google Scholar] [CrossRef]
- Li, X.-Z.; Li, Y.-L.; Yao, Y.-S.; Zhu, J.-S. Differential expression of Hirsutella sinensis genes and intraspecific genetic variation among H. sinensis strains. Biochem. Mol. Biol. J. 2022, 8, 58. [Google Scholar] [CrossRef]
- Li, X.-Z.; Li, Y.-L.; Zhu, J.-S. Differential expression of mating-type genes in Hirsutella sinensis and natural Cordyceps sinensis Proc. 2021 Intl. Conf. Info. Technol. Biomed. Engineering, 2021, pp. 318−333. https://conferences.computer.org/icitbepub/pdfs/ICITBE2021-6JNtJiThlp10s9N5Sodvuy/009900a318/009900a318.pdf. 2021. [Google Scholar]
- Li, Y.; Hsiang, T.; Yang, R.-H.; Hu, X.-D.; Wang, K.; Wang, W.-J.; Wang, X.-L.; Jiao, L.; Yao, Y.-J. Comparison of different sequencing and assembly strategies for a repeat-rich fungal genome, Ophiocordyceps sinensis. J. Microbiol. Methods. 2016, 128, 1–6. [Google Scholar] [CrossRef]
- Li, Y.; Jiao, L.; Yao, Y.-J. Non-concerted ITS evolution in fungi, as revealed from the important medicinal fungus Ophiocordyceps sinensis. Mol. Phylogenet. Evol. 2013, 68, 373–379. [Google Scholar] [CrossRef]
- Li, Y.-L.; Gao, L.; Yao, Y.-S.; Wu, Z.-M.; Lou, Z.-Q.; Xie, W.-D.; Wu, J.-Y.; Zhu, J.-S. Altered GC- and AT-biased genotypes of Ophiocordyceps sinensis in the stromal fertile portions and ascospores of natural Cordyceps sinensis. PLoS ONE. 2023, 18, e0286865. [Google Scholar] [CrossRef]
- Li, Y.-L.; Li, X.-Z.; Yao, Y.-S.; Wu, Z.-M.; Gao, L.; Tan, N.-Z.; Lou, Z.-Q.; Xie, W.-D.; Wu, J.-Y.; Zhu, J.-S. Differential cooccurrence of multiple genotypes of Ophiocordyceps sinensis in the stromata, stromal fertile portion (ascocarps) and ascospores of natural Cordyceps sinensis. PLoS ONE. 2023, 18, e0270776. [Google Scholar] [CrossRef]
- Li, Y.-L.; Li, X.-Z.; Yao, Y.-S.; Xie, W.-D.; Zhu, J.-S. Molecular identification of Ophiocordyceps sinensis genotypes and the indiscriminate use of the Latin name for the multiple genotypes and the natural insect-fungi complex. Am J BioMed Sci. 2022, 14, 115–135. [Google Scholar] [CrossRef]
- Li, Y.-L.; Yao, Y.-S.; Xie, W.-D.; Zhu, J.-S. Molecular heterogeneity of natural Cordyceps sinensis with multiple Ophiocordyceps sinensis fungi challenges the anamorph-teleomorph connection hypotheses and implementation of the Amsterdam Declaration of IMA. Am. J. Biomed. Sci. 2016, 8, 123–159. [Google Scholar] [CrossRef]
- Li, Y.-L.; Yao, Y.-S.; Zhang, Z.-H.; Xu, H.-F.; Liu, X.; Ma, S.-L.; Wu, Z.-M.; Zhu, J.-S. Synergy of fungal complexes isolated from the intestines of Hepialus lagii larvae in increasing infection potency. J. Fungal Res. 2016, 14, 96–112. [Google Scholar]
- Liang, Z.-Q.; Han, Y.-F.; Liang, J.-D.; Dong, X.; Du, W. Issues of concern in the studies of Ophiocordyceps sinensis. Microbiol. Chin. 2010, 37, 1692–1697. [Google Scholar]
- Liu, J.; Guo, L.-N.; Li, Z.-W.; Zhou, Z.; Li, Z.; Li, Q.; Xiaochen Bo, X.-C.; Wang, S.-Q.; Wang, J.-L.; Ma, S.-C.; Zheng, J.; Yang, Y. Genomic analyses reveal evolutionary and geologic context for the plateau fungus Ophiocordyceps sinensis. Clin. Med. 2020, 15, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-Q.; Lin, S.; Baker, P.J.; Wu, L.-F.; Wang, X.-R.; Wu, H.; Xu, F.; Wang, H.-Y.; Brathwaite, M.E.; Zheng, Y.-G. Transcriptome sequencing and analysis of the entomopathogenic fungus Hirsutella sinensis isolated from Ophiocordyceps sinensis. BMC Genomics. 2015, 16, 106–123. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-L.; St Leger, R.J. Chapter Seven - Insect Immunity to Entomopathogenic Fungi, Editors: Lovett B, St. Leger RJ. Advanc Genet, Academic Press, 2016. Vol. 94; pp. 251–85.
- Mao, X.-M.; Zhao, S.-M.; Cao, L.; Yan, X.; Han, R.-C. The morphology observation of Ophiocordyceps sinensis from different origins. J. Environ. Entomol. 2013, 35, 343–353. [Google Scholar]
- Meng, Q.; Yu, H.-Y.; Zhang, H.; Zhu, W.; Wang, M.-L.; Zhang, J.-H.; Zhou, G.-L.; Li, X.; Qin, Q.-L.; Hu, S.-N.; Zou, Z. Transcriptomic insight into the immune defenses in the ghost moth, Hepialus xiaojinensis, during an Ophiocordyceps sinensis fungal infection. Insect Biochem. Mol. Biol. 2015, 64, 1–15. [Google Scholar] [CrossRef]
- Nakamura, N.; Tanaka, C.; Takeuchi-Kaneko, Y. Transmission of antibiotic-resistance markers by hyphal fusion suggests partial presence of parasexuality in the root endophytic fungus Glutinomyces brunneus. Mycol. Progress 2019, 18, 453–462. [Google Scholar] [CrossRef]
- Peters, C.; Elofsson, A. Why is the biological hydrophobicity scale more accurate than earlier experimental hydrophobicity scales? Proteins. 2014, 82, 2190–2198. [Google Scholar] [CrossRef]
- Pfennig, K.S. Facultative Mate Choice Drives Adaptive Hybridization. Sci. 2007, 318, 965–967. [Google Scholar] [CrossRef]
- Qiu W-F, Chen Q-T, Qian C-R, Shen C-Y, Xu J-T, Wang L-Y, Zhang W-J. “Study on Cordyceps sinensis, a new species Paecilomyces hepiali” certificate of identification for the achievement in science and technology. In Archive of Institution of Chinese Materia Medica, Chinese Academy of Traditional Chinese Medicine (Unpublished; January 8, 1987) Docs #000021, #000044, and #000045.
- Ren, Y.; Wan, D.-G.; Lu, X.-M.; Guo, J.-L. The study of scientific name discussion for TCM Cordyceps. LisShenzhen Med. Material Medica Res. 2013, 24, 2211–2212. [Google Scholar]
- Samarasinghe, H.; You, M.; Jenkinson, T.S.; Xu, J.-P.; James, T.Y. Hybridization Facilitates Adaptive Evolution in Two Major Fungal Pathogens. Genes. 2020, 11, 101. [Google Scholar] [CrossRef]
- Seervai, R.N.H.; Jones, S.K.; Hirakawa, M.P.; Porman, A.M.; Bennett, R.J. Parasexuality and ploidy change in Candida tropicalis. Eukaryotic Cell. 2013, 12, 1629–1640. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, R.K.; Bennett, R.J. Fungal meiosis and parasexual reproduction--lessons from pathogenic yeast. Current Opinion Microbiol. 2009, 12, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Shu, R.-H.; Zhang, J.-H.; Meng, Q.; Zhang, H.; Zhou, G.-L.; Li, M.-M.; Wu, P.-P.; Zhao, Y.-N.; Chen, C.; Qin, Q. A new high-quality draft genome assembly of the Chinese cordyceps Ophiocordyceps sinensis. Genome Biol. Evol. 2020, 12, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Simm S, Einloft J, Mirus O, Schleiff E. 50 years of amino acid hydrophobicity scales: revisiting the capacity for peptide classification. Biol. Res. 2016, 49, 31. [Google Scholar] [CrossRef]
- Steensels, J.; Gallone, B.; Verstrepen, K.J. Interspecific hybridization as a driver of fungal evolution and Adaptation. Nat. Rev. Microbiol. 2021, 19, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Stensrud Ø, Hywel-Jones NL, Schumacher T. Towards a phylogenetic classification of Cordyceps: ITS nrDNA sequence data confirm divergent lineages and paraphyly. Mycol. Res. 2005, 109, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Stensrud Ø, Schumacher T, Shalchian-Tabrizi K, Svegardenib IB, Kauserud H. Accelerated nrDNA evolution and profound AT bias in the medicinal fungus Cordyceps sinensis. Mycol. Res. 2007, 111, 409–415. [Google Scholar] [CrossRef]
- Stone, R. Improbable partners aim to bring biotechnology to a Himalayan kingdom. Sci. 2010, 327, 940–941. [Google Scholar] [CrossRef]
- Sung, G.-H.; Hywel-Jones, N.L.; Sung, J.-M.; Luangsa-ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef]
- Turgeon, B.G.; Yoder, O.C. Proposed nomenclature for mating type genes of filamentous ascomycetes. Fungal Genet. Biol. 2000, 31, 1–5. [Google Scholar]
- Wang, J.-B.; St Leger, R.J.; Wang, C.-S. Advances in Genomics of Entomopathogenic Fungi. Advan. Genetics 2016, 94, 1–39. [Google Scholar] [CrossRef]
- Wang, Y.; Stata, M.; Wang, W.; Stajich, J.E.; White, M.M.; Moncalvo, J.M. Comparative genomics reveals the core gene toolbox for the fungus-insect symbiosis. mBio. 2018, 9, e00636–e18. [Google Scholar] [CrossRef]
- Wang, Y.-B.; Wang, Y.; Fan, Q.; Duan, D.-E.; Zhang, G.-D.; Dai, R.-Q.; Dai, Y.-D.; Zeng, W.-B.; Chen, Z.-H.; Li, D.-D.; Tang, D.-X.; Xu, Z.-H.; Sun, T.; Nguyen, T.-T.; Tran, N.-L.; Dao, V.-M.; Zhang, C.-M.; Huang, L.-D.; Liu, Y.-J.; Zhang, X.-M.; Yang, D.-R.; Sanjuan, T.; Liu, X.-Z.; Yang, Z.-L.; Yu, H. Multigene phylogeny of the family Cordycipitaceae (Hypocreales): new taxa and the new systematic position of the Chinese cordycipitoid fungus Paecilomyces hepiali. Fungal Diversity. 2020, 103, 1–46. [Google Scholar] [CrossRef]
- Wei, J.-C.; Wei, X.-L.; Zheng, W.-F.; Guo, W.; Liu, R.-D. Species identification and component detection of Ophiocordyceps sinensis cultivated by modern industry. Mycosystema. 2016, 35, 404–410. [Google Scholar]
- Wei, X.-L.; Yin, X.-C.; Guo, Y.-L.; Shen, N.-Y.; Wei, J.-C. Analyses of molecular systematics on Cordyceps sinensis and its related taxa. Mycosystema. 2006, 25, 192–202. [Google Scholar]
- Wilson, A.M.; Wilken, P.M.; van der Nest, M.A.; Steenkamp, E.T.; Wingfield, M.J.; Wingfield, B.D. Homothallism: an umbrella term for describing diverse sexual behaviours. IMA Fungus. 2015, 6, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.-H.; Yang, D.-R.; Jiang, J.-J.; Zhang, Q.-J.; Liu, Y.; Liu, Y.-L.; Zhang, Y.; Zhang, H.-B.; Shi, C.; Tong, Y.; Kim, C.-H.; Chen, H.; Peng, Y.-Q.; Yu, Y.; Zhang, W.; Eichler, E.E.; Gao, L.-Z. The caterpillar fungus, Ophiocordyceps sinensis, genome provides insights into highland adaptation of fungal pathogenicity. Sci. Rep. 2017, 7, 1806. [Google Scholar] [CrossRef]
- Xia, F.; Liu, Y.; Shen, G.-L.; Guo, L.-X.; Zhou, X.-W. Investigation and analysis of microbiological communities in natural Ophiocordyceps sinensis. Can. J. Microbiol. 2015, 61, 104–111. [Google Scholar] [CrossRef]
- Xiang, L.; Li, Y.; Zhu, Y.; Luo, H.; Li, C.; Xu, X.; Sun, C.; Song, J.-Y.; Shi, L.-H.; He, L.; Sun, W.; Chen, S.-L. Transcriptome analysis of the Ophiocordyceps sinensis fruiting body reveals putative genes involved in fruiting body development and cordycepin biosynthesis. Genomics. 2014, 103, 154–159. [Google Scholar] [CrossRef]
- Xiao, W.; Yang, J.-P.; Zhu, P.; Cheng, K.-D.; He, H.-X.; Zhu, H.-X.; Wang, Q. Non-support of species complex hypothesis of Cordyceps sinensis by targeted rDNA-ITS sequence analysis. Mycosystema. 2009, 28, 724–730. [Google Scholar]
- Yang, J.-L.; Xiao, W.; He, H.-H.; Zhu, H.-X.; Wang, S.-F.; Cheng, K.-D.; Zhu, P. Molecular phylogenetic analysis of Paecilomyces hepiali and Cordyceps sinensis. Acta Pharmaceut. Sinica. 2008, 43, 421–426. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Tong, X.-X.; He, C.-Y.; Bai, J.; Wang, F.; Guo, J.-L. Comparison of endogenetic microbial community diversity between wild Cordyceps sinensis, artificial C. sinensis and habitat soil. Chin. J. Chin. Materia Medica. 2021, 46, 3106–3115. [Google Scholar]
- Yao, Y.-S.; Gao, L.; Li, Y.-L.; Ma, S.-L.; Wu, Z.-M.; Tan, N.-Z.; Wu, J.-Y.; Ni, L.-Q.; Zhu, J.-S. Amplicon density-weighted algorithms for analyzing dissimilarity and dynamic alterations of RAPD polymorphisms of Cordyceps sinensis. Beijing Da Xue Xue Bao Yi Xue Bao. 2014, 46, 618–628, (http://xuebao.bjmu.edu.cn/EN/Y2014/V46/I4/618). [Google Scholar]
- Yao, Y.-S.; Zhou, Y.-J.; Gao, L.; Lu, J.-H.; Zhu, J.-S. Dynamic alterations of the differential fungal expressions of Ophiocordyceps sinensis and its mutant genotypes in stroma and caterpillar during maturation of natural Cordyceps sinensis. J. Fungal Res. 2011, 9, 37–49. [Google Scholar]
- Zhang, S.; Zhang, Y.-J. Molecular evolution of three protein-coding genes in the Chinese caterpillar fungus Ophiocordyceps sinensis. Microbiol. China. 2015, 42, 1549–1560. [Google Scholar]
- Zhang, S.; Zhang, Y.-J.; Liu, X.-Z.; Wen, H.-A.; Wang, M.; Liu, D.-S. Cloning and analysis of the MAT1-2-1 gene from the traditional Chinese medicinal fungus Ophiocordyceps sinensis. Fungal Biol. 2011, 115, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Zhang S, Zhang Y-J, Shrestha B, Xu J-P, Wang C-S, Liu X-Z. Ophiocordyceps sinensis and Cordyceps militaris: research advances, issues and perspectives. Mycosystema. 2013, 32, 577−597.
- Zhang, S.-W.; Cen, K.; Liu, Y.; Zhou, X.-W.; Wang, C.-S. Metatranscriptomics analysis of the fruiting caterpillar fungus collected from the Qinghai-Tibetan plateau. Sci. Sinica Vitae. 2018, 48, 562–570. [Google Scholar]
- Zhang Y-J, Li E-W, Wang C-S, Li Y-L, Liu X-Z. Ophiocordyceps sinensis, the flagship fungus of China: terminology, life strategy and ecology. Mycol. 2012, 3, 2–10, https://www.tandfonline.com/doi/full/10.1080/21501203.2011.654354.
- Zhang Y-J, Sun B-D, Zhang S, Wàngmŭ, Liu X-Z, Gong W-F. Mycobiotal investigation of natural Ophiocordyceps sinensis based on culture-dependent investigation. Mycosistema. 2010, 29, 518–527. [Google Scholar]
- Zhang Y-J, Xu L-L, Zhang S, Liu X-Z, An Z-Q, Wàngmŭ, et al. Genetic diversity of Ophiocordyceps sinensis, a medicinal fungus endemic to the Tibetan Plateau: implications for its evolution and conservation. BMC Evol. Biol. 2009, 9, 290. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Zhang, S.; Li, Y.-L.; Ma, S.-L.; Wang, C.-S.; Xiang, M.-C.; Liu, X.; An, Z.-Q.; Xu, J.-P.; Liu, X.-Z. Phylogeography and evolution of a fungal–insect association on the Tibetan Plateau. Mol. Ecol. 2014, 23, 5337–5355. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-N.; Zhang, J.-H.; Meng, Q.; Zhang, H.; Zhou, G.-L.; Li, M.-M.; Wu, P.-P.; Shu, R.-H.; Gao, X.-X.; Guo, L.; Tong, Y.; Cheng, L.-Q.; Guo, L.; Chen, C.; Qin, Q. Transcriptomic analysis of the orchestrated molecular mechanisms underlying fruiting body initiation in Chinese cordyceps. Gene. 2020, 763, 145061. [Google Scholar] [CrossRef]
- Zheng, P.; Wang, C.-S. Sexuality Control and Sex Evolution in Fungi. Sci Sinica Vitae. 2013, 43, 1090–1097. [Google Scholar]
- Zhong, X.; Gu, L.; Wang, H.-Z.; Lian, D.-H.; Zheng, Y.-M.; Zhou, S.; Zhou, W.; Gu, J.; Zhang, G.; Liu, X. Profile of Ophiocordyceps sinensis transcriptome and differentially expressed genes in three different mycelia, sclerotium and fruiting body developmental stages. Fungal Biol. 2018, 122, 943–951. [Google Scholar] [CrossRef]
- Zhu, J.-S.; Gao, L.; Li, X.-H.; Yao, Y.-S.; Zhou, Y.-J.; Zhao, J.-Q.; Zhou, Y.-J. Maturational alterations of oppositely orientated rDNA and differential proliferations of CG:AT-biased genotypes of Cordyceps sinensis fungi and Paecilomyces hepiali in natural C. sinensis. Am. J. Biomed. Sci. 2010, 2, 217–238. [Google Scholar] [CrossRef]
- Zhu, J.-S.; Guo, Y.-L.; Yao, Y.-S.; Zhou, Y.-J.; Lu, J.-H.; Qi, Y.; Chen, W.; Zheng, T.-Y.; Zhang, L.; Wu, Z.-M.; Zhang, L.-J.; Liu, X.-J.; Yin, W.-T. Maturation of Cordyceps sinensis associates with co-existence of Hirsutella sinensis and Paecilomyces hepiali DNA and dynamic changes in fungal competitive proliferation predominance and chemical profiles. J. Fungal Res. 2007, 5, 214–224. [Google Scholar]
- Zhu, J.-S.; Halpern, G.M.; Jones, K. The scientific rediscovery of a precious ancient Chinese herbal regimen: Cordyceps sinensis: Part I. J. Altern. Complem. Med. 1998, 4, 289–303. [Google Scholar] [CrossRef]
- Zhu, J.-S.; Halpern, G.M.; Jones, K. The scientific rediscovery of an ancient Chinese herbal medicine: Cordyceps sinensis: Part II. J. Altern. Complem. Med. 1998, 4, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-S.; Li, C.-L.; Tan, N.-Z.; Berger, J.L.; Prolla, T.A. Combined use of whole-gene expression profiling technology and mouse lifespan test in anti-aging herbal product study. Proc. TCM Products Innovation and Industrial Development Summit, Hangzhou, China (Nov 27, 2011). pp. 443–448. https://xueshu.baidu.com/usercenter/paper/show?paperid=08341c17fa58c8f85584b92572b90f75&site=xueshu_se.
- Zhu, J.-S.; Li, Y.-L. A Precious Transitional Chinese Medicine, Cordyceps sinensis: Multiple heterogeneous Ophiocordyceps sinensis in the insect-fungi complex. Lambert Academic Publishing, Saarbrüchen. Germany (.
- Zhu, J.-S.; Li, Y.-L.; Yao, Y.-S.; Xie, W.-D. Molecular evidence not supporting the use of Hirsutella sinensis strain EFCC7287 as the taxonomic standard for multiple Ophiocordyceps sinensis fungi. Intl. J. Mol. Biol. 2018, 3, 114–115. [Google Scholar]
- Zhu, J.-S.; Li, Y.-L.; Yao, Y.-S.; Xie, W.-D. The multiple genotypes of Ophiocordyceps sinensis and the “ITS pseudogene” hypothesis. Mol. Phylogenet. Evol. 2019, 139, 106322. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-S.; Wu, J.-Y. Genetic heterogeneity of natural Cordyceps sinensis with co-existence of multiple fungi. Chin. J. Cell Biol. 2015, 37, 284–298. [Google Scholar]
- Zhu, J.-S.; Zhao, J.-G.; Gao, L.; Li, X.-H.; Zhao, J.-Q.; Lu, J.-H. Dynamically altered expressions of at least 6 Ophiocordyceps sinensis mutants in the stroma of Cordyceps sinensis. J. Fungal Res. 2012, 10, 100–112. [Google Scholar]
Figure 1.
Images of immature, maturing, and mature natural C. sinensis (modified from [Zhu et al. 2010; Yao et al. 2011, 2014]). The right two panels show mature C. sinensis specimens with the expanded stromal fertile portion (SFP) close to the tip of the C. sinensis stroma, which is densely covered with numerous ascocarps.
Figure 1.
Images of immature, maturing, and mature natural C. sinensis (modified from [Zhu et al. 2010; Yao et al. 2011, 2014]). The right two panels show mature C. sinensis specimens with the expanded stromal fertile portion (SFP) close to the tip of the C. sinensis stroma, which is densely covered with numerous ascocarps.
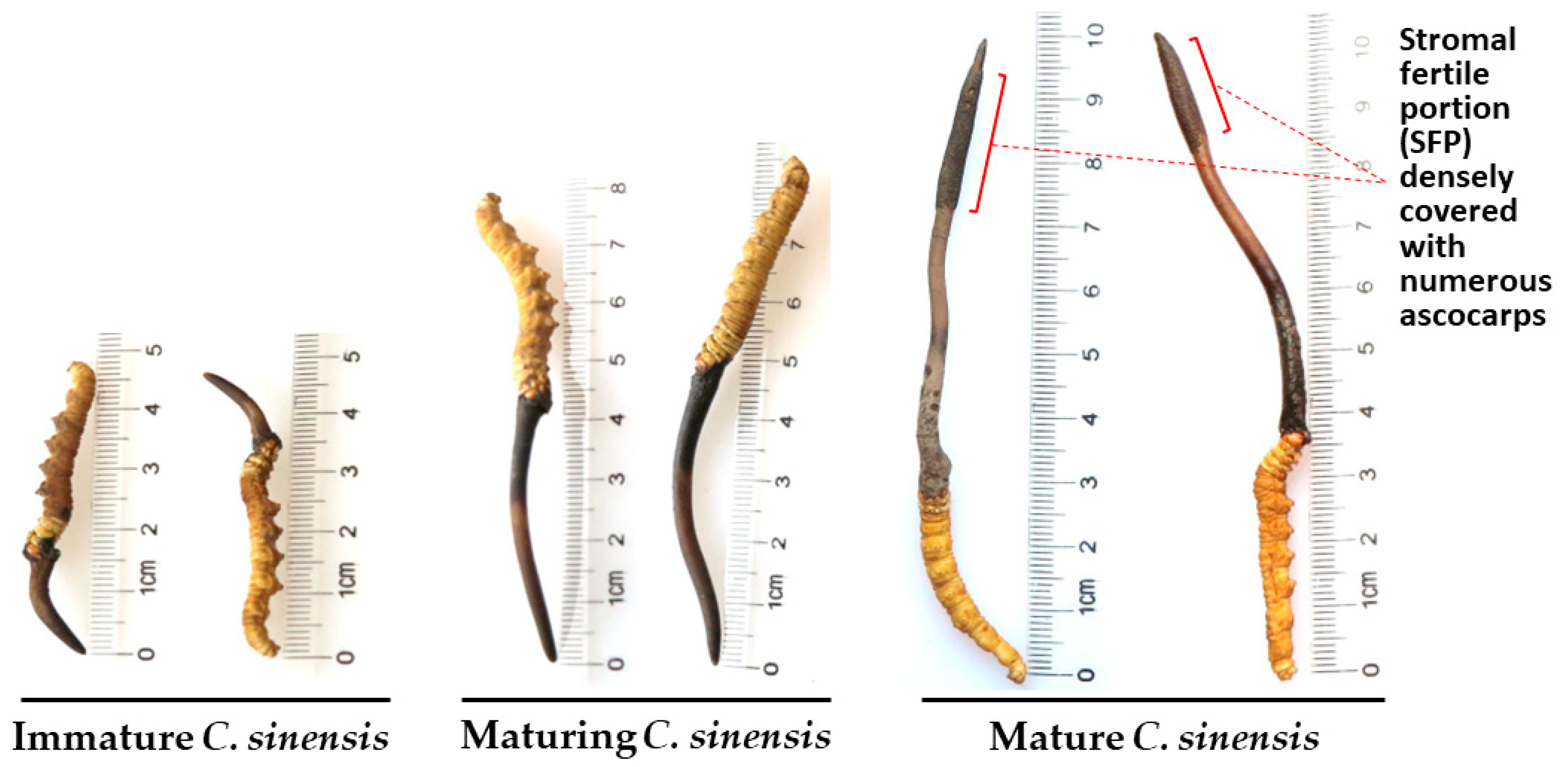
Figure 2.
Alignment of the MAT1-2-1 gene sequences with the transcriptome sequences obtained from H. sinensis Strain L0106 and natural C. sinensis. JQ325153, FJ654187, JQ325237 and KM197536 are the MAT1-2-1 gene sequences were found in H. sinensis Strains GS09_121, CS2, XZ12_16, and XZ12_16, respectively [Zhang et al. 2009, 2014]. GCQL01020543 is the transcriptome sequence obtained from H. sinensis Strain L0106 [Liu et al. 2015]. OSIN7649 is the metatranscriptome sequence obtained from natural C. sinensis collected from Deqin County, Yunnan Province, China [Xia et al. 2017]. The triplets shown in alternating brown and green indicate the open reading frame. The sequences in pink represent introns, and the underlined triplets “TAA” and “TGA” shown in blue are stop codons. The protein sequence was translated from the transcript sequences. The nucleotide and amino acid residues in red represent point mutations and variants, respectively. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.
Figure 2.
Alignment of the MAT1-2-1 gene sequences with the transcriptome sequences obtained from H. sinensis Strain L0106 and natural C. sinensis. JQ325153, FJ654187, JQ325237 and KM197536 are the MAT1-2-1 gene sequences were found in H. sinensis Strains GS09_121, CS2, XZ12_16, and XZ12_16, respectively [Zhang et al. 2009, 2014]. GCQL01020543 is the transcriptome sequence obtained from H. sinensis Strain L0106 [Liu et al. 2015]. OSIN7649 is the metatranscriptome sequence obtained from natural C. sinensis collected from Deqin County, Yunnan Province, China [Xia et al. 2017]. The triplets shown in alternating brown and green indicate the open reading frame. The sequences in pink represent introns, and the underlined triplets “TAA” and “TGA” shown in blue are stop codons. The protein sequence was translated from the transcript sequences. The nucleotide and amino acid residues in red represent point mutations and variants, respectively. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.

Figure 3.
Alignment of the MAT1-1-1 genome sequences FC437356 and KR004003 of H. sinensis Strains CS68-2-1229 and GS09_225 and the transcriptome sequences of natural C. sinensis. FC437356 is the MAT1-1-1 gene of H. sinensis Strain CS68-2-1229 [2013]. The sequence segments in pink represent introns I and II. The metatranscriptome assemblies OSIN7648 and GAGW01008880 are the MAT1-1-1 transcripts of natural C. sinensis specimens collected from Deqin County of Yunnan Province and Kangding County of Sichuan Province, China, respectively [Xiang et al. 2014; Xia et al. 2017]. The triplets shown in alternating brown and green indicate the open reading frame. The underlined “TAG” triplets in blue represent stop codons. The protein sequence was translated from the transcript sequences. Amino acid residues in red represent variants. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.
Figure 3.
Alignment of the MAT1-1-1 genome sequences FC437356 and KR004003 of H. sinensis Strains CS68-2-1229 and GS09_225 and the transcriptome sequences of natural C. sinensis. FC437356 is the MAT1-1-1 gene of H. sinensis Strain CS68-2-1229 [2013]. The sequence segments in pink represent introns I and II. The metatranscriptome assemblies OSIN7648 and GAGW01008880 are the MAT1-1-1 transcripts of natural C. sinensis specimens collected from Deqin County of Yunnan Province and Kangding County of Sichuan Province, China, respectively [Xiang et al. 2014; Xia et al. 2017]. The triplets shown in alternating brown and green indicate the open reading frame. The underlined “TAG” triplets in blue represent stop codons. The protein sequence was translated from the transcript sequences. Amino acid residues in red represent variants. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.

Figure 4.
Alignment of the MAT1-1-1 protein sequences translated from the transcripts of H. sinensis strains and metatranscriptomes of natural C. sinensis. OSIN7648 and GAGW01008880 are MAT1-1-1 protein sequences derived from metatranscriptome sequences of natural C. sinensis collected from Deqin County of Yunnan Province and Kangding County of Sichuan Province in China, respectively [Xiang et al. 2014; Xia et al. 2017]. AGW27560, ALH24995, ALH24951, ALH24992, ALH25047, ALH24948, ALH25003, ALH25005, ALH25057, ALH25001, ALH25048 and ALH24999 are the MAT1-1-1 protein sequences of Strains 1229, SC09_97, GS09_229, SC09_65, YN09_61, GS09_143, XZ05_6, XZ05_8, XZ12_16, XZ05_2, YN09_64 and XZ07_H2, respectively [Bushley et al. 2013; Zhang & Zhang 2015]. The residues in red indicate the variant amino acids. Hyphens indicate identical amino acid residues, and spaces denote unmatched sequence gaps.
Figure 4.
Alignment of the MAT1-1-1 protein sequences translated from the transcripts of H. sinensis strains and metatranscriptomes of natural C. sinensis. OSIN7648 and GAGW01008880 are MAT1-1-1 protein sequences derived from metatranscriptome sequences of natural C. sinensis collected from Deqin County of Yunnan Province and Kangding County of Sichuan Province in China, respectively [Xiang et al. 2014; Xia et al. 2017]. AGW27560, ALH24995, ALH24951, ALH24992, ALH25047, ALH24948, ALH25003, ALH25005, ALH25057, ALH25001, ALH25048 and ALH24999 are the MAT1-1-1 protein sequences of Strains 1229, SC09_97, GS09_229, SC09_65, YN09_61, GS09_143, XZ05_6, XZ05_8, XZ12_16, XZ05_2, YN09_64 and XZ07_H2, respectively [Bushley et al. 2013; Zhang & Zhang 2015]. The residues in red indicate the variant amino acids. Hyphens indicate identical amino acid residues, and spaces denote unmatched sequence gaps.

Figure 5.
ExPASy ProtScale plots for the MAT1-1-1 protein α-Helix (upper left panels), β-Sheet (upper right panels), β-Turn (lower left panels) and Coil (lower right panels). The MAT1-1-1 protein sequence AGW27560 (372 aa) of H. sinensis Strain CS68-2-1229 was compared with the metatranscriptome MAT1-1-1 sequences OSIN7648 (353 aa) and GAGW01008880 (276 aa) of natural C. sinensis. Open boxes in green indicate the variable segment regions of the MAT1-1-1 proteins, and open boxes in blue indicate the largely truncated N-terminal segment of the MAT1-1-1 protein GAGW01008880.
Figure 5.
ExPASy ProtScale plots for the MAT1-1-1 protein α-Helix (upper left panels), β-Sheet (upper right panels), β-Turn (lower left panels) and Coil (lower right panels). The MAT1-1-1 protein sequence AGW27560 (372 aa) of H. sinensis Strain CS68-2-1229 was compared with the metatranscriptome MAT1-1-1 sequences OSIN7648 (353 aa) and GAGW01008880 (276 aa) of natural C. sinensis. Open boxes in green indicate the variable segment regions of the MAT1-1-1 proteins, and open boxes in blue indicate the largely truncated N-terminal segment of the MAT1-1-1 protein GAGW01008880.
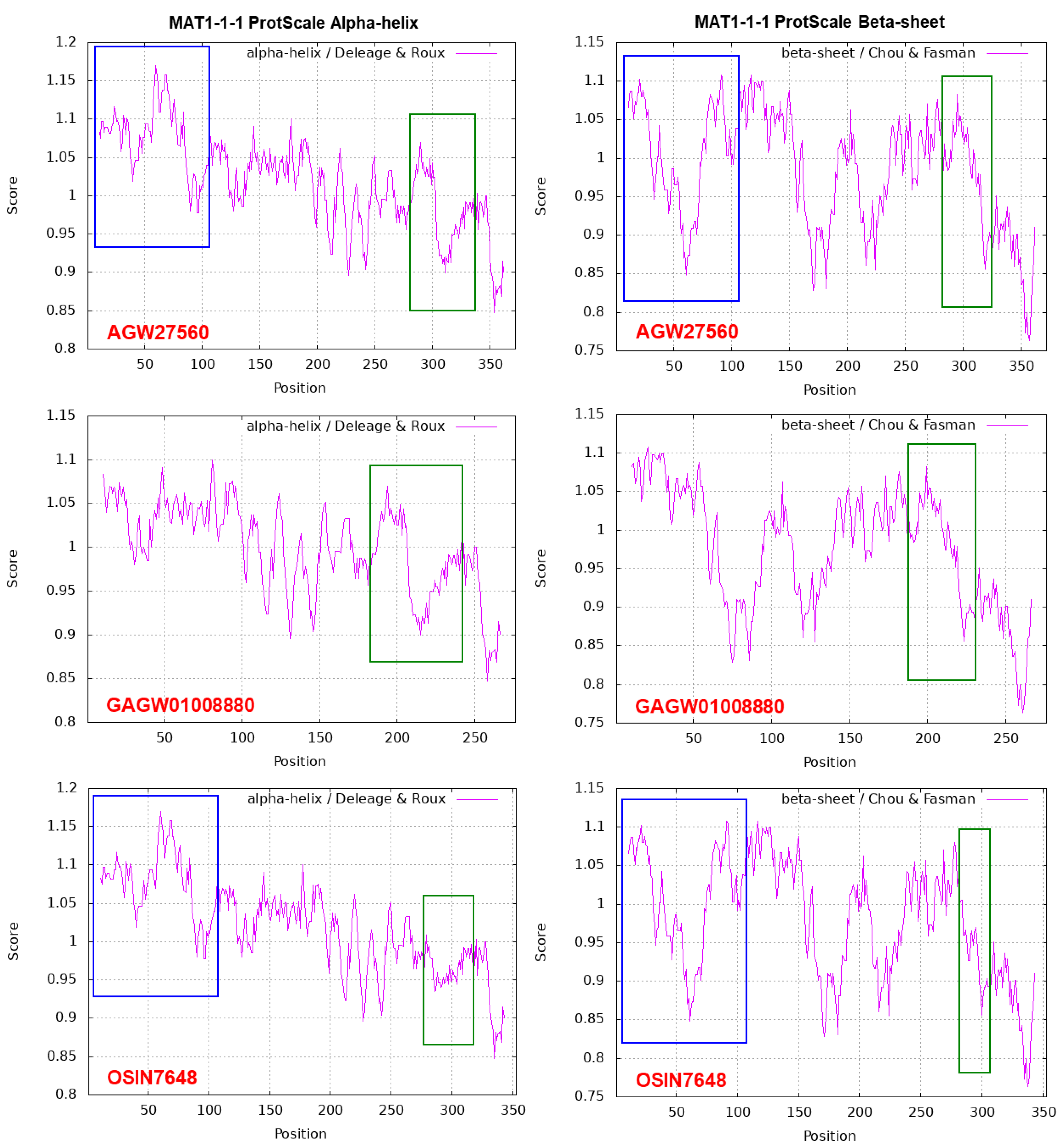
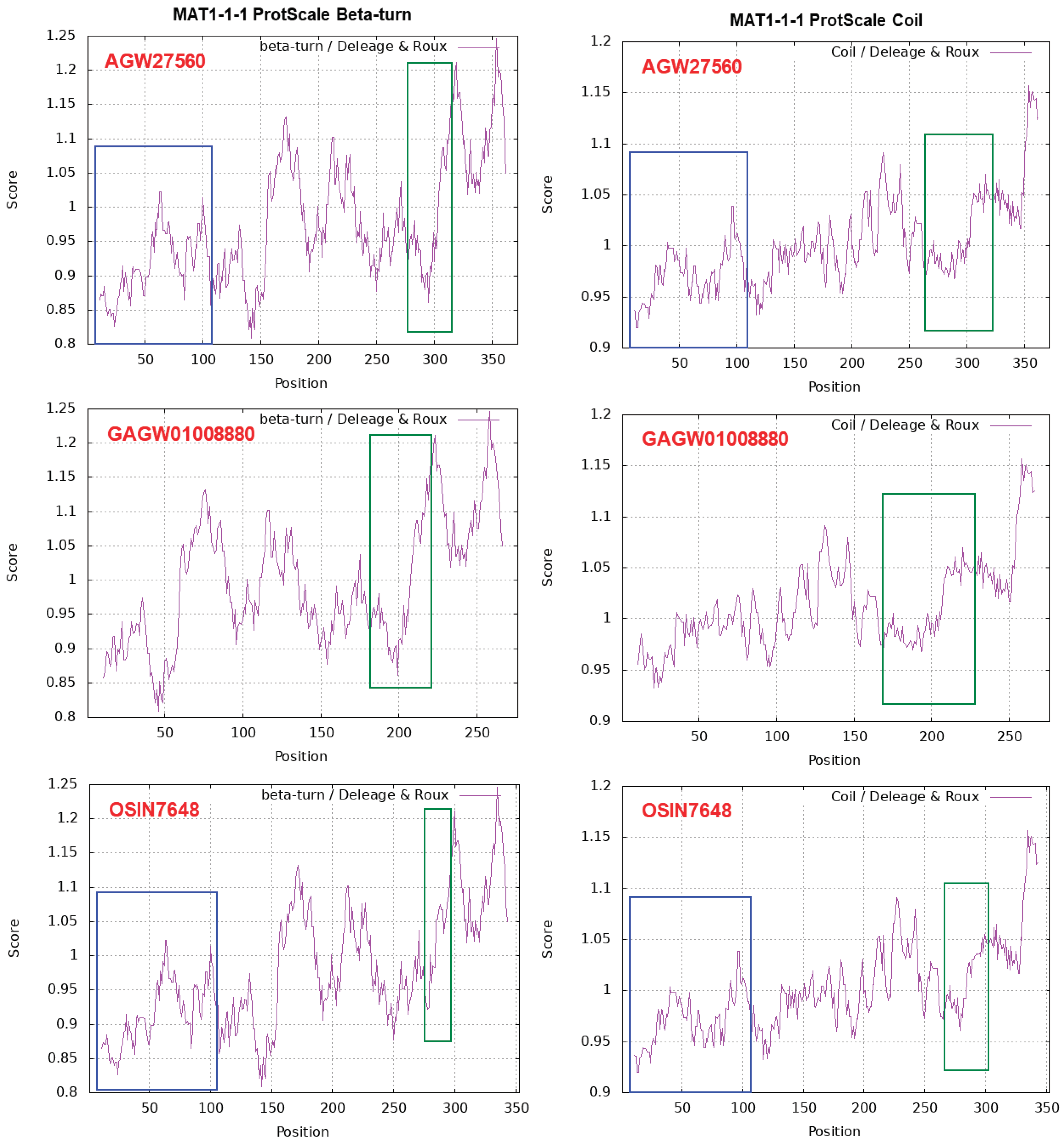
Figure 6.
Alignment of the MAT1-2-1 protein sequences translated from transcript sequences of H. sinensis strains and the metatranscriptome sequence of natural C. sinensis. OSIN7649 is the MAT1-2-1 protein sequence derived from the metatranscriptome sequence of natural C. sinensis collected from Deqin County of Yunnan Province, China [Xia et al. 2017]. AFX66389, AFX66415, ACV60375, ACV60372, AIV43040, AFX66472, AFX66473, AFX66474, AFH35020, ACV60417, ACV60418 and AFX66443 are the MAT1-2-1 protein sequences of H. sinensis Strains GS09_121, QH09_201, XZ-SN-44, XZ-LZ06-61, XZ12_16, YN09_6, YN09_22, YN09_51, XZ06-124, XZ-LZ07-H1, XZ-LZ07-H2 and XZ05_8, respectively [Zhang et al. 2009, 2011, 2014; Bushley et al. 2013]. The residues in red indicate the variant amino acids, and hyphens indicate identical amino acid residues.
Figure 6.
Alignment of the MAT1-2-1 protein sequences translated from transcript sequences of H. sinensis strains and the metatranscriptome sequence of natural C. sinensis. OSIN7649 is the MAT1-2-1 protein sequence derived from the metatranscriptome sequence of natural C. sinensis collected from Deqin County of Yunnan Province, China [Xia et al. 2017]. AFX66389, AFX66415, ACV60375, ACV60372, AIV43040, AFX66472, AFX66473, AFX66474, AFH35020, ACV60417, ACV60418 and AFX66443 are the MAT1-2-1 protein sequences of H. sinensis Strains GS09_121, QH09_201, XZ-SN-44, XZ-LZ06-61, XZ12_16, YN09_6, YN09_22, YN09_51, XZ06-124, XZ-LZ07-H1, XZ-LZ07-H2 and XZ05_8, respectively [Zhang et al. 2009, 2011, 2014; Bushley et al. 2013]. The residues in red indicate the variant amino acids, and hyphens indicate identical amino acid residues.

Figure 7.
Alignment of the genome and transcriptome sequences of a-pheromone receptor (PRE-1) from H. sinensis strains and natural C. sinensis. KE659055 is the gene sequence of H. sinensis Strains Co18 [Hu et al. 2013]. The genome assemblies NGJJ01000446, LKHE01002330, ANOV01020521, JAAVMX010000007, and LWBQ01000004 were obtained from H. sinensis strains CC1406-203, 1229, Co18, IOZ07, and ZJB12195, respectively [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Jiu et al. 2020; Shu et al. 2020]. OSIN6252 and GAGW01004735/GAGW01004736 are the transcriptome assembly sequences obtained from natural C. sinensis specimens collected from Deqin County of Yunnan Province and Kangding County of Sichuan Province of China, respectively [Xiang et al. 2014; Xia et al. 2017]. The segments in red are introns I and II. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.
Figure 7.
Alignment of the genome and transcriptome sequences of a-pheromone receptor (PRE-1) from H. sinensis strains and natural C. sinensis. KE659055 is the gene sequence of H. sinensis Strains Co18 [Hu et al. 2013]. The genome assemblies NGJJ01000446, LKHE01002330, ANOV01020521, JAAVMX010000007, and LWBQ01000004 were obtained from H. sinensis strains CC1406-203, 1229, Co18, IOZ07, and ZJB12195, respectively [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Jiu et al. 2020; Shu et al. 2020]. OSIN6252 and GAGW01004735/GAGW01004736 are the transcriptome assembly sequences obtained from natural C. sinensis specimens collected from Deqin County of Yunnan Province and Kangding County of Sichuan Province of China, respectively [Xiang et al. 2014; Xia et al. 2017]. The segments in red are introns I and II. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.

Figure 8.
Alignment of the translated protein sequences of a-pheromone receptor (PRE-1) from H. sinensis strains and natural C. sinensis. The PRE-1 protein sequences EQK97482 and KAF4506800 are translated from the gene sequence KE659055 of H. sinensis strain Co18 and the genome segment sequence JAAVMX010000007 of H. sinensis strain IOZ07 [Hu et al. 2013; Shu et al. 2020]. The PRE-1 transcriptome assembly sequences OSIN6252, GAGW01004735 and GAGW01004736 of natural C. sinensis are translated into protein sequences [Xiang et al. 2014; Liu et al. 2015]. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.
Figure 8.
Alignment of the translated protein sequences of a-pheromone receptor (PRE-1) from H. sinensis strains and natural C. sinensis. The PRE-1 protein sequences EQK97482 and KAF4506800 are translated from the gene sequence KE659055 of H. sinensis strain Co18 and the genome segment sequence JAAVMX010000007 of H. sinensis strain IOZ07 [Hu et al. 2013; Shu et al. 2020]. The PRE-1 transcriptome assembly sequences OSIN6252, GAGW01004735 and GAGW01004736 of natural C. sinensis are translated into protein sequences [Xiang et al. 2014; Liu et al. 2015]. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.

Figure 9.
Alignment of the genome and transcriptome sequences of the α-pheromone receptor of H. sinensis and natural C. sinensis. The genome sequence of KE653642 was obtained from H. sinensis Strain Co18 [Hu et al. 2013]. JAAVMX010000012, NGJJ01001310, LWBQ0100044, ANOV01006352, and LKHE01000580 are genome assemblies obtained from H. sinensis strains IOZ07, CC1406-203, ZJB12195, Co18, and 1229, respectively [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Jiu et al. 2020; Shu et al. 2020]. GCQL01007648, GCQL01017756, GCQL01015779, and OSIN6424 are the transcriptome assemblies obtained from H. sinensis strain L0106 and natural C. sinensis specimens collected from Deqin County of Yunnan Province, China [Liu et al. 2015; Xia et al. 2017]. The triplets shown in alternating brown and blue indicate the open reading frame. The sequences in pink indicate introns I and II between nucleotides 3,092→3,240 and 3,528→3,586, respectively, of the α-pheromone receptor gene KE653642, and the underlined “TAG” triplets shown in red are stop codons. The protein sequence was translated from the transcript sequences. The amino acid residues in red represent variants. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.
Figure 9.
Alignment of the genome and transcriptome sequences of the α-pheromone receptor of H. sinensis and natural C. sinensis. The genome sequence of KE653642 was obtained from H. sinensis Strain Co18 [Hu et al. 2013]. JAAVMX010000012, NGJJ01001310, LWBQ0100044, ANOV01006352, and LKHE01000580 are genome assemblies obtained from H. sinensis strains IOZ07, CC1406-203, ZJB12195, Co18, and 1229, respectively [Hu et al. 2013; Li et al. 2016a; Jin et al. 2020; Jiu et al. 2020; Shu et al. 2020]. GCQL01007648, GCQL01017756, GCQL01015779, and OSIN6424 are the transcriptome assemblies obtained from H. sinensis strain L0106 and natural C. sinensis specimens collected from Deqin County of Yunnan Province, China [Liu et al. 2015; Xia et al. 2017]. The triplets shown in alternating brown and blue indicate the open reading frame. The sequences in pink indicate introns I and II between nucleotides 3,092→3,240 and 3,528→3,586, respectively, of the α-pheromone receptor gene KE653642, and the underlined “TAG” triplets shown in red are stop codons. The protein sequence was translated from the transcript sequences. The amino acid residues in red represent variants. Hyphens indicate identical bases, and spaces denote unmatched sequence gaps.

Figure 10.
Alignment of the protein sequences of the α-pheromone receptor of H. sinensis strain Co18 and the transcriptome sequences of H. sinensis strain L0106 and natural C. sinensis. The gene sequence EQK99119 was obtained from H. sinensis strains Co18 [Hu et al. 2013]. The transcriptome assemblies GCQL01007648, GCQL01017756, GCQL01015779 and OSIN6424 were obtained from H. sinensis strain L0106 and natural C. sinensis specimens collected from Deqin County of Yunnan Province, China [Xiang et al. 2014; Xia et al. 2017]. Hyphens indicate identical amino acid residues, and spaces denote unmatched sequence gaps.
Figure 10.
Alignment of the protein sequences of the α-pheromone receptor of H. sinensis strain Co18 and the transcriptome sequences of H. sinensis strain L0106 and natural C. sinensis. The gene sequence EQK99119 was obtained from H. sinensis strains Co18 [Hu et al. 2013]. The transcriptome assemblies GCQL01007648, GCQL01017756, GCQL01015779 and OSIN6424 were obtained from H. sinensis strain L0106 and natural C. sinensis specimens collected from Deqin County of Yunnan Province, China [Xiang et al. 2014; Xia et al. 2017]. Hyphens indicate identical amino acid residues, and spaces denote unmatched sequence gaps.

Figure 11.
ExPASy ProtScale plots for the α-Helix (upper-left panels), β-Sheet (upper-right panels), β-Turn (lower-left panels) and Coil (lower-right panels) of α-pheromone receptor proteins. The α-pheromone receptor protein sequence EQK99119 (390 aa) of H. sinensis Strain Co18 was compared with the transcriptome and metatranscriptome α-pheromone receptor sequences and GCQL01017756 (301 aa) and GCQL01015779 (301 aa) of H. sinensis Strain L0106 and OSIN6424 (408 aa) of natural C. sinensis. Open boxes in green indicate the variable segment regions of the α-pheromone receptor proteins, and open boxes in blue indicate the largely truncated N-terminal segment of the α-pheromone receptor proteins GCQL01017756 and GCQL01015779.
Figure 11.
ExPASy ProtScale plots for the α-Helix (upper-left panels), β-Sheet (upper-right panels), β-Turn (lower-left panels) and Coil (lower-right panels) of α-pheromone receptor proteins. The α-pheromone receptor protein sequence EQK99119 (390 aa) of H. sinensis Strain Co18 was compared with the transcriptome and metatranscriptome α-pheromone receptor sequences and GCQL01017756 (301 aa) and GCQL01015779 (301 aa) of H. sinensis Strain L0106 and OSIN6424 (408 aa) of natural C. sinensis. Open boxes in green indicate the variable segment regions of the α-pheromone receptor proteins, and open boxes in blue indicate the largely truncated N-terminal segment of the α-pheromone receptor proteins GCQL01017756 and GCQL01015779.
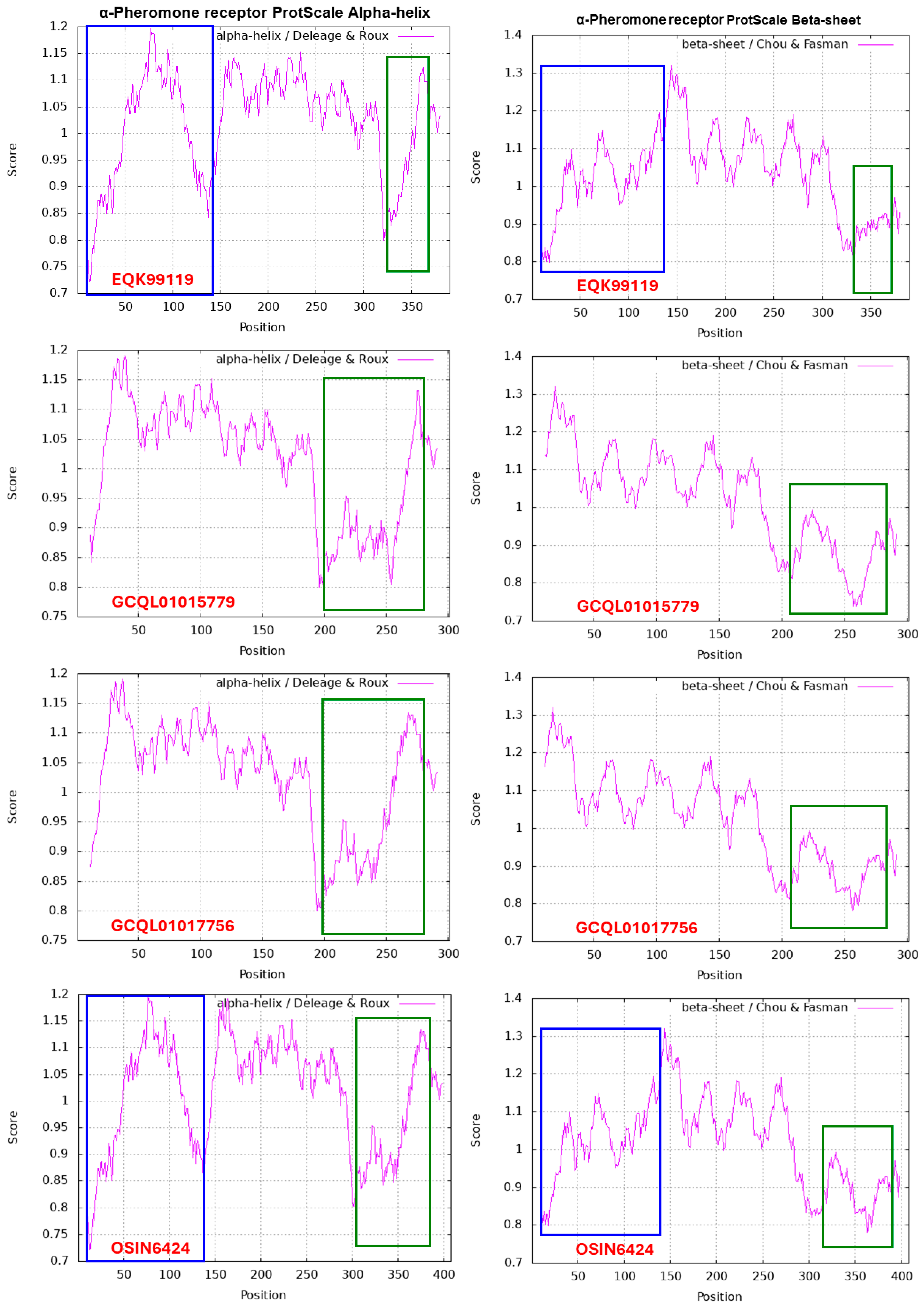
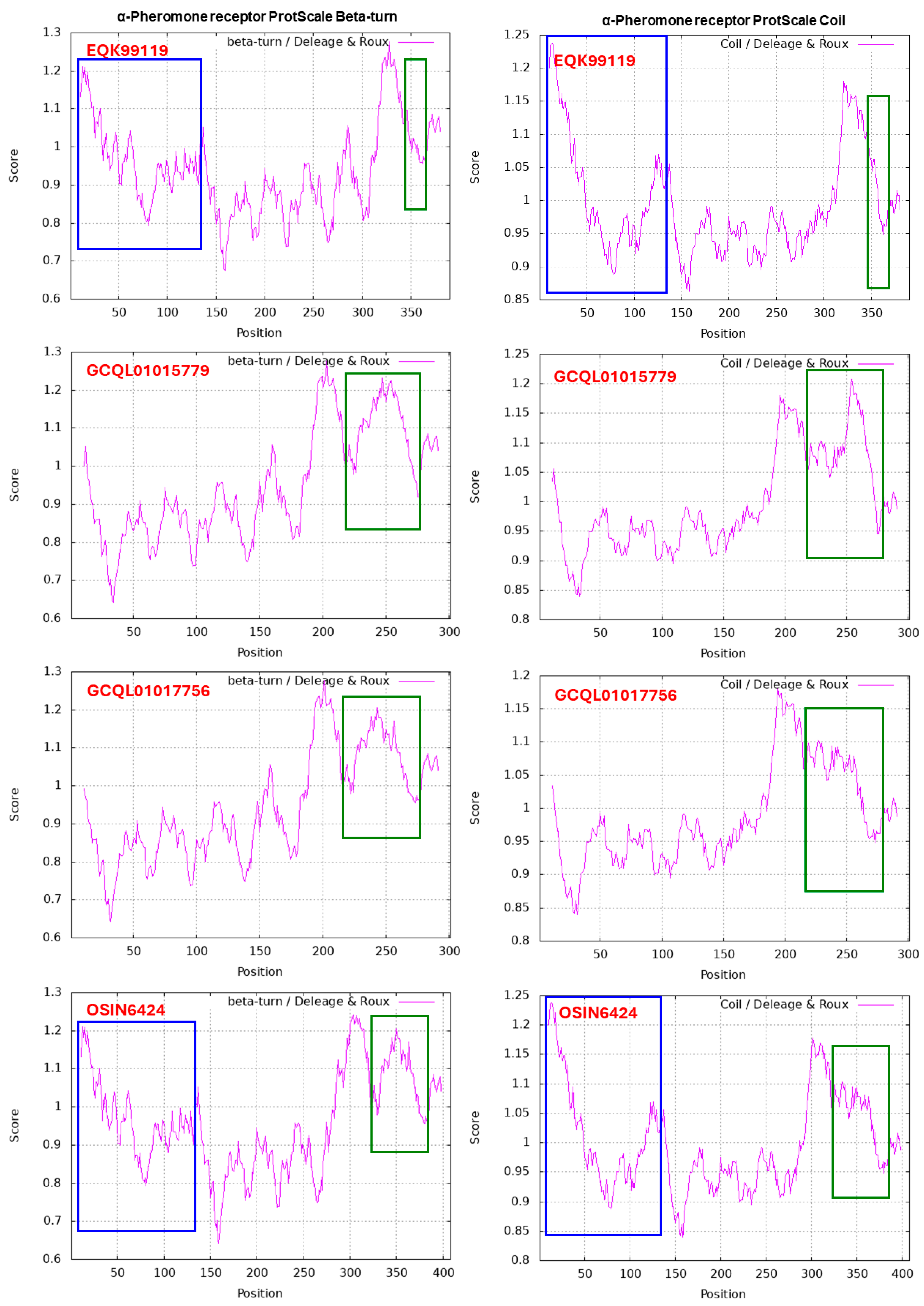
Figure 12.
Dynamic alterations in the abundance of the amplicons of GC-biased Genotypes #1 and #2 of O. sinensis in the stromata of natural C. sinensis during maturation (modified from Figure 6 of [Zhu et al. 2012]).
Figure 12.
Dynamic alterations in the abundance of the amplicons of GC-biased Genotypes #1 and #2 of O. sinensis in the stromata of natural C. sinensis during maturation (modified from Figure 6 of [Zhu et al. 2012]).
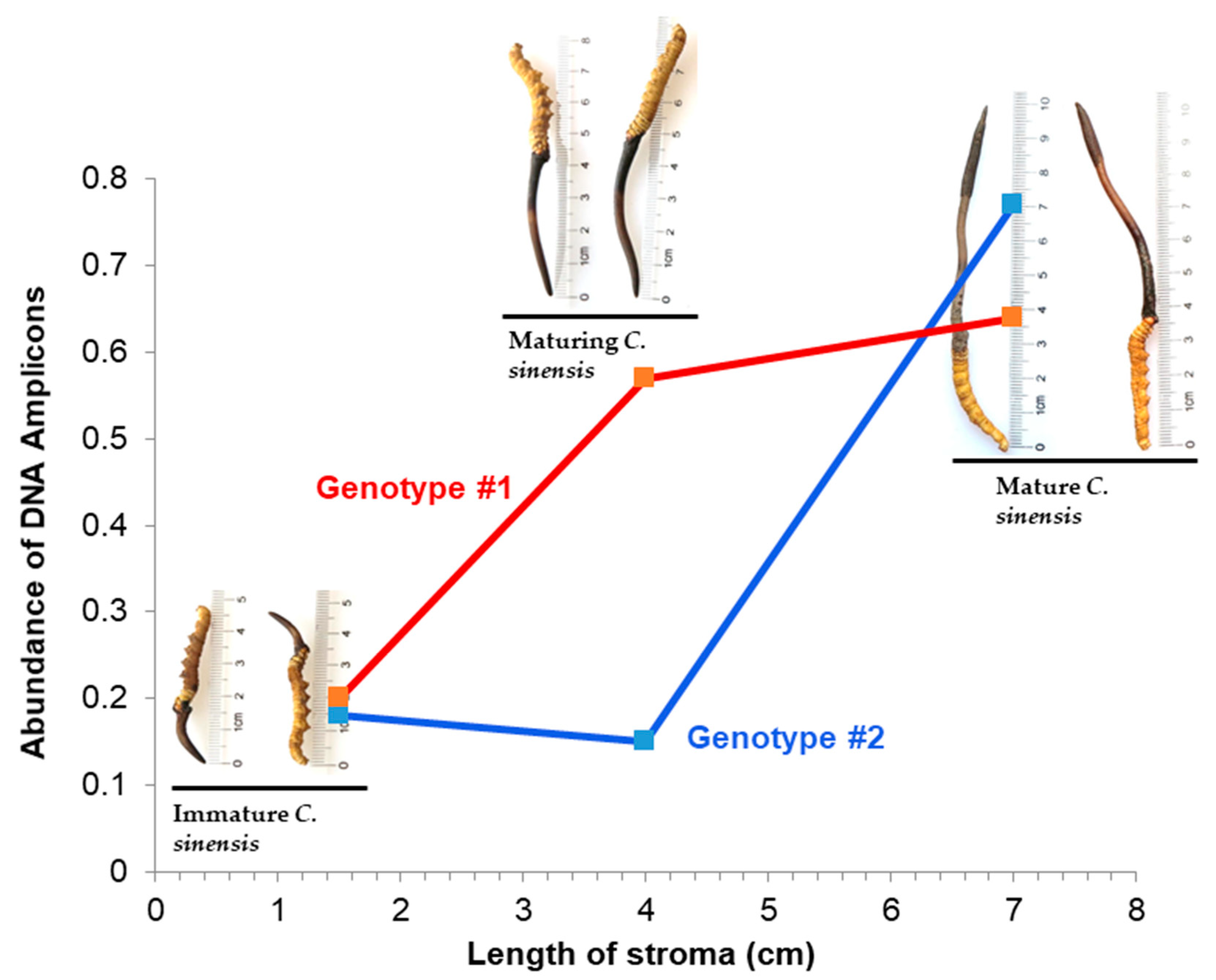
Table 1.
Differential occurrence of the MAT1-1-1 and MAT1-2-1 genes in 237 H. sinensis strains.
| 22 strains | 66 strains | 149 strains | ||||
|---|---|---|---|---|---|---|
| containing only the MAT1-1-1 gene | containing only the MAT1-2-1 gene | containing both the MAT1-1-1 and MAT1-2-1 genes | ||||
| CS09-143 | CC1406-203 | XZ-CD-41 | 1229 | NP10_1 | SC09_47 | XZ09_32 |
| CS09-229 | CS2 | XZ-CD-59 | Co18 | NP10_2 | SC09_57 | XZ09_46 |
| GS03 | GS09_337 | XZ-CD-64 | CS6-251 | QH01 | SC09_65 | XZ09_48 |
| IOZ07 | QH02 | XZ-ML-191 | CS18-266 | QH03 | SC09_77 | XZ09_59 |
| QH07_188 | QH05 | XZ-LZ05-6 | CS25-273 | QH04 | SC09_87 | XZ09_71 |
| QH07_197 | QH09_11 | XZ-LZ06_1 | CS26-277 | QH06 | SC09_97 | XZ09_80 |
| XZ05_3 | QH09_187 | XZ-LZ06_7 | CS34-291 | QH07 | SC09_107 | XZ09_95 |
| XZ05_6 | QH-LJ-214 | XZ-LZ06_21 | CS36-1294 | QH08 | SC09_117 | XZ09_100 |
| XZ07_H2 | QH-LJ-236 | XZ-LZ06_61 | CS37-295 | QH09_20L | SC09_128 | XZ09_106 |
| XZ07_108 | QH-QL-206 | XZ-LZ06_108 | CS68-2-1228 | QH09_33L | SC09_147 | XZ09_113 |
| XZ07_154 | QH-QL-207 | XZ-LZ07_30 | CS68-2-1229 | QH09_37 | SC09_157 | XZ09_118 |
| XZ07_166 | QH-YS-188 | XZ-LZ07_64 | CS68-5-1216 | QH09_46 | SC09_167 | XZ10_7 |
| XZ07_176 | QH-YS-189 | XZ-LZ07_H1 | CS70-1208 | QH09_56 | SC09_180 | XZ10_15 |
| XZ07_180 | QH-YS-196 | XZ-LZ07_H2 | CS70-1211 | QH09_66 | SC09_190 | XZ10_17 |
| XZ08_4 | QH-YS-197A | XZ-NQ_74 | CS70-1212 | QH09_78 | SC09_200 | XZ10_23 |
| XZ08_10 | QH-YS-197B | XZ-NQ_80 | CS71-1218 | QH09_93 | SC10_4 | XZ12_1 |
| XZ08_26 | QH-YS-199 | XZ-NQ_84 | CS71-1219 | QH09_122 | SC10_18 | XZ12_16 |
| XZ08_59 | SC-2 | XZ-NQ_86 | CS71-1220 | QH09_131 | SC10_21 | XZ12_33 |
| XZ08_A1 | SC-3 | XZ-NQ_92 | CS76-1284 | QH09_151 | TB01 | XZ12_43 |
| XZ08_B1 | SC-4 | XZ-NQ_139 | CS91-1291 | QH09_164 | TB02 | YN01 |
| YN07_6 | SC-5 | XZ-NQ_154 | CS560-961 | QH09_173 | TB03 | YN02 |
| YN07_8 | SC-7 | XZ-NQ_155 | CS561-964 | QH09_201 | TB04 | YN03 |
| SCK05-4-3 | XZ-NQ_156 | GS01 | QH09_210 | TB05 | YN09_3 | |
| XZ07_11 | XZ-NQ_166 | GS02 | QH10_1 | TB06 | YN09_6 | |
| XZ07_46 | XZ-NQ_176 | GS04 | QH10_4 | TB07 | YN09_22 | |
| XZ08_33 | XZ-NQ_180 | GS05 | QH10_7 | TB08 | YN09_51 | |
| XZ08_38 | XZ-SN_44 | GS09_111 | SC01 | XZ05_2 | YN09_61 | |
| XZ-CD-A1 | YN-1 | GS09_121 | SC02 | XZ05_7 | YN09_64 | |
| XZ-CD-B1 | YN-4 | GS09_131 | SC03 | XZ05_8 | YN09_72 | |
| XZ-CD-4 | YN-5 | GS09_143 | SC04 | XZ05_12 | YN09_81 | |
| XZ-CD-10 | YN-6 | GS09_201 | SC05 | XZ06_124 | YN09_85 | |
| XZ-CD-26 | YN-8 | GS09_225 | SC06 | XZ06_152 | YN09_89 | |
| XZ-CD-30 | ZJB12195 | GS09_229 | SC07 | XZ06_260 | YN09_96 | |
| GS09_281 | SC08 | XZ07_133 | YN09_101 | |||
| GS09_311 | SC09_1 | XZ08_24 | YN09_140 | |||
| GS10_1 | SC09_21 | XZ08_56 | ||||
| GS10_10 | SC09_36 | XZ09_4 | ||||
| ID10_1 | SC09_37 | XZ09_15 | ||||
Table 2.
Amino acid properties and scale: Conformational parameters for α-Helix, β-Sheet, and Coil [Deleage & Roux 1987] and β-Sheet [Chou & Fasman 1978].
Table 2.
Amino acid properties and scale: Conformational parameters for α-Helix, β-Sheet, and Coil [Deleage & Roux 1987] and β-Sheet [Chou & Fasman 1978].
| Chemical-physical property | α-Helix | β-Sheet | β-Turn | Coil | |||
|---|---|---|---|---|---|---|---|
| Aspartic acid | Asp | D | Acidic | 0.924 | 0.540 | 1.197 | 1.197 |
| Glutamic acid | Glu | E | Acidic | 1.504 | 0.370 | 1.149 | 0.761 |
| Alanine | Ala | A | Aliphatic | 1.489 | 0.830 | 0.788 | 0.824 |
| Isoleucine | Ile | I | Aliphatic | 1.003 | 1.600 | 0.240 | 0.886 |
| Leucine | Leu | L | Aliphatic | 1.236 | 1.300 | 0.670 | 0.810 |
| Valine | Val | V | Aliphatic | 0.990 | 1.700 | 0.387 | 0.772 |
| Phenylalanine | Phe | F | Aromatic | 1.195 | 1.380 | 0.624 | 0.797 |
| Tryptophan | Trp | W | Aromatic | 1.090 | 1.370 | 0.546 | 0.941 |
| Tyrosine | Tyr | Y | Aromatic | 0.787 | 1.470 | 0.795 | 1.109 |
| Arginine | Arg | R | Basic | 1.224 | 0.930 | 0.912 | 0.893 |
| Histidine | His | H | Basic | 1.003 | 0.870 | 0.970 | 1.068 |
| Lysine | Lys | K | Basic | 1.172 | 0.740 | 1.302 | 0.897 |
| Asparagine | Asn | N | with polar neutral side chains | 0.772 | 0.890 | 1.572 | 1.167 |
| Cysteine | Cys | C | with polar neutral side chains | 0.966 | 1.190 | 0.965 | 0.953 |
| Glutamine | Gln | Q | with polar neutral side chains | 1.164 | 1.100 | 0.997 | 0.947 |
| Methionine | Met | M | with polar neutral side chains | 1.363 | 1.050 | 0.436 | 0.810 |
| Serine | Ser | S | with polar neutral side chains | 0.739 | 0.750 | 1.316 | 1.130 |
| Threonine | Thr | T | with polar neutral side chains | 0.785 | 1.190 | 0.739 | 1.148 |
| Glycine | Gly | G | Unique amino acids | 0.510 | 0.750 | 1.860 | 1.251 |
| Proline | Pro | P | Unique amino acids | 0.492 | 0.550 | 1.415 | 1.540 |
Table 3.
Percent similarity between the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs and the genome and transcriptome sequences of H. sinensis and natural C. sinensis.
Table 3.
Percent similarity between the mating-type genes of the MAT1-1 and MAT1-2 idiomorphs and the genome and transcriptome sequences of H. sinensis and natural C. sinensis.
| MAT1-1-1 | MAT1-1-2 | MAT1-1-3 | MAT1-2-1 | |
|---|---|---|---|---|
| (vs. KC437356 of Strain CS68-2-1229) | (vs. JQ325153 of Strain GS09_121) | |||
| % similarity of the genome sequences | ||||
| H. sinensis Strain 1229 | 100% | 100% | 100% | 99.9% |
| H. sinensis Strain Co18 | 99.9% | 100% | 100% | 99.7% |
| H. sinensis Strain IOZ07 | 100% | 100% | 100% | ― |
| H. sinensis Strain CC1406-203 | ― | ― | ― | 99.9% |
| H. sinensis Strain ZJB12195 | ― | ― | ― | 99.9% |
|
% similarity of the coding sequences of the genes (excluding introns) and the transcriptome sequences |
||||
| H. sinensis Strain L0106 | ― | ― | ― | 99.0% |
| H. sinensis Strain 1229 | 100% | 100% | 100% | 100% |
| Natural C. sinensis (Deqin, Yunnan) | 100% | ― | ― | 100% |
| Natural C. sinensis (Kangding, Sichuan) | 100% | ― | ― | ― |
Table 4.
Percentage of amino acids in the MAT1-1-1 and MAT1-2-1 proteins of H. sinensis and natural C. sinensis.
Table 4.
Percentage of amino acids in the MAT1-1-1 and MAT1-2-1 proteins of H. sinensis and natural C. sinensis.
| % Amino acids of mating-type protein | |||||
|---|---|---|---|---|---|
| Hydrophobic | Acidic | Basic | Neutral | ||
| MAT1-1-1 protein | |||||
| AGW27560 | H. sinensis Strain CS68-2-1229 | 41.4% | 13.2% | 11.0% | 34.4% |
| GAGW01008880 | Natural C. sinensis | 41.3% | 14.1% | 9.8% | 34.8% |
| OSIN7648 | Natural C. sinensis | 41.9% | 13.1% | 11.1% | 33.7% |
| MAT1-2-1 protein | |||||
| AFX66389 | H. sinensis Strain GS09_121 | 41.2% | 10.0% | 16.5% | 33.3% |
| OSIN7649 | Natural C. sinensis | 41.2% | 10.0% | 16.5% | 33.3% |
Table 5.
Percentage of similarity between the protein sequences of pheromone receptors and the genome and transcriptome sequences of H. sinensis and natural C. sinensis.
Table 5.
Percentage of similarity between the protein sequences of pheromone receptors and the genome and transcriptome sequences of H. sinensis and natural C. sinensis.
| Percent similarity | ||
|---|---|---|
| a-Factor-like pheromone receptor | α-Factor-like pheromone receptor | |
| Between the sequences of pheromone receptor genes and the H. sinensis genomes | ||
| H. sinensis Strain 1229 | 100% | 99.8% |
| H. sinensis Strain CC1406-203 | 100% | 99.8% |
| H. sinensis Strain Co18 | 100% | 100% |
| H. sinensis Strain IOZ07 | 100% | 99.9% |
| H. sinensis Strain ZJB12195 | 95.2% | 97.5% |
| Between the coding sequences of the pheromone receptor genes (excluding introns) and the transcriptome sequences of H. sinensis and natural C. sinensis | ||
| H. sinensis Strain L0106 | ― | 99.3% |
| Natural C. sinensis (Deqin, Yunnan) | 100% | 99.7% |
| Natural C. sinensis (Kangding, Sichuan) | 98.5‒100% | ― |
| Between the pheromone receptor protein sequences translated from the gene sequences and the transcript sequences of H. sinensis and natural C. sinensis | ||
| H. sinensis Strain L0106 | ― | 86.4‒100% |
| Natural C. sinensis (Deqin, Yunnan) | 100% | 82.9% |
| Natural C. sinensis (Kangding, Sichuan) | 97.8‒100% | ― |
Table 6.
Percentages of amino acids of the pheromone receptor proteins of H. sinensis and natural C. sinensis.
Table 6.
Percentages of amino acids of the pheromone receptor proteins of H. sinensis and natural C. sinensis.
| % Amino acids of pheromone receptor protein | |||||
|---|---|---|---|---|---|
| Hydrophobic | Acidic | Basic | Neutral | ||
| a-pheromone receptor protein | |||||
| EQK97482 | H. sinensis Strain Co18 | 50.7% | 6.5% | 11.6% | 31.4% |
| GAGW01004735-GAGW01004736 | Natural C. sinensis | 50.4% | 6.2% | 11.2% | 31.2% |
| OSIN6252 | Natural C. sinensis | 50.7% | 6.5% | 11.6% | 31.4% |
| α-pheromone receptor protein | |||||
| EQK99119 | H. sinensis Strain Co18 | 55.1% | 5.6% | 7.4% | 31.8% |
| GCQL01017756 | H. sinensis Strain L0106 | 50.8% | 6.0% | 9.0% | 34.2% |
| GCQL01015779 | H. sinensis Strain L0106 | 51.8% | 6.0% | 9.0% | 33.2% |
| OSIN6424 | Natural C. sinensis | 52.7% | 5.8% | 8.6% | 32.8% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



