
Submitted:
01 April 2024
Posted:
01 April 2024
You are already at the latest version
Abstract
This article develops a multi-perspective view on motivations and methods for tobamovirus purification through the ages, and presents a novel efficient, easy-to-use approach that can be well-adapted to different species of native and functionalized virions.
We survey the various driving forces prompting researchers to enrich tobamoviruses: from the search for the causative agents of mosaic diseases in plants to their increasing recognition as versatile nanocarriers in biomedical and engineering applications. Best practice and also rarely applied options for the serial processing steps required for successful isolation of tobamoviruses are then reviewed. Adaptations for distinct particle species, pitfalls and 'forgotten' or underrepresented technologies are considered as well. The article is topped off with our own development of a method for virion preparation, rooted in historical protocols. It combines selective re-solubilization of polyethylene glycol (PEG) virion raw precipitates with density step gradient centrifugation in biocompatible iodixanol formulations, yielding ready-to-use particle suspensions. This newly established protocol and some considerations for perhaps worthwile further developments could serve as putative stepping stones towards preparation procedures appropriate for routine practical uses of the multivalent soft-matter nanorods.
Keywords:
tobacco mosaic virus (TMV)
; turnip vein clearing virus (TVCV)
; plant virus
; history
; purification
; isolation
; precipitation
; ultracentrifugation
; density gradients
; iodixanol
1. Introduction
How did the early researchers achieve the purification of virions from 'tobacco mosaic-diseased' plants until a fundamental understanding of the helical tobamovirus particle structure was reached? Have the initial protocols experienced a continuous 'evolution' and optimization throughout now 125 years of tobamovirus research, or have new technologies and scientific questions driven major changes in purification strategies? Which approaches and combinations thereof have ever been tested for the different serial processing steps, from leaf tissue homogenization up to formulation of the final virion preparations? And last but not least: Is there still room for further improvements with regard to current scientific challenges and emerging practical uses of the robust, multivalent nucleoprotein particles?
These questions have arisen from our involvement in both teaching and research, and have prompted us to work out a sketch map on tobamovirus purification strategies applied over time, in conjunction with a case study demonstrating the high potential of 'old', but topically refreshed methods for an efficient, customized isolation of engineered viruses with properties deviating from the parental ones. The principle of PEG raw precipitate re-solubilization in inverse PEG concentration gradients [1,2] has recently been taken up in our team for the fast and gentle enrichment of a tobacco vein clearing virus (TVCV) variant with S. aureus protein A-domains fused to all of its coat proteins (CPs; TVCVPA [3]). On this basis we have now implemented an advantageous, simplified procedure using iodixanol step gradients for selective virion banding, yielding stable ready-to-use particle suspensions, as described below. This article section might encourage further efforts and process developments that profit from historic achievements also in future research.
Our initial TVCVPA purification via the almost forgotten inverse PEG solubility-concentration gradients had been proposed and supported by Holger Jeske as scientific mentor, collaborator and friend. He also suggested its further streamlining, and has devoted extensive own labwork and many hours at the transmission electron microscope (EM) to the development of preparative and analytical techniques for novel types of tobacco mosaic virus (TMV)-based assemblies (e.g., [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22]). Well-known and highly active primarily in geminivirus research, Holger continuously raised awareness for the rarely opened treasure trove of early, but surprisingly sophisticated (bio-) chemical, physical and molecular methods in their historical contexts. He also appreciated and participated gladly in the establishment of our research focus on novel uses and in vitro derivatives of tobamovirus particles as a spin-off from his department in Stuttgart, for which reason we dedicate this article to the memory of him.
1.1. What Can (and Cannot) Be Expected from this Reading?
This article provides an overview of methods for isolating tobamoviruses from their hosts, which starts with the earliest experiments on tobacco showing striking leaf variegation, spot or 'mosaic' symptoms (Figure 1A). Already these initial works from the 1880s on attempted to separate the disease-inducing agent (detectable easily by modern microscopy now: Figure 1B-C) from plant sap - yet unsuccessful up to the 1930s. Notwithstanding, the era of 'fishing in increasingly clearer water' prepared the ground for viable virion purification methods. It is described with regard to technical and conceptual advances, and the first understanding of viruses as own class of biological entities (1.2). The subsequent section (1.3) spans from the first precipitation of infectious TMV paracrystals (Figure 1D-E)to the present. It firstly surveys the history of two decades, when different laboratories hunted for the compound called TMV, and eventually understood its helical organization from CP subunits protecting an RNA. This, finally, came out to be the genetic blueprint of the virus. Despite these fundamental conceptual achievements on the self-encoding but host-dependent tobamovirus structure, the methods of TMV purification had not changed substantially during the respective two decades, as described concisely. However, they enabled also the growth of further research areas on tobamoviruses, which - after mid of the 1950s - broadened fast and spread around all of the world (as the viruses did themselves). It is impossible to review all these scientific developments, so we have selected a few major upcoming fields of work and relate their objectives to typically employed virion isolation routes. Conversely, TMV has also been an ideal 'guinea pig' for testing and implementing novel purification procedures established in different contexts, which is considered as well. As novel applications of tobamoviruses in biomedicine and nanotechnology have brought along an increased need of purified particles, these developments of the last thirty years are worked out at greater length. The last introductory paragraphs (1.4) will outline the actual knowledge of biological and physicochemical particle properties and the diversity of distinct tobamoviruses, which has arisen through the ages and provides the keys to appropriate purification strategies.
Chapter 2 dives more deeply into the principles and application histories of major operations to access and store the desired whitish-opalescent colloidal tobamovirus sols. As purification protocols combine different procedures serially, a rough overview of relevant processing chains is given in Chapter 3, which also hints at potential pitfalls, a few 'neglected' and some peculiar TMV isolation approaches. Our case study on a beneficial, new combination of uncommon purification steps involving PEG precipitate re-solubilization in adapted, iodixanol-based density step gradients is presented as Chapter 4. It might add some more value on 'forgotten' methods as sources for new procedures, as introduced above. At last, an outlook in Chapter 5 dares a few further suggestions on potential future directions in tobamovirus nanoparticle production, which, however, are facing a trade-off between promising application prospects and current regulatory issues.
Originally, we set out to obtain a reliable survey of purification methods applied in different scientific contexts, labs, and time periods. The more we read, though, the more gaps in our reviewing approach became obvious so that despite a broad literature search, we will have missed relevant information. Nevertheless, more than 10,000 entries (out of a total of about ≈250,000 on the tobacco mosaic [virus] available in English or German in the Google Scholar database by Feb 2024) mentioning virion preparation by different wording, and existing collections in our teams have been the source for more than 600 handpicked articles as main starting material, covering 30 to 150 publications per decade from the 1930s on. These were sifted for tobamovirus purification methods, and complemented by articles on tobamoviruses other than TMV according to references in the International Committee on Taxonomy of Viruses (ICTV) database. Hardcopies available for selected non-digitalized historic papers as well as methodological reports and reviews were accessed in addition. This modus operandi could, however, not avoid bias due to the non-accessibility of various articles through our institutions, publications in languages other than English or German, missing or wrongly referenced descriptions of purification procedures, and unexpectedly many studies even on wildtype TMV not found via our search strings, because the virus name was not explicitly included in title, abstract or keywords. Our work has, however, carved out a multifaceted overview of the history of tobamovirus purification, which we hope to inspire many new ideas and debates - as it already did in our authors' team.
1.2. The Cryptic 'Poisonous' agent Causing Tobacco Mosaic Disease: Early 'Pre-Purification' Experiments towards Virus Separation
Experiments to isolate the agent causing 'tobacco mosaic disease' in the Netherlands may be regarded the key to the whole discipline of virology. After Adolf E. Mayer had discovered the transmissibility of the disease to previously healthy plants, and speculated about its origin already in the 1880s (as reviewed in detail [26]), attempts to separate the responsible agent from plant sap by Chamberland filter candles developed for water sterilization [27] were carried out in different labs. As the porous porcelain failed to remove infectivity, and since both cultivation and diffusion tests on agar plates did not indicate any 'corpuscular', bacterial pathogen, Martinus W. Beijerinck claimed the existence of a 'contagium vivum fluidum' inducing the 'tobacco spot disease' in 1898[28]. This was groundbreaking, because despite his misconception of a 'living fluid', Beijerinck proposed a completely new type of infectious 'virus' (i.e., 'poisonous' agent) that passed filtration pores small enough to retain microbes. His foresight of viruses as a novel class of biological agents was in clear contrast to earlier [29] and also subsequent conclusions of the Russian researcher Dmitri I. Ivanowski from similar filtration tests, who was convinced of a bacterial origin of the disease[as reviewed, e.g., in 26,30-32]. Hence, Beijerinck's study is now widely recognized as the foundation of virology[e.g., 33,34-37], the more, as it also stated 'that propagation results only when the virus is connected with the living and growing protoplasm of the host-plant' [28]. An understanding of viruses as biochemical, but non-living entities was proposed only a few years later by the German geneticist Erwin Baur[38], on the basis of his and earlier grafting experiments with ornamental, mosaic-diseased Abutilon plants (outlined in[39]). He hypothesized thereby transmissible 'viruses' to be substances produced and accumulated by the affected plant itself - a modern description valid up to now.
Approaches to isolate the infectious agent from infected leaves by filtration, cultivation on nutrient media, and passage into further plants have thus played central roles already in the earliest investigations on TMV and its relatives. To get clues about virus structure and composition, plant sap, homogenates and filtrates as well as soil from pots with diseased plants underwent microscopy and treatments with, e.g. heat, acid, glycerol, ethanol or formalin, before they were tested for residual infectivity[28,29,40,41]. Such experiments made up the majority of studies on tobacco and increasing numbers of other plant 'mosaic diseases' during the first three decades of the 20st century, once termed 'the no man's land before molecular biology got off the ground' by Lute Bos[26]. However, in those years, progress in separation technologies brought along new suggestions for getting access to pathogens that passed bacteria filters (called 'filterable' at that time, i.e., not being retained). Such were found also responsible for animal diseases including foot-and-mouth disease, rabies, or pox: As early as in 1907, gel filtration in media based on gelatin or cellulose nitrate (collodium) was proposed as 'ultrafiltration' method with high promise for fractionating those pathogens[42]. Protocols for a reproducible preparation of tobamoviruses sufficiently pure for molecular and biochemical work and for structural studies were, though, implemented only from the 1930s on. Before this, however, the well-known local lesion assay allowing a quantification of infectious TMV [33,43], and neutralizing anti-TMV antisera raised in rabbits [33,44] are further fundamental findings that have contributed to a better understanding of the virus before its purification.
1.3. Tobamovirus Purification Over Time: How and Why
1.3.1. From Crystal-Like Needles to 'Ribonucleoprotein' Helices
Right time, right person, right place - and the right portion of luck: In the early 1930s, the chemist Wendell M. Stanley accepted the challenge to extract the TMV compound at the Rockefeller Institute for Medical Research in Princeton, New Jersey - a scientific environment with experience on enzyme purification and crystallization[as reviewed in more detail elsewhere 26,33,36]. He worked on this in parallel to other researchers in a number of laboratories in the U.S., Great Britain and Australia, some of them very close to purified TMV. Carl George Vinson in the U.S. had already obtained infectious crystals[45,46]. However, Stanley was the first to publish a specific procedure giving rise to needles of a 'crystalline protein' (Figure 1E) with the 'properties of tobacco mosaic virus', ≈30 µm in length[47,48]. These were obtained initially from an ammonium sulfate-precipitate from juice from diseased Turkish tobacco plants, following repeated suspension and re-precipitation including the use of lead subacetate and diatom silica (Celite) for the removal of plant components, and finally an addition of acidified ammonium sulfate. Stanley tested a number of similar protocols (with the flowchart of a variant from 1936 shown in Figure 1D), yielding essentially the same type of needles . These could be 're-crystallized' 15 times without loss of activity - and were reproduced in several hundred batches within the next three years, with comparable outcome and 'identical physical, chemical, biological and serological properties'[49]. Although Stanley believed in the infectious nature of a protein alone, he was honored for his achievement and awarded the Nobel Prize for Chemistry in 1946. Shortly after Stanley's publication, his colleague Ralph Wyckoff achieved an even faster purification of the virus at the same institute: Based on the pioneering development of analytical ultracentrifuges more than a decade earlier by Theodor Svedberg in Sweden[50], he and co-workers had constructed a simpler air-driven prototype[51]. In accordance with results obtained in Sweden shortly before[52], Wyckoff developed a TMV isolation procedure mainly via differential centrifugation[53].
In parallel and only a few months after Stanley's major report on purified TMV 'protein', Frederick C. Bawden, Norman W. Pirie et al. at the Rothamsted Experimental Station in England identified ribonucleic acid as the second main constituent of TMV particles, which, however, was initially not accepted by Stanley[33,54,55,56]. The horserace and dispute on the initial isolation and biochemical characterization of TMV would be worth a much more detailed description, but is not feasible in the course of this review.
TMV has also been subject of many biophysical experiments up to the second world war - with important impact on the development of cutting-edge analytical technologies and instruments. Its rod-like, charged structure was deduced from double refraction observed by Takahashi and Rawlins for virus-containing plant sap already in 1932[49], from X-ray studies on the fibrous precipitates with their 'apparent crystallinity' [55] - better referred to as 'paracrystallinity' [57] -, and later also by quantitative optical double refraction measurements for virus suspensions in flow [58] or electric fields[59]. Capillary viscosimeters revealed orientation-dependent mechanical properties of the anisotropic TMV colloids in a shear flow[60,61], with their liquid-crystalline phase behavior characterized in much detail soon[62], as also their interaction with different types of colloids such as gold particles [63] - routes of tobamovirus research still up to date in novel contexts[e.g., 20,64,65-67]. Figure 1F illustrates the captivating appearance of TMV gels with crystalline status [25]. In most of these early studies, TMV was either enriched via salt-based precipitation as major separation step, similar to the procedure of Stanley[48], and/or by differential centrifugation. Ultracentrifuges had soon found their way into many labs worldwide, so that both the more laborious high-speed as well as the easier-to-handle low-speed centrifugation have been combined with biochemical methodologies[59,68,69,70].
The availability of new preparative equipment and expertise on plant virus purification fell into the same period when three different groups in Berlin constructed distinct types of analytically useful electron microscopes (EMs), after the initial invention of such an instrument with, however, only 16-fold magnification achieved in 1932 by Ernst Ruska and Max Knoll at the Technical University of Berlin[as reviewed in detail 71]. After several bacteria and three types of orthopoxviruses, TMV and potato virus X (PVX) became the first plant viruses studied initially in a pre-serial transmission EM (TEM) at 10-15 nm resolution[72], and with 7 nm resolution in a commercially distributed instrument shortly thereafter[71]. Already the first TEM visualization confirmed the rod-like shape of TMV, and indicated dimensions of 'around 300 respective 150 x 15 mµ' [i.e., nm] of the 'molecules of the TM-virus'[72]. Tobamoviruses have then been among the viral objects most intensely applied for EM analyses, as their robustness, accurate diameters, early use in immunological studies and high availability did not only ensure fruitful studies on the virus particle and its assembly itself, but also suggested TMV as excellent model object for the development of novel preparative and analytical EM methodology still these days[32,71,73].
After the first electron-optical inspection of TMV, however, it needed almost two further decades of analytical and conceptual progress before a consistent understanding of the supramolecular particle structure had been established in the late 1950s. The initial and fundamentally new assumption of TMV as helical assembly was published in 1954 by James Watson, who had collected clues for the virus structure from earlier and own X-ray diffraction patterns in Cambridge, in parallel to Francis Crick's work on the method. Watson, however, could not yet localize the RNA inside and hypothesized its longitudinal insertion in the center of the virions[74]. Building on the growing body of work in several institutes, X-ray scattering experiments of the British scientist Rosalind Franklin and the American Donald Caspar performed in London, U.K., eventually led to a model of TMV as ribonucleoprotein helix with a correctly integrated RNA: Franklin and Caspar described its position between the CP subunits at a radius of 40 nm in a 'pair' of consecutive papers in Nature in 1956, as reviewed elaborately[33,74]. The actual number of 49 CPs per three helical turns was reported about a year later by R. Franklin, Aaron Klug and Kenneth C. Holmes [75] in the course of a Ciba Foundation meeting on viruses in London, which brought together members of all four leading laboratories working on fundamental virus research in the Western countries after the second world war: Berkeley (U.S.), Cambridge and Birkbeck/London (U.K.), and Tübingen (Germany)[74].
Franklin died only a year after her precise description of TMV virions together with Klug and Holmes, without being recognized broadly[74]. Her and her co-workers' scientific route towards the TMV structure had been prepared by cutting-edge data, conceptual ideas and purified tobamovirus particles from the labs participating in the Ciba Foundation meeting. In Birkbeck, Bernal and Fankuchen had discussed initial evidence for 'repeat units' within the elongated particles already in 1941[62], although TMV was typically regarded a single molecule at that time. The TMV preparations applied in their crystallography/X-ray work were purified by their British colleagues Bawden and Pirie from pre-clarified plant sap, via extensive series of repetitive precipitations, including treatments with alcohol, HCl and NaOH, ammonium sulfate, and NaCl. The resulting 'nearly colourless and slightly opalescent' preparations contained 1 to 2 g TMV from every liter of plant sap[55]. Complementary early evidence for TMV consisting of different subunits came from Germany, where research on viruses was assembled originally at the Kaiser-Wilhelm-Institute in Berlin-Dahlem by Adolf Butenandt, Alfred Kühn, and Fritz von Wettstein with the department heads Gerhard Schramm and Georg Melchers working on TMV from 1938 on[33,76]. In 1943 during world war II, this work was relocated to Tübingen in southern Germany, and founded the Max Planck Institute of Virus Research, where fundamental molecular discoveries on tobamoviruses were achieved mainly from the mid-1950s up to the 1980s, before the institute was re-oriented towards developmental biology (as German plant virology had settled at other places, with the teams of Karl-Wolfgang Mundry and, from 1993 on, Holger Jeske, nearby in Stuttgart - the scientific origin of most of us authors of this article). Still in 1943, Schramm described the disassembly of TMV into smaller building blocks under alkaline conditions, and published their re-assembly four years later[33]. He used to purify TMV (if specified) mainly via ultracentrifugation[77]. Due to his arrangement with the Nazi regime, and the multifaceted relations between those German virologists who continued their work during the Third Reich, and the governmental authorities, as well as with international colleagues (reviewed thoroughly and rich in nuances[33,78]), recognition and impact of Schramm's and co-workers' extensive experiments in the 1940s (published mostly in German language) remained very limited.
This changed only slowly in the 1950s, during the difficult return of German researchers into international scientific communities, which profited from influential emigrated colleagues[78]. Largely in parallel to the X-ray diffraction studies eventually revealing the correct helical arrangement of the TMV building blocks, Schramm and Alfred Gierer as important co-worker, in collaboration with Melchers and Hans Friedrich-Freksa, obtained increasing evidence for the RNA as genetic information encoding the virus. It has not been a straight route at all until in 1956, Schramm and Gierer published that pure TMV RNA isolated from virions raised viral lesions on tobacco leaves - which is sometimes regarded the most fundamental German contribution to tobamovirus research[78]. A report on preliminary experiments of Schramm et al. discussed this possibility already a year earlier - nearly in parallel to a similar suggestion of Heinz Fraenkel-Conrat, competitor in the lab of Stanley in Berkeley. However, Fraenkel-Conrat published his proposal devoid of experimental evidence for an infectivity of RNA released from TMV, which he reported first also in 1956: He then had managed to assemble hybrid TMV particles in vitro, which trans-encapsidated the RNA of a distinct TMV strain and caused the latter's symptoms in plants. In this article, though, he relativized his initial interpretation of RNA as the relevant coding material[78]. Thus, both scientists may deserve equal merits for their findings, and have been honored by the New York Academy's of Sciences Lasker Prize jointly in 1958. While Schramm's work made use of virus particles enriched by ultracentrifugation, Fraenkel-Conrat has not specified their initial purification from plants, but has applied ultracentrifugation as major clue towards his results as well[79].
Until the fundamental structure of TMV had been resolved, virion isolation and analysis was not only brought forward and optimized in the 'Western world', but also in Russia, Japan and further countries. The history of TMV in the 'non-Western' regions, after Ivanowsky's debate with Beijerinck, however, is not covered by most current review articles, due to the more difficult accessibility of relevant articles and a lack in translated digital resources. Exemplary interesting work comprises the purification of distinct TMV variants in the first Japanese air-driven vacuum ultracentrifuge, including a detailed biophysical characterization of the particles in Tokyo as published in 1953[80], and electron microscopic studies on TMV and other plant viruses using ammonium sulfate-precipitated materials in Hokkaido two years later[81]. In Russia, experiments on TMV liquid crystals were reported in 1941, with, however, the details of virus purification not simply accessible[82], as it is the case for precipitation tests with TMV-containing liquids published by the USSR Academy of Sciences in 1950[83].
Understanding both composition and structure has been a primary motivation for purifying immense amounts of tobamoviruses up to mid of the last century (Stanley, 1942). Already in parallel to these investigations, however, curiosity about the principles of function of such viruses, their obviously unique physicochemical properties, and the increasing awareness of their importance as pathogens have become further driving forces in 'tobamovirology'. Hence many different types of experiments soon demanded for isolated virions, and have prompted researchers worldwide to identify purification protocols best-suited for the intended purpose or the lab where the work was performed. The following section therefore will focus on only a few of them.
1.3.2. Manifold Routes of Research and Technical Progress Thereafter
In parallel to the emergence of a consistent picture on structurally and functionally important particle properties in the late 1950s, the fascinating, widely recognized work on TMV inspired many researchers worldwide to tread novel paths in tobamovirus research (worked out thoroughly elsewhere, e.g. in [32,33,34,35,45,84]). This resulted in a plenitude of molecular investigations on the viral lifecycle, its genetically encoded interplay with plant components, and the identification of more and more tobamoviruses in various crops, ornamentals, and wild plants. Efforts to resolve the respective virion structures at increased resolution, and investigations of their behavior under different chemical and physical treatments, namely with regard to their assembly and disassembly, were directly associated with a continuous need of purified tobamovirus particles in laboratories worldwide. This soon entailed almost countless numbers of published studies, for which reason only a few exemplary lines of research can be related to typically applied virion isolation strategies. Technical progress as a complementary driving force has also affected purification protocols. The most recent increase in publications on TMV and further tobamoviruses will be treated in a separate sub-section: the use of viral nanotubes in nanotechnology and biomedical applications (see below).
The purposes for which tobamovirus particles have been purified from infected plant material have varied over time. In the early days, until the advent of molecular biological methods, virions were mostly used to determine the virus family and species by various analytical procedures [85,86,87] (references in Table S1). At the same time, newly discovered tobamovirus species were clearly distinguished from those already known, as indicated by distinct names or 'strain descriptors', and further characterized if necessary. Analytical approaches were based on the particles' serological reactivity, the amino acid composition of the CP, and on infection experiments to determine the host plant spectrum and symptom development in diagnostic host plants (syn. 'indicator plants', a concept introduced in 1931 [88]).
Tobamovirus virions have a strong immunogenic effect and are serologically different [86,87]. Therefore, purified particles were employed already in the 1960s to generate specific antisera for diagnostic applications, which are widely used still today in various test formats for rapid monitoring of tobamovirus infections [85,89,90,91]. Many current standard diagnostic protocols for the detection and identification of plant viruses including tobamoviruses, published, e.g., by the European Plant Protection Organization (EPPO), describe antisera/antibody-based techniques such as enzyme-linked immunosorbent assay (ELISA) or immunoelectron microscopy (EPPO Standards – PM 7 Diagnostics: PM 7/125(1) ELISA tests for plant viruses; PM 7/126(1) Electron microscopy in diagnosis of plant viruses).
Isolated virions have been subject of structural investigations from the early experimental stages on up to now, e.g., via X-ray crystallography, electron microscopy, light diffraction, light scattering or flow birefringence analyses [25,62,92,93,94,95,96,97,98]. They have repeatedly served as model particles for scrutinizing novel technologies, which has, e.g., given rise to a 2. 3 Å-resolved TMV structure upon testing the power of a direct electron detector in cryo-EM single-particle analysis [99]. Tobamovirus particles were also analyzed qualitatively by mass spectrometry, allowing the identification of variants with mutated CPs within the population that would be missed by other methods such as by SDS-PAGE analysis [100,101]. For structural examination, virion purification techniques often had to be adapted to special requirements of the respective analytical methods (for more details and case studies see Chapter 3.1).
Initially, strategies for particle purification were based firstly on clarification and removal of non-viral components from the plant homogenate, e.g. by filtration, salt precipitation, emulsion with chloroform and/or butanol and centrifugation, and secondly on concentration and further purification by precipitation, e.g. by salt or acid and various centrifugation techniques such as differential and density gradient centrifugation (for a more detailed presentation of the techniques, see the following chapters and Table S1 with references therein). For higher degrees of purity, various of these techniques were combined, and/or several cycles were carried out serially. Alternatively, chromatography, e.g. via a matrix of controlled-pore glass beads, was used to finalize the preparation (Chapter 2.5, Table X, Table S1). Since the development of PEG precipitation in the 1960s for virion purification procedures [102,103], it has become the preferred standard method for tobamoviruses (Chapters 1.3.2, 2.3 and 2.4.4). Its advantages are the high yield of particles obtained and the relatively simple handling. With the growing interest in using plant virus particles for biotechnological, nanotechnological and biomedical applications, the need for easy-to-perform and high-yield purification methods for tobamoviruses is increasing (Chapters 1.3.3).
Purified virions have also served as source for the isolation of single-stranded (ss) genomic RNA molecules of sense polarity (+). The obtained viral RNA was initially characterized by electrophoretic techniques, and with the development of high performance molecular biology tools it then served as starting material for cloning purposes, sequencing of full-length genomes and the generation of plant-infectious constructs. Within the last decades, the increasing amount of sequence information has become the main source of identification of new species, and for extensive phylogenetic analyses of tobamoviruses [91,104,105,106]. This has rendered virion isolation largely unnecessary for monitoring tobamovirus diversity and evolution. In particular, the establishment of powerful and sensitive methods for a direct amplification of viral RNA by RT-PCR, and high-throughput next generation sequencing (NGS) methods have opened up alternative ways for the unambiguous identification of tobamoviruses [89], and references in Table S1.
1.3.3. Novel Applications: Increasing Demand: Tobamovirus Particles as Tools
Starting in the 1980s and accelerating substantially since the turn of the millennium, a conspicuous international renaissance of tobamovirus research is observed: TMV and related taxa have been 'refurbished' in several virology research teams, and also found their ways into labs not investigating plant viruses before. This is due to the rise of novel expression and nano-technologies that make viral derivatives attractive tools and building blocks in materials and devices, with highly diverse application prospects. These have boosted the demand for tobamovirus nanoparticles considerably and are thus outlined in greater detail, before we throw a glance at the virion purification methods employed.
Already in 1989, T. Michael A. Wilson at the John Innes Institute in Norwich, U.K., has reviewed plant virus-based 'designer functions' that had developed fast in the previous few years [107]. The pioneering work included repurposed tobamovirus-derived elements such as the TMV packaging signal applied for the protection of heterologous RNAs in pseudovirus particles, the viral RNA's omega leader used as translational enhancer, TMV sequences promising for pathogen-derived plant protection strategies, and a first study on the use of TMV CP fusion proteins as carrier for foreign epitopes [see 107 for original references]. The foresighted work of Haynes et al. in Ontario, Canada has yielded the first self-assembling TMV-based polio vaccine from an E. coli-expressed CP fusion protein, inducing neutralizing antibodies in rats [108]. Since then, tobamovirus-derived particles became increasingly recognized as robust but tailorable, sustainably produced and bio-degradable multivalent nanoscaffolds, with a multitude of biomedical and technical applications tested with highly promising prospects [109]. Technology-oriented research and developments include uses of viral nanoparticles (VNPs) and virus-like particles (VLPs) as biotemplates for inorganic and organic compounds, and uses as carriers for biomolecules [11]. Thirty years of creative developments have given rise to an enormous variety of novel colloidal (particulate), layered and volume materials, either active by themselves as hybrid structures, or after an integration into technical devices [110,111,112,113,114,115,116,117,118,119,120,121]. The following showcases only a small selection out of the many exciting overviews and cutting-edge studies.
Biomedical uses of tobamovirus derivatives are evaluated for both human and veterinary treatments as well as in phytopathology, with auspicious trials on the animal and laboratory testing level [111,122,123]. They span from vaccines, adjuvants and test antigens [124,125,126,127,128,129] to the intravital delivery of tobamovirus diagnostic imaging agents and therapeutics, with cutting-edge multitasking and theranostic approaches [130,131]. Inactivated tobacco mild green mosaic virus (TMGMV) shows agronomic promise for a root-directed supply of pesticides [132], with the plus that the replicating TMGMV is also approved as herbicide against invasive plants in the U.S., due to its limited risk of spread and its natural occurrence [133]. This would make inefficient virus inactivation due to technical failures manageable. Primary advantages of tobamoviral (and other plant viral) delivery systems consist in their particle structure with cargo transport possible in the inner channel, on the outer surface of more than 2000 repetitively arranged and selectively addressable CP subunits [134], and in-between these CPs [135]. Various applications also may benefit from the intriguing options for shape design enabled through in vitro assembly of tobamovirus derivatives [114], their the high and further tailorable biocompatibility with an increasing understanding of tobamovirus pharmacology following medical administration[118,136], and many means of multifunctionalization, e.g., with cell-targeting molecules, tracer and effector compounds. Therefore, tobamoviruses also hold great opportunities in scaffold-assisted tissue and organoid engineering technologies, to generate implants and model structures for personalized treatments: The rod-like VNPs exert beneficial effects on cells cultivated on planar substrates and in 3D-hydrogels and can guide the diffentiation of progenitor cells in response to their contact with peptides presented on the viral backbones [137,138]. Recently, tobamoviral scaffolds were combined with other plant viral effectors into osteogenesis-promoting hydrogels that were able to directed the formation of mineralized bone tissue-like entities [139].
Beyond such uses with living organisms or cellular structures, tobamovirus particles are employed as high surface-area biotemplates and nanocarriers in various kinds of technical devices and composite materials, several of them in the close-to-application status. Hydrated and especially dried TMV has proven a robustness that often surpassed all initial expectations [114,140,141] so that even electrospun polymer fibers with biofunctionalized TMV [142], nanostructured nickel-coated surfaces for efficient boiling heat transfer [143], or long-term durable biosensors with TMV adapter elements [144,145,146] have become available.
In tobamovirus-aided fabrication, virions have initially served as richly available templates for the deposition of hard compounds, which has yielded mineralized tubes around TMV cores already in 1999 [147,148], nanowires inside the central channel a few years later[21], and numerous types of functional composites with different inorganic and organic matter thereafter [149]. While originally native tobamoviruses were used, chemically and genetically modified virions have soon enabled a better-controlled interaction or linkage with heterologous compounds as well as their integration into technical devices [11,112,113,121,150,151,152,153,154]. Many of these profit from a considerable surface enhancement and good steric accessibility of the nanorods equipped, e.g., with coatings of conductive or catalytically active metals, semi-conductors or alloys [18,22,155,156,157], chromophores[158], polymers [159], and even fullerenes [160] or metal-organic frameworks [161]. The resulting 'smart materials' and virus-assisted devices have been prepared for applications in fundamental research [162], energy storage and conversion [163,164], data memory [165], environmental remediation [166], as antireflective coatings [167], ferrofluids with increased magnetoviscosity [66], fibres for metal nanoparticle production, and many more [168, and above-listed exemplary review articles,169-171].
Combined with biomolecules such as peptides, antibodies and enzymes, tobamoviruses are not only applied in medical contexts as outlined above, but have an equally promising track record also in the fabrication of new types of biosensors, enzymatically active nanoparticles and materials for bioaffinity enrichment or deposition [11,114,146]. Virions displaying genetically fused or chemically installed peptides on their outer CP surfaces were, for instance, shown to nucleate a selective mineralization of different materials [12,172], or to direct self-ligation with complementary peptide tags, as demonstrated by way of the Streptococcus pyogenes-based SpyTag/SpyCatcher (ST/SC) system [173] for the immobilization of fungal enzymes on TMV [174]. Such peptide-mediated coupling of larger proteins is one of a few biomolecular workarounds for overcoming restrictions in size and charge of genetic fusions to the TMV CP, which are known to abolish systemic infectivity and/or virus assembly for most amino acid chains longer than 23 residues. Longer fluorescent proteins or receptor-binding protein domains could be installed on a sub-set of the CPs by advanced expression constructs integrating 'ribosomal skip' sequences into the TMV RNA, or by peptide linkers counteracting steric and charge constraints [reviewed in more detail in 175,176,177]. Tobacco vein clearing virus (TVCV), a subgroup 3 tobamovirus (see below), has even allowed the C-terminal extension of its CP by 148 amino acids, to directly display two IgG-binding domains of Staphylococcus aureus protein A (PA) on all CP subunits [178]. The systemically infectious, immunoadsorbent particles could be equipped with a two-enzyme system conveniently by way of enzyme-recognizing antibody conjugates captured by the PA domain shell [3].
TMV and TVCV particles with sensor enzymes installed at high surface densities this way or via chemical and bioaffinity linkage have proven excellent functionality as adapter coatings in microtiter plate-based colorimetric assays and in different label-free electrochemical biosensor layouts. The exposure of different enzymes converting antibiotics, sugars, urea or fermentation markers on tobamoviral carriers enhanced sensor performance and reusability considerably, in comparison to sensors with conventionally deposited enzymes [145,146,179,180,181,182,183]. TMV stabilized the sensor characteristics up to a year of repetitive application without loss in sensitivity in the case of a field-effect sensor for penicillin [144]. Similarly beneficial effects of tobamo-VLP-scaffolded receptor layers were observed for impedimetric on-chip sensors [184] and in an optical microdisk resonator setup [151] for the label-free detection of antibodies via VLP-exposed peptides. The detection system could be transferred into capillary flow-loaded microfluidic devices [185], paving the way towards portable TMV-enhanced detectors. All-soft, liquid-permeable biosensors are a further layout recently enabled by TMV rods, which retained signal-generating sensor enzymes in an immersed porous hydrogel matrix [186]. Last but not least, the RNA-guided in vitro assembly of TMV-like particles offers unique access to uncommon nanrod-derived straight and kinked structures with controllable 'arm' lengths, as well as to nanorods with selectively addressable longitudinal subdomainsv[reviewed in detail 187]. The latter may be generated in precise lengths by way of dynamic DNA-assisted nanotechnology [6], which allows, e.g., the display of well-defined biomolecule ensembles and has a large potential for fundamental research [188].
The huge number of rapidly developing tobamovirus applications led us to assume that only a few particle purification protocols have become standard during recent years, due to their ease, efficiency, speed, low requirements for instrumentation, or simply their good description in widely recognized studies. However, this guess has only partially been confirmed by a small set of 'test drillings' in technology-oriented publications from distinct areas of research and different countries: Virion precipitation from pre-cleared plant homogenates, often by way of PEG, in combination with differential and, in part of the studies, density gradient or cushion centrifugation, seems most frequently employed all over the world [134,138,162,164,172,177,189,190,191,192,193,194,195]. Nevertheless, a considerable variety of deviating conditions in the serial processing steps is found for one and the same tobamovirus species, and even within single research groups, different protocols are applied. This may be due to a specific need for, e.g., narrow particle length distributions obtainable, e.g., by zonal zentrifugation (i.e., separation according to sedimentation coefficients), or uses demanding for superior removal or avoidance of certain contaminants (Chapter 3.1). This might be one of the reasons accounting for the co-existence of several clearing treatments for the plant raw extract, ranging from organic solutes via heat to mineral adsorbants (see Chapter 2.2). In other cases, though, the choice of a specific protocol might also reflect the ready-to-hand literature, the availability of a certain PEG type in the cupboard, or personal experience - in order to not change a winning team (Materials and Methods sections typically do not contain too many arguments). Alternative strategies for the preparation of tobamovirus particles are not extinct either: Some researchers precipitate these by ammonium sulfate [196,197,198], others refrain from any precipitation and enrich virions by different consecutive centrifugation steps. If a background of accompanying compounds from plants is tolerated or even wanted in the work, such protocols can be quite simple [199]. In the opposite case, if highly pure virus is needed, e.g., for ultrastructural analyses [200], centrifugal enrichment may comprise extensive series of differential sedimentation and resolvation steps, essentially according to [201]. In more recent work, high-resolution cryo-EM maps have also been obtained from PEG-precipitated TMV, but after post-purification via repetitive differential centrifugation [98,202], to get rid of residual polymer on the virus particles.
Hence, our conclusions on contemporary purification protocols for tobamovirus particles may be kept short: The use of PEG is most widespread so that G.V. Gooding, Jr., and T.T. Hebert from the North Carolina State University, USA [102], and S.N. Chapman from the Scottish Crop Research Institute, Scotland [203] will remain well-known also to the next generations of plant virologists. However, many variations of the theme exist, and also considerably different virion isolation approaches are likely to survive.
1.4. Tobamovirus Properties and Variability - Essential Knowledge
Knowledge on the in-planta accumulation and biological diversity of tobamoviruses with deviating physical and biochemical properties is crucial for well-adapted and successful purification protocols, as follows.
The virions often form large para-crystalline inclusions of elongated or plate-like shape in infected plant cells, which are easily recognizable already under the light microscope, and reveal their densely packed particle arrangement under the electron microscope at high resolution [204] (references in Table S1, Figure 1B and C). Virions show per se liquid crystalline properties [25 and references therein,55,62,205]. In planta, they form liquid crystals of 2D smetic order (hexagonal crystals), whereas the crystalline phases obtained for purified virions in vitro are mostly nematic [92,205,206,207]. Kreibig and Wetter [25] investigated the crystalline properties of six different tobamovirus species by optical diffraction. They showed that purified virions from all tested species formed iridescent gels with different micro- and macrocrystalline structural phases in vitro (nematic and smectic liquid crystals, Figure 1F). Examination by optical diffraction indicated a crystalline multilayer structure similar to the immature crystal forms in living plant cells.
Tobamovirus particles have a diameter of approx. 18 nm; the predominant lengths are 300-312 nm [208,209,210,211]. Encapsidation of subgenomic RNAs also results in shorter VLPs (32-40 nm), which typically make up only a small proportion of the virus population [212,213,214,215]. The rod-shaped particles have a helical organization built from the viral RNA and a single protein of 1.70-1.80 x 10 Dalton (156-161 amino acids). The TMV particle, as an example, contains approximately 2130 CP subunits that are tightly packed into a rigid tube 300 nm long, with an inner and outer diameter of 4 nm and 18 nm, respectively. The tube has a helical structure with 16 1/3 protein subunits/helix and a clockwise base pitch of 2.3 nm [216].
Different, position-dependent types of CP-CP and CP-RNA-interactions contribute to its high stability [98,200,217]. A (+) ssRNA of 6.3 to 6.5 kbase length is 'sandwiched' in-between the CP helix of the particle at a radius of approximately 4 nm, with three nucleotides associated with each protein subunit [208,209,210,211,218]. This repetitive supramolecular arrangement is visualized clearly elsewhere, e.g., in [219,220], and described more concisely also in our previous review [114]. The end of the tobamovirus particle containing the 3'-terminus of the genome is convex in shape, while the other one is concave [221]. The genomic viral RNA constitutes ca. 5% of the particle weight and encodes at least three proteins, the 126-kDa replicase protein and its readthrough of 183-kDa, the 30-kDa movement protein and the CP. For TMV vulgare (i.e., strain U1), the full genome sequence has been published in 1982 [222].
Particles of distinct tobamovirus species differ slightly in their properties. The virions have a buoyant density in CsCl of 1.307 - 1.325 g cm-3 and a sedimentation coefficient from 187 S to 212 S [208,209] (references in Table S1). The isoelectric point (pI) varies between 3.7 and 4.64, although the values determined experimentally may be slightly lowered by PEG remnants on the virions after PEG-mediated precipitation, e.g., from pH 3.7 to pH 3.38 in the case of pepper mild mottle virus [223]. Tobamoviruses have a thermal inactivation point (10 min) of 90°C and survive in plant sap for many years, up to 50 years were reported [224].
At present, the genus Tobamovirus is one out of seven in the family Virgaviridae, and comprises 37 accepted plus two tentative species not yet classified [208; https://ictv.global/report/chapter/virgaviridae/virgaviridae/tobamovirus]. TMV represents the type member. Particles of at least 31 tobamovirus species have been purified from infected plant material (Table S1) by various methods, often based on PEG precipitation in combination with further techniques. Tobamoviruses can be categorized into at least three subgroups based on the amino acid composition of their CPs, which correlates with serological specificity, CP amino acid primary structure and the host from which the viruses were originally isolated [105,106,225,226]. Further differences between the groups exist in the position of their origins of assembly (OAs) in the RNA, and the separation or partial overlap of the open reading frames (ORFs) of the movement protein and the coat protein [106,208,218,227].
Tobamoviruses have a broad host range that includes many crop and ornamental plants such as tobacco, tomato, cucumber and orchids, and they cause major economic damage with, in some cases, open end - as it is currently the case for the tomato brown rugose fruit virus pandemic, with the virus detected initially in 2015 in Jordan now threatenting tomato cultivation worldwide [91,218,228,229,230,231,232,233].
2. Techniques for Virion Enrichment and Storage, One by One
The extraordinary stability of tobamoviruses exemplified above and detailed further below (in Section 2.6) is exploited to separate the virus particles from plant components. Prior to purification, it is a common practice to freeze TMV-infected plant material either at -20°C or in liquid nitrogen [94,99,234,235]. The integrity of the virus particles is not obviously hampered by this procedure [236], which allows to separate harvest from purification procedures - at least for experiments not relying on highest-quality, evenly suspended virions (see Section 3.1 and [25]). Likewise, fresh infected plant tissues may be used as a starting material [237].
2.1. Grinding, Blending, and More: Plant Disruption
All purification strategies will start with a homogenization step in order to disrupt plant tissue and cells, to make the virus particles accessible for purification. At this point, precipitation of the particles into an insoluble aggregate has to be prevented. If - primarily due to the release of vacuolar contents - the pH in the homogenate will drop close to the viral pI (as an example, pI of 3.3 for TMV [238,239]), the virus particles might be lost due to aggregation. Therefore, homogenization is most commonly performed in sufficiently concentrated phosphate buffer at a pH around 7. In early work, Bawden and Pirie [240] developed an extraction protocol of TMV by freezing and thawing plant tissue in pure water, with aggregation of TMV counteracted by a large liquid volume and repeated neutralization when necessary.
For mechanical disruption of the plant material, different mincer and blender-type devices are employed, including domestic meat mincers, Waring blenders or Smoothie Makers [68,235,241]. The process may lead to a substantial production of frictional heat, so that cold disruption or intermitted mixing may be advantageous [237]. Grinding with a mortar and pestle [94] with an optional addition of an abrasive generates less heat, but seems of lower importance for the purification of tobamoviruses compared to other virus genera because of the nanotubes' stability. Notably, though, for natural TVCV and especially the protein A-domain fashioned TVCVPA, mechanical damage during the early purification steps was obvious upon use of standard TMV isolation protocols [178,241], for which reason we developed an initial version of a gentle PEG precipitate resolubilisation method in inverse PEG-sucrose gradients [3], which has been the starting point for our novel iodixanol-based method described below (Chapter 4). A systematic comparison of the particle sensitivities against shear forces between distinct tobamovirus species might be worth testing, as simple predictions from structural models seem not yet possible.
In any case, however, supplementation of homogenization buffers with protease inhibitors, commonly used in the biochemical purification of various molecules and supramolecular complexes from plants, is not a necessary standard for tobamovirus isolation. Even the mechanically more sensitive TVCVPA, which exhibits some CP degradation over time, did not profit from supplementation of the extraction media with different protease inhibitors tested [3,241]. This indicates that the mechanical sensitivity of tobamovirus particles against rupture or breakaway of protruding heterologous CP portions may be an issue, but protease sensitivity usually not.
2.2. Getting Rid of Plant Stuff: Clarification
Crude extracts can be directly used for purification of tobamovirus particles by one or the other method (see Section 2.3, 2.4 and 2.5), but more often homogenates are first clarified by filtration through muslin, miracloth, cheesecloth, gauze or similar [234]. This step will eliminate a major part of the larger plant components, e.g. unbroken tissue segments, leaf ribs and fibers. As an alternative or in addition, filtration through a layer of adsorbents like Celite® (a diatom silica preparation) or Bentonite (a less defined clay mineral primariliy from silica and a considerable amount of alumina) will further facilitate clearance from plant material [242]. Also charcoal may be applied [23]. A more comprehensive view is presented in 2.5.2, as this clarification procedure can be regarded as a form of chromatography. Clarification of crude extracts is also achieved by a first low-speed centrifugation and will be described later in detail (Section 2.4).
Since the structure of TMV particles is not affected by freezing or heating up to 50 °C for about two hours [242] or about 90 °C for ten min in solution [243], these procedures can be used to precipitate plant components that can be further separated by a low-speed centrifugation step. Mundry [237] has described a method for TMV purification by a combination of cycles of freezing and heat treatment at 60°C. However, heat incubation alone at 50 to 55°C can suffice to remove unwanted plant components from pre-filtered TMV-containing homogenates, with the heat-precipitated compounds sedimented by centrifugation therafter [234,242].
A substantial amount of plant contaminants can be removed by incubation with n-butanol and/or chloroform, causing both precipitation of the unwanted material and extraction of chlorophyll into the organic phase simultaneously, while the virus particles will remain in the aqueous phase. This procedure is typically followed by centrifugation as well, to separate the precipitate and the organic phase from the TMV particles [102,203].
2.3. Precipitation of Virus Particles
Since the early days of tobamovirus purification, antichaotropic salts of the Hofmeister series like ammonium sulphate have been used successfully employed to precipitate tobamovirus particles from cleared extracts or homogenates [68,197,198,234]. If necessary, the salt-mediated precipitation is repeated to increase yield and purity. If TMV is already enriched, a slow increase in the ammonium sulphate concentration leads to the assembly of TMV into small paracrystalline needles of about 0.02 mm length ([244], Figure 1E).
An efficient and to some extent selective precipitation of virions can be achieved by the addition of the polymer PEG in concentrations above a threshold specific for the respective virus type, and is promoted by the presence of NaCl. The method has been introduced for plant virus purification by T.T. Hebert in 1963 [103], and optimized for TMV by R. Leberman [245] and further by G.V. Gooding and Hebert [102] soon afterwards. As their depletion-induced precipitation depends on shape, charge and size of the particles (worked out briefly in [3]), conditions may be found that separate virions well from the majority of accessory plant compounds. For tobamovirus purification, PEG precipitation has become a predominant method as detailed in 1.3.3). In order to increase the yield of virus particles, repeated rounds of PEG precipitation may be performed.
A major advantage of purification protocols using precipitation steps for the enrichment of viruses is the dispensibility of ultracentrifugation, because the aggregated virions can be sedimented efficiently by low g-forces. This does not only reduce the necessary instrumentation and effort, but also allows processing larger amounts of virion-containing plant extracts. The structural integrity of TMV virus particles collected via PEG precipitation is demonstrated by the high-resolution structure determined by electron cryo-microscopy and image reconstruction [98].
2.4. Centrifugation
Centrifugation stands as a cornerstone technique to separate and concentrate tobamoviruses from cellular debris of plant tissue. It capitalizes on differences in size, density, and shape to achieve purification and enrichment. Ultracentrifuges, capable of generating immensely high centrifugal forces ranging from 100,000 x g to 1,000,000 x g are instrumental to sediment particles with sedimentation coefficients in the range of 10 to 105 S. For instance, TMV, with a sedimentation coefficient of approximately s=200 S or 200 * 10-13 s in aqueous solution [52,246], can sediment at a velocity of around 10 cm/h in a gravitational field of RCF=140,000 x g (for theory see [247]).
Ultracentrifugation finds application both analytically to determine the sedimentation coefficients of a particle, or preparatively to separate the virus from cellular debris and/ or to concentrate virus preparations. Preparative ultracentrifugation may utilize different sedimentation coefficients ('differential sedimentation'), different densities ('isopycnic density gradient centrifugation') or a combination thereof known ('zonal density gradient centrifugation') to separate different particles. To scale up preparative ultracentrifugation, rotor designs and appropriate equipment enable continuous ultracentrifugation (Figure 2A-B) with or without density gradient material.
2.4.1. Differential Sedimentation
An initial documentation of tobamovirus preparation through differential sedimentation dates back to 1927 (McKinney, 1927), marking a pivotal moment in virology. Notably, at that time, the true nature of TMV remained elusive. The sedimentation of the virus was only achieved after heat coagulation of the virus. Subsequent analytical determinations of sedimentation coefficients [52,246] paved the way for the widespread use of ultracentrifugation to purify and characterize the virus from plant sap [248,249,250,251,252,253]. Early protocols involved multiple high speed centrifugations to pellet the virus, interspersed with low speed centrifugations to separate the virus into the supernatant ('differential centrifugation'). However, these early preparations often suffered from aggregation that could not be resolved. Improved buffer conditions for preparation (0.1 M phosphate) and storage (0.01 M phosphate) of the virions were subsequently elucidated [94,254]. Differential centrifugation was applied for the isolation of many tobamoviruses (e.g., [25]; Table S1). Aggregation during preparation for TEM analytics, however, remained a concern, challenging the perception of virus monodispersity in solution [255]. Tobamoviruses have also been purified by several adaptations of differential centrifugation such as parallel chloroform extraction ([256], and Table S1) and high-speed centrifugation through sucrose cushions ([257,258]; Table S1 ), which may present potential alternatives to labor-intensive density gradient centrifugation [259].
2.4.2. Continuous Ultracentrifugation
Despite centrifugation traditionally being viewed as limited in terms of scalability, advancements in rotor design have facilitated a degree of scalability that allows for the preparation of larger quantities of viral particles. This approach was first documented in 1942 [260], when it enabled the preparation of up to 15 g of TMV within ten hours. Given the substantial yield achieved through this method, it warrants reconsideration for applications necessitating high quantities of virus. Furthermore, continuous ultracentrifugation has been adapted to gradient separations for human influenza B and herpes simplex virus [261]. However, to the best of the authors' knowledge, the purification of tobamoviruses with continuous density gradient centrifugation has not been reported.
2.4.3. Density Gradient Centrifugation
Density gradient centrifugation of plant viruses has been extensively reviewed [262]. In one variant, known as rate zonal gradient centrifugation, sucrose gradients typically ranging from 10 to 40 % (corresponding to densities between 1.04 and 1.18 g/ml) are prepared in a centrifugation tube [263,264]. Solutions containing the virus, with a density of about 1.3 g/ml in the case of TMV, are layered on top of the gradient and the virus particles are centrifuged into the gradient for 1-3 h, depending on centrifugal force. As the density of the gradient material is lower than that of the virus, the duration of centrifugation must be carefully controlled. Methodological nuances include variations in the average sucrose density, gradient steepness, and virus loading quantities [265,266]. Despite its long-standing use, the method has remained prevalent in contemporary laboratories [101,189,267]. Notably, adjustments to the gradient may be necessary for varying particle lengths [268] as zonal gradient centrifugation depends not only on density but also on virus shape and size, enabling to separate particles that have been disassembled to different degrees [269].
In contrast, isopycnic gradient centrifugation employs a density gradient medium matching the density of the virus particles. During centrifugation, the virus migrates until its density aligns with that of the medium, forming a distinct band (Figure 2C). While for rate zonal gradient centrifugation, the gradient is typically pre-formed, during isopycnic gradient centrifugation the gradient is formed during centrifugation, requiring several hours to reach equilibrium. TMV, for example, achieves equilibrium in gradients formed by 32% CsCl only after approximately 16 hours [270]. Alternative alkali halides yielded comparable results to CsCl (ibid). To address CsCl’s high ionic strength and a potential for virus aggregation, non-ionic and non-chaotropic density gradient media such as Metrizamide and Nycodenz® have been explored [271,272]. Both compounds share a tri-iodinated benzamido group and multiple hydroxyl groups as common structural elements [273,274]. Metrizamide facilitated shallower gradients and improved separation of closely related tobamoviruses [272](Skotnicki et al., 1976). Nycodenz® allowed the separation of potato virus X and TMV (Figure 2C), but poses challenges in its removal from the virus particle preparations after centrifugation, thus making it attractive mainly for viruses unstable in CsCl or Metrizamide [271].
Colloidal silica presents a compelling alternative to traditional density gradient methods, offering reduced run times and mitigated osmolarities [275]. Isodensity banding with colloidal silica can be achieved in less than 1 h, with viruses banding at much lower apparent densities compared to CsCl, owing to their exclusion of colloidal silica from the surrounding medium. This exclusion effect, contingent on size and form, enables separation based not only on density but also on the exclusion phenomenon. However, silica adherence to viral particles necessitates precipitation to remove silica from the virus after centrifugation, a challenge addressed by coating silica particles with polyvinylpyrrolidone. This innovation has led to the development of Percoll—a widely used gradient material for cell separation [276]. While there is currently no reported data on TMV purification using Percoll, its potential merits further exploration in virological research.
2.4.4. Solubility Gradient Centrifugation
A refined isolation method combines precipitation with density gradient centrifugation [1,2]. This method involves centrifugation of PEG-precipitated virus through a reverse concentration gradient of PEG 6000 stabilized in a positive sucrose gradient. The solubility concentration, determining when virus particles resolubilize, relies on their surface/volume ratio and charge characteristics, and is influenced by pH and ionic strength within the solubility gradient. This technique has been successfully tested on TMV and plant viruses from various genera [2]. Recently, it was adapted to gently isolate the tobamoviruses TVCV and its mutant TVCVPA [3], resulting in a well-preserved particle length compared to that of virions obtained via methods involving PEG-precipitation and differential centrifugation [178,241]; see also Section 2.1.
2.5. Chromatography
Chromatography is employed in two distinct modes for tobamovirus purification. In the flow-through mode, the virus traverses the chromatographic matrix while contaminants are retained. This mode is frequently applied in the initial stages of virus isolation to clarify plant saps. In the bind/elute mode, the virus is selectively captured under specific binding conditions by the column medium, while contaminants are not. Subsequent elution facilitates direct virus purification (Table 1).
2.5.1. Flow-through Chromatography
Initially, chromatography found application in the purification of tobamoviruses through its capacity to bind plant components to the chromatographic material, thereby clarifying the virus-containing extracts. For a comparison of flow-through chromatography with other clarification methods, please refer to chapter 2.2. One of the earliest materials utilized for flow-through chromatography was Celite, derived from pulverized diatom shells and predominantly composed of amorphous silicon dioxide [244]. Other materials such as Bentonite, a complex aluminum silicate, charcoal and DEAE-Cellulose were similarly employed [280,281]. These materials effectively remove plant components such as ribosomes, Rubisco, or phytoferritin from extracts containing the virus, thereby facilitating the purification of several tobamoviruses [25,282]; Table S1).
The use of paper chromatography marked an interesting development in TMV purification by flow-through chromatography, employing a simple yet effective principle where the virus could migrate through paper at pH values of 4.5 or higher, leaving behind plant components like chlorophyll at the application site [283]. In a notable modification to this technique, an electric field was applied during paper chromatography at pH 7, resulting in the separation of plant components and a distinct fraction containing the virus, demonstrating an early instance of electrophoretic mobility enhancing chromatographic separation [284].
Chromatography, typically effective for protein separation, faces limitations with tobamoviruses due to their size. Standard methods often use porous beads, but tobamoviruses' larger size prevents efficient diffusion into these pores, reducing interaction with the stationary phase and complicating isolation efforts. Still, methods based on chromatography have been developed for clarification or purification of the virus.
Size exclusion chromatography, utilizing porous particles, differentiates molecules based on size. Small molecules and buffer components enter the pores and traverse the column slowly, whereas larger entities like viruses bypass these pores, enabling faster movement. This method effectively acts as a 'flow through' mode for viruses. Advances in isolation techniques have introduced materials such as controlled pore glass [285] or microgranular Spheron gels [286] for efficient separation of TMV and other plant viruses through size exclusion column chromatography ([287,288]; Table S1).
2.5.2. Bind/Elute Chromatography
Bind/Elute chromatography is the main chromatographic method employed for virus purification (Table 1). Early chromatography media for tobamovirus enrichment included cellulose derivatives [289,290,291]. With the advent of monolithic columns [292,293], tobamoviruses found compatibility with supports fashioned as continuous homogeneous phases [292,294,295]. Unlike individual particles, monolithic columns (Figure 2D) feature large pores resembling channels throughout the entire column, rendering the entire stationary surface accessible to tobamoviruses. Convective Interaction Media (CIM) monolithic materials with anion-exchanging capabilities have been particularly successful in tobamovirus studies (Table 1).
Though chromatography being probably one of the most important purification methods for bioproduction, it is still not commonly applied in routine labs, and no state-of-the-art chromatography material has been accepted. Density gradient ultracentrifugation still is the method of choice when high purity of the virus is required. However, promising materials such as monolithic columns become more and more recognized.
2.6. Finishing
2.6.1. Ultrafiltration
Ultrafiltration has been primarily utilized to concentrate tobamoviruses and for buffer exchange, especially after the functionalization of the virus surface [101].
2.6.2. Lyophilization and Storage of Virus Preparations
The stability of viruses during storage is crucial for their use in research and biotechnological applications. Tobamoviruses, particularly TMV, have been extensively studied for their robustness and persistence under a variety of storage conditions. Early observations noted TMV's exceptional stability, able to retain infectivity after decades in liquid form. This review delves into the seminal works that have contributed to the methodologies for storing tobamoviruses, emphasizing the evolution of liquid and dry storage techniques.
Historically, the liquid storage of tobamoviruses has revealed their extraordinary stability. A pivotal observation was that TMV could remain stable for up to 50 years in liquid formulations, a testament to its robust nature [224]. Recently, an over 40 year old preparation stored in a cold room was analyzed and still predominantly contained 300 nm long TMV rods [195]. This longevity in liquid medium not only underscores the virus's structural stability, but also its potential for long-term studies without significant loss of infectivity.
One study meticulously investigated the stability of TMV at different pH levels and ionic strengths at an elevated temperature of 68 °C [296]. This investigation revealed that a minimum pH of 6.8 (the lowest pH value tested), combined with a 0.2 M phosphate buffer (the highest concentration tested), conferred the highest stability to the virus. Another study showed that ultracentrifugation properties and infectivity of TMV remained stable at 4 °C for up to 28 weeks both in water and 0.02 M phosphate buffer at pH 7.5 [297]. In buffered solution at pH 4, the virus retained its infectivity for up to twelve years at 4°C (the maximum duration tested) [298]. Alternatives to aqueous solutions were also investigated [299]: The CD spectrum of TMV stored at room temperature in ethylammonium mesylate, a protic ionic liquid, was stable for up to four months (the maximum reported), while it decreased within three weeks when stored in 10 mM phosphate buffer pH 7.2.
The transition to dry storage methods presented new avenues for preserving tobamoviruses, with techniques such as freezing and lyophilization (freeze-drying) being explored. No changes in ultracentrifugation properties when frozen in water or 0.1 M sodium acetate pH 6 have been observed [236], indicating that freezing did not adversely affect the structural integrity of the virus. When frozen in phosphate buffer, slight changes in ultracentrifugation properties were noticed in one study, but without effects on infectivity [297]. These findings have been pivotal in establishing freezing as a viable method for long-term storage of tobamoviruses.
The drying of viruses presents unique challenges, notably the risk of reduced infectivity. Drying TMV with P2O5 decreased its infectivity to 50% or less, regardless of whether the sample had been frozen before drying or whether the pH of the solution was neutral or acidic. However, drying with Na2SO4/ Na2SO4*10 H2O or saturated ammonium sulphate retained the infectivity of TMV [55]. This indicates a certain level of stability during the drying process, but also underscores a propensity for infectivity loss. Freeze-drying was explored as a method to retain stability of the virus: When dried under very low pressures (<0.1 mm Hg, corresponding to 0.13 mbar), only few particles broke [300]. However, significant particle breakage occurred at higher pressures, or if the sample had thawed before water sublimation, suggesting that conditions during freeze-drying critically influence virus integrity. A systematic optimization of freeze-drying conditions for tobamoviruses identified specific temperature and pressure profiles as well as cryoprotectants that sustained up to 89% infectivity [301]. This work significantly advanced the understanding of freeze-drying as a viable method for the long-term storage of tobamoviruses, offering insights into the delicate balance required to maintain viral integrity during the lyophilization process.
The historical advancements in the storage of tobamoviruses, from liquid formulations to dry storage methods, have provided a foundation for current practices in virology research and biotechnology. The remarkable stability of tobamoviruses, particularly TMV, under various conditions has not only facilitated their use in scientific studies but also highlighted the importance of specific storage conditions to preserve viral infectivity and integrity. As research continues, the insights gained from historical studies will undoubtedly contribute to further innovations in the storage and utilization of viruses for a range of applications.
3. Combined Recipes: Stirred, Shaken and Poured
3.1. Adaptating Purification Strategies to Specific Needs: Combined Protocols and a Few Pitfalls to Keep in Mind
Purification strategies generally have to be adapted to needs and requirements of the intended follow-up experiments with the isolated virus particles. Some aspects to be considered as well as an instructive example highlighting the relevance of optimized purification protocols are summarized in this chapter.
The early methods used for the purification of tobamovirus particles were based primarily on precipitation and centrifugation, and aimed to obtain sample material for understanding the identity of the mosaic-inducing agent, for the identification of virus species, and for elucidation of the underlying structures by X-ray diffraction (Section 1.3.1 and 1.3.2 and Table S1). Around thirty years later, PEG precipitation was established in virus purification procedures [102,103,203,245] including protocols for tobamoviruses, which have evolved into the standard methods due to several advantages (Section 2.3). They yield virus particles of sufficient quality for most subsequent applications, though not for every purpose: It should be borne in mind that properties of the virions may be altered by PEG residues adhering to the surface, as indicated by shifted pI [223] and high-resolution EM (according to personal communication with experts). Additionally, it should be considered that divalent cations such as Ca2+ and Mg2+ strongly favor end-to-end aggregation of virions, which might interfere with subsequent applications planned. This phenomenon can be avoided by adding EDTA, and by replacing the divalent ions with Na+ in the first steps of the purification process [302,303,304].
PEG precipitation is often combined with other methods to multi-stage purification processes that are best-suited for the intended down-stream applications of the VNPs, such as antiserum production via immunization of rabbits [305] (Table S1). The combination of a PEG-based isolation protocol with zonal sucrose density gradient centrifugation, or centrifugation through a sucrose cushion, respectively, may be used to fractionate tobamoviral particles according to their aspect ratio, in order to examine and narrow down their size distribution towards relatively uniform preparations [214 according to Otsuki et al., 1977 see Table S1,306]. Such fractionation according to the particle size (corresponding with its sedimentation coefficient, see 2.4.3) can also be achieved by successive centrifugation steps of the pre-cleared homogenate, including separation in a density gradient, to recover the original particle population without PEG precipitation and resuspension [257 see Table S1]. From nanorod fractions of different average lengths, obtained through such protocol, the encapsidated genomic and subgenomic tobamoviral RNAs were isolated and further characterized, e.g., according to their size, coding capacity as revealed by in vitro and in vivo translation, or localization of the origin of assembly sequence [214,257,306].
An avoidance of PEG precipitation has been a rationale also in even simpler purification protocols that aimed at well-dispersed high quality virions. Under carefully adjusted pH and ionic strength conditions, preparations of near-uniform particles are also accessible by well-controlled differential centrifugation of cleared plant crude extract alone. In this case, full-length tubes sedimented prior to the spin-down of shorter ones - which are removed with the supernatants - undergo a number of successive centrifugation cycles each with extensive re-solvation of the pellets overnight [94 and Table S1]. The excellent dispersion and purity of such virion preparations has made them first choice for certain subsequent applications, e.g., for biophysical and structural investigations [25].
The work of Kreibig and Wetter [25] is an informative example of how the choice of a method for particle purification and its precise implementation may determine the outcome of subsequent experiments. As described above (Chapter 1.4), the authors purified virions of six distinct tobamovirus species from infected plant material to study their assembly into different liquid-crystalline phases in vitro by optical diffraction. However, the successful generation of the starting materials, the iridescent gels formed by the virions, depended strikingly on the virus isolation protocol. Three different purification methods were tested comparatively, and only the one according to Boedetker and Simons [94] (see Table S1 for further details) proved reliably successful, after the necessary numbers and sedimentation parameters of the differential centrifugation cycles had been optimized thoroughly. The particle purity required for an optimal formation of iridescent gels showing diffraction colours was examined in much detail. It was found that the highest purity was achieved through four cycles of centrifugation, whereas the ability of the virions to form the desired gels was optimal after three cycles. This indicated monodispersity - compromised by too excessive repetitive resuspension - as important factor for crystallization. In addition, the study found that storage conditions of the plant material before virion isolation were also crucial for the success of the follow-up experiments. Only plant material frozen for short (overnight) periods led to a reliable applicability of the isolated particles in optical diffraction tests, whereas virions obtained from plant tissues after long-term storage (i.e., frozen for one to five years) yielded 'poor results' [25].
3.2. Methods Worth Being Explored in Future Work
While this review has explored various alternative strategies tested and implemented for tobamovirus purification, it is worth looking at a few innovative techniques that, to our knowledge, seem not to play considerable roles in the enrichment of TMV and related plant viral particles up to now.
Asymmetrical Flow Field-Flow Fractionation (AF4) is one such technology [307]. AF4 instruments employ a channel flow in combination with a cross flow, the latter directed towards a permeable ultrafiltration membrane with a molecular weight cut-off (MWCO) chosen according to the desired particles. Sample components with small diffusion coefficients and higher hydrodynamic diameters travel closer to the membrane along the channel flow, while those with higher diffusion coefficients and lower hydrodynamic diameters travel faster due to the parabolic shape of the laminar channel flow. This method, not requiring virus aggregation or interaction with a stationary phase, offers gentle purification leading to homogeneous particles. Although primarily applied analytically, preparative approaches are also described [308]. Aqueous two-phase systems (ATPS) represent another technique utilizing the separation of two aqueous systems into two phases for liquid-liquid extraction. First experiments with TMV in aqueous two-phase systems are available [309], and continuous ATPS has already enabled the purification of mammalian viral particles (Figure 2E) with high contaminant partitioning capacity [279]. Continuous purification methods are especially interesting because they allow facile scale-up. Last but not least, despite most frequent applications in final polishing steps with discardible devices, membrane chromatography is increasingly valued for virus manufacturing due to its scalability, and may also be used in different flow-through modes depending on the type of device [310]. Membrane chromatography, compared to traditional resin column chromatography, reduces pressure drop and mass transfer resistance [311,312]. This is similar to the advantages of the interconnected channels in monolithic columns, which - apart from uses in bind/elute chromatography (see 2.5.2) - may also be integrated into advanced radial flow devices for upscaling, thereby evading problems with the mechanical instability of disk-shaped monolithic CIM media or pressure increase in elongated columns [310]. An instructive comparison between monolithic, membrane, and conventional chromatography may be found online as application guide [313].
Depending on the specific need, it will be required to test and adapt such new methods for tobamovirus purification to yield highly homogenous particle suspensions or allow scale-up, respectively. The pioneering fundamental research on, and with, tobamoviruses in many labs worldwide has brought along a fast, straightforward method development in the mid-20th century, but only a limited number of main strategies has been followed since then. PEG precipitation remains the most widely adopted method for TMV purification in contemporary laboratories, in combination with a number of centrifugation modes. These strategies were rooted in the favorable virion properties described above, which give rise to efficient separation strategies for the rod-shaped nucleoprotein colloids from most plant components with relatively low effort, in comparison to purification protocols for individual proteins or small biomolecule oligomers. Nonetheless, it should be worth testing the abovementioned non-established methods in combination with high throughput-compatible equipment, as processed and in vitro assembled non-infectious tobamovirus-based particles are attracting increasing attention as biogenic multivalent nanomaterials for many potential uses. Upscaling opportunities would expand the application prospects further.
4. The Proven and the New - Back to the Future? A Case Study
4.1. Motivation and Challenge
Old methods for new virus mutants: In order to find out if novel approaches with engineered tobamoviruses might profit from historic methodology, we performed a case study with TVCVPA. This particular TVCV mutant features an extensive polypeptide, comprised of two protein A domains (E and D, 133 amino acids) attached to its CPs. The size of these extensions is significant enough to be observed in transmission electron microscopy as a granular seam surrounding TVCVPA virions (see below). The PA-domains retain their IgG binding ability when fused C-terminally to the CPs via a long flexible linker ([GGGGS]3) [178]). TVCVPA thus represents a rod-shaped nanoparticle with a predicted length of 317 nm, exhibiting immunosorbent properties and amenable to facile production through molecular farming. Previous studies have demonstrated its capacity to bind morethan 500 IgGs per virion [3], allowing multiple applications in biotechnology, including IgG capture in pull-down assays or magnetic bead separation [241], enzyme immobilization, and potentially biosensors [3].
Given the high potential applications of TVCVPA, we considered it an intriguing specimen for developing a new VNP isolation technique, especially for challenging, potentially more sensitive tobamovirus mutants in future studies. Initially, TVCVPA was isolated from mechanically pressed leaf juice using chloroform extraction, double PEG-precipitation, and low-speed centrifugation [178]. This method yielded approximately 3 g TVCVPA per kg Nicotiana benthamiana leaves with an average virion length of 200–220 nm, mirroring the length of TVCV wildtype isolated by the same procedure. These observed average lengths of both virion types were not in agreement with the anticipated ones of 317 nm and 296 nm, respectively.
Subsequent modifications of the isolation process to enhance scalability included omitting chloroform, conducting a single PEG precipitation, and comparing leaf homogenization techniques using mortar and pestle aided by liquid nitrogen, or a blender [241]. These modifications revealed the susceptibility of both TVCV and TVCVPA (abbreviated 'TVCV(PA)' in the following, if both are addressed) virion integrity to mechanical stress: Particles isolated by this adapted method displayed notably increased average lengths (239 nm) when leaves were ground frozen in a mortar, compared to virions released from the tissues in a blender (217 nm). An iteration of the double PEG precipitation, integrating butanol extraction and ultracentrifugation for the ultimate virion collection, resulted in equally high yields of TVCVPA (2.5 g TVCVPA kg-1 N. benthamiana leaves) of high purity, as evidenced by a minimal background in SDS-PAGE, and a median virion length of approximately 270 nm [3]. However, this length still deviated from the predicted one. Recently, this inconsistency was overcome by using inverse PEG sucrose solubility gradients, as introduced by Clark [1]. In contrast to protocols relying on PEG precipitation/sedimentation, PEG solubility gradient-based isolation of TVCV(PA) avoids pelleting of virions at the tubes' bottom. Virions are precipitated directly in the plant raw homogenate, and pass the gradient up to its region with highest PEG concentration allowing selective resolubilization, where they accumulate in a largely pure band. The early formation of the particle precipitate in the course of the treatments, and its gentle resolvation probably mitigate TVCV(PA)'s susceptibility to mechanically-induced shortening. This resulted in TVCV(PA) particles exhibiting a length distribution closer to their predicted dimensions (largest classes of rod lengths: TVCV 280–300 nm; TVCVPA 300–320 nm), albeit with a slightly reduced yield (1-2 g TVCV(PA) kg-1 N.b. leaves) compared to PEG-precipitation/sedimentation methods [3].
While the inverse PEG-sucrose solubility gradients yielded TVCVPA and TVCV preparations with elevated purity and integrity, a drawback was that virions were distributed along the gradients quite broadly. Consequently, the harvest of virus-containing gradient fractions produced relatively high sample volumes and rather dilute samples, necessitating gradient fractionation and the testing of each fraction for both purity and virus concentration. To improve the isolation of TVCV(PA) in regard to these aspects, we departed from the PEG-solubility gradient approach and developed a method based on density barriers and precipitate re-solvation in an advantageous separation medium instead, as follows. The detailed description of the method can be found in the Supplemental.
4.2. Strategy
Drawing inspiration from Clark und Lister [2], a hand-drawn sketch from Holger Jeske, whom this article is dedicated to, as well as from animal VLP isolations from plants by Thuenemann et al. [314,315] and Dennis et al. [316], we integrated elements from the classical PEG-precipitation technique introduced by Hebert [103] with contemporary non-ionic density gradient ultracentrifugation. In the new method, an initial PEG precipitation facilitates the processing of extract volumes that were impractical for direct application onto phase systems as in our earlier work [3]. A subsequent phase system was designed to resolubilize the PEG-precipitate, eliminate associated contaminants, concentrate the virus on a density cushion in an overnight ultracentrifugation, and yield preparations ready-to-use for different purposes.
Iodixanol was selected as density medium due to its high density, low viscosity as well as its biocompatible, inert and uncharged characteristics [317]. Employing iodixanol, our primary objectives were to achieve two major improvements over the previous PEG solubility gradient-based isolation method: i) simplify the gradient architecture to reduce isolation workload, and ii) obtain a more concentrated yet clean and undegraded virus sample. We simplified the concept of 'sample-on-top-of-phases', with a number of increasingly dense phases from top to bottom [315,316], to a three-layer density barrier phase system. This rationalization to only three phases was possible by designing the phases' densities accordingly to the buoyant density of the viruses, which had to be determined first within the iodixanol compound. On this basis, specific functionalities and dimensions could be assigned to the three phases of the barrier system: the upper one was adjusted to a density slightly above that of the PEG-precipitated, resuspended sample, to act as a loading support, filler and pre-purification zone that facilitates sample application and retains mainly very low-density constituents. The middle phase served as a retention phase primarily for constituents with a buoyant density lower than that of the virus, other contaminants with low sedimentation velocity. The lower phase acted as a cushion, retaining and concentrating the virus without pelleting, and allowing passage/removal of constituents with higher density than the virus particles. Albeit in our application, all three phases were set up in the same buffer, it is conceivable to prepare every phase with a different buffer constitution, imparting the entire phase system advanced washing and buffer exchange capabilities. Even zones of considerably distinct composition and function might be added by placing them into the phase-system via their density adjusted by iodixanol as needed.
4.3. Results
To ascertain appropriate iodixanol concentrations for the system's phases as schematically depicted in Figure 3 A (middle), it was imperative to determine the buoyant densities of TVCV and TVCVPA first. The buoyant density of TMV in iodixanol was also determined. Isopycnic centrifugation (Figure S1) revealed a buoyant density of 1.205 g/ml (equivalent to 38% iodixanol) for both TVCV and TVCVPA. Notably, despite their significant structural differences, there was no discernible impact on buoyant density. TMV exhibited a buoyant density of 1.215 g/ml (equivalent to 40% iodixanol) in the iodixanol gradient, considerably differing from the reported value of 1.32 g/ml in isopycnic CsCl gradients [16,270].
In accordance with these determined buoyant densities, the cushion designed to retain TVCV(PA) was established at a concentration of 41% iodixanol. The retention phase was initially set to 34% iodixanol in the first runs, as estimated from our former experiences. However, under these conditions, the white opaque TVCV(PA) band formed in the retention phase had migrated only a few millimetres down, with low separation from a greenish band held back on top of the retention phase after centrifugation (13.5 h, 25,000 rpm [106,464 x g]), making it challenging to yield the virus. Thus, the iodixanol concentration in the retention phase was reduced to 32%, which resulted in a well-defined, distinct, white opaque band atop the cushion (Figure 3A). SDS-PAGE analysis confirmed that this band was rich in TVCVPA CP and low in proteinaceous contaminants (Figure 3B, sample 2 of twin tubes 1 & 2). In contrast, the greenish band (Figure 3A, arrow 1) contained a large amount of contaminants and only few CP. Although contaminants could penetrate deeper into the retention phase containing 32% iodixanol, the ample space between the white virus-containing and the greenish, contaminant-rich band facilitated convenient sample collection. Further adjustments, such as increasing the volume of the retention phase and fine-tuning centrifugation speed and time, might potentially improve separation and ease yielding of the virus further. For researchers working with viruses or VLPs exhibiting distinct characteristics, this method may be appropriately adapted by exploring their buoyant density and adjusting the density barrier phases accordingly. Additional purification, if necessary, could probably be accomplished by subjecting the iodixanol-containing virus preparation directly to subsequent isopycnic isolation again in iodixanol, without any intermediate sample treatment.
Our density barrier phase systems of the aforementioned composition yielded 1-2 mg of high-purity TVCVPA, or 1-4 mg TVCV g-1 symptomatic N. benthamiana leaves. This range aligns with the observations for TVCVPA isolated via stepwise enrichment through butanol extraction, two-fold PEG precipitation, and ultracentrifugation. Virion purity as indicated by SDS-PAGE analysis (Figure 3 B), and particle integrity as obvious from TEM images (Figure 3 C, D), were both high. Imporantly, the storage stability of TVCVPA in iodixanol-containing solutions aspirated from the ultracentrifugation tubes was substantially superior compared to previously described methods. Those experienced a significant decrease of the TVCVPA-CP size within several days, not preventable by protease inhibitors [178, and own experience]. Upon storage in iodixanol-supplemented media, we did not observe such a size shift in SDS-PAGE after more than five months of storage at 4 °C without preservative additives.
Iodixanol thus has emerged in this work as a convincing candidate for tuning densities of purification phases in gradients, eliminating the necessity for a removal in the downstream applications tested. For example, TVCVPA diluted from the resulting stock solutions of approximately 32% iodixanol, were successfully immobilized in 96-well plates directly, and retained their IgG-binding capabilities, as activity measurements of HRP immobilized on TVCVPA via IgGs demonstrated (Figure S2). Immobilisation at high surface densities on polyelectrolyte-(polyallylamine hydrochloride, PAH)-coated Ta2PO5 chips for potential biosensor applications was possible, similar to previous observations for TVCVPA isolated via PEG sucrose solubility gradients [3]. The residual iodixanol did not affect TEM image quality either (Figure 3 C, D). A minor constraint is the high UV-absorbance of iodixanol, which precludes direct spectrometric determination of TVCV(PA)-concentrations so that indirect methods via protein quantification, or comparative band intensity analyses on SDS-PA gels are needed.
4.4. Perspectives for the Novel Method
Out of the increasing number of tobamovirus mutants and VLPs gaining prominence in nanobiotechnology, some demonstrate sensitivity to conventional isolation approaches, as seen with integrity issues in TVCV(PA) or reduced yield for engineered TMV-VLPs [195]. The method described here may offer an alternative route to establish optimized and convenient, versatile protocols. It facilitates isolations with fair yield and good quality, is customizable in terms of additives, ensures compatibility of residual components with most downstream applications, and offers scalability by using two to six ultracentrifugation rotor buckets available in capacities from a few milliliters up to several dozen milliliters per tube.
5. An Attempt to a Conclusion
This review has highlighted significant advancements in tobamovirus purification, demonstrating a wide array of techniques and their combinations, among which PEG-mediated virion precipitation is notable for its cost-effectiveness and ability to yield a highly purified virus, a method in use for now over 60 years [103]. Despite the vast potential applications of tobamoviruses in fields ranging from medicine to energy storage, the prospects for their commercialization remains elusive and will depend on both regulatory issues and industrial maturity of individual approaches. Many patents have been granted in the area of tobamovirus-assisted technologies, clinical trials are promising, and the urgent worldwide need for 'green', sustainable processes and novel biodegradable materials has increased the efforts for repurposing plant viruses as tools and biogenic building blocks further, with budding prospects [126,318,319,320]. For biomedical applications, the development of glycosylation-engineered 'humanized' N. benthamiana lines is an important additional milestone achieved in recent years [321].
All novel uses and market success of plant-harvested virues will be affected by their specific purification history, which determines particle quality and availability, underscoring the importance of this comprehensive review of methods. For industrial-scale production, scalability becomes paramount. Historical approaches like continuous ultracentrifugation, capable of producing up to 15 g of four-fold precipitate virus from plant juice within 10 h [260], are now seen as difficult to scale, and maintenance-intensive. Potential alternative, extensively scalable methods might include solvent extraction with countercurrent flow for clarification, straightforward large-scale PEG precipitation using tangential flow filtration, and subsequent purification by membrane or monolithic column chromatography. Preservation and fomulation techniques such as spray drying or spray-freeze-drying may also be employed.
Taken together, the methods described in this review can provide solid foundations for a development of large-scale purification processes once commercially viable products are identified. Beyond technical feasibility and product quality, cost-effectiveness remains crucial. An initial calculation suggests surprisingly low production costs already for conventional TMV purification at pilot-scale, sufficient for several potential real-world applications of functionalized virions, e.g., in biosensors or for medical drug delivery [114]. Re-visiting the continuously expanded field of options for customized large-scale processes might thus lay the groundwork for economically competitive uses of tobamoviral particles.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization - review sections, C.W., T.W., K.H., T.K., F.E. - case study, T.W.; methodology and investigation - experimental work, T.W., B.B., K.H. - review sections, T.W., K.H., T.K., F.E., C.W.; writing - original draft preparation, T.W., B.B., K.H., T.K., F.E., C.W.; writing - review and editing T.W., K.H., T.K., F.E., C.W.; supervision, C.W.; project administration, C.W.; funding acquisition, C.W.. All authors have read and agreed to the published version of the manuscript.
Funding
Part of this work was funded by the Deutsche Forschungsgemeinschaft (DFG: German Research Foundation), project number 446507449.
Acknowledgments
The authors thank Annika Allinger, Nadja Beuttenmüller and Laura Markle for taking care of the plants, and Rebecca Hummel for her assistance in lab maintenance, all at the University of Stuttgart. Special thanks go to Heinz Schwarz (MPI for Biology Tübingen) for providing fluorescence and electron microscopy images of infected plant leaves.
References
- Clark, F. Polyethylene glycol solubility gradients, a new and rapid technique for investigations of plant viruses. Virology 1970, 42, 246–247. [Google Scholar] [CrossRef]
- Clark, M.F.; Lister, R.M. The application of polyethylene glycol solubility-concentration gradients in plant virus research. Virology 1971, 43, 338–351. [Google Scholar] [CrossRef]
- Wendlandt, T.; Koch, C.; Britz, B.; Liedek, A.; Schmidt, N.; Werner, S.; Gleba, Y.; Vahidpour, F.; Welden, M.; Poghossian, A.; et al. Facile purification and use of tobamoviral nanocarriers for antibody-mediated display of a two-enzyme system. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Altintoprak, K.; Seidenstücker, A.; Krolla-Sidenstein, P.; Plettl, A.; Jeske, H.; Gliemann, H.; Wege, C. RNA-stabilized protein nanorings: high-precision adapters for biohybrid design. Bioinspired, Biomimetic Nanobiomater. 2017, 6, 208–223. [Google Scholar] [CrossRef]
- Schenk, A.; Eiben, S.; Goll, M.; Reith, L.; Kulak, A.N.; Meldrum, F.; Jeske, H.; Wege, C.; Ludwigs, S. Virus-directed formation of electrocatalytically active nanoparticle-based Co3O4 tubes. Nanoscale 2017, 9, 6334–6345. [Google Scholar] [CrossRef]
- Schneider, A.; Eber, F.J.; Wenz, N.L.; Altintoprak, K.; Jeske, H.; Eiben, S.; Wege, C. Dynamic DNA-controlled "stop-and-go" assembly of well-defined protein domains on RNA-scaffolded TMV-like nanotubes. Nanoscale 2016, 8, 19853–19866. [Google Scholar] [CrossRef]
- Wenz, N.; Piasecka, S.; Kalinowski, M.; Schneider, A.; Richert, C.; Wege, C. Building expanded structures from tetrahedral DNA branching elements, RNA and TMV protein. Nanoscale 2018, 10, 6496–6510. [Google Scholar] [CrossRef]
- Eber, F.J.; Eiben, S.; Jeske, H.; Wege, C. RNA-controlled assembly of tobacco mosaic virus-derived complex structures: from nanoboomerangs to tetrapods. Nanoscale 2015, 7, 344–355. [Google Scholar] [CrossRef]
- Eber, F.J.; Eiben, S.; Jeske, H.; Wege, C. Bottom-up-assembled nanostar colloids of gold cores and tubes derived from tobacco mosaic virus. Angew. Chem. Int. Ed. Engl. 2013, 52, 7203–7207. [Google Scholar] [CrossRef]
- Eiben, S.; Stitz, N.; Eber, F.; Wagner, J.; Atanasova, P.; Bill, J.; Wege, C.; Jeske, H. Tailoring the surface properties of tobacco mosaic virions by the integration of bacterially expressed mutant coat protein. Virus Res. 2014, 180, 92–96. [Google Scholar] [CrossRef]
- Koch, C.; Eber, F.J.; Azucena, C.; Foerste, A.; Walheim, S.; Schimmel, T.; Bittner, A.; Jeske, H.; Gliemann, H.; Eiben, S.; et al. Novel roles for well-known players: from tobacco mosaic virus pests to enzymatically active assemblies. Beilstein J. Nanotechnol. 2016, 7, 613–629. [Google Scholar] [CrossRef]
- Altintoprak, K.; Seidenstücker, A.; Welle, A.; Eiben, S.; Atanasova, P.; Stitz, N.; Plettl, A.; Bill, J.; Gliemann, H.; Jeske, H.; et al. Peptide-equipped tobacco mosaic virus templates for selective and controllable biomineral deposition Beilstein J. Nanotechnol. 2015, 6, 1399–1412. [Google Scholar] [CrossRef]
- Geiger, F.C.; Eber, F.J.; Eiben, S.; Mueller, A.; Jeske, H.; Spatz, J.P.; Wege, C. TMV nanorods with programmed longitudinal domains of differently addressable coat proteins. Nanoscale 2013, 5, 3808–3816. [Google Scholar] [CrossRef]
- Atanasova, P.; Rothenstein, D.; Schneider, J.J.; Hoffmann, R.C.; Dilfer, S.; Eiben, S.; Wege, C.; Jeske, H.; Bill, J. Virus-templated synthesis of ZnO nanostructures and formation of field-effect transistors. Adv. Mater. 2011, 23, 4918–4922. [Google Scholar] [CrossRef]
- Mueller, A.; Eber, F.J.; Azucena, C.; Petershans, A.; Bittner, A.M.; Gliemann, H.; Jeske, H.; Wege, C. Inducible site-selective bottom-up assembly of virus-derived nanotube arrays on RNA-equipped wafers. ACS Nano 2011, 5, 4512–4520. [Google Scholar] [CrossRef]
- Kadri, A.; Maiss, E.; Amsharov, N.; Bittner, A.M.; Balci, S.; Kern, K.; Jeske, H.; Wege, C. Engineered Tobacco mosaic virus mutants with distinct physical characteristics in planta and enhanced metallization properties. Virus Res. 2011, 157, 35–46. [Google Scholar] [CrossRef]
- Mueller, A.; Kadri, A.; Jeske, H.; Wege, C. In vitro assembly of Tobacco mosaic virus coat protein variants derived from fission yeast expression clones or plants. J. Virol. Methods 2010, 166, 77–85. [Google Scholar] [CrossRef]
- Balci, S.; Bittner, A.M.; Schirra, M.; Thonke, K.; Sauer, R.; Hahn, K.; Kadri, A.; Wege, C.; Jeske, H.; Kern, K. Catalytic coating of virus particles with zinc oxide. Electrochimica Acta 2009, 54, 5149–5154. [Google Scholar] [CrossRef]
- Balci, S.; Leinberger, D.M.; Knez, M.; Bittner, A.M.; Boes, F.; Kadri, A.; Wege, C.; Jeske, H.; Kern, K. Printing and aligning mesoscale patterns of tobacco mosaic viruses on surfaces. Adv. Mater. 2008, 20, 2195–2200. [Google Scholar] [CrossRef]
- Balci, S.; Noda, K.; Bittner, A.M.; Kadri, A.; Wege, C.; Jeske, H.; Kern, K. Self-assembly of metal-virus nanodumbbells. Angew. Chem. Int. Ed. Engl. 2007, 46, 3149–3151. [Google Scholar] [CrossRef]
- Knez, M.; Bittner, A.M.; Boes, F.; Wege, C.; Jeske, H.; Maiss, E.; Kern, K. Biotemplate synthesis of 3-nm nickel and cobalt nanowires. Nano Lett. 2003, 3, 1079–1082. [Google Scholar]
- Knez, M.; Sumser, M.; Bittner, A.M.; Wege, C.; Jeske, H.; Kooi, S.; Burghard, M.; Kern, K. Electrochemical modification of individual nano-objects. Journal of Electroanalytical Chemistry 2002, 522, 70–74. [Google Scholar] [CrossRef]
- Steere, R.L. The purification of plant viruses. In Adv. Virus Res., Smith, K.M., Lauffer, M.A., Eds.; Academic Press: 1959; Volume 6, pp. 1–73.
- Stanley, W.M. Chemical studies on the virus of tobacco mosaic: VIII. The isolation of a crystalline protein possessing the properties of aucuba mosaic virus. J. Biol. Chem. 1937, 117, 325–340. [Google Scholar]
- Kreibig, U.; Wetter, C. Light diffraction of in vitro crystals of six tobacco mosaic viruses. Z. Naturforsch. Sect. C 1980, 35, 750–762. [Google Scholar] [CrossRef]
- Bos, L. Beijerinck's work on tobacco mosaic virus: historical context and legacy. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 1999, 354, 675–685. [Google Scholar] [CrossRef]
- Denyer, S.P.; Hodges, N.A. 12.4 Filtration sterilization. Russell, Hugo & Ayliffe's Principles and Practice of Disinfection, Preservation and Sterilization.
- Beijerinck, M.W. Über ein contagium vivum fluidum als Ursache der Fleckenkrankheit der Tabaksblätter. Verhandel. Koninkl. Akad. van Wettenschappen te Amsterdam 1898, 5, 3–21. [Google Scholar]
- Ivanowski, D. Ueber die Mosaikkrankheit der Tabakpflanze / Concerning the mosaic disease of the tobacco plant (translation). Bulletin de l’Académie Impériale des Sciences de Saint-Pétersbourg/ translated in: Phytopathological Classics, 1892; 35. [Google Scholar]
- Lecoq, H. Découverte du premier virus, le virus de la mosaïque du tabac : 1892 ou 1898 ? Comptes Rendus de l'Académie des Sciences - Series III - Sciences de la Vie 2001, 324, 929–933. [Google Scholar]
- Lustig, A.; Levine, A.J. One hundred years of virology. J. Virol. 1992, 66, 4629–4631. [Google Scholar] [CrossRef]
- Scholthof, K.B. Tobacco mosaic virus: a model system for plant biology. Annu. Rev. Phytopathol. 2004, 42, 13–34. [Google Scholar] [CrossRef]
- Creager, A.N.H. Tobacco Mosaic Virus and the History of Molecular Biology. Annual Review of Virology 2022, 9, 39–55. [Google Scholar] [CrossRef]
- Scholthof, K.-B.G.; Shaw, J.G.; Zaitlin, M. , (Eds.) Tobacco mosaic virus: One hundred years of contributions to virology. APS Press, The American Phytopathological Society: St Paul, Minnesota, USA, 1999.
- Bos, L. 100 years of virology: from vitalism via molecular biology to genetic engineering. Trends Microbiol. 2000, 8, 82–87. [Google Scholar] [CrossRef]
- van der Want, J.P.H.; Dijkstra, J. A history of plant virology. Arch. Virol. 2006, 151, 1467–1498. [Google Scholar] [CrossRef]
- Zerbini, F.M.; Kitajima, E.W. From Contagium vivum fluidum to Riboviria: A tobacco mosaic virus-centric history of virus taxonomy. Biomolecules 2022, 12, 1363. [Google Scholar] [CrossRef]
- Baur, E. Über die infektiöse Chlorose der Malvaceen. Kgl. Preuss. Akad. Wiss. 1906, 11–29. [Google Scholar]
- Wege, C.; Gotthardt, R.D.; Frischmuth, T.; Jeske, H. Fulfilling Koch's postulates for Abutilon mosaic virus. Arch. Virol. 2000, 145, 2217–2225. [Google Scholar] [CrossRef]
- Smith, E.F. Ueber die Mosaikkrankheit des Tabaks by Adolf Mayer. The Journal of Mycology 1894, 7, 382–385. [Google Scholar] [CrossRef]
- Koning, C.J. Die Flecken- oder Mosaikkrankheit des holländischen Tabaks. Zeitschrift für Pflanzenkrankheiten 1899, 9, 65–80. [Google Scholar]
- Bechhold, H. Die Gallertfiltration (Ultrafiltration.). Zeitschrift für Chemie und Industrie der Kolloide 1907, 2, 33–41. [Google Scholar]
- Holmes, F.O. Local lesions in tobacco mosaic. Bot. Gaz. 1929, 87, 39–55. [Google Scholar] [CrossRef]
- Purdy, H.A. Immunologic reactions with tobacco mosaic virus. J. Exp. Med. 1929, 49, 919–935. [Google Scholar] [CrossRef]
- Zaitlin, M.; Palukaitis, P. Advances in understanding plant viruses and virus diseases. Annu. Rev. Phytopathol. 2000, 38, 117–143. [Google Scholar] [CrossRef]
- Vinson, C. Possible chemical nature of tobacco mosaic virus. Science 1934, 79, 548–549. [Google Scholar] [CrossRef]
- Stanley, W.M.; Loring, H.S. The isolation of crystalline tobacco mosaic virus protein from diseased tomato plants. Science 1936, 83, 85. [Google Scholar] [CrossRef]
- Stanley, W.M. Isolation of a crystalline protein possessing the properties of tobacco-mosaic virus. Science 1935, 81, 644–645. [Google Scholar] [CrossRef]
- Stanley, W. Isolation and properties of tobacco mosaic and other virus proteins: harvey lecture, March 17, 1938. Bulletin of the New York Academy of Medicine 1938, 14, 398. [Google Scholar]
- Svedberg, T.; Rinde, H. The ultra-centrifuge, a new instrument for the determination of size and distribution of size of particle in amicroscopic colloids. J. Am. Chem. Soc. 1924, 46, 2677–2693. [Google Scholar]
- Biscoe, J.; Pickels, E.G.; Wyckoff, R.W. An air-driven ultracentrifuge for molecular sedimentation. J. Exp. Med. 1936, 64, 39–45. [Google Scholar] [CrossRef]
- Eriksson-Quensel, I.-B.; Svedberg, T. Sedimentation and electrophoresis of the tobacco mosaic virus protein. J. Am. Chem. Soc. 1936, 58, 1863–1867. [Google Scholar] [CrossRef]
- Wyckoff, R.W.G. The ultracentrifugal study of virus proteins. Proceedings of the American Philosophical Society 1937, 77, 455–462. [Google Scholar]
- Bawden, F.C.; Pirie, N.W.; Bernal, J.D.; Fankuchen, I. Liquid crystalline substances from virus infected plants. Nature 1936, 138, 1051–1052. [Google Scholar]
- Bawden, F.C.; Pirie, N.W. The isolation and some properties of liquid crystalline substances from solanaceous plants infected with three strains of tobacco mosaic virus. Proceedings of the Royal Society of London. Series B, Biological sciences 1937, 123, 274–320. [Google Scholar] [CrossRef]
- Pennazio, S.; Roggero, P. The discovery of the chemical nature of tobacco mosaic virus. Riv Biol 2000, 93, 253–281. [Google Scholar]
- Bernal, J.D.; Fankuchen, I. Structure types of protein 'crystals' from virus infected plants. Nature 1937, 139, 923–924. [Google Scholar]
- Kausche, G.; Guggisberg, H.; Wissler, A. Quantitative Untersuchung der Strömungsdoppel-brechung von Tabakmosaik-und Kartoffel-X-Virus. Naturwissenschaften 1939, 27, 303–304. [Google Scholar]
- Lauffer, M.A. The electro-optical effect in certain viruses. J. Am. Chem. Soc. 1939, 61, 2412–2416. [Google Scholar]
- Robinson, J. Shape of tobacco mosaic virus particles in solution. Nature 1939, 143, 923–926. [Google Scholar] [CrossRef]
- Robinson, J. Studies in the viscosity of colloids. I. The anomalous viscosity of dilute suspensions of rigid anisometric particles. Proceedings of the Royal Society of London. Series A. Mathematical and Physical Sciences 1939, 170, 519–550. [Google Scholar]
- Bernal, J.D.; Fankuchen, I. X-Ray and crystallographic studies of plant virus preparations : I. Introduction and preparation of specimens ii. Modes of aggregation of the virus particles. The Journal of General Physiology 1941, 25, 111–146. [Google Scholar] [CrossRef]
- Kausche, G. Über die Charakterisierung von pflanzlichen Virussolen mit kolloidem Gold. Naturwissenschaften 1938, 26, 445–445. [Google Scholar]
- Adams, M.; Fraden, S. Phase behavior of mixtures of rods (tobacco mosaic virus) and spheres (polyethylene oxide, bovine serum albumin). Biophys. J. 1998, 74, 669–677. [Google Scholar] [CrossRef]
- Liljeström, V.; Ora, A.; Hassinen, J.; Rekola, H.T.; Nonappa; Heilala, M. ; Hynninen, V.; Joensuu, J.J.; Ras, R.H.A.; Törmä, P.; et al. Cooperative colloidal self-assembly of metal-protein superlattice wires. Nature Communications 2017, 8, 671. [Google Scholar] [CrossRef]
- Wu, Z.; Mueller, A.; Degenhard, S.; Ruff, S.E.; Geiger, F.; Bittner, A.; Wege, C.; Krill III, C. Enhancing the magnetoviscosity of ferrofluids by the addition of biological nanotubes. ACS Nano 2010, 4, 4531–4538. [Google Scholar] [CrossRef]
- Khan, A.A.; Fox, E.K.; Gorzny, M.L.; Nikulina, E.; Brougham, D.F.; Wege, C.; Bittner, A.M. pH Control of the Electrostatic Binding of Gold and Iron Oxide Nanoparticles to Tobacco Mosaic Virus. Langmuir 2013, 29, 2094–2098. [Google Scholar] [CrossRef]
- Bawden, F.C.; Pirie, N.W. Methods for the purification of tomato bushy stunt and tobacco mosaic viruses. Biochem. J. 1943, 37, 66–70. [Google Scholar] [CrossRef]
- Pfankuch, E.; Hagenguth, K. Isolierung einiger pflanzlicher Viren. Naturwissenschaften 1943, 31, 370–371. [Google Scholar]
- Lauffer, M.A. The size and shape of tobacco mosaic virus particles1. J. Am. Chem. Soc. 1944, 66, 1188–1194. [Google Scholar]
- Gelderblom, H.R.; Krüger, D.H. Helmut Ruska (1908–1973): his role in the evolution of electron microscopy in the life sciences, and especially virology. In Advances in imaging and electron physics, Hawkes, P.W., Ed.; Elsevier: 2014; Volume Volume 182, pp. 1–94.
- Kausche, G.A.; Pfankuch, E.; Ruska, H. Die Sichtbarmachung von pflanzlichem Virus im Übermikroskop. Naturwissenschaften 1939, 27, 292–299. [Google Scholar]
- Creager, A.N. The life of a virus: Tobacco mosaic virus as an experimental model, 1930-1965; University of Chicago Press: 2002.
- Creager, A.N.; Morgan, G.J. After the double helix: Rosalind Franklin's research on Tobacco mosaic virus. Isis 2008, 99, 239–272. [Google Scholar] [CrossRef]
- Franklin, R.E.; Klug, A.; Holmes, K.C. X-ray diffraction studies of the structure and morphology of tobacco mosaic virus. In Proceedings of the Ciba Foundation Symposium on the Nature of Viruses, London; 1957; pp. 39–55. [Google Scholar]
- Klein, J. Max-Planck-Institut für Biologie, Tübingen: Die Geschichte des Instituts von 1912 bis 1983; Tübingen, 1983; p. 71.
- Schramm, G. Zur Chemie des Mutationsvorganges beim Tabakmosaik-Virus. Zeitschrift für Naturforschung B 1948, 3, 320–327. [Google Scholar]
- Lewis, J. From virus research to molecular biology: Tobacco mosaic virus in Germany, 1936-1956. Journal of the History of Biology 2004, 37, 259–301. [Google Scholar] [CrossRef]
- Fraenkel-Conrat, H.; Singer, B.; Williams, R. Infectivity of viral nucleic acid. Biochim. Biophys. Acta 1957, 25, 87–96. [Google Scholar] [CrossRef]
- Watanabe, I.; Kawade, Y. Purification and characterization of tobacco mosaic virus. Bulletin of the Chemical Society of Japan, 2006; 26. [Google Scholar] [CrossRef]
- Fukushi, T.; Shikata, E. Electron microscope studies on plant viruses I. Journal of the Faculty of Agriculture, Hokkaido University 1955, 50, 74–94. [Google Scholar]
- Ryjkoff, V.; Smirnova, M.V. Liquid crystals of the virus of the tobacco mosaic (Nicotiana virus 1 Allard). Doklady Akademii nauk SSSR 1941, 31, 930–932. [Google Scholar]
- Ryzkov, V.; Tarasevic, L.; Loïdina, G. The effect of strong solutions of sodium nucleinate on the virus of the mosaic disease of tobacco (VTM) and on albumen. Doklady Akademii nauk SSSR 1950, 74, 1023–1024. [Google Scholar]
- Carr, J.P. Tobacco mosaic virus. In Plant-Pathogen Interactions, Talbot, N.J., Ed.; Annual Plant Reviews; Blackwell Publishers: Oxford, U.K, 2004; pp. 25–67. [Google Scholar]
- Hamilton, R.I.; Edwardson, J.R.; Francki, R.I.B.; Hsu, H.T.; Hull, R.; Koenig, R.; Milne, R.G. Guidelines for the identification and characterization of plant viruses. J. Gen. Virol. 1981, 54, 223–241. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H.V. Antigenic relationships between strains of tobacco mosaic virus. Virology 1975, 64, 415–420. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H.V. Tobacco mosaic virus antigenic structure. In The plant viruses: The rod-shaped plant viruses, Van Regenmortel, M.H.V., Fraenkel-Conrat, H., Eds.; Springer US: Boston, MA, 1986; pp. 79–104. [Google Scholar]
- Smith, K.M. On the composite nature of certain potato virus diseases of the mosaic group as revealed by the use of plant indicators and selective methods of transmission. Proceedings of the Royal Society of London. Series B, Containing Papers of a Biological Character 1931, 109, 251–267. [Google Scholar] [CrossRef]
- Mehetre, G.T.; Leo, V.V.; Singh, G.; Sorokan, A.; Maksimov, I.; Yadav, M.K.; Upadhyaya, K.; Hashem, A.; Alsaleh, A.N.; Dawoud, T.M.; et al. Current developments and challenges in plant viral diagnostics: A systematic review. Viruses 2021, 13, 412. [Google Scholar] [CrossRef]
- Bernabé-Orts, J.M.; Torre, C.; Méndez-López, E.; Hernando, Y.; Aranda, M.A. New resources for the specific and sensitive detection of the emerging tomato brown rugose fruit virus. Viruses 2021, 13, 1680. [Google Scholar] [CrossRef]
- Mandal, S.; Mandal, B.; Haq, Q.; Varma, A. Properties, diagnosis and management of Cucumber green mottle mosaic virus. Plant viruses 2008, 2, 25–34. [Google Scholar]
- Wilkins, M.; Stokes, A.; Seeds, W.; Oster, G. Tobacco mosaic virus crystals and three-dimensional microscopic vision. Nature 1950, 166, 127–129. [Google Scholar] [CrossRef]
- Klug, A. The tobacco mosaic virus particle: structure and assembly. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 1999, 354, 531–535. [Google Scholar] [CrossRef]
- Boedtker, H.; Simmons, N.S. The preparation and characterization of essentially uniform tobacco mosaic virus particles. J. Am. Chem. Soc. 1958, 80, 2550–2556. [Google Scholar] [CrossRef]
- Kendall, A.; Stubbs, G. Fibre diffraction in the analysis of filamentous virus structure. Crystallogr. Rev. 2016, 22, 84–101. [Google Scholar] [CrossRef]
- Gebhardt, R.; Teulon, J.-M.; Pellequer, J.-L.; Burghammer, M.; Colletier, J.-P.; Riekel, C. Virus particle assembly into crystalline domains enabled by the coffee ring effect. Soft Matter 2014, 10, 5458–5462. [Google Scholar] [CrossRef]
- Wang, H.; Planchart, A.; Allen, D.; Pattanayek, R.; Stubbs, G. Preliminary X-ray diffraction studies of ribgrass mosaic virus. J. Mol. Biol. 1993, 234, 902–904. [Google Scholar] [CrossRef]
- Fromm, S.A.; Bharat, T.A.; Jakobi, A.J.; Hagen, W.J.; Sachse, C. Seeing tobacco mosaic virus through direct electron detectors. Journal of Structural Biology 2015, 189, 87–97. [Google Scholar] [CrossRef]
- Song, B.; Lenhart, J.; Flegler, V.J.; Makbul, C.; Rasmussen, T.; Böttcher, B. Capabilities of the Falcon III detector for single-particle structure determination. Ultramicroscopy 2019, 203, 145–154. [Google Scholar]
- Lewis, J.K.; Bendahmane, M.; Smith, T.J.; Beachy, R.N.; Siuzdak, G. Identification of viral mutants by mass spectrometry. Proc. Natl. Acad. Sci. USA 1998, 95, 8596–8601. [Google Scholar] [CrossRef]
- Lumata, J.L.; Ball, D.; Shahrivarkevishahi, A.; Luzuriaga, M.A.; Herbert, F.C.; Brohlin, O.; Lee, H.; Hagge, L.M.; D'Arcy, S.; Gassensmith, J.J. Identification and physical characterization of a spontaneous mutation of the tobacco mosaic virus in the laboratory environment. Sci. Rep. 2021, 11, 15109. [Google Scholar] [CrossRef]
- Gooding, G.V., Jr.; Hebert, T.T. A simple technique for purification of tobacco mosaic virus in large quantities. Phytopathology 1967, 57, 1285. [Google Scholar]
- Hebert, T.T. Precipitation of plant viruses by polyethylene glycol. Phytopathology 1963, 53, 362. [Google Scholar]
- Fraile, A.; Escriu, F.; Aranda, M.A.; Malpica, J.M.; Gibbs, A.J.; García-Arenal, F. A century of tobamovirus evolution in an Australian population of Nicotiana glauca. J. Virol. 1997, 71, 8316–8320. [Google Scholar] [CrossRef]
- Gibbs, A. Evolution and origins of tobamoviruses. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 1999, 354, 593–602. [Google Scholar] [CrossRef]
- Gibbs, A.J.; Wood, J.; Garcia-Arenal, F.; Ohshima, K.; Armstrong, J.S. Tobamoviruses have probably co-diverged with their eudicotyledonous hosts for at least 110 million years. Virus Evol. 2015, 1, vev019. [Google Scholar] [CrossRef]
- Wilson, T.M.A. Plant viruses: A tool-box for genetic engineering and crop protection. Bioessays 1989, 10, 179–186. [Google Scholar] [CrossRef]
- Haynes, J.R.; Cunningham, J.; von Seefried, A.; Lennick, M.; Garvin, R.T.; Shen, S.-H. Development of a genetically–engineered, candidate polio vaccine employing the self–assembling properties of the tobacco mosaic virus coat protein. Bio/Technology 1986, 4, 637. [Google Scholar] [CrossRef]
- Lomonossoff, G.P.; Wege, C. Chapter Six - TMV particles: the journey from fundamental studies to bionanotechnology applications. In Adv. Virus Res., Palukaitis, P., Roossinck, M.J., Eds.; Academic Press: 2018; Volume 102, pp. 149–176.
- Wen, A.M.; Lee, K.L.; Steinmetz, N.F. Plant Virus-Based Nanotechnologies. In Women in Nanotechnology: Contributions from the Atomic Level and Up, Norris, P.M., Friedersdorf, L.E., Eds.; Springer International Publishing: Cham, 2020; pp. 57–69. [Google Scholar]
- Zhang, J.; He, H.; Zeng, F.; Du, M.; Huang, D.; Chen, G.; Wang, Q. Advances of structural design and biomedical applications of tobacco mosaic virus coat protein. Adv. Nanobiomed Res. 2024, n/a, 2300135. [Google Scholar] [CrossRef]
- Fan, X.Z.; Pomerantseva, E.; Gnerlich, M.; Brown, A.; Gerasopoulos, K.; McCarthy, M.; Culver, J.; Ghodssi, R. Tobacco mosaic virus: A biological building block for micro/nano/biosystems. J. Vac. Sci. Technol., A 2013, 31, 050815. [Google Scholar] [CrossRef]
- Culver, J.N.; Brown, A.D.; Zang, F.; Gnerlich, M.; Gerasopoulos, K.; Ghodssi, R. Plant virus directed fabrication of nanoscale materials and devices. Virology 2015, 479–480, 200–212. [Google Scholar] [CrossRef]
- Wege, C.; Koch, C. From stars to stripes: RNA-directed shaping of plant viral protein templates—structural synthetic virology for smart biohybrid nanostructures. WIREs Nanomedicine and Nanobiotechnology 2020, 12, e1591. [Google Scholar] [CrossRef]
- Dragnea, B. Virus-based devices: prospects for allopoiesis. ACS Nano 2017, 11, 3433–3437. [Google Scholar] [CrossRef]
- Steele, J.F.C.; Peyret, H.; Saunders, K.; Castells-Graells, R.; Marsian, J.; Meshcheriakova, Y.; Lomonossoff, G.P. Synthetic plant virology for nanobiotechnology and nanomedicine. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2017, 9, e1447. [Google Scholar] [CrossRef]
- Lin, B.; Ratna, B. Virus hybrids as nanomaterials : methods and protocols; Humana Press - Springer: New York, Heidelberg, Dordrecht, London, 2014. [Google Scholar]
- Nkanga, C.I.; Steinmetz, N.F. The pharmacology of plant virus nanoparticles. Virology 2021, 556, 39–61. [Google Scholar]
- Czapar, A.E.; Steinmetz, N.F. Plant viruses and bacteriophages for drug delivery in medicine and biotechnology. Curr. Opin. Chem. Biol. 2017, 38, 108–116. [Google Scholar] [CrossRef]
- Khudyakov, Y.; Pumpens, P. Viral Nanotechnology; CRC Press, Taylor & Francis: Boca Raton, London, New York, 2016. [Google Scholar]
- Arul, S.S.; Balakrishnan, B.; Handanahal, S.S.; Venkataraman, S. Viral nanoparticles: Current advances in design and development. Biochimie 2024, 219, 33–50. [Google Scholar]
- Eiben, S.; Koch, C.; Altintoprak, K.; Southan, A.; Tovar, G.; Laschat, S.; Weiss, I.; Wege, C. Plant virus-based materials for biomedical applications: trends and prospects. Adv. Drug Del. Rev. 2019, 145, 96–118. [Google Scholar] [CrossRef]
- Chariou, P.L.; Ortega-Rivera, O.A.; Steinmetz, N.F. Nanocarriers for the Delivery of Medical, Veterinary, and Agricultural Active Ingredients. ACS Nano 2020, 14, 2678–2701. [Google Scholar] [CrossRef]
- Takeyama, N.; Kiyono, H.; Yuki, Y. Plant-based vaccines for animals and humans: recent advances in technology and clinical trials. Therapeutic Advances in Vaccines 2015, 3, 139–154. [Google Scholar] [CrossRef]
- Kolotilin, I.; Topp, E.; Cox, E.; Devriendt, B.; Conrad, U.; Joensuu, J.; Stoger, E.; Warzecha, H.; McAllister, T.; Potter, A.; et al. Plant-based solutions for veterinary immunotherapeutics and prophylactics. Vet. Res. 2014, 45, 117. [Google Scholar] [CrossRef]
- Chung, Y.H.; Church, D.; Koellhoffer, E.C.; Osota, E.; Shukla, S.; Rybicki, E.P.; Pokorski, J.K.; Steinmetz, N.F. Integrating plant molecular farming and materials research for next-generation vaccines. Nature Reviews Materials 2021, 1–17. [Google Scholar] [CrossRef]
- Crisci, E.; Bárcena, J.; Montoya, M. Virus-like particles: The new frontier of vaccines for animal viral infections. Vet. Immunol. Immunopathol. 2012, 148, 211–225. [Google Scholar] [CrossRef]
- Nikitin, N.; Vasiliev, Y.; Kovalenko, A.; Ryabchevskaya, E.; Kondakova, O.; Evtushenko, E.; Karpova, O. Plant Viruses as Adjuvants for Next-Generation Vaccines and Immunotherapy. Vaccines 2023, 11, 1372. [Google Scholar] [CrossRef]
- Williams, L.; Jurado, S.; Llorente, F.; Romualdo, A.; González, S.; Saconne, A.; Bronchalo, I.; Martínez-Cortes, M.; Pérez-Gómez, B.; Ponz, F.; et al. The C-Terminal Half of SARS-CoV-2 Nucleocapsid Protein, Industrially Produced in Plants, Is Valid as Antigen in COVID-19 Serological Tests. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Azizi, M.; Shahgolzari, M.; Fathi-Karkan, S.; Ghasemi, M.; Samadian, H. Multifunctional plant virus nanoparticles: An emerging strategy for therapy of cancer. WIREs Nanomedicine and Nanobiotechnology 2023, 15, e1872. [Google Scholar]
- Venkataraman, S.; Hefferon, K. Application of plant viruses in biotechnology, medicine, and human health. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Chariou, P.L.; Ma, Y.; Hensley, M.; Rosskopf, E.N.; Hong, J.C.; Charudattan, R.; Steinmetz, N.F. Inactivated plant viruses as an agrochemical delivery platform. ACS Agricultural Science & Technology 2021, 1, 124–130. [Google Scholar] [CrossRef]
- Charudattan, R. Use of plant viruses as bioherbicides: the first virus-based bioherbicide and future opportunities. Pest Manage. Sci. 2024, 80, 103–114. [Google Scholar]
- Schlick, T.L.; Ding, Z.B.; Kovacs, E.W.; Francis, M.B. Dual-surface modification of the tobacco mosaic virus. J. Am. Chem. Soc. 2005, 127, 3718–3723. [Google Scholar] [CrossRef]
- González-Gamboa, I.; Caparco, A.A.; McCaskill, J.; Fuenlabrada-Velázquez, P.; Hays, S.S.; Jin, Z.; Jokerst, J.V.; Pokorski, J.K.; Steinmetz, N.F. Inter-coat protein loading of active ingredients into Tobacco mild green mosaic virus through partial dissociation and reassembly of the virion. Sci. Rep. 2024, 14, 7168. [Google Scholar] [CrossRef]
- Gulati, N.M.; Pitek, A.S.; Steinmetz, N.F.; Stewart, P.L. Cryo-electron tomography investigation of serum albumin-camouflaged tobacco mosaic virus nanoparticles. Nanoscale 2017, 9, 3408–3415. [Google Scholar] [CrossRef]
- Dickmeis, C.; Kauth, L.; Commandeur, U. From infection to healing: The use of plant viruses in bioactive hydrogels. WIREs Nanomed Nanobiotechnol. 2021, 13, e1662. [Google Scholar] [CrossRef]
- Maturavongsadit, P.; Luckanagul, J.A.; Metavarayuth, K.; Zhao, X.; Chen, L.M.; Lin, Y.; Wang, Q. Promotion of in vitro chondrogenesis of mesenchymal stem cells using in situ hyaluronic hydrogel functionalized with rod-like viral nanoparticles. Biomacromolecules 2016, 17, 1930–1938. [Google Scholar] [CrossRef]
- Schuphan, J.; Stojanović, N.; Lin, Y.-Y.; Buhl, E.M.; Aveic, S.; Commandeur, U.; Schillberg, S.; Fischer, H. A combination of flexible modified plant virus nanoparticles enables additive effects resulting in improved osteogenesis. Adv. Healthc. Mater. 2024, n/a, 2304243. [Google Scholar]
- Alonso, J.M.; Górzny, M.Ł.; Bittner, A.M. The physics of tobacco mosaic virus and virus-based devices in biotechnology. Trends Biotechnol. 2013, 31, 530–538. [Google Scholar] [CrossRef]
- Riekel, C.; Burghammer, M.; Snigirev, I.; Rosenthal, M. Microstructural metrology of tobacco mosaic virus nanorods during radial compression and heating. Soft Matter 2018, 14, 194–204. [Google Scholar] [CrossRef]
- Wu, L.Y.; Zang, J.F.; Lee, L.A.; Niu, Z.W.; Horvatha, G.C.; Braxtona, V.; Wibowo, A.C.; Bruckman, M.A.; Ghoshroy, S.; zur Loye, H.C.; et al. Electrospinning fabrication, structural and mechanical characterization of rod-like virus-based composite nanofibers. J Mater Chem 2011, 21, 8550–8557. [Google Scholar] [CrossRef]
- Rahman, M.M.; Ölçeroğlu, E.; McCarthy, M. Biotemplates: Scalable nanomanufacturing of virus-templated coatings for enhanced boiling. Adv. Mater. Interfaces 2014, 1. [Google Scholar] [CrossRef]
- Poghossian, A.; Jablonski, M.; Koch, C.; Bronder, T.S.; Rolka, D.; Wege, C.; Schöning, M.J. Field-effect biosensor using virus particles as scaffolds for enzyme immobilization. Biosensors Bioelectron. 2018, 110, 168–174. [Google Scholar] [CrossRef]
- Koch, C.; Poghossian, A.; Schoening, M.J.; Wege, C. Penicillin detection by tobacco mosaic virus-assisted colorimetric biosensors. Nanotheranostics 2018, 2, 184–196. [Google Scholar] [CrossRef]
- Poghossian, A.; Jablonski, M.; Molinnus, D.; Wege, C.; Schoning, M.J. Field-effect sensors for virus detection: from Ebola to SARS-CoV-2 and plant viral enhancers. Front. Plant Sci. 2020, 11, 598103. [Google Scholar]
- Douglas, T.; Young, M. Virus particles as templates for materials synthesis. Adv. Mater. 1999, 11, 679–681. [Google Scholar] [CrossRef]
- Shenton, W.; Douglas, T.; Young, M.; Stubbs, G.; Mann, S. Inorganic-organic nanotube composites from template mineralization of tobacco mosaic virus. Adv. Mater. 1999, 11, 253–256. [Google Scholar] [CrossRef]
- Young, M.; Willits, D.; Uchida, M.; Douglas, T. Plant viruses as biotemplates for materials and their use in nanotechnology. Annu. Rev. Phytopathol. 2008, 46, 361–384. [Google Scholar] [CrossRef]
- Bittner, A.M.; Alonso, J.M.; Górzny, M.L.; Wege, C. Nanoscale science and technology with plant viruses and bacteriophages. In Structure and physics of viruses: an integrated textbook, Mateu, M.G., Ed.Harris, J.R., Ed.; Subcellular Biochemistry (); Springer Science+Business Media: Dordrecht, 2013; Volume 68, pp. 667–702. ISSN 0306-0225. [Google Scholar]
- Fan, X.Z.; Naves, L.; Siwak, N.P.; Brown, A.; Culver, J.; Ghodssi, R. Integration of genetically modified virus-like-particles with an optical resonator for selective bio-detection. Nanotechnology 2015, 26, 205501. [Google Scholar] [CrossRef]
- Soto, C.M.; Ratna, B.R. Virus hybrids as nanomaterials for biotechnology. Curr. Opin. Biotechnol. 2010, 21, 426–438. [Google Scholar] [CrossRef]
- Chen, Z.; Li, N.; Li, S.; Dharmarwardana, M.; Schlimme, A.; Gassensmith, J.J. Viral chemistry: the chemical functionalization of viral architectures to create new technology. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2016, 8, 512–534. [Google Scholar] [CrossRef]
- Chu, S.; Brown, A.D.; Culver, J.N.; Ghodssi, R. Tobacco mosaic virus as a versatile platform for molecular assembly and device fabrication. Biotechnol. J. 2018, 13, 1800147. [Google Scholar] [CrossRef]
- Balci, S.; Hahn, K.; Kopold, P.; Kadri, A.; Wege, C.; Kern, K.; Bittner, A.M. Electroless synthesis of 3 nm wide alloy nanowires inside Tobacco mosaic virus. Nanotechnology 2012, 23. [Google Scholar] [CrossRef]
- Bittner, A.M. TMV-Templated Formation of Metal and Polymer Nanotubes. In Virus-Derived Nanoparticles for Advanced Technologies: Methods and Protocols, Wege, C., Lomonossoff, G.P., Eds.; Springer New York: New York, NY, 2018; pp. 383–392. [Google Scholar]
- Shah, S.N.; Heddle, J.G.; Evans, D.J.; Lomonossoff, G.P. Production of Metallic Alloy Nanowires and Particles Templated Using Tomato Mosaic Virus (ToMV). Nanomaterials 2023, 13, 2705. [Google Scholar]
- Miller, R.A.; Stephanopoulos, N.; McFarland, J.M.; Rosko, A.S.; Geissler, P.L.; Francis, M.B. Impact of assembly state on the defect tolerance of TMV-based light harvesting arrays. J. Am. Chem. Soc. 2010, 132, 6068–6074. [Google Scholar] [CrossRef]
- Rong, J.H.; Oberbeck, F.; Wang, X.N.; Li, X.D.; Oxsher, J.; Niu, Z.W.; Wang, Q. Tobacco mosaic virus templated synthesis of one dimensional inorganic-polymer hybrid fibres. J Mater Chem 2009, 19, 2841–2845. [Google Scholar] [CrossRef]
- Dönmez Güngüneş, Ç.; Başçeken, S.; Elçin, A.E.; Elçin, Y.M. Fabrication and molecular modeling of navette-shaped fullerene nanorods using tobacco mosaic virus as a nanotemplate. Mol. Biotechnol. 2022, 64, 681–692. [Google Scholar]
- Riccò, R.; Liang, W.; Li, S.; Gassensmith, J.J.; Caruso, F.; Doonan, C.; Falcaro, P. Metal–organic frameworks for cell and virus biology: a perspective. ACS Nano 2018, 12, 13–23. [Google Scholar] [CrossRef]
- Moreira Da Silva, C.; Ortiz-Peña, N.; Boubekeur-Lecaque, L.; Dušek, J.; Moravec, T.; Alloyeau, D.; Ha-Duong, N.-T. In situ insights into the nucleation and growth mechanisms of gold nanoparticles on tobacco mosaic virus. Nano Lett. 2023, 23, 5281–5287. [Google Scholar] [CrossRef]
- Gerasopoulos, K.; McCarthy, M.; Royston, E.; Culver, J.N.; Ghodssi, R. Nanostructured nickel electrodes using the Tobacco mosaic virus for microbattery applications. J Micromech Microeng 2008, 18. [Google Scholar] [CrossRef]
- Royston, E.; Ghosh, A.; Kofinas, P.; Harris, M.T.; Culver, J.N. Self-assembly of virus-structured high surface area nanomaterials and their application as battery electrodes. Langmuir 2008, 24, 906–912. [Google Scholar] [CrossRef]
- Tseng, R.J.; Tsai, C.; Ma, L.P.; Ouyang, J.; Ozkan, C.S.; Yang, Y. Digital memory device based on tobacco mosaic virus conjugated with nanoparticles. Nature nanotechnology 2006, 1, 72–77. [Google Scholar] [CrossRef]
- Yang, C.X.; Choi, C.H.; Lee, C.S.; Yi, H.M. A facile synthesis-fabrication strategy for integration of catalytically active viral-palladium nanostructures into polymeric hydrogel microparticles via replica molding. ACS Nano 2013, 7, 5032–5044. [Google Scholar] [CrossRef]
- Chiang, C.-Y.; Epstein, J.; Brown, A.; Munday, J.N.; Culver, J.N.; Ehrman, S. Biological templates for antireflective current collectors for photoelectrochemical cell applications. Nano Lett. 2012, 12, 6005–6011. [Google Scholar] [CrossRef]
- Lee, K.Z.; Basnayake Pussepitiyalage, V.; Lee, Y.-H.; Loesch-Fries, L.S.; Harris, M.T.; Hemmati, S.; Solomon, K.V. Engineering tobacco mosaic virus and its virus-like-particles for synthesis of biotemplated nanomaterials. Biotechnol. J. 2021, 16, 2000311. [Google Scholar]
- Zhou, Q.; Liu, X.; Tian, Y.; Wu, M.; Niu, Z. Mussel-inspired polydopamine coating on tobacco mosaic virus: one-dimensional hybrid nanofibers for gold nanoparticle growth. Langmuir 2017, 33, 9866–9872. [Google Scholar] [CrossRef]
- Love, A.J.; Makarov, V.; Yaminsky, I.; Kalinina, N.O.; Taliansky, M.E. The use of tobacco mosaic virus and cowpea mosaic virus for the production of novel metal nanomaterials. Virology 2014, 449, 133–139. [Google Scholar] [CrossRef]
- Hou, C.; Xu, H.; Jiang, X.; Li, Y.; Deng, S.; Zang, M.; Xu, J.; Liu, J. Virus-based supramolecular structure and materials: concept and prospects. ACS Appl.. Bio Mater. 2021, 4, 5961–5974. [Google Scholar] [CrossRef]
- Love, A.J.; Makarov, V.V.; Sinitsyna, O.V.; Shaw, J.; Yaminsky, I.V.; Kalinina, N.O.; Taliansky, M. A genetically modified tobacco mosaic virus that can produce gold nanoparticles from a metal salt precursor. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Zakeri, B.; Fierer, J.O.; Celik, E.; Chittock, E.C.; Schwarz-Linek, U.; Moy, V.T.; Howarth, M. Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proceedings of the National Academy of Sciences 2012, 109, E690–E697. [Google Scholar] [CrossRef]
- Schuphan, J.; Commandeur, U. Analysis of engineered tobacco mosaic virus and potato virus X nanoparticles as carriers for biocatalysts. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Lomonossoff, G.P.; Wege, C. Chapter Six - TMV Particles: The Journey From Fundamental Studies to Bionanotechnology Applications. In Advances in Virus Research, Vol. 102, Palukaitis, P., Roossinck, M.J., Eds.; Academic Press: 2018; Volume 102, pp. 149–176.
- Röder, J.; Fischer, R.; Commandeur, U. Adoption of the 2A ribosomal skip principle to tobacco mosaic virus for peptide display. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Frolova, O.Y.; Petrunia, I.V.; Komarova, T.V.; Kosorukov, V.S.; Sheval, E.V.; Gleba, Y.Y.; Dorokhov, Y.L. Trastuzumab-binding peptide display by Tobacco mosaic virus. Virology 2010, 407, 7–13. [Google Scholar] [CrossRef]
- Werner, S.; Marillonnet, S.; Hause, G.; Klimyuk, V.; Gleba, Y. Immunoabsorbent nanoparticles based on a tobamovirus displaying protein A. Proceedings of the National Academy of Sciences USA 2006, 103, 17678–17683. [Google Scholar] [CrossRef]
- Bäcker, M.; Koch, C.; Eiben, S.; Geiger, F.; Eber, F.J.; Gliemann, H.; Poghossian, A.; Wege, C.; Schöning, M.J. Tobacco mosaic virus as enzyme nanocarrier for electrochemical biosensors. Sens. Actuators, B 2017, 238, 716–722. [Google Scholar] [CrossRef]
- Koch, C.; Wabbel, K.; Eber, F.J.; Krolla-Sidenstein, P.; Azucena, C.; Gliemann, H.; Eiben, S.; Geiger, F.; Wege, C. Modified TMV particles as beneficial scaffolds to present sensor enzymes. Frontiers in Plant Science 2015, 6, 1137. [Google Scholar] [CrossRef]
- Welden, M.; Poghossian, A.; Vahidpour, F.; Wendlandt, T.; Keusgen, M.; Christina, W.; Schöning, M.J. Capacitive field-effect biosensor modified with a stacked bilayer of weak polyelectrolyte and plant virus particles as enzyme nanocarriers. Bioelectrochemistry 2023, 151, 108397. [Google Scholar]
- Welden, M.; Poghossian, A.; Vahidpour, F.; Wendlandt, T.; Keusgen, M.; Wege, C.; Schöning, M.J. Towards multi-analyte detection with field-effect capacitors modified with tobacco mosaic virus bioparticles as enzyme nanocarriers. Biosensors 2022, 12, 43. [Google Scholar] [CrossRef]
- Welden, M.; Severins, R.; Poghossian, A.; Wege, C.; Bongaerts, J.; Siegert, P.; Keusgen, M.; Schoning, M.J. Detection of acetoin and diacetyl by a tobacco mosaic virus-assisted field-effect biosensor. Chemosensors 2022, 10, 218. [Google Scholar]
- Zang, F.; Gerasopoulos, K.; Fan, X.Z.; Brown, A.D.; Culver, J.N.; Ghodssi, R. Real-time monitoring of macromolecular biosensing probe self-assembly and on-chip ELISA using impedimetric microsensors. Biosensors Bioelectron. 2016, 81, 401–407. [Google Scholar]
- Zang, F.; Gerasopoulos, K.; Brown, A.D.; Culver, J.N.; Ghodssi, R. Capillary microfluidics-assembled virus-like particle bionanoreceptor interfaces for label-free biosensing. ACS Appl. Mater. Interfaces 2017, 9, 8471–8479. [Google Scholar] [CrossRef]
- Grübel, J.; Wendlandt, T.; Urban, D.; Jauch, C.O.; Wege, C.; Tovar, G.E.M.; Southan, A. Soft sub-structured multi-material biosensor hydrogels with enzymes retained by plant viral scaffolds. Macromol Biosci 2023, e2300311. [Google Scholar] [CrossRef]
- Wege, C.; Koch, C. From stars to stripes: RNA-directed shaping of plant viral protein templates—structural synthetic virology for smart biohybrid nanostructures. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2020, 12, e1591. [Google Scholar]
- Paiva, T.O.; Schneider, A.; Bataille, L.; Chovin, A.; Anne, A.; Michon, T.; Wege, C.; Demaille, C. Enzymatic activity of individual bioelectrocatalytic viral nanoparticles: dependence of catalysis on the viral scaffold and its length. Nanoscale 2022, 14, 875–889. [Google Scholar]
- Yi, H.; Rubloff, G.W.; Culver, J.N. TMV microarrays: Hybridization-based assembly of DNA-programmed viral nanotemplates. Langmuir 2007, 23, 2663–2667. [Google Scholar] [CrossRef]
- Bruckman, M.A.; Steinmetz, N.F. Chemical Modification of the Inner and Outer Surfaces of Tobacco Mosaic Virus (TMV). In Virus Hybrids as Nanomaterials: Methods and Protocols, Lin, B., Ratna, B., Eds.; Humana Press: Totowa, NJ, 2014; pp. 173–185. [Google Scholar]
- Niu, Z.; Bruckman, M.A.; Li, S.; Lee, L.A.; Lee, B.; Pingali, S.V.; Thiyagarajan, P.; Wang, Q. Assembly of tobacco mosaic virus into fibrous and macroscopic bundled arrays mediated by surface aniline polymerization. Langmuir 2007, 23, 6719–6724. [Google Scholar] [CrossRef]
- Mukerrem, D.; Michael, H.B.S. A chemoselective biomolecular template for assembling diverse nanotubular materials. Nanotechnology 2002, 13, 541. [Google Scholar] [CrossRef]
- Dickmeis, C.; Altintoprak, K.; van Rijn, P.; Wege, C.; Commandeur, U. Bioinspired silica mineralization on viral templates. In Virus-Derived Nanoparticles for Advanced Technologies, Wege, C., Lomonossoff, G.P., Eds.Walker, J.M., Ed.; Methods in Molecular Biology; Humana Press, Springer Science+Business Media: Heidelberg, London, New York, 2018; Volume 1776, pp. 337–362. [Google Scholar]
- Sacco, A.; Barzan, G.; Matić, S.; Giovannozzi, A.M.; Rossi, A.M.; D’Errico, C.; Vallino, M.; Ciuffo, M.; Noris, E.; Portesi, C. Raman-dielectrophoresis goes viral: towards a rapid and label-free platform for plant virus characterization. Front. Microbiol. 2023, 14. [Google Scholar] [CrossRef]
- Saunders, K.; Thuenemann, E.C.; Shah, S.N.; Peyret, H.; Kristianingsih, R.; Lopez, S.G.; Richardson, J.; Lomonossoff, G.P. The use of a replicating virus vector for in planta generation of tobacco mosaic virus nanorods suitable for metallization. Frontiers in Bioengineering and Biotechnology 2022, 10, 877361. [Google Scholar] [CrossRef]
- Negrouk, V.; Eisner, G.; Midha, S.; Lee, H.-i.; Bascomb, N.; Gleba, Y. Affinity purification of streptavidin using tobacco mosaic virus particles as purification tags. Anal. Biochem. 2004, 333, 230–235. [Google Scholar]
- Dobrov, E.N.; Nikitin, N.A.; Trifonova, E.A.; Parshina, E.Y.; Makarov, V.V.; Maksimov, G.V.; Karpova, O.V.; Atabekov, J.G. β-structure of the coat protein subunits in spherical particles generated by tobacco mosaic virus thermal denaturation. J. Biomol. Struct. Dyn. 2014, 32, 701–708. [Google Scholar] [CrossRef]
- Atabekov, J.; Nikitin, N.; Arkhipenko, M.; Chirkov, S.; Karpova, O. Thermal transition of native tobacco mosaic virus and RNA-free viral proteins into spherical nanoparticles. J. Gen. Virol. 2011, 92, 453–456. [Google Scholar]
- Ambrico, M.; Ambrico, P.F.; Minafra, A.; De Stradis, A.; Vona, D.; Cicco, S.R.; Palumbo, F.; Favia, P.; Ligonzo, T. Highly sensitive and practical detection of plant viruses via electrical impedance of droplets on textured silicon-based devices. Sensors 2016, 16, 1946. [Google Scholar]
- Sachse, C.; Chen, J.Z.; Coureux, P.D.; Stroupe, M.E.; Fandrich, M.; Grigorieff, N. High-resolution electron microscopy of helical specimens: a fresh look at tobacco mosaic virus. J. Mol. Biol. 2007, 371, 812–835. [Google Scholar]
- Gregory, J.; Holmes, K.C. Methods of preparing orientated tobacco mosaic virus sols for X-ray diffraction. J. Mol. Biol. 1965, 13, 796–IN715. [Google Scholar] [CrossRef]
- Weis, F.; Beckers, M.; von der Hocht, I.; Sachse, C. Elucidation of the viral disassembly switch of tobacco mosaic virus. EMBO reports 2019, 20, e48451. [Google Scholar]
- Chapman, S.N. Tobamovirus Isolation and RNA Extraction. In Plant Virology Protocols: From Virus Isolation to Transgenic Resistance, Foster, G.D., Taylor, S.C., Eds.; Humana Press: Totowa, NJ, 1998; pp. 123–129. [Google Scholar]
- Martelli, G.P.; Russo, M. Plant virus inclusion bodies. In Adv. Virus Res., Lauffer, M.A., Bang, F.B., Maramorosch, K., Smith, K.M., Eds.; Academic Press: 1977; Volume 21, pp. 175–266.
- Dierking, I.; Al-Zangana, S. Lyotropic Liquid Crystal Phases from Anisotropic Nanomaterials. Nanomaterials 2017, 7, 305. [Google Scholar] [CrossRef]
- Bawden, F.C.; Sheffield, F.M.L. The intracellular inclusions of some plant virus diseases. Ann. Appl. Biol. 1939, 26, 102–115. [Google Scholar] [CrossRef]
- Wehrmeyer, W. Darstellung und Strukturordnung eines Tabakmosalkvirus-Einschlußkörpers in der Zelle. Naturwissenschaften 1957, 44, 519–520. [Google Scholar]
- Adams, M.J.; Adkins, S.; Bragard, C.; Gilmer, D.; Li, D.; MacFarlane, S.A.; Wong, S.-M.; Melcher, U.; Ratti, C.; Ryu, K.H.; Consortium, I.R. ICTV virus taxonomy profile: Virgaviridae. J. Gen. Virol. 2017, 98, 1999–2000. [Google Scholar] [CrossRef]
- Gibbs, A.J. Tobamovirus group CMI/AAB descriptions of plant viruses. Association of Applied Biologicts 1977, 184. [Google Scholar]
- Van Regenmortel, M.H.; Fraenkel-Conrat, H. The Plant Viruses: The rod-shaped plant viruses; Van Regenmortel, M.H., Fraenkel-Conrat, H., Eds.; New York Plenum Press: 1986.
- Melcher, U.; Lewandowski, D.J.; Dawson, W.O. Tobamoviruses (Virgaviridae). In Encyclopedia of Virology (Fourth Edition), Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Oxford, 2021; pp. 734–742. [Google Scholar]
- Higgins, T.J.; Goodwin, P.B.; Whitfeld, P.R. Occurrence of short particles in beans infected with the cowpea strain of TMV. II. Evidence that short particles contain the cistron for coat-protein. Virology 1976, 71, 486–497. [Google Scholar] [CrossRef]
- Beachy, R.N.; Zaitlin, M. Characterization and in vitro translation of the RNAs from less-than-full-length, virus-related, nucleoprotein rods present in tobacco mosaic virus preparations. Virology 1977, 81, 160–169. [Google Scholar] [CrossRef]
- Fukuda, M.; Meshi, T.; Okada, Y.; Otsuki, Y.; Takebe, I. Correlation between particle multiplicity and location on virion RNA of the assembly initiation site for viruses of the tobacco mosaic virus group. Proc. Natl. Acad. Sci. USA 1981, 78, 4231–4235. [Google Scholar] [CrossRef]
- Kim, S.-M.; Lee, J.-M.; Yim, K.-O.; Oh, M.-H.; Park, J.-W.; Kim, K.-H. Nucleotide sequences of two korean isolates of cucumber green mottle mosaic virus. Molecules and Cells 2003, 16, 407–412. [Google Scholar] [CrossRef]
- Finch, J.T. The hand of the helix of tobacco mosaic virus. J. Mol. Biol. 1972, 66, 291–294. [Google Scholar] [CrossRef]
- Namba, K.; Pattanayek, R.; Stubbs, G. Visualization of protein-nucleic acid interactions in a virus. Refined structure of intact tobacco mosaic virus at 2.9 A resolution by X-ray fiber diffraction. J. Mol. Biol. 1989, 208, 307–325. [Google Scholar]
- Ilyas, R.; Rohde, M.J.; Richert-Pöggeler, K.R.; Ziebell, H. To be seen or not to be seen: Latent infection by tobamoviruses. Plants 2022, 11, 2166. [Google Scholar] [CrossRef]
- Stubbs, G. Tobacco mosaic virus particle structure and the initiation of disassembly. Phil. Trans. R. Soc. Lond. B 1999, 354, 551–557. [Google Scholar]
- Culver, J.N. Tobacco mosaic virus assembly and disassembly: Determinants in pathogenicity and resistance. Annu. Rev. Phytopathol. 2002, 40, 287–308. [Google Scholar]
- Wilson, T.M.A.; Perham, R.N.; Finch, J.T.; Butler, P.J.G. Polarity of the RNA in the tobacco mosaic virus particle and the direction of protein stripping in sodium dodecyl sulphate. FEBS Lett. 1976, 64, 285–289. [Google Scholar] [CrossRef]
- Goelet, P.; Lomonossoff, G.P.; Butler, P.J.; Akam, M.E.; Gait, M.J.; Karn, J. Nucleotide sequence of tobacco mosaic virus RNA. Proceedings of the National Academy of Sciences of the United States of America 1982, 79, 5818–5822. [Google Scholar]
- Wetter, C.; Conti, M.; Altschuh, D.; Tabillion, R.; Van Regenmortel, M. Pepper mild mottle virus, a tobamovirus infecting pepper cultivars in Sicily. Phytopathology 1984, 74, 405–410. [Google Scholar] [CrossRef]
- Silber, G.; Burk, L.G. Infectivity of tobacco mosaic virus stored for fifty years in extracted, ‘unpreserved’ plant juice. Nature 1965, 206, 740–741. [Google Scholar] [CrossRef]
- Fraile, A.; García-Arenal, F. Chapter four - Tobamoviruses as models for the study of virus evolution. In Adv. Virus Res., Palukaitis, P., Roossinck, M.J., Eds.; Academic Press: 2018; Volume 102, pp. 89–117.
- Lartey, R.T.; Voss, T.C.; Melcher, U. Tobamovirus evolution: gene overlaps, recombination, and taxonomic implications. Mol. Biol. Evol. 1996, 13, 1327–1338. [Google Scholar] [CrossRef]
- Min, B.E.; Chung, B.N.; Kim, M.J.; Ha, J.H.; Lee, B.Y.; Ryu, K.H. Cactus mild mottle virus is a new cactus-infecting tobamovirus. Arch. Virol. 2006, 151, 13–21. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Smith, E. Seed transmission of Tobamoviruses: Aspects of global disease distribution; 2017; Volume 12, pp. 233–260.
- Dombrovsky, A.; Tran-Nguyen, L.T.T.; Jones, R.A.C. Cucumber green mottle mosaic virus: Rapidly increasing global distribution, etiology, epidemiology, and management. Annu. Rev. Phytopathol. 2017, 55, 231–256. [Google Scholar] [CrossRef]
- Ishibashi, K.; Kubota, K.; Kano, A.; Ishikawa, M. Tobamoviruses: old and new threats to tomato cultivation. J. Gen. Plant Pathol. 2023, 89, 305–321. [Google Scholar] [CrossRef]
- 231. Anses. Expert Committee on "Biological risks for plant health" ToBRFV Working Group Request No. 2019-SA-0080 ToBRFV Express pest risk analysis of tomato brown rugose fruit virus for France. Opinion Collective Expert Appraisal Report.
- Zhang, S.; Griffiths, J.S.; Marchand, G.; Bernards, M.A.; Wang, A. Tomato brown rugose fruit virus: An emerging and rapidly spreading plant RNA virus that threatens tomato production worldwide. Mol. Plant Pathol. 2022, 23, 1262–1277. [Google Scholar] [CrossRef]
- Salem, N.M.; Jewehan, A.; Aranda, M.A.; Fox, A. Tomato brown rugose fruit virus pandemic. Annu. Rev. Phytopathol. 2023, 61, 137–164. [Google Scholar] [CrossRef]
- Matthews, R.E. Effects of some purine analogues on tobacco mosaic virus. Journal of General Microbiology 1954, 10, 521–532. [Google Scholar] [CrossRef]
- Commoner, B.; Shearer, G.B.; Yamada, M. Linear biosynthesis of tobacco mosaic virus: changes in rod length during the course of infection. Proc. Natl. Acad. Sci. USA 1962, 48, 1788–1795. [Google Scholar] [CrossRef]
- Kaper, J.M.; Siberg, R.A. The effect of freezing on the structure of turnip yellow mosaic virus and a number of other simple plant viruses: An ultracentrifugal analysis. Cryobiology 1969, 5, 366–374. [Google Scholar] [CrossRef]
- Mundry, K.-W. Die Abhängigkeit des Auftretens neuer Virusstämme von der Kulturtemperatur der Wirtspflanzen. Zeitschrift für Induktive Abstammungs- und Vererbungslehre 1957, 88, 407–426. [Google Scholar] [CrossRef]
- Wetter, C. Die Flüssigkristalle des Tabakmosaikvirus. Biologie in unserer Zeit 1985, 15, 81–89. [Google Scholar] [CrossRef]
- Zaitlin, M. Tobacco mosaic virus CMI/AAB descriptions of plant viruses. Association of Applied Biologicts 2000, 370. [Google Scholar]
- Bawden, F.; Pirie, N. The separation and properties of tobacco mosaic virus in different states of aggregation. B.r J. Exp. Pathol. 1945, 26, 294. [Google Scholar]
- McNulty, M.J.; Schwartz, A.; Delzio, J.; Karuppanan, K.; Jacobson, A.; Hart, O.; Dandekar, A.; Giritch, A.; Nandi, S.; Gleba, Y.; McDonald, K.A. Affinity sedimentation and magnetic separation with plant-made immunosorbent nanoparticles for therapeutic protein purification. Frontiers in Bioengineering and Biotechnology 2022, 10, 865481. [Google Scholar] [CrossRef]
- Devash, Y.; Hauschner, A.; Sela, I.; Chakraburtty, K. The antiviral factor (AVF) from virus-infected plants induces discharge of histidinyl-TMV-RNA. Virology 1981, 111, 103–112. [Google Scholar] [CrossRef]
- Zaitlin, M.; Israel, H.W. Tobacco mosaic virus (type strain). AAB Descriptions of Plant Viruses.
- Stanley, W.M. Chemical studies on the virus of tobacco mosaic. VI. The isolation from diseased Turkish tobacco plants of a crystalline protein possessing the properties of tobacco mosaic virus. Phytopathology 1936, 26, 305–320. [Google Scholar]
- Leberman, R. The isolation of plant viruses by means of “simple” coacervates. Virology 1966, 30, 341–347. [Google Scholar]
- Wyckoff, R.W.G.; Biscoe, J.; Stanley, W.M. An ultracentrifugal analysis of the crystalline virus proteins isolated from plants diseased with different strains of tobacco mosaic virus. J. Biol. Chem. 1937, 117, 57–71. [Google Scholar] [CrossRef]
- Mächtle, W.; Börger, L. Analytical ultracentrifugation of polymers and nanoparticles; Springer laboratory: Springer, Berlin, New York, 2006. [Google Scholar]
- Wyckoff, R.W.G.; Corey, R.B. The ultracentrifugal crystallization of tobacco mosaic virus protein. Science 1936, 84, 513–513. [Google Scholar] [CrossRef]
- Stanley, W.M. Chemical studies on the virus of tobacco mosaic: X. The activity and yield of virus protein from plants diseased for different periods of time. J. Biol. Chem. 1937, 121, 205–217. [Google Scholar] [CrossRef]
- Stanley, W.M. Chemical studies on the virus of tobacco mosaic: IX. Correlation of virus activity and protein on centrifugation of protein from solution under various conditions. J. Biol. Chem. 1937, 117, 755–770. [Google Scholar] [CrossRef]
- Stanley, W.M.; Wyckoff, R.W.G. The isolation of tobacco ring spot and other virus proteins by ultracentrifugation. Science 1937, 85, 181–183. [Google Scholar] [CrossRef]
- Wyckoff, R.W.G. An ultracentrifugal study of the pH stability of tobacco mosaic virus protein. J. Biol. Chem. 1937, 122, 239–247. [Google Scholar] [CrossRef]
- Lauffer, M.A. Ultracentrifugation studies on tobacco mosaic and bushy stunt viruses. The Journal of Physical Chemistry 1940, 44, 1137–1146. [Google Scholar] [CrossRef]
- Schachman, H.K.; Kauzmann, W.J. Viscosity and sedimentation studies on tobacco mosaic virus. The Journal of Physical and Colloid Chemistry 1949, 53, 150–161. [Google Scholar] [CrossRef]
- Williams, R.C.; Steere, R.L. Electron microscopic observations on the unit of length of the particles of tobacco mosaic virus1. J. Am. Chem. Soc. 1951, 73, 2057–2061. [Google Scholar] [CrossRef]
- Francki, R.; McLean, G. Purification of potato virus X and preparation of infectious ribonucleic acid by degradation with lithium chloride. Australian J. biol. Sci. 1968, 21, 1311–1318. [Google Scholar] [CrossRef]
- Bruening, G.; Beachy, R.N.; Scalla, R.; Zaitlin, M. In vitro and in vivo translation of the ribonucleic acids of a cowpea strain of tobacco mosaic virus. Virology 1976, 71, 498–517. [Google Scholar] [CrossRef]
- Gardner, R.C.; Shepherd, R.J. A procedure for rapid isolation and analysis of cauliflower mosaic virus DNA. Virology 1980, 106, 159–161. [Google Scholar] [CrossRef]
- Jensen, S.M.; Nguyen, C.T.; Jewett, J.C. A gradient-free method for the purification of infective dengue virus for protein-level investigations. J. Virol. Methods 2016, 235, 125–130. [Google Scholar] [CrossRef]
- Stanley, W.M. The concentration and purification of tobacco mosaic virus by means of the sharples super-centrifuge. J. Am. Chem. Soc. 1942, 64, 1804–1806. [Google Scholar] [CrossRef]
- McAleer, W.J.; Hurni, W.; Wasmuth, E.; Hilleman, M.R. High-resolution flow-zonal centrifuge system. Biotechnol. Bioeng. 1979, 21, 317–321. [Google Scholar] [CrossRef]
- Brakke, M.K. Density gradient centrifugation and its application to plant viruses. In Adv. Virus Res.; Advances in Virus Research; Elsevier: 1961; Volume 7, pp. 193–224.
- Brakke, M.K. Density gradient centrifugation: a new separation technique1. J. Am. Chem. Soc. 1951, 73, 1847–1848. [Google Scholar] [CrossRef]
- Hills, G.J. Partial crystallization of tobacco mosaic virus. Virology 1959, 7, 239–240. [Google Scholar] [CrossRef]
- Brakke, M.K. Nonideal sedimentation and the capacity of sucrose gradient columns for virus in density-gradient centrifugation. Archives of Biochemistry and Biophysics 1964, 107, 388–403. [Google Scholar] [CrossRef]
- Brakke, M.K.; Van Pelt, N. Linear-log sucrose gradients for estimating sedimentation coefficients of plant viruses and nucleic acids. Anal. Biochem. 1970, 38, 56–64. [Google Scholar] [CrossRef]
- Saunders, K.; Thuenemann, E.C.; Peyret, H.; Lomonossoff, G.P. The tobacco mosaic virus origin of assembly sequence is dispensable for specific viral rna encapsidation but necessary for initiating assembly at a single site. J. Mol. Biol. 2022, 434, 167873. [Google Scholar] [CrossRef]
- Liu, X.; Wu, F.; Tian, Y.; Wu, M.; Zhou, Q.; Jiang, S.; Niu, Z. Size Dependent Cellular Uptake of Rod-like Bionanoparticles with Different Aspect Ratios. Sci. Rep. 2016, 6, 24567. [Google Scholar] [CrossRef]
- Perham, R.N. Sucrose density-gradient analysis of the alkaline degradation of tobacco mosaic virus. J. Mol. Biol. 1969, 45, 439–441. [Google Scholar] [CrossRef]
- Siegel, A.; Hudson, W. Equilibrium centrifugation of two strains of tobacco mosaic virus in density gradients. Biochim. Biophys. Acta 1959, 34, 254–255. [Google Scholar] [CrossRef]
- Gugerli, P. Isopycnic centrifugation of plant viruses in Nycodenz® density gradients. J. Virol. Methods 1984, 9, 249–258. [Google Scholar] [CrossRef]
- Skotnicki, A.; Scotti, P.D.; Gibbs, A. On the nature of the difference in the densities of the particles of two tobamoviruses. Intervirology 1976, 7, 292–302. [Google Scholar] [CrossRef]
- Rickwood, D.; Ford, T.; Graham, J. Nycodenz: A new nonionic iodinated gradient medium. Anal. Biochem. 1982, 123, 23–31. [Google Scholar] [CrossRef]
- Rickwood, D.; Birnie, G. Metrizamide, a new density-gradient medium. FEBS Lett. 1975, 50, 102–110. [Google Scholar]
- Pertoft, H.; Philipson, L.; Oxelfelt, P.; Höglund, S. Gradient centrifugation of viruses in colloidal silica. Virology 1967, 33, 185–196. [Google Scholar] [CrossRef]
- Pertoft, H. Fractionation of cells and subcellular particles with Percoll. J. Biochem. Biophys. Methods 2000, 44, 1–30. [Google Scholar] [CrossRef]
- Prucha, M.J.; Tanner, F.W. TIME SAVING BACTERIOLOGICAL APPARATUS. J. Bacteriol. 1920, 5, 559–563. [Google Scholar] [CrossRef]
- Arrua, R.D.; Strumia, M.C.; Alvarez Igarzabal, C.I. Macroporous Monolithic Polymers: Preparation and Applications. Materials 2009, 2, 2429–2466. [Google Scholar]
- Turpeinen, D.G.; Joshi, P.U.; Kriz, S.A.; Kaur, S.; Nold, N.M.; O'Hagan, D.; Nikam, S.; Masoud, H.; Heldt, C.L. Continuous purification of an enveloped and non-enveloped viral particle using an aqueous two-phase system. Separation and Purification Technology 2021, 269, 118753. [Google Scholar] [CrossRef]
- McLean, G.D.; Francki, R.I.B. Purification of lettuce necrotic yellows virus by column chromatography on calcium phosphate gel. Virology 1967, 31, 585–591. [Google Scholar] [CrossRef]
- Mahy, B.W.J. Virology: A practical approach, Reprint. ed, ed.; IRL Press: Oxford, 1985. [Google Scholar]
- Francki, R.; Zaitlin, M.; Grivell, C. An unusual strain of tobacco mosaic virus from Plumeria acutifolia. Australian J. biol. Sci. 1971, 24, 811–814. [Google Scholar] [CrossRef]
- Cochran, G. A chromatographic method for the detection of tobacco-mosaic virus in juice from diseased Turkish tobacco plants. Phytopathology 1947, 37, 850–851. [Google Scholar]
- Gray, R.A. The electrophoresis and chromatography of plant viruses on filter paper. Arch. Biochem. Biophys. 1952, 38, 305–316. [Google Scholar] [CrossRef]
- Barton, R.J. An examination of permeation chromatography on columns of controlled pore glass for routine purification of plant viruses. J. Gen. Virol. 1977, 35, 77–87. [Google Scholar] [CrossRef]
- Čech, M.; Jelinkova, M.; Čoupek, J. High-pressure chromatography of tobacco mosaic virus on Spheron gels. J. Chromatogr. 1977, 135, 435–440. [Google Scholar] [CrossRef]
- Brunt, A.; Phillips, S.; Jones, R.; Kenten, R. Viruses detected in Ullucus tuberosus (Basellaceae) from Peru and Bolivia. Ann. Appl. Biol. 1982, 101, 65–71. [Google Scholar] [CrossRef]
- Orita, H.; Sakai, J.-I.; Kubota, K.; Okuda, M.; Tanaka, Y.; Hanada, K.; Imamura, Y.; Nishiguchi, M.; Karasev, A.V.; Miyata, S.-I. Molecular and serological characterization of cucumber mottle virus, a new cucurbit-infecting tobamo-like virus. Plant Dis. 2007, 91, 1574–1578. [Google Scholar] [CrossRef]
- Cochran, G.W.; Chidester, J.L.; Stocks, D.L. Chromatography of tobacco mosaic virus on a cellulose cation exchange adsorbent. Nature 1957, 180, 1281–1282. [Google Scholar] [CrossRef]
- Commoner, B.; Lippincott, J.A.; Shearer, G.B.; Richman, E.E.; Wu, J.-H. Reconstitution of tobacco mosaic virus components. Nature 1956, 178, 767–771. [Google Scholar] [CrossRef]
- Levin, Ö. Chromatography of tobacco mosaic virus and potato virus X. Arch. Biochem. Biophys. 1958, 78, 33–45. [Google Scholar] [CrossRef]
- Ruščić, J.; Gutiérrez-Aguirre, I.; Tušek Žnidarič, M.; Kolundžija, S.; Slana, A.; Barut, M.; Ravnikar, M.; Krajačić, M. A new application of monolithic supports: The separation of viruses from one another. J. Chromatogr. 2015, 1388, 69–78. [Google Scholar] [CrossRef]
- Krajacic, M.; Ravnikar, M.; Štrancar, A.; Gutiérrez-Aguirre, I. Application of monolithic chromatographic supports in virus research. Electrophoresis 2017, 38, 2827–2836. [Google Scholar] [CrossRef]
- Kramberger, P.; Petrovič, N.; Štrancar, A.; Ravnikar, M. Concentration of plant viruses using monolithic chromatographic supports. J. Virol. Methods 2004, 120, 51–57. [Google Scholar] [CrossRef]
- Kramberger, P.; Peterka, M.; Boben, J.; Ravnikar, M.; Štrancar, A. Short monolithic columns-A breakthrough in purification and fast quantification of tomato mosaic virus. J. Chromatogr. 2007, 1144, 143–149. [Google Scholar] [CrossRef]
- Fraser, D.; Johnson, F. The influence of buffer composition, pH and aggregation on the thermal denaturation of tobacco mosaic virus. Arch. Biochem. 1949, 24, 338–349. [Google Scholar]
- Desjardins, P.R.; French, J.V. Storage of purified plant viruses in the unfrozen state free of microbial contamination. Experientia 1969, 25, 444–446. [Google Scholar] [CrossRef]
- Best, R.J. Longevity of tobacco mosaic virus: Part I. in vitro life of the pure virus in buffer solution at pH 4. Aust. J. Exp. Bio.l Med. Sci. 1948, 26, 163–170. [Google Scholar] [CrossRef]
- Byrne, N.; Rodoni, B.; Constable, F.; Varghese, S.; Davis, J.H. Enhanced stabilization of the tobacco mosaic virus using protic ionic liquids. Physical Chemistry Chemical Physics 2012, 14, 10119–10121. [Google Scholar] [CrossRef]
- Rice, R.V.; Kaesberg, P.; Stahmann, M.A. Further studies concerning the breaking of tobacco mosaic virus. Biochim. Biophys. Acta 1956, 20, 488–496. [Google Scholar] [CrossRef]
- Donev, T.; Yordanova, A.; Stoimenova, E.; Damjanova, S. Determination of parameters for tobacco mosaic virus cryogenic treatment and freeze-drying. Biotechnol. Tech. 1996, 10, 971–976. [Google Scholar] [CrossRef]
- Wacker, W.E.; Gordon, M.P.; Huff, J.W. Metal content of tobacco mosaic virus and tobacco mosaic virus RNA. Biochemistry 1963, 2, 716–719. [Google Scholar] [CrossRef]
- Hulett, H.R.; Loring, H.S. Effect of particle length distribution on infectivity of tobacco mosaic virus. Virology 1965, 25, 418–430. [Google Scholar] [CrossRef]
- Loring, H.S.; Fujimoto, Y.; Tu, A.T. Tobacco mosaic virus—A calcium-magnesium coordination complex. Virology 1962, 16, 30–40. [Google Scholar] [CrossRef]
- Ladipo, J.L.; Koenig, R.; Lesemann, D.E. Nigerian tobacco latent virus: a new tobamovirus from tobacco in Nigeria. Eur. J. Plant Pathol. 2003, 109, 373–379. [Google Scholar] [CrossRef]
- Whitfeld, P.R.; Higgins, T.J. Occurrence of short particles in beans infected with the cowpea strain of TMV. I. Purification and characterization of short particles. Virology 1976, 71, 471–485. [Google Scholar] [CrossRef]
- Eskelin, K.; Poranen, M.M.; Oksanen, H.M. Asymmetrical flow field-flow fractionation on virus and virus-like particle applications. Microorganisms 2019, 7, 555. [Google Scholar] [CrossRef]
- Lampi, M.; Oksanen, H.M.; Meier, F.; Moldenhauer, E.; Poranen, M.M.; Bamford, D.H.; Eskelin, K. Asymmetrical flow field-flow fractionation in purification of an enveloped bacteriophage ϕ6. Journal of Chromatography B 2018, 1095, 251–257. [Google Scholar] [CrossRef]
- Albertsson, P.A.; Frick, G. Partition of virus particles in a liquid two-phase system. Biochimica et Biophysica Acta 1960, 37, 230–237. [Google Scholar] [CrossRef]
- Teepakorn, C.; Fiaty, K.; Charcosset, C. Comparison of Membrane Chromatography and Monolith Chromatography for Lactoferrin and Bovine Serum Albumin Separation. Processes 2016, 4, 31. [Google Scholar]
- Yang, Z.; Xu, X.; Silva, C.A.T.; Farnos, O.; Venereo-Sanchez, A.; Toussaint, C.; Dash, S.; González-Domínguez, I.; Bernier, A.; Henry, O.; Kamen, A. Membrane chromatography-based downstream processing for cell-culture produced influenza vaccines. Vaccines 2022, 10, 1310. [Google Scholar] [CrossRef]
- Chen, J.; Yu, B.; Cong, H.; Shen, Y. Recent development and application of membrane chromatography. Anal. Bioanal. Chem. 2023, 415, 45–65. [Google Scholar] [CrossRef]
- 313. Sartorius. Resins, monoliths, or membranes. Which chromatographic method should you use? Sartorius AG Science-Snippets.
- Thuenemann, E.C.; Le, D.H.T.; Lomonossoff, G.P.; Steinmetz, N.F. Bluetongue virus particles as nanoreactors for enzyme delivery and cancer therapy. Mol. Pharmacol. 2021, 18, 1150–1156. [Google Scholar] [CrossRef]
- Thuenemann, E.C.; Meyers, A.E.; Verwey, J.; Rybicki, E.P.; Lomonossoff, G.P. A method for rapid production of heteromultimeric protein complexes in plants: assembly of protective bluetongue virus-like particles. Plant Biotechnology Journal 2013, 11, 839–846. [Google Scholar] [CrossRef]
- Dennis, S.J.; Meyers, A.E.; Guthrie, A.J.; Hitzeroth, II; Rybicki, E. P. Immunogenicity of plant-produced African horse sickness virus-like particles: implications for a novel vaccine. Plant Biotechnol. J. 2018, 16, 442–450. [Google Scholar] [CrossRef]
- Ford, T.; Graham, J.; Rickwood, D. Iodixanol: a nonionic iso-osmotic centrifugation medium for the formation of self-generated gradients. Analytical Biochemistry 1994, 220, 360–366. [Google Scholar] [CrossRef]
- Rybicki, E.P. Plant molecular farming of virus-like nanoparticles as vaccines and reagents. WIREs Nanomedicine and Nanobiotechnology 2020, 12, e1587. [Google Scholar] [CrossRef]
- Marsian, J.; Lomonossoff, G.P. Molecular pharming — VLPs made in plants. Curr. Opin. Biotechnol. 2016, 37, 201–206. [Google Scholar] [CrossRef]
- Zahmanova, G.; Aljabali, A.A.; Takova, K.; Toneva, V.; Tambuwala, M.M.; Andonov, A.P.; Lukov, G.L.; Minkov, I. The Plant Viruses and Molecular Farming: How Beneficial They Might Be for Human and Animal Health? International Journal of Molecular Sciences 2023, 24, 1533. [Google Scholar]
- Margolin, E.; Allen, J.D.; Verbeek, M.; Van Diepen, M.; Ximba, P.; Chapman, R.; Meyers, A.; Williamson, A.-L.; Crispin, M.; Rybicki, E. Site-specific glycosylation of recombinant viral glycoproteins produced in Nicotiana benthamiana. Front. Plant Sci. 2021, 12, 709344. [Google Scholar]
Figure 1.
Arrangement of TMV virions into highly ordered aggregates (syn. (para)crystals, liquid crystals) in infected plant cells and after purification in vitro. (A) Title page of popular scientific booklet on 'Viruses and Miasmata', published 1949 in Stuttgart, Germany: contemporary drawing of N. tabacum leaf showing symptoms of the tobacco mosaic disease. Copyright 2024 by Franckh-Kosmos Verlags-GmbH & Co. KG on behalf of Franckh'sche Verlagshandlung, W. Keller & Co., Stutgart. (B) and (C) Aggregates of TMV virions arranged in layered rows in infected plant cells can be visualized by both fluorescence and electron microscopy. (B) TMV is detected by immunofluorescence (yellow) on sections prepared for electron microscopy (scale bar 10 µm, cell nuclei are stained with DAPI, blue). (C) Transmission electron microscopy of ultrathin sections showing the regular arrays of TMV particles in leaf cells (scale bar 500 nm). (B) and (C) are kindly provided by Heinz Schwarz, MPI for Biology Tübingen, Germany. (D) Overview of a classical protocol applied by Stanley (1936) for TMV virion isolation via series of salt and acid precipitation. Reproduced from Steere, 1959 [23] with minor changes. Copyright 2024 by Copyright Clearance Center, Inc. on behalf of Elsevier Science & Technology Journals. (E) Photomicrograph of crystalline aggregates formed in vitro from purified TMV (strain acuba) particles at 393x magnification. Reproduced from Stanley 1937 [24] with minor changes. This work is made available under the terms of the Creative Commons Attribution-NonCommercial 4.0 Unported License, https://creativecommons.org/licenses/by/4.0/ (F) Iridescent TMV gels with different micro- and macrocrystalline structural phases (nematic and smectic liquid crystals) formed in vitro from purified virions (Kreibig and Wetter 1980). Diffraction of visible light by iridescent gel (upper panel) and by gel of randomly oriented macrocrystals with different diffraction colors (lower panel); scale bar 0.5 mm. Reproduced from Kreibig and Wetter 1980 [25] with slight modifications. This work is made available under the terms of the Creative Commons Attribution-NonCommercial 4.0 Unported License, https://creativecommons.org/licenses/by/4.0/.
Figure 1.
Arrangement of TMV virions into highly ordered aggregates (syn. (para)crystals, liquid crystals) in infected plant cells and after purification in vitro. (A) Title page of popular scientific booklet on 'Viruses and Miasmata', published 1949 in Stuttgart, Germany: contemporary drawing of N. tabacum leaf showing symptoms of the tobacco mosaic disease. Copyright 2024 by Franckh-Kosmos Verlags-GmbH & Co. KG on behalf of Franckh'sche Verlagshandlung, W. Keller & Co., Stutgart. (B) and (C) Aggregates of TMV virions arranged in layered rows in infected plant cells can be visualized by both fluorescence and electron microscopy. (B) TMV is detected by immunofluorescence (yellow) on sections prepared for electron microscopy (scale bar 10 µm, cell nuclei are stained with DAPI, blue). (C) Transmission electron microscopy of ultrathin sections showing the regular arrays of TMV particles in leaf cells (scale bar 500 nm). (B) and (C) are kindly provided by Heinz Schwarz, MPI for Biology Tübingen, Germany. (D) Overview of a classical protocol applied by Stanley (1936) for TMV virion isolation via series of salt and acid precipitation. Reproduced from Steere, 1959 [23] with minor changes. Copyright 2024 by Copyright Clearance Center, Inc. on behalf of Elsevier Science & Technology Journals. (E) Photomicrograph of crystalline aggregates formed in vitro from purified TMV (strain acuba) particles at 393x magnification. Reproduced from Stanley 1937 [24] with minor changes. This work is made available under the terms of the Creative Commons Attribution-NonCommercial 4.0 Unported License, https://creativecommons.org/licenses/by/4.0/ (F) Iridescent TMV gels with different micro- and macrocrystalline structural phases (nematic and smectic liquid crystals) formed in vitro from purified virions (Kreibig and Wetter 1980). Diffraction of visible light by iridescent gel (upper panel) and by gel of randomly oriented macrocrystals with different diffraction colors (lower panel); scale bar 0.5 mm. Reproduced from Kreibig and Wetter 1980 [25] with slight modifications. This work is made available under the terms of the Creative Commons Attribution-NonCommercial 4.0 Unported License, https://creativecommons.org/licenses/by/4.0/.
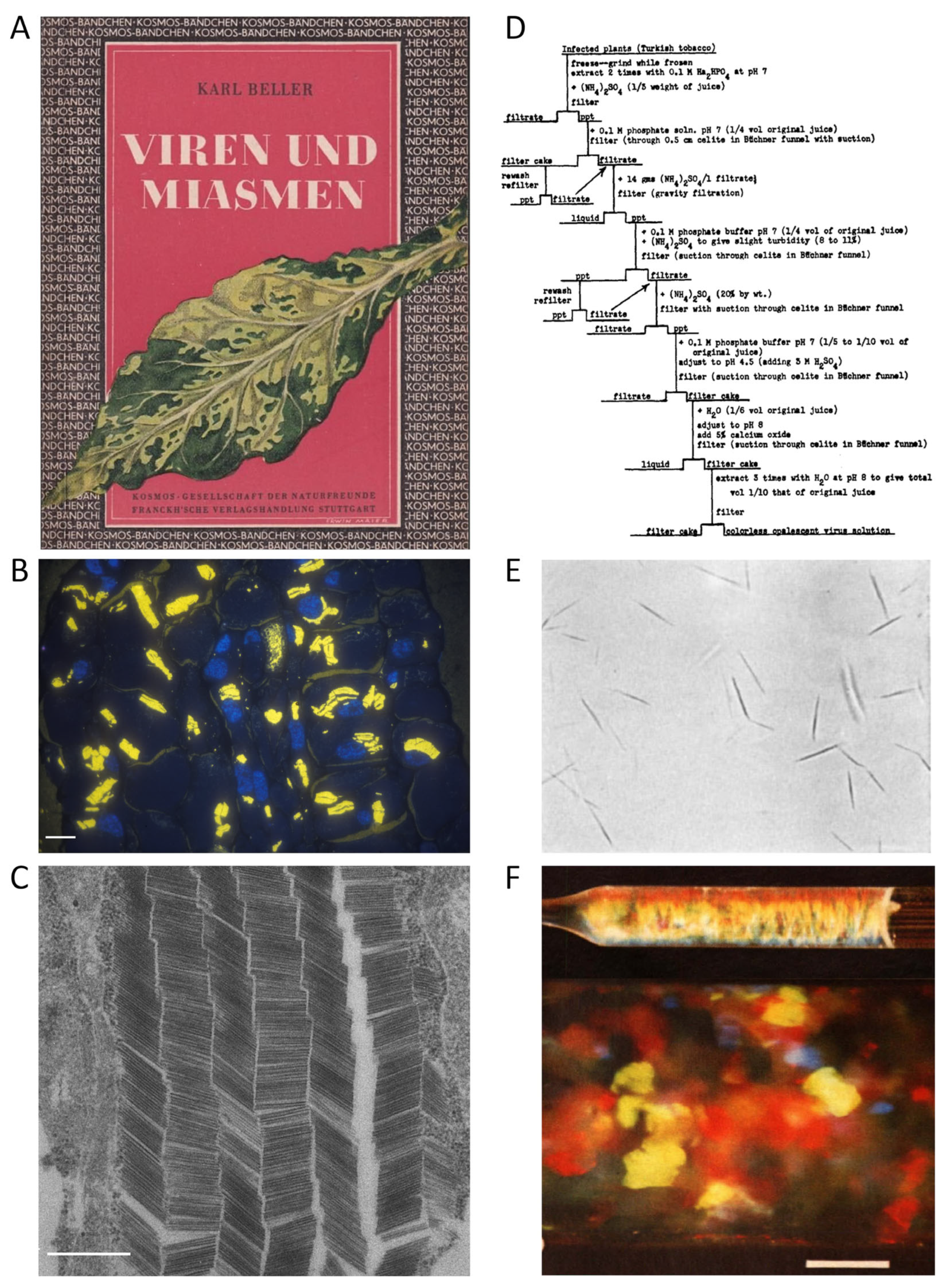
Figure 2.
Laboratory instruments for virus purification. (A) Supercentrifuges were run in a continuous mode to pellet virus or virus precipitates. Rotor (2) within a chassis (3) is connected to a drive (5). Feed liquid (1) enters the rotor and and separates into a heavy and light fraction, which leave the centrifuge through separate outlets (4). (B) Glassware was cleaned for example with electrically driven test tube and flask washing machines. A and B were reproduced from Prucha and Tanner 1920 [277] with minor changes. Copyright 2024 by Copyright Clearance Center, Inc. on behalf of the American Society for Microbiology Journals. (C) Later, viruses were separated by density gradient centrifugation, for example with isopycnic banding in Nycodenz® gradients. Reproduced from Gugerli 1984 [271] with minor changes. Copyright 2024 by Copyright Clearance Center on behalf of Elsevier Science & Technology Journals. (D) For the past 20 years, monolithic materials have been evaluated for virus purification by chromatography. In monolithic materials, viruses can easily access active sites on surfaces by convection, which allows increased flow rates. Reproduced from Arrua 2009 [278] with minor changes. This work is made available under the terms of the Creative Commons Attribution-NonCommercial 3.0 Unported License, http://creativecommons.org/licenses/by/4.0/ (E) For continuous purification of viruses, aqueous two phase systems are currently explored: A virus solution and both phases are combined and mixed in a first part of the instrument (marked with 1 and shown in the lower left inset). The two phases are then separated in a second part of the instrument (marked with 2 and shown in the lower right inset). Adapted from Turpeinen 2021 [279] with minor changes. Copyright 2024 by Copyright Clearance Center on behalf of Elsevier.
Figure 2.
Laboratory instruments for virus purification. (A) Supercentrifuges were run in a continuous mode to pellet virus or virus precipitates. Rotor (2) within a chassis (3) is connected to a drive (5). Feed liquid (1) enters the rotor and and separates into a heavy and light fraction, which leave the centrifuge through separate outlets (4). (B) Glassware was cleaned for example with electrically driven test tube and flask washing machines. A and B were reproduced from Prucha and Tanner 1920 [277] with minor changes. Copyright 2024 by Copyright Clearance Center, Inc. on behalf of the American Society for Microbiology Journals. (C) Later, viruses were separated by density gradient centrifugation, for example with isopycnic banding in Nycodenz® gradients. Reproduced from Gugerli 1984 [271] with minor changes. Copyright 2024 by Copyright Clearance Center on behalf of Elsevier Science & Technology Journals. (D) For the past 20 years, monolithic materials have been evaluated for virus purification by chromatography. In monolithic materials, viruses can easily access active sites on surfaces by convection, which allows increased flow rates. Reproduced from Arrua 2009 [278] with minor changes. This work is made available under the terms of the Creative Commons Attribution-NonCommercial 3.0 Unported License, http://creativecommons.org/licenses/by/4.0/ (E) For continuous purification of viruses, aqueous two phase systems are currently explored: A virus solution and both phases are combined and mixed in a first part of the instrument (marked with 1 and shown in the lower left inset). The two phases are then separated in a second part of the instrument (marked with 2 and shown in the lower right inset). Adapted from Turpeinen 2021 [279] with minor changes. Copyright 2024 by Copyright Clearance Center on behalf of Elsevier.

Figure 3.
Isolation of TVCV and TVCVPA using a novel three-layer density barrier phase system (5/32/42% iodixanol) for virion purification from PEG raw precipitates. (A) Photograph and scheme (middle) depicting a three-layer density barrier phase system loaded with a resuspended PEG precipitate (17.5 ml TVCVPA-containing sample per SW 32 Ti ultracentrifugation tube). One of the two corresponding tubes (tube 1) was photographed before (left) and after (right) centrifugation (4 °C, 15 h, 24,000 rpm [rmax 98,381 × g]). Visible bands formed upon centrifugation in the 32 % iodixanol-containing retention phase (arrows 1 and 2) were sampled by puncturing the tube's wall with a syringe and needle. Band 1 (arrows 1) was partially aspirated (300 µl) for later analysis by SDS-PAGE (sample 1 in subfigure B), and band 2 (arrows 2) taken completely (1.3 ml per twin tube, sample 2 in subfigure B). (B) SDS-PAGE (15% polyacrylamide) of samples taken from leaf raw extract before PEG precipitation, and samples 1 and 2 taken from tube 1 (shown in A) and twin tube 2. 9 µl of samples 1, 9 µl of raw lysate, and 2 µl of samples 2 were applied to the gel. Staining: Coomassie Blue-G-250. (C) and (D) Purified wildtype TVCV (C) and TVCVPA (D). Particles (0.1 mg/ml) were negatively stained with 1% uranyl acetate on glow-discharged carbon-coated copper grids. Grids were examined with a Tecnai G2 Spirit BioTwin (Thermo Fisher Scientific) operated at 120 kV and equipped with a TemCam XF-416 (TVIPS GmbH).
Figure 3.
Isolation of TVCV and TVCVPA using a novel three-layer density barrier phase system (5/32/42% iodixanol) for virion purification from PEG raw precipitates. (A) Photograph and scheme (middle) depicting a three-layer density barrier phase system loaded with a resuspended PEG precipitate (17.5 ml TVCVPA-containing sample per SW 32 Ti ultracentrifugation tube). One of the two corresponding tubes (tube 1) was photographed before (left) and after (right) centrifugation (4 °C, 15 h, 24,000 rpm [rmax 98,381 × g]). Visible bands formed upon centrifugation in the 32 % iodixanol-containing retention phase (arrows 1 and 2) were sampled by puncturing the tube's wall with a syringe and needle. Band 1 (arrows 1) was partially aspirated (300 µl) for later analysis by SDS-PAGE (sample 1 in subfigure B), and band 2 (arrows 2) taken completely (1.3 ml per twin tube, sample 2 in subfigure B). (B) SDS-PAGE (15% polyacrylamide) of samples taken from leaf raw extract before PEG precipitation, and samples 1 and 2 taken from tube 1 (shown in A) and twin tube 2. 9 µl of samples 1, 9 µl of raw lysate, and 2 µl of samples 2 were applied to the gel. Staining: Coomassie Blue-G-250. (C) and (D) Purified wildtype TVCV (C) and TVCVPA (D). Particles (0.1 mg/ml) were negatively stained with 1% uranyl acetate on glow-discharged carbon-coated copper grids. Grids were examined with a Tecnai G2 Spirit BioTwin (Thermo Fisher Scientific) operated at 120 kV and equipped with a TemCam XF-416 (TVIPS GmbH).
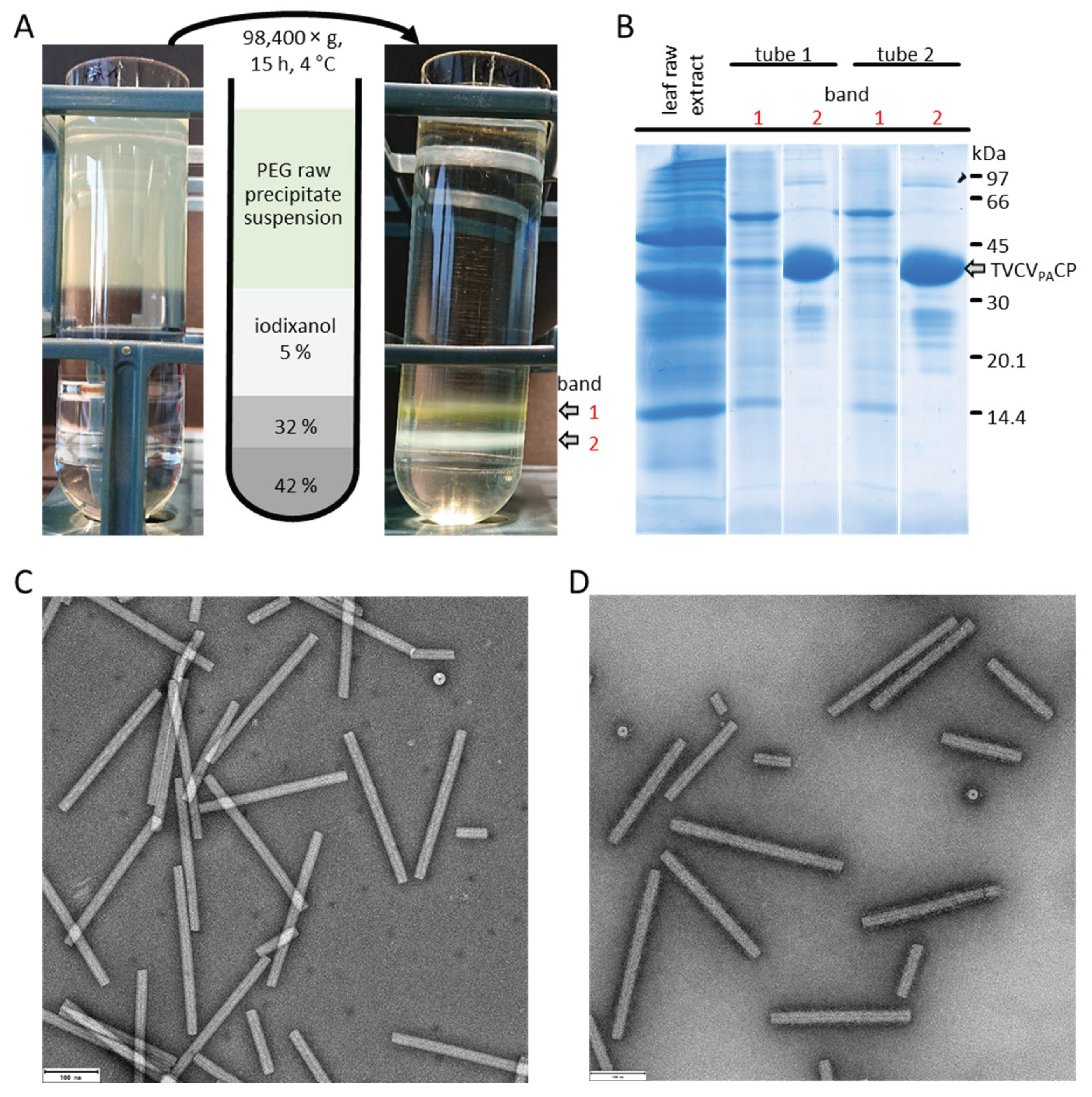

Table 1.
Chromatographic media and conditions for purification TMV with the bind/elute mode. Arrows indicate gradients with starting conditions on the left side and final conditions at the right side of the arrow.
Table 1.
Chromatographic media and conditions for purification TMV with the bind/elute mode. Arrows indicate gradients with starting conditions on the left side and final conditions at the right side of the arrow.
| Type | Stationary phase | Mobile Phase | Comment | Citation |
|---|---|---|---|---|
| Cation exchange | Carboxymethyl-cellulose | 5 mM Citric acid pH 3 50 mM Na2HPO4 /0.1 M NaCl pH 9.05 |
-- | (Cochran et al., 1957) |
| Cation exchange/ Ca2+ affinity | Hydroxyapatite | 0.001 M Phosphate pH 6.8 0.3 M Phosphate pH 6.8 |
Used for separation of partially and completely in vitro reconstituted particles | (Guilley et al., 1972) |
| Anion exchange | Cellulose modified with epichlorhydrin and triethanolamine | 0.01 M Phosphate pH 7 0.01 M Phosphate / 1 M NaCl pH 7 |
Used for separation of in vitro reconstituted particles | (Commoner et al., 1956) |
| Anion exchange | DEAE-cellulose | 0.01 M Tris-HCl pH 7.3 0.8 M Tris-HCl pH 7.3 |
-- | (Levin, 1958) |
| Anion exchange | CIM - QA | 1.20 mM NaOAc pH 5.5 2.20 mM NaOAc / 1.5 M NaCl pH 5.5 |
Yields 70-90 % Tomato mosaic virus only |
(Kramberger et al., 2004) |
| Anion exchange | CIM - QA + CIM - DEAE | Various; best performing with CIM QA: 1. 20 mM NaOAc pH 5.5 2. 20 mM NaOAc / 0.45 M NaCl pH 5.5 3. 20 mM NaOAc / 1.5 M NaCl pH 5.5 |
Highest yield with CIM – QA Up to 98 % yield, on average 75 % Allows purification from clarified extract within one step; tomato mosaic virus only |
(Kramberger et al., 2007) |
| Anion exchange | CIM - QA + CIM - DEAE | 20 mM NaOAc pH 5.5 20 mM NaOAc / 1 M NaCl pH 5.5 |
Separation of different viruses from their mixtures | (Ruščić et al., 2015) |
| various | Chitin | 1. 0.01 M Tris-HCl pH 6.8 2. Water 0.5 M K2HPO4 |
-- | (Townsley, 1961) |
| various | Methylated albumin on kieselgur (MAK) | 0.05 M Phosphate / 0.2 M NaCl 0.05 M Phosphate / 1 M NaCl |
Separation RNA, TMV CP and TMV particles | (Kubo et al., 1965) |
| Size exclusion | Spheron (Hydroxyalkyl methacrylate gel) | 0.05 M Tris-HCl pH 7.5 | Various particle sizes and exclusion limits tested Yield about 90% |
(Čech et al., 1977) |
| Size exclusion | Controlled pore glass | 0.05 M phosphate pH 7 | Recovery close to 100 % Comparison with Sucrose gradient centrifugation; Yield of centrifugation: 80% Comparison of different viruses |
(Barton, 1977) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



