
Submitted:
04 April 2024
Posted:
05 April 2024
You are already at the latest version
Abstract
Tanning, crucial for leather production, relies heavily on chromium yet poses risks due to chromium's oxidative conversion, leading to significant wastewater and solid waste generation. Physico-chemical methods are typically used for heavy metal removal, but they have drawbacks, prompting interest in eco-friendly biological remediation techniques like biosorption, bioaccumulation, and biotransformation. The EU Directive (2018/850) mandates alternatives to landfilling or incineration for industrial textile waste management, highlighting the importance of environmentally conscious practices for leather products' end-of-life management, with com-posting being the most researched and viable option. This study aimed to isolate microorganisms from tannery wastewater and identify those responsible for different types of tanned leather biodegradation. Using a leather biodegradation assay (ISO 20136) with tannery and municipal wastewaters as inoculum, bacterial shifts during leather biodegradation were observed. Over 10,000 bacterial species were identified in all analyzed samples, with 8 bacterial strains isolated from tannery wastewater. Identification of bacterial genera like Acinetobacter, Brevundimo-nas, and Mycolicibacterium provides insights into potential microbial candidates for enhancing leather biodegradability, wastewater treatment, and heavy metal bioremediation in industrial applications.
Keywords:
heavy metal reduction
; leather biodegradation
; microbial strain characterization
; tannery wastewater
1. Introduction
The leather industry is vital in the global economy, providing high-quality fashion, automotive, and upholstery products [1]. Tanning is the chemical process by which collagen fibres are stabilized, preventing their putrefaction and ultimately forming durable leather [2]. For over 160 years, conventional chrome tanning has been employed in leather manufacturing [3], endowing leather with excellent shrinkage temperature (Ts) [4] and mechanical properties like flexibility, durability, and resistance to environmental factors [5]. Currently, 90% of hides are tanned using chrome [6], where chromium ions (Cr3+) crosslink with the carboxyl and amino groups in the collagen fibres [7]. However, chrome tanning agents face significant limitations due to potential hazards, with the most concerning being the oxidative conversion of Cr(III) into hazardous and carcinogenic Cr(VI)[8]. This oxidation process could occur during tanning process due to elevated pH levels, temperature fluctuations, exposure to UV radiation, improper storage conditions, and the use of lubricants containing double bonds in their molecular structure[9]. At the micro-level, Cr(III) is an essential trace element for multiple physiological processes in the human body, such as glucose, fat, and protein metabolism, by enhancing insulin activity [10]. While Cr(III) complexes face challenges in penetrating cell membranes [11], Cr(VI) is highly soluble in water and toxic. It can pass rapidly through cell membranes, accumulating and, eventually, interacting with proteins and nucleic acids, ultimately damaging DNA [12].
The leather goods industry was worth USD 245 billion in 2022 [13] and stands out as one of the most environmentally impactful and resource-intensive sectors. From every 1,000 kg of raw material, 250 kg of leather is produced, leaving a substantial water footprint ranging from 15,000 to 120,000 cubic meters [14]. This process results in the generation of 15 to 50 metric tons of wastewater and 400 to 700 kg of solid waste, greenhouse gases (such as CO2, H2S, NH3), as well as volatile organic compounds like amines, aldehydes, and hydrocarbons [12]. The emission of chemicals is strongly influenced by the treatment type and the technological processes employed in tanneries [15].
Various methods are employed to remove heavy metals from inorganic eluents [16]. Physico-chemical approaches include precipitation through the use of metal hydroxides, sulfides, carbonates, and phosphates [17]. Ion exchange utilizes solid resins for reversible ion exchange [18]. Membrane filtration methods include nanofiltration (NF) for molecules within the 300-500 Da molecular weight range [19] and reverse osmosis (RO) through a pressure-driven separation [20]. Other recent alternative techniques have also been employed, such as coagulation/flocculation, electrocoagulation, electro-floatation, and electro-deposition [21]. However, these methods may have drawbacks, such as incomplete metal removal, sludge generation, high reagent and energy requirements, and membrane fouling [22]. As a result, attention has turned towards biological remediation methods, such as biosorption, bioaccumulation and biotransformation, which offer cost-effective and environmentally friendly solutions for efficiently removing heavy metals like chromium from industrial waste [23]. Microorganisms eliminate heavy metals through enzyme-catalyzed metabolic pathways for toxic substance degradation, transforming them into carbon dioxide, methane, water, and biomass [24]. A wide range of microorganisms, bacteria [25], algae [26], fungi [27] and phyto species [28] have already been identified and isolated for potential heavy metal bioremediation and wastewater treatment.
On the other hand current Directive 1999/31/EC of April 26th 1999 [29] on the landfill of waste, as well as Directive 2008/98/EC [30] on garbage [16], allows the incineration or disposal in landfills of leather waste, chrome shavings, and solid waste. However, starting January 1st, 2025, the new Directive (EU) 2018/850 on landfills of waste [20] will prohibit landfilling or incineration as management methods for industrial textile waste. The European leather industry to increasingly focus on sustainability initiatives to reduce environmental impact and promote responsible sourcing [31]. Through the implementation of leather processing guides to improve and implement sustainable manufacturing practices [32], but most importantly by developmping more biodegradable leather products which could be composted. Leather biodegradability directly depends on the nature of the tanning agents and chemicals used in the manufacturing process, therefore the leather industry has also focused on developping more biodegradable leathers with the aim of giving their end products an sustainable, non-contaminating second life or feasable disposal. These include the use of chrome-free leather manufacturing using non-metal tanning materials such as vegetable tannins (polyphenolic agents known to bind to collagen) [33], aldehydes compounds (phenolic synthetic compounds bind to amine groups of collagens forming ionic bonds), calixarene [34], and other metal-free tanning agents [35]. Also the developpment of faux leather substitutes, mainly consist of polyvinyl chloride (PVC) or polyurethane (PU) layered onto a backing fabric (synthetic or natural, such as cotton or organic waste), undergo a surface coating procedure to boost their resilience and longevity. A recent study on the composting capacity of leathers[36] found that bovine leather treated with alginate derivatives degraded completely within 21 to 25 days, conventionally produced wet-blue leather degraded within 31 to 35 days, vegetable-tanned bovine leather showed initial signs of degradation after 60 days but did not fully disintegrate even after 90 days, whreas alternative materials, containing non-biodegradable components like PU and PVC, showed no degradation after 90 days.
Current compostability standard ISO 17088:2021[37] developped specifically for plastic compostability evaluation and has been commonly used to evaluate leather compostability. Leather and plastic have different physico-chemical characteristics and show different ways of degradation. Plastics degrades into smaller particles and poses a microplastic generation hazzard whereas leather posses chemical contanimation hazzard of the final compost. In this context International Standard ISO 20136:2020 “Leather. Determination of degradability by micro-organisms”[38] was developed specifically for leather. This methodology uses complex consortium of microorganisms from the tannery and urban wastewaters as inoculum in liquid medium to measure the biodegradation potential of leather as a meassure of CO2 generated during leather degradation in 28 days.
This article aims to present recent research discoveries regarding various front lines; the identification and isolation of microorganisms from tannery wastewaters; identification of microbial composition in the starting inoculum used for a leather biodegradation assay according to ISO 20136 (tannery and municipal wastewater); and identifying microbial diversity shifts in initial inoculum during the process of different tanned leather degradation. The aim is to identify which genera is capable of biodegrading what type of tanning agent as well as which genera is acting at the different stages of degradation.
The focus is exploring their potential applications in enhancing leather biodegradability, wastewater treatment within the leather industry, and facilitating bioremediation processes for heavy metals. This study provides crucial insights into sustainable solutions for addressing environmental challenges within the leather industry, paving the way for developing efficient and environmentally friendly treatments for tannery wastewater. It will also contribute to optimizing leather composting processes, ultimately reducing the environmental impact of tannery operations and reinforcing the leather industry’s commitment to responsible and sustainable production.
2. Materials and Methods
2.1. Species Identification from Tannery Wastewaters
2.1.1. Tannery Wastewater Collection and Preparation
A two-litre water sample was taken from an aeration pond at a wastewater treatment plant (Curtidos Serpiel S.A., Caudete, Spain) at a pH of 4.5 and estimated chromium concentration of ~100 ppm. The water sample was filtered through a paper filter to remove macroscopic precipitates. Part of the filtrate was supplemented with 0.5% yeast to promote growth, and bacteriological agar was added up to 1%. A set of dilutions was prepared from the remaining filtered water up to 10-4 to the initial concentration. 100 μL of each dilution was used to seed the solid media initially prepared. Two plates were seeded for each dilution and incubated at room temperature for 10 days.
2.1.2. Species Isolation
Colonies from the 10-3 dilution were resuspended in LB medium and reseeded on fresh plates at different dilutions. Up to 9 microorganisms were isolated and maintained through successive reseeding in standard LB medium.
2.1.3. Species Identification
Species identification was carried out by 16S/18S rRNA amplification and sequencing. Eight colonies were randomly selected and resuspended in 20 μL of ultrapure H2O, incubated at 99 °C for 15 minutes to lyse the cells. The NZY Microbial gDNA Isolation kit (NZYtech, Lisboa, Portugal) [39] was used to extract the DNA sample for PCR with Taq DNA polymerase. Three pairs of degenerate universal oligonucleotides specific for the 16S/18S gene of bacteria, archaea, and eukaryotes were designed. - BACF: 5′-AGAGTTTGATCCTGGCTCAG-3′ - BACR: 5′-GGYTACCTTGTTACGACTT-3′ - ARCF: 5′-TCCGGTTGATCCYGCBRG-3′ - ARCR: 5′-TTMGGGGCATRCIKACCT-3′ - EUKF: 5′-GGTTGATYCTGCCAGTAG-3′ - EUKR: 5′-GTACACACCGCCCGTCGCT-3′.
The PCR product obtained was sequenced using an automated sequencer and bacterial oligonucleotides, following two strategies: cloning the PCR products into the pGEM-T easy vector (Promega, Madison, United States) and sequencing the PCR product directly after purifying the bands with the GFX PCR DNA and Gel Band Purification Kits (Cytiva, United States) [40].
2.2. Microorganism Identification from Leather Biodegradation Assay
2.2.1. ISO:20136:2020: Determination of Leather Degradability by Microorganisms’ Assay
A leather biodegradation assay (method B) was performed as described in ISO20136:2020 [38]. The inoculum was a mixture of 50:50 (ratio) of tannery and municipal sewage wastewater. Municipal wastewaters were collected from the local area treatment plant and tannery wastewaters were collected from Curtidos Segorbe S.L. (Segorbe, Spain) [41] a different source of tannery waste water to that mention in 2.1.1. Pure collagen from bovine Achilles tendon (Sigma-Aldrich®, Missouri, United States) [42]was used as a positive control (control sample). The assay was run for approximately 800 hours (33 days). Five differently tanned leather samples were used for the assay; 0.5 g of each sample was placed in each Erlenmeyer flask containing minimal salts and the inoculum, as shown in Table 1. The total organic carbon content of the material being tested is determined by elemental analysis. This allows the theoretical maximum quantity of carbon dioxide evolution to be calculated as biodegradation.
2.2.2. Wastewater and Leather Biodegradation Assay Sample Collection
Table 2 shows the samples taken from each Erlenmeyer flask, the hour at which they were withdrawn after the start of the assay, and the leather biodegradation stage registered at that time. According to the standard either tannery or municipal wastewater can be used in the assay. Sample M1 was municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50) from M1 and M2. This was the inoculum used in the assay and microorganism profile identified in samples M4 to M29 shift from this sample (M3). The aim of microorganism identification in samples M1 and M2 was merely informative to confirm and compare the final microorganism profile in sample M3. The remaining samples were drawn from an ongoing leather biodegradation assay (ISO 20136 assay) [43] at different stages of the degradation process (initial, exponential, and final) using a syringe and 0.1 microlitre filters. All samples were stored and transported at -20 °C.
2.2.3. DNA Extraction and Quality Control
Cells were recovered from filters followed by DNA isolation using the Qiagen QIAsymphony PowerFecal Pro DNA Kit (Qiagen, Hilden, Germany) [44]. That involved cellular lysis through mechanical disruption and enzymatic treatment. Subsequently, DNA purification from the sample was performed using a silica/gel column, allowing DNA isolation and removing contaminants and inhibitors for future reactions using the QIAamp DNA Micro Kit (Qiagen, Hilden, Germany) [45]. Cellular lysis was carried out through mechanical disruption and enzymatic treatment. Subsequently, DNA purification from the sample was performed using a silica/gel column, which enables DNA isolation and removal of contaminants and inhibitors for future reactions using the Qiagen QIAamp DNA Micro Kit. The quality and concentration of the DNA were evaluated using Nanodrop.
2.2.4. Sequence Library Preparation
50 ng of extracted DNA was amplified using a two-stage PCR protocol, the 16S Metagenomic Sequencing Library Preparation protocol Illumina 15044223. Primers were designed with the following structure: 1) an amplicon consisting of a universal linker sequence that allows the incorporation of indexes and sequencing primers using the Nextera XT Index kit [46]; and 2) universal primers for the 16S rRNA gene [47].
2.2.5. Sequencing
The sequencing libraries were prepared and loaded onto the Illumina MiSeq platform following a 300 bp x 2 paired-end design. In the second and final step of the assay, amplification indexes were included. The resulting 16S libraries were quantified using fluorimetry with the Quant-iT™ PicoGreen™ dsDNA Assay kit (Thermo Fisher scientific, Waltham, Massachusetts, United States) ([48]. Libraries were pooled before sequencing on the MiSeq platform (Illumina) following 300 paired end-reads design cycles. The size and concentration of the pool were evaluated using Agilent Bioanalyzer 2100 (Agilent, California, United States) [49]. The PhiX Control library (v3) (Illumina) was combined with the amplicon library (with an expected percentage of 20%). The sequencing data became available in approximately 56 hours. Image analysis, base calling, and data quality control were performed on the MiSeq platform (MiSeq Control Software (MCS v3.1)). The raw sequences, forward (R1) and reverse (R2), were merged to obtain the complete sequence using the BBMerge package of the BBMap software V.38. With this approach, the ends of the sequences overlapped to obtain complete sequences. Sequencing adapters were searched for and removed using the Cutadapt program (v 1.8.1) to reduce bias in the subsequent annotation stage.
2.2.6. Bioinformatic Analysis
Identifying and removing non-genomic regions or those with poor quality were carried out. Initially, the BBMerge module of BBMap software V.38 was employed to merge each pair of sequences (R1 and R2) from the sequencing platform, ensuring a minimum overlap of 70 nts at each end. This process resulted in a unique and complete sequence. Subsequently, the Cutadapt v1.8.1 program was used to detect and eliminate sequencing adapters from both ends in each sample. Once adapter-free sequences were obtained, reads with quality below Q20 and lengths less than 200 bp were removed. The Reformat module of BBMap V.38 was utilized for this analysis, enabling the trimming of nucleotides with a quality value below Q20 from both ends.
The final step in quality processing involved eliminating potential chimaera sequences resulting from incomplete extension during PCR amplification. This step was performed using the cd-hit-dup module, part of the cd-hit 4.8.1 software, predicting these amplicons de novo from the sample. The resulting sequences were then used for annotation. Sequences sharing 99% similarity were grouped into a single sequence using the “cd-hit” program. The outcomes were applied to the sequence group represented by the analyzed one, referred to as the Operational Taxonomic Unit (OTU). Subsequently, each sequence group was compared against the RefSeq 16S rRNA gene database (NCBI) using the local alignment BLAST strategy to associate each group with a taxonomic group from the database.
3. Results
3.1. Species Identification from Tannery Wastewaters Treatment Plant (Curtidos Serpiel S.A., Caudete, Spain)
3.1.1. Species Identification
Agarose gel electrophoresis (1%) of PCR products shown in Figure 1 was obtained from eight colonies using oligonucleotides for bacteria, archaea, and eukaryotes. The bands that can be seen from B1 to B8 in Figure 1 are of the same size as those seen in Figure 2.
All PCR products were sequenced with the corresponding oligonucleotides at the Genomics and Proteomics Service of the University of Alicante Technical Services. The sequences obtained in each case were compared in the NCBI databases and EZBioCloud. The identified bacterial strains are shown in Table 3. Strains have been identified by sequencing the whole bacterial 16S rRNA gene, therefore species indicating a similarity percentage over 99% are considered to be an exact match [51]. Other species would have to be identified by whole genome sequencing.
3.2. Microorganism Identification from Leather Biodegradation Assay Using ISO 20136:2020
3.2.1. ISO:20136: Leather- Determination of Degradability by Microorganisms
Biodegradation results for leather samples described in section 2.2.1 are shown in Figure 3. These biodegradation curves show the % of leather biodegradation throughout the assay (30 days); the times shown in Table 2 (section 2.2.2) correspond to the time (hours) in this graph. There is an initial, exponential, and final phase of leather biodegradation for all the samples at different levels of biodegradation percentage. Collagen is used as a positive control since it fully degrades in approximately 30 days [38]. At the final stage of the assay (747 h since time 0), when the last samples were taken, the biodegradation percentage for collagen was 81.5%, oxazolidine 59.4%, glutaraldehyde 5.2%, chromium 7.6% and aluminium 23.2%.
3.2.2. Sequencing
The concentration and purity of the extracted DNA reached enough levels to proceed with the preparation of sequencing libraries. The sequencing results, including the number of raw sequences obtained, average length, total sequenced bases measured in Megabases (Mb), and the mean quality for forward (R1) and reverse (R2) sequences, are shown in Table S1 within the supplementary materials. The read count exceeded 50,000 in all analysed samples, except for sample 210527A-M4, which yielded 43,447 reads.
3.2.3. Bioinformatics and Species Identification
Although this sample did not reach the desired read count during the sequencing process, this quantity proved optimal as it reached the plateau zone in the rarefaction curve. The profiles are studied at a maximum genus level, providing species-level information for discussion on potentially implicated species. However, due to the product size of around 400 nts for bacteria, precise species identification cannot be guaranteed.
It can be determined that it is close to the plateau zone with the inspected number of sequences and the microbial profile. As shown in the rarefaction curves (alpha-diversity index) in Figure 4, most samples are in the plateau zone, indicating that an increase in sequences would not yield a significant rise in the number of newly detected genera.
The statistical analysis employing the vegan package in R [52] allows for the examination of the organism-abundance relationship. These results are presented in the Table 4. The Shannon index estimates the specific biodiversity present in the sample, providing a positive numeric value starting from 0 (indicating a single species) and increasing as diversity increases [53]. The Chao 1 value estimates the total number of species that may be present in the sample based on the number of less represented species in the sample [54]. In taxon-based approaches, accurately estimating the number of microbial species in a sample is challenging due to the complexity of microbial diversity, therefore assessing species richness is crucial for understanding biological communities effectively [55]. Richness estimators like Chao 1 are employed to deduce the total richness of a microbial community based on observed Operational Taxonomic Units (OTUs). Unlike rarefaction, which compares observed richness among samples, richness estimators predict total richness from a single sample. For all the samples, Shannon index is around 4 and Chao 1 index was around 2500. Sample M4 has shown to be the only one that differs from other samples showing a Shannon index of 2.70 and a Chao 1 of 266.
Local BLAST alignment is conducted to associate each of the obtained reads (after cleaning and filtering) with an organism. Working with a well-curated database is crucial, as databases may be incomplete or lack certain organisms due to being sequenced or not being taxonomically assigned. In cases where a sequence is associated with multiple hits with the same e-value and identity, the first hit is considered the best. It’s important to note that these taxonomic identifications represent the best outcome in local alignment, not guaranteeing the organism’s presence in the sample.
Short sequences are excluded from the analysis to reduce the likelihood of duplicating sequences in the database and encountering false positives or misassignments. Sequencing errors that can alter assignments to a specific organism by modifying sequence similarity are also eliminated from the analysis. For each sequence, details such as percentage identity with the database hit, e-value, absolute number of identical positions in the alignment, relative number of identities considering the total number of alignment bases, absolute number of gaps in the alignment, and relative number of gaps considering the total number of alignment bases are provided.
Once each read is associated with an organism, various analyses and filters are applied. Reads are grouped based on family, genera, and species, with associations typically made at the family or genera level, but Species-level associations are also provided. The database is curated according to different taxonomic levels described in the NCBI database [56]. Some levels may remain null if they are not characterised, cultivated, or unknown. The presence of different taxonomic groups in the analysed samples is summarized in tables and graphs. A rarefaction curve is generated for each analysed sample, comparing the number of analysed sequences with the number of detected taxa at different levels, as shown in Figure 4. This curve helps determine if the detection has reached saturation, indicating that all organisms have been detected or if more sequences are needed to capture the total variability of the sample.
Sample M4 (collagen at 17% biodegradation at 52 h since start of the assay) has shown to be the only one that differs from other samples showing a Shannon index of 2.70 and a Chao 1 of 266, considerably lower to the rest of the reads. This is also shown in Figure 4, where the alpha-diversity index of M4 is considerably lower than the rest of the samples. The reduction of these is caused by the immense presence of Acinetobacter in the sample. This bacterium shows to be a great collagen degrader, as its presence grows exponentially within the first 48 hours within the assay. On the other hand, M3, being the mix of both wastewaters, shows the highest alpha-diversity index as well as one the highest Shannon index Chao parameters, 5.05 and 3383, respectively.
Bacterial abundance detected in the analysed samples at genera level is shown in Figure 5, M1-M29 samples refer to those shown previously in Table 2 and Table 4. Additionally, this diagram incorporates the LCBD beta diversity index, revealing diversity patterns. High LCBD values indicate that the bacterial composition of the sample significantly differs from the rest of the studied samples [57]. Bacterial strains identified in highest percentages in municipal wastewaters (sample M1) were Nakamurella endophytica (10.76%), Clostridium saudiense (9.87%), Romboutsia timonensis (9.21%) and Mycolicibacterium peregrinum (3.41%). On the other hand, the bacterial strains most present in tannery wastewaters (sample M2) were Pseudonocardia rhizophila (13.02%), Variibacter gotjawalensis (11.46%), Pseudorhodoplanes sinuspersici (6.42%), Hyphomicrobium aestuarii (3.46%) and M. peregrinum (3.38%). The inoculum used for the assay was sample M3 a 50:50 mixture of M1 and M2 and, as it would be expected, the composition in terms of bacterial presence is halfway between M1 and M2.
For comparison purposes, the results of bacterial identification have been categorized according to the different tanned leather samples. This allows for a comparison between different tanning agents, as well as between various stages of biodegradation within the same type of leather. M1, M2 and M3 have been left in all the following bar plot graphs since they represent the initial bacterial composition of the inoculum. M3 was the inoculum used in the assay, therefore all the bacterial shift in the following samples diverge from sample M3. Figure 6 (a) shows the bacterial abundance at species level detected in the analysed samples for collagen (control sample). As collagen biodegradation takes place (samples M4 – M25) the most abundant species present are Brevundimonas terrae, A. johnsonii and M. peregrinum. Figure 6 (b) shows a line graph representing bacterial presence shift in the sample as a percentage of the top six bacterial species identified at the different stages of collagen biodegradation. Within the first 52 hours after the start of tha assay the most predominant genius is Acinetobacter Some of the species, such as Chryseobacterium indoltheticum, start at very high concentrations, but quickly decrease as they phase no possibility to proliferate using collagen as the only carbon source.
Figure 7 (a) shows the bacterial abundance at species level detected in the analysed samples for chromium tanned leather. As leather biodegradation takes places (samples M4 – M25) the most abundant species present are Brevundimonas terrae, Acinetobacter johnsonii and Mycolicibacterium peregrinum. Figure 7 (b) shows a line graph representing bacterial presence shift in the sample as a percentage of the top six bacterial species identified at the different stages of chromium leather biodegradation. In this case, Brevundimonas is the predominant bacterial genus, Brevidumonas terrae shows to be the bacterial strain most present in samples M11 to M4 shifting to Brevundimonas kwangchunensis which is present as a 15.9% in the sample.
Figure 8 (a) shows the bacterial abundance at species level detected in the analysed samples for glutaraldehyde tanned leather. As leather biodegradation takes place (samples M10 – M27) the most abundant species present are A. johnsonii, B. terrae and M. peregrinum. Figure 8 (b) shows a line graph representing bacterial presence shift in the sample as a percentage of the top six bacterial species identified at the different stages of glutaraldehyde leather biodegradation. In this case, A. johnsonii is the predominant bacterial strain being present in above 10% in all the samples M10 to M27.
Figure 9 (a) shows the bacterial abundance at species level detected in the analysed samples for oxazolidine tanned leather. As leather biodegradation takes place (samples M6 – M26) the most abundant species present are B. terrae, A. johnsonii and M. peregrinum. Figure 9 (b) shows a line graph representing bacterial presence shift in the sample as a percentage of the top six bacterial species identified at the different stages of oxazolidine leather biodegradation. In this case, B. terrae is the predominant bacterial strain in all samples M6 to M26, always present in around 20% in all the samples.
Figure 10 (a) shows the bacterial abundance at species level detected in the analysed samples for aluminium tanned leather. As leather biodegradation takes place (samples M7 – M29) the most abundant species present are B. terrae, A. johnsonii and M. peregrinum. Figure 10 (b) shows a line graph representing bacterial presence shift in the sample as a percentage of the top six bacterial species identified at the different stages of aluminium leather biodegradation. In this case, B.terrae is the predominant bacterial strain in all samples M6 to M26, always present in around 20% in all the samples.
4. Discussion
4.1. Species Identification from Tannery Wastewaters
Though the years there has been different standards fixation for the use of 16S rRNA genes in taxonomy identification[58]. The latest cutoff value at the species level has been evaluated at 98.7%[59], however several authors have shown that these thresholds are not applicable to multiple genera[60]. Isolated species 1 Dietzia maris has been previously isolated from soil for zinc bioremediation [61], for petroleum hydrocarbons and crude oil degradation [62,63] and tolerance to heavy metals such as cadmium or cobalt [64] and multiple-extreme resistance [65]. Species 2 T. pasteurii has been previously identified for the reduction of hexavalent chromium [66], showing two potential pathways for Cr (VI) removal; sulfidogenesis-induced Cr (VI) reduction pathway, 90% Cr (VI) removal by sulfide generated from biological reduction of sulfate. The second being direct 10% Cr (VI) removal by bacteria as the electron acceptor [67]. Corynebacterium lubricantis has been previously isolated from chromite mine seepage of Odisha as a heavy metal tolerant and chromate reducing bacterium [68], as well as isolated from a chromium-polluted soil, tested for chromate reduction capability and multiple heavy metal tolerance up to a concentration of 22 mM [69]. Microbacterium strains have been previously studied for chromium waste biocementation [70] and isolated from tannery wastewaters for hexavalent chromium reduction [71]. B. safensis has been isolated from tannery effluent as a chromium (Cr) and tannic acid (TA) resistance bacterial strain [72], isolated from contaminated coal mining soil for chromium reduction [73], and isolated from rare-earth ore for hexavalent chromium conversion to trivalent chromium, where an gene nfrA is involved [74]. All these species represent good candidates for further investigation in which they are submitted to stress condition and evaluation of the functional groups in heavy metal bioremediation.
Identification of the specific functional groups responsible for metal ion binding to microbial biomass is crucial for understanding the biosorption mechanism in efficient methods for removing heavy metals from contaminated environments [75]. The type, structure, and arrangement of functional groups can vary significantly between microorganisms[76], with many of these groups primarily identified on microbial cell walls[77]. Functional groups such as aldehydes, alkyl chains, amides, amines, alcohols/phenols, carboxylic acids, esters, organic halides, phosphates, sulfoxides, and aliphatic organic chains of cellulose have been identified as key players in the biosorption of chromium[78]. Multiple spectroscopic and microscopic techniques, such as infrared and Raman spectroscopy, electron dispersive spectroscopy and nuclear magnetic resonance (NMR) have been identified for active sites involved in binding of heavy metal ions identification[79]. However, most studies employ Fourier-transform infrared spectroscopic (FT-IR) technique to identify and characterize certain functional groups present in microbial biomass for uptake of toxic heavy metals such as hexavalent chromium[80]. Specific functional groups and certain mechanisms for Cr biosorption already identified for certain species such as Bacillus marisflavis and Bacillus arthrobacter [81] as well Klebsiella sp.[82] include _OH, –NH acetamido group, amide bond, C=O of COO-, free phosphates, phosphate groups, –CN and NH2, O-H, -CONH-, -COOH, C = C, -CH2 (Freundlich adsorption isotherm) respectively.
4.2. Microorganism Identification from Leather Biodegradation Assay
4.2.1. ISO:20136:2020: Leather- Determination of Degradability by Microorganisms
Leather biodegradation process is heavily dependent on the tanning agent employed within the tanning process giving leather specific physical-chemical properties[1]. Pure collagen’s exponential degradation phase shows to be in the first 16 h of assay. Oxazolidines are cyclic condensation products of β-amino alcohols and aldehydes or ketone [64]. The ease of hydrolysis of oxazolidines in aqueous solution relates directly to their structural features, a cyclic ring structure that contains both a nitrogen atom and a carbonyl group [65]. These functional groups are susceptible to nucleophilic attack during hydrolysis, leading to the cleavage of the ring and the formation of the corresponding β-amino alcohol and aldehyde or ketone [66]. This leads into a higher biodegradation potential, like mineral based tanning methods such as zeolites, which have shown 81% biodegradation potential in 30 days [67]. Metal-tanned hides form stable cross-links between the collagen fibres, rendering the material less susceptible to enzymatic attack by microorganisms. These can exhibit toxicity to microorganisms, thereby inhibiting their ability to degrade the leather.[68] The presence of toxic metal ions can disrupt microbial activity, slowing down the biodegradation process [69].
4.2.2. Bioinformatics and Species Identification
More than 10,000 bacterial species have been identified to be present within all the analysed samples of the leather biodegradation assay and 8 bacterial strains have been isolated from tannery wastewaters. Bacterial genera identified and isolated from the first tannery wastewater sample, described in Table 3 have been identified in low percentages in sample M2 (tannery wasterwater) is as follows: D. maris 0% T. pasteurii 0.012%, C. lubricantis 0%, M. laevaniformans 0% and B. safensis 0%. Tannery wastewaters are generally characterised by its dark brown colour and high levels of pollutants total dissolved solids (TDS), chromium, phenolics, and high pH [83]. Bacterial diversity difference amongst these samples could be given due to changes in these conditions, which vary depending at which stage and time at which the sample is taken as well as the type of tanning that has been performed [84].
The most common identified microorganisms throughout the assay are the following: Acinetobacter, Brevundimonas and Mycolicibacterium. Acinetobacter genus is commonly found in aquatic environments such as wastewater and river waters [85]. Four Acinetobacter strains (A. johnsonii, A. modestus, A. tjernbergiae, A. junii) have been identified. These species have previously been identified and isolated from tannery and residual wastewater, characterised and evaluated for chromium bioremediation [86,87,88,89] and Cr6+ transformation to Cr3+ [90,91]. Brevundimonas has been mainly identified and isolated from sedimented waters [92], rivers [93] and soil samples [94,95,96]. Brevundimonas strains have also been identified and isolated for heavy metal bioremediation [97,98], cadmium and zinc bioremediation [99], and arsenic resistance [100,101]. Mycolicibacterium is a non-tuberculous identified and isolated in cotton fields [102], peat bog [103], sea coast [104] and mangrove sediment [105]. This strain has been previously identified and isolated for zinc-lead bioremediation [106].
Within the first 16 hours since the start of the assay, at the exponential phase of collagen biodegradation (M4 and M5) 60% and 37% of the inoculum consists of the genius Acinetobacter along with Brevundimonas present in 6.75% and 16% respectively. Within the bacterial profile for chromium samples, Variibacter gotjawalensis has been found to be present over or around 5% in all samples and 11.6% in samples M28 at the final stages of biodegradation assay (7.5% of leather biodegradation). This strain only appears significantly in samples from chromium tanned leather assay. This strain has been previously isolated from soil in a lava forest in Korea[107] and tested in Cd-Zn-Pb-contaminated soil for phytoextraction[108].
It must be noted that these identified genera (Acinetobacter, Brevundimonas and Mycolicibacterium) are merely present in initial inoculum (M3), Mycolicibacterium is around 5% and the other two do not reach 1%. There is an immense jump to such higher percentages as leather biodegradation takes places. The amount of leather sample and therefore of tanning agents are not significant to evaluate from these results the capability of the identified species to degrade and/or tolerate these molecules in higher concentrations. It is clear from the results that they have the capability of bond breakage between the tanning agent and the collagen fibres and biodegrade collagen itself. For certain tanning agents such as glutaraldehyde (Figure 8b) and oxazolidine (Figure 9b), Acinetobacter johnsonii and Brevundimonas terrae respectively, are the present species and therefore the most active species in terms of leather biodegradation. Further investigation would have to be carried out for a deeper understanding of species biodegradation mechanisms.
5. Conclusions
This study has contributed to the understanding of the microbial diversity and abundance in the context of leather biodegradation and heavy metal resistance. Despite the great abundance of identified bacterial species, the identification of bacterial genera such as Acinetobacter, Brevundimonas, and Mycolicibacterium in the samples has provided valuable insights into the potential microbial candidates showcasing their potential applications in enhancing leather biodegradability, wastewater treatment, and bioremediation processes for heavy metals.
Overall, the findings of this study underscore the importance of sustainable solutions for addressing environmental challenges within the leather industry. The selective isolation and identification of microorganisms with dual capabilities offer promising prospects for the development of efficient and environmentally friendly treatments for tannery wastewater, leather biodegradation, and heavy metal remediation, thereby contributing to the advancement of sustainable practices in the leather industry.
Author Contributions
Conceptualization, Marcelo Bertazzo and María-José Bonete; Data curation, Manuela Bonilla-Espadas, Basilio Zafrilla and Irene Lifante-Martínez; Formal analysis, Manuela Bonilla-Espadas; Funding acquisition, Elena Orgilés-Calpena and María-José Bonete; Investigation, Manuela Bonilla-Espadas and Basilio Zafrilla; Methodology, Manuela Bonilla-Espadas, Basilio Zafrilla and Irene Lifante-Martínez; Project administration, Elena Orgilés-Calpena, Francisca Arán-Aís and María-José Bonete; Resources, Mónica Camacho; Supervision, Elena Orgilés-Calpena, Francisca Arán-Aís, Marcelo Bertazzo and María-José Bonete; Validation, Manuela Bonilla-Espadas; Writing – original draft, Manuela Bonilla-Espadas; Writing – review & editing, Mónica Camacho, Marcelo Bertazzo and María-José Bonete.
Funding
This research was co-financed by the European Union through the European Regional Development Fund, within the Operational Programme of the Valencian Community 2014-2020 within the BIOREQ project with grant number IMDEEA/2021/11; Project UAIND21-02B from University of Alicante.
Data Availability Statement
The datasets presented in this study are available on request to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Covington, A.D. Tanning Chemistry: The Science of Leather; Royal Society of Chemistry, 2009; ISBN 978-0-85404-170-1.
- Muthukrishnan, L. Nanotechnology for Cleaner Leather Production: A Review. Environ Chem Lett 2021, 19, 2527–2549. [Google Scholar] [CrossRef]
- Hao, D.; Wang, X.; Liang, S.; Yue, O.; Liu, X.; Hao, D.; Dang, X. Sustainable Leather Making — An Amphoteric Organic Chrome-Free Tanning Agents Based on Recycling Waste Leather. Science of The Total Environment 2023, 867, 161531. [Google Scholar] [CrossRef] [PubMed]
- Rosu, L.; Varganici, C.; Crudu, A.; Rosu, D.; Bele, A. Ecofriendly Wet–White Leather vs. Conventional Tanned Wet–Blue Leather. A Photochemical Approach. Journal of Cleaner Production 2018, 177, 708–720. [Google Scholar] [CrossRef]
- Gao, D.; Li, X.; Cheng, Y.; Lyu, B.; Ma, J. The Modification of Collagen with Biosustainable POSS Graft Oxidized Sodium Alginate Composite. International Journal of Biological Macromolecules 2022, 200, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Ariram, N.; Madhan, B. Development of Bio-Acceptable Leather Using Bagasse. Journal of Cleaner Production 2020, 250, 119441. [Google Scholar] [CrossRef]
- Hassan, M.M.; Harris, J.; Busfield, J.J.C.; Bilotti, E. A Review of the Green Chemistry Approaches to Leather Tanning in Imparting Sustainable Leather Manufacturing. Green Chem. 2023, 25, 7441–7469. [Google Scholar] [CrossRef]
- Hansen, É.; de Aquim, P.M.; Gutterres, M. Environmental Assessment of Water, Chemicals and Effluents in Leather Post-Tanning Process: A Review. Environmental Impact Assessment Review 2021, 89, 106597. [Google Scholar] [CrossRef]
- Basaran, B.; Ulaş, M.; Bitlisli, B.; Aslan, A. Distribution of Cr (III) and Cr (VI) in Chrome Tanned Leather. Indian Journal of Chemical Technology 2008, 15, 511–514. [Google Scholar]
- Anderson, R.A. Nutritional Role of Chromium. Science of The Total Environment 1981, 17, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, H.; Gu, Y.; Song, X.; Zhao, J. Carcinogenicity of Chromium and Chemoprevention: A Brief Update. OncoTargets and Therapy 2017, 10, 4065–4079. [Google Scholar] [CrossRef]
- Chojnacka, K.; Skrzypczak, D.; Mikula, K.; Witek-Krowiak, A.; Izydorczyk, G.; Kuligowski, K.; Bandrów, P.; Kułażyński, M. Progress in Sustainable Technologies of Leather Wastes Valorization as Solutions for the Circular Economy. Journal of Cleaner Production 2021, 313, 127902. [Google Scholar] [CrossRef]
- Leather Goods Market - Industry Analysis Report 2032. Available online: https://www.gminsights.com/industry-analysis/leather-goods-market (accessed on 30 January 2024).
- Oruko, R.O.; Selvarajan, R.; Ogola, H.J.O.; Edokpayi, J.N.; Odiyo, J.O. Contemporary and Future Direction of Chromium Tanning and Management in Sub Saharan Africa Tanneries. Process Safety and Environmental Protection 2020, 133, 369–386. [Google Scholar] [CrossRef]
- Fela, K.; Wieczorek-Ciurowa, K.; Konopka, M.; Woźny, Z. Present and Prospective Leather Industry Waste Disposal. Polish Journal of Chemical Technology 2011, 13, 53–55. [Google Scholar] [CrossRef]
- Alam, N.; Sayid Mia, Md.A.; Ahmad, F.; Rahman, M. An Overview of Chromium Removal Techniques from Tannery Effluent. Applied Water Science 2020, 10. [Google Scholar] [CrossRef]
- Benalia, M.C.; Youcef, L.; Bouaziz, M.G.; Achour, S.; Menasra, H. Removal of Heavy Metals from Industrial Wastewater by Chemical Precipitation: Mechanisms and Sludge Characterization. Arab J Sci Eng 2022, 47, 5587–5599. [Google Scholar] [CrossRef]
- Kocaoba, S.; Cetin, G.; Akcin, G. Chromium Removal from Tannery Wastewaters with a Strong Cation Exchange Resin and Species Analysis of Chromium by MINEQL+. Sci Rep 2022, 12, 9618. [Google Scholar] [CrossRef] [PubMed]
- Younas, F.; Niazi, N.K.; Bibi, I.; Afzal, M.; Hussain, K.; Shahid, M.; Aslam, Z.; Bashir, S.; Hussain, M.M.; Bundschuh, J. Constructed Wetlands as a Sustainable Technology for Wastewater Treatment with Emphasis on Chromium-Rich Tannery Wastewater. Journal of Hazardous Materials 2022, 422, 126926. [Google Scholar] [CrossRef] [PubMed]
- Boussouga, Y.-A.; Okkali, T.; Luxbacher, T.; Schäfer, A.I. Chromium (III) and Chromium (VI) Removal and Organic Matter Interaction with Nanofiltration. Science of The Total Environment 2023, 885, 163695. [Google Scholar] [CrossRef] [PubMed]
- Villaseñor-Basulto, D.L.; Kadier, A.; Singh, R.; Navarro-Mendoza, R.; Bandala, E.; Peralta-Hernández, J.M. Post-Tanning Wastewater Treatment Using Electrocoagulation: Optimization, Kinetics, and Settlement Analysis. Process Safety and Environmental Protection 2022, 165, 872–886. [Google Scholar] [CrossRef]
- Krishna, R.; Chintalpudi, V.; Muddada, S. Application of Biosorption for Removal of Heavy Metals from Wastewater. In; 2018 ISBN 978-1-78923-472-5.
- Fernández, P.M.; Viñarta, S.C.; Bernal, A.R.; Cruz, E.L.; Figueroa, L.I.C. Bioremediation Strategies for Chromium Removal: Current Research, Scale-up Approach and Future Perspectives. Chemosphere 2018, 208, 139–148. [Google Scholar] [CrossRef] [PubMed]
- GracePavithra, K.; Jaikumar, V.; Kumar, P.S.; SundarRajan, P. A Review on Cleaner Strategies for Chromium Industrial Wastewater: Present Research and Future Perspective. Journal of Cleaner Production 2019, 228, 580–593. [Google Scholar] [CrossRef]
- Pradhan, D.; Sukla, L.B.; Sawyer, M.; Rahman, P.K.S.M. Recent Bioreduction of Hexavalent Chromium in Wastewater Treatment: A Review. Journal of Industrial and Engineering Chemistry 2017, 55, 1–20. [Google Scholar] [CrossRef]
- Sutkowy, M.; Kłosowski, G. Use of the Coenobial Green Algae Pseudopediastrum Boryanum (Chlorophyceae) to Remove Hexavalent Chromium from Contaminated Aquatic Ecosystems and Industrial Wastewaters. Water 2018, 10, 712. [Google Scholar] [CrossRef]
- Bakshi, A.; Panigrahi, A.K. Chromium Contamination in Soil and Its Bioremediation: An Overview. In Advances in Bioremediation and Phytoremediation for Sustainable Soil Management: Principles, Monitoring and Remediation; Malik, J.A., Ed.; Springer International Publishing: Cham, 2022; pp. 229–248. ISBN 978-3-030-89984-4. [Google Scholar]
- Cunningham, S.D.; Ow, D.W. Promises and Prospects of Phytoremediation. Plant Physiology 1996, 110, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Landfill Waste - European Commission. Available online: https://environment.ec.europa.eu/topics/waste-and-recycling/landfill-waste_en (accessed on 29 January 2024).
- Directive 2008/98/EC of the European Parliament and of the Council of 19 November 2008 on Waste and Repealing Certain Directives (Text with EEA Relevance); 2008; Vol. 312;
- Ding, W.; Liu, H.; Remón, J.; Jiang, Z.; Chen, G.; Pang, X.; Ding, Z. A Step-Change toward a Sustainable and Chrome-Free Leather Production: Using a Biomass-Based, Aldehyde Tanning Agent Combined with a Pioneering Terminal Aluminum Tanning Treatment (BAT-TAT). Journal of Cleaner Production 2022, 333, 130201. [Google Scholar] [CrossRef]
- A Guide to Modern Leather Making. Available online: https://www.leathernaturally.org/a-guide-to-modern-leather-making/ (accessed on 31 March 2024).
- Yu, Y.; Lin, Y.; Zeng, Y.; Wang, Y.; Zhang, W.; Zhou, J.; Shi, B. Life Cycle Assessment for Chrome Tanning, Chrome-Free Metal Tanning, and Metal-Free Tanning Systems. ACS Sustainable Chem. Eng. 2021, 9, 6720–6731. [Google Scholar] [CrossRef]
- Zhou, Y.; Ma, J.; Gao, D.; Li, W.; Shi, J.; Ren, H. A Novel Chrome-Free Tanning Approach Based on Sulfonated Tetraphenyl Calix [4]Resorcinarene: Preparation and Application. Journal of Cleaner Production 2018, 201, 668–677. [Google Scholar] [CrossRef]
- China, C.R.; Maguta, M.M.; Nyandoro, S.S.; Hilonga, A.; Kanth, S.V.; Njau, K.N. Alternative Tanning Technologies and Their Suitability in Curbing Environmental Pollution from the Leather Industry: A Comprehensive Review. Chemosphere 2020, 254, 126804. [Google Scholar] [CrossRef]
- Sardroudi, N.P.; Sorolla, S.; Casas, C.; Bacardit, A. A Study of the Composting Capacity of Different Kinds of Leathers, Leatherette and Alternative Materials. Sustainability 2024, 16, 2324. [Google Scholar] [CrossRef]
- ISO ISO 17088:2021. Available online: https://www.iso.org/standard/74994.html (accessed on 19 March 2024).
- IULTCS ISO 20136:2020. Available online: https://www.iso.org/standard/75892.html (accessed on 20 November 2023).
- NZYMicrobial gDNA Isolation Kit. Available online: https://www.nzytech.com/en/mb21702-nzy-microbial-gdna-isolation-kit/ (accessed on 16 November 2023).
- GFX PCR DNA and Gel Band Purification Kits. Available online: https://www.cytivalifesciences.com/en/us/shop/molecular-and-immunodiagnostics/pcr-cleanup-and-size-selection/illustra-gfx-pcr-dna-and-gel-band-purification-kits-p-00386 (accessed on 1 February 2024).
- Español - Curtidos Segorbe S.L. Available online: http://www.curtidosegorbe.com/curtidos-segorbe-s-l/espanol/ (accessed on 22 February 2024).
- Collagen from bovine achilles tendon powder, suitable for substrate for collagenase | 9007-34-5. Available online: http://www.sigmaaldrich.com/ (accessed on 20 November 2023).
- Ake, A.H.J.; Hafidi, M.; Ouhdouch, Y.; Jemo, M.; Aziz, S.; El Fels, L. Microorganisms from Tannery Wastewater: Isolation and Screening for Potential Chromium Removal. Environmental Technology & Innovation 2023, 31, 103167. [Google Scholar] [CrossRef]
- QIAsymphony PowerFecal Pro DNA Kit. Available online: https://www.qiagen.com/us/products/discovery-and-translational-research/dna-rna-purification/dna-purification/microbial-dna/qiasymphony-powerfecal-pro-dna-kit?catno=938036 (accessed on 20 November 2023).
- QIAamp DNA Accessory Set, Micro and Mini Kits - QIAGEN. Available online: https://www.qiagen.com/us/products/discovery-and-translational-research/dna-rna-purification/dna-purification/genomic-dna/qiaamp-dna-kits?catno=56304 (accessed on 20 November 2023).
- Nextera XT DNA Library Prep Kit | Sequence Small Genomes, Plasmids, cDNA. Available online: https://emea.illumina.com/products/by-type/sequencing-kits/library-prep-kits/nextera-xt-dna.html (accessed on 20 November 2023).
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Research 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Quant-iTTM PicoGreenTM dsDNA Assay Kits and dsDNA Reagents. Available online: https://www.thermofisher.com/order/catalog/product/es/en/P7589 (accessed on 20 November 2023).
- Bioanalyzer Instruments for Sample Quality Control | Agilent. Available online: https://www.agilent.com/en/product/automated-electrophoresis/bioanalyzer-systems/bioanalyzer-instrument (accessed on 8 February 2024).
- Thermo Scientific™ Estándar de ADN GeneRuler Mix, listo para su uso - Marcadores de peso molecular de ácidos nucleicos Productos bioquímicos y reactivos. Available online: https://www.fishersci.es/shop/products/fermentas-generuler-ready-to-use-dna-ladder-mix-1/11531605 (accessed on 8 February 2024).
- Church, D.L.; Cerutti, L.; Gürtler, A.; Griener, T.; Zelazny, A.; Emler, S. Performance and Application of 16S rRNA Gene Cycle Sequencing for Routine Identification of Bacteria in the Clinical Microbiology Laboratory. Clinical Microbiology Reviews 2020, 33. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Preprint at 2022, 3–1.
- Graciano-Ávila, G.; Aguirre-Calderón, Ó.A.; Alanís-Rodríguez, E.; Lujan-Soto, J.E. Composición, Estructura y Diversidad de Especies Arbóreas En Un Bosque Templado Del Noroeste de México. Ecosistemas y recursos agropecuarios 2017, 4, 535–542. [Google Scholar] [CrossRef]
- Moreno, C.E. Métodos Para Medir La Biodiversidad. M&T–Manuales y Tesis SEA, Vol. 1. Zaragoza 2001, 84, 2. [Google Scholar]
- Kim, B.-R.; Shin, J.; Guevarra, R.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J Microbiol Biotechnol 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 20 February 2024).
- Ijaz, U.Z.; Sivaloganathan, L.; McKenna, A.; Richmond, A.; Kelly, C.; Linton, M.; Stratakos, A.C.; Lavery, U.; Elmi, A.; Wren, B.W. Comprehensive Longitudinal Microbiome Analysis of the Chicken Cecum Reveals a Shift from Competitive to Environmental Drivers and a Window of Opportunity for Campylobacter. Frontiers in microbiology 2018, 9, 2452. [Google Scholar] [CrossRef] [PubMed]
- Beye, M.; Fahsi, N.; Raoult, D.; Fournier, P.-E. Careful Use of 16S rRNA Gene Sequence Similarity Values for the Identification of Mycobacterium Species. New Microbes New Infect 2017, 22, 24–29. [Google Scholar] [CrossRef]
- Stackebrandt, E. Taxonomic Parameters Revisited: Tarnished Gold Standards. Microbial Today 2006, 33, 152. [Google Scholar]
- Rossi-Tamisier, M.; Benamar, S.; Raoult, D.; Fournier, P.-E. Cautionary Tale of Using 16S rRNA Gene Sequence Similarity Values in Identification of Human-Associated Bacterial Species. International Journal of Systematic and Evolutionary Microbiology 2015, 65, 1929–1934. [Google Scholar] [CrossRef]
- Rani, N.; Kaur, G.; Kaur, S.; Mutreja, V.; Pandey, N. Plant Growth-Promoting Attributes of Zinc Solubilizing Dietzia Maris Isolated from Polyhouse Rhizospheric Soil of Punjab. Curr Microbiol 2022, 80, 48. [Google Scholar] [CrossRef] [PubMed]
- Gharibzahedi, S.M.T.; Razavi, S.H.; Mousavi, M. Potential Applications and Emerging Trends of Species of the Genus Dietzia: A Review. Ann Microbiol 2014, 64, 421–429. [Google Scholar] [CrossRef]
- Venil, C.K.; Malathi, M.; Devi, P.R. Characterization of Dietzia Maris AURCCBT01 from Oil-Contaminated Soil for Biodegradation of Crude Oil. 3 Biotech 2021, 11, 291. [Google Scholar] [CrossRef] [PubMed]
- Gillard, B.; Chatzievangelou, D.; Thomsen, L.; Ullrich, M.S. Heavy-Metal-Resistant Microorganisms in Deep-Sea Sediments Disturbed by Mining Activity: An Application Toward the Development of Experimental in Vitro Systems. Frontiers in Marine Science 2019, 6. [Google Scholar] [CrossRef]
- Gholami, M.; Etemadifar, Z. Isolation and Characterization of a Novel Strain of Genus Dietzia Capable of Multiple-Extreme Resistance. Microbiology 2015, 84, 389–397. [Google Scholar] [CrossRef]
- Tandukar, M.; Huber, S.J.; Onodera, T.; Pavlostathis, S.G. Biological Chromium(VI) Reduction in the Cathode of a Microbial Fuel Cell. Environ. Sci. Technol. 2009, 43, 8159–8165. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Wei, L.; Liu, R.; Jiang, F.; Hao, X.; Chen, G.-H. An Exploratory Study on the Pathways of Cr (VI) Reduction in Sulfate-Reducing Up-Flow Anaerobic Sludge Bed (UASB) Reactor. Sci Rep 2016, 6, 23694. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Paul, A.K.; Dey, S.; Paul, A.K. Assessment of Heavy Metal Tolerance and Hexavalent Chromium Reducing Potential of <em>Corynebacterium Paurometabolum</Em> SKPD 1204 Isolated from Chromite Mine Seepage. AIMSBOA 2016, 3, 337–351. [Google Scholar] [CrossRef]
- Viti, C.; Pace, A.; Giovannetti, L. Characterization of Cr(VI)-Resistant Bacteria Isolated from Chromium-Contaminated Soil by Tannery Activity. Curr Microbiol 2003, 46, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lun, L.; Li, D.; Yin, Y.; Li, D.; Xu, G.; Zhao, Z.; Li, S. Characterization of Chromium Waste Form Based on Biocementation by Microbacterium Sp. GM-1. Indian J Microbiol 2016, 56, 353–360. [Google Scholar] [CrossRef]
- Mishra, S.; Chen, S.; Saratale, G.D.; Saratale, R.G.; Romanholo Ferreira, L.F.; Bilal, M.; Bharagava, R.N. Reduction of Hexavalent Chromium by Microbacterium Paraoxydans Isolated from Tannery Wastewater and Characterization of Its Reduced Products. Journal of Water Process Engineering 2021, 39, 101748. [Google Scholar] [CrossRef]
- Chaudhary, P.; Beniwal, V.; Umar, A.; Kumar, R.; Sharma, P.; Kumar, A.; Al-Hadeethi, Y.; Chhokar, V. In Vitro Microcosm of Co-Cultured Bacteria for the Removal of Hexavalent Cr and Tannic Acid: A Mechanistic Approach to Study the Impact of Operational Parameters. Ecotoxicology and Environmental Safety 2021, 208, 111484. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.; Vishwakarma, K.; Singh, J.; Mishra, M.; Kumar, V.; Rani, R.; Mishra, R.K.; Chauhan, D.K.; Tripathi, D.K.; Sharma, S. Tolerance and Reduction of Chromium(VI) by Bacillus Sp. MNU16 Isolated from Contaminated Coal Mining Soil. Front Plant Sci 2017, 8, 778. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Li, Y.; Zhang, X.; Liu, P.; Ren, J.; Wu, G.; Zhang, Y.; Chen, Y.; Li, X. A Bacillus Subtilis Strain Can Reduce Hexavalent Chromium to Trivalent and an nfrA Gene Is Involved. International Biodeterioration & Biodegradation 2015, 97, 90–96. [Google Scholar] [CrossRef]
- Ayele, A.; Godeto, Y. Bioremediation of Chromium by Microorganisms and Its Mechanisms Related to Functional Groups. Journal of Chemistry 2021, 2021, 1–21. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Gupta, A.; Kaur, A.; Malik, D. Alleviation of Hexavalent Chromium by Using Microorganisms: Insight into the Strategies and Complications. Water Science and Technology 2019, 79, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, Y.; Zhang, C.; Zhang, D.; Yao, C.; Meng, Q.; Zhao, R.; Wei, Z. Speciation, Toxicity Mechanism and Remediation Ways of Heavy Metals during Composting: A Novel Theoretical Microbial Remediation Method Is Proposed. Journal of Environmental Management 2020, 272, 111109. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Chang, J.-S. Bioremediation of Heavy Metals Using Microalgae: Recent Advances and Mechanisms. Bioresource Technology 2020, 303, 122886. [Google Scholar] [CrossRef] [PubMed]
- Javanbakht, V.; Alavi, S.A.; Zilouei, H. Mechanisms of Heavy Metal Removal Using Microorganisms as Biosorbent. Water Sci Technol 2014, 69, 1775–1787. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, C. Biosorbents for Heavy Metals Removal and Their Future. Biotechnology Advances 2009, 27, 195–226. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Doble, M. Novel Chromium Tolerant Microorganisms: Isolation, Characterization and Their Biosorption Capacity. Ecotoxicol Environ Saf 2008, 71, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Hossan, S.; Hossain, S.; Islam, M.R.; Kabir, M.H.; Ali, S.; Islam, M.S.; Imran, K.M.; Moniruzzaman, M.; Mou, T.J.; Parvez, A.K.; et al. Bioremediation of Hexavalent Chromium by Chromium Resistant Bacteria Reduces Phytotoxicity. Int J Environ Res Public Health 2020, 17, 6013. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, Q.; Tang, Y.; Zhou, J.; Guo, H. Tannery Wastewater Treatment: Conventional and Promising Processes, an Updated 20-Year Review. J Leather Sci Eng 2022, 4, 10. [Google Scholar] [CrossRef]
- Hong, S.-H.; Bunge, J.; Jeon, S.-O.; Epstein, S.S. Predicting Microbial Species Richness. Proceedings of the National Academy of Sciences 2006, 103, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.; Sahoo, R.K.; Gaur, M.; Behera, D.U.; Sahu, A.; Das, A.; Dey, S.; Dixit, S.; Subudhi, E. Environmental Carbapenem-Resistant Acinetobacter Baumannii in Wastewater Receiving Urban River System of Eastern India: A Public Health Threat. Int. J. Environ. Sci. Technol. 2023, 20, 9901–9910. [Google Scholar] [CrossRef]
- Hu, L.; Liu, B.; Li, S.; Zhong, H.; He, Z. Study on the Oxidative Stress and Transcriptional Level in Cr(VI) and Hg(II) Reducing Strain Acinetobacter Indicus Yy-1 Isolated from Chromium-Contaminated Soil. Chemosphere 2021, 269, 128741. [Google Scholar] [CrossRef] [PubMed]
- Sevak, P.; Pushkar, B.; Mazumdar, S. Mechanistic Evaluation of Chromium Bioremediation in Acinetobacter Junii Strain B2w: A Proteomic Approach. J Environ Manage 2023, 328, 116978. [Google Scholar] [CrossRef]
- Zakaria, Z.A.; Zakaria, Z.; Surif, S.; Ahmad, W.A. Biological Detoxification of Cr(VI) Using Wood-Husk Immobilized Acinetobacter Haemolyticus. Journal of Hazardous Materials 2007, 148, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Fadhil, G.; Alhadithi, H.; E. A.Al-Razzaq Bioremediation of Polycyclic Aromatic Hydrocarbon by Acinetobacter Species Isolated from Ecological Source. Journal of Environmental Biology 2017, 38, 785–789. [Google Scholar] [CrossRef]
- Abdulmalik, A.F.; Yakasai, H.M.; Usman, S.; Muhammad, J.B.; Jagaba, A.H.; Ibrahim, S.; Babandi, A.; Shukor, M.Y. Characterization and Invitro Toxicity Assay of Bio-Reduced Hexavalent Chromium by Acinetobacter Sp. Isolated from Tannery Effluent. Case Studies in Chemical and Environmental Engineering 2023, 8, 100459. [Google Scholar] [CrossRef]
- Montes-Robledo, A.; Baena-Baldiris, D.; Baldiris-Avila, R. Reduction of Cr(VI) by Planktonic Cells and Biofilm of Acinetobacter Sp. (ADHR1) Isolated from Electroplating Wastewater. Environmental Technology & Innovation 2024, 33, 103521. [Google Scholar] [CrossRef]
- Ghosh, A.; Sah, D.; Chakraborty, M.; Rai, J.P.N. Bio-Mediated Detoxification of Heavy Metal Contaminated Soil and Phytotoxicity Reduction Using Novel Strain of Brevundimonas Vancanneytii SMA3. Heliyon 2023, 9, e22344. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Lee, K.H.; Lee, S.Y.; Im, W.-T. Brevundimonas Fluminis Sp. Nov., Isolated from a River. International Journal of Systematic and Evolutionary Microbiology 2020, 70, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Feng, Y.; Wei, L.; Zong, Z. Genome-Based Taxonomy of Brevundimonas with Reporting Brevundimonas Huaxiensis Sp. Nov. Microbiology Spectrum 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Zhao, Z.; Liang, Z. Biodegradation of Ochratoxin A and Ochratoxin B by Brevundimonas Naejangsanensis Isolated from Soil. Food Control 2022, 133, 108611. [Google Scholar] [CrossRef]
- Naqqash, T.; Imran, A.; Hameed, S.; Shahid, M.; Majeed, A.; Iqbal, J.; Hanif, M.K.; Ejaz, S.; Malik, K.A. First Report of Diazotrophic Brevundimonas Spp. as Growth Enhancer and Root Colonizer of Potato. Sci Rep 2020, 10, 12893. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Chaturvedi, P.; Chandra, R.; Kumar, S. Identification of Heavy Metals Tolerant Brevundimonas Sp. from Rhizospheric Zone of Saccharum Munja L. and Their Efficacy in in-Situ Phytoremediation. Chemosphere 2022, 295, 133823. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gong, Z.; Allinson, G.; Li, X.; Jia, C. Joint Effects of Bacterium and Biochar in Remediation of Antibiotic-Heavy Metal Contaminated Soil and Responses of Resistance Gene and Microbial Community. Chemosphere 2022, 299, 134333. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Li, M.; Su, J.; Li, Y.; Wang, Z.; Bai, Y.; Ali, E.F.; Shaheen, S.M. Brevundimonas Diminuta Isolated from Mines Polluted Soil Immobilized Cadmium (Cd2+) and Zinc (Zn2+) through Calcium Carbonate Precipitation: Microscopic and Spectroscopic Investigations. Science of The Total Environment 2022, 813, 152668. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, Y.; Feng, R.; Chen, J.; Alwathnani, H.A.; Xu, W.; Rensing, C. Characterization of Two Highly Arsenic-Resistant Caulobacteraceae Strains of Brevundimonas Nasdae: Discovery of a New Arsenic Resistance Determinant. International Journal of Molecular Sciences 2022, 23, 5619. [Google Scholar] [CrossRef]
- Soto, J.; Charles, T.C.; Lynch, M.D.J.; Larama, G.; Herrera, H.; Arriagada, C. Genome Sequence of Brevundimonas Sp., an Arsenic Resistant Soil Bacterium. Diversity 2021, 13, 344. [Google Scholar] [CrossRef]
- Huang, R.-R.; Yang, S.-R.; Zhen, C.; Ge, X.-F.; Chen, X.-K.; Wen, Z.-Q.; Li, Y.-N.; Liu, W.-Z. Genomic Molecular Signatures Determined Characterization of Mycolicibacterium Gossypii Sp. Nov., a Fast-Growing Mycobacterial Species Isolated from Cotton Field Soil. Antonie van Leeuwenhoek 2021, 114, 1735–1744. [Google Scholar] [CrossRef] [PubMed]
- Dahl, J.L.; Gatlin III, W.; Tran, P.M.; Sheik, C.S. Mycolicibacterium Nivoides Sp. Nov Isolated from a Peat Bog. Int J Syst Evol Microbiol 2021, 71, 004438. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Ahn, S.; Truong, T.C.; Kim, J.-H.; Weerawongwiwat, V.; Lee, J.-S.; Yoon, J.-H.; Sukhoom, A.; Kim, W. Description of Mycolicibacterium Arenosum Sp. Nov. Isolated from Coastal Sand on the Yellow Sea Coast. Curr Microbiol 2024, 81, 73. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Li, Z.; Huang, S.; Huang, Y.; Wang, Q.; Tao, Z.; Hu, W. Mycolicibacterium Aurantiacum Sp. Nov. and Mycolicibacterium Xanthum Sp. Nov., Two Novel Actinobacteria Isolated from Mangrove Sediments. International Journal of Systematic and Evolutionary Microbiology 2022, 72, 005595. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.-X.; Chen, X.; Wu, Y.-J.; Wang, H.-L.; Lu, C.-M.; Wang, X.-M.; Zhang, Y.; Liu, Z.-C.; He, J.-B.; Tang, S.-K.; et al. Mycolicibacterium Arseniciresistens Sp. Nov., Isolated from Lead–Zinc Mine Tailing, and Reclassification of Two Mycobacterium Species as Mycolicibacterium Palauense Comb. Nov. and Mycolicibacterium Grossiae Comb. Nov. International Journal of Systematic and Evolutionary Microbiology 2024, 74, 006221. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Lee, K.C.; Eom, M.K.; Kim, J.-S.; Kim, D.-S.; Ko, S.-H.; Kim, B.-H.; Lee, J.-S. Variibacter Gotjawalensis Gen. Nov., Sp. Nov., Isolated from Soil of a Lava Forest. Antonie van Leeuwenhoek 2014, 105, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wang, S.; Ni, W.; Rensing, C.; Xing, S. Enhanced Cd-Zn-Pb-Contaminated Soil Phytoextraction by Sedum Alfredii and the Rhizosphere Bacterial Community Structure and Function by Applying Organic Amendments. Plant Soil 2019, 444, 101–118. [Google Scholar] [CrossRef]
Figure 1.
M: GeneRuler DNA Ladder Mix (Thermo Fisher scientific) [50]. A1-A8: PCR products using archaea oligonucleotides. B1-B8: PCR products using bacteria oligonucleotides. E1-E8: PCR products using eukaryotes oligonucleotides.
Figure 1.
M: GeneRuler DNA Ladder Mix (Thermo Fisher scientific) [50]. A1-A8: PCR products using archaea oligonucleotides. B1-B8: PCR products using bacteria oligonucleotides. E1-E8: PCR products using eukaryotes oligonucleotides.
Figure 2.
Agarose gel electrophoresis at 1% of the PCR products purified from PCR reactions performed using oligonucleotides for bacteria. Band sizes in bp are indicated. M: GeneRuler DNA Ladder Mix (Thermo Fisher scientific) [48].
Figure 2.
Agarose gel electrophoresis at 1% of the PCR products purified from PCR reactions performed using oligonucleotides for bacteria. Band sizes in bp are indicated. M: GeneRuler DNA Ladder Mix (Thermo Fisher scientific) [48].
Figure 3.
Biodegradation results of the ISO20136 assay performed with four different types of leather samples. The graph shows the biodegradation % vs time of each sample. Collagen is the positive control.
Figure 3.
Biodegradation results of the ISO20136 assay performed with four different types of leather samples. The graph shows the biodegradation % vs time of each sample. Collagen is the positive control.
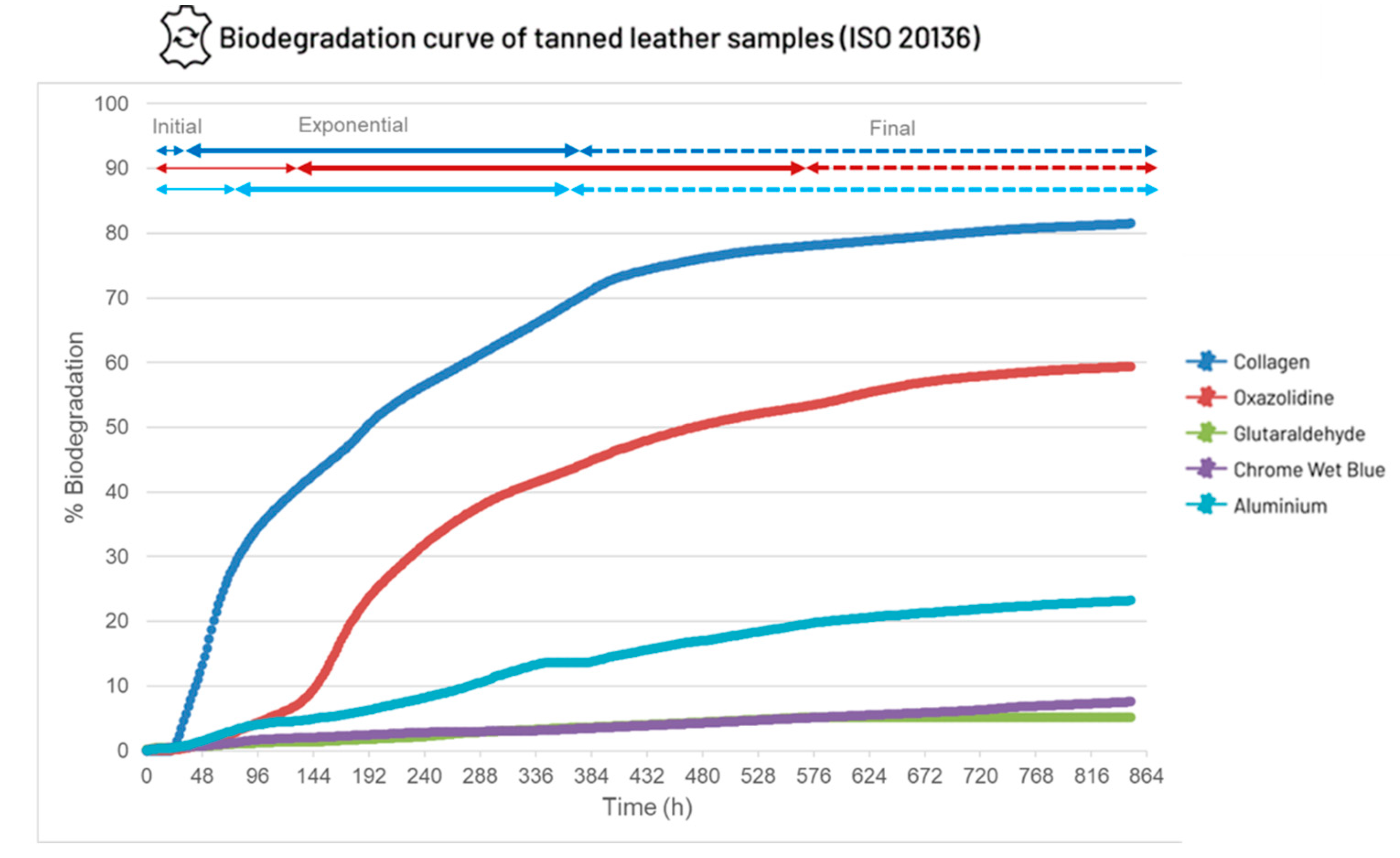
Figure 4.
Rarefaction curves of the amplified samples for bacteria detection. Samples shown M1 to M29.
Figure 4.
Rarefaction curves of the amplified samples for bacteria detection. Samples shown M1 to M29.
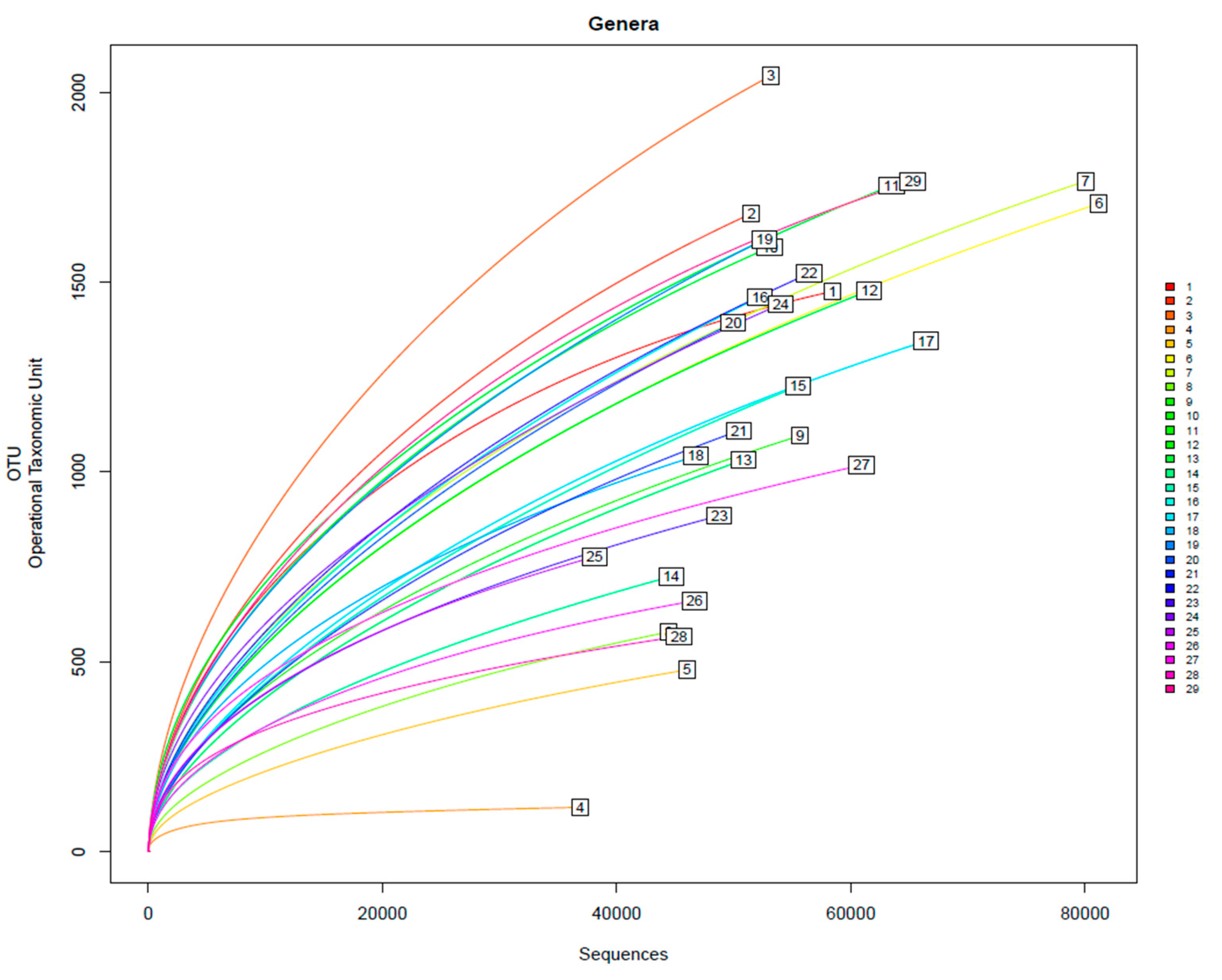
Figure 5.
Bar plot figures representing the proportions of detected bacterial genera in the studied samples.
Figure 5.
Bar plot figures representing the proportions of detected bacterial genera in the studied samples.
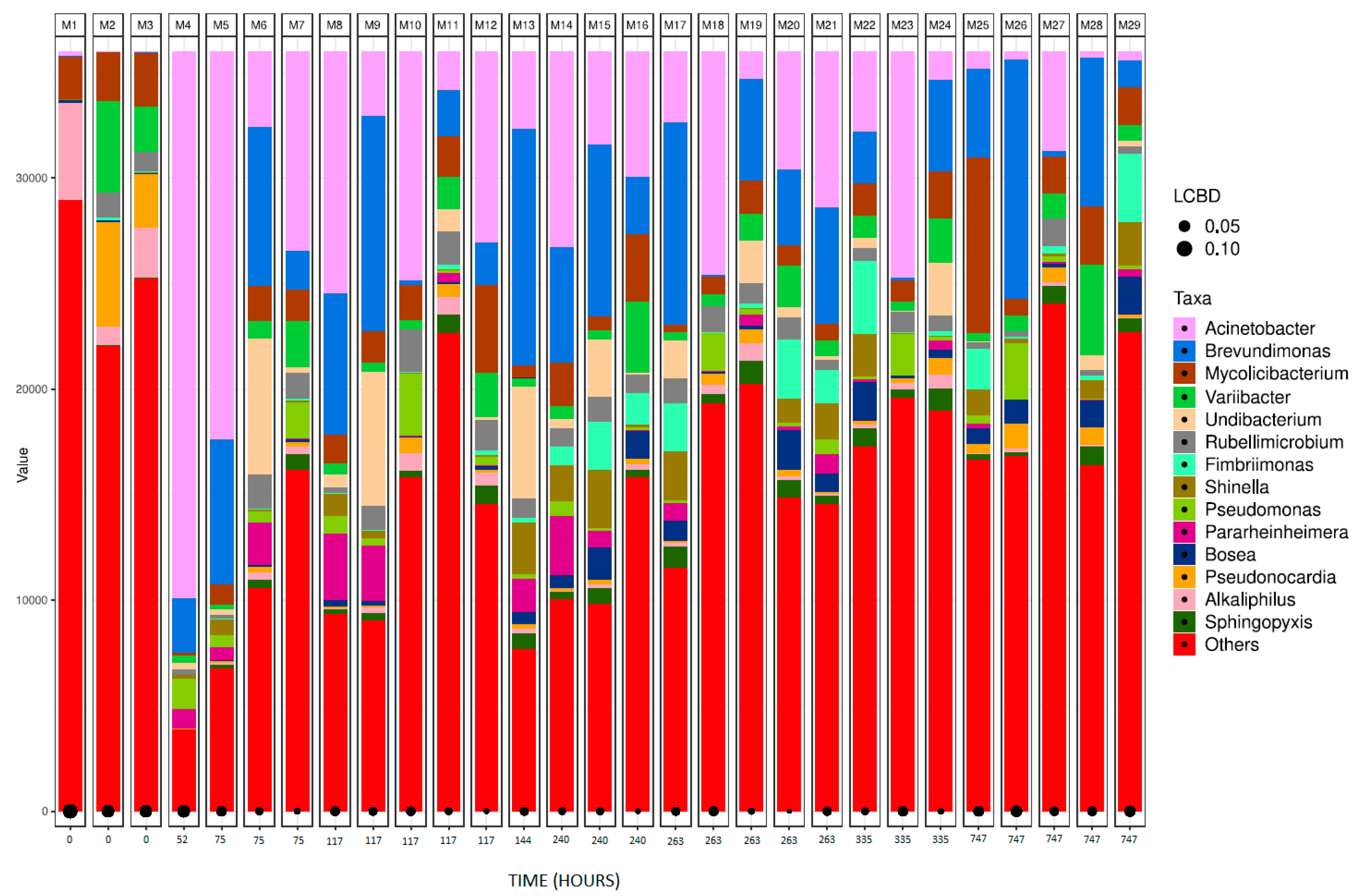
Figure 6.
Bacterial species detected for collagen samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M4, M5, M8, M14, M21 and M25). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all collagen samples.
Figure 6.
Bacterial species detected for collagen samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M4, M5, M8, M14, M21 and M25). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all collagen samples.
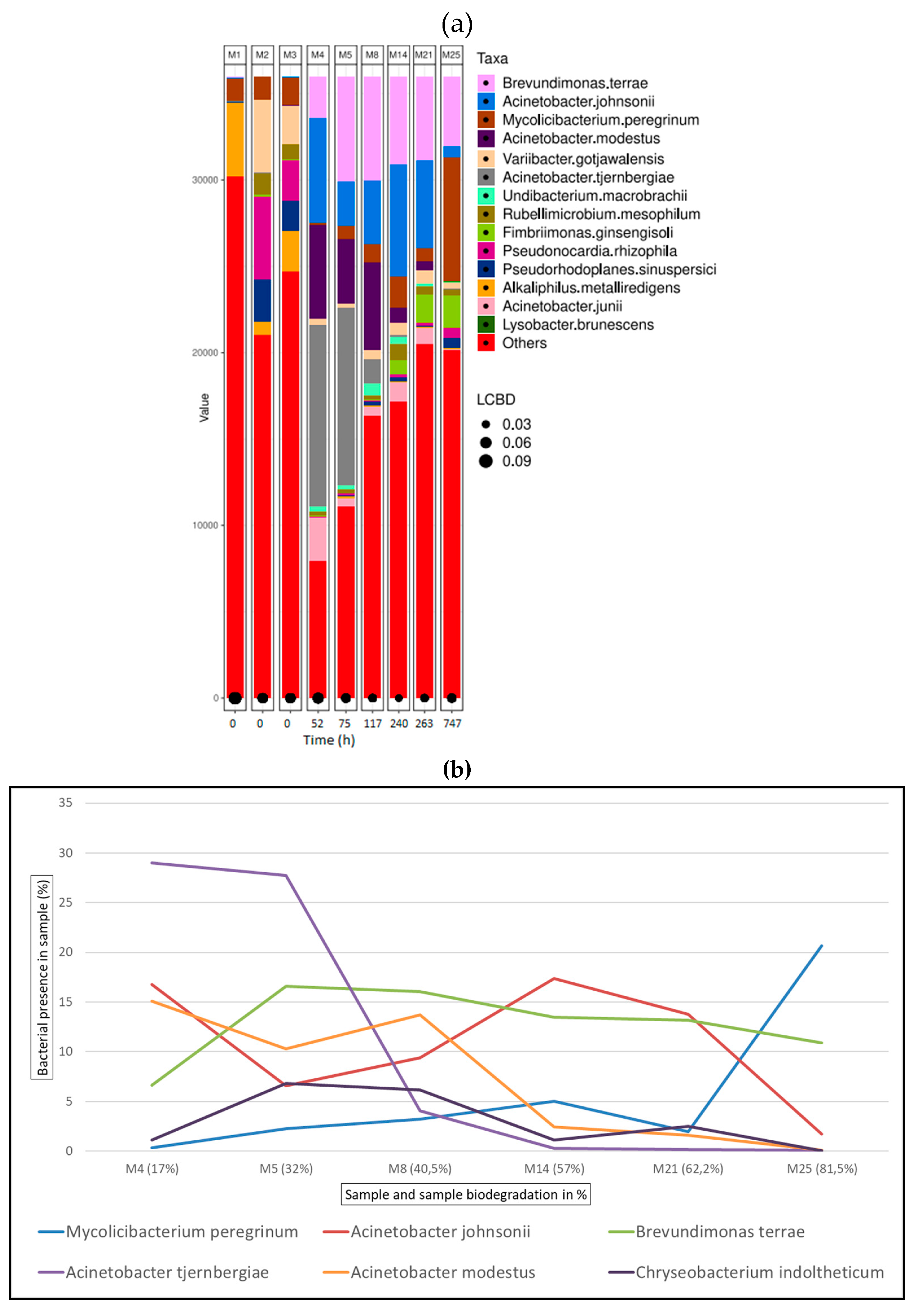
Figure 7.
Bacterial species detected for chromium samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M11, M19, M24, M28). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top 6 bacterial species found for all chromium samples.
Figure 7.
Bacterial species detected for chromium samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M11, M19, M24, M28). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top 6 bacterial species found for all chromium samples.
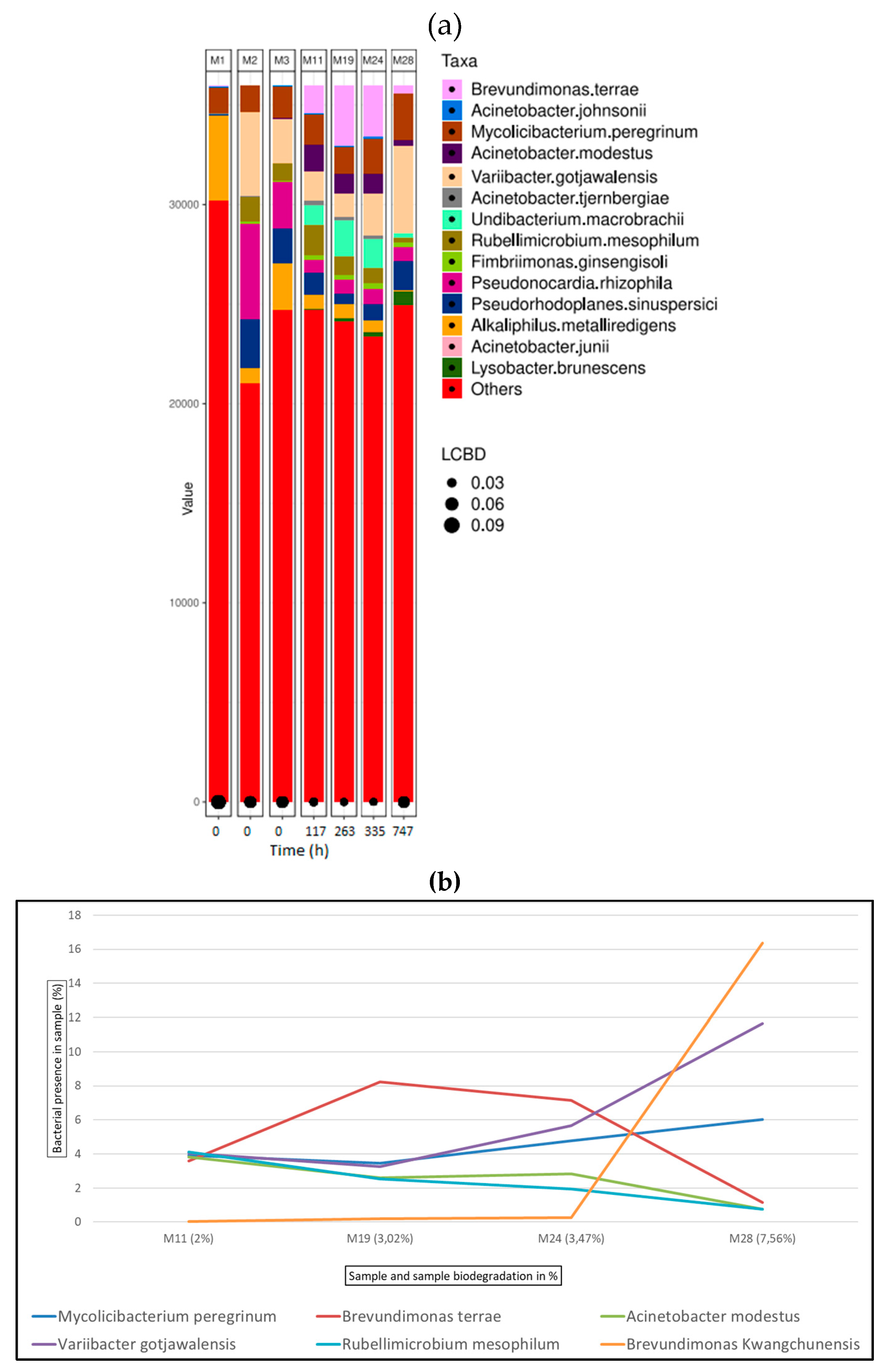
Figure 8.
Bacterial species detected for glutaraldehyde samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M10, M18, M23, M27). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all glutaraldehyde samples.
Figure 8.
Bacterial species detected for glutaraldehyde samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M10, M18, M23, M27). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all glutaraldehyde samples.
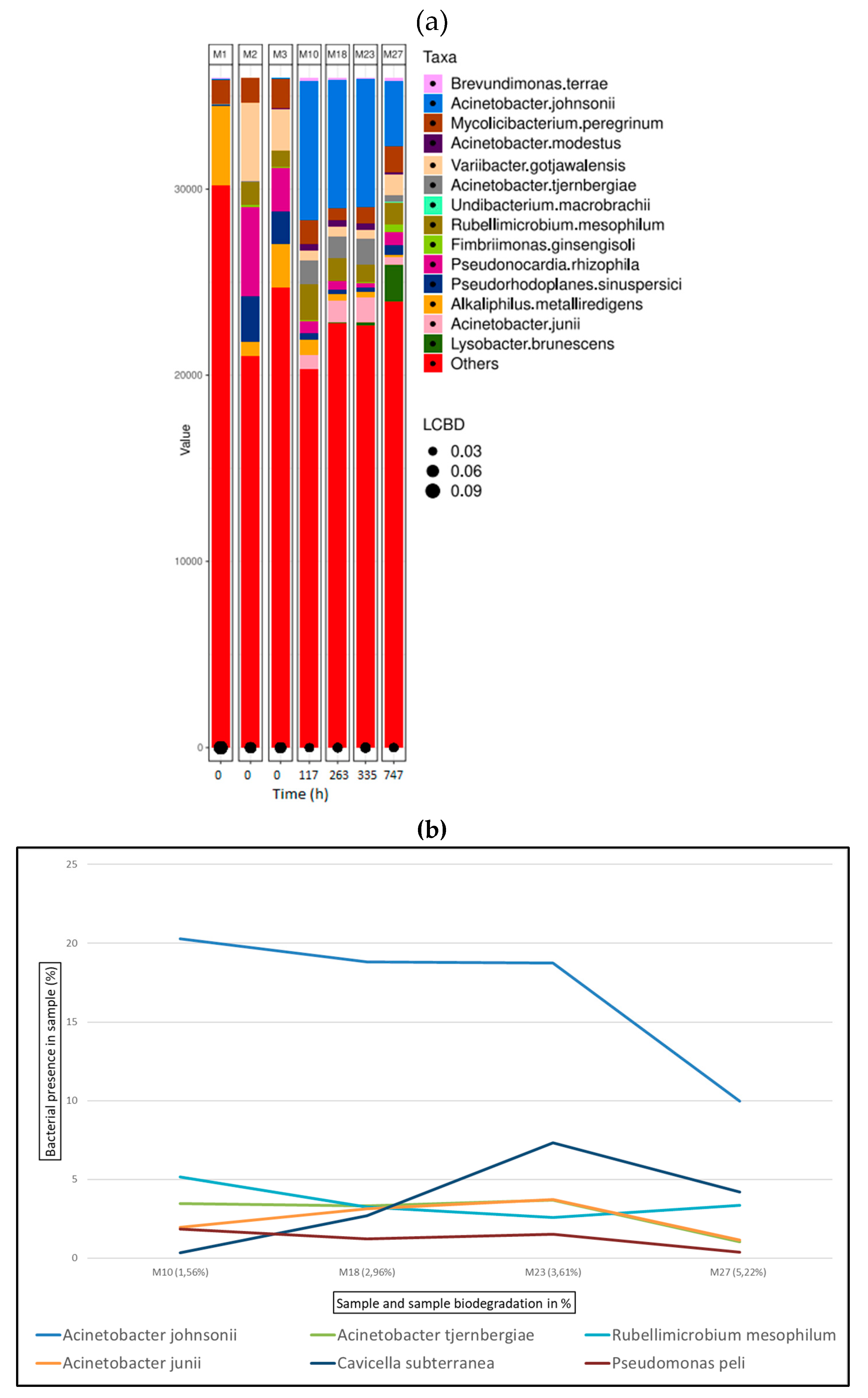
Figure 9.
Bacterial species detected for oxazolidine samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M6, M9, M13, M15, M17, M26). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all oxazolidine samples.
Figure 9.
Bacterial species detected for oxazolidine samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M6, M9, M13, M15, M17, M26). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all oxazolidine samples.
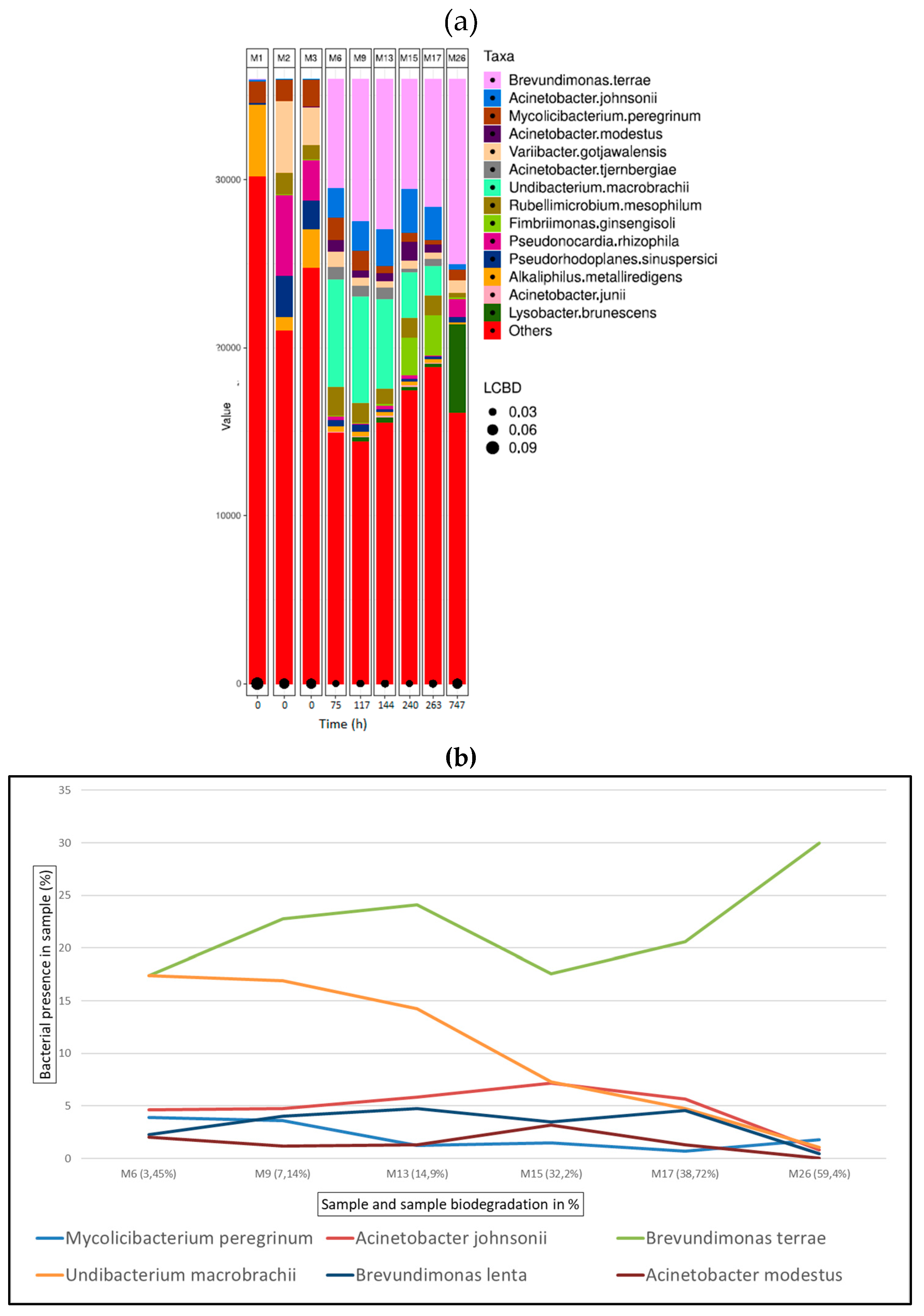
Figure 10.
Bacterial species detected for aluminium samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M7, M12, M16, M20, M22, M29). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all aluminium samples.
Figure 10.
Bacterial species detected for aluminium samples in ISO20136; (a) Bar plot figures representing the proportions of detected bacterial species in the studied samples (M7, M12, M16, M20, M22, M29). M1 being municipal residual wastewater, sample M2 was tannery wastewater, and sample M3 was a mixed inoculum (50:50). (b) Line graph representing bacterial shift as bacterial presence in sample (%) of the top six bacterial species found for all aluminium samples.
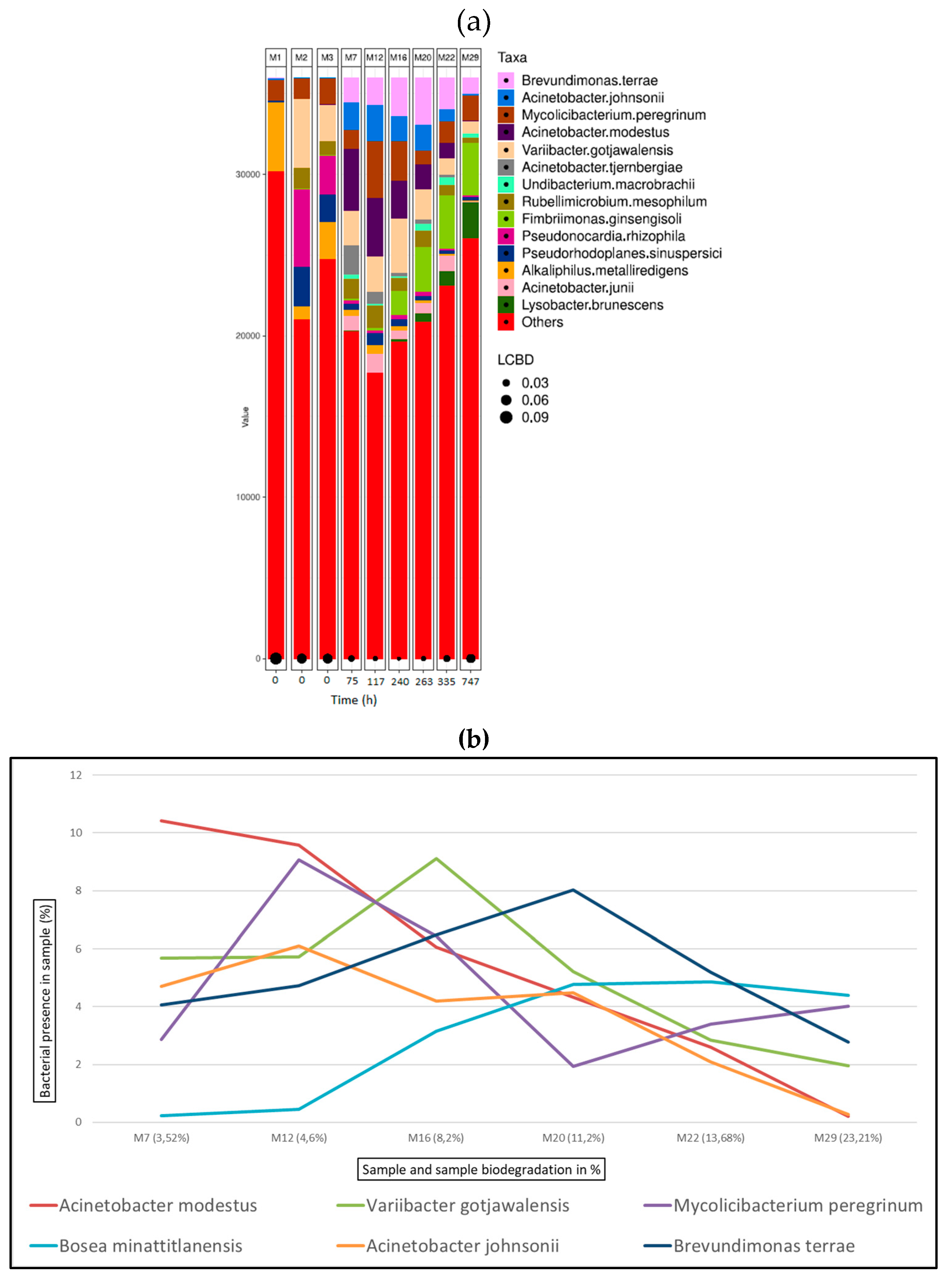

Table 1.
Leather samples (S1 to S4) and control (Pure Collagen) used in leather biodegradation assay.
Table 1.
Leather samples (S1 to S4) and control (Pure Collagen) used in leather biodegradation assay.
| Sample | Tanning agent | Carbon% | Weight (g) | Erlenmeyer Flask Ref |
|---|---|---|---|---|
| Control | None | 50.60 | 0.5047 | 2 |
| S1 | Oxazolidine | 44.76 | 0.5006 | 4 |
| S2 | Glutaraldehyde | 47.76 | 0.5002 | 7 |
| S3 | Chromium | 36.11 | 0.5012 | 10 |
| S4 | Aluminium | 41.45 | 0.5036 | 14 |
Table 2.
Extracted samples from an ongoing leather biodegradation assay.
| Sample | Time (h)1 | E. Flask Ref | Leather Sample | Volume (ml)2 | Biodegradation (%)3 |
|---|---|---|---|---|---|
| M1 | 0 | - | None | 70 | 0 |
| M2 | 0 | - | None | 50 | 0 |
| M3 | 0 | - | None | 50 | 0 |
| M4 | 52 | 2 | Control | 70 | 17 |
| M5 | 75 | 2 | Control | 50 | 32 |
| M6 | 75 | 4 | S1 | 60 | 3.45 |
| M7 | 75 | 14 | S4 | 60 | 3.52 |
| M8 | 117 | 2 | Control | 60 | 40.5 |
| M9 | 117 | 4 | S1 | 50 | 7.14 |
| M10 | 117 | 7 | S2 | 60 | 1.56 |
| M11 | 117 | 10 | S3 | 50 | 2 |
| M12 | 117 | 14 | S4 | 50 | 4.6 |
| M13 | 144 | 4 | S1 | 50 | 14.9 |
| M14 | 240 | 2 | Control | 60 | 57 |
| M15 | 240 | 4 | S1 | 60 | 32.2 |
| M16 | 240 | 14 | S4 | 60 | 8.2 |
| M17 | 263 | 4 | S1 | 60 | 38.72 |
| M18 | 263 | 7 | S2 | 60 | 2.96 |
| M19 | 263 | 10 | S3 | 50 | 3.02 |
| M20 | 263 | 14 | S4 | 50 | 11.2 |
| M21 | 263 | 7 | S2 | 50 | 62.2 |
| M22 | 335 | 14 | S4 | 50 | 13.68 |
| M23 | 335 | 7 | S2 | 50 | 3.61 |
| M24 | 335 | 10 | S3 | 50 | 3.47 |
| M25 | 747 | 2 | Control | 50 | 81.5 |
| M26 | 747 | 4 | S1 | 50 | 59.4 |
| M27 | 747 | 14 | S4 | 50 | 5.22 |
| M28 | 747 | 10 | S3 | 50 | 7.56 |
| M29 | 747 | 14 | S4 | 50 | 23.21 |
1Time (h) since time 0 of the assay. 2Volume (ml) extracted from the Erlenmeyer Flask. 3Biodegradation stage at which sample was extracted. M1: municipal wastewater, M2: tannery wastewater, M3: 50:50 mixture of M1 and M2.
Table 3.
Bacterial strains identified from tannery wastewater.
| Name | Top-hit taxon | Similarity (%) | Completeness (%) | Length (bp) |
|---|---|---|---|---|
| Species 1 | Dietzia maris | 99.48 | 94.4 | 1355 |
| Species 2 | Trichococcus pasteurii | 99.21 | 94.3 | 1396 |
| Species 3 | Corynebacterium lubricantis | 97.86 | 97.7 | 1034 |
| Species 4 | Microbacterium laevaniformans | 99.47 | 95.8 | 1370 |
| Species 5 | Bacillus safensis | 99.36 | 96.2 | 1416 |
| Species 6 | ProteiniphilumAB243818_s | 99.26 | 98 | 1419 |
| Species 7 | ProteiniphilumAB243818_s | 95.80 | 97 | 1405 |
Table 4.
Bacterial diversity for each sample according to the Shannon and Chao 1 parameters.
| Sample | Shannon | Chao 1 |
|---|---|---|
| M1 | 4.56 | 2127 |
| M2 | 4.62 | 2816 |
| M3 | 5.05 | 3383 |
| M4 | 2.70 | 266 |
| M5 | 3.07 | 1205 |
| M6 | 4.05 | 3296 |
| M7 | 4.65 | 3177 |
| M8 | 3.68 | 1344 |
| M9 | 3.79 | 2232 |
| M10 | 4.59 | 2817 |
| M11 | 5.24 | 3295 |
| M12 | 4.55 | 3053 |
| M13 | 3.77 | 2284 |
| M14 | 3.96 | 1821 |
| M15 | 4.16 | 2818 |
| M16 | 4.68 | 3068 |
| M17 | 4.10 | 2553 |
| M18 | 4.33 | 2040 |
| M19 | 5.11 | 3406 |
| M20 | 4.72 | 2972 |
| M21 | 4.16 | 2198 |
| M22 | 4.84 | 3049 |
| M23 | 4.10 | 1837 |
| M24 | 4.99 | 3031 |
| M25 | 4.01 | 1506 |
| M26 | 3.32 | 1217 |
| M27 | 4.73 | 1903 |
| M28 | 4.15 | 1105 |
| M29 | 4.94 | 2727 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



