
Submitted:
05 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
Genetic engineering has become an essential element in developing climate-resilient crops and environmental sustainable solutions to respond to the increasing need for global food security. Genome editing using CRISPR/Cas [Clustered regulatory interspaced short palindromic repeat (CRISPR)-associated protein (Cas)] technology is being applied to a variety of organisms, including plants. This technique has become popular because of its high specificity, effectiveness, and low production cost. Therefore, this technology has the potential to revolutionize agriculture and contribute to global food security. Over the past few years, increasing efforts have been seen in its application in developing higher-yielding, nutrition rich, disease resistant, stress-tolerant “crops” fruits and vegetables. Cas proteins such as Cas9, Cas12, Cas13, and Cas14, among others, have distinct architectures and have been used to create new genetic tools that improve features that are important for agriculture. The versatility of Cas has accelerated genomic analysis and facilitated the use of CRISPR/Cas to manipulate and alter nucleic acid sequences in cells of different organisms. This review provides the evolution of CRISPR technology exploring its mechanisms and contrasting it with traditional breeding and transgenic approaches to improve different stress tolerance. We have also discussed the CRISPR/Cas system and explored three Cas proteins that are currently known to exist: Cas12, Cas13, and Cas14 and their potential to generate foreign DNA free or non-transgenic crops that could be easily regulated for commercialization in most countries.
Keywords:
CRISPR-Cas system
; crop improvement
; genome engineering
; prime editing
1. Introduction
The world population is about to reach 10 billion in the coming years. This increased rate of population rise would result in increased global need for food followed by production [1]. However, the reduction of agricultural land caused by rapid urbanization and industrialization and reduction in fresh produce due to extreme weather accompanied by environmental stressors like climate change and global warming limit agriculture and food production, which may threaten food security [2]. Breeding practices have been helpful over decades in providing us improved crops. However, one of the barriers to de novo domestication through gene editing is the lack of expertise of botanists in wild-plant biology and in recent times, we are in need of novel and efficient tools that could improve crop traits in a shorter time frame. Various techniques have been developed that allow us to modify genetic sequences specifically such as, transcription activator-like effector nucleases (TALENs), RNAi (RNAi), zinc finger nucleases (ZFNs) and CRISPR/Cas9 have revolutionized crop engineering by making it easier to create crops with higher yield and other desirable characteristics. Among them, the best state-of-the-art technology is CRISPR/Cas genome editing that is gaining popularity because of its boarder application across different organisms [3]. Different genome editing methods have been well compared and the potential use of CRISPR/Cas9 technology in precision plant breeding has been reviewed in detail by Sun et al., 2016 and Chen et al., 2019 [4,5]. CRISPR–Cas system actually originated from the defense mechanism that bacteria use based on RNA, and they recognize and eliminate foreign DNA from invading plasmids and bacteriophages and are thus considered a type of bacterial “immune system” [6]. Overall, the CRISPR/Cas9 modifies genes through exact cutting of DNA, followed by the natural mechanisms of DNA repair [3,7]. The adoption of genome-edited plants by the general public and regulatory agencies is also impacted by these strategies [8].
In this review, we provide an overview of the most recent developments in CRISPR/Cas technology, the functional processes of all identified Cas proteins, and their use in contemporary horticulture and agriculture.
2. History of CRISPR/Cas System
Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) was first identified in E.coli in 1987 by a Japanese scientist, Yoshizumi Ishino, and his team, as unique repetitive DNA sequences interspersed with spacer sequences [9]. Later, such sequences have been identified in Salmonella enterica and Shigella dysenteries, two types of enterobacteria, as well as in other E. coli strains. Similar to this, when examining Mycobacterium TB strains, researchers found 36 bp repetitions scattered with unique 35–41 bp spacers. In the ensuing investigations, this CRISPR array was found in the genomes of 40% of archaea and 90% of bacteria. The biological purpose of CRISPR was a mystery for some time due to a lack of required spacer or protospacer genome sequence data. It is only in early 2000s that it was discovered that the bacteria possessing these homologous spacer sequences like bacteriophages and viruses were immune to the attack, suggesting their role in adaptive immunity in prokaryotes. Following this research, it has been found that when a bacterium is challenged by a virus, spacer sequences in CRISPR array are transcribed and guided by Cas protein to cleave the viral DNA or RNA to prevent further infection. In addition to the requirement of Cas protein, the CRISPR system also needs a short 2-6 nucleotide protospacer adjacent motif sequence (PAM) placed adjacent to the sequence identified by Cas protein (Figure 1). This ability of the bacterial adaptive immune system and the ease of selecting the target genomic location based on shorter PAM requirement has resulted in precise genome editing in various organisms for varied applications [10].
3. Nomenclature and Mechanism of CRISPR/Cas System
The CRISPR-Cas system has been divided into two main categories and several subtypes based on the complexity of the effector proteins, genomic location, pre-crRNA processing and interference. The diversity of CRISPR-Cas has increased over the past years and the evolutionary classification of this system and Cas genes has been well documented by Makarova et al., 2019 [11]. In this review, we have discussed the two main classes of CRISPR-Cas system with focus on Cas proteins that have the potential for use in agriculture.
CRISPR-Cas system is mainly classified in two main classes-I and II. In the Class 1 system, multiple effector proteins are required for the RNA-guided target cleavage, while in Class 2, only one RNA-guided endonuclease is required for the DNA sequence cleavage. Three types of CRISPR are present in the Class 1 system: I, III, and IV, while three types are present in the Class 2 system: II, V, and VI. The Cas3 signature gene, which codes for an immune protein containing a helicase to unravel DNA-DNA and RNA-DNA duplexes, is found in the type I system’s CRISPR/Cas locus. The multidomain Cas9 (CRISPR-associated protein 9) that the type II locus generates cleaves and targets dsDNA. The cas10 signature gene is present in type III CRISPR/Cas, along with a multidomain protein with a palm domain that can target ssDNA. Other Cas proteins like csf1 (large subunit, cas8-like) signature protein belongs to type IV locus and the RuvC gene, encoded by the CRISPR from Prevotella and Francisella 1 (Cpf1), C2c1 or C2c3 protein, which is carried by the type V locus, carries the Cas12 signature gene (a DNA repair-related E. coli protein) domain. This domain cleaves either dsDNA or ssDNA. Type VI has a nucleotide-binding domain (HEPN) called Cas13 (C2c2) that is responsible for cleaving ssRNA in higher eukaryotes and prokaryotes. (Table 1)
Different groups of bacteria and many Archaea use different CRISPR system to defend themselves against foreign nucleic acids, such as viruses and plasmids. The taxanomic distribution of the major classes of CRISPR-Cas system in different groups of bacteria and Archaea have been well documented to show the distribution of different Cas proteins in these organisms [12]. The CRISPR-Cas system could be explained to act in three stages: adaptation, expression and interference. Briefly, following the infection by the viruses, the bacterial host genome’s CRISPR repeat sequences are decoded as arrays, and type II CRISPR systems integrate sequences from overrunning DNA between them. The CRISPR reprise arrays are repeated to form crRNA, each containing a different sequence taken from the DNA it overlaps, called the “protospacer” sequence. This sequence makes up a part of the CRISPR reprise. In type II system, the trans-activating crRNA (tracrRNA), an alternative RNA hybridizes with each crRNA, forming a complex with the Cas9 nuclease. Only when PAMs are located conterminous to the protospacer-decoded portion of the crRNA will Cas9 be directed to stick reciprocal target-DNA sequences to make breaks in the DNA [12]. A detailed classification of these CRISPR-Cas systems and Cas proteins would provide valuable information for modifying and adapting them for various applications.
4. Cas proteins of the CRISPR System
Cas proteins are an essential component of CRISPR system and their varied abilities to edit DNA or RNA make them a potential tool in genome editing. Cas proteins, including Cas9, Cas12, Cas13, and Cas14, have been exploited for genetic engineering [6] and are discussed further below (Table 2 and Table 3)
4.1. Cas 1 and Cas 2 Proteins
Cas1-Cas2 exonuclease system is ubiquitous in all the CRISPR-Cas systems and it helps in cleaving both the target DNA and CRISPR array, a crucial step in CRISPR-Cas mechanism. Cas1 and Cas2 proteins are found in E. coli K12, forming a hexameric complex. Cas1 is an asymmetric homodimer with a central ferredoxin fold, while Cas2 is a symmetric homodimer with a central ferredoxin fold [13]. The complex produces crystals diffracting X-rays with a resolution of 2.3 Å. Both are heterozygous molecules that catalyze spacer integration via transesterification reactions. Recent studies have shown that Cas1-Cas2 proteins can recognize the PAM containing prespacer in the reannealing DNA in repair complexes, and could flexibly coordinate with other accessory proteins to process prespacers and directionality of integration. Cas1-Cas2 protects the host CRISPR system by cleaving the PAM after initial integration of the prespacer into the host CRISPR array and hence could potentially be used as an alternative to systems lacking Cas4, a well-studied protein that acts as a PAM-processing endonuclease. In spite of these advancements, it is still unclear on the upstream CRISPR substrate biogenesis and future investigations into their precise role in adaptation of CRISPR-Cas system would open avenues for their wide spread application in genome editing (Wang et al., 2023, [14].
4.2. Cas9 Protein
Cas9, formerly known as Csn1 or Csx12, is a protein connected to the Streptococcus pyogenes CRISPR mechanisms of adaptive immunity. Cas9 protein is the first Cas protein to be employed in genome editing (SpCas9) and its mechanism of action is extensively researched and applied in various organisms [15]. In natural and synthetic CRISPR/Cas systems, the SpCas9 protein functions as a DNA endonuclease and consists of 1368 amino acids in its big multifunctional domain. Cas9 endonuclease consists of six domains of which two are nuclease domains, RuvC and HNH and one is a PAM-interacting domain. The primary function of Cas9 protein is to cut three base pairs upstream of the PAM sequence in dsDNA creating double stranded breaks. Since the CRISPR/Cas9 system was created by fusing twin tracrRNA: crRNA into a single guide RNA (sgRNA), it could cut specific target dsDNA or ssDNA sequences (Figure 2). The cuts generated by CRISPR-Cas9 system could be repaired either by non-homologous end joining (NHEJ) repair or homology-directed repair (HDR). HDR usually requires a template that could be a sister chromatid or exogenously supplied DNA (gene knock-in) with homology arms to DSBs [16]. CRISPR-Cas9 is a preferred tool over other genome editing tools like ZFN and TALEN systems due to its simple design and higher effectiveness. The ability to create and use several sequence-specific gRNAs simultaneously allows for multiplex genome editing and makes it a prime trait development tool. Researchers have widely adopted this system in various fields, including microorganisms, plants, animals, insects, and human cell lines [17. Despite numerous advances, CRISPR/ Cas9 systems have numerous failings that raise numerous questions about the pitfalls involved in editing. Few major concerns of Cas9 application are on- and off-target mutations and restriction of edits to regions of high GC content because of a “G” rich PAM sequence requirement (Bernabé-Orts et al., 2019, [18]) of this protein. Indeed, the sophisticated CRISPR/ Cas systems with HDR produce unwanted mutations. Still, these off-target mutations can be eased by using a modified interpretation of Cas9 called Null-Cas9 (dCas9), which lacks the endonuclease activity [19]. Cas9 could be used for epigenome editing rather than creating irreversible genome modifications. However, further studies are necessary to improve the specificity of Cas9 function and to reduce off-target mutations [20].
4.3. Cas12 Protein
The CRISPR-Cas system, which was partially repurposed as a programmable genome-editing tool, was identified as an adaptively susceptible mechanism in prokaryotes [21]. In programmable genome editing, the Cas9 and Cas12a proteins are widely utilized. In contrast to Cas9, Cas12a protein has the ability to recognize “T” rich PAM sequences and generate staggered ends that could promote efficient site-directed integration in comparison [18]. As most of the regulatory elements are “AT” rich, Cas12a could serve as an efficient tool for engineering the epigenome modification [22]. Cas12a consists of two main components-protein/effector nuclease and a single crRNA, which is sufficient to process the crRNA unlike Cas9 that requires a tracrRNA to form a mature crRNA [23]. Three homologs of Cas12a viz., FnCas12a (from Francisella novicida), LbCas12a (from Lachnospiraceae bacterium), and AsCas12a (from Acidaminococcus sp.) share similar domain architecture and are widely utilized in plant genome editing technologies [24]). In a recent report, two other homologs of Cas12a viz., Ev1Cas12a and Hs1Cas12a have been shown to produce efficient multiplex genome editing in rice and tomato protoplasts [25]. Cas12a editing has been applied to various crops including rice, wheat, tomato, citrus, soybean and the model plant Arabidopsis thaliana [26] with varied editing efficiencies. Recently miniature variants, CRISPR/Cas12j (CasΦ) and Cas12f systems and were discovered with less than half the size of Cas9 and their orthologs were successfully tested in plants [27]. Efforts are underway to generate improved versions of Cas12a with altered PAM specificities [28; 29] and improved temperature sensitivity for its flexible application and efficient delivery into plant systems [30].
4.4. Cas13 Protein
Cas13 proteins are displayed in at least 21 bacterial genomes and it consists of two distinct HEPN (higher eukaryote and prokaryote nucleotide binding) domains and a single protein effector [31] (Figure 3). CRISPR-Cas13 is the only known system among others to target single stranded RNA and owing to this ability, Cas13 has a potent application in plants to target RNA (coding and non-coding) and silence defense responses of RNA viruses [32]. Recent studies have shown that Cas13 independent guide-induced gene silencing (GIGS) could substantially reduce the viral load in tobacco, tomato, and Arabidopsis, even in the absence of Cas13 protein offering a potential system for studying tissue or time specific expression studies which are otherwise difficult to manipulate with other CRISPR-Cas systems [33]. Cas13a orthologs vis., LwaCas13a (Cas13a from Leptotrichia wadei) and LshCas13a (Cas13a from Leptotrichia shahii) were successfully tested in plants with moderate efficiency. Cas13d subtype, which is a smaller protein than Cas13a has been discovered to perform efficiently at a broad temperature range of 24–41 °C. Modified and programmable Cas13 mutant forms (dCas13, Cas13x) were generated to effectively target specific effectors to specific RNAs in order to elicit specific modifications [34]. RNAs can be selectively targeted to use Cas13 entirely because of the innate crRNA biosynthesis. In contrast to RNA interference, genome changes induced by CRISPR/Cas13 are not restricted to focusing on cytoplasmic transcripts. In expansion, Cas13 selectively knocks off cytoplasmic mRNA transcripts, enabling quicker downregulation of expression [34]. These finding suggests a promising opportunity to deploy this system for generating multigene silencing in polypoid plants [31,35].
4.5. Cas14 Protein
Cas14, a protein in the CRISPR-Cas framework, has been characterized for its unique biochemical properties. This protein originated in extreme thermophile and shows high affinity to single stranded DNA (ssDNA) without the requirement of PAM sequence for its activity [36]. (Fig-4). The CRISPR/Cas14 system has been used to detect and genotype single nucleotide polymorphisms (SNPs) and improve resistance to ssDNA viruses in crops [37] Cas14 protein offers several advantages over traditional Cas9 protein. For example, Cas14 is incredibly small, with only 500 amino acids and hence could be delivered to any target organism more efficiently than the Cas9 protein. (Table 4 and Table 5). Moreover, Cas14 based biosensors (HARRY) are getting more focus than others like Cas12 and Cas13 proteins because of its improved sensitivity in the detection of diverse targets [38].
The major CRISPR-Cas systems which are currently in use have different pros and cons and there is always a scope to improve their efficiency and build up new approaches to use these systems for varied application across different fields.
5. Prime Editing
The double stranded breaks (DSB) caused by CRISPR-Cas9 system usually generates a complex mix of unintended in-del byproducts, translocations and chromosome fragmentations. Although attempts have been made to improve these issues [39,40], they are not accurate enough and with advancements in knowledge of the CRISPR-Cas mechanism, researchers are continuously working on improving and optimizing this gene editing technology. One such technology is named Prime editing, developed by David Liu’s group in 2019. This technology has been shown to have reduced off-targets because of its unique ability to search and replace target sequences without the need of an exogenous donor repair template [41,42]. Prime editing system mainly consists of two components: prime editor (PE-nCas9-MMLV) and PE guide RNA (pegRNA) and its mechanism of action is well studied and improved versions of prime editors have been developed for better accuracy [review by 43]. Because of this unique feature of Prime editing system, it has been successfully used to develop resistance against biotic stress by integrating a 30-nucleotide cis-regulatory element through knock-in with high efficiency in rice [44]. Various methods used to improve prime editing technology have been summarized by [45 & 43]. Improved versions of PE system have been successfully used to generate crop germplasm resources, herbicide tolerance germplasm in rice and wheat by generating specific edits and also designing mutations at existing variant sites [6,43,45]. By using modified prime editor, pCXPE03 in tomato, three genes viz., GAI, ALS2, and PDS1 were edited efficiently and produced lower frequency of off-target byproducts of 0.5%–4.9% [46]. In addition to these, improved plant GRAND pegRNA strategy from mammalian cells was successfully used to generate efficient insertion of protein tags in plants with higher insertion efficiency of 25% [25]. Although all these studies promote its wide spread application in plants, its low editing efficiency is still a concern and improved prime editors are needed to resolve these issues for a better use of this technology (Table 9).
6. Application of CRISPR in Plant Abiotic and Biotic Stress Tolerance
CRISPR-Cas is a robust tool that can be used to knock-out, knock-in or replace a gene element at a target genomic sequence to regulate the expression of a gene at the genome and epigenome level. This inexpensive and efficient tool has been employed to improve the traits of various crops.
6.1. CRISPR in Abiotic Stresses Tolerance
Abiotic stresses like drought, salinity, heavy metal and extreme temperatures greatly affect the plant growth and development and could lead to 50% of crop losses [47]. According to a report, 20% of the aerable land is affected by salinization and the increasing percentage of Earth’s land is expected to get affected in coming years at a faster pace [48. Plants under salt stress may have negative effects that lower yield and quality. This is due to the induction of osmotic, ionic, and secondary stress. Using CRISPR-Cas9 system, Alam et al. in 2022 [49] developed a rice variety by knocking out OsbHLH024 gene and enhancing the expression of the ion transporter genes OsHKT1-3, OsHAK7, and OsSOS1. 27 [50]. CRISPR/Cas9-induced OsRR22 gene mutation in rice increased its resistance to salt without affecting other agronomic characteristics [51]. It was also employed successfully to alter OsRAV2, and the resultant mutant exhibited increased survivability under salt stress. Furthermore, it has been reported that CRISPR-Cas9 system can significantly increase the resistance of different crops to salt stress by deleting or overexpressing the genes for example tomato SlARF4, rice OsDST [52], OsNAC041 and OsmiR535 [53], and barley HvITPK1 [54], Arabidopsis AtC/VIF1 [55] and Soybean GmMYB118 transcription factor [56]. Increasing impact of global warming has led to increased drought and temperature stress conditions and by targeting the drought response genes using CRISPR-Cas9 system enhanced drought tolerance has been achieved in various plant species. In rice improved drought tolerance was achieved by targeted mutagenesis of OsERA1 using CRISPR/Cas9 [57]. Other genes of rice viz., OsDST, SRL1 and SRL2, SAPK2 have been edited to improve the plant’s ability to have better water retention capacity, lower stomatal density, improved scavenging of reactive oxygen species and have improved drought tolerance [52,58,59]. The ospyl9 mutant, produced using CRISPR/Cas9, was shown to boost up rice yield and drought tolerance [58]. Additional studies involving CRISPR/Cas editing of ERF family members from rice (OsBIERF1, OsBIERF3 and OsBIERF4), maize (ARGOS8) and wheat (TaERF3) have also shown to improve drought tolerance [60,61]. Ding et al. 2020 suggest that cold stress, encompassing temperatures lower than 20°C and 0°C, obstructs growth and plant development and significantly restricts plant geographical distribution and agricultural output [62]. Low temperatures directly decrease plants’ metabolic response, causing osmotic stress, oxidative stress, and other types of stress. CRISPR/Cas9 editing was used to create rice with pyl1/4/6 triple knockdown. The mutant showed less germination before harvest, a higher yield, and a higher tolerance to temperature than the natural variety. Genes involved in ABA signaling and cell membrane biosynthesis have been targeted in rice using CRISPR-Cas9 system (CBF1, OsANN3, SAPK2, and OsMYB30) and have been found to prevent electrolyte leakage, improve relative electrical conductivity and improved the drought tolerance efficiency by nearly 63% [63 & 60]. Other major abiotic stresses like high temperature and heavy metal stress have also been addressed successfully using CRISPR gene editing in various crops like tomato (BZR1, AGL6 for heat stress tolerance), maize (TGMS5 for heat tolerance) and rice (OsNRAMP1 and OsNRAMP5-Cadmium tolerance, OsARM1-arsenic tolerance, OsHAK1- Caesium tolerance) [60,64]. The list of crops edited using CRISPR-Cas system for improvement of abiotic stress tolerance have been summarized in Table 7.
6.2. CRISPR in Biotic Stress Tolerance
Crop yield up to 20-40% worldwide are accounted to various biotic factors like viruses, fungi and bacteria [65]. CRISPR-Cas system has proven as a robust tool that could specifically knockout undesirable genes and confer plant tolerance to various diseases. Powdery mildew is one of the most destructive types of fungal diseases that crops can suffer, as it significantly reduces crop yield. Higher resilience to powdery mildew in wheat was achieved by knocking out three MILDEW-RESISTANCE LOCUS (MLO) genes which are known to be responsible for the infection, using CRISPR/Cas9. Similarly, in grapes [66] and tomatoes [67] resistance to powdery mildew was achieved through CRISPR/Cas9-mediated reduction of SlMLO and VvMLO. Furthermore, tomato powdery mildew resistance was markedly enhanced by the CRISPR/Cas9-mediated SlPMR4 mutation, although immunity was not entirely restored [68]. Several other devastating fungal diseases like Fusarium wilt and blast cause severe crop losses and lack of any resistant germplasm restrict the possibility of traditional breeding for developing resistant varieties [69]. CRISPR-Cas system was successfully used to edit genes and confer resistance to these pathogens. In rice, OsERF922 and OsSEC3A genes were mutated using CRISPR/Cas9 and the plants tested were shown to be significantly resistant to blast disease at both the seedling and tillering stages [70,71]. A method enabled by CRISPR/Cas9 technology allows for the mutation of the acetylegenase-encoding genes ACER1a and ACET1b to generate distinct resistant materials with increased resistance to bacterial and fungal diseases. Tomato susceptibility gene SlDMR6-1 mutations produced by CRISPR/Cas9 offer resistance against a variety of diseases, such as bacteria, oomycetes, and fungus [72]. Broad-spectrum resistance to bacteria and fungi was demonstrated by CRISPR/Cas9-mediated osnramp1 mutants, which showed decreased catalase (CAT) activity but elevated hydrogen peroxide (H2O2) content and superoxide dismutase (SOD) activity [73]. The list of crops and the genes edited in different crops for abiotic stress tolerance are summarized in Table 7 and Table 8.

Table 8.
CRISPR/Cas genes intended to confer tolerance against biotic stress.
| Stress | Pathogen Factor | Crop | The name of the target gene | References |
|---|---|---|---|---|
|
Insect disease |
Plant hopper | Rice (Oryza sativa) | CYTOCHROME P450 71A1 (OsCYP71A1) | [109] |
| Stem borer | Rice (Oryza sativa) | OsCYP71A1 | [109] | |
| Common cutworm | Soybean (Glycine max) | CALCIUM-DEPENDENT PROTEIN KINASE 38 (GmCDPK38) |
[110] |
|
|
Virus disease |
Rice tungro spherical virus | Rice (Oryza sativa) | eIF4G | [111] |
| Cucumber vein yellowing virus | Cucumber (Cucumis sativus) | EUKARYOTIC TRANSLATION INITIATION FACTOR 4E(eIF4E) | [112] | |
| Zucchini yellow mosaic (Cucumis virus) | Cucumber (Cucumis sativus) | eIF4E | [112] |
|
| Papaya ring spot mosaic (Cucumis virus) | Cucumber (Cucumis sativus) | eIF4E | [112] | |
| Tomato mosaic virus | Tomato (Solanum lycopersicum) |
DICER-LIKE 2b (SlDCL2b) | [113] | |
| Potato virus X | Tomato (Solanum lycopersicum) | SlDCL2a and SlDCL2b | [113] | |
|
Fungus disease |
Rice Blast | Rice (Oryza sativa) | OsERF922 SUBUNIT OF THE |
[113] |
| Rice (Oryza sativa) | EXOCYST COMPLEX 3A (OsSEC3A) | [70] |
||
| Rice (Oryza sativa) | Pi21 and Bsr-d1 | [71] ,[50] |
||
| Powdery mildew | Tomato (Solanum lycopersicum) |
MILDEW RESISTANT LOCUS O (SlMLO) | [114] | |
| Wheat (Triticum aestivum) | TaMLO-A1, TaMLO-B1 and TaMLO-D1 | [115] | ||
| Grapevine (Vitis vinifera) | VvMOL3 | [115] | ||
| Tomato (Solanum lycopersicum) | POWDERY MILDEW RESISTANCE 4 (SlPMR4) | [67] | ||
|
Late blight |
Tomato (Solanum lycopersicum) | miR482b and miR482c | [113] | |
| Gray mould | Tomato (Solanum lycopersicum) | PECTATE LYASE (SlPL) | [116] | |
|
Bacterial disease |
Bacterial blight | Rice (Oryza sativa) | SUGARS WILL EVENTUALLY BE EXPORTED TRANSPORTER 13 (OsSWEET13) | [117] |
| Citrus bacterial canker | Orange (Citrus sinensis) | LATERAL ORGAN BOUNDARY 1 (CsLOB1) | [118] |
|
| Bacterial leaf spot disease | Tomato (Solanum lycopersicum) | JASMONATE ZIM-DOMAIN 2 (SlJAZ2) | [119] |
Table 9.
List of crops modified by Prime editing technology.
| Crop Type | Monocotyledon Crops | Dicotyledon Crops |
|---|---|---|
| Prime Technologies | Examples | Examples |
|
Genomic Selection |
Rice (Oryza sativa) | Soybean (Glycine max) |
| Wheat (Triticum aestivum) | Cotton (Gossypium hirsutum) | |
| Maize (Zea mays) | Sunflower (Helianthus annuus) | |
|
CRISPR-Cas9 |
Barley (Hordeum vulgare) | Tomato (Solanum lycopersicum) |
| Sugarcane (Saccharum officinarum) | Potato (Solanum tuberosum) | |
| Sorghum (Sorghum bicolor) | Canola (Brassica napus) | |
|
Marker-Assisted Breeding |
Barley (Hordeum vulgare) | Common bean (Phaseolus vulgaris) |
| Sugarcane (Saccharum officinarum) | Pea (Pisum sativum) | |
|
High-Throughput Sequencing (NGS) |
Maize (Zea mays) | Soybean (Glycine max) |
| Rice (Oryza sativa) | Cotton (Gossypium hirsutum) | |
|
Biotechnology |
Sugarcane (Saccharum officinarum) | Cotton (Gossypium hirsutum) |
| Maize (Zea mays) | Soybean (Glycine max) | |
| Rice (Oryza sativa) | Canola (Brassica napus) | |
|
Precision Agriculture and Data Analytics |
Wheat (Triticum aestivum) | Sunflower (Helianthus annuus) |
| Rice (Oryza sativa) | Tomato (Solanum lycopersicum) | |
|
Traditional Breeding Methods |
Barley (Hordeum vulgare) | Pea (Pisum sativum) |
| Sugarcane (Saccharum officinarum) | Potato (Solanum tuberosum) | |
|
Grafting and Hybridization |
Bamboo (Various species) | Apple (Malus domestica) |
| Banana (Musa spp.) | Grape (Vitis vinifera) | |
|
Organic and Sustainable Farming Practices |
Bamboo (Various species) | Common bean (Phaseolus vulgaris) |
| Rice (Oryza sativa) | Pea (Pisum sativum) | |
|
Seed Enhancement Technologies |
Maize (Zea mays) | Cotton (Gossypium hirsutum) |
| Rice (Oryza sativa) | Soybean (Glycine max) |
7. Crop Improvement, Possible Risks and Ethical Concerns of CRISPR-Cas Based Genome Editing
The applications of genetic modification technologies based on CRISPR are applicable in many domains, such as agricultural enhancement and plant functional genomics [8]. Because of its ease of application, CRISPCas9 technology is easily accessible and relatively inexpensive to use. Public sector institutions, including the Consultative Group for International Agricultural Research (CGIAR) are using this technology to help the smallholder farmers to improve crops of less importance to the profit-driven private sectors and the current genome editing projects conducted by CGIAR to improve several crop traits are detailed by Pixley et al., 2022[74]. Genome editing is being applied to more than 40 crops to improve food and feed quality or stress tolerance in more than 25 countries but only six genome edited crops for different traits have been approved for commercialization to date [75]. Although the CRISPR-Cas system has a great potential for use, many countries are unsure about the regulations to be used for growing these genome edited crops [76]. Other major risk factors include generation of off-target mutations [77] and possible break down of natural reproductive barriers that prevent some mutations from occurring in nature [45]. However, much efforts are made to improve computational and bioinformatic tools to minimize off-target edits and to better understand the nature and frequency of these non-target mutations generated using CRISPR technology. Moreover, it is also important to consider that natural mutations occur during every generational advance during traditional plant breeding and the frequency of mutations generated through chemical or radiation are 1000 times higher than the natural mutation frequency and the non-target mutations frequency generated by current genome editing tools [45,78,79]. Hence it is important to assess the risk of using the CRISPR based technology over other traditional methods of plant breeding.
The remarkable potential that CRISPR-Cas9 technology unlocks also raises important ethical and regulatory issues. The act of modifying the genetic code of live beings gives rise to important inquiries on the limits of genetic engineering and the possible consequences of modifying inherent biological functions. The moral implications of using CRISPR-Cas9 for germline editing, in which genetic modifications that are inherited by offspring can be passed down to them, have long been a topic of discussion. The complexities of germline modifications require careful thought in order to avoid unexpected outcomes and guarantee the highest level of responsibility in its implementation. Moreover, the regulatory environment surrounding CRISPR-Cas9 technology is constantly changing, with various nations and authorities taking distinct stances on its supervision. It becomes essential to establish clear rules and regulations governing the use of CRISPR-Cas9 in order to ensure its ethical use, responsible execution, and steadfast dedication to environmental and human safety.
8. Conclusion and Future Perspectives
Nowadays, CRISPR/Cas9 technology is being used to improve the different traits such as abiotic stress tolerance, disease resistance, quality and yield of both the monocots and dicots. Crop genomic sequences can be modified to give higher yields by diversifying the approaches used to characterize the activities of individual genes. The Cas proteins has boosted the basic, therapeutics and diagnostic research. Because these CRISPR/Cas systems are inexpensive and simple to use, many researchers are using them to investigate the functions of different organisms’ genes. Furthermore, identifying the unexplained evolution of Cas masteries, which persist in a variety of microscopic organisms or archaea, would transform a variety of fields, such as the diagnosis of new illnesses, medicines, agriculture, breeding, and so forth. If these proteins’ advanced genome-editing potential is fully explored, it may ignite an untapped CRISPR “fever” that might lead to the widespread adoption of compelling and revolutionary CRISPR technologies in the near future.
Author Contributions
NT, SP, DSM, RKS conceived the manuscript. SM, SN and SP wrote the manuscript with the inputs from NT, DSM and RKS.
Data Availability Statement
The findings of this study are available from the corresponding author.
Acknowledgements
The author thanks to Centurion University of Technology and Management, Bhubaneswar, Odisha, India for providing the necessary support for the completion of this manuscript.
Conflicts of Interest
There is no conflict of Interest
References
- Clarke, J. L., & Zhang, P. (2013). Plant biotechnology for food security and bioeconomy. Plant molecular biology, 83(1-2), 1–3. [CrossRef]
- Gill, S. S., & Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant physiology and biochemistry: PPB, 48(12), 909–930. [CrossRef]
- Akram, F., Sahreen, S., Aamir, F., Haq, I. U., Malik, K., Imtiaz, M., Naseem, W., Nasir, N., & Waheed, H. M. (2023). An Insight into Modern Targeted Genome-Editing Technologies with a Special Focus on CRISPR/Cas9 and its Applications. Molecular biotechnology, 65(2), 227–242. [CrossRef]
- Sun, Y., Li, J., & Xia, L. (2016). Precise Genome Modification via Sequence-Specific Nucleases-Mediated Gene Targeting for Crop Improvement. Frontiers in plant science, 7, 1928. [CrossRef]
- Chen K, Wang Y, Zhang R, Zhang H, Gao C (2019b) CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu Rev Plant Biol 70:667–697. [CrossRef]
- Xu, Y., & Li, Z. (2020). CRISPR-Cas systems: Overview, innovations and applications in human disease research and gene therapy. Computational and structural biotechnology journal, 18, 2401–2415. [CrossRef]
- Rasheed, A., Gill, R. A., Hassan, M. U., Mahmood, A., Qari, S., Zaman, Q. U., Ilyas, M., Aamer, M., Batool, M., Li, H., & Wu, Z. (2021). A Critical Review: Recent Advancements in the Use of CRISPR/Cas9 Technology to Enhance Crops and Alleviate Global Food Crises. Current issues in molecular biology, 43(3), 1950–1976. [CrossRef]
- Yang B. (2020). Grand Challenges in Genome Editing in Plants. Frontiers in genome editing, 2, 2. [CrossRef]
- Ishino, Y., Krupovic, M., & Forterre, P. (2018). History of CRISPR-Cas from Encounter with a Mysterious Repeated Sequence to Genome Editing Technology. Journal of bacteriology, 200(7), e00580-17. [CrossRef]
- Gostimskaya I. (2022). CRISPR-Cas9: A History of Its Discovery and Ethical Considerations of Its Use in Genome Editing. Biochemistry. Biokhimiia, 87(8), 777–788. [CrossRef]
- Makarova, K.S., Wolf, Y.I., Iranzo, J. et al. Evolutionary classification of CRISPR–Cas systems: a burst of class 2 and derived variants. Nat Rev Microbiol 18, 67–83 (2020). Makarova, K.S., Wolf, Y.I., Iranzo, J. et al. Evolutionary classification of CRISPR–Cas systems: a burst of class 2 and derived variants. Nat Rev Microbiol 18, 67–83 (2020). [CrossRef]
- Makarova, K., Haft, D., Barrangou, R. et al. Evolution and classification of the CRISPR–Cas systems. Nat Rev Microbiol 9, 467–477 (2011). [CrossRef]
- Kim, E., Koo, T., Park, S. et al. In vivo genome editing with a small Cas9 orthologue derived from Campylobacter jejuni. Nat Commun 8, 14500 (2017). [CrossRef]
- Wang, J.Y., Tuck, O.T., Skopintsev, P. et al. Genome expansion by a CRISPR trimmer-integrase. Nature 618, 855–861 (2023). [CrossRef]
- Liu, Z., Dong, H., Cui, Y., Cong, L., & Zhang, D. (2020). Application of different types of CRISPR/Cas-based systems in bacteria. Microbial cell factories, 19(1), 172. [CrossRef]
- Dangquan Zhang, Zhiyong Zhang, Turgay Unver, Baohong Zhang, CRISPR/Cas: A powerful tool for gene function study and crop improvement, Journal of Advanced Research, Volume 29, 2021, Pages 207-221. [CrossRef]
- Ceasar, S. A., Maharajan, T., Hillary, V. E., & Ajeesh Krishna, T. P. (2022). Insights to improve the plant nutrient transport by CRISPR/Cas system. Biotechnology advances, 59, 107963. [CrossRef]
- Bernabé-Orts, J. M., Casas-Rodrigo, I., Minguet, E. G., Landolfi, V., Garcia-Carpintero, V., Gianoglio, S., Vázquez-Vilar, M., Granell, A., & Orzaez, D. (2019). Assessment of Cas12a-mediated gene editing efficiency in plants. Plant biotechnology journal, 17(10), 1971–1984. [CrossRef]
- Rodríguez-Rodríguez, D. R., Ramírez-Solís, R., Garza-Elizondo, M. A., Garza-Rodríguez, M. L., & Barrera-Saldaña, H. A. (2019). Genome editing: A perspective on the application of CRISPR/Cas9 to study human diseases (Review). International journal of molecular medicine, 43(4), 1559–1574. [CrossRef]
- Moradpour, M., & Abdulah, S. N. A. (2020). CRISPR/dCas9 platforms in plants: strategies and applications beyond genome editing. Plant biotechnology journal, 18(1), 32–44. [CrossRef]
- Zhang, B., Ye, Y., Ye, W., Perčulija, V., Jiang, H., Chen, Y., Li, Y., Chen, J., Lin, J., Wang, S., Chen, Q., Han, Y. S., & Ouyang, S. (2019). Two HEPN domains dictate CRISPR RNA maturation and target cleavage in Cas13d. Nature communications, 10(1), 2544. [CrossRef]
- Wolter, F., & Puchta, H. (2019). In planta gene targeting can be enhanced by the use of CRISPR/Cas12a. The Plant journal : for cell and molecular biology, 100(5), 1083–1094. [CrossRef]
- Safari, F., Zare, K., Negahdaripour, M., Barekati-Mowahed, M., & Ghasemi, Y. (2019). CRISPR Cpf1 proteins: structure, function and implications for genome editing. Cell & bioscience, 9, 36. [CrossRef]
- Zetsche, B., Gootenberg, J. S., Abudayyeh, O. O., Slaymaker, I. M., Makarova, K. S., Essletzbichler, P., Volz, S. E., Joung, J., van der Oost, J., Regev, A., Koonin, E. V., & Zhang, F. (2015). Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell, 163(3), 759–771. [CrossRef]
- Li, G., Zhang, Y., Dailey, M., & Qi, Y. (2023). Hs1Cas12a and Ev1Cas12a confer efficient genome editing in plants. Frontiers in genome editing, 5, 1251903. [CrossRef]
- Bandyopadhyay, A., Kancharla, N., Javalkote, V. S., Dasgupta, S., & Brutnell, T. P. (2020). CRISPR-Cas12a (Cpf1): A Versatile Tool in the Plant Genome Editing Tool Box for Agricultural Advancement. Frontiers in plant science, 11, 584151. [CrossRef]
- Gong, Z., Previtera, D.A., Wang, Y. et al. Geminiviral-induced genome editing using miniature CRISPR/Cas12j (CasΦ) and Cas12f variants in plants. Plant Cell Rep 43, 71 (2024). [CrossRef]
- Gao, L., Cox, D. B. T., Yan, W. X., Manteiga, J. C., Schneider, M. W., Yamano, T., Nishimasu, H., Nureki, O., Crosetto, N., & Zhang, F. (2017). Engineered Cpf1 variants with altered PAM specificities. Nature biotechnology, 35(8), 789–792. [CrossRef]
- Tóth, E., Varga, É., Kulcsár, P. I., Kocsis-Jutka, V., Krausz, S. L., Nyeste, A., Welker, Z., Huszár, K., Ligeti, Z., Tálas, A., & Welker, E. (2020). Improved LbCas12a variants with altered PAM specificities further broaden the genome targeting range of Cas12a nucleases. Nucleic acids research, 48(7), 3722–3733. [CrossRef]
- Schindele, P., & Puchta, H. (2020). Engineering CRISPR/LbCas12a for highly efficient, temperature-tolerant plant gene editing. Plant biotechnology journal, 18(5), 1118–1120. [CrossRef]
- Zhang, F., Neik, T. X., Thomas, W. J. W., & Batley, J. (2023). CRISPR-Based Genome Editing Tools: An Accelerator in Crop Breeding for a Changing Future. International journal of molecular sciences, 24(10), 8623. [CrossRef]
- Kordyś, M., Sen, R., & Warkocki, Z. (2022). Applications of the versatile CRISPR-Cas13 RNA targeting system. Wiley interdisciplinary reviews. RNA, 13(3), e1694. [CrossRef]
- Sharma, V. K., Marla, S., Zheng, W., Mishra, D., Huang, J., Zhang, W., Morris, G. P., & Cook, D. E. (2022). CRISPR guides induce gene silencing in plants in the absence of Cas. Genome biology, 23(1), 6. [CrossRef]
- Lotfi, M., & Rezaei, N. (2020). CRISPR/Cas13: A potential therapeutic option of COVID-19. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie, 131, 110738. [CrossRef]
- Dong, G., Fan, Z. CRISPR/Cas-mediated germplasm improvement and new strategies for crop protection. Crop Health 2, 2 (2024). [CrossRef]
- Harrington, L. B., Burstein, D., Chen, J. S., Paez-Espino, D., Ma, E., Witte, I. P., Cofsky, J. C., Kyrpides, N. C., Banfield, J. F., & Doudna, J. A. (2018). Programmed DNA destruction by miniature CRISPR-Cas14 enzymes. Science (New York, N.Y.), 362(6416), 839–842. [CrossRef]
- Khan, M. Z., Haider, S., Mansoor, S., & Amin, I. (2019). Targeting Plant ssDNA Viruses with Engineered Miniature CRISPR-Cas14a. Trends in biotechnology, 37(8), 800–804. [CrossRef]
- Bin Zhou, Runlin Yang, Muhammad Sohail, Xiaoxue Kong, Xing Zhang, Ninghua Fu, Bingzhi Li. CRISPR/Cas14 provides a promising platform in facile and versatile aptasensing with improved sensitivity, Talenta, Volume 254, 2023, 124120. [CrossRef]
- Richardson, C. D., Ray, G. J., DeWitt, M. A., Curie, G. L., and Corn, J. E. (2016). Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA. Nat. Biotechnol. 34 (3), 339–344. [CrossRef]
- Quadros, R. M., Miura, H., Harms, D. W., Akatsuka, H., Sato, T., Aida, T., et al. (2017). Easi-CRISPR: a robust method for one-step generation of mice carrying conditional and insertion alleles using long ssDNA donors and CRISPR ribonucleoproteins. Genome Biol. 18 (1), 92. [CrossRef]
- Anzalone, A. V., Randolph, P. B., Davis, J. R., Sousa, A. A., Koblan, L. W., Levy, J. M., et al. (2019). Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 576 (7785), 149–157. [CrossRef]
- Marzec, M., Brąszewska-Zalewska, A., and Hensel, G. (2020). Prime editing: A new way for genome editing. Trends Cell Biol. 30 (4), 257–259. [CrossRef]
- Liu Tingting, Zou Jinpeng, Yang Xi, Wang Kejian, Rao Yuchun, Wang Chun. Development and Application of Prime Editing in Plants, Rice Science, Volume 30, Issue 6, 2023, Pages 509-522. [CrossRef]
- Gupta, A., Liu, B., Chen, Q. J., & Yang, B. (2023). High-efficiency prime editing enables new strategies for broad-spectrum resistance to bacterial blight of rice. Plant biotechnology journal, 21(7), 1454–1464. [CrossRef]
- Huang, S., Weigel, D., Beachy, R. N., & Li, J. (2016). A proposed regulatory framework for genome-edited crops. Nature genetics, 48(2), 109–111. [CrossRef]
- Lu, Y., Tian, Y., Shen, R., Yao, Q., Zhong, D., Zhang, X., et al. (2021). Precise genome modification in tomato using an improved prime editing system. Plant Biotechnol. J. 19 (3), 415–417. [CrossRef]
- Liu, Z., Ma, C., Hou, L., Wu, X., Wang, D., Zhang, L., et al. (2022). Exogenous SA affects rice seed germination under salt stress by regulating Na(+)/K(+) balance and endogenous GAs and ABA homeostasis. Int. J. Mol. Sci. 23 (6), 3293. [CrossRef]
- Al Murad, Musa, Abdul Latif Khan, and Sowbiya Muneer. 2020. „Silicon in Horticultural Crops: Cross-talk, Signaling, and Tolerance Mechanism under Salinity Stress” Plants 9, no. 4: 460. [CrossRef]
- Alam, M. S., Kong, J., Tao, R., Ahmed, T., Alamin, M., Alotaibi, S. S., Abdelsalam, N. R., & Xu, J. H. (2022). CRISPR/Cas9 Mediated Knockout of the OsbHLH024 Transcription Factor Improves Salt Stress Resistance in Rice (Oryza sativa L.). Plants (Basel, Switzerland), 11(9), 1184. [CrossRef]
- Nawaz, G., Usman, B., Peng, H., Zhao, N., Yuan, R., Liu, Y., & Li, R. (2020). Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-Based Proteomic Analysis of Mutants Revealed New Insights into M. oryzae Resistance in Elite Rice Line. Genes, 11(7), 735. [CrossRef]
- Han, X., Chen, Z., Li, P., Xu, H.; Liu, K., Zha, W., Li, S., Chen, J., Yang, G., Huang, J., et al. Development of Novel Rice Germplasm for Salt-Tolerance at Seedling Stage Using CRISPR-Cas9. Sustainability 2022, 14, 2621. [CrossRef]
- Santosh Kumar, V. V., Verma, R. K., Yadav, S. K., Yadav, P., Watts, A., Rao, M. V., & Chinnusamy, V. (2020). CRISPR-Cas9 mediated genome editing of drought and salt tolerance (OsDST) gene in indica mega rice cultivar MTU1010. Physiology and molecular biology of plants : an international journal of functional plant biology, 26(6), 1099–1110. [CrossRef]
- Yue, E., Cao, H., & Liu, B. (2020). OsmiR535, a Potential Genetic Editing Target for Drought and Salinity Stress Tolerance in Oryza sativa. Plants (Basel, Switzerland), 9(10), 1337. [CrossRef]
- Vlčko, T., & Ohnoutková, L. (2020). Allelic Variants of CRISPR/Cas9 Induced Mutation in an Inositol Trisphosphate 5/6 Kinase Gene Manifest Different Phenotypes in Barley. Plants (Basel, Switzerland), 9(2), 195. [CrossRef]
- Yang B. (2020). Grand Challenges in Genome Editing in Plants. Frontiers in genome editing, 2, 2. [CrossRef]
- Du Y.T., Zhao M.J., Wang C.T., Gao Y., Wang Y.X., Liu Y.W., Ma Y.Z. Identification and Characterization of GmMYB118 Responses to Drought and Salt Stress. BMC Plant Biol. 2018;18:320. [CrossRef]
- Ogata, T., Ishizaki, T., Fujita, M., & Fujita, Y. (2020). CRISPR/Cas9-targeted mutagenesis of OsERA1 confers enhanced responses to abscisic acid and drought stress and increased primary root growth under nonstressed conditions in rice. PloS one, 15(12), e0243376. [CrossRef]
- Usman, B., Nawaz, G., Zhao, N., Liao, S., Liu, Y., & Li, R. (2020). Precise Editing of the OsPYL9 Gene by RNA-Guided Cas9 Nuclease Confers Enhanced Drought Tolerance and Grain Yield in Rice (Oryza sativa L.) by Regulating Circadian Rhythm and Abiotic Stress Responsive Proteins. International journal of molecular sciences, 21(21), 7854. [CrossRef]
- Lou D., Wang H., Liang G., Yu D. OsSAPK2 Confers Abscisic Acid Sensitivity and Tolerance to Drought Stress in Rice. Front. Plant Sci. 2017;8:993. [CrossRef]
- Erdoğan İ, Cevher-Keskin B, Bilir Ö, Hong Y, Tör M. Recent Developments in CRISPR/Cas9 Genome-Editing Technology Related to Plant Disease Resistance and Abiotic Stress Tolerance. Biology (Basel). 2023 Jul 22;12(7):1037. [CrossRef]
- Nascimento FDS, Rocha AJ, Soares JMDS, Mascarenhas MS, Ferreira MDS, Morais Lino LS, Ramos APS, Diniz LEC, Mendes TAO, Ferreira CF, Santos-Serejo JAD, Amorim EP. Gene Editing for Plant Resistance to Abiotic Factors: A Systematic Review. Plants (Basel). 2023 Jan 9;12(2):305 . [CrossRef]
- Ding, Y., Shi, Y., & Yang, S. (2020). Molecular Regulation of Plant Responses to Environmental Temperatures. Molecular plant, 13(4), 544–564. [CrossRef]
- Zeng, Y., Wen, J., Zhao, W., Wang, Q., & Huang, W. (2020). Rational Improvement of Rice Yield and Cold Tolerance by Editing the Three Genes OsPIN5b, GS3, and OsMYB30 With the CRISPR-Cas9 System. Frontiers in plant science, 10, 1663. [CrossRef]
- Li, J., Ding, J., Zhu, J., Xu, R., Gu, D., Liu, X., Liang, J., Qiu, C., Wang, H., Li, M., Qin, R., & Wei, P. (2023). Prime editing-mediated precise knockin of protein tag sequences in the rice genome. Plant communications, 4(3), 100572. [CrossRef]
- Savary S., Ficke A., Aubertot J.N., Hollier C. Crop Losses Due to Diseases and Their Implications for Global Food Production Losses and Food Security. Food Secur. 2012;4:519–537. [CrossRef]
- Wan, D. Y., Guo, Y., Cheng, Y., Hu, Y., Xiao, S., Wang, Y., & Wen, Y. Q. (2020). CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Horticulture research, 7, 116. [CrossRef]
- Nekrasov, V., Staskawicz, B., Weigel, D., Jones, J. D., & Kamoun, S. (2013). Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nature biotechnology, 31(8), 691–693. [CrossRef]
- Santillán Martínez, M.I., Bracuto, V., Koseoglou, E. et al. CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew. BMC Plant Biol 20, 284 (2020). [CrossRef]
- Zhang, M., Liu, Q., Yang, X., Xu, J., Liu, G., Yao, X., Ren, R., Xu, J., & Lou, L. (2020). CRISPR/Cas9-mediated mutagenesis of Clpsk1 in watermelon to confer resistance to Fusarium oxysporum f.sp. niveum. Plant cell reports, 39(5), 589–595. [CrossRef]
- Wang, F., Wang, C., Liu, P., Lei, C., Hao, W., Gao, Y., Liu, Y. G., & Zhao, K. (2016). Enhanced Rice Blast Resistance by CRISPR/Cas9-Targeted Mutagenesis of the ERF Transcription Factor Gene OsERF922. PloS one, 11(4), e0154027. [CrossRef]
- Ma, J., Chen, J., Wang, M., Ren, Y., Wang, S., Lei, C., Cheng, Z., & Sodmergen (2018). Disruption of OsSEC3A increases the content of salicylic acid and induces plant defense responses in rice. Journal of experimental botany, 69(5), 1051–1064. [CrossRef]
- Thomazella, D. P. T., Seong, K., Mackelprang, R., Dahlbeck, D., Geng, Y., Gill, U. S., Qi, T., Pham, J., Giuseppe, P., Lee, C. Y., Ortega, A., Cho, M. J., Hutton, S. F., & Staskawicz, B. (2021). Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. Proceedings of the National Academy of Sciences of the United States of America, 118(27), e2026152118. [CrossRef]
- Chu, C., Huang, R., Liu, L., Tang, G., Xiao, J., Yoo, H., & Yuan, M. (2022). The rice heavy-metal transporter OsNRAMP1 regulates disease resistance by modulating ROS homeostasis. Plant, cell & environment, 45(4), 1109–1126. [CrossRef]
- Pixley, K.V., Falck-Zepeda, J.B., Paarlberg, R.L. et al. Genome-edited crops for improved food security of smallholder farmers. Nat Genet 54, 364–367 (2022). [CrossRef]
- Menz, J., Modrzejewski, D., Hartung, F., Wilhelm, R., and Sprink, T. (2020). Genome edited crops touch the market: a view on the global development and regulatory environment. Front. Plant Sci. 11:586027. [CrossRef]
- Schmidt, S. M., Belisle, M., & Frommer, W. B. (2020). The evolving landscape around genome editing in agriculture: Many countries have exempted or move to exempt forms of genome editing from GMO regulation of crop plants. EMBO reports, 21(6), e50680. [CrossRef]
- Zess, E., Begemann, M. CRISPR-Cas9 and beyond: what’s next in plant genome engineering. In Vitro Cell.Dev.Biol.-Plant 57, 584–594 (2021). [CrossRef]
- Podevin, N., Davies, H. V., Hartung, F., Nogué, F., and Casacuberta, J. M. (2013). Site-directed nucleases: a paradigm shift in predictable, knowledge-based plant breeding. Trends Biotechnol. 31, 375–383. [CrossRef]
- Agricultural Chief Scientists (G20) Communique: G20 Italy 2021, 10th Meeting of Agricultural Chief Scientists (MACS) (2021); https://www.macs-g20.org/fileadmin/macs/Annual_Meetings/2021_Italy/Documents/Communique.pdf.
- Brouns, S. J., Jore, M. M., Lundgren, M., Westra, E. R., Slijkhuis, R. J., Snijders, A. P., Dickman, M. J., Makarova, K. S., Koonin, E. V., & van der Oost, J. (2008). Small CRISPR RNAs guide antiviral defense in prokaryotes. Science (New York, N.Y.), 321(5891), 960–964. [CrossRef]
- Marraffini, L. A., & Sontheimer, E. J. (2008). CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA. Science (New York, N.Y.), 322(5909), 1843–1845. [CrossRef]
- Koonin, E. V., & Makarova, K. S. (2017). Mobile Genetic Elements and Evolution of CRISPR-Cas Systems: All the Way There and Back. Genome biology and evolution, 9(10), 2812–2825. [CrossRef]
- Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., & Charpentier, E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science (New York, N.Y.), 337(6096), 816–821. [CrossRef]
- Zetsche, B., Gootenberg, J. S., Abudayyeh, O. O., Slaymaker, I. M., Makarova, K. S., Essletzbichler, P., Volz, S. E., Joung, J., van der Oost, J., Regev, A., Koonin, E. V., & Zhang, F. (2015). Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell, 163(3), 759–771. [CrossRef]
- Abudayyeh, O. O., Gootenberg, J. S., Essletzbichler, P., Han, S., Joung, J., Belanto, J. J., Verdine, V., Cox, D. B. T., Kellner, M. J., Regev, A., Lander, E. S., Voytas, D. F., Ting, A. Y., & Zhang, F. (2017). RNA targeting with CRISPR-Cas13. Nature, 550(7675), 280–284. [CrossRef]
- Mojica, F. J., Díez-Villaseñor, C., García-Martínez, J., & Soria, E. (2005). Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. Journal of molecular evolution, 60(2), 174–182. [CrossRef]
- Cong, L., Ran, F. A., Cox, D., Lin, S., Barretto, R., Habib, N., Hsu, P. D., Wu, X., Jiang, W., Marraffini, L. A., & Zhang, F. (2013). Multiplex genome engineering using CRISPR/Cas systems. Science (New York, N.Y.), 339(6121), 819–823. [CrossRef]
- Esvelt, K. M., Mali, P., Braff, J. L., Moosburner, M., Yaung, S. J., & Church, G. M. (2013). Orthogonal Cas9 proteins for RNA-guided gene regulation and editing. Nature methods, 10(11), 1116–1121. [CrossRef]
- Horvath, P., Romero, D. A., Coûté-Monvoisin, A. C., Richards, M., Deveau, H., Moineau, S., Boyaval, P., Fremaux, C., & Barrangou, R. (2008). Diversity, activity, and evolution of CRISPR loci in Streptococcus thermophilus. Journal of bacteriology, 190(4), 1401–1412. [CrossRef]
- Yang, H., Gao, P., Rajashankar, K. R., & Patel, D. J. (2016). PAM-Dependent Target DNA Recognition and Cleavage by C2c1 CRISPR-Cas Endonuclease. Cell, 167(7), 1814–1828.e12. [CrossRef]
- Yamano, T., Nishimasu, H., Zetsche, B., Hirano, H., Slaymaker, I. M., Li, Y., Fedorova, I., Nakane, T., Makarova, K. S., Koonin, E. V., Ishitani, R., Zhang, F., & Nureki, O. (2016). Crystal Structure of Cpf1 in Complex with Guide RNA and Target DNA. Cell, 165(4), 949–962. [CrossRef]
- Walton, R. T., Christie, K. A., Whittaker, M. N., & Kleinstiver, B. P. (2020). Unconstrained genome targeting with near-PAMless engineered CRISPR-Cas9 variants. Science (New York, N.Y.), 368(6488), 290–296. [CrossRef]
- Miller, S. M., Wang, T., Randolph, P. B., Arbab, M., Shen, M. W., Huang, T. P., Matuszek, Z., Newby, G. A., Rees, H. A., & Liu, D. R. (2020). Continuous evolution of SpCas9 variants compatible with non-G PAMs. Nature biotechnology, 38(4), 471–481. [CrossRef]
- Chen, J. S., Dagdas, Y. S., Kleinstiver, B. P., Welch, M. M., Sousa, A. A., Harrington, L. B., Sternberg, S. H., Joung, J. K., Yildiz, A., & Doudna, J. A. (2017). Enhanced proofreading governs CRISPR-Cas9 targeting accuracy. Nature, 550(7676), 407–410. [CrossRef]
- Casini, A., Olivieri, M., Petris, G., Montagna, C., Reginato, G., Maule, G., Lorenzin, F., Prandi, D., Romanel, A., Demichelis, F., Inga, A., & Cereseto, A. (2018). A highly specific SpCas9 variant is identified by in vivo screening in yeast. Nature biotechnology, 36(3), 265–271. [CrossRef]
- Lee, J. K., Jeong, E., Lee, J., Jung, M., Shin, E., Kim, Y. H., Lee, K., Jung, I., Kim, D., Kim, S., & Kim, J. S. (2018). Directed evolution of CRISPR-Cas9 to increase its specificity. Nature communications, 9(1), 3048. [CrossRef]
- Hu, J. H., Miller, S. M., Geurts, M. H., Tang, W., Chen, L., Sun, N., Zeina, C. M., Gao, X., Rees, H. A., Lin, Z., & Liu, D. R. (2018). Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature, 556(7699), 57–63. [CrossRef]
- Nishimasu, H., Shi, X., Ishiguro, S., Gao, L., Hirano, S., Okazaki, S., Noda, T., Abudayyeh, O. O., Gootenberg, J. S., Mori, H., Oura, S., Holmes, B., Tanaka, M., Seki, M., Hirano, H., Aburatani, H., Ishitani, R., Ikawa, M., Yachie, N., Zhang, F., … Nureki, O. (2018). Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science (New York, N.Y.), 361(6408), 1259–1262. [CrossRef]
- Slaymaker, I. M., Gao, L., Zetsche, B., Scott, D. A., Yan, W. X., & Zhang, F. (2016). Rationally engineered Cas9 nucleases with improved specificity. Science (New York, N.Y.), 351(6268), 84–88. [CrossRef]
- Kleinstiver, B. P., Pattanayak, V., Prew, M. S., Tsai, S. Q., Nguyen, N. T., Zheng, Z., & Joung, J. K. (2016). High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects. Nature, 529(7587), 490–495. [CrossRef]
- Kleinstiver, B. P., Prew, M. S., Tsai, S. Q., Nguyen, N. T., Topkar, V. V., Zheng, Z., & Joung, J. K. (2015). Broadening the targeting range of Staphylococcus aureus CRISPR-Cas9 by modifying PAM recognition. Nature biotechnology, 33(12), 1293–1298. [CrossRef]
- Hirano, H., Gootenberg, J. S., Horii, T., Abudayyeh, O. O., Kimura, M., Hsu, P. D., Nakane, T., Ishitani, R., Hatada, I., Zhang, F., Nishimasu, H., & Nureki, O. (2016). Structure and Engineering of Francisella novicida Cas9. Cell, 164(5), 950–961. [CrossRef]
- Ran, F. A., Cong, L., Yan, W. X., Scott, D. A., Gootenberg, J. S., Kriz, A. J., Zetsche, B., Shalem, O., Wu, X., Makarova, K. S., Koonin, E. V., Sharp, P. A., & Zhang, F. (2015). In vivo genome editing using Staphylococcus aureus Cas9. Nature, 520(7546), 186–191. [CrossRef]
- Hou, Z., Zhang, Y., Propson, N. E., Howden, S. E., Chu, L. F., Sontheimer, E. J., & Thomson, J. A. (2013). Efficient genome engineering in human pluripotent stem cells using Cas9 from Neisseria meningitidis. Proceedings of the National Academy of Sciences of the United States of America, 110(39), 15644–15649. [CrossRef]
- Magadán, A. H., Dupuis, M. È., Villion, M., & Moineau, S. (2012). Cleavage of phage DNA by the Streptococcus thermophilus CRISPR3-Cas system. PloS one, 7(7), e40913. [CrossRef]
- Fonfara, I., Richter, H., Bratovič, M., Le Rhun, A., & Charpentier, E. (2016). The CRISPR-associated DNA-cleaving enzyme Cpf1 also processes precursor CRISPR RNA. Nature, 532(7600), 517–521. [CrossRef]
- Li, SY., Cheng, QX., Liu, JK. et al. CRISPR-Cas12a has both cis- and trans-cleavage activities on single-stranded DNA. Cell Res 28, 491–493 (2018). [CrossRef]
- Hillary, V. E., & Ceasar, S. A. (2023). A Review on the Mechanism and Applications of CRISPR/Cas9/Cas12/Cas13/Cas14 Proteins Utilized for Genome Engineering. Molecular biotechnology, 65(3), 311–325. [CrossRef]
- Lu, H. P., Luo, T., Fu, H. W., Wang, L., Tan, Y. Y., Huang, J. Z., Wang, Q., Ye, G. Y., Gatehouse, A. M. R., Lou, Y. G., & Shu, Q. Y. (2018). Resistance of rice to insect pests mediated by suppression of serotonin biosynthesis. Nature plants, 4(6), 338–344. [CrossRef]
- Li, R., Char, S.N., Yang, B. (2019). Creating Large Chromosomal Deletions in Rice Using CRISPR/Cas9. In: Qi, Y. (eds) Plant Genome Editing with CRISPR Systems. Methods in Molecular Biology, vol 1917. Humana, New York, NY. [CrossRef]
- Macovei, A., Sevilla, N. R., Cantos, C., Jonson, G. B., Slamet-Loedin, I., Čermák, T., Voytas, D. F., Choi, I. R., & Chadha-Mohanty, P. (2018). Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant biotechnology journal, 16(11), 1918–1927. [CrossRef]
- Chandrasekaran, J., Brumin, M., Wolf, D., Leibman, D., Klap, C., Pearlsman, M., Sherman, A., Arazi, T., & Gal-On, A. (2016). Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Molecular plant pathology, 17(7), 1140–1153. [CrossRef]
- Wang, T., Deng, Z., Zhang, X., Wang, H., Wang, Y., Liu, X., Liu, S., Xu, F., Li, T., Fu, D., Zhu, B., Luo, Y., & Zhu, H. (2018). Tomato DCL2b is required for the biosynthesis of 22-nt small RNAs, the resulting secondary siRNAs, and the host defense against ToMV. Horticulture research, 5, 62. [CrossRef]
- Wan, D. Y., Guo, Y., Cheng, Y., Hu, Y., Xiao, S., Wang, Y., & Wen, Y. Q. (2020). CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Horticulture research, 7, 116. [CrossRef]
- Wang, Y., Cheng, X., Shan, Q., Zhang, Y., Liu, J., Gao, C., & Qiu, J. L. (2014). Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nature biotechnology, 32(9), 947–951. [CrossRef]
- Silva, C. J., van den Abeele, C., Ortega-Salazar, I., Papin, V., Adaskaveg, J. A., Wang, D., Casteel, C. L., Seymour, G. B., & Blanco-Ulate, B. (2021). Host susceptibility factors render ripe tomato fruit vulnerable to fungal disease despite active immune responses. Journal of experimental botany, 72(7), 2696–2709. [CrossRef]
- Zhou, J., Peng, Z., Long, J., Sosso, D., Liu, B., Eom, J. S., Huang, S., Liu, S., Vera Cruz, C., Frommer, W. B., White, F. F., & Yang, B. (2015). Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice. The Plant journal : for cell and molecular biology, 82(4), 632–643. [CrossRef]
- Peng, A., Chen, S., Lei, T., Xu, L., He, Y., Wu, L., Yao, L., & Zou, X. (2017). Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant biotechnology journal, 15(12), 1509–1519. [CrossRef]
- Ortigosa, A., Gimenez-Ibanez, S., Leonhardt, N., & Solano, R. (2019). Design of a bacterial speck resistant tomato by CRISPR/Cas9-mediated editing of SlJAZ2. Plant biotechnology journal, 17(3), 665–673. [CrossRef]
Figure 1.
The CRISPR/Cas adaptive immunity system involves three stages: CRISPR adaptation, CRISPR RNA biogenesis, and CRISPR interference. In the adaption stage, viruses trigger Cas1 and Cas2 modules, cleaving invading sequences. In the biogenesis stage, the CRISPR array is transcribed into mature crRNA molecules, forming effector complexes with Cas proteins.
Figure 1.
The CRISPR/Cas adaptive immunity system involves three stages: CRISPR adaptation, CRISPR RNA biogenesis, and CRISPR interference. In the adaption stage, viruses trigger Cas1 and Cas2 modules, cleaving invading sequences. In the biogenesis stage, the CRISPR array is transcribed into mature crRNA molecules, forming effector complexes with Cas proteins.
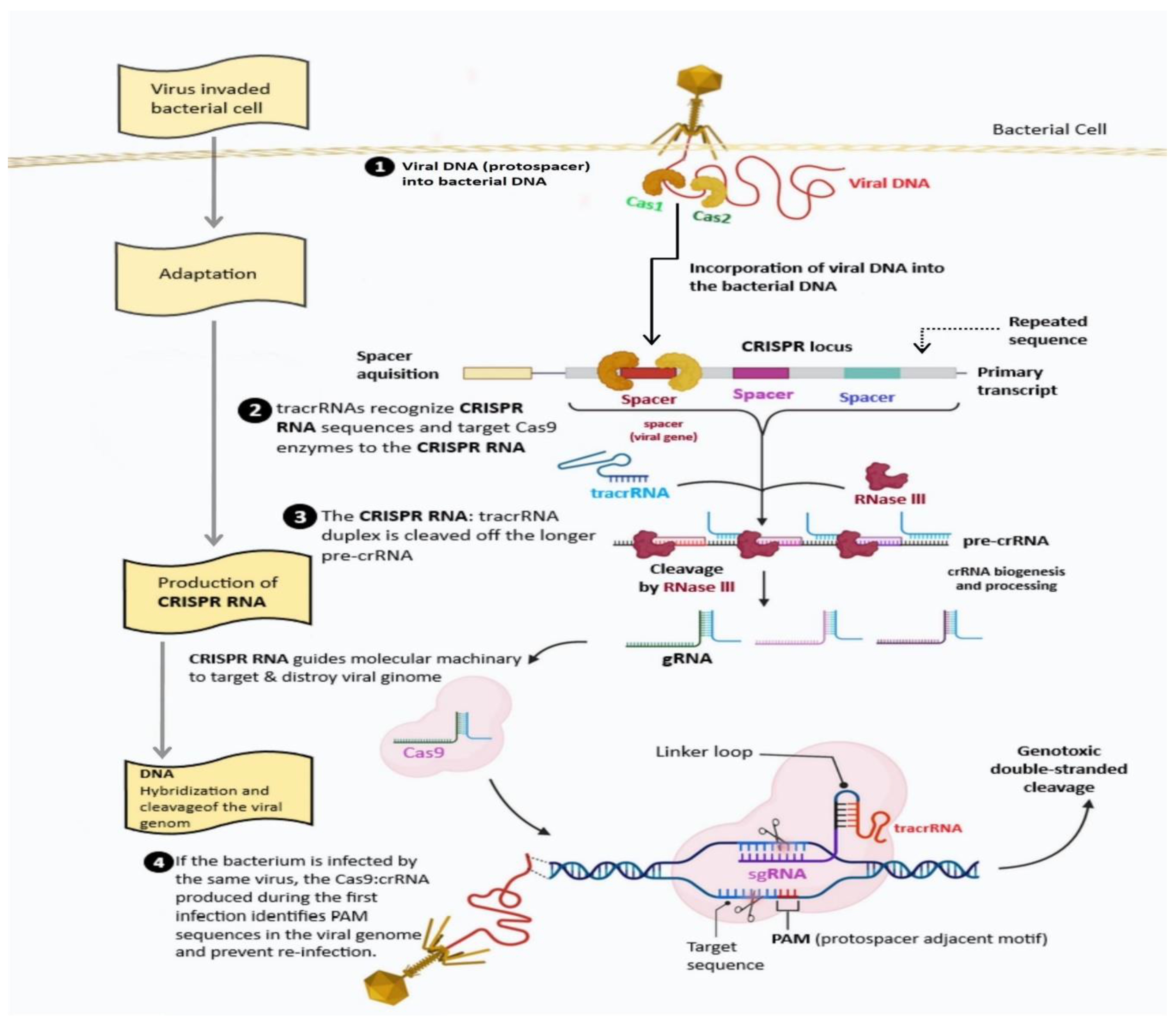
Figure 2.
The CRISPR/Cas9 mechanism involves six domains: recognition flap (REC I), arginine-rich bridging helix, PAM interaction, HNH, and RuvC. REC I is responsible for hRNA binding, while HNH and RuvC initiate cleavage. B-programmed gRNA binds to Cas9, transforming it into an active form. Cas9 generates a 3bp DSP using HNH and RuvC.
Figure 2.
The CRISPR/Cas9 mechanism involves six domains: recognition flap (REC I), arginine-rich bridging helix, PAM interaction, HNH, and RuvC. REC I is responsible for hRNA binding, while HNH and RuvC initiate cleavage. B-programmed gRNA binds to Cas9, transforming it into an active form. Cas9 generates a 3bp DSP using HNH and RuvC.
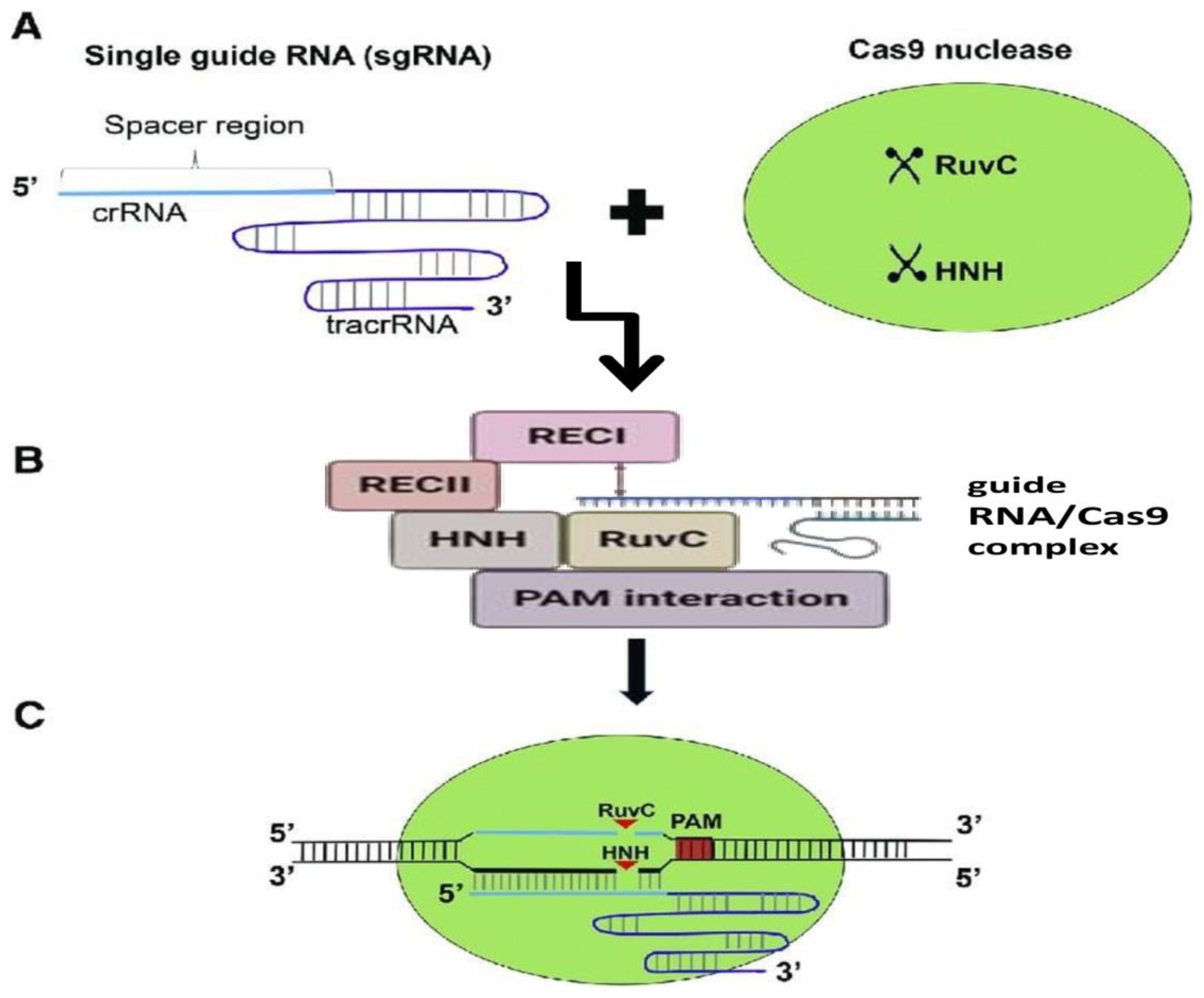
Figure 3.
The CRISPR/Cas12 mechanism involves cleaving the target region near the PAM sequence, creating an R-loop and R cycle. The CRISPR/Cas13a mechanism activates Cas13a protein, which encodes crRNA, NUC particles, and HEPN domains for target RNA. The CRISPR/Cas14 system consists of Cas14 protein, TracRNA and crRNA, Case 14, and target proteins of ssDNA. The PAM domain meets usability and application criteria for future genetic engineering.
Figure 3.
The CRISPR/Cas12 mechanism involves cleaving the target region near the PAM sequence, creating an R-loop and R cycle. The CRISPR/Cas13a mechanism activates Cas13a protein, which encodes crRNA, NUC particles, and HEPN domains for target RNA. The CRISPR/Cas14 system consists of Cas14 protein, TracRNA and crRNA, Case 14, and target proteins of ssDNA. The PAM domain meets usability and application criteria for future genetic engineering.
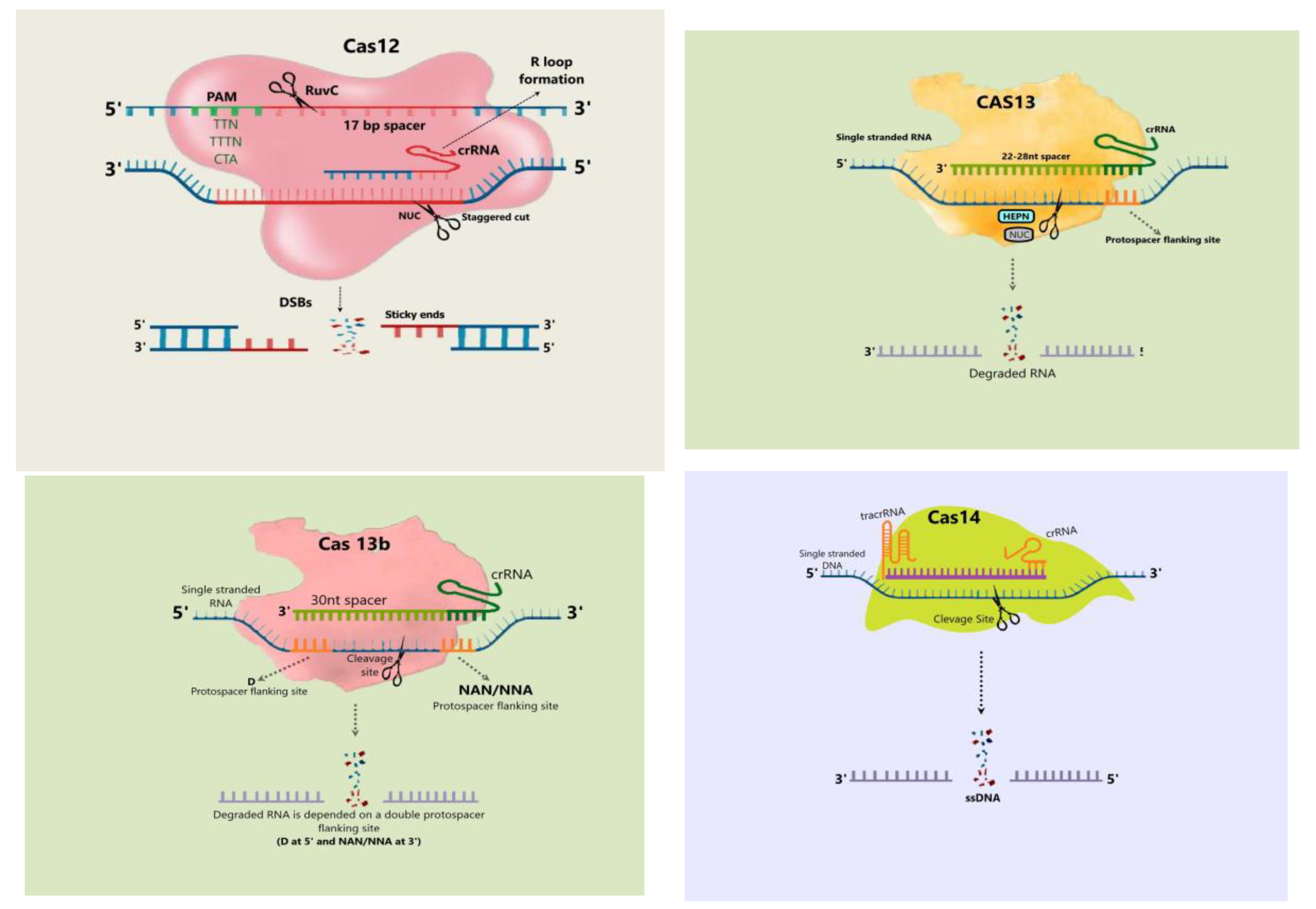
Table 1.
Classification of CRISPR-Cas system: Types of Cas proteins with their target molecule.
| CRISPR/Cas System Used |
Host Organism | Name of the Effector | Types of Protein | Target Molecule | Reference |
|---|---|---|---|---|---|
| Cas7, Cas5, Cas8, and Cas3 |
E. coli |
Cas3, Cascade, and crRNA | Cas 3 | ssDNA | [80] |
| Cas7, Cas5, and Cas1 |
S. epidermics |
Cmr/Csm, crRNA, and Cas10 | Cas 6 | ssDNA | [81] |
| Cas7, Cas5, and Csf1 | - | - | Csf1 | - | [82] |
|
Cas9 |
S. thermophilus and S. pyogenes | Cas9, tracrRNA, and crRNA | Cas9 | dsDNA | [83] |
|
Cas12 |
F. novicida |
Cpf1, crRNA and tracrRNA | Cpf1 | ssDNA and ds DNA | [84] |
|
Cas13 |
- | C2c1, and crRNA | C2c2 | ssRNA | [85] |
Table 2.
Host organisms with their cas protein and target molecules.
| Host Organism | Target Molecule | Protein Name | References |
|---|---|---|---|
| C.bacterjejuna | DNA | Cas9 | [13] |
| S. thermophilus | DNA | Cas9 | [89] |
| S. thermophilus | DNA | Cas9 | - |
| N.meningitidis | DNA | Cas9 | [88] |
| S. aureus | DNA | Cas9 | [88] |
| F. novicida | DNA | Cas9 | - |
| S. pyogenes | DNA | Cas9 | [87] |
| S. pyogenes | dsDNA | Cas9 | [86] |
| Acidaminococcus sp. | DNA | Cpf1 | [91] |
| P.Francisella | DNA | Cpf1 | [84] |
| A.bacillusacidoterrestris | DNA | C2c1 | [90] |
| Acidaminococcus sp. | DNA | Cas12a | [13] |
| L. bacterium | ssRNA | Cas13 | [85] |
| Uarchaea | ssDNA | Cas14 | [36] |
Table 3.
Major naturally occurring and genetically modified Cas enzymes used for genome editing.
| Name | Cas | CRISPR/Cas | PAM | PAM location | Resources | Reference |
|---|---|---|---|---|---|---|
| SpRY SpG Cas9-NRNH HypaCas9 evoCas9 Sniper-Cas9 xCas9 SpCas9-NG eSpCas9 SpCas9-HF SaCas9-KKH Modified SpCas9 FnCas9variant SpCas9 SaCas9 FnCas9 NmCas9 CjCas9 St1Cas9 St1Cas9 FnCas12a LbCas12a AsCas12a LsCas13# Cas14 |
Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas9 Cas12a(cpf1) Cas12a(cpf1) Cas12a(cpf1) Cas13(C2c2) Cas14 |
Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II Type II TypeVI NA |
NRN or NYN NGN NRNH NGG NGG NGG NG NG NGG NGG NNNRRT NGA or NAG YG NGG NNGRRT NGG NNNNGATT NNNNRYAC NNAGAAW NGGNG TTTN or YTN TTTV TTTV NA NA |
3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 3′ 5′ 5′ 5′ NA NA. |
Engineered SpCas9 Engineered SpCas9 Engineered SpCas9 Mutated SpCas9-HF Mutated SpCas9 Engineered SpCas9 Engineered SpCas9 Engineered SpCas9 Engineered SpCas9 Engineered SpCas9 Engineered SaCas9 Engineered SpCas9 Modified FnCas9 S. pyogenes S. aureus F. Novicida N. meningitidis C. jejuni S. thermophilus S. thermophilus F. novicida L. bacterium Acidaminococcus sp. L. shahii Archaea |
[92] [92] [93] [94] [95] [96] [97] [98] [99] [100] [101] [102] [103] [83] [103] [102] [104] [13] [105] [105] [84] [84] [106] [85] [36] |
Table 4.
Comparison of Cas9 vs Cas12 vs Cas13 vs Cas14.
| Parameter | Cas9 | Cas12 | Cas13 | Cas14 |
|---|---|---|---|---|
|
Size of Protein (Amino Acid) |
~1000-1600 |
~1300 |
~1400 |
~400-700 |
|
Target |
DNA |
DNA |
RNA |
DNA |
|
RNA |
Two RNA molecules |
Single RNA molecules |
Two RNA molecules |
Single RNA molecules |
|
Nuclease Site |
2 nuclease domains HNH and RuvC | Single nuclease RuvC-Nuc |
Target RNA domain HEPN |
DNA binding domain RuvC |
|
Pattern of cut |
Blunt |
Sticky-ended |
Degraded |
NA |
|
Spacer Size |
16-20nt |
16-25nt |
25-35nt |
NA |
|
Protospacer restriction |
PAM |
PAM |
PFS |
PAM |
|
Single guide molecular size (Nucleotides, nt) |
17-24nt |
42-44nt |
-64nt |
-140nt |
|
Non-specifically cut nucleic acids (DNA or RNA) |
DNA(SS) | DNA(SS) | RNA(SS) | DNA(SS) |
Table 5.
Mechanism and its merits of Cas protein.
| Cas Proteins | Mechanism | Applications | Merits | Demerits | Reference |
|---|---|---|---|---|---|
| Cas 9 |
|
|
|
|
[83] |
| Cas 12 |
|
|
|
|
[107] |
| Cas 13 |
|
|
|
|
[85] |
| Cas 14 |
|
|
|
|
[108] |
| Non-specifically cut nucleic acids (DNA or RNA) | DNA(SS) | DNA(SS) | RNA(SS) | DNA(SS) |
Table 6.
CRISPR/Cas genes reported to confer tolerance against different abiotic stresses.
| Stress | Cas Enzmyes | Crop | The Name of the Target Gene | References |
|---|---|---|---|---|
|
Salinity |
Cas9 | Rice (Oryza sativa) | BASIC HELIX-LOOP-HELIX 024 (OsbHLH024) | [49] |
| Rice (Oryza sativa) | RESPONSE REGULAT ORS 22 (OsRR22) | [51] | ||
| Rice (Oryza sativa) | RELATED TO ABI3/VP1 2 (OsRAV2) | [43] | ||
| Rice (Oryza sativa) | DROUGHT AND SALT TOLERANCE (OsDST) | [43] | ||
| Rice (Oryza sativa) | NAM, ATAF and CUC 041 (OsNAC041) | [70] | ||
| Rice (Oryza sativa) | OsmiR535 | [53] | ||
| Barley (Hordeum vulgare) | INOSITOLTRISPHOSPHATE 5/6 KINASES 1 (HvITPK1) | [54] | ||
| Tomato (Solanum lycopersicum) | HYBRID PROLINE-RICH PROTEIN 1 (SlHyPRP1) | [43] |
||
| Tomato (Solanum lycopersicum) | Auxin Response Factor 4 (SlARF4) | [43] | ||
|
Drought |
Rice (Oryza sativa) | ENHANCED RESPONSE TO ABA1 (OsERA1) | [57] | |
| Rice (Oryza sativa) | OsDST | [52] | ||
| Rice (Oryza sativa) | PYRABACTIN RESISTANCE-LIKE 9 (OsPYL9) | [58] | ||
| Rice (Oryza sativa) | SEMI-ROLLED LEAF 1 (SRL1) and SEMI-ROLLED LEAF 2 (SRL2) | [58] | ||
| Maize (Zea mays) | AUXIN-REGULATED GENE INVOLVED IN ORGAN SIZE 8 (ZmARGOS8) | [43] | ||
| Cas9 | Wheat (Triticum aestivum) | DEHYDRATION RESPONSIVE ELEMENT BINDING PROTEIN 2 (TaDREB2) | [13] |
|
| Wheat (Triticum aestivum) | ETHYLENE-RESPONSE FACTOR 3 (TaERF3) | [13] | ||
| Tomato (Solanum lycopersicum) | GA-INSENSITIVE DWARF1 1 (SlGID1) | [43] | ||
| Tomato (Solanum lycopersicum) | LATERAL ORGAN BOUNDARIES DOMAIN 40 (SlLBD40) | [15] | ||
| Arsenic Caesium | Cas 9 | Rice (Oryza sativa) | HIGH-AFFINITY POTASSIUM TRANSPORTER 1 (OSHAK1) | [43] |
| Rice (Oryza sativa) | ARSENITE-RESPONSIVE MYB1 (OsARM1) | [43] | ||
|
Low temperature |
Rice (Oryza sativa) | PIN-FORMED 5b (OsPIN5b) | [43] | |
| Cas9 | Rice (Oryza sativa) | GRAIN SIZE (GS3) | [43] | |
| Rice (Oryza sativa) | V-MYB AVIAN MYELOBLASTOSIS VIRAL ONCOGENE HOMOLOG 30 (OsMYB30) |
[43] | ||
|
High temperature |
Rice (Oryza sativa) | PYRABACTIN RESISTANCE-LIKE 1/4/6 (OsPYL1/4/6) | [43] | |
| Cas9 | Tomato (Solanum lycopersicum) | MITOGEN-ACTIVATED PROTEIN KINASES 3 (SlMAPK3) | [43] | |
|
Cadmium |
Rice (Oryza sativa) | NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN 5 (OsNRAMP5) |
[73] | |
| Cas9 | Rice (Oryza sativa) | LOW-AFFINITY CATION TRANSPORTER 1 (OsLCT1) | [43] | |
| Rice (Oryza sativa) | NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN 1 (OsNRAMP1) |
[73] |
Table 7.
Achievement of different targeted gene.
| Crop Species | Trait | Gene Targeted | Achievement made | Reference |
|---|---|---|---|---|
|
O. sativa |
|
|
|
[79] |
| M. balbisiana | Enhanced resistance | Mutation in downy mildew resistance 6 (DMR6) | Musa dmr6 transgenic mutants of banana showed enhanced resistance to BXW, and did not show any detrimental effect on plant growth | [80] |
| M. domestica | Disease resistance | Mutation in apple DIPM-1, DIPM-2 and DIPM-4 | Resistance to fire blast disease in non-transgenic but mutant apple lines | [81] |
| S. lycopersicum | Disease resistance | Loss of function mutation in SlDMR6-1 gene | Mutants do not have detrimental effects on growth and had multiple disease resistance P. syringae, P. capsici and Xanthomonas spp. | [82] |
| C. sinensis Osbeck | Canker resistance | Mutation and loss of function in CsWRKY22 | Mutant orange plants showed decreased susceptibility to citrus canker | [83] |
| C. sinensis Osbeck | Canker resistance | CRISPR/Cas9-targeted mutation in CsLOB1 promoter in citrus | Promoter editing of CsLOB1 alone was sufficient to enhance citrus canker resistance in citrus. | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



