
Submitted:
05 April 2024
Posted:
07 April 2024
You are already at the latest version
Abstract
Morphological and phylogenetic (45S nrDNA+RPB1 gene) analyses of three glomoid spore-producing arbuscular mycorrhizal fungi revealed the presence of two new species be-longing to the genus Glomus in the family Glomeraceae. In the field, Glomus highlandensis sp. nov. was found in a saltmarsh of the Scottish Highlands and in maritime sand dunes of the Baltic Sea in Poland, both saline environments, while G. mongioie sp. nov. originates from the Italian Alps. Phylogenetic placement analysis using environmental sequences indicated that G. highlandensis could have likely worldwide distribution while G. mongioie seems to be a rare species in the world. The molecular and phylogenetic analysis provided important insights about the presence of ribosomal variants in Glomus, with potential negative implication in phylogeny and species recognition.
Keywords:
Glomeromycota
; molecular phylogeny
; ribosomal variants
; taxonomy
; two new Glomus species
1. Introduction
Arbuscular mycorrhizal fungi (AMF) are key components of the soil microbiome. They live in close relationship with the vast majority of plant species in forest or agricultural systems, benefiting them by increasing water and nutrient absorption and mitigating biotic and abiotic stress. They also enhance soil structure and stability by stimulating root expansion and developing a hyphal network that contributes to the formation of stable macroaggregates, thereby decreasing erosion [1,2]. Finally, AMF hyphae are able to recruit specific microbial populations with multiple plant growth promoting traits such as phosphate solubilization, nitrogen fixation, plant growth regulation and biocontrol activity, increasing the functional diversity and richness of rhizosphere microorganisms [3,4].
Because of the benefits they provide, AMF have high ecological and agronomic potential, and several species, especially from the Glomeraceae family, are frequently used in commercial inoculants [5]. However, the richness of species that make up this group is still poorly understood, with several species potentially highly effective in forming symbioses yet to be discovered, as evidenced by the disparity between the number of environmental sequences available and the number of species formally described. Currently, around 360 AMF species are recognized, distributed in 49 genera and 17 families within the phylum Glomeromycota [6,7,8].
The description of Glomus macrocarpum and G. microcarpum by [9] marked the recognition of the first fungal genus within Glomeromycota, despite seven years later both species were transferred to the genus Endogone [10]. Glomus was reinstated by [11] with the revision of the family Endogonaceae, entailing the reestablishment of the original species (G. macrocarpum and G. microcarpum), the description of new Glomus species, and the transfer of other species from Endogone to Glomus. Subsequently, for at least four decades, all species with glomoid-type spores (spores produced terminally, sub-terminally or intercalary from subtending hyphae) were included in Glomus. Until 2010, Glomus encompassed nearly 50% of the described richness within Glomeromycota. However, phylogenetic analysis revealed that the genus was polyphyletic, and several Glomus species were reorganized into other taxonomic categories from genus to class level [7,12,13,14,15,16].
The phylogeny of Glomus has been constrained by the absence of molecular data from the type species, G. macrocarpum, lectotypified by [17]. [12] established a molecular epitype of G. macrocarpum and defined it as the only species in Glomus sensu stricto, while the other species were categorized as Glomus sensu lato due to their uncertain or unknown phylogenetic data. [15] described G. tetrastratosum Błaszk., Chwat & Góralska based on its phylogenetic proximity to G. macrocarpum, and other four new species (G. bareae Błaszk., Niezgoda, B.T. Goto & Kozłowska, G. ibericum A. Guillén, F.J. Serrano-Tamay, J.B. Peris & I. Arrillaga, G. atlanticum Błaszk., Niezgoda, B.T. Goto, Moreira & Magurno, and G. chinense F.X. Yu, B.T. Goto, H.Y. Feng & Yong Jun Liu) were recently described [18,19,20,21], enriching the Glomus sensu stricto clade. Morphologically, this clade is represented by glomoid species occurring in loose clusters or rarely in hypogeous or epigeous unorganized glomerocarps, forming a single spore wall with two to four layers.
During investigations of AMF assemblages associated with plants growing in a saltmarsh area, sandy dunes and mountains of Europe, we obtained single-spore cultures of three glomoid fungi, thereafter recognized as new members of the genus Glomus. The new species are here described under the name G. highlandensis and G. mongioie spp. nov., based on the morphology of the spores and the phylogenetic analysis of 45S nrDNA and the RPB1 genes.
Moreover, the analyses performed showed the presence of two ribosomal variants in G. highlandensis and G. tetrastratosum, highly divergent and with potential negative implication in phylogeny and species recognition.
2. Materials and Methods
2.1. Sampling and Single Species Pot Cultures
The field inoculum containing Glomus highlandensis was collected in the saltmarsh habitat of Beauly Firth (57°30'13.9" N 4°19'06.6" W), Scotland, UK, which is a part of Moray Firth and together with Inverness Firth form the estuarine component of the Moray Basin system. The soils are mixosaline to saline, mainly mineral, and pH is circumneutral [22]. The climate of the area is temperate oceanic, with a mean annual temperature of 9.3°C. The mean annual rainfall is relatively low (624.4 mm) due to the “rain shadow” effect caused by surrounding mountains [22,23]. The plant communities consist of Zostera angustifolia Rchb., Z. noltii Hornem., Carex recta Boott as well as Juncus gerardi Loisel., Plantago maritima L., Armeria maritima (Mill.) Willd. and Festuca rubra L. [22,24]. The field samples were collected by F. Magurno on 29th October 2018.
The second occurrence of G. highlandensis was found associated with the roots of Leymus arenarius Hochst. growing on the sand dunes of the Baltic Sea near Jastarnia, Poland (54°42'23.2452" N 18°40'7.7988 E") on the Hel Peninsula. The coastal climate of the peninsula is shaped by oceanic and continental influences, with an average annual temperature of about 9°C and mean annual rainfall of 750 mm. The soils developed on the dunes consist of unconsolidated or partly consolidated material [25]. The material was collected on 9th July 2021 by K. Bierza from yellow dune communities, representing earlier succession stages of tall-grass perennial swards with the domination of L. arenarius and Ammophila arenaria (L.) Link.
The field inoculum containing Glomus mongioie was collected on the foothills of Monte Mongioie in the Ligurian Alps, Italy (44°09'34.7” N 7°46'29.2 E"), characterized by rich mesophilous grasslands and calcitic-dolomitic soil. The plant species composition included Aquilegia atrata W.D.J.Koch, Gentiana ligustica R.Vilm. & Chopinet, Leontopodium nivale subsp. alpinum (Cass.) Greuter, Nigritella sp., Potentilla sp., Helictotrichon sedenense (DC.) Holub, Aster alpinus L., Pulsatilla alpina (L.) Delarbre, Linum alpinum L., Carex firma Host, Saxifraga caesia L., S. cernua L., Trifolium thalii Vill., Trollius europaeus L., Polygala alpestris Rchb, and Festucetum sp. The site is characterized by low temperature climate with mean annual values of 3.0 to 5.9°C and mean annual precipitation of 1001-1500 mm [26]. The samples were collected by F. Magurno on 17th August 2019.
Field-collected mixtures of rhizosphere soils and root fragments collected from the three locations were used for inoculation of pot trap cultures with Plantago lanceolata L. as host plant. After approx. five months, spores and loose clusters were isolated from the substrate and used to establish pure species pot cultures in a mixture of sterile river sand and bentonite clay (9:1, v/v). The cultures were checked at regular intervals of three months to detect the presence of mycorrhizal structures (data not shown).
2.2. Morphological Analysis
Spore clusters and spores were mounted in water, polyvinyl alcohol-lactic acid-glycerol (PVLG), and a mixture of PVLG and Melzer’s reagent (1:1, v/v). Morphological characteristics of spores and their sub-cellular structure were characterized based on at least 50‒100 spores, examined and photographed using dissecting and compound microscopes. Color names were from [27]. The terminology of spore characters follows [21,28,29,30]. Types of spore wall layers are those defined by [31,32]. Nomenclature of fungi and the authors of fungal names are from the Index Fungorum database (www.indexfungorum.org). The terms “glomerospores” and “glomerocarps'' were used for spores and fruit bodies (sporocarps) produced by AM fungi, respectively, as [33,34] proposed. Voucher specimens of the two new species were deposited in the ZT Myc (ETH Zu-rich, Switzerland; holotypes), and in the UFRN herbarium, Brazil (isotypes).
2.3. Molecular Analysis
For both species, genomic DNA was extracted from single clusters of spores with the DNeasy PowerSoil Pro kit (Qiagen), following the producer’s instructions with the modifications as follows: the first step of vortexing was replaced by crushing the spores with a micropestle inside a vial containing 100 μl of CD1 solution; the kit’s solutions volume was arranged proportionally till the first washing step; final elution was performed in 30 μl volume. Amplicons of 45S nrDNA partial genes (thereafter 45S) were obtained by nested PCR with the primer pairs SSUmAf–LSUmAr and SSUmCf–LSUmBr [35] using the Phusion Plus DNA Polymerase (Thermo Fisher Scientific) with universal annealing temperature of 60°C, according to producer’s instructions. RPB1 amplicons were obtained from Glomus highlandensis by PCR with the primers RPB1 3F alfa and RPB1 5R beta [36] targeting the region from exon 3 to 5, and from G. mongioie with the primers RPB1-4F1 in combination with RPB1-5R as in [30] targeting the region from exon 4 to 5. PCR amplicons were purified with GeneJET PCR Purification Kit (Thermo Fisher Scientific), cloned with CloneJET PCR Cloning Kit (Thermo Fisher Scientific) and sequenced at Genomed S.A. (Warsaw). Both 45S and RPB1 sequences were deposited in GenBank (accession numbers will be added after revision of the manuscript).
2.4. Phylogenetic Analyses
For the phylogenetic accommodation of the here presented new species, two datasets of 45S sequences and one dataset of RPB1 partial gene sequences were prepared, representing all the sequenced members of the genus Glomus, upon verification by blastn analysis of the affiliation of the new species to genus. Sequences of the two described Complexispora species [36] were included to represent the outgroup in the phylogenetic analysis.
The 45S datasets contained 49 (ds1) and 41(ds2) sequences respectively, overlapping the partial 18S - ITS - partial 28S regions. The datasets differed by the presence of alternative gene variant sequences, to test their influence on the stability of species clades in the phylogenetic inference. Additionally, the ds1 included 4 sequences representing both variants previously obtained from an isolate of Glomus tetrastratosum from the AMF collection of the University of Silesia in Katowice.
The RPB1 dataset contained 29 sequences, the longest spanning from exon 1 to 5, the shortest from exon 4 to 5.
The datasets were aligned separately with the online version of MAFFT 7 using the E-INS-i iterative refinement methods (http://mafft.cbrc.jp/alignment/server/). A concatenated alignment was prepared combining the 45S (ds2) and the RPB1 alignments using a custom Python script.
The two 45S and the concatenated alignments were used as input for the Bayesian inference (BI) and maximum likelihood (ML) phylogenetic inference, performed via CIPRES Science Gateway 3.1 [37]. The 45S alignments were divided into five partitions: 18S, ITS1, 5.8S, ITS2, 28S. Nine additional RPB1 partitions, representing 5 exons and four introns, were employed in the concatenated alignment. In both BI and ML analyses GTR+I+G was used as nucleotide substitution model for each partition [38]. Four Markov chains were run over one million generations in MrBayes 3.2 [39], sampling every 1,000 generations, with a burn-in at 30% sampled trees. The ML phylogenetic tree inference was performed with RAxML-NG 1.0.1 [40], using a maximum likelihood/1000 bootstrapping run, and ML estimated proportion of invariable sites and base frequencies. For each alignment, the resulting phylogenetic trees were visualized, merged and edited in TreeGraph 2 [41]. Any discrepancy in the topology between BI and ML trees was recorded. Clades were considered supported with Bayesian posterior probabilities ≥ 0.95 and ML bootstrap values ≥ 70%.
The percentages of identity intraspecies and between the new species and the closest relatives was calculated in Mothur v.1.44.3 using the function dist.seqs (calc=eachgap, countends=F) using the aligned 45S and RPB1 datasets as input.
To detect other possible occurrences of the two species, blastn was used to retrieve uncultured Glomeromycota sequences from GenBank with percentage of identity > 96% with at least one of the 45S sequences of the species of interest. The environmental sequences were aligned with the db1 and their phylogenetic placement was established by RAxML-EPA [42] followed by the placement mass accumulation in GAPPA v0.8.0 [43] of each sequence upwards the 45S ML tree used as reference, with threshold > 0.9, similarly to [44].
3. Results
3.1. Phylogenetic Analyses
Overall, 16 sequences for the 45S locus and 7 sequences for the RPB1 locus were obtained in the study. Among the 45S sequences, 11 were obtained for Glomus highlandensis (7 for the isolate “Highlands - Scotland”, 4 for the isolate “Hel-Poland”) and 5 for G. mongioie (Italian Alps). About the RPB1 locus, 5 sequences were obtained for G. highlandensis (2 for the isolate “Highlands”, 3 for the isolate “Hel”) and 2 for G. mongioie.
The visual inspection of the two 45S alignments allowed to recognize the presence of two variants, S (=short) and L (=long) among the sequences of G. highlandensis (present in both isolates) and G. tetrastratosum, differing mostly in the ITS1 motif by an insertion of ca 40-50 bp (Figure S1).
Both the phylogenies generated from the concatenated alignment (Figure 1) and the two 45S alignments (Figure 2 and Figure S2) showed identical topology. All parameters of the convergence diagnostic (potential scale reduction factor and standard deviation of split frequencies) indicated that the convergence was obtained in the BI analysis. The sequences of G. highlandensis and G. mongioie populated independent, significantly supported clades, both sister to G. macrocarpum.
The phylogenetic inference of the db1 (Figure 2) showed all 4 G. highlandensis sequences of the L variant clustering in a fully supported “long branch” clade inside the species clade that received full BI support (1.00) and moderate ML support (71%). In the consensus tree generated from the db2 alignment (without the L variant sequences) (Figure S2) and in the concatenated tree (Figure 1), the species clade ML support increased to 84% and 88%, respectively.
The G. mongioie clade received in all analysis full BI support and ML support ranging from 88-90% in the 45S phylogenies (db2 and db1 alignments, respectively) to 98% in the 45S-RPB1 phylogeny.
The L variant sequence of G. tetrastratosum clustered with the S variant sequences of the species in an almost fully supported clade (BI=1.00, ML=99%), while the S variant clade received only low ML support (63%).
Considering the two variants together, the 45S sequences of G. highlandensis displayed a high intraspecific variability, with a range of identity as 91.5-100%. The clade hosting the L variant shared 91.5-94.4% of identity with the sequences of the S variant. The S variant was considered as “conventional” since the visual inspection of the alignment showed a similar ITS1 organization with the sequences representing the other Glomus species (therefore used for species comparison). Considering the S variant, the percentage of identity between the Highlands and Hel isolates ranged between 96 and 97.9%. G. highlandensis differed from the neighbour species G. macrocarpum and G. mongioie by 95.9-97.6 and 94.5-98.3%, respectively. In G. mongioie the percentage of identity among 45S sequences was higher than 98.5%. The species differed from the neighbour species G. macrocarpum by 95.2 and 96.1%.
The percentage of identity of RPB1 sequences was higher than 99.6% and 99.3% in G. highlandensis and G. mongioie, respectively. The two species differed by 99-99.2%, and from G. macrocarpum by 99.1-99.4 (G. highlandensis) and 98.4-98.6 (G. mongioie). In G. tetrastratosum the L variant shared ca 94% of identity with the S variant sequences.
Regarding possible environmental occurrences of the two species, 34 “Uncultured Glomus/Glomeromycota” sequences were retrieved from Genbank. RAxML-EPA and GAPPA analysis assigned six sequences to G. highlandensis, while no environmental occurrences were found for G. mongioie (Figure S3; see also Ecology and distribution in Taxonomy section).
3.2. Taxonomy
Glomus highlandensis B.T. Goto, Magurno, Uszok, M.B. Queiroz, sp. nov.
Figure 3A‒G.
MycoBank No. MB ...… (will be added after reviews)
Etymology: English, highlandensis, referring to the Scottish Highlands, location where this species was originally found.
Typification: Scotland. Spores from a single-species culture established from a trap culture inoculated with a field-collected mixture of rhizospheric soil from a saltmarsh of the Beauly Firth, Scotland, UK (57°30'13.9" N 4°19'06.6" W), October 2018, F. Magurno. Holotype: slide with spores no. XXXX - will be added after reviews, collection name. Isotype: slides with spores no. XXXX - will be added after reviews, UFRN-Fungos.
Diagnosis: Differs from G. macrocarpum, the closest phylogenetic relative together with G. mongioie (Figure 1), in: (i) the spore wall and subtending hyphal wall structure, as well as (ii) nucleotide composition of sequences of the 45S nuc rDNA gene and the RPB1 gene.
Description: Glomerospores formed in soil, in loose clusters with 2‒18 spores or singly, arise blastically at tips of sporogenous hyphae directly continuous with extraradical mycorrhizal hyphae (Figure 3A‒G). Spores pale yellow (4A3) to greyish yellow (4B3); globose to subglobose; (30‒)62(‒110) µm diam; rarely ovoid; 70‒98 × 81‒110 µm; with one subtending hypha (Figure 3A‒F). Spore wall composed of four layers (Figure 3A,C‒G).
Figure 3.
A‒G. Glomus highlandensis. A‒B. Cluster with sporogenous hyphae, spores, and a spore subtending hypha (sh). A, C‒G. Spore wall layers (swl) 1‒4. A, F. Subtending hyphal wall layers (shwl) 1‒4 continuous with spore wall layers (swl) 1‒4. A, C, E. Spores in PVLG. B, D, F, G. Spores in PVLG+Melzer’s reagent. A‒G. Differential interference microscopy.
Figure 3.
A‒G. Glomus highlandensis. A‒B. Cluster with sporogenous hyphae, spores, and a spore subtending hypha (sh). A, C‒G. Spore wall layers (swl) 1‒4. A, F. Subtending hyphal wall layers (shwl) 1‒4 continuous with spore wall layers (swl) 1‒4. A, C, E. Spores in PVLG. B, D, F, G. Spores in PVLG+Melzer’s reagent. A‒G. Differential interference microscopy.
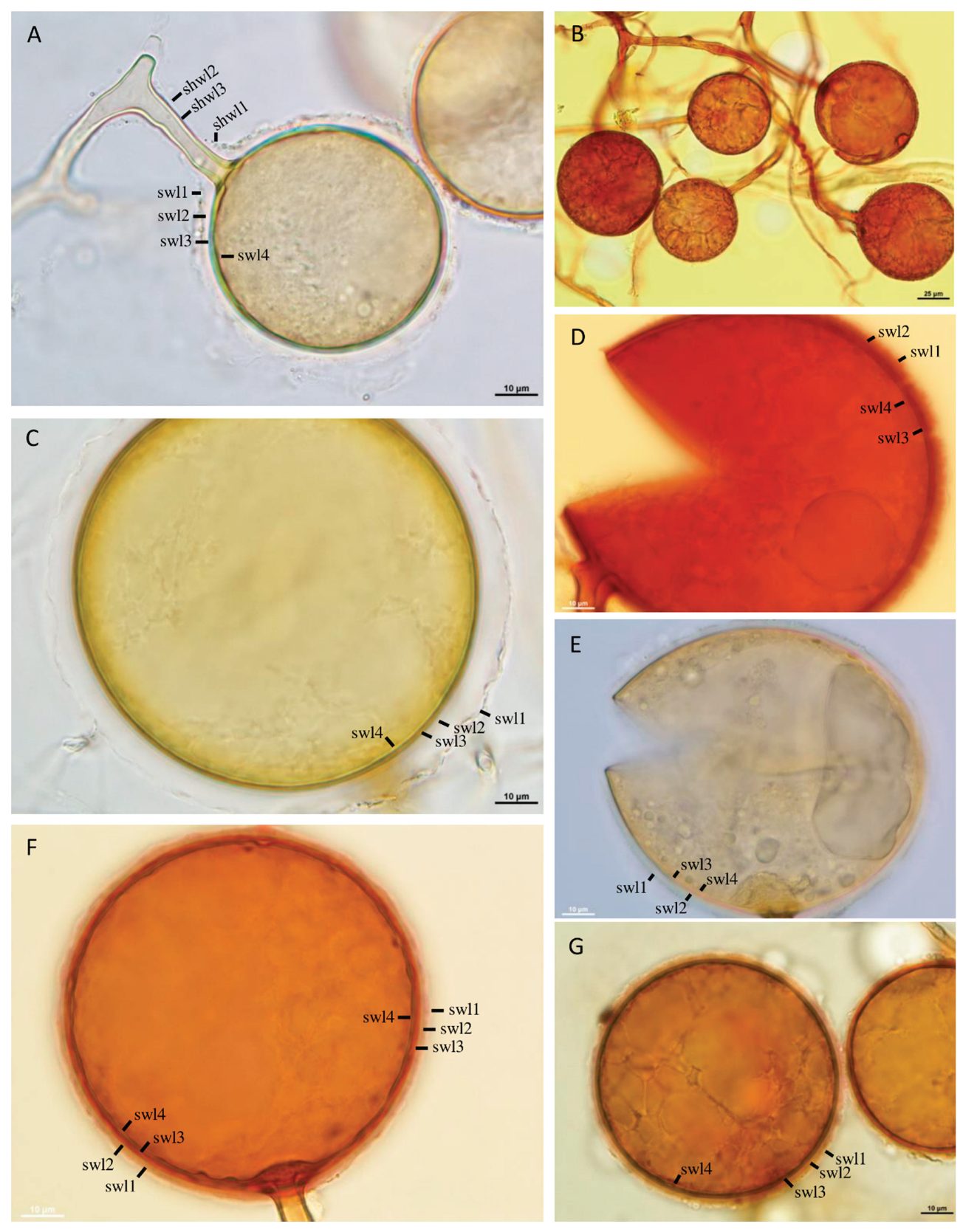
Layer 1, forming the spore surface, evanescent, mucilaginous, short-lived, hyaline, (0.6‒)0.7(‒0.8) µm thick, often highly expanding in spores mounted in PVLG and separating from the upper surface of spore wall layer 2 by up to 15.0 µm, frequently entirely sloughed off in mature spores (Figure 3A,C). Layer 2 uniform, permanent, smooth, semi-flexible, hyaline, (0.5‒)0.8(‒1.3) µm thick, tightly adherent to the upper surface of layer 3, not separating from this layer in even vigorously crushed spores (Figure 3C‒G). Layer 3 laminate, permanent, smooth, semi-flexible to semi-rigid, pale yellow (4A3) to greyish yellow (4B3), (0.8‒)1.5(‒3.0) µm thick, consisting of very thin, <0.5 µm thick, laminae, tightly adherent to and not separating from each other in even vigorously crushed spores (Figure 3C‒F). Layer 4 permanent, flexible, hyaline, 0.5‒1.0 µm thick, detected in mature spores and difficult to see in young spores. In Melzer’s reagent, spore wall layers 1 and 3 stains reddish white (7A2) to deep red (8A3) (Figure 3B,D,F,G). Subtending hypha pale yellow (4A3) to greyish yellow (4B3); straight or recurved, cylindrical to slightly funnel-shaped at the spore base; (7.6‒)9.8(‒13.7) µm wide at the spore base (Figure 3A,B,D,F). Wall of subtending hypha pale yellow (4A3) to greyish yellow (4B3); (1.7‒)2.7(‒3.8) µm thick at the spore base; consisting of four layers continuous with spore wall layers 1‒4; subtending hyphal wall layer 1 swelling in PVLG and usually highly deteriorated or, occasionally, entirely sloughed off in mature spores (Figure 3C). Pore (1.5‒)4.5(‒8.0) µm wide at the spore base, open, rarely closed by a curved septum connecting the inner surface of spore wall layer 4 (Figure 3A,F). Spore content of hyaline oily substance. Germination unknown.
Ecology and distribution: In the field, G. highlandensis probably lived in arbuscular mycorrhizal symbiosis with roots of salt-tolerant wetland plant species in Scotland, and with roots of L. arenarius in sand dunes of the Baltic sea, Poland, but no molecular analyses were performed to confirm this assumption. In single-species cultures with P. lanceolata as the host plant, G. highlandensis formed mycorrhiza with arbuscules, vesicles, as well as intra- and extraradical hyphae (data not shown). The RAxML-EPA and GAPPA affiliation analysis using environmental sequences indicated that G. highlandensis was putatively detected in prairie grasses of Canada (EU380107), in an ash sedimentation pond of Czech Republic (HG425911, HG425912) and in forest and groves areas in the USA (JQ029746, HQ895795, JX848925).
Glomus mongioie Magurno, B.T. Goto, Uszok, M.B. Queiroz, sp. nov.
Figure 4A‒F.
MycoBank No. MB ...… (will be added after revision)
Etymology: mongioie, referring to the Monte Mongioie, where this species was originally found.
Typification: Italy. Spores from a single-species culture established from a trap culture inoculated with a field-collected mixture of rhizospheric soil from Monte Mongioie, a mountain of the Ligurian Alps located in the southern Piedmont (44°09'34.7” N 7°46'29.2 E"), August 2019, F. Magurno. Holotype: slide with spores no. XXXX - will be added after reviews, collection name. Isotype: slides with spores no. XXXX - will be added after reviews, UFRN-Fungos.
Diagnosis: Differs from G. highlandensis, the closest phylogenetic relative together with G. macrocarpum (Figure 1), in: (i) the spore wall structure, (ii) morphometric features of the spore wall, the subtending hyphal wall and pore, and (iii) nucleotide composition of sequences of the 45S nuc rDNA region and the RPB1 gene.
Description: Glomerospores formed in soil, in loose clusters with 2‒21 spores or singly, arise blastically at tips of (i) sporogenous hyphae branched from a parent hypha continuous with an extraradical mycorrhizal hypha (spores in clusters) or (ii) sporogenous hyphae directly continuous with extraradical mycorrhizal hyphae (single spores), Figure 4A‒F. Spores yellowish white (4A2) to greyish yellow (4B5); globose to subglobose; (45‒)86(‒110) µm diam; rarely ovoid; 37‒73 × 45‒83 µm; with one subtending hypha (Figure 4A‒F). Spore wall composed of three layers (Figure 4A,C‒F). Layer 1, forming the spore surface, uniform (not containing visible sublayers), mucilaginous, short-lived, flexible, hyaline, (0.8‒)0.9(‒1.1) µm thick, always highly swelling in spores mounted in PVLG and separating from the upper surface of spore wall layer 2 by up to 14.0 µm, frequently entirely sloughed off in mature spores (Figure 4C‒E). Layer 2 uniform (without visible sublayers), permanent, semi-flexible, smooth, hyaline, (0.8‒)1.1(‒1.4) µm thick, tightly adherent to the upper surface of spore wall layer 3 (swl3), not separating from this layer in even vigorously crushed spores, occasionally difficult to detect when very thin, especially in young spores with a semi-hyaline or brightly coloured swl3 due to the lack of contrast.
Figure 4.
A‒F. Glomus mongioie. A‒B. Cluster with sporogenous hyphae, spores, and a spore subtending hypha (sh). A, C‒F. Spore wall layers (swl) 1‒3. A, F. Subtending hyphal wall layers (shwl) 1‒3 continuous with spore wall layers (swl) 1‒3. A, C‒F. Spores in PVLG. B. Spores in PVLG+Melzer’s reagent. A‒F. Differential interference microscopy.
Figure 4.
A‒F. Glomus mongioie. A‒B. Cluster with sporogenous hyphae, spores, and a spore subtending hypha (sh). A, C‒F. Spore wall layers (swl) 1‒3. A, F. Subtending hyphal wall layers (shwl) 1‒3 continuous with spore wall layers (swl) 1‒3. A, C‒F. Spores in PVLG. B. Spores in PVLG+Melzer’s reagent. A‒F. Differential interference microscopy.
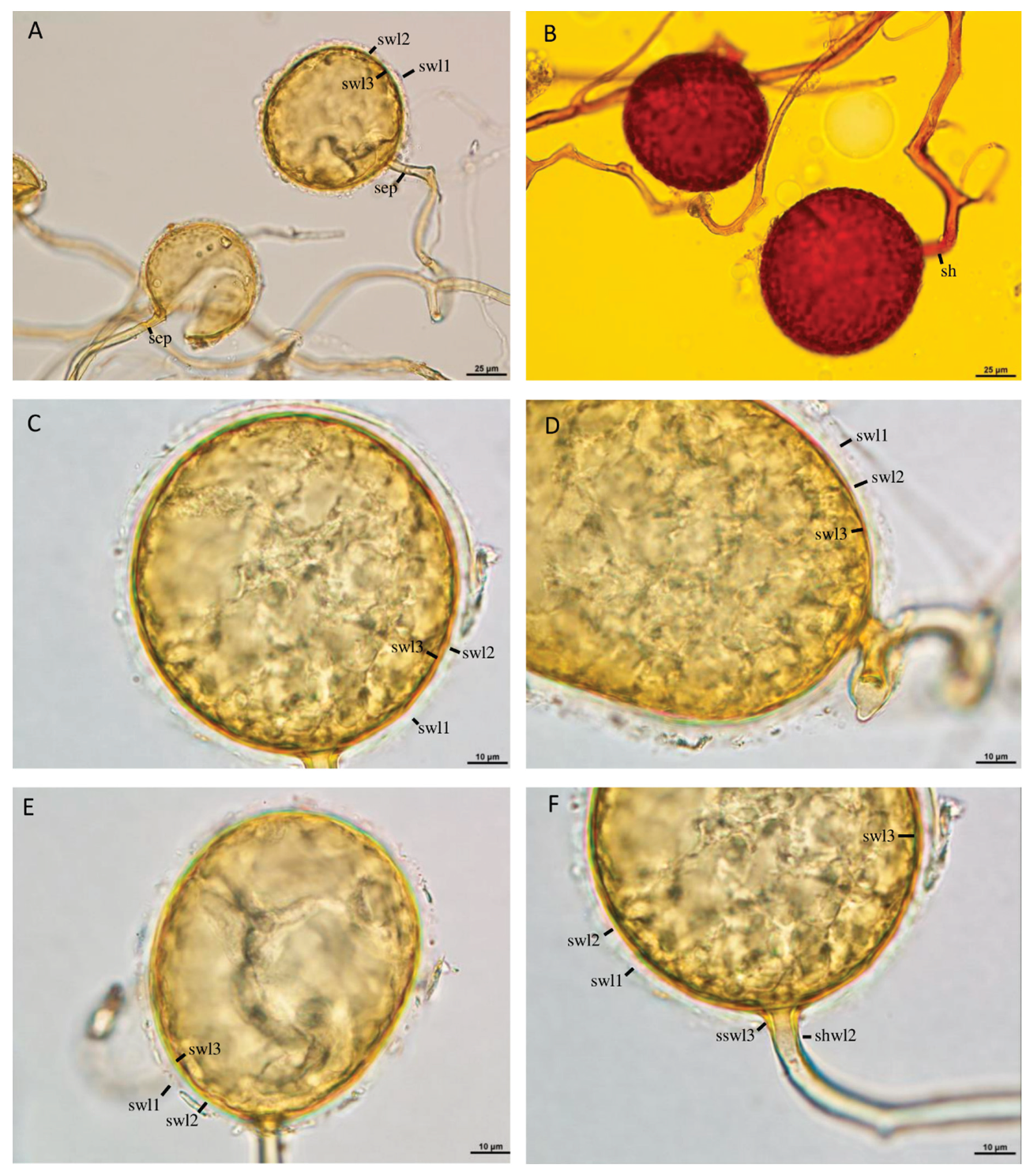
Layer 3 laminate, permanent, smooth, semi-flexible, yellowish white (4A2) to greyish yellow (4B5), (1.1‒)1.3(‒1.6) µm thick, consisting of very thin, <0.5 µm thick, laminae, tightly adherent to and not separating from each other in even vigorously crushed spores (Figure 4C‒F). In Melzer’s reagent, spore wall layer 1 and 3 usually stains reddish white (11A2) to brownish violet (11D8; Figure 4B). Subtending hypha yellowish white (4A2) to greyish yellow (4B5); straight or recurved, usually slightly funnel-shaped, rarely cylindrical or slightly constricted at the spore base; (5.3‒)8.4(‒12.4) µm wide at the spore base (Figure 4A‒F); not braking in crushed spores. Wall of subtending hypha yellowish white (4A2) to greyish yellow (4B5); (1.3‒)2.4(‒3.2) µm thick at the spore base; consisting of three layers continuous with spore wall layers 1‒3; subtending hyphal wall layer 1 swelling in PVLG and usually highly deteriorated or, occasionally, entirely sloughed off in mature spores (Figure 4D,F). Pore (2.5‒)3.9(‒6.4) µm wide at the spore base, usually open, very rarely closed by a curved septum, 0.6‒1.0 µm thick, continuous with some innermost laminae of spore wall layer 3 (Figure 4A). Spore content of hyaline oily substance. Germination unknown.
Ecology and distribution: In the field, G. mongioie lived in arbuscular mycorrhizal symbiosis with roots of herbaceous mountain vegetation in the Ligurian Alps, Italy, but no molecular analyses were performed to confirm this assumption. In single-species cultures with P. lanceolata as the host plant, G. mongioie formed mycorrhiza with arbuscules, vesicles, as well as intra- and extraradical hyphae (data not shown). The RAxML-EPA and GAPPA affiliation analysis using environmental sequences showed no sequences potentially representing G. mongioie, suggesting that this is probably a rare species in the world.
4. Discussion
The morphological analysis supported by the phylogenetic inference using single and concatenated loci confirmed the three AMF isolates obtained from saltmarshes (Highlands, Scotland) and coastal sand dunes (Hel Peninsula, Poland), and from the Mountain Alps (Italy) represent two new species of the genus Glomus. The analysis accommodated the first two isolates, both from hypersaline environments, in G. highlandensis, while the third isolate from Alps represented G. mongioie. The phylogenetic analysis placed the two new species in a supported clade together with G. macrocarpum.
Finally, the molecular and phylogenetic analysis showed the presence of two ribosomal variants in G. highlandensis and G. tetrastratosum, highly divergent and negatively affecting the support of the species clade in G. highlandensis.
4.1. Comparison with Glomus Species
Morphologically, G. highlandensis differs substantially from G. mongioie. In G. highlandensis, the spore wall consists of four layers (Figure 3A,C‒G), while the spore wall of G. mongioie contains three layers (Figure 4A,C‒F), lacking the inner, hyaline, flexible swl4 of the former species. Both species differ fundamentally from G. macrocarpum, their closest species in the phylogenetic analysis. The most significant difference resides in the spore wall. Glomus macrocarpum contains only two layers [11,17], lacking the inner, hyaline, flexible swl4 of the G. highlandensis and the second permanent, uniform, hyaline layer of the G. mongioie. At the moment G. macrocarpum represents a species producing spores in unorganized glomerocarps without convincing evidence of their capacity to grow in single or trap culture. In contrast, G. highlandensis and G. mongioie produced hundreds of spores both in trap and single species culture. The only other glomoid spore-producing species without ornamentation and with a four-layered spore wall is G. tetrastratosum [45]. However, spores of G. highlandensis are clearly lighter (vs. brownish yellow in G. tetrastratosum), 2.2‒2.4-fold smaller when globose.
The only species non-sequenced with similar morphology to that of G. mongioie is G. kerguelense Dalpé & Strullu. Both species produce yellow coloured glomoid-like spores with a three-layered spore wall and a subtending hypha [7,46]. The main differences between these species reside in the phenotypic and histochemical characters of the spore wall layers. Instead of the uniform, hyaline, thin (0.8‒1.4 µm) spore wall layer 2 of G. mongioie, G. kerguelense has a laminate, hyaline to pale yellow, thicker (4.3‒6.8 µm) layer. Spore wall layer 3 in both species is laminate and yellow coloured. But, in G. kerguelense it is clearly thicker (1.6‒2.2 µm), and, most importantly, its inner surface in maturing spores is covered with granular processes, forming a granular/spongiform appearance that disappear with spore maturing (vs. always smooth in G. mongioie). None of the spore wall layers of G. kerguelense stains in Melzer’s reagent like spore wall layer 1 of G. mongioie. Glomus atlanticum, G. bareae and G. chinense, all species of Glomus sensu strictu, also present three layers in the spore wall [18,20,21]. Similarly to G. mongioie, in G. atlanticum and G. bareae the Melzer's reaction is observed in the swl1, but these species differ significantly. The swl3 in G. mongioie is laminated and greyish yellow (vs. laminated and orange brown in G. atlanticum and flexible and hyaline in G. bareae) [20]. In G. chinense, the second, uniform, hyaline slw2 is missing as in G. mongioie. In addition, the Melzer's reaction in G. chinense is observed in swl1‒2 (vs. in swl1 and swl3 in G. mongioie).
4.2. Ribosomal Variants: A “Forgotten” Issue
The existence of ribosomal variants and their possible implication in the phylogenetic placement of AMF species have been known for a long time. Already 20 years ago, [47] was warning about the possibility of misleading phylogenetic information using ribosomal markers in environmental studies. Based on analysis of the 28S D2 region, [47] detected several variants in isolates of Glomus claroideum and G. etunicatum (currently Entrophospora claroidea and E. etunicata; [7]) with a level of divergence up to 19% bp differences among the sequences. [48] investigated the rRNA gene diversity amplifying the 5.8S-partial 28S region from single spores in nine populations of Claroideoglomus (=Entrophospora) species (E. claroidea, E etunicata, E. lutea). According to the study two groups of sequences, called S and L variants, were detected, clustering in two distinct supported clades as result of ancestral polymorphism. Each clade was populated by sequences from the three species investigated. The S (=short) variant differed from the L (=long) variant primarily by a 100 bp deletion in the ITS2 and by multiple nucleotide polymorphisms along the 28S region. Both variants were considered functional after verification of gene expression and estimation of the structural stability. Finally, recent comparative analysis of genomes from several isolates of Rhizoglomus irregulare confirmed the presence in this species of up to 11 divergent rRNA copies on each haplotype investigated, located on different chromosomes [49,50]. These late findings expanded our knowledge about the presence of divergent rRNA variants even outside the genus Entrophospora.
In the present study we had further confirmation, detecting two highly divergent variants among G. highlandensis and G. tetrastratosum sequences, probably as a consequence of ancestral polymorphism. Differently from what was observed by [48], the differences among the variants were observed mostly in the ITS1 locus, with an insertion of ca 40-60 bp. Furthermore, the variants were not forming independent clades trespassing the species boundaries, but they had a negative influence on the species clades’ support of G. highlandensis. Further investigations, principally employing genome sequencing, are needed to understand the magnitude of this phenomenon in the Glomeromycota, how it could affect the phylogeny of closely related species, and how it could mislead taxonomists in recognizing “artificial” species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Bird’s-Eye View of ITS1 alignment; Figure S2: 50% majority-rule consensus tree from the Bayesian analysis of dataset ds2 (S-variant sequences); Figure S3: Phylogenetic placement of environmental sequences potentially related to the two new Glomus species.
Author Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by Franco Magurno, Sylwia Uszok, Bruno Tomio Goto, Mariana Bessa de Queiroz, and Karolina Bierza. The first draft of the manuscript was written by Franco Magurno, Bruno Tomio Goto, and Mariana Bessa de Queiroz and all authors commented on previous versions of the manuscript. Conceptualisation: Franco Magurno, Bruno Tomio Goto; methodology: Franco Magurno, Sylwia Uszok, Bruno Tomio Goto, and Mariana Bessa de Queiroz; formal analysis and investigation: Franco Magurno, Sylwia Uszok, Bruno Tomio Goto, and Mariana Bessa de Queiroz; writing original draft preparation: Franco Magurno, Bruno Tomio Goto and Mariana Bessa de Queiroz; writing—review and editing: Franco Magurno, Sylwia Uszok, Bruno Tomio Goto, Mariana Bessa de Queiroz, Karolina Bierza, Zoltan Kende, and Jawdat Bakr. The funding acquisition: Franco Magurno and Bruno Tomio Goto; resources: Franco Magurno and Bruno Tomio Goto; Supervision: Franco Magurno and Bruno Tomio Goto. All authors read and approved the final manuscript.
Funding
This research was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico, proc. 306632/2022–5.
Data Availability Statement
Datasets generated during and/or analyzed during the current study are available from the corresponding author upon request. Sequences obtained in the study are available in Genbank with the accession numbers: XXX will be added after review.
Acknowledgments
We thank Mrs Paola Galluzzo for her precious contribution in the description of the floristic characteristics of Monte Mongioie.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wilkes, T.I.; Warner, D.J.; Edmonds-Brown, V.; Davies, K.G.; Denholm, I. Zero Tillage Systems Conserve Arbuscular Mycorrhizal Fungi, Enhancing Soil Glomalin and Water Stable Aggregates with Implications for Soil Stability. Soil Syst 2021, 5, 4. [Google Scholar] [CrossRef]
- Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Apori, S.O.; Ndiaye, A.; Badji, A.; Ngom, K. Roles of Arbuscular Mycorrhizal Fungi on Soil Fertility: Contribution in the Improvement of Physical, Chemical, and Biological Properties of the Soil. Front Fungal Biol 2022, 3, 723892. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xiao, Q.; Geng, X.; Lin, K.; Li, Z.; Li, Y.; Chen, J.; Li, X. Arbuscular mycorrhizal fungi alter rhizosphere bacterial diversity, network stability and function of lettuce in barren soil. Sci Hortic 2024, 323, 112533. [Google Scholar] [CrossRef]
- Pandit, A.; Kochar, M.; Srivastava, S.; Johny, L.; Adholeya, A. Diversity and Functionalities of Unknown Mycorrhizal Fungal Microbiota. Microbiol Res 2022, 256, 126940. [Google Scholar] [CrossRef] [PubMed]
- Salomon, M.J.; Demarmels, R.; Watts-Williams, S.J.; McLaughlin, M.J.; Kafle, A.; Ketelsen, C.; Soupir, A.; Bücking, H.; Cavagnaro, T.R.; van der Heijden, M.G.A. Global evaluation of commercial arbuscular mycorrhizal inoculants under greenhouse and field conditions. Appl Soil Ecol 2022, 169, 104225. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.; Dai, D.; Sánchez-García, M.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Sánchez-García, M.; Niezgoda, P.; Zubek, S.; Fernández, F.; Vila, A.; Al-Yahya’ei, M.N.; Symanczik, S.; Milczarski, P.; Malinowski, R.; Cabello, M.; Goto, B.T.; Casieri, L.; Malicka, M.; Bierza, W.; Magurno, F. A new order, Entrophosporales, and three new Entrophospora species in Glomeromycota. Front Microbiol 2022, 13, 962856. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.A.; Corazon-Guivin, M.A.; de Assis, D.M.A.; Oehl, F. Blaszkowskia, a new genus in Glomeraceae. Mycol Prog 2023, 22, 74. [Google Scholar] [CrossRef]
- Tulasne, L.R.; Tulasne, C. Fungi nonnulli hipogaei, novi v. minus cognito act. Giornale Botanico Italiano 1845, 2, 55–63. [Google Scholar]
- Tulasne, L.R.; Tulasne, C. Fungi hypogaei. Histoire et monographie des champignons hypogds. Paris, 1851.
- Gerdemann, J.W.; Trappe, J.M. The Endogonaceae of the Pacific Northwest. Micologie Memoir 1974, 5, 1–76. [Google Scholar]
- Schüßler, A.; Walker, C. The Glomeromycota. A species list with new families and new genera. Gloucester, England, 2010.
- Oehl, F.; Silva, G.A.; Goto, B.T.; Maia, L.C.; Sieverding, E. Glomeromycota: two new classes and a new order. Mycotaxon 2011, 116, 365–379. [Google Scholar] [CrossRef]
- Oehl, F.; Silva, G.A.; Goto, B.T.; Sieverding, E. Glomeromycota: three new genera and glomoid species reorganized. Mycotaxon 2011, 116, 75–120. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Chwat, G.; Góralska, A.; Ryszka, P.; Kovács, G.M. Two new genera, Dominikia and Kamienskia, and D. disticha sp. nov. in Glomeromycota. Nova Hedwigia 2015, 1-2, 225–238. [Google Scholar]
- Sieverding, E.; Silva, G.A.; Reinhard, B.; Oehl, F. Rhizoglomus, a new genus of the Glomeraceae. Mycotaxon 2015, 129, 373–386. [Google Scholar] [CrossRef]
- Berch, S.M.; Fortin, J.A. Lectotypification of Glomus macrocarpum and proposal of new combinations: Glomus australe, Glomus versiforme, and Glomus tenebrosum (Endogonaceae). Canad J Bot 1983, 61, 2608–2617. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Niezgoda, P.; Goto, B.T.; Kozłowska, A. Halonatospora gen. nov. with H. pansihalos comb. nov. and Glomus bareae sp. nov. (Glomeromycota; Glomeraceae). Botany 2018, 96, 737–748. [Google Scholar] [CrossRef]
- Guillén, A.; Serrano-Tamay, F.J.; Peris, J.B.; Arrillaga, I. Glomus ibericum, Septoglomus mediterraneum, and Funneliformis pilosus, three new species of arbuscular mycorrhizal fungi. Mycologia 2020, 1–10. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Niezgoda, P.; Zubek, S.; Meller, E.; Milczarski, P.; Malicka, M.; Goto, B.T.; Woźniak, G.; Moreira, H.; Magurno, F. Dominikia bonfanteae and Glomus atlanticum, two new species in the Glomeraceae (phylum Glomeromycota) with molecular phylogenies reconstructed from two unlinked loci. Mycol Prog 2021, 20, 131–148. [Google Scholar] [CrossRef]
- Yu, F.; Goto, B.T.; Magurno, F.; Błaszkowski, J.; Wang, J.; Ma, W.; Feng, H.; Liu, Y. Glomus chinense and Dominikia gansuensis, two new Glomeraceae species of arbuscular mycorrhizal fungi from high altitude in the Tibetan Plateau. Mycol Prog 2022, 21, 32. [Google Scholar] [CrossRef]
- Firth, C.; Fleet, L. Information Sheet on Ramsar Wetlands (RIS), 2006.
- Gunn, G.F. Measuring nocturnal near-surface urban heat island intensity in the small, mid-latitude city of Inverness, Scotland. Scott Geogr J 2023, 1–18. [Google Scholar] [CrossRef]
- Polderman, P.J.; Polderman-Hall, R.A. Algal communities in Scottish saltmarshes. Brit Phycol J 1980, 15, 59–71. [Google Scholar] [CrossRef]
- Woch, M.W.; Kapusta, P.; Stanek, M.; Możdżeń, K.; Grześ, I.M.; Rożej-Pabijan, E.; Stefanowicz, A.M. Effects of invasive Rosa rugosa on Baltic coastal dune communities depend on dune age. NeoBiota 2023, 82, 163–187. [Google Scholar] [CrossRef]
- Fratianni, S.; Acquaotta, F. The climate of Italy. In: Landscapes and landforms of Italy, Eds. Soldati M., Marchetti M., Springer, 2017, pp 29-38.
- Kornerup, A.; Wanscher, J. H. Methuen handbook of colour, 3rd ed.; E. Methuen: London, UK, 1983. [Google Scholar]
- Furrazola, E.; Torres-Arias, Y.; Ferrer, R.L.; Herrera, R.A.; Berbara, R.; Goto, B.T. Glomus crenatum (Glomeromycetes), a new ornamented species from Cuba. Mycotaxon 2011, 116, 143–132. [Google Scholar] [CrossRef]
- Goto, B.T.; Jardim, J.G.; da Silva, G.A.; Furrazola, E.; Torres-Arias, Y.; Oehl, F. Glomus trufemii (Glomeromycetes), a new sporocarpic species from Brazilian sand dunes. Mycotaxon 2012, 120, 1–9. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Jobim, K.; Niezgoda, P.; Meller, E.; Malinowski, R.; Milczarski, P.; Zubek, S.; Magurno, F.; Casieri, L.; Bierza, W.; Błaszkowski, J.; Crossay, T.; Goto, B.T. New Glomeromycotan Taxa, Dominikia glomerocarpica sp. nov. and Epigeocarpum crypticum gen. nov. et sp. nov. From Brazil, and Silvaspora gen. nov. From New Caledonia. Front Microbiol 2021, 12, 655910. [Google Scholar] [CrossRef] [PubMed]
- Walker, C. Taxonomic concepts in the Endogonaceae: spore wall characteristics in species descriptions. Mycotaxon 1983, 18, 443–455. [Google Scholar]
- Błaszkowski, J. Glomeromycota; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2012. [Google Scholar]
- Goto, B.T.; Maia, L.C. Glomerospores: a new denomination for the spore of Glomeromycota, a group molecularly distinct from the zygomycota. Mycotaxon 2006, 96, 129–132. [Google Scholar]
- Jobim, K.; Błaszkowski, J.; Niezgoda, P.; Kozłowska, A.; Zubek, S.; Mleczko, P.; Chachuła, P.; Ishikawa, N.K.; Goto, B.T. New sporocarpic taxa in the phylum Glomeromycota: Sclerocarpum amazonicum gen. et sp. nov. in the family Glomeraceae (glomerales) and Diversispora sporocarpia sp. nov. in the Diversisporaceae (Diversisporales). Mycol Prog 2019, 18, 369–384. [Google Scholar] [CrossRef]
- Krüger, M.; Stockinger, H.; Krüger, C.; Schüßler, A. DNA-based species level detection of Glomeromycota: one PCR primer set for all arbuscular mycorrhizal fungi. New Phytol 2009, 183, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Błaszkowski, J.; Yamato, M.; Niezgoda, P.; Zubek, S.; Milczarski, P.; Malinowski, R.; Meller, E.; Malicka, M.; Goto, B.T.; Uszok, S.; Casieri, L.; Magurno, F. A new genus, Complexispora, with two new species, C. multistratosa and C. mediterranea, and Epigeocarpum japonicum sp. nov. Mycol Prog 2023, 22, 34. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In: Proceedings of the Gateway Computing Environments Workshop, New Orleans, LA: IEEE 2010, 1-8.
- Abadi, S.; Azouri, D.; Pupko, T.; Mayrose, I. Model selection may not be a mandatory step for phylogeny reconstruction. Nat Commun 2019, 10, 934. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinformatics 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.A.; Krompass, D.; Stamatakis, A. Performance, accuracy, and web server for evolutionary placement of short sequence reads under maximum likelihood. Syst Biol 2011, 60, 291–302. [Google Scholar] [CrossRef]
- Czech, L.; Barbera, P.; Stamatakis, A. Genesis and Gappa: processing, analyzing and visualizing phylogenetic (placement) data. Bioinformatics 2020, 36, 3263–3265. [Google Scholar] [CrossRef] [PubMed]
- Malicka, M.; Magurno, F.; Piotrowska-Seget, Z. Phenol and Polyaromatic Hydrocarbons are Stronger Drivers Than Host Plant Species in Shaping the Arbuscular Mycorrhizal Fungal Component of the Mycorrhizosphere. Int J Mol Sci 2022, 23, 12585. [Google Scholar] [CrossRef] [PubMed]
- Błaszkowski, J.; Chwat, G.; Góralska, A.; Bobrowska-Chwat, A. Glomus tetrastratosum, a new species of arbuscular mycorrhizal fungi (Glomeromycota). MycoScience 2015, 56, 280–286. [Google Scholar] [CrossRef]
- Dalpé, Y.; Plenchette, C.; Frenot, Y.; Gloaguen, J.C.; Strullu, D.G. Glomus kerguelense, a new Glomales species from sub-Antarctic. Mycotaxon 2002, 84, 51–60. [Google Scholar]
- Rodriguez, A.; Clapp, J.P.; Robinson, L.; Dodd, J.C. Studies on the diversity of the distinct phylogenetic lineage encompassing Glomus claroideum and Glomus etunicatum. Mycorrhiza 2005, 15, 33–46. [Google Scholar] [CrossRef]
- VanKuren, N.W.; den Bakker, H.C.; Morton, J.B.; Pawlowska, T.E. Ribosomal RNA gene diversity, effective population size, and evolutionary longevity in asexual Glomeromycota. Evolution 2013, 67, 207–224. [Google Scholar] [CrossRef]
- Sperschneider, J.; Yildirir, G.; Rizzi, Y.S.; Malar, M.C.; Nicol, AM.; Sorwar, E.; Villeneuve-Laroche, M.; Chen, E.C.H.; Iwasaki, W.; Brauer, E.K.; Bosnich, W.; Gutjahr, C.; Corradi, N. Arbuscular mycorrhizal fungi heterokaryons have two nuclear populations with distinct roles in host–plant interactions. Nat Microbiol 2023, 8, 2142–2153. [Google Scholar] [CrossRef] [PubMed]
- Yildirir, G.; Sperschneider, J.; Malar, M.C.; Chen, E.C.H.; Iwasaki, W.; Cornell, C.; Corradi, N. Long reads and Hi-C sequencing illuminate the two-compartment genome of the model arbuscular mycorrhizal symbiont Rhizophagus irregularis. New Phytol 2022, 233, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
50% majority-rule consensus tree from the Bayesian analysis of 45S sequences concatenated with RPB1 sequences of Glomus highlandensis, G. mongioie, the 6 species of Glomus sensu strictu in possession of molecular data, and two Complexispora species (the closest taxa to Glomus in the family Glomeraceae) serving as outgroup. The two new species are in bold coloured font. Different shades of blue are used to distinguish the two isolates of G. highlandensis. Bayesian posterior probabilities ≥0.95 and ML bootstrap values ≥60% are shown near the branches, respectively. Bar indicates 0.1 expected change per site per branch.
Figure 1.
50% majority-rule consensus tree from the Bayesian analysis of 45S sequences concatenated with RPB1 sequences of Glomus highlandensis, G. mongioie, the 6 species of Glomus sensu strictu in possession of molecular data, and two Complexispora species (the closest taxa to Glomus in the family Glomeraceae) serving as outgroup. The two new species are in bold coloured font. Different shades of blue are used to distinguish the two isolates of G. highlandensis. Bayesian posterior probabilities ≥0.95 and ML bootstrap values ≥60% are shown near the branches, respectively. Bar indicates 0.1 expected change per site per branch.
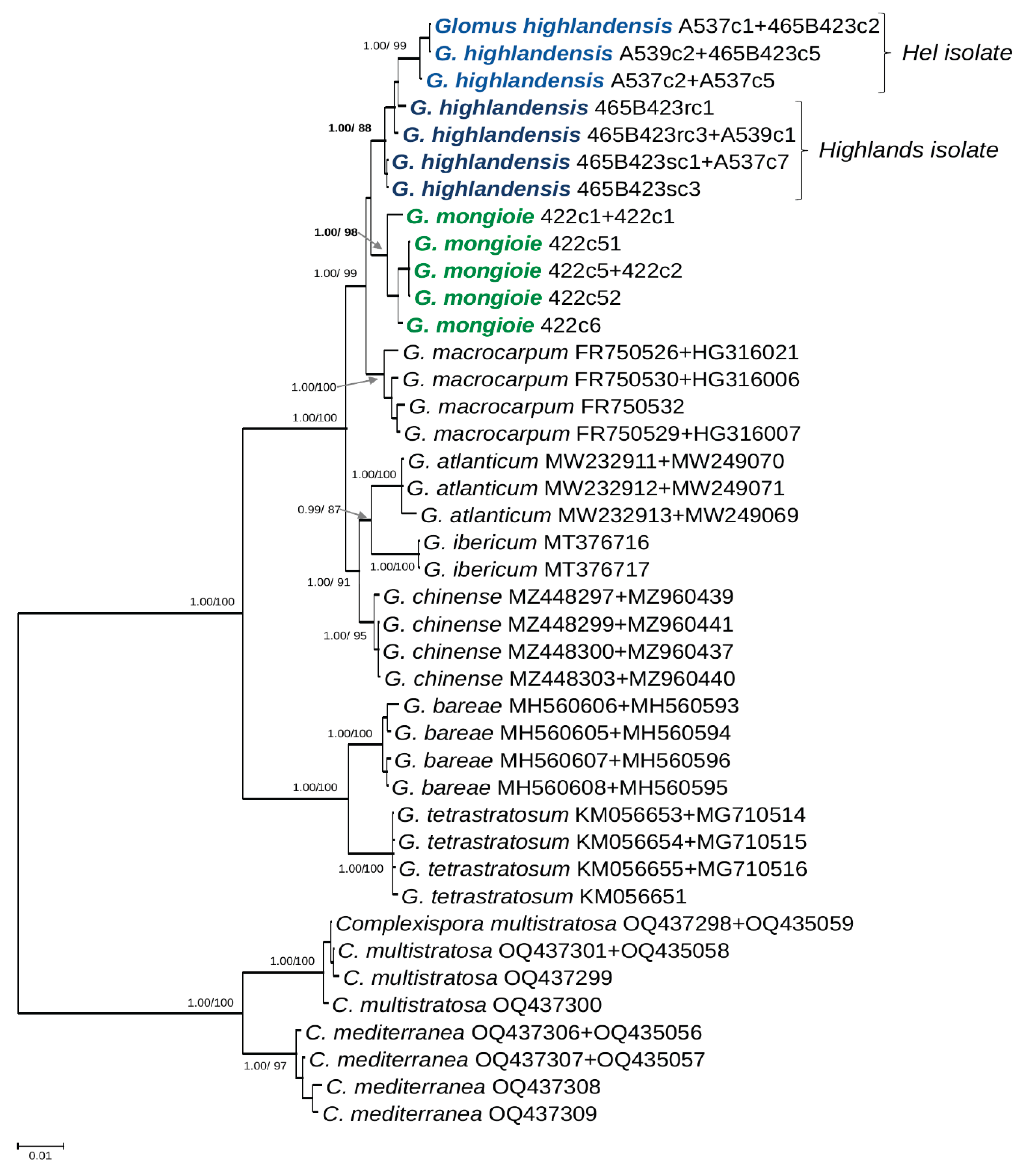
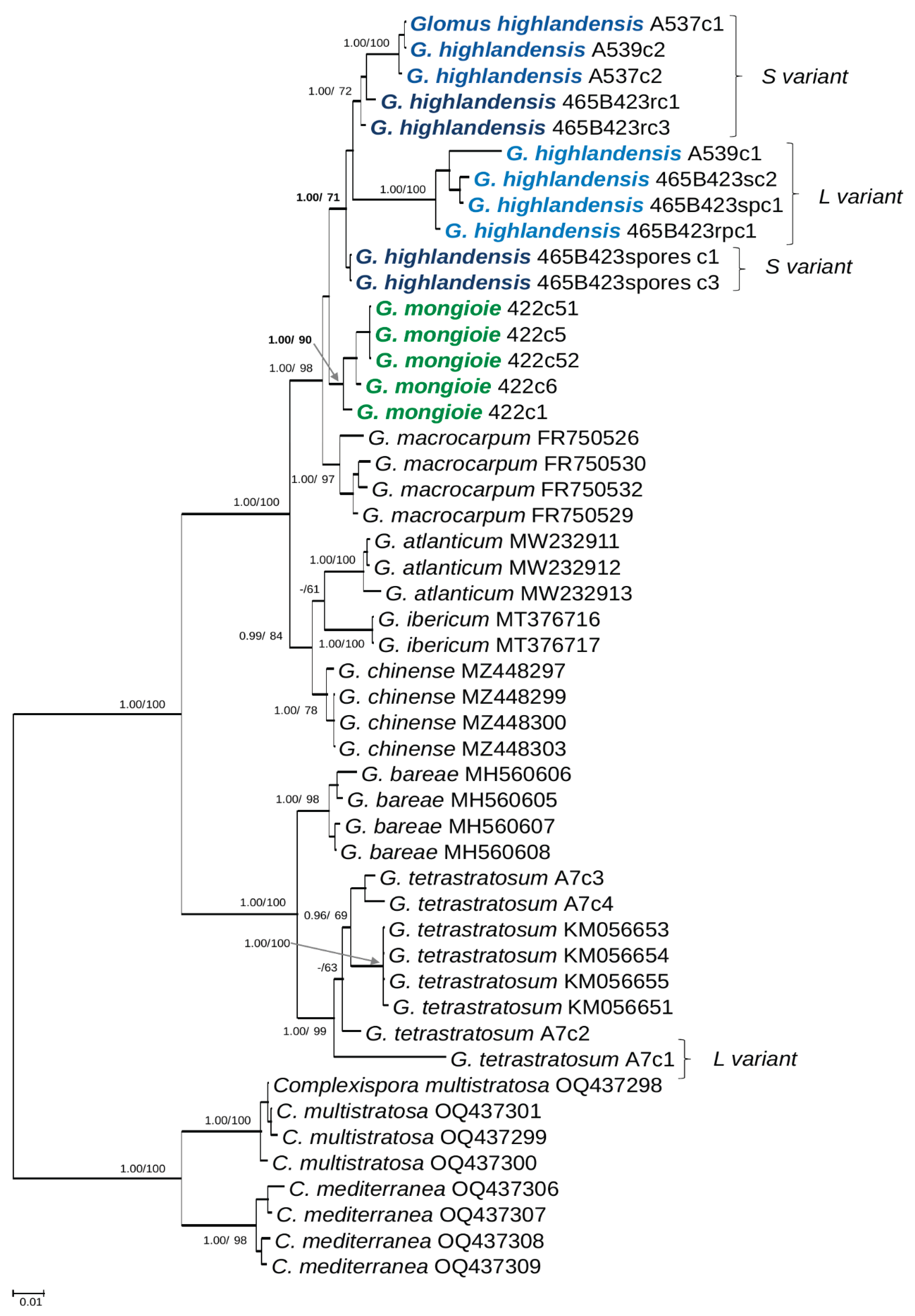
Figure 2.
50% majority-rule consensus tree from the Bayesian analysis of dataset ds1 (S- and L-variant sequences). The dataset consisted in 45S sequences of Glomus highlandensis, G. mongioie, six species of Glomus sensu strictu in possession of molecular data, and two Complexispora species (the closest taxa to Glomus in the family Glomeraceae) serving as outgroup. The two new species are in bold coloured font. Different shades of blue are used to distinguish the two isolates of G. highlandensis, and their L-variant sequences. Bayesian posterior probabilities ≥0.95 and ML bootstrap values ≥60% are shown near the branches, respectively. Bar indicates 0.1 expected change per site per branch.
Figure 2.
50% majority-rule consensus tree from the Bayesian analysis of dataset ds1 (S- and L-variant sequences). The dataset consisted in 45S sequences of Glomus highlandensis, G. mongioie, six species of Glomus sensu strictu in possession of molecular data, and two Complexispora species (the closest taxa to Glomus in the family Glomeraceae) serving as outgroup. The two new species are in bold coloured font. Different shades of blue are used to distinguish the two isolates of G. highlandensis, and their L-variant sequences. Bayesian posterior probabilities ≥0.95 and ML bootstrap values ≥60% are shown near the branches, respectively. Bar indicates 0.1 expected change per site per branch.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



