
Submitted:
08 April 2024
Posted:
09 April 2024
You are already at the latest version
Abstract
Ecuador is an important center of biodiversity of the plant family Amaryllidaceae, specially the subfamily Amaryllidoideae, known as a source of important bioactive molecules. The aim of this study was to assess the chemical and biological potential of four different species of Amaryllidoideae collected in Ecuador: Urceolina formosa, Urceolina ruthiana, Clinanthus incarnatus, and Stenomesson aurantiacum. Sixteen alkaloids were identified in the bulb extract of these species by GC-MS, which the extract of S. aurantiacum exhibiting the greatest structural diversity. The potential of these three genera against Alzheimer’s disease was evaluated by measuring their AChE and BuChE inhibitory activity, revealing that C. incarnatus and U. formosa (from Sucumbíos province) showed the best results with IC50 values of 1.73 ± 0.25 and 30.56 ± 1.56 g·ml-1, respectively. Molecular dynamic assays were conducted to elucidate the possible interactions that occurs among 2-hydroxyanhydrolycorine and AChE enzyme, suggesting some similarity with those observed for galanthamine. This study enhances our understanding of the biodiversity of Amaryllidoideae species from Ecuador, highlighting their potential as a source of chemical compounds with pharmaceutical potential.
Keywords:
Amaryllidoideae
; alkaloids
; Amazon lily
; Alzheimer’s disease
; computational experiments
1. Introduction
The Amaryllidaceae family, specifically the subfamily Amaryllidoideae, is an important source of an exclusive and still expanding group of alkaloids, known as Amaryllidaceae alkaloids, that shows a broad spectrum of biological activities [1]. This subfamily comprises ca. 900 species and 75 genera, bringing up about 700 Amaryllidaceae alkaloids, which are mainly classified into nine skeleton types, represented by norbelladine-, lycorine-, homolycorine-, crinine-, haemanthamine-, narciclasine-, tazettine-, montanine- and galanthamine-type structures [2,3].
The subfamily Amaryllidoideae is found in different zones, being South America, South Africa, and the Mediterranean region, the most representative [2]. Cultures worldwide have utilized Amaryllidaceae plants for centuries in traditional medicine, recognizing their pharmaceutical potential often linked to the production of specific alkaloids [4]. Ecuador is the major center of diversity in the tribe Eucharideae (Amaryllidaceae) [5], including the genus Urceolina Planch. This genus, known as Amazon lily, is mainly distributed in the western Amazon basin and the adjoining lower slopes of the eastern Andean cordillera [6]. The genus Stenomesson Herb. (Amaryllidaceae) is mainly found in Peru, with only S. aurantiacum Herb. previously reported in Ecuador [7]. These plants usually occur in seasonally dry, grassy vegetation or at the margins of cloud forests above 2000 m elevation, but are also found in Peruvian inter-Andean valleys below 2000 m [8], and in the loma formations along the coast of this country, being all the petiolate-leafed Stenomesseae more closely related to Eucharideae than to the lorate leafed Stenomesseae [5,9]. The genus Clinanthus Herb. (Amaryllidaceae), endemic to Peru and Ecuador [10], was separated from Stenomesson, and is primarily known in its same locations [8,11,12].
Regarding the traditional use of these plants, Urceolina species have been documented as compresses applied to sores and tumors by native people in Ecuador, and the Jíbaro indigenous people in Peru have utilized them for treating facial blemishes and acne [13]. Additionally, archeological findings at Inca ruins in South America suggest that certain Amaryllidaceae genera, such as Stenomesson, were depicted on ceremonial drinking vessels, and may have held significance within popular culture [8,14]. In recent years, the Amaryllidaceae species from Ecuador have garnered significant attention due to their alkaloid profiling and potential therapeutic benefits against Alzheimer's disease. The description and understanding of these species have progressively expanded in the literature in the last 10 years [15,16,17,18].
Dementia is a significant contributor to disability and dependency in the elderly population [19]. This condition gives rise to physical, psychological, social, and economic impacts, with Alzheimer's disease being the most prevalent form of this disorder [19]. The development of Alzheimer's disease appears to involve various mechanisms, one of which is a decrease in the neurotransmitter acetylcholine in the inter-synaptic space [20]. Currently, galanthamine, donepezil, and rivastigmine are the only cholinesterase inhibitor drugs approved by FDA for the clinical treatment of mild to moderate Alzheimer’s disease symptomatology. The first one, galanthamine - an Amaryllidaceae alkaloid originally isolated from Galanthus woronowii in 1952, has been commercialized under its salt form since 2001 [21]. However, the synthesis of this molecule remains expensive, as pharmaceutical companies continue to obtain it from Amaryllidaceae species [22].
The aim of this study was to evaluate the chemical and biological potential of five samples of Amaryllidaceae collected in Ecuador. The alkaloid extracts from the bulbs of Urceolina formosa Meerow, Urceolina ruthiana L. Jost, Oleas & Meerow, Clinanthus incarnatus (Kunth) Meerow, and Stenomesson aurantiacum (Kunth) Herb. were analyzed using gas chromatography coupled to mass spectrometry (GC-MS). Additionally, their potential in combating Alzheimer's disease was estimated by evaluating their inhibitory activity against cholinesterase enzymes, acetylcholinesterase (AChE) and butyrylcholinesterase (BuChE). Computational experiments were also conducted to gain further insights into the molecular interactions among alkaloid-AChE.
2. Results and Discussion
2.1. Alkaloid Profile
The alkaloid extracts of five samples of Amaryllidaceae collected in Ecuador were analyzed, resulting in the quantification of sixteen alkaloids. These molecules are listed in Table 1 and visually represented in Figure 1. The quantities of each compound are reported in milligrams of galanthamine (mg GAL) per gram of alkaloid extract (g AE).
Some similarities among the alkaloid profiling of the two samples of Urceolina formosa (collected in Tungurahua’s and Sucumbíos’ provinces) and Urceolina ruthiana have been observed in this scientific report (Table 1). All these samples showed the presence of alkaloids from haemanthamine- and lycorine-type skeleton in their chemical composition. The alkaloid haemanthamine (10) was reported in all the samples of Urceolina evaluated in this work, and presented a large abundance in U. formosa (sample B) and U. ruthiana (sample C), 172.5 and 101.4 mg GAL.g−1 AE, respectively. The alkaloid 11-hydroxyvittatine (13) was found in both samples of U. formosa (samples A and B), but not in U. ruthiana. The alkaloid lycorine (15) also has been detected in all the extracts of Urceolina evaluated herein, being the highest concentration observed in the species U. formosa (from Sucumbíos province, sample B), followed by U. formosa (from Tungurahua province, sample A), with 273.9 and 251.6 mg GAL.g−1 AE, respectively.
Galanthamine (2) was identified in the bulbs extract of both samples of U. formosa (sample A and B), being the highest amount in sample A, 46.2 mg GAL.g−1 AE. The alkaloid tazettine (11), which belongs to pretazettine-type scaffolds, was reported only in the extract of U. ruthiana, 32.6 mg GAL.g−1 AE. This compound is also known as 6-deoxytazzetine, which is an artifact from the alkaloid pretazettine [24]. No unidentified compound has been observed in the species of Urceolina evaluated herein (Table 1).
The alkaloid profiling of Urceolina bonplandii (published as Eucharis bonplandii) collected in Colombia showed the presence of structures from narciclasine-, galanthamine-, haemanthamine-, and lycorine-type scaffolds, comprising twelve Amaryllidaceae alkaloids, together with one unidentified compound [25,26]. The same authors also reported the alkaloid composition of Urceolina caucana (published as Eucharis caucana) collected in two different locations from Colombia - Risaralda and Chocó, describing the presence of structures belonging to narciclasine-, galanthamine-, and lycorine-type alkaloids in both samples, as well as structures from haemanthamine/crinine-, pretazettine-, and montanine-type which were observed just in the second species [25].
As shown in Table 1, C. incarnatus presented three alkaloids among its chemical profile, being lycorine (15) the most abundant (149.9 mg GAL·g−1 AE). A great amount of the compound 16, which must be identified as 2-hydroxyanhydrolycorine by Soto-Vásquez and co-workers [23], has been quantified in this species, 60 mg GAL·g-1 AE (Table 1). According to the literature, C. incarnatus collected in Peru presented a high amount of lycorine (15), as well as seven other different alkaloids [27]. A current publication revealed the presence of two lycorine-type alkaloids in C. incarnatus collected in Ecuador, which are lycorine and 11,12-dehydroanhydrolycorine, and 4 unidentified structures, that probably belong to lycorine- and ismine-type skeleton [28].
According to Table 1, Stenomesson aurantiacum, sample E, showed the highest content of alkaloids, totalizing 502.3 mg GAL·g−1 AE. Among the samples evaluated herein, this plant extract presented the most diversity of structures, totalizing 11 compounds, which two of them were not identified, 4 and 14, and seem to belong to galanthamine- and pretazettine-type scaffold, respectively. The molecules 11,12-dehydroanhydrolycorine (12), 11-hydroxyvittatine (13), and galanthamine (2) were the most representative in this species, with 181.6, 108.1, and 74.1 mg GAL·g−1 AE, respectively. Lycorine (15) has been quantified in all the samples evaluated in this work, except in S. aurantiacum (Table 1). In a previous work, 22 alkaloids were detected in S. aurantiacum collected in Ecuador, being haemanthamine the most abundant [15].
2.2. AChE and BuChE Inhibitory Activity
The bulbs alkaloid extract of the five samples of Amaryllidaceae collected in Ecuador presented inhibitory activity against AChE and BuChE (Figure 2). The samples of C. incarnatus (sample D) and both samples of U. formosa (sample A and B) showed the best results toward AChE, with IC50 values of 1.73 ± 0.25, 2.63 ± 0.83, and 2.81 ± 0.48 μg·ml-1, respectively. The samples of U. formosa (sample A and B) presented the best results against BuChE, with IC50 values of 34.22 ± 1.46 and 30.56 ± 1.56 μg·ml-1, respectively, while C. incarnatus (sample D) showed IC50 values of 50.51 ± 3.93 μg·ml-1 against this enzyme. The IC50 values obtained with the alkaloid extract of U. ruthiana and S. aurantiacum against AChE were 8.15 ± 0.07 and 3.41 ± 0.05 μg·ml-1, respectively, and 57.26 ± 1.69 and 81.60 ± 9.97 μg·ml-1, respectively, against BuChE. Galanthamine was used as control and showed IC50 values of 0.57 ± 0.05 and 3.77 ± 0.20 μg·ml-1 against AChE and BuChE, respectively (Figure 2). As shown in Table 1, the presence of this alkaloid was observed in the bulbs extract of both samples of U. formosa (sample A and B), and S. aurantiacum (sample E), and it must be contributing for the cholinesterase inhibitory effect of these species.
According to the literature, the bulbs extract of U. bonplandii, U. caucana (Risaralda) and U. caucana (Chocó) collected in Colombia showed IC50 values lower than 10 μg·ml-1 against the enzymes Electrophorus electricus and human AChE, and higher than 15 μg·ml-1 against horse serum and human BuChE [26]. Among these samples, the authors reported that the best results were obtained with the bulbs extract of U. bonplandii which exhibited IC50 values of 0.72 ± 0.05 μg·ml-1 against hAChE [26]. Both leaves and bulbs extracts of Urceolina bouchei Woodson & P.H. Allen collected in Panama and evaluated by Calderón and co-authors showed activity against AChE though TLC bioautographic methodology, while the whole plant extract was inactive [29]. A currently publication described the IC50 values of the alkaloid extract of C. incarnatus as higher than 40 μg·ml-1 against AChE and BuChE, representing the low activity of this species against both enzymes [28]. Although the alkaloid profiling of S. aurantiacum have been previously reported in the literature, this is the first report about its anti-cholinestarase potential.
Previously works about six species of Phaedranassa (Amaryllidaceae) from Ecuador report their potential against Alzheimer’s disease [16,17]. According to the literature, the species P. cuencana was the most active against AChE, with IC50 values of 0.88 ± 0.11 μg·ml-1, while the best activity against BuChE was observed for P. dubia, which showed IC50 values of 14.26 ± 2.71 μg·ml-1 [16,17]. Recently, the bulbs and leaves alkaloid extract of Crinum x amabile also collected in Ecuador have been evaluated against the same enzymes, being the bulbs the most active against AChE, with IC50 values of 1.35 ± 0.13 μg·ml-1, while the leaves showed better activity against BuChE, with IC50 values of 8.50 ± 0.76 and 45.42 ± 3.72 μg·ml-1, respectively [18].
2.3. Molecular Docking Results
According to the results shown in Table 1 and Figure 2, the species C. incarnatus presented a high concentration of the compound 16, 2-hydroxyanhydrolycorine, and the best activity against AChE, respectively. Then, to know how 16 could work in the inhibition of this enzyme, a theoretical inhibition experiment was carried out through a molecular docking procedure using galantamine as a control. The Tetronarce californica AChE, a classical structure for biochemical studies of cholinergic neurotransmission [30,31], was used for all in-silico analysis. Molecular docking results provided binding energies for 2-hydroxyanhydrolycorine of -8.94 kcal·mol-1 vs. galanthamine, -9.43 kcal·mol-1. As a first approximation, our results indicate 2-hydroxyanhydrolycorine is not as active as galanthamine. Nevertheless, molecular docking data are only approximate and a molecular dynamics investigation must be performed to determine the true behavior within the active site of AChE.
The TcAChE protein's active site is about 20 nm deep and is composed by three important regions: (i) peripheral anionic site (PAS), which is composed predominantly by Tyr70, Tyr121, Asp72, Trp279 and Tyr334 residues, all located at the gate of the active site; ii) alpha-anionic site, place made up by Trp84 and Phe330 residues and are located in the middle of active site; and iii) catalytic anionic site (CAS), mainly constituted by Ser200, His440, and Glu 327 residues [32,33]. The ligand’s position in TcAChE enzyme is reported in Figure 3. The 2-hydroxyanhydrolycorine molecule is located at the entrance of peripheral anionic site, PAS, and exhibits stabilizing interactions with Leu76, Trp86, Phe295, Tyr337, Tyr341 and Hsd447. Galantamine is located at the catalytic site bottom and shows stabilizing interactions with Trp86, Glu202, Ser203, Phe295, Tyr341, and Tyr337. Although 2-hydroxyanhydrolycorine and galanthamine inhibit differently the TcAChE, the results indicate that there are some residues which stabilize both ligands (Trp86, Phe295, Tyr337, Tyr341). Additionally, experimental reports have found synergistic effects of Amaryllidaceae alkaloids on inhibition of AChE, so this process could be carried out successfully [34].
2.3. Molecular dynamics (MD) simulations
Molecular dynamics simulations were performed on the poses obtained by molecular docking experiments with the aim of elucidating how the structure 2-hydroxyanhydrolycorine could be inhibiting TcAChE. The free energy of binding for both complexes were performed using molecular mechanics generalized Born solvent accessibility (MM-GBSA). A small variation in root-mean-square deviation (RMSD) study indicates the stability of protein-ligand conformations during MD (Figure 4).
The binding free energies reported in Table 2 suggest 2-hydroxyanhydrolycorine complex is significantly less stable than galantamine complex at 13 kcal.mol-1. These results were consistent with those obtained in molecular docking experiments and indicate that, although 2-hydroxyanhydrolycorine is not as active as galanthamine, it could be a potential inhibitor of TcAChE.
A frequency graph of contact between the most significant residues and 2-hydroxyanhydrolycorine and galantamine molecules along the MD is presented in Figure 4. A radius of 3 Å has been chosen due to this being the average distance between hydrogen bond variations in the analyzed systems. The MD results show galantamine remaining mostly in the bottom of the active site, near Asn87, Ser125, Gly126, Tyr133, Glu202, Ser203, and Phe295 residues, whereas 2-hydroxyanhydrolycorine is localized at the PAS site near to Leu76, Thr83, Tyr77, Trp286, Ser293, Val294, Glu334, Phe338, and Pro446 residues. Finally, Figure 5 shows both ligands have a significant frequency of contact with Phe297 residue, which is involved in the formation of the acyl-binding pocket.
3. Materials and Methods
3.1. Plant Material
Five samples of Amaryllidaceae have been evaluated in this study, all of them collected in Ecuador in 2019 (Figure 6 and Figure 7). Urceolina formosa (Ravenna) Ravenna (sample A) was collected at Sucumbíos province, at Cantón Shushufindi, Recinto el Mirador, kilometer 5, Ecuador. Urceolina formosa (Ravenna) Ravenna (sample B) was collected at Tungurahua province at Reserva Rio Zuñag. Urceolina ruthiana L. Jost, Oleas & Meerow (sample C) was obtained from Zamora-Chinchipe province, Copalinga private reserve, near Zamora. Clinanthus incarnatus (Kunth) Meerow (sample D) was collected at Chimborazo province, Guasuntos Panamericana highway. Stenomesson aurantiacum (Kunth) Herb. (sample E) was collected at Loja Province, close to the limit to Azuay. The species were authenticated by Alan Meerow from Arizona State University, USA.
3.2. Alkaloid Extraction
The bulbs of Urceolina, Clinanthus, and Stenomesson species were dried at 40 °C for 7 days. Then, the samples were triturated, and their powder was macerated with methanol at room temperature for 3 days, changing the solvent daily (3 × 100 mL), and, trying to increase the extraction potential, and ultrasonic baths was applied for 20 min, 8 times per day. The mash was filtered and evaporated to dryness under reduced pressure. The crude extracts of the bulbs were acidified with an aqueous solution of sulfuric acid (2%, v/v) to pH 2 and, then cleaned with diethyl ether to remove neutral material. The aqueous solution was basified with ammonium hydroxide at 25% (v/v) to pH 9–10 and extracted with ethyl acetate to obtain the alkaloid extract (AE) of each sample.
3.3. GC-MS Analysis
Two mg of each sample were dissolved in 1 ml of methanol: chloroform (0.5:0.5, v/v) and evaluated using GC-MS (Agilent Technologies 6890N coupled with MSD5975 inert XL; Santa Clara, CA, USA) equipment operating in electron ionization (EI) mode at 70 eV. A Sapiens-X5 MS column (30 m × 0.25 mm i.d., film thickness 0.25 µm; Teknokroma, Barcelona, Spain) was used. One µL of each sample was injected into the equipment using the splitless mode. Codeine (0.05 mg.ml−1) was used as the internal standard. The injector and detector temperatures were 250 and 280 °C, respectively, and the flow-rate of carrier gas (He) was 1 ml.min-1. The temperature gradient was 12 min at 100 °C, 100–180 °C at 15 °C·min−1, 180–300 °C at 5 °C·min−1, and 10 min hold at 300 °C.
A calibration curve of galanthamine (10, 20, 40, 60, 80, and 100 µg.ml-1 was applied to quantify every single constituent detected in the chromatogram, using codeine (0.05 mg·ml−1) as the internal standard. Peak areas were manually obtained, considering selected ions for each compound (usually the base peak of their MS, i.e., m/z at 286 for galanthamine and 299 for codeine). The ratio between the values obtained for galanthamine and codeine in each solution was plotted against the corresponding concentration of galanthamine to obtain the calibration curve and its equation (y = 0.0112x − 0.0469; R2 = 0.9995). All data were standardized to the area of the internal standard (codeine), and the equation obtained for the calibration curve of galanthamine was used to calculate the amount of each alkaloid. Results are presented as mg GAL (galanthamine), which was finally related to the alkaloid extract (AE). As the peak area does not only depend on the corresponding alkaloid concentration but also on the intensity of the mass spectra fragmentation, the quantification is not absolute, but it is considered suitable to compare the specific alkaloid amount in Amaryllidaceae samples [35].
3.4. AChE and BuChE Inhibitory Activity
Cholinesterases inhibitory activities were determined according to Ellman and co-workers [36] with some modifications as by López and co-workers [37]. Stock solutions with 500U of AChE from Electrophorus electricus (Merck, Darmstadt, Germany) and BuChE from equine serum (Merck, Darmstadt, Germany), respectively, were prepared and kept at −20 °C. Acetylthiocholine iodide (ATCI), S-butyrylthiocholine iodide (BTCI), and 5,50-dithiobis (2-nitrobenzoic) acid (DTNB) were obtained from Merck (Darmstadt, Germany). Fifty microliters of AChE or BuChE in phosphate buffer (8 mM K2HPO4, 2.3 mM NaH2PO4, 0.15 NaCl, pH 7.5) and 50 µl of the sample dissolved in the same buffer were added to the wells. The plates were incubated for 30 min at room temperature. Then, 100 µl of the substrate solution (0.1 M Na2HPO4, 0.5 M DTNB, and 0.6 mM ATCI or 0.24 mM BTCI in Millipore water, pH 7.5) was added. These reagents were obtained from Merck (Darmstadt, Germany). After 10 min, the absorbance was read at 405 nm in a Labsystem microplate reader (Helsinki, Finland). Enzymes activities were calculated as percent compared to a control using a buffer without any inhibitor. Galanthamine served as a positive control. In the first step, the activities of each sample against AChE and BuChE were assessed at 10, 100, and 200 µg·ml−1. Then, the calibration curves of bulbs alkaloid extracts of Urceolina, Clinanthus, and Stenomesson species towards both enzymes were calculated and applied to obtain IC50 values. The cholinesterase inhibitory data were analyzed with the Prism 10 software.
3.5. Statistical Analysis
Three independent assays were used to evaluate the cholinesterase activity of the alkaloid extracts from the Amaryllidaceae species collected in Ecuador. Results were analyzed by ANOVA, using the Prism 10 software. Data are expressed as the mean ± standard deviation (SD). Significant results are marked as follows: **** p < 0.0001 and ** p < 0.01. For AChE and BuChE, one-way ANOVA with Dunnett’s multiple comparison test was used to compare the mean of each column with the mean of a control column (galanthamine).
3.5. Molecular docking
The binding mechanisms of 2-hydroxyanhydrolycorine galantamine alkaloids to the active site in Torpedo californica acetylcholinesterase (TcAChE) protein, PDB code 1DX6 [38], were investigated using the AutoDock v.4.2 tool [39]. The use of TcAChE protein has been reported as a good instrument in the search for new cholinesterase inhibitors [40]. As a protein preparation procedure for the molecular docking experiment water, cofactors, and ions were removed from X-ray crystallographic structure. The polar hydrogen atoms were added, the atomic charges computed toward the Gasteiger procedure, and the non-polar hydrogen atoms merged. The grid maps required by AutoDock were computed using the auxiliary program AutoGrid, choosing a 60 Å × 60 Å × 60 Å grid box around the active site. The Lamarckian Genetic Algorithm (LGA) was used to conduct the docking searches [41] using 2000 individuals as a population, obtaining 2,500,000 energy evaluations for each 100 LGA run. The best docking complex poses were analyzed according to intermolecular interactions (ligand/enzyme), such as hydrogen bonding, hydrophobic interactions, and the cation–π, π–π stacking.
3.5. Molecular dynamics simulations (MD)
MD experiments were achieved on the ligand-TcAChE (PDBID:1DX6) complex, in aqueous solutions with galantamine and 2-hydroxyanhydrolycorine as ligands employing TIP3P water model as an explicit solvent [42] (≈16.000 water molecules). Also, Na+ and Cl- ions were added to neutralize the systems and remain an ionic concentration of 0.15 mol·L-1. A general AMBER force field (GAFF) has been used to parametrize galantamine and 2-hydroxyanhydrolycorine molecules [43,44]. The protein structure was modeled with CHARMM27 par_all27_prot_lipid.inp parameter [45]. The simulations were carried out using a standard MD protocol as follows: (I) minimization and structural relaxation of water molecules utilizing 2000 stages of minimization (downward step) and MD simulations with an NPT (310 K) assembly by 1000 ps employing harmonic constraints of 10 kcal⋅mol Å-2 on the protein and ligand; (II) minimization of the complete structure considering 2000 downstream minimization steps and 6500 steps of conjugate gradient minimization; (III) the minimized systems were progressively heated to 310 K, with harmonic restrictions of 10 kcal⋅mol Å-2 in the carbon skeleton and ligand during 0.5 ns; (IV) The system was then balanced for 0.5 ns while adhering to the constraints, and then for 5 ns without constraints to 310 K in a canonical assembly (NVT); and (V) a production dynamic was conducted for 50 ns without constraints at 310 K and 1 atm with a temporary passage of 2 fs using an isothermal isobaric assembly (NPT). In the MD simulation, the temperature was controlled by the Langevin dynamics with a collision frequency of 1 ps-1 (NVT) and the pressure with the Berendsen barostat (NPT). In addition, the Particle Mesh Ewald (PME) method with a cut-off value of 10 Å was used to treat nonbonding and long-range electrostatic interactions. All MD simulation calculations were performed using NAMD, software developed by the Theoretical and Computational Biophysics Group in the Beckman Institute for Advanced Science and Technology at the University of Illinois at Urbana-Champaign [46,47]. Molecular visualization of the systems and MD trajectory analysis were carried out with the VMD software package [48].
3.5.1. Free energy calculations
The binding free energy of TcAChE-ligand complexes was estimated using the molecular MM/GBSA technique. For computations, the first 40 ns of MD were isolated and the explicit water molecules and ions were deleted. Three subsets of each system were analyzed using MM/ GBSA: the protein alone, the ligand alone, and the complex (protein-ligand). The total free energy (ΔGtot) was computed as follows for each of these subsets:
Where EMM denotes the bonded and Lennard–Jones energy components; GSolv denotes the polar and nonpolar contributions to the solvation energy, respectively; T denotes the temperature; and ΔSconf denotes the conformational entropy [49]. Both EMM and GSolv were calculated using NAMD program with the generalized Born implicit solvent model [50]. ΔGtot was calculated as a linear function of the solvent-accessible surface area, which was calculated with a probe radius of 1.4 Å [51]. The difference between the binding free energies of TcACh and ligand complexes (ΔGbind) was used to compute the binding free energy of TcACh and ligand complexes (ΔGbind) where values represent the simulation’s averages.
4. Conclusions
In summary, this study was the first report about the alkaloid profiling of Urceolina species collected in Ecuador, as well as the first work about the anti-cholinesterase potential of the species studied herein, excepted C. incarnatus. The extract of Stenomesson aurantiacum presented unidentified structures among its profiling, suggesting this species as a possible source of new Amaryllidaceae alkaloids. The samples C. incarnatus and Urceolina formosa (collected in Sucumbíos province) provided the best results against AChE and BuChE, respectively. Computational experiments suggest some similarity in the interactions observed among 2-hydroxyanhydrolycorine and TcAChE with those observed for galanthamine. In vitro assays with 2-hydroxyanhydrolycorine must be carried out to corroborate the in silico findings. Our results highlight the importance of Amaryllidaceae species in the search of new bioactive molecules.
Author Contributions
Conceptualization, L.R.T. and N.H.O.; methodology, L.R.T., K.A.L., R.C., and E.H.O.; software, L.R.T. and E.H.O.; validation, E.H.O.; formal analysis, L.R:T., R.C., and E.H.O.; investigation, L.R.T. and E.H.O.; resources, K.A.L., E.H.O., J.B., and N.H.O.; data curation, L.R.T., K.A.L., and E.H.O.; writing—original draft preparation, L.R.T., E.H.O., and N.H.O.; writing—review and editing, K.A.L. and J.B.; visualization, L.J.; supervision, N.H.O.; project administration, J.B. and N.H.O.; funding acquisition, N.H.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Programa CYTED, grant number, 416RT0511 and Universidad TecnológicaIndoamérica, grant number Biocamb-02-(2016–2020). and “The APC was funded by Universidad Indoamérica Grant number INV-0019-003-006 ”.
Data Availability Statement
Data is contained within the article.
Acknowledgments
The authors thank the Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo (CYTED, 223RT0140) for providing the framework and economic support. L.R.T. and J.B. thank CCiTUB for technical support. The authors thank the Ministerio del Ambiente of Ecuador for collecting permits MAE-DNB-CM-2018-0086.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bastida, J.; Lavilla, R.; Viladomat, F. Chemical and biological aspects of Narcissus alkaloids. In: Cordell GA (ed), The Alkaloids: Chemistry and Physiology. 63, Elsevier, 2006, Amsterdam, pp. 87–179.
- Berkov S, Osorio E, Viladomat F, Bastida J (2020) Chemodiversity, chemotaxonomy and chemoecology of AA. In: Knölker H-J (ed), The Alkaloids: Chemistry and Biology. 83, Elsevier, Amsterdam, pp. 113–185. [CrossRef]
- Meerow, A.W.; Gardner, E.M.; Nakamura, K. Phylogenomicsof the Andean Tetraploid Clade of theAmerican Amaryllidaceae (SubfamilyAmaryllidoideae): Unlockinga Polyploid Generic Radiation Abettedby Continental Geodynamics. Front. Plant Sci. 2020, 11, 582422. [Google Scholar] [CrossRef] [PubMed]
- Kornienko, A.; Evidente, A. Chemistry, biology and medicinal potential of narciclasine and its congeners. Chem. Rev. 2008, 108, 1982–2014. [Google Scholar] [CrossRef] [PubMed]
- Meerow, A.W.; Jost, L.; Oleas, N. Two new species of endemic Ecuadorean Amaryllidaceae (Asparagales,Amaryllidaceae, Amarylloideae, Eucharideae). Phytokeys 2015, 48, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Meerow, A.W. Biosyntematics of two sympatric species of Eucharis (Amaryllidaceae). Pl Syst Evol 1989, 166, 11–30. [Google Scholar] [CrossRef]
- Meerow, A.W. 202. Amaryllidaceae. In: Harling G, Andersson L (Eds) Flora of Ecuador 41: 1–52. Univ. Göteborg/Riksmuseum, Stockholm/Pontificia Univ. Católica del Ecuador, Quito, 1990.
- Meerow, A.W.; Werff, H. van der. Pucara (Amaryllidaceae) Reduced to Synonymy with Stenomesson on the Basis of Nuclear and Plastid DNA Spacer Sequences, and a New Related Species of Stenomesson. Syst. Bot. [CrossRef]
- Meerow, A.W.; Guy, C.L. ; Li, Q-B.; Yang, S-L. Phylogeny of he American Amaryllidaceae based on nrDNA ITS sequences. Syst. Bot. :4. [CrossRef]
- Alzate, F.; Lesmes, M.; Cortés, N.; Varela, S.; Osorio, E. Sinopsis de la família Amaryllidaceae em Colombia. Biota Colombiana, 1: 20; :1.
- León, B.; Sagástegui, A.; Sánchez, I.; Zapata, M.; Meerow, A.; Cano, A. Amaryllidaceae endémicas del Perú. Rev. Peru. Biol. 2: 13, 1727; :2. [Google Scholar]
- Leiva, S.; Meerow, A.W. A new species of Clinanthus from northern Peru (Asparagales, Amaryllidaceae, Amarylloideae, Clinantheae). PhytoKeys, 2016, 63, 99–106. [Google Scholar] [CrossRef]
- Cabezas, F.; Ramírez, A.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Eucharis amazonica (Amaryllidaceae). Chem Pharm Bull.
- Vargas, C.C. Phytomorphic representations of the ancient Peruvians. Econ. Bot. 1962, 16, 106–115. [Google Scholar] [CrossRef]
- Acosta, K.; Pigni, N.; Oleas, N.; Bastida, J. Identification of the alkaloids of Steno- messon aurantiacum (Kunth) Herb., an Amaryllidaceae species from the Ecuadorian Andes. PharmacologyOnline 2014, 3, 178–183. [Google Scholar]
- Acosta, K.; Inca, A.; Tallini, L.R.; Osorio, E.H.; Robles, J.; Bastida, J.; Oleas, N.H. Alkaloids of Phaedranassa dubia (Kunth) J.F. Macbr. and Phaedranassa brevifolia Meerow (Amaryllidaceae) from Ecuador and its cholinesterase-inhibitory activity. S Afri J Bot. [CrossRef]
- Moreno, R.; Tallini, L.R.; Salazar, C.; Osorio, E.H.; Montero, E.; Bastida, J.; Oleas, N.H.; León, K.A. Chemical profiling and cholinesterase inhibitory activity of five Phaedranassa Herb. (Amaryllidaceae) species from Ecuador. Molecules 2020, 25, 2092. [Google Scholar] [CrossRef]
- Tallini, L.R.; Carrasco, A.; Acosta, K.L.; Vinueza, D.; Bastida, J.; Oleas, N.H. Alkaloid Profiling and Cholinesterase Inhibitory Potential of Crinum × amabile Donn. (Amaryllidaceae) Collected in Ecuador. Plants 2021, 10, 2686. [Google Scholar] [CrossRef]
- WHO, 2023. Acessible online: https://www.who.int/health-topics/dementia#tab=tab_1 Accessed on 20 Nov, 2023.
- Konrath, E.L.; dos Santos, C.P.; Klein-Júnior, L.C.; Henriques, A.T. Alkaloids as a source of potential anticholinesterase inhibitors for the treatment of Alzheimer’s disease. J. Pharm. Pharm. 2013, 65, 1701–1725. [Google Scholar] [CrossRef]
- Berkov, S.; Georgieva, L.; Kondakova, V.; Atanassov, A.; Viladomat, F.; Bastida, J.; Codina, C. Plant sources of galanthamine: Phytochemical and biotechnological aspects. Biotechnol. Biotechnol. Equip. 2009, 23, 1170–1176. [Google Scholar] [CrossRef]
- Berkov, S.; Georgieva, L.; Sidjimova, B.; Bastida, J. Evaluation of Hippeastrum papilio (Ravenna) Van Scheepen potencial as a new industrial source of galanthamine. Ind. Crop. Prod., 2022, 178, 114619. [Google Scholar] [CrossRef]
- Soto-Vásquez, M.R.; Alvarado-García, P.A.A.; Osorio, E.H.; Tallini, L.R.; Bastida, J. Antileishmanial activity of Clinanthus milagroanthus S. Leiva & Meerow (Amaryllidaceae) collected in Peru. Plants, 2023, 12, 322. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, J.P.; Pigni, N.B.; Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Bioactive alkaloid extracts from Narcissus broussonetii: mass spectral studies. J Pharmaceut Biomed 2012, 70, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Cortes, N.; Castañeda, C.; Osorio, E.H.; Cardona-Gomez, G.P.; Osorio, E. Amaryllidaceae alkaloids as agents with protective effects against oxidative neural cell injury. Life Sci. 2018, 203, 54–65. [Google Scholar] [CrossRef]
- Cortes, N.; Sierra, K.; Alzate, F.; Osorio, E.H.; Osorio, E. Alkaloids of Amaryllidaceae as Inhibitors of Cholinesterases (AChEs and BChEs): An Integrated Bioguided Study. Phytochem Anal, :2. [CrossRef]
- Soto-Vásquez, M.R.; Horna-Pinedo, M.V.; Tallini, L.R.; Bastida, J. Chemical Composition and In Vitro Antiplasmodial Activity of the Total Alkaloids of the Bulbs of Two Amaryllidaceae Species from Northern Peru. Pharmacogn. J. 2021, 13, 1046–1052. [Google Scholar] [CrossRef]
- Rodríguez-Escobar, M.L.; Tallini, L.R.; Lisa-Molina, J.; Berkov, S.; Viladomat, F.; Meerow, A.; Bastida, J.; Torras-Claveria, L. Chemical and Biological Aspects of Different Species of the Genus Clinanthus Herb. (Amaryllidaceae) from South America. Molecules 2023, 28, 5408. [Google Scholar] [CrossRef]
- Calderón, A.I.; Cubilla, M.; Espinosa, A.; Gupta, M.P. Screening of plants of Amaryllidaceae and related families from Panama as sources of acetylcholinesterase inhibitors. Pharmaceutical Biology, :9.
- Lindstrom, J.; Cooper, J.; Tzartos, S. Acetylcholine receptors from Torpedo and Electrophorus have similar subunit structures. Biochemistry. [CrossRef]
- Stavrianakou, M.; Perez, R.; Wu, C.; Sachs, M.S.; Aramayo, R.; Harlow, M. Draft de novo transcriptome assembly and proteome characterization of the electric lobe of Tetronarce californica: a molecular tool for the study of cholinergic neurotransmission in the electric organ. BCM Genomics 2017, 18, 611. [Google Scholar] [CrossRef]
- Silman, I.; Sussman, J.L. Acetylcholinesterase: How is structure related to function? Chem-Biol. Interact. 2008, 175, 3–10. [Google Scholar] [CrossRef]
- Dvir, H.; Silman, I.; Harel, M.; Rosenberry, T.L.; Sussman, J.L. Acetylcholinesterase: From 3D structure to function. Chem-Biol. Interact. 2010, 187, 10–22. [Google Scholar] [CrossRef]
- Orhan, I.; Sener, B. Bioactivity-directed fractionation of alkaloids from some Amaryllidaceae plants and their anticholinesterase activity. Chem. Nat. Compd. 2003, 39, 383–386. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Metabolomic analysis of bioactive Amaryllidaceae alkaloids of ornamental varieties of Narcissus by GC-MS combined with k-means cluster analysis. Ind. Crop. Prod. 2014, 56, 211–222. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, Jr.V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- López, S.; Bastida, J.; Viladomat, F.; Codina, C. Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci. 2002, 71, 2521–2529. [Google Scholar] [CrossRef]
- Greenblatt, H.M.; Kryger, G.; Lewis, T.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with (-)galanthamine at 2. 3 Å resolution. FEBS Lett. 1999, 463, 321–326. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2785; :16. [Google Scholar] [CrossRef]
- Sierra, K.; de Andrade, J.P.; Tallini, L.R.; Osorio, E.H.; Yañéz, O.; Osorio, M.I.; Oleas, N.H.; García-Beltrán, O.; Borges, W. de S.; Bastida, J.; Osorio, E.; Cortes, N. In vitro and in silico analysis of galanthine from Zephyranthes carinata as an inhibitor of acetylcholinesterase. Biomed. Pharmacother. 2022, 150, 113016. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated Docking Using a Lamarckian Genetic Algorithm and an Empirical Binding Free Energy Function. J. Comput. Chem. 1639; :14. [Google Scholar] [CrossRef]
- Neria, E.; Fischer, S.; Karplus, M. Simulation of activation free energies in molecular systems. J. Chem. Phys. 1996, 105, 1902–1921. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 1157; :9. [Google Scholar] [CrossRef]
- Özpinar, G.A.; Peukert, W.; Clark, T. An improved generalized AMBER force field (GAFF) for urea. J. Mol. Model. 2016, 16, 1427–1440. [Google Scholar] [CrossRef] [PubMed]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An overview of the Amber biomolecular simulation package. J. Comput. Chem. 1668; :16. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable Molecular Dynamics with NAMD. J. Comput. Chem. 1781; :16. [Google Scholar] [CrossRef]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graphics. :1. [CrossRef]
- Hayes, J.M.; Archontis, G. MM-GB(PB)SA Calculations of Protein-Ligand Binding Free Energies. Molecular Dynamics - Studies of Synthetic and Biological Macromolecules. 2012. [CrossRef]
- Götz, A.W.; Williamson, M.J.; Xu, D.; Ducan, P.; Grand, S.L.; Walker, R.C.; Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 1. Generalized Born. J. Chem. Theory. Comput. 2012, 8, 1542–1555. [Google Scholar] [CrossRef]
- Abroshan, H.; Akbarzadeh, H.; Parsafar, G.A. Molecular dynamics simulation and MM–PBSA calculations of sickle cell hemoglobin in dimer form with Val, Trp, or Phe at the lateral contact. J. Phys. Org. Chem. :9. [CrossRef]
Figure 1.
Structures of the alkaloids identified in Amaryllidaceae species collected in Ecuador.
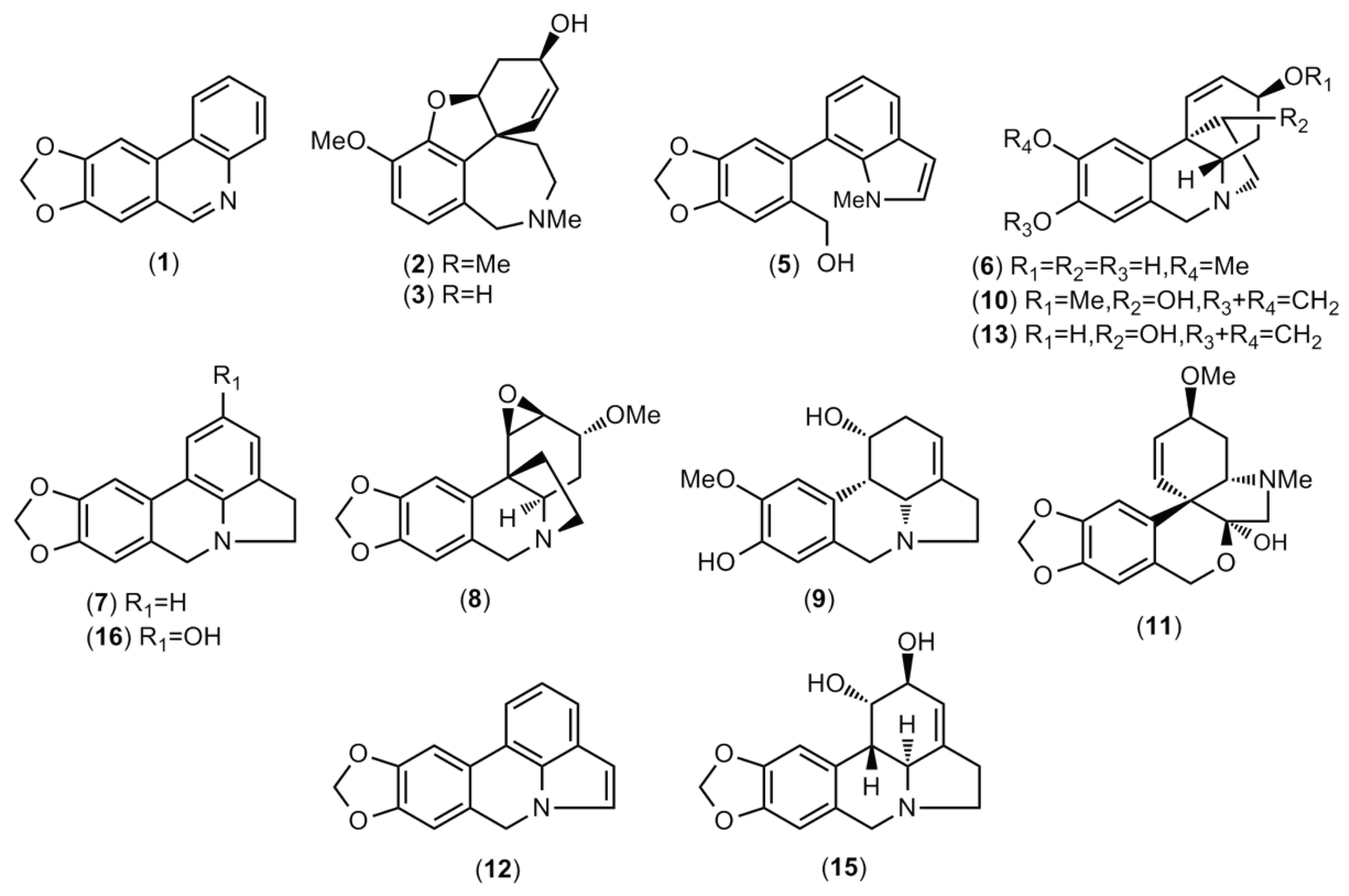
Figure 2.
AChE and BuChE inhibitory activity of different species of Amaryllidaceae collected in Ecuador. A: Urceolina formosa (from Tungurahua province); B: Urceolina formosa (from Sucumbíos province); C: Urceolina ruthiana; D: Clinanthus incarnatus; E: Stenomesson aurantiacum; GAL: galanthamine (control).
Figure 2.
AChE and BuChE inhibitory activity of different species of Amaryllidaceae collected in Ecuador. A: Urceolina formosa (from Tungurahua province); B: Urceolina formosa (from Sucumbíos province); C: Urceolina ruthiana; D: Clinanthus incarnatus; E: Stenomesson aurantiacum; GAL: galanthamine (control).
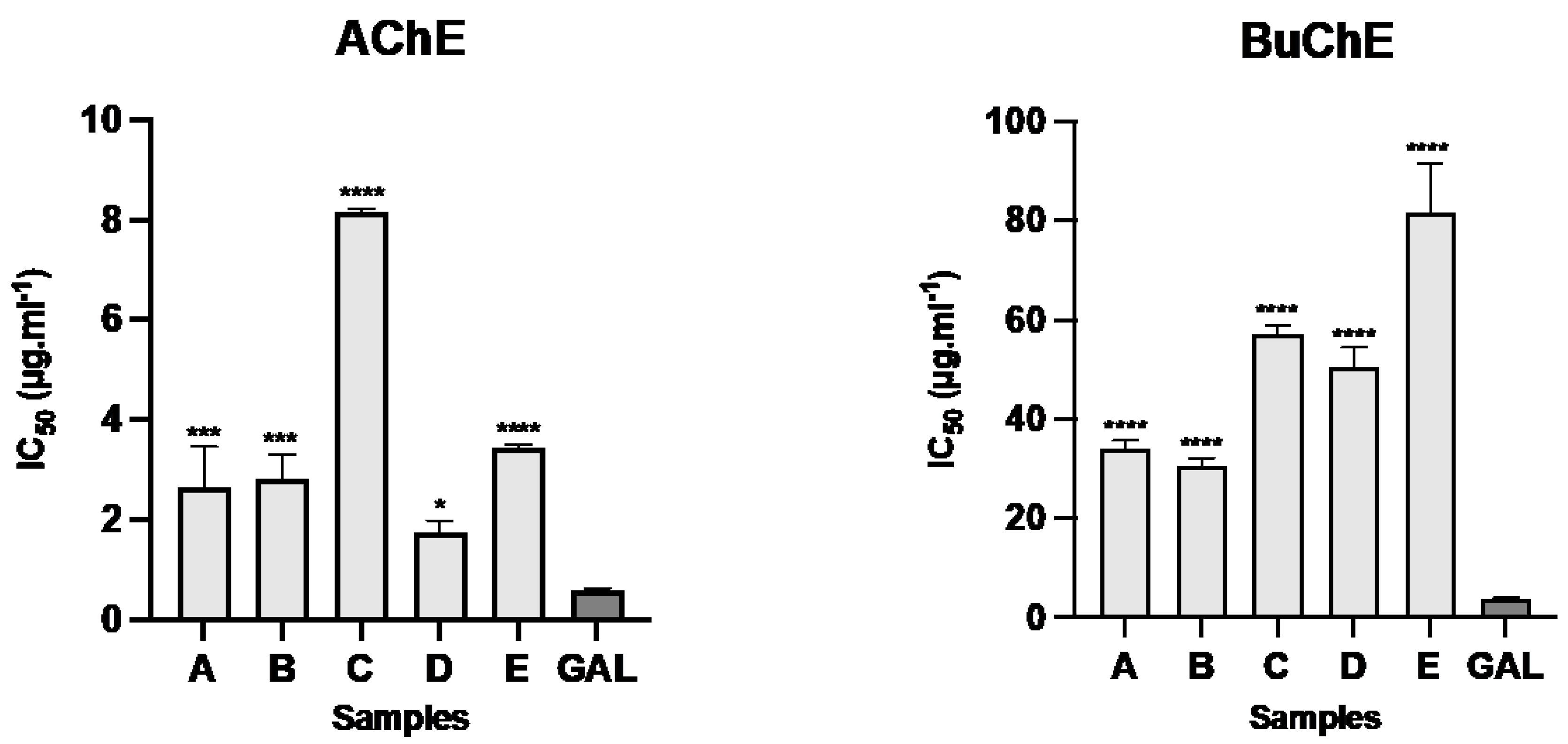
Figure 3.
Ligand conformations in the TcAChE enzyme obtained by molecular docking experiments.
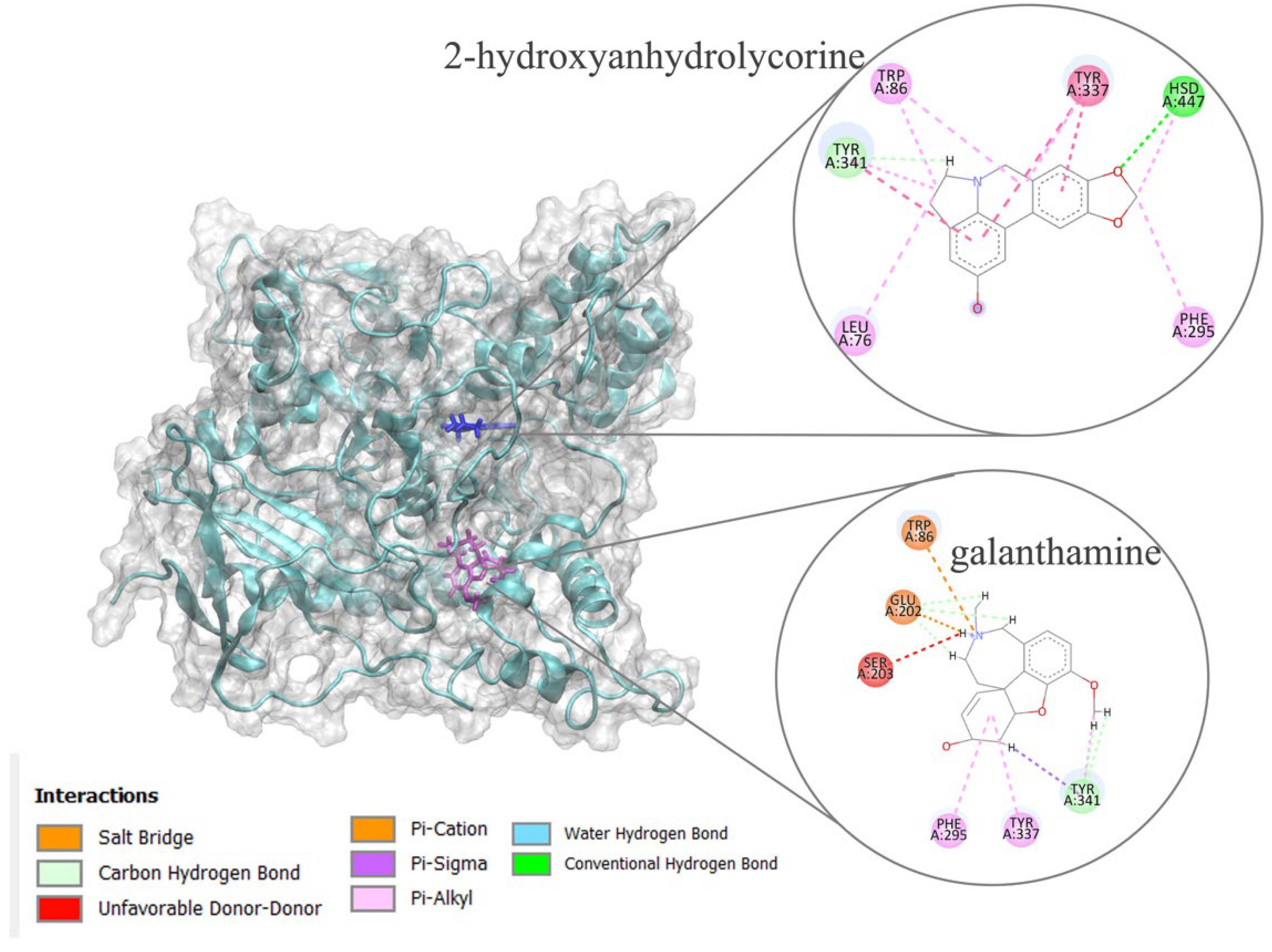
Figure 4.
RMSD analysis for 2-hydroxyanhydrolycorine-TcAChE and galanthamine- TcAChE complexes.
Figure 5.
Residue occurrence frequencies at 3 Å or less from the ligand within the ligand-TcAChE complex calculated by molecular dynamics procedures.
Figure 5.
Residue occurrence frequencies at 3 Å or less from the ligand within the ligand-TcAChE complex calculated by molecular dynamics procedures.
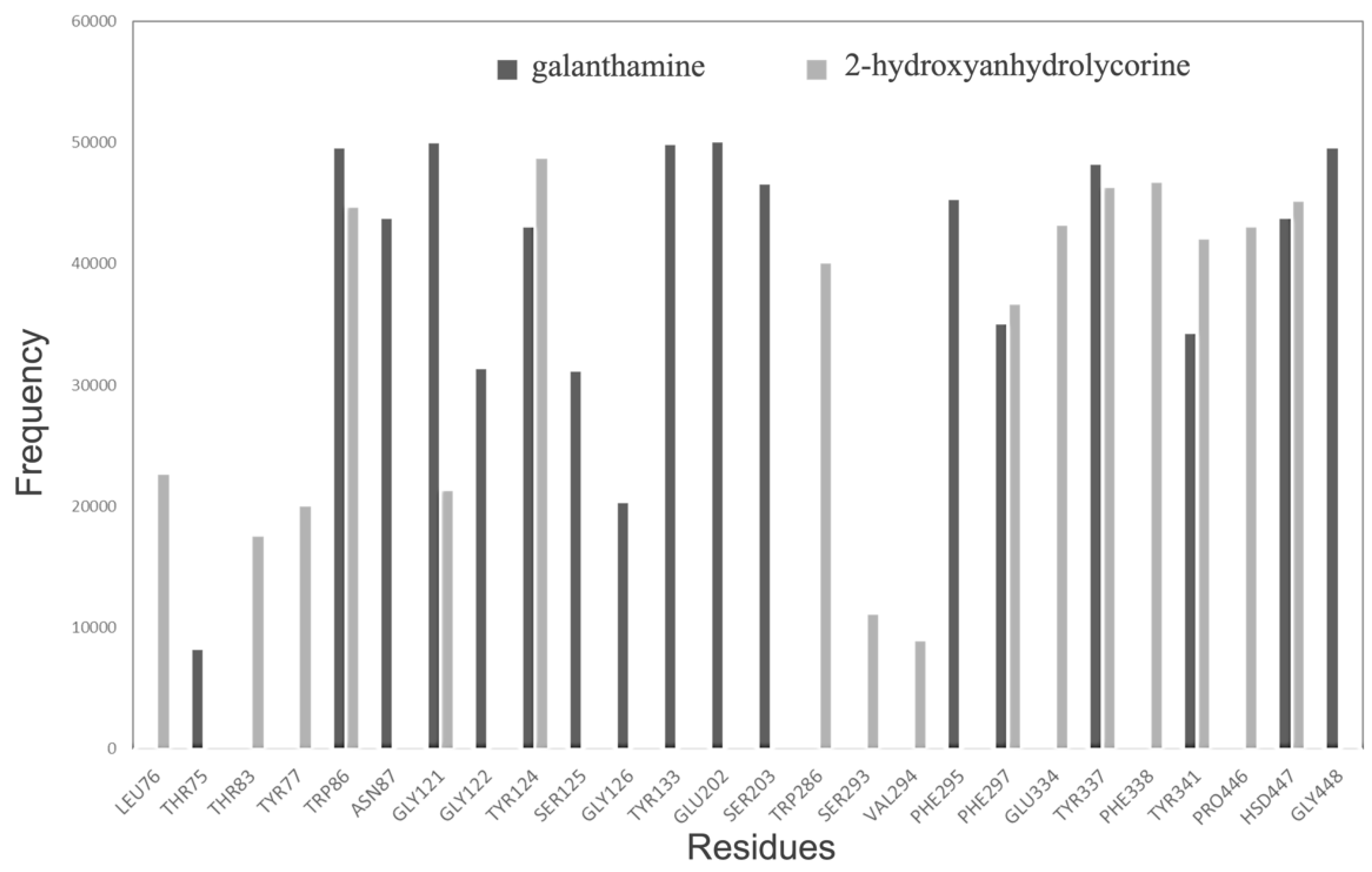
Figure 6.
Map of Ecuador showing the sites of the samples. A: Urceolina formosa (from Tungurahua province); B: Urceolina formosa (from Sucumbíos province); C: Urceolina ruthiana; D: Clinanthus incarnatus; E: Stenomesson aurantiacum. Map source: Google Earth.
Figure 6.
Map of Ecuador showing the sites of the samples. A: Urceolina formosa (from Tungurahua province); B: Urceolina formosa (from Sucumbíos province); C: Urceolina ruthiana; D: Clinanthus incarnatus; E: Stenomesson aurantiacum. Map source: Google Earth.
Figure 7.
Flowering Amaryllidaceae species collected in Ecuador. A, B: Urceolina formosa; C: Urceolina ruthiana; D: Clinanthus incarnatus; E: Stenomesson aurantiacum. Pictures source: Nora H. Oleas.
Figure 7.
Flowering Amaryllidaceae species collected in Ecuador. A, B: Urceolina formosa; C: Urceolina ruthiana; D: Clinanthus incarnatus; E: Stenomesson aurantiacum. Pictures source: Nora H. Oleas.


Table 1.
Alkaloids identified in Amaryllidaceae species collected in Ecuador by GC-MS. Values are expressed as mg GAL·g-1 AE.
Table 1.
Alkaloids identified in Amaryllidaceae species collected in Ecuador by GC-MS. Values are expressed as mg GAL·g-1 AE.
| Alkaloid | RI | [M+] | BP | A | B | C | D | E |
|---|---|---|---|---|---|---|---|---|
| trisphaeridine (1) | 2323.1 | 223 | 223 | - | - | - | - | 8.1 |
| galanthamine (2) | 2386.1 | 287 | 286 | 46.2 | 28.2 | - | - | 74.1 |
| sanguinine (3) | 2452.4 | 273 | 273 | - | - | - | - | 7.8 |
| unidentified (galanthamine-type)1 (4) | 2474.4 | 287 | 286 | - | - | - | - | 24.0 |
| galanthindole (5) | 2535.0 | 281 | 281 | - | - | - | - | 15.8 |
| 8-O-demethylmaritidine (6) | 2539.9 | 273 | 273 | - | - | - | - | 6.1 |
| anhydrolycorine (7) | 2543.6 | 251 | 250 | - | - | - | 24.5 | - |
| augustine (8) | 2576.1 | 301 | 301 | - | - | - | - | 45.4 |
| kirkine (9) | 2590.1 | 273 | 252 | - | - | - | - | 23.1 |
| haemanthamine (10) | 2611.5 | 301 | 272 | 172.5 | 37.2 | 101.4 | - | - |
| tazettine (11) | 2620.4 | 331 | 247 | - | - | 32.6 | - | - |
| 11,12-dehydroanhydrolycorine (12) | 2644.4 | 249 | 248 | - | - | - | - | 181.6 |
| 11-hydroxyvittatine (13) | 2679.4 | 287 | 258 | 16.9 | 9.8 | - | - | 108.1 |
| unidentified (pretazettine-type)1 (14) | 2697.3 | 331 | 261 | - | - | - | - | 8.2 |
| lycorine (15) | 2716.5 | 287 | 226 | 251.6 | 273.9 | 168.1 | 149.9 | - |
| 2-hydroxyanhydrolycorine 2 (16) | 2891.6 | 267 | 266 | - | - | - | 60.2 | - |
| Total | 487.2 | 349.1 | 302.1 | 234.6 | 502.3 |
1 proposed alkaloid-type according to fragmentation pattern; 2 proposed alkaloid structure according to fragmentation pattern by [23]; A: Urceolina formosa (from Tungurahua province); B: Urceolina formosa (from Sucumbíos province); C: Urceolina ruthiana; D: Clinanthus incarnatus; E: Stenomesson aurantiacum.
Table 2.
Free energy calculations of ligand-TcAChE complexes calculated through molecular dynamics and molecular docking procedures.
Table 2.
Free energy calculations of ligand-TcAChE complexes calculated through molecular dynamics and molecular docking procedures.
| Substrate | MM-GBSA (kcal·mol-1)* |
Molecular Docking (kcal·mol-1) |
|---|---|---|
| 2-hydroxyanhydrolycorine | -23.695 (2.106) | -8.94 |
| galanthamine | -36.710 (2.842) | -9.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



