
Submitted:
08 April 2024
Posted:
09 April 2024
You are already at the latest version
Abstract
Fiber-rich green river Aonori alga is mass-cultivated in Japan; it has been reported to reduce inflammation and improve metabolic health in humans. Peroxisome proliferation-activated receptor (PPARs) are associated with metabolic disorders such as obesity and type 2 diabetes (T2DM). We evaluated the effects of supplementation with NBF2, a formula made of algal and junos citrus biomaterials, on metabolic markers in obese and diabetic rats. An experimental study was performed using 18 obese and hyperglycemic Otsuka Long-Evans Tokushima Fatty (OLETF) rats randomly assigned to high dose NBF2 (20mg/kg; n=6), low dose (10mg/kg; n=6) NBF2 drink for 6 weeks, whereas OLETF control rats received similar amount of tap water (vehicle; n=6). LETO rats were used as normal controls (n=8). Biospecimens were taken for ELISA and real-time PCR. Both high dose and low dose NBF2 increased brown adipose tissue (BAT) mass and adiponectin (APN), improved insulin sensitivity, and significantly improved glycemic and lipid (TG, LDL-c) profiles in NBF2-supplemented OLETF rats after a 6-week supplementation with NBF2 (vs. OLETF controls), as well as LDL-c (p<0.05). During the glucose tolerance test (OGTT), both NBF2 groups and LETO rats had normal glycemia by 90-minute time-point (vs. OLETF controls; p<0.01). Furthermore, intake of NBF2 up-regulated PPARa/g-mRNA and Sirt2-mRNA gene expressions in BAT, and improved blood pressure profile in OLETF rats. Findings from this research suggest that intake of NBF2 formula induces dual PPAR-a/g and Sirt2 activation, providing insights on beneficial prophylactic and therapeutic effects of this NB2 formula on dyslipidemia and hyperglycemia in T2DM.
Keywords:
Algal formula
; Dyslipidemia
; Obese rat
; Type 2 diabetes
; Peroxisome proliferation-activated receptor
Introduction
Peroxisome proliferation-activated receptor (PPARs) are associated with metabolic disorders such as obesity and type 2 diabetes (T2DM)1). T2DM is a metabolic disease characterized by an inadequate pancreatic beta cells response to the progressive insulin resistance. Over the last decade, T2DM management has undergone a major conceptual change [1,2,3]. In T2DM, the loss of insulin function is closely related to APN deficiency. This suggests that replenishment of APN may reverse T2DM and improve the metabolic health. Discovered in the 1990′s, APN has been reported to stimulate fatty acids oxidation, inhibit glucose production in the liver and improve insulin sensitivity, resulting in an improvement in energy homeostasis [4]. Insulin sensitizing effect of adiponectin is reported to be mediated by the activation of peroxisome proliferator-activated receptor (PPAR)−α [5,6]. Furthermore, in addition to its function in relation to glucose and lipid metabolism, the interest of researchers in the field of cardiometabolic health for APN is also due to its role in tissue regeneration [7,8].
Endogenous upregulation of APN production to correct its deficiency and the restoration of the function of cells involved in energy homeostasis have motivated the APN modulators research. In the literature, a number of APN modulators with beneficial metabolic health effects have been reported. Radix astragali is reported to induce endogenous increase in APN in experimental studies [9,10]. Omega 3 poly-unsaturated fatty acids are also reported to modulate APN and improve metabolic biomarkers in high-risk subjects and T2DM patients [11]; however, optimal APN level that can reverse the course of metabolic disorders is almost out of reach. Furthermore, we have discovered APN modulating formulas (NBF) containing bio-materials from an edible marine algal species mass-cultivated in Japan, known as Sujiaonori or Aosanori, with beneficial metabolic health effects [12,13]. This study evaluated the effects of NBF2 formula composed of Aonori alga and junos citrus peel-derived biomaterials on metabolic health markers in obesity and T2DM model rats.
Methods
Animals and Intervention
The experimental protocol, as well as animal care procedures, were carried out according to the guidelines for the care and use of animals of Experimental Animal Center of Kagawa University (ethical approval number: 20656-1), Japan. Six-week-old male Otsuka Long-Evans Tokushima Fatty (OLETF) and genetic control Long-Evans Tokushima Otsuka (LETO) rats were purchased from Japan SLC Inc. (Shizuoka, Japan) and maintained under a controlled temperature (24 ± 2 °C) and humidity (55 ± 5%), with a 12 h light/dark cycle, at the Animal Center of Kagawa University in Japan. Rats were fed with MF chow and had access to tap water ad libitum. They were used when they reached 10 weeks of age. It has been reported that OLETF rats exhibit metabolic syndrome and prediabetes from the age of 10 weeks of age and, later on, the type-2 diabetic phase.
NBF2-based drink was a gift from Kochi Ice Co.Ltd. (Kochi, Japan), a food processing factory located in Kochi prefecture, Japan. It is food-based formula containing 60% of ulvan-rich dried alga powder and 40% of dried junos Tanaka citrus peel poweder. The chemical composition of the two biomaterials that make NBF2 formula is as follows: Aosanori algal biomaterial used in this study is rich in fiber (~65%), whose main bioactive component is ulvan [12], protein (9~14%), omega-3 and omega-6 fatty acids (10.4~10.9%), and small amounts of flavonoids, carotenoids and minerals; junos citrus peel biomaterial is rich in limonene (68~70%), other terpenoids such as γ-terpinene (11.4~12.5%), β−phellandrene (4.6~5.4%), myrcene (3.0~3.2%), α−pinene (2.3~21.7%) and a few amounts of aromatic compounds [14,15].
Experimental Protocol
In this experiment, a total of 26 rats were used, including 18 OLETF and 8 LETO (normal) rats. All eight LETO were used as normal controls; whereas prediabetic OLETF rats were randomly divided into 3 groups at baseline (10 weeks of age) based on body weight and fasting blood glucose (FBG):
(1) 2 mL H2O (vehicle)-treated OLETF or OLETF control (n = 6);
(2) 20 mg/kg/day) NBF2-treated OLETF rats (n = 6);
(3) 10 mg/kg/day NBF2-treated OLETF rats (n = 6).
NBF2-treated groups received 2 mL of drink containing the corresponding dose diluted in tap water, whereas OLETF control and normal control rats received the same amount of tap water daily for five days a week for six weeks. Food intakes were measured for 3 consecutive days every week; the average daily food intake was calculated for each animal, and total daily calorie intake was estimated based on energy provided by MF chow and NBF2-contained nutrients. Fasting blood glucose (FBG) was measured with the use of a freestyle glucose meter (Glucotest AI, Sanwa Biochemical Research Center, Japan) from the tip of the tail once a week. Rat body weight was also measured on a weekly basis.
Measurements
Oral glucose tolerance test (OGTT) assay was carried in duplicate within the last week of the experiment when rats were 16 weeks old, as described previously [16]. Briefly, rats were fasted for 12 h, and 0-min blood glucose was measured. Then, rats were each fed 2 g/kg of 50% glucose solution by gavage, and blood glucose was measured again at 30, 60 and 120-min. OGTT was performed in duplicate.
Blood pressure (BP) and heart rate (HR) were measured using an automatic device (BP-98A, Softron, Tokyo), following the tail-cuff method. Briefly, a photo-elective sensor, which is attached to the animal tail by a piece of rubber tubing, detects the pulse volume oscillations by cuff-pressure and pulse waves. The pulse waves are detected as systolic BP (SBP) and mean BP (MBP); these two values are used to calculate diastolic BP (DBP) automatically with the use of the following formula: DBP = (3 x MBP - SBP)/2. Homeostatic model assessment of insulin resistance (HOMA-IR) was calculated according to the following formula:
HOMA-IR = glycemia (mg/dL) x plasma insulin (UI/mL)/405.
Biospecimen Sampling, Histological Examination and Real-Time PCR
On the day of animal sacrifice, rats were fasted for 12 h and anesthetized; blood was drawn. Blood lipids were measured using a portable lipid analyzer MLA-1 (Changsha Zealson Biotech Co.Ltd., Hunan, China). Plasma adiponectin and insulin measurements were performed by enzyme-linked immunosorbant assay (ELISA; SRL biochemical company, Kobe, Japan). Brown adipose tissue (BAT) was collected at the scapula area, whereas white adipose tissue (WAT) was collected from the mesenteric, retroperitoneal and epididymal spaces. All necessary organs taken from rats were weighted and stored at −80 °C prior to histological examination. The latter was performed by a pathologist. Visceral adipose tissue and liver specimen were formalin-fixed and paraffin-embedded. They were stained with hematoxylin-eosin (HE), prior to observation of the specimen with all-in-one fluorescence microscope BZ-X800.
Collected adipose tissues were snap frozen in liquid nitrogen, then stored at −80 °C until processing for RNA extraction. Real-time reverse- transcription PCR (RT-PCR) was performed at the laboratory unit of the department of Environmental Medicine, Kochi University Medical School, Japan. RNA was isolated from WAT, FAT and liver tissue by the phenol-chloroform extraction method and cDNA prepared. Messenger RNA (mRNA) expressions of PPAR−α, PPAR−γ, leptin, APN were analyzed in adipose tissue, whereas that of three markers of lipid metabolism, namely fatty acid synthase (FAS), sirtuin2 (Sirt2), ATPase Sarcoplasmic/Endoplasmic Reticulum Ca2+ Transporting 2 (ATP2a2) were analyzed in liver tissue by RT-PCR using ABI Prism 7000 with Power SYBR Green PCR Master Mix (Invitrogen, Japan) and primer sequences from Eurofins genomics (Japan).Group comparisons for mRNA gene expressions of different biomarkers were performed using the relative quantification of genes.
Ethical Consideration and Statistical Analysis of Data
Ethical approval was obtained from the “Ethical review board the Experimental Animal Center of Kagawa University” (ethical approval number: 20656-1). Animal research procedures were performed according to internationally guidelines for the care and use of laboratory animals. All values related to continuous variables are presented as mean +/- standard deviation (SD). For data related to the experimental study, two tailed t test was employed to compare two groups for normally distributed outcome variables. To compare more than two groups, one-way analysis of variance (ANOVA), followed by Mann-Whitney test or a post-hoc Tukey’s test was used when appropriate. The significance level was set at p-value less than 0.05. Stata statistical software version 15 was used for all statistical analyses (SataCorp, College Station, TX, USA).
Results
Obesity and Diabetic Hyperglycemia Establishment in OLETF Rat Model and Effects of Daily Intake of NBF2 on Body Weight (BW) and Fasting Blood Glucose (FBG)
OLETF rats get obese spontaneously and develop hyperglycemia by 10 weeks of age. BW and FBG were measured in rats on weekly basis; glucose tolerance test (OGTT) was performed in duplicate at 16 weeks of age. Prior to rat sacrifice, at week17, blood pressure was measured in rats using an automatic device (BP-98A); afterwards, blood and organ (adipose tissue, liver) samples were taken for ELISA, real time PCR assays, and histopathological examination (Figure 1).
Figure 1 shows the procedures used to implement the study as well as measurements performed to obtain clinical, laboratory (ELISA, RT-PCR assays) and pathological data
It was observed that treatment with NBF2 significantly reduced weight gain in OLETF rats that received both high and low dose of NBF2, as compared with vehicle-treated OLETF controls (p < 0.05) (Figure 2A). Additionally, both low and high dose NBF2-treated OLETF rats had a reduced fasting blood glucose (FBG) as compared with OLETF controls (p < 0.01), which was normalized within the third week of treatment (Figure 2B). Furthermore, it was observed that high dose NBF2 group had higher daily calorie intake as compared with OLETF controls (p < 0.05) (not shown).
-Legend: *, p-value less than 0.05; #, p-value less than 0.01 (by one-way ANOVA); OLETF, Otsuka Long Evans Tokushima Fatty rats; LETO, Long Evans Tokushima Otsuka rat.
The Figure 2 shows reduced weight in both low dose and high dose NBF2-treated OLETF rat groups (vs. vehicle-treated OLETF controls; p < 0.05) (2A), reduced fasting blood glucose (FBG) level in both low dose (p < 0.05) and high dose (p < 0.01) NBF2-treated OLETF rats (vs. OLETF control rats) across the experiment (2B) and during glucose tolerance test (2C).
OGTT was performed by administering 2 g of glucose per kg of body weight (0.002 g of glucose/1 g body weight); FBG was measured at baseline (T0), 15 min (T15), 30 min (T30), 60 min (T60) and 120 min (T120). The decrease of FBG was greater in normal control rats, high dose NBF2 (p < 0.01) as well as low dose (p < 0.05) NBF2-treated OLETF rat groups, as compared with OLETF controls (Figure 2C).
3.2. Effects of NBF2 on Lipid and Blood Pressure Profiles, and Plasma Adiponectin, Insulin and Insulin Resistance Index (HOMA-IR)
Rat plasma lipid and blood pressure profiles are shown in Figure 3. High dose NBF2 showed a better lipid lowering effect for total cholesterol (Figure 3A), LDL-c (p < 0.05) (Figure 3B), and triglycerides (TG) (p < 0.01) (Figure 3C), as compared with the vehicle in OLETF control rats. Similar trend was observed for low dose NBF2 for total cholesterol, TG and LDL-c (p < 0.05). HDL-c level was markedly higher in high dose NBF2-treated rat group as compared with vehicle-treated OLETF controls (p < 0.05). Though low dose NBF2 rat group and normal (LETO) rat group had higher plasma HDL-c, the differences did not reach statistical significance level (Figure 3D). Additionally, markedly reduced blood pressure was observed in high and low dose NBF2-treated OLETF rats for both DBP and MBP, as compared with vehicle-treated OLETF controls (p<0.05) (Figure 3E-F-G).
-Legend: *, p-value less than 0.05; #, p-value less than 0.01 (by one-way ANOVA); OLETF, Otsuka Long Evans Tokushima Fatty rats; LETO, Long Evans Tokushima Otsuka rat; T-chol, total cholesterol; LDL-c, low-density lipoprotein cholesterol; HDL-c, high-density lipoprotein cholesterol; TG, triglycerides.Figure 3shows significant reduction of blood T-chol, LDL-c in high (p < 0.01) and low dose NBF2 (p < 0.05) treated OLETF rats (3A), reduction of TG (p < 0.05) in low and high dose NBF2-treated rat groups (3B-C), and significant increase of blood HDL-c in high dose NBF2-treated rats (3D). Furthermore, marked reduction of diastolic blood pressure (DBP) and mean BP (MBP) in NBF2-supplemented OLETF rats (p < 0.05), and in normal rats (p < 0.01) vs. OLETF controls (p < 0.05) (3E-G).
Additionally, ELISA assay performed using plasma from blood samples drawn at baseline and at the end of study showed markedly increased APN level in NBF2-treated OLETF rats as compared with OLETF controls (p < 0.05), whereas a light but not significant change was noted in plasma insulin level (Figure 4A-B). Furthermore, a significant decrease in HOMA-IR value was observed in the group of OLETF rats supplemented with NBF2 (high and low dose-treated rats combined), as compared to vehicle-treated OLETF rats (−17% versus −3% of change, respectively; p < 0.05) (Figure 4C).
-Legend:*, p < 0.05; IU, international unit; APN, adiponectin; NBF, Ngatu Bio Formula; HOMA-IR, homeostasis model assessment for insulin resistance.
Figure 4 shows a marked increase of plasma APN level in high dose NBF2-treated OLETF rats (vs. OLETF controls; p < 0.05), and a significant reduction of HOMA-IR value in NBF2-treated OLETF rats as compared with vehicle-treated OLETF controls (p < 0.05).
3.3. Messenger RNA (mRNA) Gene Expression of Biomarkers (White/Brown Adipose Tissue, Liver Biopsies) and Histopathological Feature of Biospecimens
The real time PCR assays were performed to determine the mRNA gene expression levels of metabolic biomarkers. “Table 1” presents the list of primers of rat biomarkers and the corresponding mRNA gene sequences. Leptin and fatty acid synthase (FASN) are linked to the development of obesity and related metabolic disorders. We found that rats treated with NBF2 had significantly reduced leptin-mRNA expression in brown (BAT) and white adipose tissue (WAT), as well as in liver specimens as compared with vehicle-treated OLETF controls (p < 0.05) (Figure 5A-B-C). Similarly, NBF2-treated rats had reduced FASN-mRNA gene expression in both BAT and WAT specimen (Figure 5D-E), suggesting an inhibitory effect on fat accumulation.
Unlike FASN and leptin, sirtuin 2 (Sirt2) and peroxisome proliferator- activated receptors alpha and gamma (PPAR-α/γ) are known to improve obesity-associated metabolic disorders and considered as therapeutic targets for diseases such as T2DM. We observed that treatment with NBF2 caused a marked increase in Sirt2-mRNA expression in the rat BAT (p < 0.01) and WAT (p < 0.05), as well as an increase of PPAR−γ and PPAR−α mRNA expressions in OLETF rat BAT as compared with vehicle in control rats (Figure 5G-H).
Similar finding was observed in low dose NBF2-treated rats for BAT APN-mRNA and WAT APN-mRNA. ATPase sarcoplasmic/endoplasmic reticulum Ca2+, also known as ATPase 2a2 or SERCA2, is an enzyme that plays an important metabolic role as glucose transport regulator; it is downregulated in adipocytes in the adipocytes of diabetic animals and humans. We found that BAT ATP2a2-mRNAexpression was up-regulated in NBF2 treated rats (Figure 5M-N-O) (vs. OLETF control; p < 0.05). Consequently, dual PPARα/PPARγ and Sirt2 modulatory effects of NBF2 observed in this study suggests an improvement of lipid and glucose metabolism.
Figure 5.
Messenger RNA (mRNA) gene expression of biomarkers in white/brown adipose tissue and liver biopsies.
Figure 5.
Messenger RNA (mRNA) gene expression of biomarkers in white/brown adipose tissue and liver biopsies.
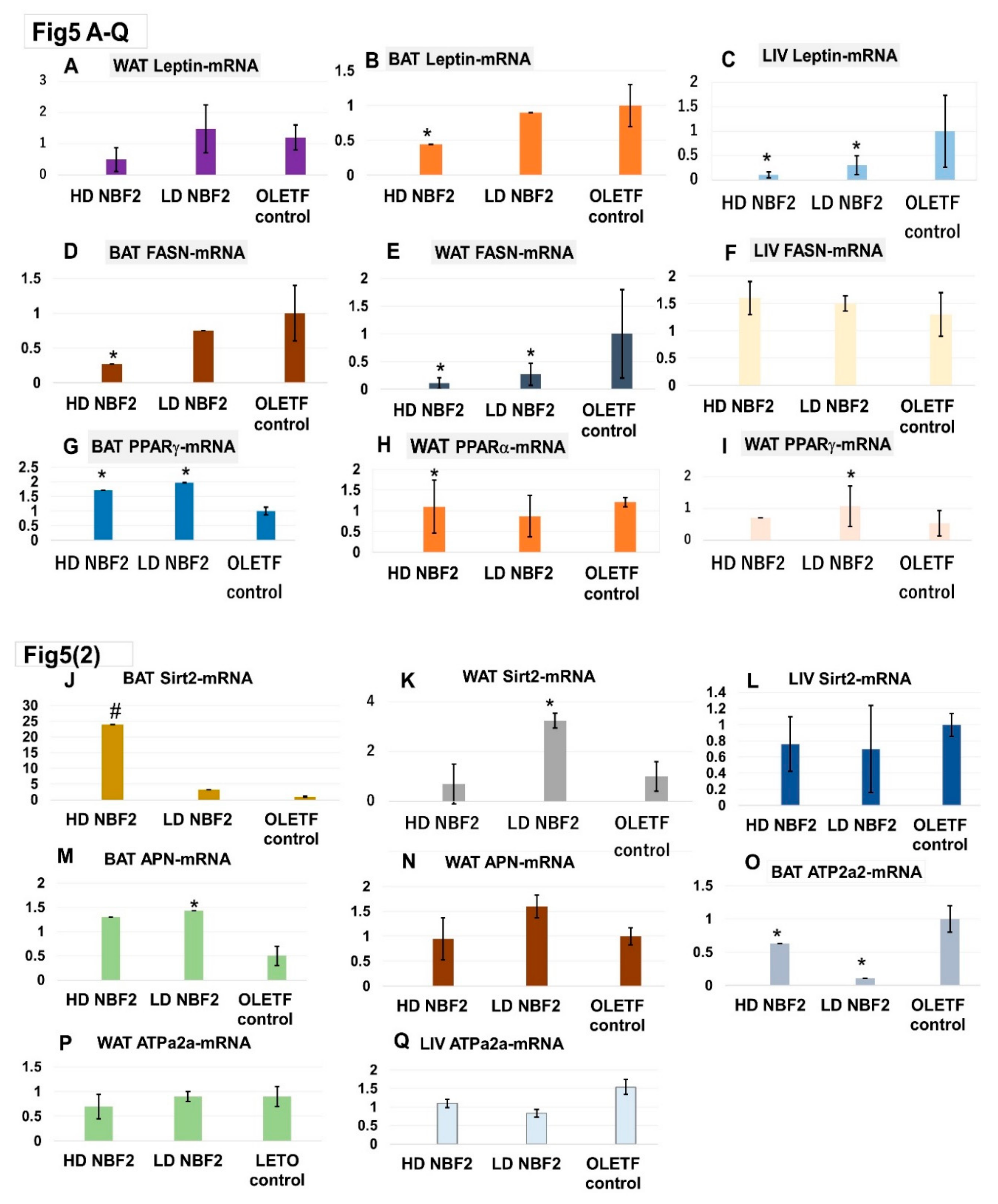
-Legend: *, p-value less than 0.05; #, p-value less than 0.01 (vs. OLET control; by Student t test);OLETF, Otsuka Long Evans Tokushima Fatty rats; LETO, Long Evans Tokushima Otsuka rat; mRNA, messenger ribonucleic acid; BAT, brown adipose tissue; WAT, white adipose tissue; LIV, liver tissue; APN, adiponectin; FASN, fatty acid synthase; PPAR, peroxisome proliferator-activated receptor; Sirt2; sirtuin 2; ATPa2a or SERCA2, ATPase sarcoplasmic/ endoplasmic reticulum Ca2+.Figure 4shows downregulation of leptin-mRNA gene expression in BAT and liver (4A, 4C) and FASN-mRNA in BAT and WAT (4D, 4E) by treatment with NBF2, and up-regulation of PPARγ-mRNA and PPARα-mRNA gene expression in BAT and WAT of OLET rats by NBF2 treatment (4G, 4H). Additionally, up-regulation of Sirt2-mRNA gene expression in BAT and WAT (4J, 4K) and reduction of ATP2a2-mRNA gene expression (4O) were observed in NBF2-treated OLETF rats vs. vehicle-treated control rats.
Histopathological examination of hematoxylin-eosin (HE) stained paraffin-embedded BAT, WAT and liver sections, results were as follows: large adipocytes were mostly observed in WAT specimen from OLETF control rats (Suppl. Figure 1C-D) than in specimens from normal rats (Suppl. Figure 1A-B) and NBF2-treated animals (Suppl. Figure 1E-F). Regarding BAT specimens, the presence of smaller adipocytes mixed with large adipocytes observed in OLETF controls (Suppl. Figure 1I-J), whereas larger ones were predominant in BAT from NBF2-treated OLET (Suppl. Figure 1K-L) and LETO rats (Suppl. Figure 1G-H). Images of blood sinusoids dilation, hepatic steatosis with lipid droplets were mostly observed in liver specimen from OLETF control rats (Suppl. Figure 1O-P), whereas these changes were either reduced and inexistent in liver specimens from NBF2-treated OLETF (Suppl. Figure 1Q-R) and normal rats (Suppl. Figure 1M-N).
Discussion
The present study explored the beneficial effects of a low dose and high or normal dose of edible marine alga and citrus-derived formula, NBF2, on glucose and lipid metabolism in obese and diabetic OLETF rats. NBF2 treatment induced an increase in circulating APN, as shown in results from ELISA assay. More interestingly, NBF2 treatment also resulted in increased APN-mRNA gene expression, particularly in rats that received a low dose of this biomaterial. Additionally, the RT-PCR assay of rat adipose tissues showed an improvement of PPAR-α/γ mRNA gene expressions, which corroborates with the improved plasma adiponectin observed in our study.
In fact, we previously reported that marine alga-derived biomaterial, which is the main component of NBF2, induced a 2.5 to 3-fold increase of APN in apparently health adult Japanese in a clinical investigation [12]. On the other hand, junos citrus peel, another component of NBF2, is mainly composed of limonene. We previously reported on the anti-inflammatory property of limonene [14]. Thus, the formula used in this study can be considered a potent PPARs modulator, APN modulator and anti-inflammatory agent that exerts beneficial effects on glucose and lipid metabolism.
In our study, NBF2 intake prevented weight gain in OLETF rats than the vehicle. Moreover, NBF2 could improve insulin sensitivity, and also normalize blood glucose in OLETF rats to a level similar to that of lean LETO rats across the experiment. The latter finding can be explained by the increased brown fat mass, the relatively low FASN-mRNA and enhanced APN production in NBF2-treated OLETF rats. APN is known to ameliorate insulin resistance and glucose intolerance. By restoring insulin sensitivity through upregulation of APN production, NBF2 could reverse hyperglycemia. Recently, we have reported that, in addition to its anti-inflammatory effect, NBF-contained algal biomaterial has a potential to improve glycemic profile in individuals with high cardiometabolic risk, and in diabetic patients as well13). The same finding was observed in the case of diabetic patient described in this report, whose FPG was controlled after 8 weeks of NBF2 supplementation.
Our study also showed that NBF2 supplementation improved lipid and blood pressure profiles in OLETF rats, with high dose NBF2 exerting better effects particularly in regard to decreasing plasma TG, DBP, MBP and increasing HDL-c. The improved lipid profile, possibly intertwined by Sirt2 activation, may explain the slowing of weight gain and improved blood pressure profile in NBF2-treated OLETF rats. Recently, new research-based evidences suggested that, not only PPAR but also sirtuin activators can be utilized as novel therapeutic targets for metabolic disorders [15,16]. Sirt2, known as a NAD+-dependent deacetylase and which was activated by NBF2 in our study, plays a critical role in the regulation of lipid metabolic homeostasis and in sustaining liver integrity [17,18]. Furthermore, hepatic steatosis or white fat accumulation in the liver is one of the complications of obesity and T2DM. OLETF rats are known to naturally develop liver steatosis; this process was attenuated in rats treated with NBF2.
This study showed that NBF2 simultaneously activated PPAR−α and PPAR−γ, thus behaving like a dual PPAR agonist. Previously, it has been suggested that dual PPARα/γ agonists, which are reported to reduce hypertriglyceridemia and improve insulin sensitivity in T2DM patients with hyperlipidemia [19], can help to overcome the efficacy and safety limitations associated with other PPAR agonists such as thiazolidinediones and fibrates. Therefore NBF2, which acts as a dual PPAR agonist, modulates APN and improves glucose and lipid metabolism [20,21].
Despite the impressive health effects of NBF2 observed in the present study, there is some limitation. The effects of NBF2 on other metabolic markers such as glucagon-like protein-1 receptor (GLP-1R). Drugs that activate GLP-1R have recently emerged as effective therapeutic agents for T2DM, as they stimulate insulin secretion, causing the decrease in blood glucose [22]. We plan to include this metabolic marker in future investigations.
Conclusion
As a conclusion, the present research, which is related to one of newly discovered alga-based adiponectin modulating formulas, NBF2, showed that its intake caused a Sirt2 and dual PPAR activation, upregulated APN, and improved glucose and lipid metabolism in a rodent model of obesity and T2DM. These findings suggest that NBF2 may be beneficial in preventing and reversing hyperlipidemia and T2DM.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conception: NRN, AH, AR, AN. Acquisition, analysis of data: NRN, TH, KK, ME. Interpretation of data: KDT, LSN, NS. Drafting the manuscript: NRN, SA, KDT, ME. Final approval of the manuscript: all authors.
Funding
No external fund was received. This study was conducted with internal fund of the department of public health, Kagawa University Faculty of Medicine, Japan.
Data Availability Statement
The datasets used in the study are available from the corresponding authors.
Acknowledgments
We also thank the manager of Kochi ice company (Kochi, Japan), Mrs Yukimi Nakamura from Takamatsu Health Center, Japan, and Dr Joel Ngoie, president of FICE (Congo Diaspora Forum) international, for supporting this research project.
Conflicts of Interest
The authors declare they have no conflict of interest.
Abbreviations
APN, adiponectin; HOMA-IR, homeostasis model assessment of insulin resistance; NBF, ngatu bio formula; T2DM, type-2 diabetes mellitus; RT-PCR, real-time.
References
- Luo W, Guo Z, Wu M, Hao C, Hu X, Zhou Z, et al. Association of peroxisome proliferator-activated receptor α/δ/γ with obesity, and gene-gene interaction, in the Chinese Han population. J Epidemiol 2013, 23, 187–194.
- American Diabetes Association. Pharmacologic approaches to glycemic treatment: Standards of medical care in diabetes-2021. Diabetes care, 2021; 44, S111–S124.
- Lingvay, I. Obesity management as a primary treatment goal for type 2 diabetes: Time to reframe the conversation. The Lancet 2022, 399, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J Clin Investig. 2006, 116, 1784–92.
- Kersten S, Desvergne B, Wahli W. Roles of PPARs in health and disease. Nature 2000, 405, 421–424.
- Yamauchi T, Kamon J, Minokoshi Y, Ito Y, Waki H, Ychida S, et al. Adiponectin stimulates glucose utilization and fatty acid oxidation by activating AMP-activated protein kinase. Nat Med 2002, 8, 1288–1295.
- Karbowska J, Kochan Z. Role of adiponectin in the regulation of carbohydrate and lipid metabolism. J Physiol Pharmacol 2006, 57, 103–113, Available online: Role of adiponectin in the regulation of carbohydrate and lipid metabolism—PubMed (nih.gov).
- Fiaschi T, Magherini F, Gamberi T, Modesti PA, Modesti A. Adiponectin as a tissue regenerating hormone: More than a metabolic function. Cell Mol Life Sci 2014, 71, 1917–1925.
- Xu A, Wang H, Hoo RLC, Sweeney G, Vanhoutte PM, Wang Y, et al. Selective elevation of adiponectin production by the natural compounds derived from medicinal herb alleviates insulin resistance and glucose intolerance in obese mice. Endocrinol 2009, 150, 625–633.
- Hoo RLC, Wong JYL, Qiao CF, Xu A, Xu HX, Lam KSL. The effective fraction isolated from Radix astragali alleviates glucose intolerance, insulin resistance and hyperglyceridemia in diabetic db/db mice through its anti-inflammatory activity. Nutr Metab (Lond) 2010, 7, 67–79.
- O’Mahony LL, Matu J, Price OJ, Birch KM, Ajjan RA, Farrar D, et al. Omega-3 polyunsaturated fatty acids favourably modulate cardiometabolic biomarkers in type 2 diabetes: A meta-analysis and meta-regression of randomized controlled trials. Cardiovasc. Diabetol 2018, 17, 98–110.
- Ngatu RN, Ikeda M, Watanabe H, Tanaka M, Inoue M, Kanbara S, et al. Uncovering adiponectin replenishing property of Sujiaonori algal biomaterial in humans. Mar. Drugs 2017, 15, 32–41.
- Ngatu RN, Ikeda M, Tonduangu DK, Nangana SL, Hirao T. Modulatory effects of NBF1 algal-rich bioformula on adiponectin and c-reactive protein levels and its therapeutic prospects for metabolic syndrome and type 2 diabetes. Biomedicines 2022, 10, 272–2581.
- Hirota R, Ngatu-Nlandu R, Nakamura H, Songo HS, Sawamura M. Anti-inflammatory effects of limonene from yuzu (citrus junos Tanaka) essential oil on eosinophils. J Food Sci 2010, 75, H87–92.
- Dosoky NS, Setzer WN. Biological activities and safety of citrus spp. Essential oils. Int J Mol Sci 2018, 19, 1966–1991.
- Hossain A, Yamaguchi F, Hirose K, Matsunaga T, Sui L, Hirata Y, et al. Rare sugar d-psicose prevents progression and development of diabetes in T2DM model Otsuka Long-Evans Tokushima Fatty rats. Drug Des Devel Ther 2015, 9, 525–535.
- Watanabe H, Inata Y, Kimura K, Matsumoto M, Kaneko S, Kasuga M et al. Sirt2 facilitates hepatic glucose uptake by deacetylating glucokinase regulatory protein. Nat Commun 2018, 9, 30.
- Ren H, Hu F, Wang D, Kang X, feng X, Zhang L, et al. Sirtuin 2 prevents liver steatosis and metabolic disorders by deacetylation of hepatocyte nuclear factor 4a. Hepatology 2021, 74, 723–740.
- Park SY, Chung MJ, Son JY, Yun HH, Park JM, Yim JH, et al. The role of sirtuin2 in sustaining functional integrity of the liver. Life Sci 2021, 285, 119997–120004.
- Kim SH, Hong SH, Park YJ, Sung JH, Lee KW, Jung K, et al. MD001, a novel peroxisome proliferator-activated receptor α/γ agonist, improves glucose and lipid metabolism. Sci Rep 2019, 9, 1656–1667.
- Jain N, Bhansali S, Kurpad AV, Hawkins M, Sharma A, Kaur S, et al. Effect of a dual PPAR α/γ agonist on insulin sensitivity in patients of type 2 diabetes with hyperglycemia–randomized double-blind placebo-controlled trial. Sci Rep 2019, 9, 19017–19026.
- Gallwitz, B. Clinical perspectives on the use of the GIP/GLP-1 receptor agonist tirzepatide for the treatment of type-2 diabetes and obesity. Front Endocrinol (Lausanne) 2022, 13, 1004044. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental protocol.
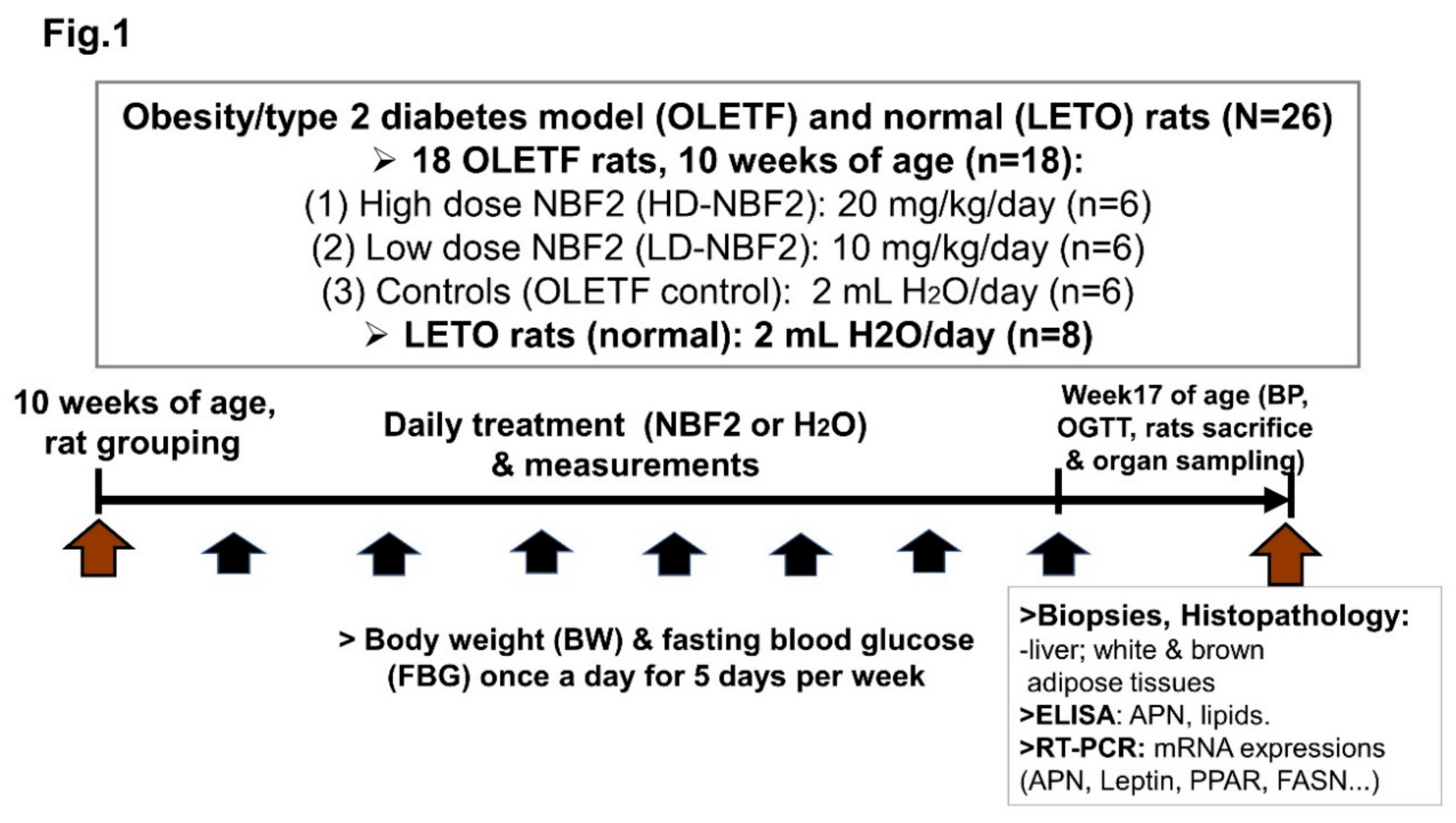
Figure 2.
A-B-C: Trend in rat body weight (A), fasting blood glucose across the study period (B) and blood glucose level during glucose tolerance test (oGTT) (C).
Figure 2.
A-B-C: Trend in rat body weight (A), fasting blood glucose across the study period (B) and blood glucose level during glucose tolerance test (oGTT) (C).
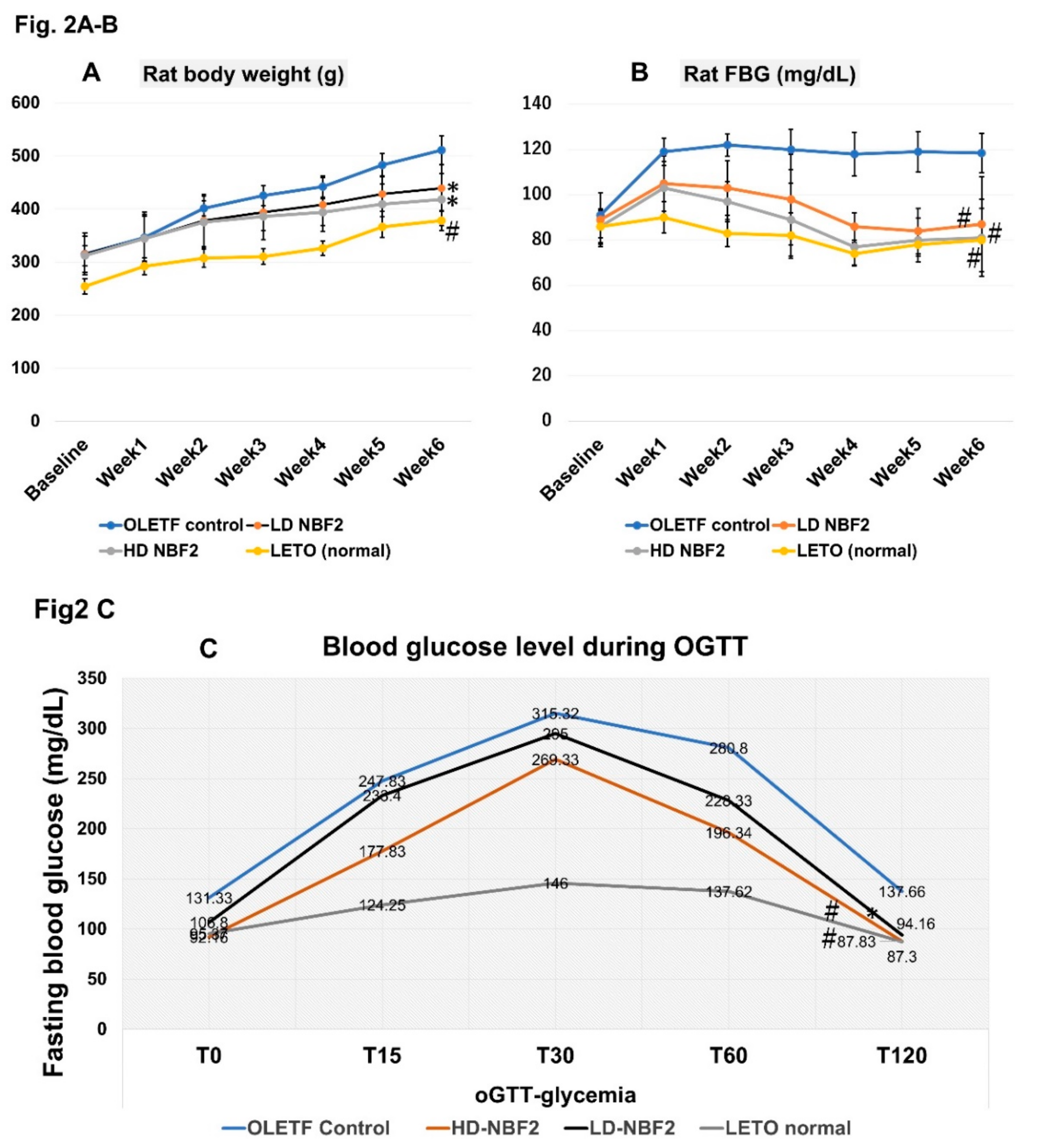
Figure 3.
A-B-C-D-E-F-G: Blood lipid and blood pressure profiles in rats.
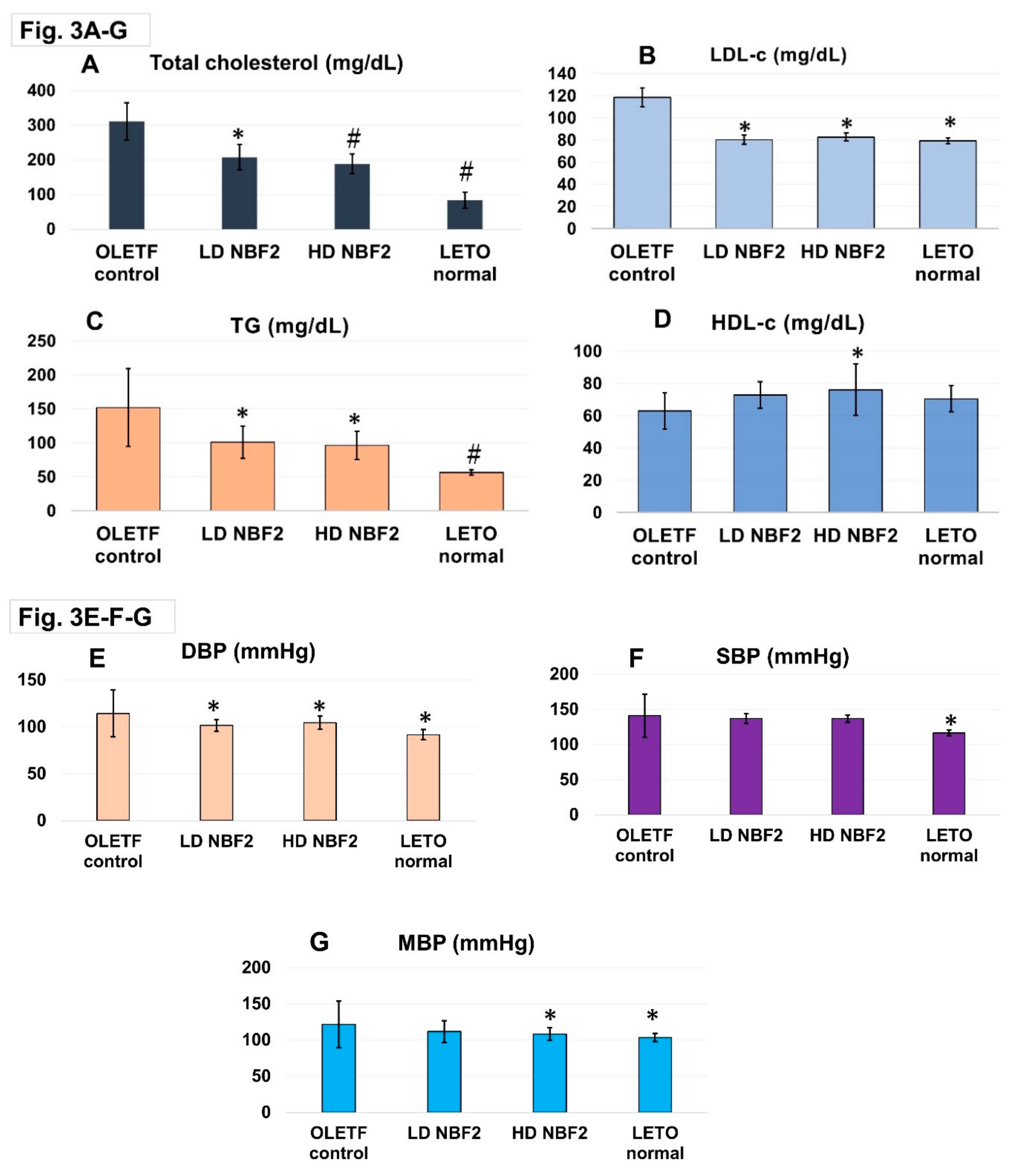
Figure 4.
A-B-C: Plasma adiponectin (A), insulin (B), and insulin resistance index (HOMA-IR) (C).
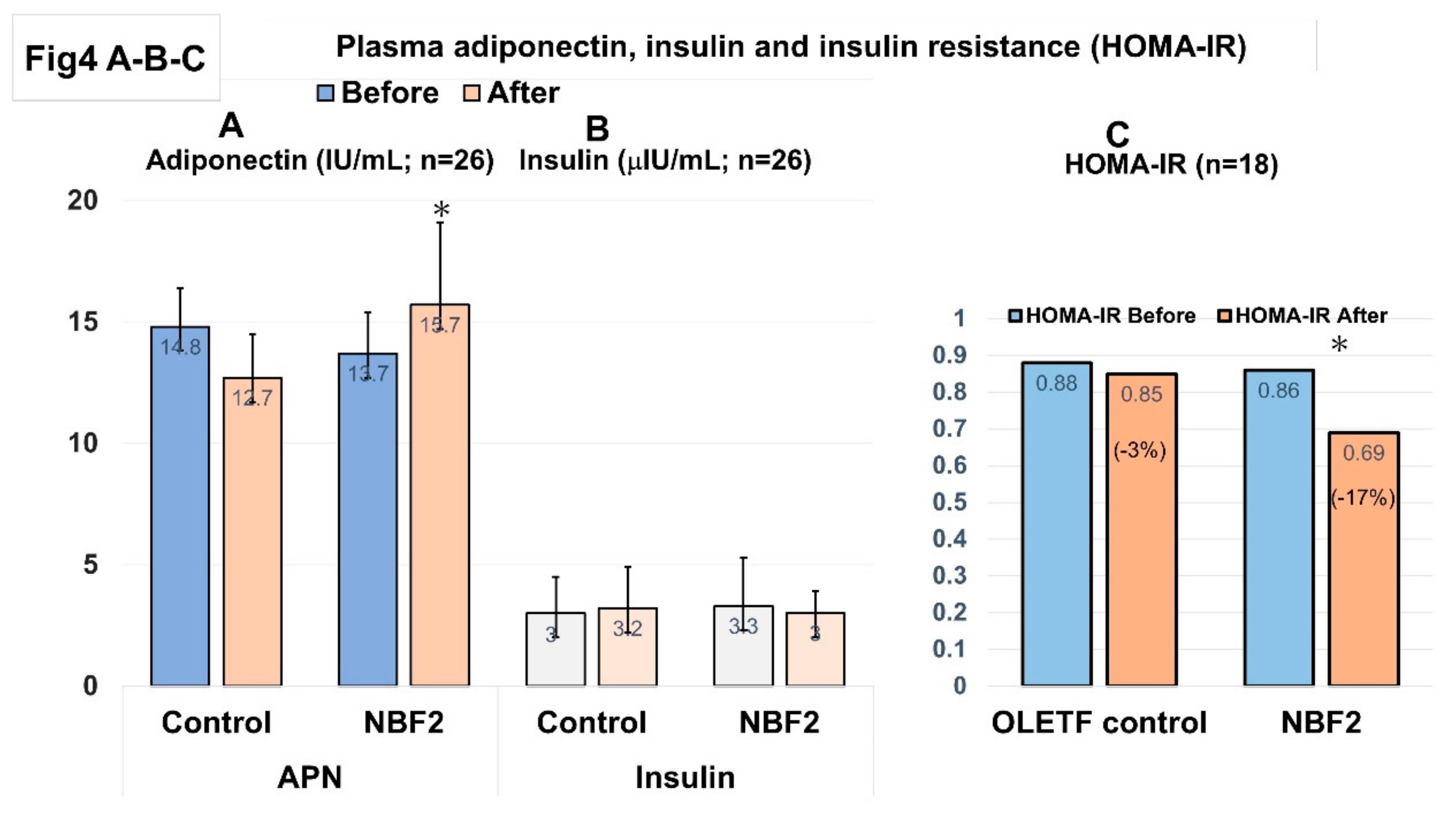

Table 1.
Statistical description of the CS parameters.
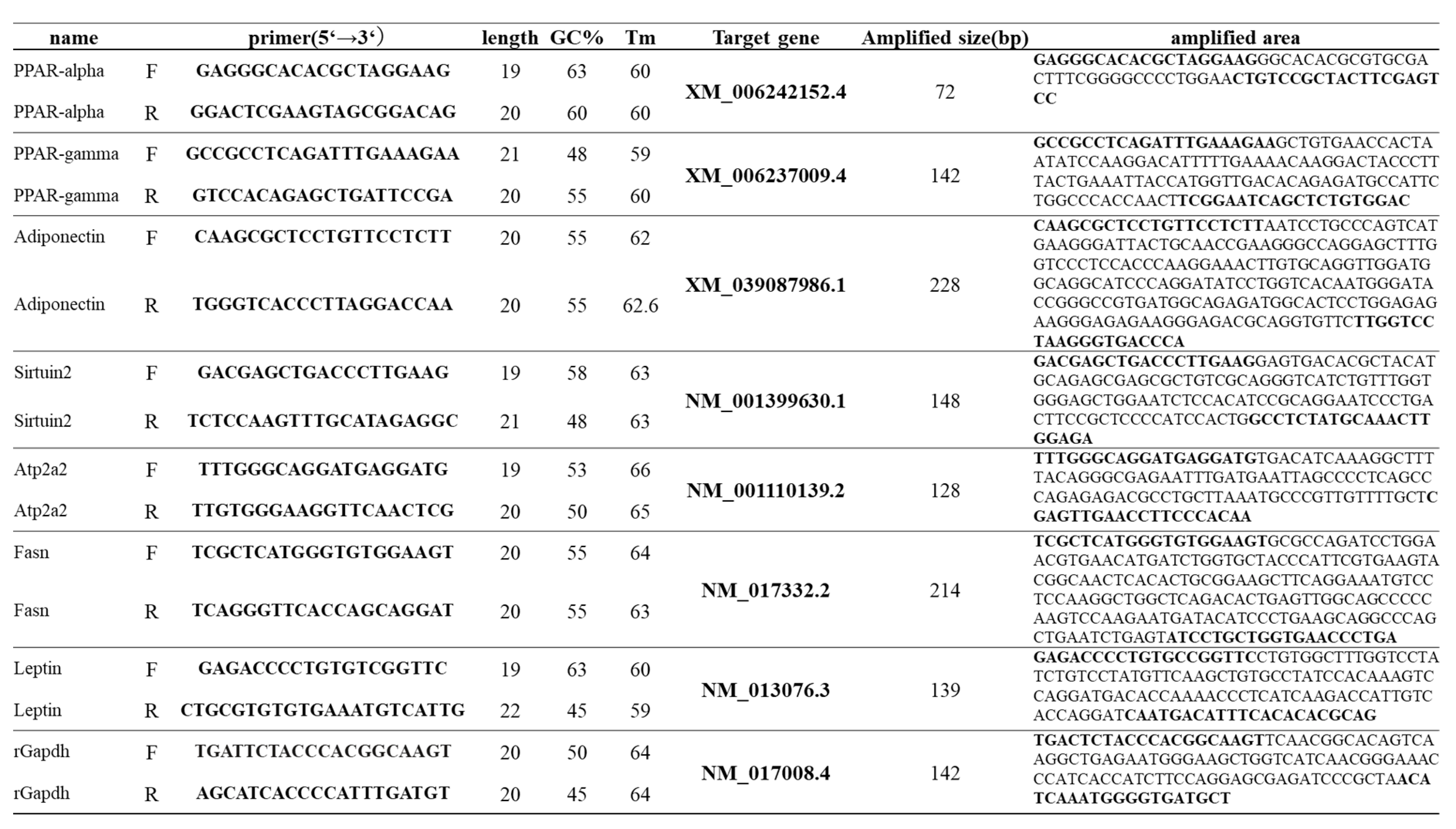 |
*Notes: PPAR, peroxisome proliferator-activated receptor; FASN, fatty acid synthase; ATP2a2, ATPase sarcoplasmic/endoplasmic reticulum Ca2+ transporting 2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



