
Submitted:
09 April 2024
Posted:
11 April 2024
You are already at the latest version
Abstract
Dirofilaria immitis and D. repens are the two most widespread and important species of mosquito-borne nematodes, posing a significant threat to veterinary health, particularly affecting canines and felines. While D. immitis causes cardiopulmonary dirofilariasis, D. repens causes subcutaneous infections in dogs and other carnivores. Despite extensive knowledge about the aforementioned parasites, little is known about their natural vectors in Serbia. Thus, the aim of this study was to: i) further map out Dirofilaria sp. hotspots in the Vojvodina Province, ii) detect positive mosquito species which can provide insights in how the nematodes spread and adapt to the environmental conditions, and iii) analyze the blood fed mosquitoes females of species found infected to identify the potential source of parasite infection. A total of 2,902 female mosquitoes were collected across 73 locations during 2021 and 2022. Molecular biology methods, based on conventional PCR, were used to analyze non-blood fed (2,521 specimens) and blood fed (381 specimens) mosquito females, in order to detect filarial nematode presence and identify blood meal sources respectively. When the parasite genome was detected, the amplicon (COI gene, 650 bp fragment) was sent for Sanger sequencing, further confirming the presence of nematodes and species assignation. D. immitis was detected in three Culex pipiens mosquitoes collected in XXXX (location and date), while Setaria tundra was detected in Aedes vexans and Aedes caspius. This work adds two new locations where D. immitis occurs in Vojvodina, and is the first report of S. tundra on the territory of Serbia. Blood meal analysis provided insights into the preferences of mosquitoes that were positive for Dirofilaria sp. and S. tundra.
Keywords:
Dirofilaria sp.
; Setaria tundra
; mosquito surveillance
; Cox1 gene
1. Introduction
Parasitic filarial nematodes (Nematoda: Filarioidea) of the genus Dirofilaria, represent a severe threat to veterinary and public health, particularly affecting dogs and cats, and in rare occasions, humans as well [1,2,3,4]. Besides canines and felines, these cosmopolitan parasitic worms [5] might also infect other carnivores as well, such as wolves (Canis lupus), red foxes (Vulpes vulpes) and golden jackals (Canis aureus) [6,7,8,9].
Dirofilaria immitis (Leidy 1856), an important mosquito-borne nematode, known as the dog heartworm, causes cardiopulmonary dirofilariasis, invading the heart and large blood vessels [10]. The damage caused by this parasite to arteries and right cardiac chambers of infected hosts might have a fatal outcome, especially if not treated or if treatment is delayed. Another dirofilarial worm is D. repens Railliet et Henry, 1911, which causes subcutaneous infections in dogs and other carnivores [11]. Both Dirofilaria species can accidentally be transmitted to humans [12,13,14,15]. Although humans are dead-end hosts to these filarial nematodes (as they cannot proliferate in the human body), they can still cause health issues depending on the invaded body part. The infection may manifest superficially with the adult nematodes appearing subcutaneously and subconjunctival [16]. However, the major concerns in human populations are the benign pulmonary nodules caused by D. immitis in human lungs, frequently mistaken for malignant lung tumors [13,17,18,19].
Nowadays, cases of dirofilarial infections have been detected worldwide [1]. The process of parasite transmission to hosts is very complex. Successful transmission requires the presence of competent mosquito vectors. Once a mosquito female intakes blood infected with microfilariae, in the following two or more weeks nematodes are going to molt to the infective third larval stage. The infective stage moves from the tubules via the hemocoel to the lumen of the labial sheath in the mosquito’s mouthparts [20]. The duration of this period, measured in the body of several mosquito species (Aedes vexans, Ae. triseratus, Ae. trivittatus, and Anopheles quadrimaculatus), lasted 14 days and is directly temperature-dependent [21,22,23,24]. Subsequent blood meal intake of an infected female mosquito will result in the parasite transmission to the bitten host [10,13].
Regardless of the fact, around 70 mosquito species classified to the Anopheles, Aedes, Culex, Culiseta, and Coquillettidia genera have been considered as potential vectors of animal and human dirofilariasis, where only a few species have been proven as competent vectors [10,25].
Serbia is considered as an endemic country of Dirofilaria sp. in animals and humans for many years [10]. Several studies have been conducted targeting Dirofilaria in reservoirs (animals) and humans [26,27,28,29,30,31,32]. Between 2006 and 2007 the reported prevalence for D. immitis was 7.2% in the Vojvodina and 3.2% in Branicevo regions [26,27]. In the region of Belgrade, a few years later, the prevalence was 22.01%, with 3.97% of dogs showing co-infections with D. repens [10].
Despite all the knowledge about the presence of Dirofilaria sp. in Serbia, little is known about their vectors. So far, only one publication has focused on the vectors of Dirofilaria [33]. Kurucz et al. [33] showed that 8.3 % of tested mosquito pools were positive for Dirofilaria. Positive mosquitoes belonged to five mosquito species: Aedes vexans, Ae. caspius, Ae. sticticus, Culex pipiens and Coquillettidia richiardii. Mosquitoes were found positive for both D. immitis and D. repens at several localities throughout the entire mosquito breeding season.
Therefore, the aim of the present study is to contribute to the mapping of Dirofilaria hotspots in the Vojvodina Province and Mačva region, Serbia. Detecting positive mosquitoes can provide insights into the distribution of the parasite in Serbia. This information could help us better understand how the parasite spreads and adapts to temperate environmental conditions. Analyzing the blood meals of vectors could help create a list of animal species that may be at risk due to potential Dirofilaria infections.
2. Materials and Methods
2.1. Mosquito Sampling and Vector Identification
Mosquito sampling was conducted in Vojvodina Province, Serbia (65 locations), covering an area of 21,506 km2. In addition, eight locations belonging to the Mačva region (612 km²) were included. Sampling was carried out at 73 locations in total (Figure 1), during the summer season of mosquito activity in 2021, starting from May till October. Due to the low number of Aedes albopictus collected in 2021 and the high significance in filarial transmission of this invasive species, we included the samples from 2022 to increase the likelihood of parasite detection. The geo-coordinates of locations are shown in Appendix A. This study only included adult female mosquitoes. Females were collected using CO2 baited (dry ice) adult traps (NS2 trap type). Traps were set up in the afternoon hours and operated overnight. Mosquito samples were then kept in dry ice until transferred to the laboratory within the Centre of Excellence – One Health at the Faculty of Agriculture, University of Novi Sad, Serbia. When the samples arrived in the laboratory, mosquitoes were morphologically identified to species level, using identification key Becker et al. [34].
All collected females per location were categorized based on the presence of blood meal in their abdomen as non-blood fed and blood fed. Females were separated in pools of up to 100 individuals per species per tube. From each mosquito trap only one pool per species was taken. Samples were conserved dry in 2 ml tubes (Eppendorf, Hamburg, Germany) and stored in the freezer on -20 ⁰C until analyzed.
Due to the regularly high number (> 200 per trap) of non-blood fed mosquitoes in traps in the majority of locations, a selection of mosquito species (aimed for further analysis) from this category was based on vector competence to transmit Dirofilaria sp. Selected mosquito species were: Aedes vexans (Meigen 1830), Aedes caspius (Pallas 1771), Aedes albopictus Skuse 1894, and Anopheles maculipennis Complex Meigen 1818.
The number of blood fed females in traps was usually very low (< 5 per trap), therefore we analyzed all captured blood fed mosquito species for the presence of Dirofilaria sp. Because of this low number of blood fed specimens, we also included mosquitoes collected in 2022.
After screening non-blood fed and blood fed mosquitoes for the presence of parasites, we analyzed the blood meal source in blood fed females to identify the putative host species. The following selection for host detection included: a) females from the positive locations belonging to the same species as the positive ones, b) females from locations in the close vicinity to the positive locations. Additionally, non-blood fed females which belonged to the same species and same locations (refers to a and b from above) were also added to try to detect the host (it was assumed that some females might have already digested a blood meal and it was not visible in the abdomen).
2.2. DNA extraction
Extractions and the molecular analysis of all samples were conducted at the Institute of Research and Development, within the Mivegec research unit, Montpellier, France.
Extraction of DNA was carried out by using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturers’ instructions.
For parasite detection, non-blood fed mosquitoes were pooled in tubes by up to 20 individuals for DNA extraction. Therefore, pools with the number of mosquitoes higher than 20 had to be divided. While for blood fed females we put one mosquito per tube, in order to be further analyzed (if positive) for blood meal source detection.
Positive controls of Dirofilaria repens and Dirofilaria immitis were extracted from infected dogs’ blood and were provided by dr. Ettore Napoli (University of Messina, Department of Veterinary Sciences). DNA extraction of positive controls was also done using the Dneasy Blood and Tissue Kit.
2.3. Identification of Dirofilaria sp.
Screening of mosquito pools for the presence of Dirofilaria sp. was conducted using a conventional PCR approach based on the amplification of the COI (Cox1) gene of parasites. The COI gene was targeted using the primer pair COIintF (5′-TGATTGGTGGTTTTGGTAA-3′) and COIintR (5′-ATAAGTACGAGTATCAATATC-3′) under the modified PCR conditions described in Casiraghi et al. [35,36], Gabrielli et al. [37] and Tasić-Otašević et al. [38].
Polymerase chain reaction (PCR) was performed in 25 µl volumes of mix under the following final conditions: 16.05 µl of water, Tp 10x 2.5 µl (Eurogentec, Seraing, Belgium) including 50mM MgCl2 0.75 µl (Eurogentec), 10mM dNTP 0.5 µl (Eurogentec), primer COI-int-F (10pmol/µl = 10µM) 1.5 µl, primer COI-int-R (10pmol/µl = 10µM) 1.5 µl and TAQ Platinum (5U/µl) 0.2 µl (Invitrogen, Waltham, Massachusetts, USA). Two µL of sample DNA were added to 23 µl of Master mix.
The thermal profile used was 94 °C 10 min, and then 5 cycles of 94 °C, 30 sec, 52 °C 45 sec, 72 °C 1 min, afterward 30 cycles of 94 °C 30 sec, 58 °C 45 sec, 72 °C 1 min, and the final was 72 °C 7 min. These conditions provided PCR products of 650 bp.
PCR products were separated by TAE 0.5X and 1.3% agarose gel electrophoresis (Eurogentec) stained with gelred (Biotium, San Francisco, California, USA) and sized with 4.5 µl ladder (Generuler 100 bp, Thermo Scientific, Waltham, Massachusetts, USA). The quantity used for the preparation of gel was as follows: 50 ml of TAE 0.5X, 0.65 g of agarose and 10 µl of stain gelred. The product was then migrated for 35 min at 100 V.
Samples which produced non-specific bands were further processed by sequencing (Eurofins Genomics, Germany). Results of the Blast analysis showing only the highest percent identity (98-100%) were considered in this study.
The consensus sequences were made and cleaned in BioEdit. Sequence alignment was performed using the ClustalW method. Same was done for the five samples which were positive for D. immitis aimed to validate previously detected parasites.
2.4. Identification of Blood Meal Host
Molecular identification of blood meal source species was performed following the protocol by Boessenkool et al. [39]. The primers used were 16Smam1 (CGGTTGGGGTGACCTCGGA) and 16Smam2 (GCTGTTATCCCTAGGGTAACT). PCR was performed in a final volume of 50 µl under the following conditions: water 36 µl, Tp 10X 5 µl (Eurogentec), Mgcl2 50mM 2 µl (Eurogentec), dNTP 10mM 0.2 µl (Eurogentec), primer 16Smam1 (10pmol/µl =10µM) 0.8 µl, primer 16Smam2 (10pmol/µl = 10µM) 0.8 µl and TAQ Platinum (5U/µl) 0.2 µl (Invitrogen). We added 45 µl of Master mix + 5 µL of DNA.
Thermal profile consisted of 55 cycles with the temperatures as follows: 94 °C 2 min, 94 °C 30 sec, 60 °C 30 sec, 72 °C 30 sec, 72 °C 10 min. These conditions provided PCR products of 150 bp.
PCR products were separated by TAE 0.5X and 2% agarose gel electrophoresis stained with gelred and sized with 4.5 µl ladder. The quantity used for the preparation of gel was: 50 ml of TAE 0.5X, 1 g of agarose and 10 µl of stain gelred. The product was then migrated for 35 min at 100 V. Amplicons were sent for sequencing to Eurofins.
Regarding the results of the Blast analysis, only those with the highest percent identity (98-100%) were included in this study.
3. Results
3.1. Presence of Dirofilaria immitis and Setaria tundra in Mosquitoes
The total number of analyzed mosquitoes was 2,902, of which 2,521 were non-blood fed, and 381 were blood fed mosquitoes. For non-blood fed, analyzed specimens belonged to An. maculipennis complex, Ae. vexans, Ae. caspius and Ae. albopictus. For blood fed mosquitoes, in addition to the species mentioned, we also analyzed Aedes sticticus (Meigen 1835), Culex pipiens Linnaeus 1758, Culiseta annulata (Schrank 1776) and Coquillettidia richiardii (Ficalbi 1889) specimens.
For the filarial worms screening we analyzed: 398 An. maculipennis complex (383 non-blood fed and 15 blood fed), 1,340 Ae. vexans specimens (1,253 non-blood fed and 87 blood fed), 316 Ae. caspius (305 non-blood fed and 11 bloodfed), 8 Ae. sticticus (all blood fed), 580 Ae. albopictus (all non-blood fed), 225 Cx. pipiens (all blood fed), 7 Cs. annulata (all blood fed) and 28 Cq. richiardii (all blood fed) specimens. Out of 2,902 screened mosquitoes, the genome of filaria was found in only five mosquito pools (in total six mosquitoes, one pool consisted of two mosquitoes) (Figure 2). After sequencing and Blast analysis, three were found corresponding to D. immitis and two to Setaria tundra. All positive mosquitoes were collected in the Vojvodina Province. Mosquitoes from the Mačva region were not positive for the aimed parasites.
The sequencing and the Blast Analysis confirmed the presence of Dirofilaria immitis in three samples, all of which were detected in Cx. pipiens mosquitoes. Positive Cx. pipiens were collected in three different locations: Glogonj, Svetozar Miletić and Zrenjanin. Positive mosquitoes in Glogonj and Svetozar Miletić were collected in July 2021, while in Zrenjanin Cx. pipens was positive at the end of August 2021. D. immits was present only in blood fed Cx. pipiens.
The results also showed that two out of five positive samples were positive for Setaria tundra (Issaitshikoff & Rajewskaya, 1928), a species of nematode which has not been detected before on the territory of Serbia. In this study, S. tundra was detected in two mosquito species, Ae. caspius and Ae. vexans. Aedes caspius was collected in the location named Mali Iđoš, at the end of July 2021, while Ae. vexans was collected at the location Iđoš, during mid-August of 2021. This parasite was detected in non-blood fed mosquitoes.
All five locations with positive mosquitoes are shown in Figure 3.
3.2. Blood Meal Host Detection
Out of five positive locations for parasites, blood fed females were collected only in four. Besides these four, additional four neighboring locations were included in the analyses. In total blood fed females from eight locations were analyzed.
Out of 30 selected females, 22 were blood fed and eight were non-blood fed females. We analyzed 19 Cx. pipiens (blood fed), seven Ae. vexans (three blood fed and four non-blood fed), and four Ae. caspius (non-blood fed).
In total, 16 mosquitoes resulted in successful host detection. One mosquito was non-blood fed, and the rest of them were blood fed. Identified hosts are presented in Figure 3. The host was not identified in any of the analyzed Ae. caspius females.
3.3. Phylogenetic analysis of Setaria tundra
Phylogenetic analysis of S. tundra nucleotide sequnces were analysed using BLAST NCBI and MEGA v. 11.0 software [40] to align sequences and determine phylogenetic relationships. Maximum Likelihood with the Jones-Taylor-Thornton substitution model was used as the tree construction method. Additionaly, BLAST searches were performed in GenBank (https://www.ncbi.nlm.nih.gov), and S. tundra matches showing a high genetic affinity were downloaded and incorporated into the alignment. Bootstrap analysis of 1000 randomly generated sample trees were performed to assess the stability of the inferred phylogenies. The selected outgroup was D. immitis.
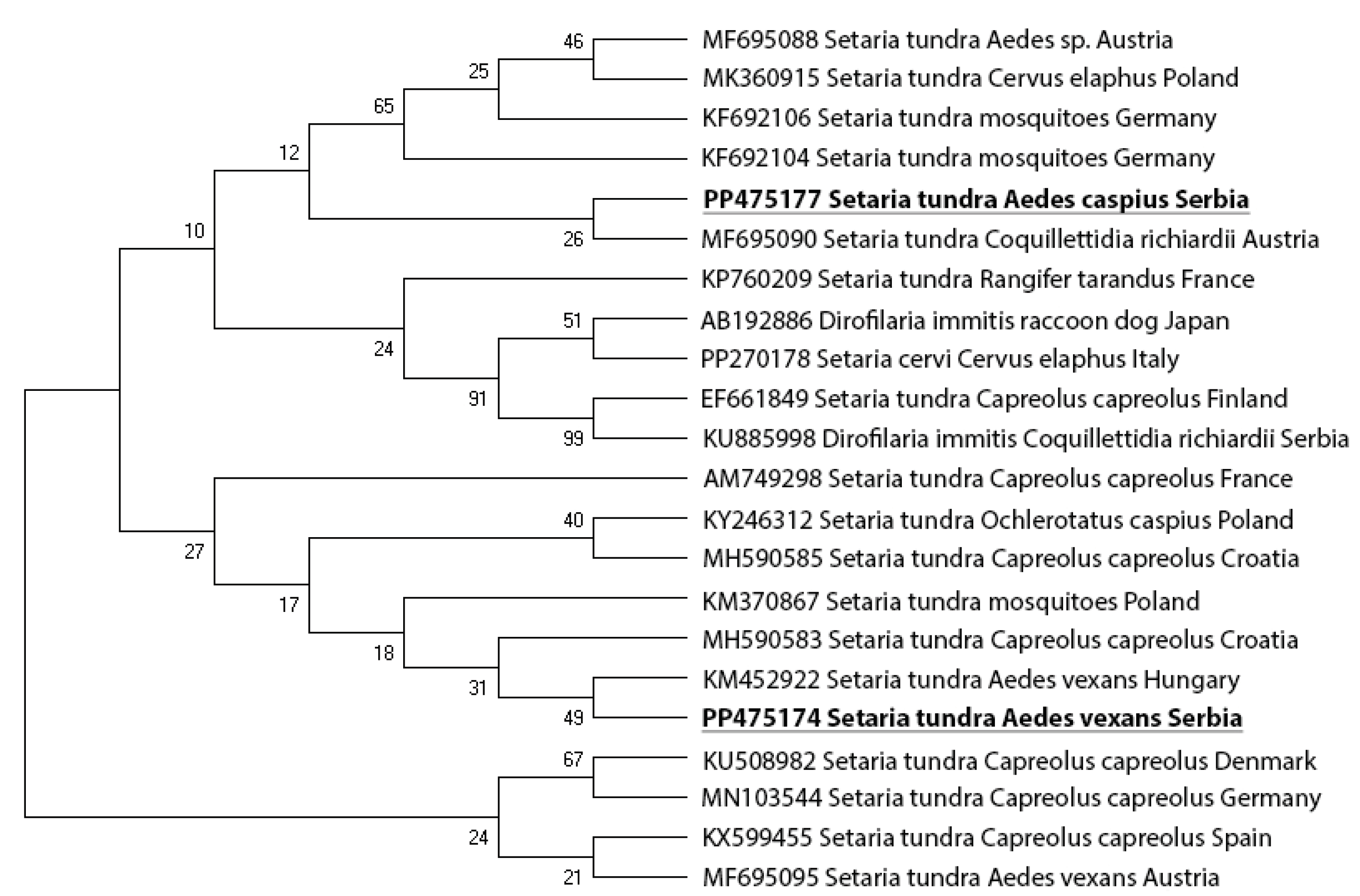
All new nucleotide sequences in this study have been deposited in GenBank NCBI with the accession numbers: PP475177 (S. tundra isolated from Ae. caspius) and PP475174 (S. tundra isolated from Ae. vexans).
The approximate 650 bp fragment of the COX-1 gene was analyzed in two isolates. S. tundra isolated from Ae. caspius has shown similarity with S. tundra originally isolated from Cq. richiardii in Austria (MF695090), while S. tundra isolated from Ae. vexans has shown a similarity with S. tundra isolated from Ae. vexans in Hungary (KM452922) (Figure 4).
4. Discussion
This study represents contribution to the distribution of Dirofilaria spp. and the first record of S. tundra in Serbia. Also, it provides valuable insight in the species of mosquito vectors and their host preference in the temperate climate.
The previous paper published by Kurucz et al. [33] provided the first molecular evidence of D. immitis and D. repens nematodes from mosquito samples in Serbia. However, out of 73 locations, the present study only confirmed D. immitis in 3 locations in Vojvodina Province. Considering that the previous study treated a high number of Cx. pipiens, our study presented more information on other vectors such as Ae. vexans, Ae. caspius, Ae. albopictus and An. maculipennis complex. European studies have confirmed infections by D. immitis in the following mosquito species: Cx. pipiens in Spain [41], Italy [25], and Turkey [42]; Cx. theileri in Madeira, Portugal [43], and on the Canary Islands, Spain [44]; Ae. vexans in Turkey [42,45] and Ae. albopictus, Ae. caspius, An. maculipennis, and Cq. richiardii in Italy [25,46,47].
In our study, only D. immitis was detected in analyzed mosquitoes collected at 73 locations. Although D. repens was earlier detected by Kurucz et al. [33], in this research, it was not found. All three positive samples in the present study belonged to Cx. pipiens. These mosquitoes were collected in three different locations (Svetozar Miletić, Glogonj and Zrenjanin), not close to each other (Glogonj vs Zrenjanin 53 km, Svetozar Miletić vs Zrenjanin 135 km and Glogonj vs Svetozar Miletić 172 km). Two of these locations are villages and one is an urban settlement. The study of Kurucz et al. (2016) [33] detected these parasitic worms in mosquitoes at six locations, and also their positive locations were very distant. Bearing in mind that Cx. pipiens is a very bad flier, it is indicative that D. immitis is a widely spread parasite in Vojvodina Province. One location selected by Kurucz et al. [33] (Zrenjanin) overlaps with our results, demonstrating the persistent circulation of D. immitis in this city (from 2014 till 2021).
Our analysis of blood meal sources from mosquitoes collected in Zrenjanin and Glogonj (both locations positive for D. immitis) demonstrated that Cx. pipiens took the blood (at least the last blood meal) from humans, two mosquitoes being from Zrenjanin and one from Glogonj. In Zrenjanin, other blood meals were identified from various animals including a dog, raven, wild boar, sparrow and pigeon (2 times). In the location close to Zrenjanin, it was demonstrated that Cx. pipiens was feeding on a pigeon. These findings could also represent a contribution to the understanding of the West Nile virus circulation which is very frequently detected in these locations [48]. Interestingly, blood meals of other collected mosquitoes (two Cx. pipiens) were cats’ blood. Earlier studies demonstrated that cats could get infected with Dirofilaria sp., but it does not cause severe disease in them. Cats are not considered as good hosts for Dirofilaria because the infections are cleaned by their immune system before the nematodes can become adults [2]. It is estimated that the prevalence of feline infections in Europe is between 5 and 20% of the total canine prevalence in the same region [5].
The first systematic studies of dirofilariasis in dogs in Serbia were initiated at the beginning of the 21st century. The study was performed in Vojvodina Province showing endemic status for D. repens and D. immitis infection in dogs [27,28]. The climatic conditions in Serbia, coupled with the long activity periods of competent vectors such as Cx. pipiens and Ae. albopictus (Kavran et al., unpub. data), are considered suitable for the transmission of D. immitis and D. repens to humans and animals for at least half of the year (sometimes even more), depending on the air temperature [49,50]. Findings of Savić et al. [17] showed a prevalence of 26.30% for D. immitis infections in dogs, with 25.72% showing microfilariae. The prevalence of D. repens larvae was 1.45%. An earlier study showed a prevalence of 22.9% for D. immitis, while for D. repens it was 39.34% [28]. Several studies conducted in Serbia demonstrated an increasing trend of D. immitis infections and a decreasing trend of D. repens [17,27,51,52].
Setaria tundra is a new species on the list of parasites in Serbia. In this study, S. tundra was found in two locations (Iđoš and Mali Iđoš), which are almost 93 km apart from each other.
Setaria nematodes are classified to the Filarioidea superfamily, family Onchocercidae and are parasites of different ungulates. At least four species of the genus Setaria are present in Europe: S. equina [53], S. cervi [54], S. labiatopapillosa [55] and S. tundra [56]. Setaria tundra was first described in Russia in 1928 [57] and up to now it has been reported in many European countries [58]. The reports from the European countries are given chronologically: Russia 1928 [57], Austria 1969 [59], Finland 1970 [60], Sweden 1973 [61], Norway 1973 [62], Bulgaria 1973 [63], Switzerland 1974 [64], Germany 1975 [65], Italy 2003 [66], France 2006 [67], Denmark 2011 [68], Poland 2010 [69], Hungary 2013 [70], Spain 2016 [71], Croatia 2018 [72], Slovakia 2022 [73].
Olos et al. [74] hypothesized that geographical expansion of Setaria nematodes may be indirectly related to wet and warm summers. This is because intermediate hosts are found in abundance, along with the high density of possible definitive hosts as well as wild and domesticated ungulates. These authors stated that the recent focus on S. tundra has been due to its spreading range to the southern regions of Europe. This species of nematode has expanded its geographical range by hundreds of kilometers and is known to be a major cause of mass mortality of wild and semi domesticated reindeer in Fennoscandia, Finland [75,76]. In northern Europe, the reindeer (Rangifer tarandus) is the major definitive host, yet the moose can serve as an asymptomatic carrier [56,77,78], while roe deer and red deer (Cervus elaphus) serve as the definitive hosts in central and southern Europe [74,76,79]. In the review of Olos et al. [58] it was stated that domestic species such as sheep, goats, cattle, and horses are also potentially at risk [80,81,82,83]. Over the past decade, the populations of wild ruminants and wild boars have increased across Europe [84,85]. This expansion is accompanied by an apparent negative relationship between their abundance in the wild and their health status [86]. Considering that wild animals often enter cattle pastures and spread parasites to livestock, it is of great importance to maintain surveillance and control wildlife diseases [87].
This parasite can be transmitted by several species of mosquitoes, but particularly by those of the genus Aedes [88,89,90]. Microfilariae of this parasite have been reported in Ae. vexans, Ae. caspius, Cx. pipiens, Culex torretium, Aedes annulipes, Ae. sticticus, A. rossicus and Cq. richiardii, in the following countries: Poland [91,92], Hungary [70,93] and in Germany [94,95].
In the present study S. tundra was found in Vojvodina Province in two analyzed mosquito species: Ae. vexans and Ae. caspius. When the blood meals of other mosquitoes from the same location and a nearby one were analyzed, the results showed that two mosquitoes of Ae. vexans were feeding on roe deer, and one had fed on a sheep. The DNA from the blood meal of Ae. caspius was not successfully identified. It is interesting to note that, upon analyzing the locations where these mosquitoes were collected, their traps were not very near to the forests. One trap is located in the middle of a human settlement, while the other one is approximately one km away from the settlement. The second trap was actually placed between a field of sunflower and a vineyard. Bearing in mind that the tested mosquitoes contained the blood of deer, we can assume two possibilities. Either mosquitoes flew from the forest to the humans’ vicinity (Ae. vexans and Ae. caspius have good flight capacities and can fly long distances) or the deer did it.
The number of analyzed mosquitoes did not yield a high number of positive cases of either Dirofilaria or S. tundra. Therefore, we cannot determine the prevalence. According to previous studies that focused on the detection of Dirofilaria in animals and humans, the expected positivity in mosquitoes was much higher than what was demonstrated in this study. It is necessary to perform a systematic screening of mosquitoes, at least in the locations with positive animals and humans, to better understand the prevalence and behavior/preferences of the parasite and to determine potential risks for human and animal populations.
5. Conclusion
The present study provided an update of D. immitis in mosquitoes in the Vojvodina Province and Mačva region. Two new locations of D. immitis presence in vectors in Vojvodina were provided, along with the confirmation of the previously detected positive location where the circulation of the parasite is still active. Setaria tundra was confirmed on the territory of Serbia for the first time in this research. The analysis of blood meals provided insight into the preferences of the species that were positive for Dirofilaria and S. tundra. This opened many questions that would only be answered by systematic research of the determined hotspots, reservoirs and detected mosquito vector species.
Author Contributions
Conceptualization, MK, DP, CP, AIĆ; methodology, DP and CP; formal analysis, MK, SŠ, TM; investigation, MK, SŠ, TM, DJ; data curation, MK, SŠ, TM, DJ, AV; writing—original draft preparation, MK, SŠ, TM; writing—review and editing, MK, SŠ, TM, AIĆ, DJ, AV, CP; visualization, MK, SŠ, TM, AV; supervision, DP and CP; funding acquisition, DP and CP. All authors have read and agreed to the published version of the manuscript.
Funding
This study was done under the COST Action CA17108 and supported by the Ministry of Science and Technology, Serbia (project number 451-03-65/2024-03/ 200117), Secretariat for Urbanism and Environment Protection of Vojvodina Province (Serbia) and Loznica Municipality.
Data Availability Statement
The data presented in this study are available in the article on the site of MDPI Journal.
Acknowledgments
The authors would like to thank the technicians who assisted in the field and laboratory work. Special acknowledgement is referred to Dr Ettore Napoli for positive controls of D. immitis and D. repens, and to Dr Marta Loc for the assistance in the phylogenetic analysis.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The sampling Locations with the Location Name, Geo-Coordinates and Number of Analyzed Females Per Sampling Period
References
- R. Morchón, J. A. Montoya-Alonso, I. Rodríguez-Escolar, and E. Carretón, “What Has Happened to Heartworm Disease in Europe in the Last 10 Years?,” Pathogens, vol. 11, no. 9, p. 1042, Sep. 2022. [CrossRef]
- J. A. Montoya-Alonso et al., “First epidemiological report of feline heartworm infection in the Barcelona metropolitan area (Spain),” Parasit Vectors, vol. 7, no. 1, p. 506, Dec. 2014. [CrossRef]
- W. Tarello, “Clinical Aspects of Dermatitis Associated with Dirofilaria repens in Pets: A Review of 100 Canine and 31 Feline Cases (1990–2010) and a Report of a New Clinic Case Imported from Italy to Dubai,” J Parasitol Res, vol. 2011, pp. 1–7, 2011. [CrossRef]
- D. Traversa, A. Di Cesare, and G. Conboy, “Canine and feline cardiopulmonary parasitic nematodes in Europe: emerging and underestimated,” Parasit Vectors, vol. 3, no. 1, p. 62, 2010. [CrossRef]
- C. Genchi and L. H. Kramer, “The prevalence of Dirofilaria immitis and D. repens in the Old World,” Vet Parasitol, vol. 280, p. 108995, Apr. 2020. [CrossRef]
- Sadighian, “Helminth Parasites of Stray Dogs and Jackals in Shahsavar Area, Caspian Region, Iran,” J Parasitol, vol. 55, no. 2, p. 372, Apr. 1969. [CrossRef]
- J. W. McCall, C. Genchi, L. H. Kramer, J. Guerrero, and L. Venco, “Chapter 4 Heartworm Disease in Animals and Humans,” 2008, pp. 193–285. [CrossRef]
- D. Ćirović et al., “First records of Dirofilaria repens in wild canids from the region of Central Balkan,” Acta Vet Hung, vol. 62, no. 4, pp. 481–488, Dec. 2014. [CrossRef]
- M. Ionică et al., “Role of golden jackals (Canis aureus) as natural reservoirs of Dirofilaria spp. in Romania,” Parasit Vectors, vol. 9, no. 1, p. 240, Dec. 2016. [CrossRef]
- R. Morchón, E. Carretón, J. González-Miguel, and I. Mellado-Hernández, “Heartworm Disease (Dirofilaria immitis) and Their Vectors in Europe – New Distribution Trends,” Front Physiol, vol. 3, 2012. [CrossRef]
- D. Otranto et al., “Vector-borne helminths of dogs and humans in Europe,” Parasit Vectors, vol. 6, no. 1, p. 16, Dec. 2013. [CrossRef]
- F. Rivasi, R. Boldorini, P. Criante, M. Leutner, and S. Pampiglione, “Detection of Dirofilaria (Nochtiella) repens DNA by polymerase chain reaction in embedded paraffin tissues from two human pulmonary locations,” APMIS, vol. 114, no. 7–8, pp. 566–573, Aug. 2006. [CrossRef]
- F. Simón et al., “Human and Animal Dirofilariasis: the Emergence of a Zoonotic Mosaic,” Clin Microbiol Rev, vol. 25, no. 3, pp. 507–544, Jul. 2012. [CrossRef]
- B. K. Saha et al., “Human Pulmonary Dirofilariasis: A Review for the Clinicians,” Am J Med Sci, vol. 363, no. 1, pp. 11–17, Jan. 2022. [CrossRef]
- T. Sako et al., “Human pulmonary dirofilariasis presenting as a small nodule with a cavity.,” J Med Invest, vol. 47, no. 3–4, pp. 161–3, Aug. 2000.
- Muro, C. Genchi, M. Cordero, and F. Simón, “Human Dirofilariasis in the European Union,” Parasitology Today, vol. 15, no. 9, pp. 386–389, Sep. 1999. [CrossRef]
- S. Savić et al., “Seroepidemiological Study of Canine and Human Dirofilariasis in the Endemic Region of Northern Serbia,” Front Vet Sci, vol. 7, Sep. 2020. [CrossRef]
- L. Zumaquero, F. Simón, E. Carretón, I. Hernández, C. Sandoval, and R. Morchón, “Prevalence of canine and human dirofilariosis in Puebla, Mexico,” Vet Parasitol, vol. 282, p. 109098, Jun. 2020. [CrossRef]
- P. A. Ferrari, A. Grisolia, S. Reale, R. Liotta, A. Mularoni, and A. Bertani, “A rare case of human pulmonary dirofilariasis with nodules mimicking malignancy: approach to diagnosis and treatment,” J Cardiothorac Surg, vol. 13, no. 1, p. 65, Dec. 2018. [CrossRef]
- J. M. Medlock, I. Barrass, E. Kerrod, M. A. Taylor, and S. Leach, “Analysis of Climatic Predictions for Extrinsic Incubation of Dirofilaria in The United Kingdom,” Vector-Borne and Zoonotic Diseases, vol. 7, no. 1, pp. 4–14, Mar. 2007. [CrossRef]
- J. Fortin and J. Slocombe, “Temperature requirements for the development of Dirofilaria immitis in Aedes triseriatus and Ae. vexans.,” Mosq News, vol. 41, no. 4, pp. 625–633, 1981.
- Bruce., M., Christensen, and A. Hollander, “Effect of temperature on vector-parasite relationships of Aedes trivittatus and Dirofilaria immitis.,” Proceedings of Helminthology Society, vol. 45, pp. 115–119, 1978.
- T. J. Jankowski and W. E. Bickley, “The Mosquitoes, Aedes canadensis and A. vexans as Potential Vectors of Dirofilaria immitis in Maryland 1,” Ann Entomol Soc Am, vol. 69, no. 5, pp. 781–783, Sep. 1976. [CrossRef]
- F. W. Kutz and R. C. Dobson, “Effects of Temperature on the Development of Dirofilaria immitis (Leidy) in Anopheles quadrimaculatus Say1 and on Vector Mortality Resulting from this Development2,3,” Ann Entomol Soc Am, vol. 67, no. 3, pp. 325–331, May 1974. [CrossRef]
- G. Cancrini et al., “Natural vectors of dirofilariasis in rural and urban areas of the Tuscan region, central Italy.,” J Med Entomol, vol. 43, no. 3, pp. 574–9, May 2006. [CrossRef]
- S. Dimitrijević, A. Tasić, S. Tasić, V. Adamović, T. Ilić, and N. Miladinović-Tasić, “Filariosis in dogs in Serbia,” in Dirofilaria immitis and D. repens in dog and cat and human infections, Mappe parassitologiche, vol. 8, C. Genchi, L. Rinaldi, and G. Cringoli, Eds., 2007, p. 201.
- Tasić, L. Rossi, S. Tasić, N. Miladinović-Tasić, T. Ilić, and S. Dimitrijević, “Survey of canine dirofilariasis in Vojvodina, Serbia,” Parasitol Res, vol. 103, no. 6, pp. 1297–1302, Nov. 2008. [CrossRef]
- Tasić et al., “Canine Dirofilaria Infections in Two Uninvestigated Areas of Serbia: Epidemiological and Genetic Aspects,” Vector-Borne and Zoonotic Diseases, vol. 12, no. 12, pp. 1031–1035, Dec. 2012. [CrossRef]
- Tasić, S. Tasić, N. Miladinović-Tasić, D. Zdravković, and J. Đorđević, “Prevalence of Dirofilaria repens-cause of zoonosis in dogs,” Acta Fac Med Naiss, vol. 24, no. 2, pp. 71–74, 2007.
- Penezić, S. Selaković, I. Pavlović, and D. Ćirović, “First findings and prevalence of adult heartworms (Dirofilaria immitis) in wild carnivores from Serbia,” Parasitol Res, vol. 113, no. 9, pp. 3281–3285, Sep. 2014. [CrossRef]
- P. Gavrilović, G. Blitva-Robertson, J. Özvegy, F. Kiskároly, and Z. Becskei, “Case Report of dirofilariasis in grey wolf in Serbia.,” Acta Parasitol, vol. 60, no. 1, pp. 175–8, Mar. 2014. [CrossRef]
- Potkonjak et al., “Molecular survey of Dirofilaria species in stray dogs, red foxes and golden jackals from Vojvodina, Serbia,” Comp Immunol Microbiol Infect Dis, vol. 68, p. 101409, Feb. 2020. [CrossRef]
- K. Kurucz et al., “First molecular identification of Dirofilaria spp. (Onchocercidae) in mosquitoes from Serbia,” Parasitol Res, vol. 115, no. 8, pp. 3257–3260, Aug. 2016. [CrossRef]
- N. Becker et al., Mosquitoes, 3rd ed. Cham: Springer International Publishing, 2020. [CrossRef]
- M. Casiraghi, T. J. C. Anderson, C. Bandi, C. Bazzocchi, and C. Genchi, “A phylogenetic analysis of filarial nematodes: comparison with the phylogeny of Wolbachia endosymbionts,” Parasitology, vol. 122, no. 1, pp. 93–103, Jan. 2001. [CrossRef]
- M. Casiraghi et al., “Mapping the presence of Wolbachia pipientis on the phylogeny of filarial nematodes: evidence for symbiont loss during evolution,” Int J Parasitol, vol. 34, no. 2, pp. 191–203, Feb. 2004. [CrossRef]
- S. Gabrielli et al., “Molecular Identification of New Cases of Human Dirofilariosis (Dirofilaria repens) in Italy,” Pathogens, vol. 10, no. 2, p. 251, Feb. 2021. [CrossRef]
- S. Tasić-Otašević et al., “Molecular Survey of Dirofilaria and Leishmania Species in Dogs from Central Balkan,” Animals, vol. 12, no. 7, p. 911, Apr. 2022. [CrossRef]
- S. Boessenkool et al., “Blocking human contaminant DNA during PCR allows amplification of rare mammal species from sedimentary ancient DNA,” Mol Ecol, vol. 21, no. 8, pp. 1806–1815, Apr. 2012. [CrossRef]
- K. Tamura, G. Stecher, and S. Kumar, “MEGA11: Molecular Evolutionary Genetics Analysis Version 11,” Mol Biol Evol, vol. 38, no. 7, pp. 3022–3027, Jun. 2021. [CrossRef]
- . Morchón et al., “Haplotype H1 of Culex pipiens Implicated as Natural Vector of Dirofilaria immitis in an Endemic Area of Western Spain,” Vector-Borne and Zoonotic Diseases, vol. 7, no. 4, pp. 653–658, Dec. 2007. [CrossRef]
- \Yildirim, A. Inci, O. Duzlu, Z. Biskin, A. Ica, and I. Sahin, “Aedes vexans and Culex pipiens as the potential vectors of Dirofilaria immitis in Central Turkey,” Vet Parasitol, vol. 178, no. 1–2, pp. 143–147, May 2011. [CrossRef]
- M. Santa-Ana, M. Khadem, and R. Capela, “Natural Infection of Culex theileri (Diptera: Culicidae) with Dirofilaria immitis (Nematoda: Filarioidea) on Madeira Island, Portugal,” J Med Entomol, vol. 43, no. 1, pp. 104–106, Jan. 2006. [CrossRef]
- R. Morchón et al., “Molecular Characterization of Culex Theileri from Canary Islands, Spain, a Potential Vector of Dirofilaria Immitis,” J Clin Exp Pathol, vol. 01, no. S3, pp. 1–6, 2011. [CrossRef]
- Z. Bışkın, O. Düzlü, A. Yildirim, and A. Incı, “The molecular diagnosis of Dirofilaria immitis in vector mosquitoes in Felahiye district of Kayseri.,” Turkiye Parazitol Derg, vol. 34, no. 3, pp. 200–5, 2010.
- G. Cancrini, M. Pietrobelli, A. F. Frangipane di Regalbono, M. P. Tampieri, and A. della Torre, “Development of Dirofilaria and Setaria nematodes in Aedes albopictus.,” Parassitologia, vol. 37, no. 2–3, pp. 141–5, Dec. 1995.
- G. Cancrini, A. Frangipane di Regalbono, I. Ricci, C. Tessarin, S. Gabrielli, and M. Pietrobelli, “Aedes albopictus is a natural vector of Dirofilaria immitis in Italy,” Vet Parasitol, vol. 118, no. 3–4, pp. 195–202, Dec. 2003. [CrossRef]
- D. Petrić et al., “West Nile virus ‘circulation’ in Vojvodina, Serbia: Mosquito, bird, horse and human surveillance.,” Mol Cell Probes, vol. 31, pp. 28–36, Feb. 2017. [CrossRef]
- G. Capelli et al., “Recent advances on Dirofilaria repens in dogs and humans in Europe,” Parasit Vectors, vol. 11, no. 1, p. 663, Dec. 2018. [CrossRef]
- M. Krstić et al., “An appraisal of canine and human cases reveals an endemic status of dirofilariosis in parts of Serbia,” Mol Cell Probes, vol. 31, pp. 37–41, Feb. 2017. [CrossRef]
- L. Spasojević-Kosić, V. Lalošević, D. Lalošević, S. Simin, I. Vasić, and L. Kuruca, “Prevalence of Dirofilariosis in pet dogs in Novi Sad,” Contemporary Agriculture, vol. 61, pp. 247–254, 2012, Accessed: Mar. 20, 2024. [Online]. Available: https://www.researchgate.net/publication/259962443_Prevalence_of_Dirofilariosis_in_pet_dogs_in_Novi_Sad.
- L. Spasojević Kosić, V. Lalošević, S. Simin, and L. Kuruca, “Dirofilariosis and angiostrongilosis in pet and hunting dogs in Novi Sad, Vojvodina, Serbia,” Archives of Veterinary Medicine, vol. 9, no. 2, pp. 53–62, Mar. 2017. [CrossRef]
- J. J. Gawor, “The prevalence and abundance of internal parasites in working horses autopsied in Poland,” Vet Parasitol, vol. 58, no. 1–2, pp. 99–108, May 1995. [CrossRef]
- M. T. Manfredi, G. Piccolo, C. Fraquelli, and F. Perco, “Elmintofauna del cervo nel Parco Nazionale dello Stelvio,” J Mt Ecol, vol. 7, pp. 245–249, 2003.
- W. Demiaszkiewicz, J. Lachowicz, and G. Karbowiak, “Wzrost zarażenia żubrów w Puszczy Białowieskiej nicieniami Setaria labiatopapillosa,” Wiadomoœci Parazytologiczne, vol. 53, no. 4, pp. 335–338, 2007.
- W. Demiaszkiewicz, I. Kuligowska, A. M. Pyziel, and J. Lachowicz, “First cases of nematodes Setaria tundra invasion in elk (Alces alces) in Poland,” Med Weter, vol. 71, no. 8, pp. 510–512, 2015.
- Z. A. Raevskaya, “Setaria and their pathogenic significance,” Trudy Gosudarstvennogo Instituta Eksperimental’noi Veterinarii, vol. 5, pp. 1–58, 1928.
- G. Oloś, J. Nowakowska, and R. Welc-Falęciak, “Setaria tundra, what do we know, what is still to be discovered?,” Ann Parasitol, vol. 67, no. 1, pp. 1–10, 2021. [CrossRef]
- E. Kutzer and H. K. Hinaidy, “Die Parasiten der wildlebenden Wiederkäuer Österreichs,” Z Parasitenk , vol. 32, pp. 354–368, 1969.
- E. Savonen, “The implementation of domestic reindeer meat inspection,” Suomen Eläinlääkärilehti, vol. 84, pp. 651–653, 1978.
- Rehbinder, D. Christensson, and V. Glatthard, “Parasitic Granulomas in Swedish Forest Reindeer (Rangifer Tarandus),” in Wildlife Diseases, Boston, MA: Springer US, 1976, pp. 597–607. [CrossRef]
- T. T. Poppe, “Parasitic diseases recorded in reindeer during meat inspection in Kautokeino (Norway) in autumn and winter,” Norsk Veterinaertidsskrift, vol. 89, pp. 791–795, 1977.
- Y. Yanchev, “The helminth fauna of roe deer (Capreolus capreolus) in Bulgaria. 3. Material on helminth fauna in roe deer (Capreolus capreolus L.) in the mountains of southern Bulgaria,” Izvestiya naTsentralnata Khelmintologichna Laboratoriya, vol. 16, pp. 205–220, 1973.
- J. R. Andrews, B. Hörning, and A. Wandeler, “Endoparasites of roe deer (Capreolus capreolus L.) from Switzerland with special reference to hosts from the Emmental region of Canton Berne,” Revue Suisse de Zoologie, Annales de la Societe Zoologique Suisse et du Museum d’Histoire Naturelle de Geneve, vol. 81, p. 13, 1974.
- K. Büttner, “Untersuchungen zur Parasitierung des Rehwildes bei steigendem Jagddruck,” Zeitschrift für Jagdwissenschaft, vol. 24, pp. 139–155, 1978.
- G. Favia, “Molecular assay for the identification of Setaria tundra,” Vet Parasitol, vol. 117, no. 1–2, pp. 139–145, Nov. 2003. [CrossRef]
- E. Ferri et al., “Integrated taxonomy: traditional approach and DNA barcoding for the identification of filarioid worms and related parasites (Nematoda),” Front Zool, vol. 6, no. 1, p. 1, Dec. 2009. [CrossRef]
- H. L. Enemark, A. Oksanen, M. Chriél, J. le Fèvre Harslund, I. D. Woolsey, and M. N. S. Al-Sabi, “Detection and molecular characterization of the mosquito-borne filarial nematode Setaria tundra in Danish roe deer (Capreolus capreolus),” Int J Parasitol Parasites Wildl, vol. 6, no. 1, pp. 16–21, Apr. 2017. [CrossRef]
- M. Bednarski, T. Piasecki, M. Bednarska, and Z. Soltysiak, “Invasion of Setaria tundra in roedeer (Capreolus capreolus) - case report,” Acta Scientiarum Polonorum. Medicina Veterinaria, vol. 3, no. 09, pp. 21–25, 2010, Accessed: Mar. 20, 2024. [Online]. Available: https://www.infona.pl//resource/bwmeta1.element.agro-efb22ab5-15e9-47fa-9f42-6ad3af19da0f.
- Zittra et al., “Screening blood-fed mosquitoes for the diagnosis of filarioid helminths and avian malaria,” Parasit Vectors, vol. 8, no. 1, pp. 1–6, Dec. 2015. [CrossRef]
- S. Angelone-Alasaad et al., “First report of Setaria tundra in roe deer (Capreolus capreolus) from the Iberian Peninsula inferred from molecular data: epidemiological implications,” Parasit Vectors, vol. 9, no. 1, p. 521, Dec. 2016. [CrossRef]
- J. Čurlík, D. Konjević, M. Bujanić, Ž. Sabol, F. Martinković, and M. Sindičić, “The first description of Setaria tundra (Issaitshikoff & Rajewskaya, 1928) in roe deer from Croatia,” Helminthologia, vol. 56, no. 3, pp. 252–255, Sep. 2019. [CrossRef]
- J. Čurlík et al., “The first report of Setaria tundra (Issaitshikoff & Rajewskaya, 1928) in Slovakia by using of molecular methods,” Vet Res Commun, vol. 47, no. 4, pp. 2247–2251, Dec. 2023. [CrossRef]
- G. Oloś, J. Nowakowska, S. Rojewska, and R. Welc-Falęciak, “New findings of Setaria tundra and Setaria cervi in the red deer (Cervus elaphus) in Poland,” Parasitology, vol. 146, no. 10, pp. 1333–1337, Sep. 2019. [CrossRef]
- S. Laaksonen, J. Kuusela, S. Nikander, M. Nylund, and A. Oksanen, “Outbreak of parasitic peritonitis in reindeer in Finland,” Veterinary Record, vol. 160, no. 24, pp. 835–841, Jun. 2007. [CrossRef]
- S. Laaksonen, M. Solismaa, R. Kortet, J. Kuusela, and A. Oksanen, “Vectors and transmission dynamics for Setaria tundra (Filarioidea; Onchocercidae), a parasite of reindeer in Finland,” Parasit Vectors, vol. 2, no. 1, p. 3, 2009. [CrossRef]
- J. Kowal, S. Kornaś, P. Nosal, M. Basiaga, and M. Lesiak, “Setaria tundra in roe deer (Capreolus capreolus)--new findings in Poland.,” Ann Parasitol, vol. 59, no. 4, pp. 179–82, 2013.
- K. Tomczuk et al., “Internal parasites in roe deer of the Lubartów Forest Division in postmortem studies,” Med Weter, vol. 73, no. 11, pp. 726–730, 2017. [CrossRef]
- S. Laaksonen and A. Oksanen, “Status and review of the vector-borne nematode Setaria tundra in Finnish cervids,” A Journal Devoted to the Biology and Management of Moose, vol. 45, pp. 81–84, 2009, Accessed: Mar. 20, 2024. [Online]. Available: https://www.researchgate.net/publication/277218607_Status_and_review_of_the_vector-borne_nematode_Setaria_tundra_in_Finnish_cervids.
- T. Bazargani et al., “Cerebrospinal Nematodiasis of Cattle, Sheep and Goats in Iran,” Iran J Parasitol, vol. 3, no. 1, pp. 16–20, 2008, Accessed: Mar. 20, 2024. [Online]. Available: https://www.researchgate.net/publication/228483310_Cerebrospinal_Nematodiasis_of_Cattle_Sheep_and_Goats_in_Iran.
- G. Sharhuu and T. Sharkhuu, “The helminth fauna of wild and domestic ruminants in Mongolia-a review,” Eur J Wildl Res, Aug. 2004. [CrossRef]
- S. T. B. Sundar and P. E. D’Souza, “Morphological characterization of Setaria worms collected from cattle,” Journal of Parasitic Diseases, vol. 39, no. 3, pp. 572–576, Sep. 2015. [CrossRef]
- Kuchboev, O. Amirov, R. R. Karimova, and M. Asakawa, “Nematodes in the digestive tracts of domestic ruminants in Uzbekistan,” Jpn J Vet Parasitol, vol. 15, no. 2, pp. 124–128, 2016, Accessed: Mar. 20, 2024. [Online]. Available: https://www.researchgate.net/publication/313399954_Kuchboev_et_al_Jpn_J_Vet_Parasitol_Vol_15_No_2_2016.
- V. Lavadinovic, Z. Popovic, D. Beukovic, and K. Cokoski, “Wild boar management (Sus scrofa L.) in the Republic of Serbia,” Glasnik Sumarskog fakulteta, no. 121, pp. 47–60, 2020. [CrossRef]
- V. Milićević et al., “Cross-sectional serosurvey of selected infectious diseases in wild ruminants in Serbia,” Res Vet Sci, vol. 170, p. 105183, Apr. 2024. [CrossRef]
- S. Lanková et al., “Setaria cervi (Filarioidea, Onchocercidae) undressing in ungulates: altered morphology of developmental stages, their molecular detection and complete sequence cox 1 gene,” Parasitology, vol. 148, no. 5, pp. 598–611, Apr. 2021. [CrossRef]
- S. Nechybová et al., “Long-term occurrence of Trichuris species in wild ruminants in the Czech Republic,” Parasitol Res, vol. 117, no. 6, pp. 1699–1708, Jun. 2018. [CrossRef]
- R. C. Anderson, Nematode parasites of Vertebrates, their development and transmission, 2nd ed. New York: CABI Publishing, 2000. Accessed: Mar. 20, 2024. [Online]. Available: https://books.google.rs/books?hl=en&lr=&id=lEERbfsvP1EC&oi=fnd&pg=PR9&dq=The+superfamily+Filarioidea.+Nematode+parasites+of+Vertebrates,+their+development+and+transmission&ots=uWqQLNtQfb&sig=JfyKt0sUeJcCXIuApfj9SlKDUJU&redir_esc=y#v=onepage&q=The%20superfamily%20Filarioidea.%20Nematode%20parasites%20of%20Vertebrates%2C%20their%20development%20and%20transmission&f=false.
- Genchi, L. Rinaldi, M. Mortarino, M. Genchi, and G. Cringoli, “Climate and Dirofilaria infection in Europe,” Vet Parasitol, vol. 163, no. 4, pp. 286–292, Aug. 2009. [CrossRef]
- S. Laaksonen et al., “Climate Change Promotes the Emergence of Serious Disease Outbreaks of Filarioid Nematodes,” Ecohealth, vol. 7, no. 1, pp. 7–13, Aug. 2010. [CrossRef]
- K. Rydzanicz, E. Lonc, A. Masny, and E. Golab, “Detection of Setaria tundra microfilariae in mosquito populations from irrigated fields in Wroclaw (Poland,” Ann Parasitol, vol. 62, no. 132, 2016, Accessed: Mar. 20, 2024. [Online]. Available: https://annals-parasitology.eu/archive_2001_2022/2016-62-Suppl-S4_132.pdf.
- Masny, W. Rożej-Bielicka, and E. Gołąb, “Setaria tundra invasive larvae in a mosquito vector in Poland,” Annales of Parasitology, vol. 59, p. 178, 2013, Accessed: Mar. 20, 2024. [Online]. Available: yadda.icm.edu.p.
- G. Kemenesi et al., “Circulation of Dirofilaria repens, Setaria tundra, and Onchocercidae species in Hungary during the period 2011–2013,” Vet Parasitol, vol. 214, no. 1–2, pp. 108–113, Nov. 2015. [CrossRef]
- Czajka, N. Becker, S. Poppert, H. Jöst, J. Schmidt-Chanasit, and A. Krüger, “Molecular detection of Setaria tundra (Nematoda: Filarioidea) and an unidentified filarial species in mosquitoes in Germany,” Parasit Vectors, vol. 5, no. 1, p. 14, Dec. 2012. [CrossRef]
- M. Kronefeld, H. Kampen, R. Sassnau, and D. Werner, “Molecular detection of Dirofilaria immitis, Dirofilaria repens and Setaria tundra in mosquitoes from Germany,” Parasit Vectors, vol. 7, no. 1, p. 30, Dec. 2014. [CrossRef]
Figure 1.
Sampling locations in Vojvodina Province and the Mačva region.
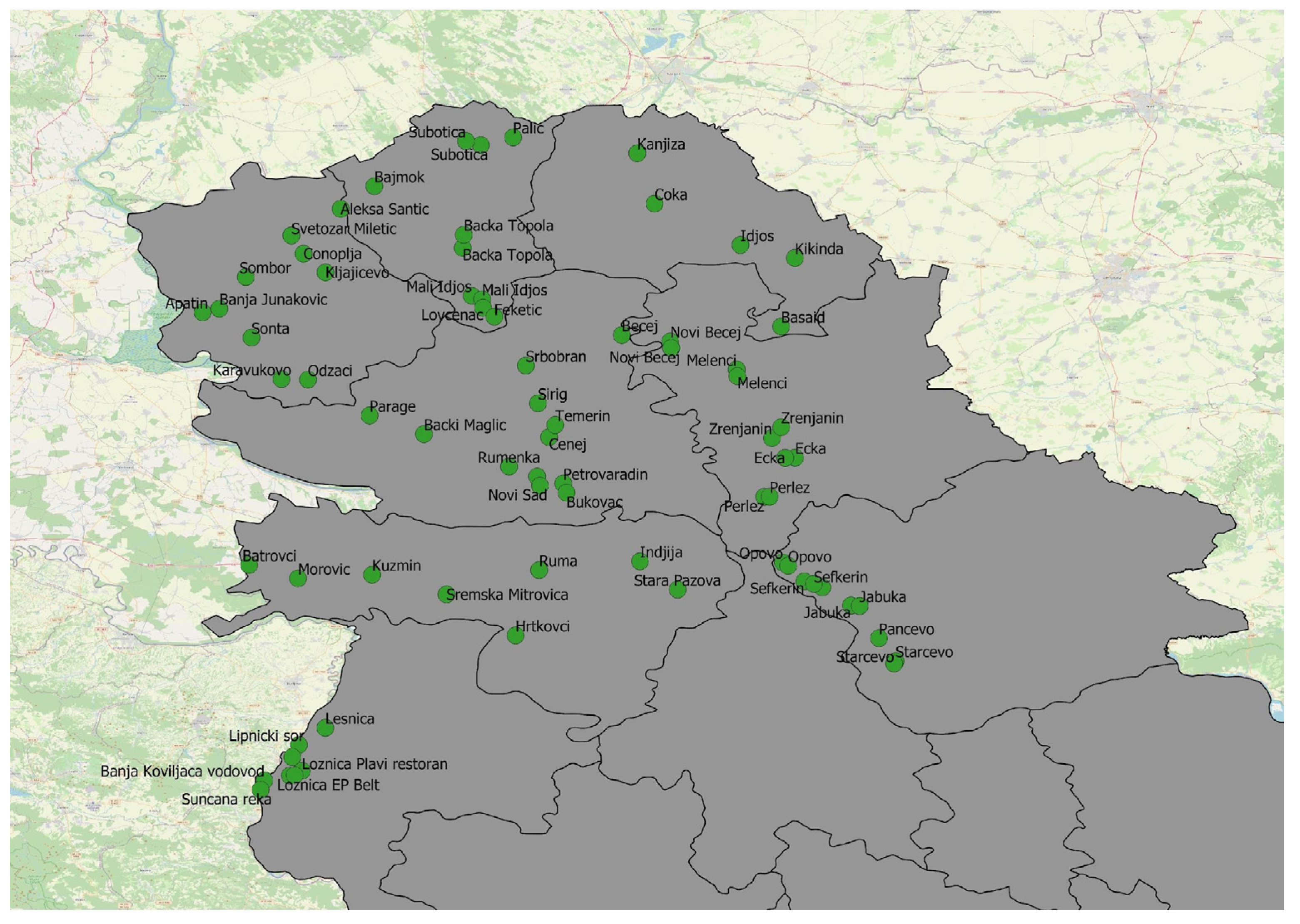
Figure 2.
Top image – Cx. pipiens infected with D. immitis (product size 650 bp); Bottom image - Ae. vexans and Ae. caspius infected with S. tundra (650 bp).
Figure 2.
Top image – Cx. pipiens infected with D. immitis (product size 650 bp); Bottom image - Ae. vexans and Ae. caspius infected with S. tundra (650 bp).
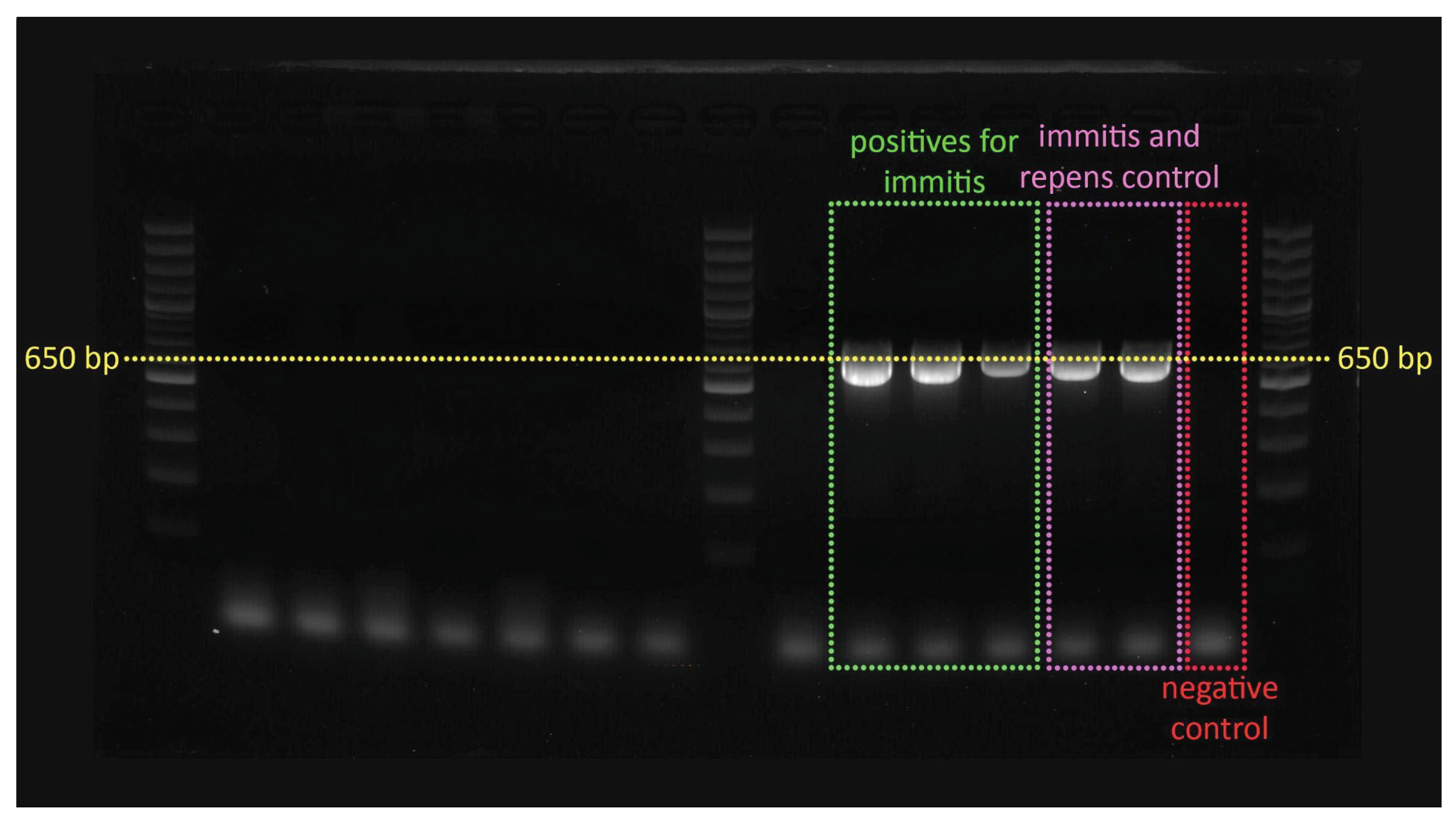
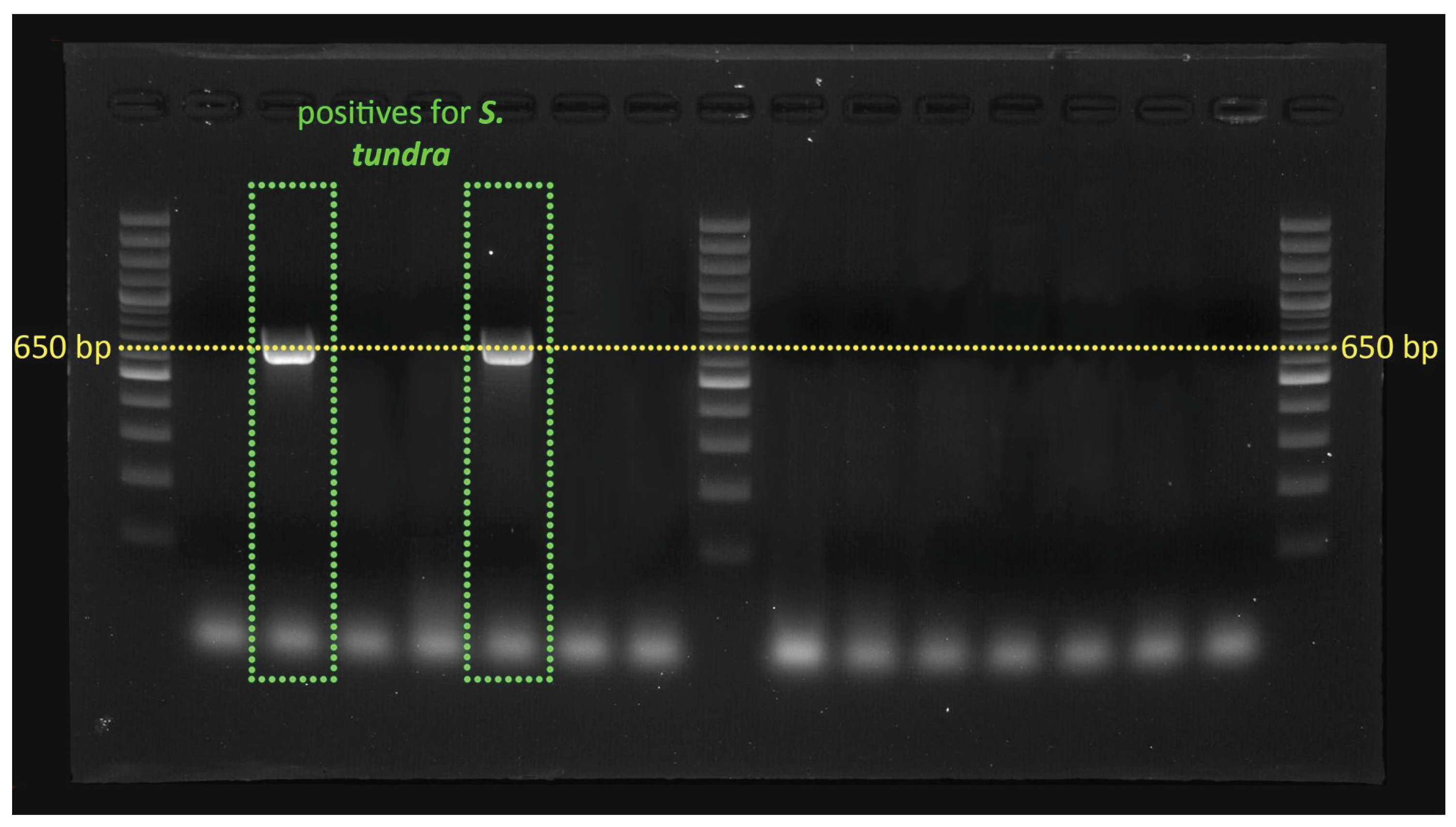
Figure 3.
Locations with Dirofilaria immits and Setaria tundra positive mosquitoes, and detected blood meal hosts.
Figure 3.
Locations with Dirofilaria immits and Setaria tundra positive mosquitoes, and detected blood meal hosts.
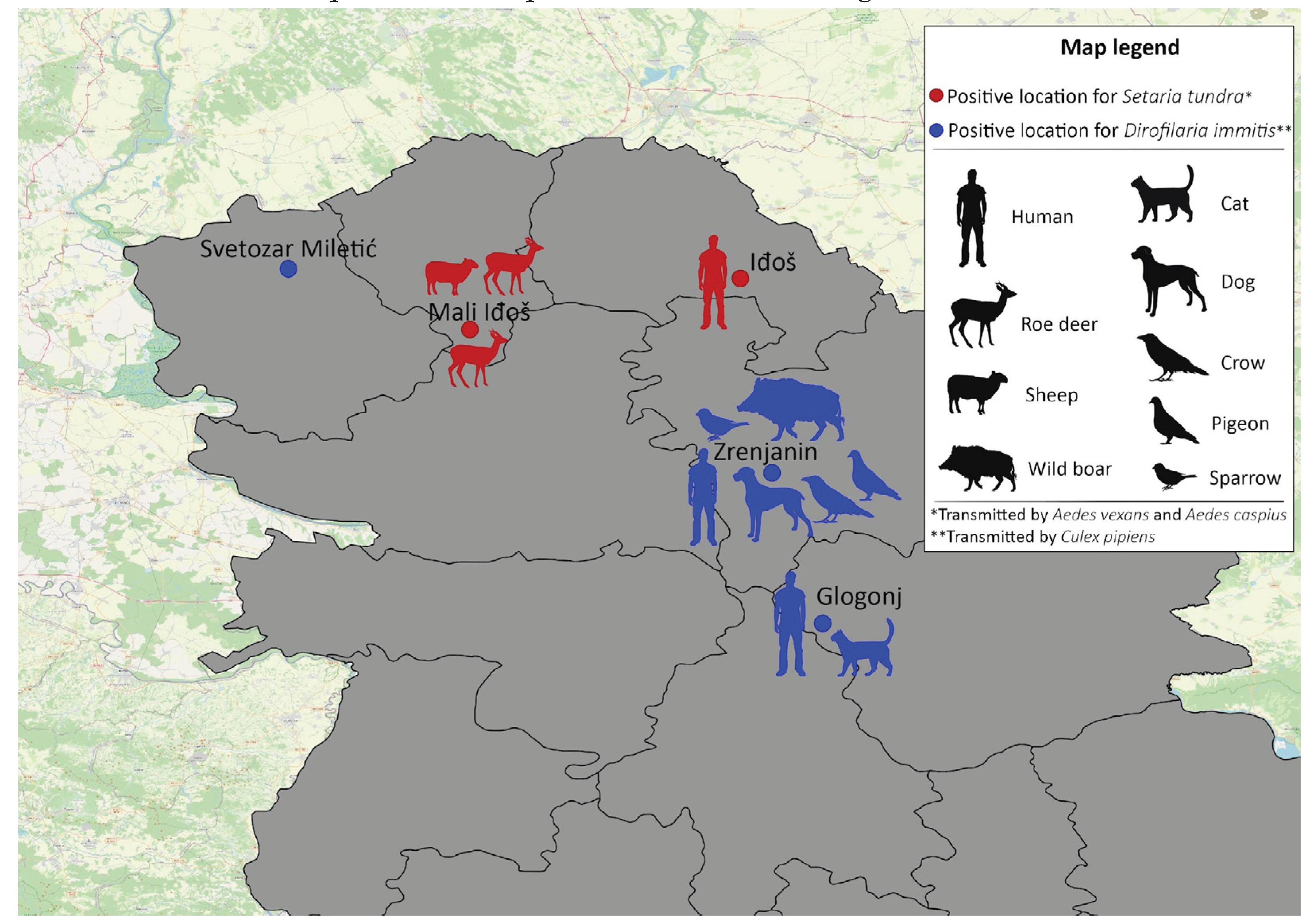
Figure 4.
Maximum likelihood (ML) tree of the Setaria isolates identified in two mosquito species (Ae. vexans and Ae. caspius) from Serbia (bolded and underlined) and selected isolates from GenBank, based on a fragment of the COX-1 gene. The numbers shown at the tree nodes represent bootstrap values based on 1000 replicates.
Figure 4.
Maximum likelihood (ML) tree of the Setaria isolates identified in two mosquito species (Ae. vexans and Ae. caspius) from Serbia (bolded and underlined) and selected isolates from GenBank, based on a fragment of the COX-1 gene. The numbers shown at the tree nodes represent bootstrap values based on 1000 replicates.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



