
Submitted:
10 April 2024
Posted:
11 April 2024
You are already at the latest version
Abstract
The focus of our study was the changes in the composition of semi-natural dry grasslands in Hungary. Since these valuable habitats were created with the help of humans, their preservation also requires regular treatment. Our mowing experiment aimed to investigate the suppression of the native Calamagrostis epigejos L. Roth. In Hungary, in the Cserhát Mountains, eight permanent plots were mowed twice a year. The vegetation was sampled annually between 2001 and 2011. The effects of mowing were investigated with repeated measures analysis of variance (ANOVA). After 10 years, the C. epigejos cover of the mowed plots decreased significantly, from the initial average of 62.38 to 7.50%. Surprisingly, we also experienced a similar decrease in the control plots. Regular treatment also had a positive effect on the number of species and diversity. Species richness increased continuously in both treatment types, which indicates the combined effect of vegetation succession and treatment. The biomass growth of other Poaceae and Fabaceae species, which are important from a grassland management perspective, was also facilitated by mowing. Our results suggest that long-term in situ experiments and continuous botanical studies are needed to establish a reliable utilization of grasslands.
Keywords:
Calamagrostis epigejos L. Roth
; conservation management
; grassland management
; land abandonment
; dry grassland
; loes steppe
; long-term data
; nature conservation
1. Introduction
Grasslands in the Carpathian Basin belong to natural forest-steppe and steppe vegetation [1] on the western border of palearctic steppe zone in Europe [2,3].
For the maintenance of grasslands a traditional land use is needed such as regular burning, grazing, or mowing and making hay [4]. These human activities contributed to species exchange between grasslands, balancing propagule availability [5,6] and they created plat communities that are today the most important sites of European biodiversity [7]. The traditional management of abandoned vineyards and orchards maintained the pattern and scale of the landscape, however, a consequence of the intensification of agriculture is that some areas are overused, while others are abandoned [8].
Land use changes often cause the loss of diversity of man-made habitats [9,10,11,12]. The diversity of grasslands is significantly reduced by the abandonment of pastures and the decline of livestock [13,14]. Overused and abandoned habitats are exposed to the rapid spreading of non-native invasive species [15]. Nevertheless, native species, especially grasses, are also capable of monotonously increasing, invasion-like expansion. Deschampsia [16], Sesleria [17] Brachypodium [18,19], and Molinia [20] are widespread dominant grass species in Europe, which fast colonizing leads to changes in the availability of nutrients, as well as to changes in temporal and spatial niche divisions. This significantly reduces the germination, establishment, and growth success of other plant species, which ultimately results in the loss of species richness [21,22].
Calamagrostis epigejos (L.) Roth (see below C. epigejos) is a strong, fast-spreading perennial grass species [23,24,25] which spread successfully in areas where previous human activity was abandoned [26,27,28]. It has extremely high morphological and physiological plasticity [25] and resistance to various harmful environmental factors. C. epigejos is widespread in Europe, rarely occurs in undisturbed close-to-nature and natural grasslands, however, it can invade these habitats as well [29]. Occurs in forests [30,31] in river floodplains [32,33] and in disturbed sites, on barren wastelands, in reclaimed mines [34,35,36]. It can be observed in many habitats [37], e.g., secondary habitats developing after deforestation or fallow lands and set-aside [38,39]. According to Rothmaler [40] it occurs mainly in clearings of forest habitats and often indicates soil degradation. The nutritional value of the species is extremely low, due to its strong leaves and high level of accumulation of dead matter. Its use as fodder is extremely rare, although there are trials, mainly with goats [41].
The most important goal of our present study was to investigate the effect of multi-year mowing on the restoration of the botanical composition of the grassland dominated by C. epigejos.
In the tenth year from the start of the continuously conducted experiment, we evaluated the effect of mowing on the dominant C. epigejos cover and the occurrence of other species, as well as the changes in species richness and biomass.
We examined the following questions:
- How can the spread of dominant grass be controlled? Can mowing reduce C. epigejos coverage, and if so, how long does it take?
- How does the species number and diversity of the sward change during the experiment?
- Does the proportion of species important for grassland management change? If so, in what direction?
2. Materials and Methods
2.1. Study Area
The experimental area is located in the western part of the Cserhát Mountains, on the border of the villages of Vác, Rád, and Vácduka. It belongs to the territory of Duna-Ipoly National Park, center coordinates are: 47° 45’ 38,23” N, 19°12’ 47,53” E. The climate of the region is temperate. The annual precipitation is 520–590 mm and the average annual temperature is 8–10 °C [42].
The substrate is loess of various thicknesses deposited during the Tertiary period. Mountain ridges emerge from the loess layer, such as the Bükkös-hill (190 m above sea level). The study sites are located on the west-facing slope (15 ha) of this hill (Foto 01).
The earlier natural grassland vegetation was Salvio nemorosae-Festucetum rupicolae Zólyomi ex Soó 1964 community [43] which corresponds to Natura 2000 habitat type 6240 “Sub-pannonic steppic grasslands” [44].
In this mainly agricultural landscape, large areas have been used for centuries as arable fields, vineyards, and orchards. These areas were abandoned from the 1960s and 1970s for various economic reasons [45].
2.2. Experimental Design and Data Collecting
In our present work, we focused on the fast-spreading, perennial rhizome grass, which strongly changes the composition of the plant community. With the mowing treatment, we aimed to reduce the vegetative and generative growth potential of C. epigejos.
Eight quadrats were designated, the corners of which were permanently marked. These permanent plots were 3 × 3 m large, positioned randomly along the west slope. We arranged mown and control plots in a split-plot design. Vegetation data were monitored in 2 × 2 m large permanent quadrats placed in the middle of each plot, i.e., there was a 2 m buffer zone between the paired (mown and control) quadrats.
This experimental layout was justified by two reasons: (1) avoidance of the forest edge effect (2) selection of contiguous, homogeneous patches. The chosen small plot size allowed us to minimize topography, vegetation and land use heterogeneity, i.e., to ensure the most similar initial conditions. During our work, we performed stratified random sampling. We did not consider patches dominated by shrubs and Robinia pseudoacacia and ignored patches with C. epigejos cover less than 60%. All plots, except for the controls were mown by hand, twice a year, in June and September.
In each plot, we visually estimated the cover of the vascular plant species present, with an accuracy of 1 percent. After the plots were mowed, the biomass was removed. The coenological survey and treatments were carried out every year between 2001 and 2011. The removed biomass was subjected to further examination in 2009. The cut plant material was collected, always using the same method. The biomass tests were carried out twice a year, in June and September. After the coenological surveys were completed, the biomass was sampled from the central 1 × 1 m grassland area with a hand-held cutter. Imitating sheep grazing, since it is the most typical grazing animal, we left 4-5 cm high stubble. The biomass collected in this way was sorted according to groups important for grazing. The values and designations of the grassland management categories were as follows:
- 4.
- 1. dominant grass species ie: C. epigejos
- 5.
- 2. subordinate grass species important for grassland management
- 6.
- 3. Fabaceae species important for grassland management
- 7.
- other Dicotyledonous species neutral for grassland management
- 8.
- prickly plants
- 9.
- litter
Wet biomass was dried to constant weight in a drying cabinet. The dry biomass values of each turf management category were given in grams.
2.3. Statistical Analyses
Most of the statistical analyses were performed with R statistical software (version 4.0.5, [48]). We used “vegan”, “ tidyverse”,” ggpubr”, ” rstatix” packages. The effects of mowing were tested by repeated–measures analyses of variance (ANOVA). For pair-wise comparisons, paired t-tests were conducted to compare the treatment to the controls and to compare all treatments to each other. Shannon -diversity values was calculated with PAST 3.11 statistical software [49].
3. Results
3.1. Effects of Mowing on the Cover of C. epigejos
C. epigejos was the most important plant species in all plots at the beginning of the investigation with similar average cover (62,38% in the plots that were assigned to be mown and 69,38% in control plots). After two years of mowing treatment (in 2003) the difference became significant (p=0,0019) (Figure 1.). The significant difference that emerged in the third year of the study remained throughout the experiment. If we compare it to the starting year as a control, we can conclude that the difference in the mown plots is also significant starting from the third year. At the same time, this is not the case with control plots. (Figure 2.)
After ten years of our study, in 2011, in the mown plots there was a considerable decline of the average cover of C. epigejos (from initial 62,38% to 7,50%). In the control plots the average cover also decreased to 56,88%. The total cover of all species has not substantially altered. (Table 1.)
In 2011 in the mown plots there was a considerable increasing of the average cover of subordinated species (from initial 33,16% to 104,41%). In the control plots the average cover also increased from 38,71% to 54.85%.
3.2. Effects of Mowing on the Number of Species and Diversity
The total number of species increased during the study period. Comparing the data on the number of species on the west slope of the Bükkös hill, we found a significant difference between the mown and control plots only in the last year of the study, in 2011, p=0.00726, 10 years after the start of the experiment (Figure 3).
A significant change in the control plots compared to the initial value in 2001 was first observed in 2009, p=0.000328, and in 2011, p=0.00592.
In the mown quadrats, the treatment in the first years did not bring significant changes in the number of species. However, a significant difference (p=0.0396042) compared to the initial state was observed in 2006, and after that throughout the study. (Figure 3.)
The most important subordinate species characteristic for the study area is the Bothriochloa ischaemum (Keng.), which became the 5th with an average coverage of 7% in the mown plots after 10 years of the treatment. At the same time, it also reached the 4th rank in the control plots based on its average coverage, so a general drying and closing of the grassland can be observed in the whole study area.
At the same time, Shannon-diversity of mown plots increased only slightly. Comparing the control quadrats to 2001, we already noticed a significant difference in 2004, which, however, disappeared by the following year. It appeared again in 2007 and has been detectable since then. In the mown plots, the significant difference appeared as early as 2003, and remained throughout the entire period of the experiment. (Figure 4.)
3.3. Effects of Mowing on Biomass Composition
The distribution of the important groups from the point of view of grassland management, it can be said that on the Bükkös-hill a large amount of thorny, unpalatable species only rarely occurred, these were Eryngium campestre and Carlina vulgaris. Otherwise, this component was not typical, which occurs mainly in grazed areas. Mowing for 8 years (till 2009) has uniformly removed it from the area, in contrast to the grazing animals, which leave it and can thus promote its reproduction. The debris accumulation is significant in the case of plots 4, 5 and 6, which are located in the central region of the mountain. Dorycnium herbaceum, Astragalus glycyphyllos, Securigeria varia species of the Fabaceae family account for a large proportion, in more disturbed conditions, Vicia cracca was the most abundant (Figure 5.)
Festuca rupicola and Brachypodium pinnatum are the most important grass species that were present in greater proportion as a result of mowing. Since their forage value is greater than that of C. epigejos, the total value of the grassland has also increased. The cover of Bothriochloa ischaemum has also increased, but this does not contribute to the usability of the grassland, as its forage value is the same as that of C. epigejos.
In the control samples, the large amount of litter is noticeable, the average measured amount is 256 g in the western area of Bükkös-hill (Figure 6). Regarding the proportion of dicotyledons, the group of Fabaceae as well as other Poaceae species, we measured a smaller biomass. In contrast, C. epigejos was present in large quantities in the control samples.
4. Discussion
In our mowing experiment, treatment twice a year effectively reduced the cover of C. epigejos. However, the significant decrease of the dominant species only occurred in the 3rd year. A similar phenomenon was also reported from Germany, Rebele & Lehmann [24] in their article wrote about a 2-year delay. The slow decline of C. epigejos coverage can be attributed to the nutrient reserves accumulated in the rhizomes [32,50,51]. The dominant species lost a large amount of its biomass and thus its competitiveness as a result of the frequent cutting [50]. Several studies reported that C. epigejos becomes established in the early stages of succession [39,52,53,54]. These results suggest that regular mowing (twice a year) is likely to be a disturbance of sufficient magnitude to deplete storage organs and cause a state of nutrient deficiency.
In this study we showed that mowing increased species number and Shannon diversity in regenerating, secondary grasslands. However, the response to the mowing treatment was slow: increases in species number occurred after several years and increase of Shannon diversity took even more years. We got a similar result in another survey, where the exposition was different [47]. Most studies reported that mowing increases the species richness of abandoned grasslands [55,56,57,58]. C. epigejos is a tall, clonal spreading grass with a large amount of standing dead matter. This dead biomass inhibits the germination and growth of other species. By removing the litter, subordinate species could grow. This is reflected in both the number of species and the values of diversity. Our results supported the results of previous similar studies and indicated that grasslands can adapt very successfully to different forms of human land use [54].
A rapid rearrangement of the dominance ranking of the species could be observed on the treated plots. The importance of long-term monitoring was highlighted also and has been previously reported by Sierka & Kopczynska [59]. Parallel to the suppression of the dominant species that make up the matrix of the grassland, we observed the spread of subordinate grass and dicotyledonous species. It is an interesting phenomenon that the increase in the absolute coverage of the other (subordinate) species was also observed in the control plots. This was mainly due to the increase in the cover of shrubs. Analyzing the change in the proportion compared to the total cover (relative cover), we revealed that the subordinate species occurred in relatively greater numbers on the mown plots. Due to mowing, the microclimate of the area also changes, it becomes drier. Because of this, the broad-leaved grass species lose their dominance, so the cover of narrow-leaved species, e.g., Festuca, increased. This observation is consistent with the results of other studies, e.g., [60,61]. Due to regular mowing, the height of the grassland was reduced, therefore more sunlight reached the soil surface, influencing drying [21,62].
Mowing twice a year significantly increased the number of plant species and to a lesser extent the diversity. Several authors reported similar results, e.g., Szépligeti et al. [63] and Piseddu et al. [64]. suggested that grassland vegetation has adapted to the traditional human land-use. Other surveys show that moderate disturbance increases species richness [65]. Increasing diversity, species richness and grassland yield is an important element in the usability of grasslands, and maintaining a healthy structure plays an important role in the risk of invasion.
5. Conclusions
- In our 10-year experiment, we found significant impact of treatment on the abundance of C. epigejos, along with the changes in the order of importance of the main grassland species.
- With this study we confirmed our observation that the retraction of C. epigejos also occurs spontaneously in later stages of succession. Although C. epigejos began to decline spontaneously, its dominance without treatment remained for a longer period.
- Species richness increased faster in the case, when plots were mowed.
- Regular mowing can improve the biomass composition and forage value of secondary dry grasslands.
Author Contributions
Conceptualization, S.B. and J. H; Methodology, S.B.,., D.P. and J.H.; Software, S.B. K.P. and J. H; Formal analysis, J.H. and S.B.; Investigation, S.B., D.P., and K.P; Data curation, K.P., J.H., D.P.; Writing—original draft preparation, S.B., D.P and J.H.; Writing—review and editing: All authors; Visualization, D.P. and J.H.; All authors have read and agreed to the published version of the manuscript.
Funding
There was no financial support.
Acknowledgments
We thank Margit Dávid for assistance during the field work, Bernadett Zsinka for helping with the statistical analysis. We acknowledge the general support of the Duna-Ipoly National Park Directorate.
Data Availability Statement
Data are available from the first author upon reasonable request.
Nomenclature
For species nomenclature we used Vascular plants of Hungary: ferns - flowering plants (Király 2009)
Conflicts of Interest
The authors declare no conflict of interest.
References
- Molnár, Z.; Biró, M.; Bartha, S.; Fekete, G. Past trends, present state and future prospects of Hungarian forest-steppes. In Eurasian steppes. Ecological problems and livelihoods in a changing world. Werger, M.J.A. and Staalduinen, M.A. (Eds.). New York: Springer Science & Business Media, 2012, 6, 209–252. [Google Scholar]
- Dengler, J.; Janišová, M.; Török, P.; Wellstein, C. Biodiversity of Palaearctic grasslands: A synthesis. Agric. Ecosyst. Environ. 2014, 182, 1–14. [Google Scholar] [CrossRef]
- Willner, W.; Kuzemko, A.; Dengler, J.; Chytrý, M.; Bauer, N.; Becker, T.; Biţă-Nicolae, C.; Botta-Dukát, Z.; Čarni, A.; Csiky, J.; et al. A higher-level classification of the Pannonian and western Pontic steppe grasslands (Central and Eastern Europe). Appl. Veg. Sci. 2016, 20, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Török, P.; Penksza, K.; Tóth, E.; Kelemen, A.; Sonkoly, J.; Tóthmérész, B. Vegetation type and grazing intensity jointly shape grazing effects on grassland biodiversity. Ecol. Evol. 2018, 8, 10326–10335. [Google Scholar] [CrossRef]
- Poschlod, P.; Wallis de Vries, M.F. The historical and socioeconomic perspective of calcareous grasslands–lessons from the distant and recent past. Biol. Conserv. 2002, 104, 361–376. [Google Scholar] [CrossRef]
- Ruprecht, E.; Szabó, A.; Enyedi, M.Z.; Dengler, J. Steppe-like grasslands in Transylvania (Romania): Characterisation and influence of management on species diversity and composition. Tuexenia 2009, 29, 353–368. [Google Scholar]
- Pärtel, M.; Bruun, H.H.; Sammul, M. Biodiversity in temperate European grasslands: Origin and conservation. Grassland science in Europe, 2005, 10 (1), 14.
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45. [Google Scholar] [CrossRef]
- Fiala, K.; Holub, P.; Sedláková, I.; Tůma, I.; Záhora, J.; Tesařová, M. Reasons and consequences of expansion of Calamagrostis epigejos in alluvial meadows of landscape affected by water control measures. Ekologia 2003, 22 (Suppl. S2), 242–252. [Google Scholar]
- Bartha, S. Composition, differentiation and dynamics in the forest steppe biome. In Slope Steppes, Loess Steppes and Forest Steppe Meadows in Hungary. Illyés, E.; Bölöni, J. Eds. Budapest, Hungary: MTA ÖBKI ISBN: 978-963-06-3673-5; 2007; pp. 194–211.
- Virágh, K.; Horváth, A.; Bartha, S.; Somodi, I. A multi scale methodological approach novel in monitoring the effectiveness of grassland management. Commun. Ecol. 2008, 9, 237–246. [Google Scholar] [CrossRef]
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; Chan, K.M.A.; et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 2019, 366, 1327. [Google Scholar] [CrossRef]
- Lindborg, R. Recreating Grasslands in Swedish Rural Landscapes – Effects of Seed Sowing and Management History. Biodivers. Conserv. 2005, 15, 957–969. [Google Scholar] [CrossRef]
- Mendenhall, C.D.; Karp, D.S.; Meyer, C.F.J.; Hadly, E.A.; Daily, G.C. Predicting biodiversity change and averting collapse in agricultural landscapes. Nature 2014, 509, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Semenchuk, P.; Essl, F.; Lenzner, B.; Moser, D.; Blackburn, T.M.; Cassey, P.; Biancolini, D.; Capinha, C.; Dawson, W.; et al. The impact of land use on non-native species incidence and number in local assemblages worldwide. Nat. Commun. 2023, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jendrisakova, S.; Kováčiková, Z.; Vargová, V.; Michalec, M. The impact of cattle and sheep grazing on grassland in Veľká Fatra National Park. J. Water Land Dev. 2011, 15, 61–67. [Google Scholar] [CrossRef]
- Kuzmanović, N.; Šinžar-Sekulić, J.; Lakušić, D. Leaf anatomy of the Sesleria rigida Heuffel ex Reichenb. (Poaceae) in Serbia. Botanica Serbica 2009, 33 (1), 51-67.
- Bonanomi, G.; Incerti, G.; Allegrezza, M. Assessing the impact of land abandonment, nitrogen enrichment and fairy-ring fungi on plant diversity of Mediterranean grasslands. Biodivers. Conserv. 2013, 22, 2285–2304. [Google Scholar] [CrossRef]
- Tardella, F.M.; Malatesta, L.; Goia, I.G.; Catorci, A. Effects of long-term mowing on coenological composition and recovery routes of a Brachypodium rupestre-invaded community: Insight into the restoration of sub-Mediterranean productive grasslands. Rendiconti Lince- Sci. Fis. e Nat. 2018, 29, 329–341. [Google Scholar] [CrossRef]
- Hejcman, M.; Češková, M.; Schellberg, J.; Pätzold, S. The Rengen Grassland Experiment: Effect of Soil Chemical Properties on Biomass Production, Plant Species Composition and Species Richness. Folia Geobot. 2010, 45, 125–142. [Google Scholar] [CrossRef]
- Catorci, A.; Ottaviani, G.; Ballelli, S.; Cesaretti, S. Functional differentiation of Central Apennine grasslands under mowing and grazing disturbance regimes. Pol. J. Ecol. 2011, 59, 115–128. [Google Scholar]
- Szentes, S.; Sutyinszki, Z.; Szabó, G.; Zimmermann, Z.; Járdi, I.; Házi, J.; Bartha, S.; Penksza, K. Studies on the effects of old world bluestem (Bothriochloa ischaemum (L. ) Keng 1936) on species composition of grassland with microcoenological methods. Anim. Welf. Ethol. Hous. Syst. 2012, 8, 88–102. [Google Scholar]
- Rebele, F. Calamagrostis epigejos (L. ) Roth auf anthropogenen Standorten–ein Überblick. Verh. Der Ges. Für Okol. 1996, 26, 753–763. [Google Scholar]
- Rebele, F.; Lehmann, C. Biological Flora of Central Europe: Calamagrostis epigejos (L.) Roth. Flora 2001, 196, 325–344. [Google Scholar] [CrossRef]
- Gloser, V.; Košvancová, M.; Gloser, J. Changes in growth parameters and content of N-storage compounds in roots and rhizomes of Calamagrostis epigejos after repeated defoliation. Biol. Bratisl. 2004, 59, 179–184. [Google Scholar]
- Prach, K.; Pyšek, P. Using spontaneous succession for restoration of human-disturbed habitats: Experience from Central Europe. Ecol. Eng. 2001, 17, 55–62. [Google Scholar] [CrossRef]
- Huhta, A.; Pasi, R.; Tuomi, J.; Laine, K. Restorative mowing on an abandoned semi-natural meadow: Short-term and predicted long-term effects. J. Veg. Sci. 2001, 12, 677–686. [Google Scholar] [CrossRef]
- Sedláková, I.; Fiala, K. Ecological degradation of alluvial meadows due to expanding Calamagrostis epigejos. Ekologia 2001, 20 (Suppl. S3), 226–333. [Google Scholar]
- Kompała-Bąba, A.; Sierka, E.; Dyderski, M.K.; Bierza, W.; Magurno, F.; Besenyei, L.; Błońska, A.; Ryś, K.; Jagodziński, A.M.; Woźniak, G. Do the dominant plant species impact the substrate and vegetation composition of post-coal mining spoil heaps? Ecol. Eng. 2019, 143, 105685. [Google Scholar] [CrossRef]
- Zhukovskaya, O.V.; Ulanova, N.G. Influence of brushing frequency on birch population structure after felling. Écoscience 2006, 13, 219–225. [Google Scholar] [CrossRef]
- Csontos, P. Light ecology and regeneration on clearings of sessile oak-Turkey oak forests in the Visegrád Mountains, Hungary. Acta Bot. Hung. 2010, 52, 265–286. [Google Scholar] [CrossRef]
- Fiala, K.; Tůma, I.; Holub, P. Effect of nitrogen addition and drought on above-ground biomass of expanding tall grasses Calamagrostis epigejos and Arrhenatherum elatius. Biologia 2011, 66, 275–281. [Google Scholar] [CrossRef]
- Błońska, E.; Kempf, M.; Lasota, J. Woody debris as a substrate for the growth of a new generation of forest trees. For. Ecol. Manag. 2022, 525. [Google Scholar] [CrossRef]
- Prach, K. Succession of vegetation on dumps from strip coal mining, N. W. Bohemia, Czechoslovakia. Folia Geobot. et Phytotaxon. 1987, 22, 339–354. [Google Scholar] [CrossRef]
- Bartha, S. Preliminary scaling for multi-species coalitions in primary succession. Abstr. Bot. 1992, 16, 31–41. [Google Scholar]
- Baasch, A.; Tischew, S.; Bruelheide, H. Twelve years of succession on sandy substrates in a post-mining landscape: A Markov chain analysis. Ecol. Appl. 2010, 20, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Hultén E, Fries M. Atlas of North European vascular plants: North of theTropic of Cancer I-III. 1986, Koeltz Scientific Books, Königstein,.
- Csecserits, A.; Rédei, T. Secondary succession on sandy old-fields in Hungary. Appl. Veg. Sci. 2001, 4, 63–74. [Google Scholar] [CrossRef]
- Bartha, S.; Szentes, S.; Horváth, A.; Házi, J.; Zimmermann, Z.; Molnár, C.; Dancza, I.; Margóczi, K.; Pál, R.W.; Purger, D.; et al. Impact of mid-successional dominant species on the diversity and progress of succession in regenerating temperate grasslands. Appl. Veg. Sci. 2013, 17, 201–213. [Google Scholar] [CrossRef]
- Rothmaler,W. Exkursionsflora von Deutschland. Aufl. n Volk und Wissen Verlag. Berlin,1988, Bd. 3. n 7.
- Hajnáczki, S.; Pajor, F.; Péter, N.; Bodnár. ; Penksza, K.; Póti, P. Solidago gigantea Ait. and Calamagrostis epigejos (L) Roth invasive plants as potential forage for goats. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12197. [Google Scholar] [CrossRef]
- Marosi, S.; Somogyi, S. (Eds.) Magyarország Kistájainak Katasztere (Cadastral of Microregions of Hungary); MTA Földrajztudományi Kutatóintézet: Budapest, Hungary, 1991; pp. 379–388. [Google Scholar]
- Borhidi, A.; Kevey, B.; Lendvai, G. Plant communities of Hungary. Akadémiai Kiadó, Budapest, Hungary, 2012, 544 pp.
- European Environment Agency. 6250 Pannonic loess steppic grasslands. Report under the Article 17 of the Habitats Directive Period 2007-2012 Available online:. Available online: https://eunis.eea.europa.eu/habitats/10124#sites (accessed on 20 December 2022).
- Házi, J.; Bartha, S.; Szentes, S.; Wichmann, B.; Penksza, K. ; Seminatural grassland management by long-term mowing of Calamagrostis epigejos in western Cserhát, Hungary (Management sekundärer Trockenrasen durch Langzeit-Mahd von Calamagrostis epigejos im westlichen Cserhát, Ungarn). In. Steppenlebensräume Europas–Gefährdung, Erhaltungsmassnahmen und Schutz; Thüringer Ministerium für Landwirtschaft, Forsten, Umwelt und Naturschutz (TMLFUN), Erfurt, ISBN 978-3-00-044248-3. Baumbach, H., Pfützenreuter, S. (eds.); 2013; pp. 331–340.
- Házi, J.; Bartha, S.; Szentes, S.; Wichmann, B.; Penksza, K. Seminatural grassland management by mowing of Calamagrostis epigejos in Hungary. Plant Biosyst. 2011, 145, 699–707. [Google Scholar] [CrossRef]
- Md. Nazmul Islam, Kusay Faisal Al-tabatabaie, Md. Ahasan Habib, Sheikh Sharif Iqbal, Khurram Karim Qureshi and Eid M. Al-Mutairi, Design of a Hollow-Core Photonic Crystal Fiber Based Edible Oil Sensor, Crystals 2022, 7, 101. [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, Vienna, Austria, 2013, ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 20 February 2018).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Klimeš, L.; Klimešova, J. The effects of mowing and fertilisation on carbohydrate reserves and regrowth of grasses: Do they promote plant coexistence in species-rich meadows? Evol Ecol 2002, 15, 363–382. [Google Scholar] [CrossRef]
- Kavanová, M.; Gloser, V. The Use of Internal Nitrogen Stores in the Rhizomatous Grass Calamagrostis epigejos During Regrowth After Defoliation. Ann. Bot. 2004, 95, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Prach, K. Succession of vegetation on dumps from strip coal mining, N. W. Bohemia, Czechoslovakia. Folia Geobot. et Phytotaxon. 1987, 22, 339–354. [Google Scholar] [CrossRef]
- Bartha, S. Preliminary scaling for multi-species coalitions in primary succession. Abstr. Bot. 1992, 16, 31–41. [Google Scholar]
- Gerwin, W.; Raab, T.; Birkhofer, K.; Hinz, C.; Letmathe, P.; Leuchner, M.; Roß-Nickoll, M.; Rüde, T.; Trachte, K.; Wätzold, F.; et al. Perspectives of lignite post-mining landscapes under changing environmental conditions: What can we learn from a comparison between the Rhenish and Lusatian region in Germany? Environ. Sci. Eur. 2023, 35, 1–24. [Google Scholar] [CrossRef]
- Bobbink, R.; During, H.J.; Schreurs, J.; Willems, J.; Zielman, R. Effects of selective clipping and mowing time on species diversity in chalk grassland. Folia Geobot. et Phytotaxon. 1987, 22, 363–376. [Google Scholar] [CrossRef]
- Bobbink, R.; Willems, J. Impact of different cutting regimes on the performance of Brachypodium pinnatum in Dutch chalk grassland. Biol. Conserv. 1991, 56, 1–21. [Google Scholar] [CrossRef]
- Fenner, M. & Palmer, L. Grassland management to promote diversity: Creation of patchy sward by mowing and fertiliser regimes. Field Studies 1998, 9: 313–324.
- Valkó, O.; Zmihorski, M.; Biurrun, I.; Loos, J.; Labadessa, R.; Venn, S. Ecology and Conservation of Steppes and Semi-Natural Grasslands. Hacquetia 2016, 15, 5–14. [Google Scholar] [CrossRef]
- Sierka, E.; Kopczynska, S. Participation of Calamagrostis epigejos (L. ) Roth in plant communities of the River Bytomka valley in terms of its biomass use in power industry. Environ. Socio-Econ. Stud. 2014, 2, 38–44. [Google Scholar]
- Neuhaus, Ryan Allan, "A comparison of the effects of burning, haying and mowing on plants and small mammals in a tallgrass prairie reconstruction" (2015). Dissertations and Theses @ UNI. 216. https://scholarworks.uni.edu/etd/216.
- Tardella, F.M.; Bricca, A.; Piermarteri, K.; Postiglione, N.; Catorci, A. Context-dependent variation of SLA and plant height of a dominant, invasive tall grass (Brachypodium genuense) in sub-Mediterranean grasslands. Flora 2017, 229, 116–123. [Google Scholar] [CrossRef]
- Oelmann, Y.; Broll, G.; Hölzel, N.; Kleinebecker, T.; Vogel, A.; Schwartze, P. Nutrient impoverishment and limitation of productivity after 20 years of conservation management in wet grasslands of north-western Germany. Biol. Conserv. 2009, 142, 2941–2948. [Google Scholar] [CrossRef]
- Szépligeti, M.; Kőrösi, Á.; Szentirmai, I.; Házi, J.; Bartha, D.; Bartha, S. Evaluating alternative mowing regimes for conservation management of Central European mesic hay meadows: A field experiment. Plant Biosyst. - Int. J. Deal. all Asp. Plant Biol. 2018, 152, 90–97. [Google Scholar] [CrossRef]
- Piseddu, F.; Bellocchi, G.; Picon-Cochard, C. Mowing and warming effects on grassland species richness and harvested biomass: Meta-analyses. Agron. Sustain. Dev. 2021, 41, 1–21. [Google Scholar] [CrossRef]
- Sheil, D.; Burslem, D.F. Defining and defending Connell's intermediate disturbance hypothesis: A response to Fox. Trends Ecol. Evol. 2013, 28, 571–572. [Google Scholar] [CrossRef] [PubMed]
Foto 01.
The study site: Bükkös-hill (Cserhát Mountains) (Foto: Judit Házi).

Figure 1.
Change of cover of C. epigejos in the control and mown plots in Bükkös hill west slopes, during the 2001-2011 period, M = mown plot (grayish boxes), C = control plot (open boxes). Significant differences between mown and control plots in the same year are marked by * (p<0.05).
Figure 1.
Change of cover of C. epigejos in the control and mown plots in Bükkös hill west slopes, during the 2001-2011 period, M = mown plot (grayish boxes), C = control plot (open boxes). Significant differences between mown and control plots in the same year are marked by * (p<0.05).
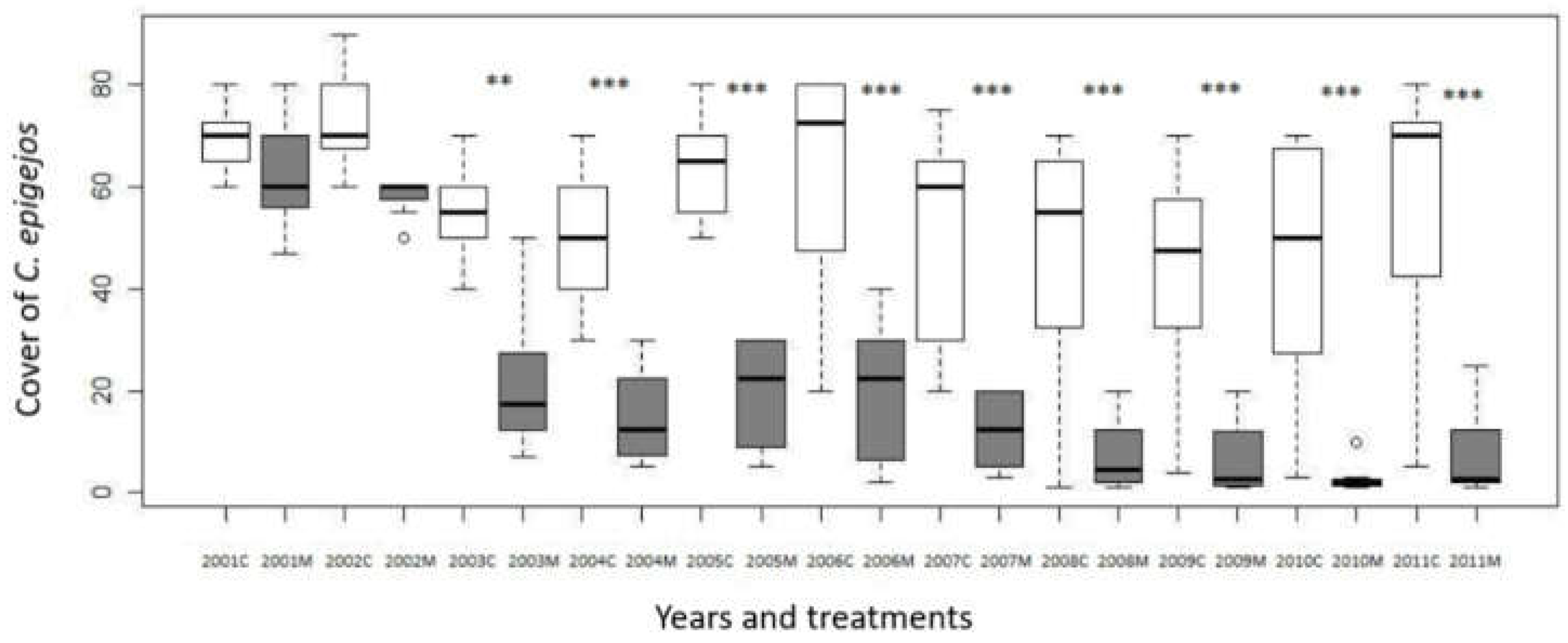
Figure 2.
Change of cover of C. epigejos in the mown and control plots in Bükkös-hill west slopes during the 2001-2011 period between mown and control plots and staring 2001 years and subsequent years. Significant differences are marked by * (p<0.05).
Figure 2.
Change of cover of C. epigejos in the mown and control plots in Bükkös-hill west slopes during the 2001-2011 period between mown and control plots and staring 2001 years and subsequent years. Significant differences are marked by * (p<0.05).
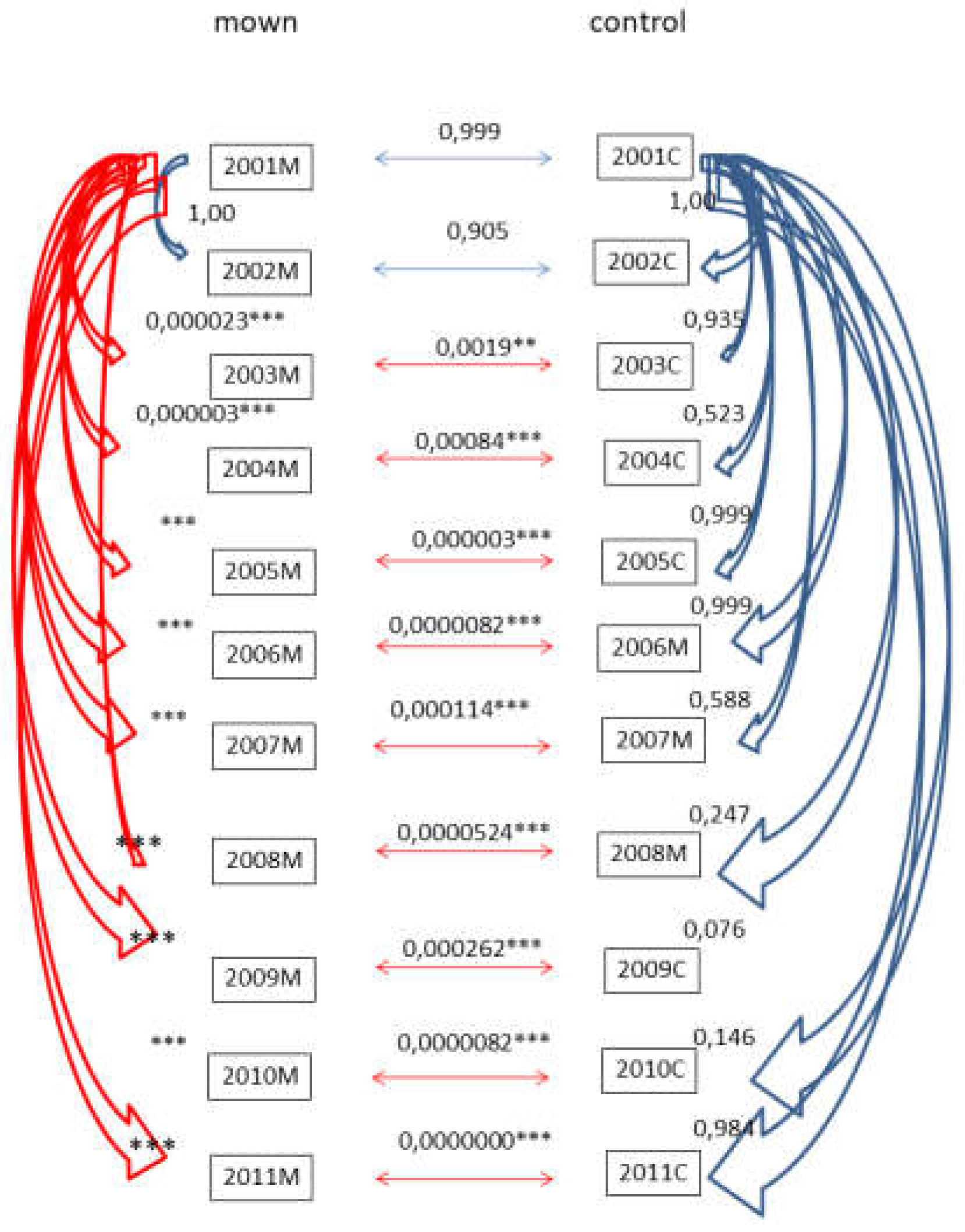
Figure 3.
Change of number of species in the mown and control plots in Bükkös-hill west slopes during the 2001-2011 period, M = mown plot (grayish boxes), C = control plot (open boxes). Significant differences between mown and control plots in the same year are marked by * (p<0.05).
Figure 3.
Change of number of species in the mown and control plots in Bükkös-hill west slopes during the 2001-2011 period, M = mown plot (grayish boxes), C = control plot (open boxes). Significant differences between mown and control plots in the same year are marked by * (p<0.05).
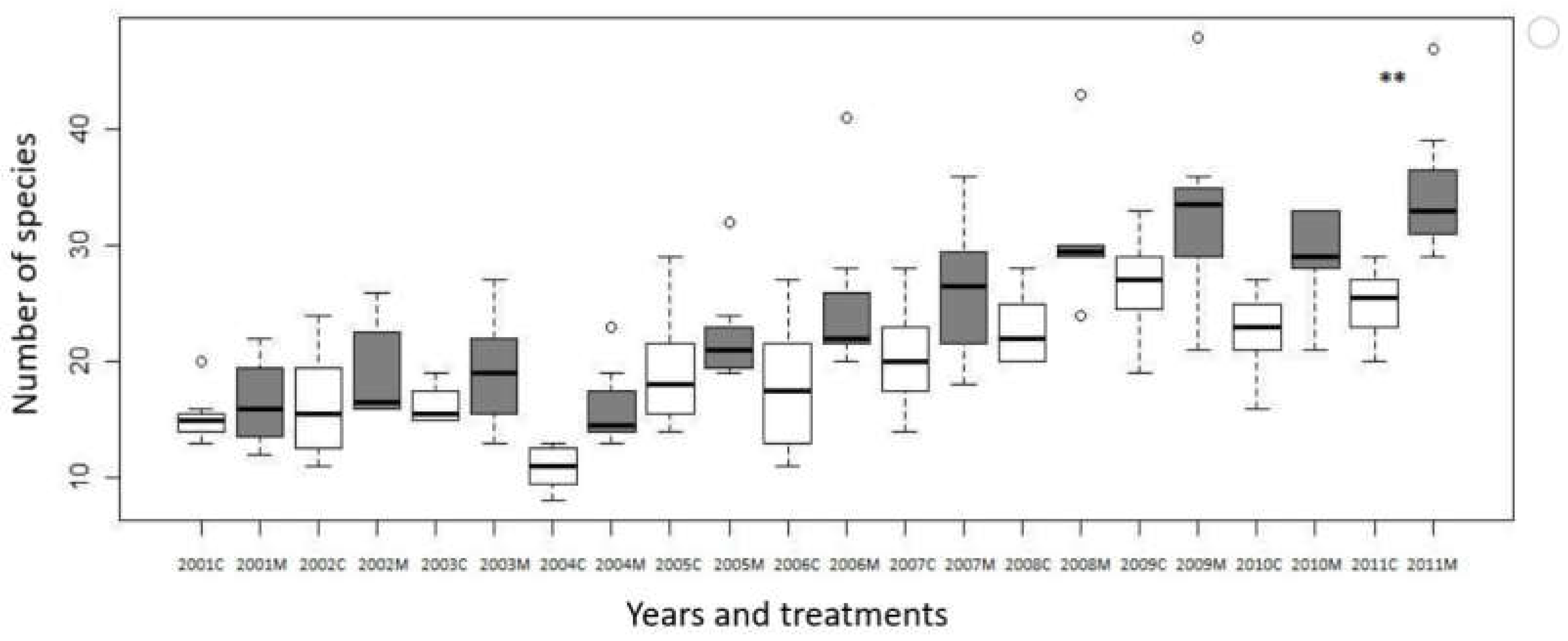
Figure 4.
Change of Shannon-diversity in the mown and control plots in Bükkös-hill west slopes during the 2001-2011 period, M = mown plot (red boxes), C = control plot (open boxes).
Figure 4.
Change of Shannon-diversity in the mown and control plots in Bükkös-hill west slopes during the 2001-2011 period, M = mown plot (red boxes), C = control plot (open boxes).
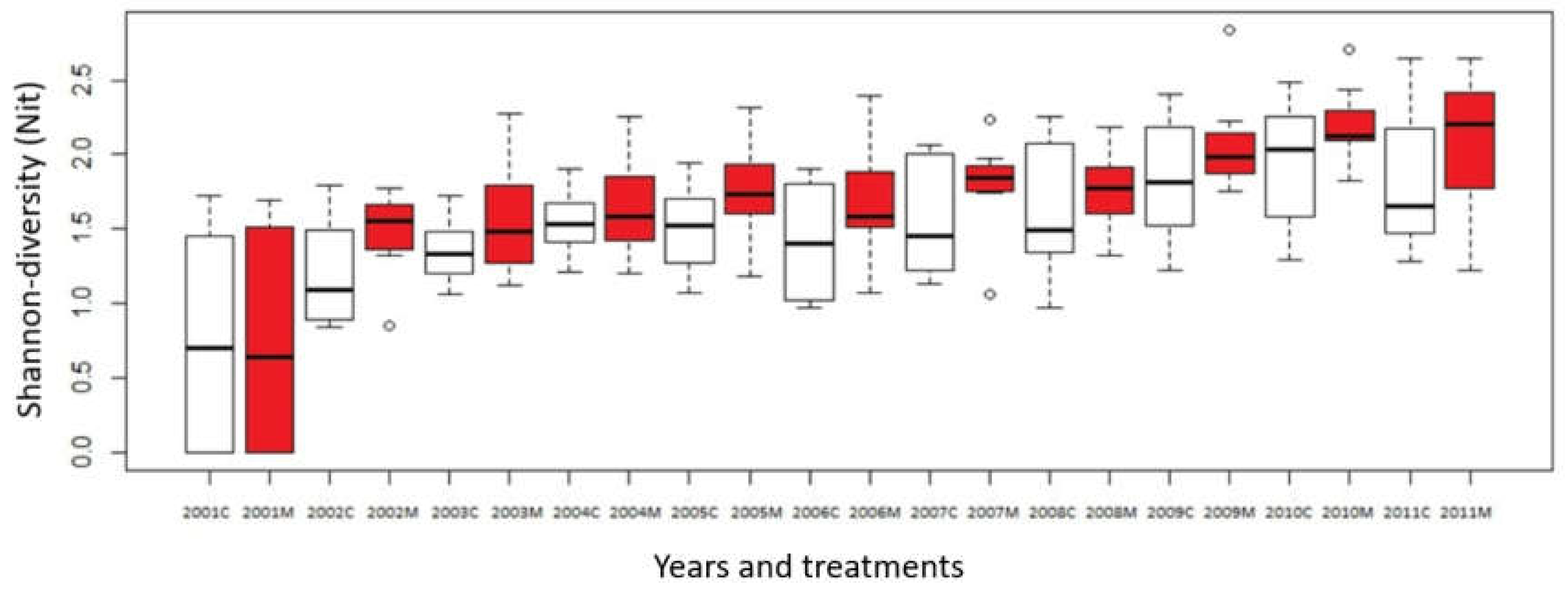
Figure 5.
Biomass composition of the mown plots in 2009.
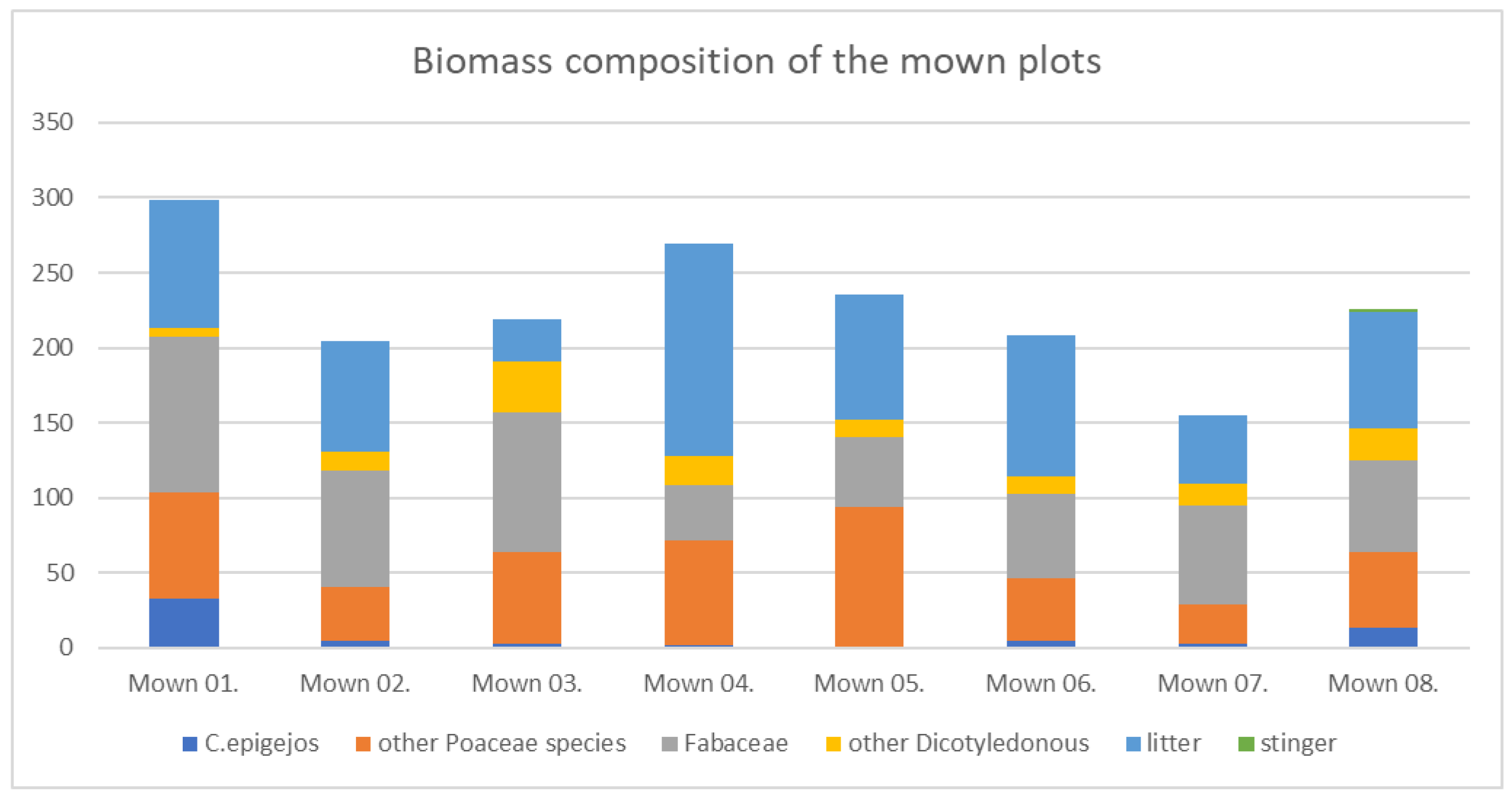
Figure 6.
Biomass composition of the mown and control plots.
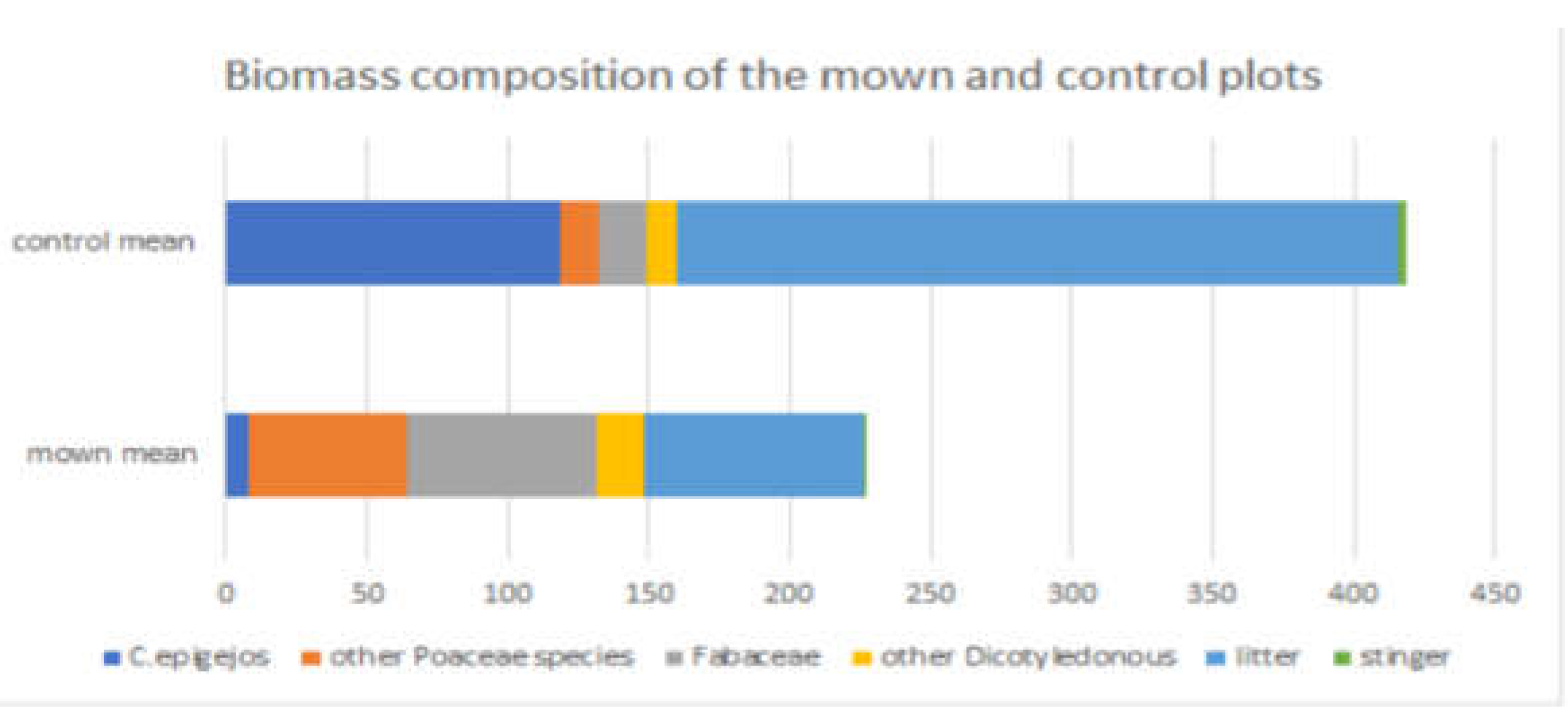

Table 1.
The average cover of different parameters of mown and control plots. “Total cover absolute” = cover of all vascular plant species; “Cover of C. epigejos absolute” = the cover of Calamagrostis epigejos; “Cover of C. epigejos relative” = the cover of Calamagrostis epigejos relative to the overall coverage; “the cover of subordinate species absolute and relative” = cover of all species except Calamagrostis epigejos, “Number of species” = number of all vascular plant species.
Table 1.
The average cover of different parameters of mown and control plots. “Total cover absolute” = cover of all vascular plant species; “Cover of C. epigejos absolute” = the cover of Calamagrostis epigejos; “Cover of C. epigejos relative” = the cover of Calamagrostis epigejos relative to the overall coverage; “the cover of subordinate species absolute and relative” = cover of all species except Calamagrostis epigejos, “Number of species” = number of all vascular plant species.
| MOWN | CONTROL | |||||||||||
| 2001 | 2006 | 2011 | 2001 | 2006 | 2011 | |||||||
| Mean | Mean | Mean | Mean | Mean | Mean | |||||||
| Total cover absolute | 95,54 | 102,18 | 111,91 | 108,09 | 108,03 | 111,71 | ||||||
| Cover of C. epigejos absolute | 62,38 | 20,00 | 7,50 | 69,38 | 62,50 | 56,88 | ||||||
| Cover of C. epigejos relative | 0,65 | 0,19 | 0,06 | 0,64 | 0,58 | 0,51 | ||||||
| Cover of subordinated species absolute | 33,16 | 82,18 | 104,41 | 38,71 | 45,53 | 54,84 | ||||||
| Cover of subordinated species relative | 0,35 | 0,81 | 0,94 | 0,36 | 0,42 | 0,49 | ||||||
| Number of species | 16,50 | 25,00 | 34,63 | 15,25 | 17,75 | 25,00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



