
Submitted:
11 April 2024
Posted:
15 April 2024
You are already at the latest version
Abstract
Food-borne transmission is a recognized route for many viruses associated with gastrointestinal, hepatic or neurological diseases. Therefore, it is essential to identify new bioactive compounds with broad-spectrum antiviral activity is needed to exploit innovative solutions against these hazards. Lately, antimicrobial peptides (AMPs) have been recognized as promising antiviral agents. Indeed, while the antibacterial and antifungal effects of these molecules have been widely reported, their use as potential antiviral agents has not yet been fully investigated. Herein, the antiviral activity of previously identified or newly designed AMPs, was evaluated against the non-enveloped RNA viruses, Hepatitis A virus (HAV), and Murine Norovirus (MNV), a surrogate for human Norovirus. Moreover, specific assays were performed to recognize at which stage of the viral infection cycle the peptides could function. Results showed that almost all peptides displayed virucidal effects with about 90% of infectivity reduction on HAV or MNV. However, the decapeptide RiLK1 demonstrated, together with its antibacterial and antifungal properties, a notable reduction in viral infection for both HAV and MNV, possibly through direct interaction with viral particles or inhibition of the viral bonding to the cell receptors. Hence, RiLK1 could represent a versatile ‘antimicrobial agent effective against various foodborne pathogens including viruses, bacteria, and fungi.
Keywords:
Antimicrobial peptide
; Enteroviruses
; Non-enveloped virus
; Virucidal activity
; Foodborne viral infections
; Hepatitis A Virus
; Murine Norovirus
1. Introduction
According to the World Health Organization (WHO), foodborne diseases are a major safety concern worldwide, affecting approximately 30% of the population in developed countries annually [1]. Each year, foodborne illnesses affect over 600 million people worldwide and result in over 420,000 deaths, 125,000 of which are in children [2].
Most food-related diseases are a result of microbial contamination, principally from bacteria, fungi, and viruses. Pathogens can be transferred to food products through contact with contaminated surfaces, especially with biofilms, which are more resistant to biocidal chemicals and conventional methods for microbial cleaning than free-living cells, as they are generally composed of multiple species showing a special physiology and physical matrix barrier [3,4,5,6]. As a result, preventing or eliminating contamination in food-processing environments, especially when ready-to-eat foods are handled, represents an arduous challenge. In this context, virus contamination, that is one of the important causes of foodborne illness worldwide, is recognized as a top priority among the microbiological food safety risks [7]. Specifically, NoroVirus (NoV) and Hepatitis A Virus (HAV) are considered to have a very relevant public health impact [8,9]. They are leading agents of acute gastroenteritis due to its low infection dose, wide transmission route, strong environmental resistance, and ease of contaminating products such as fruit, lettuce and shellfish causing foodborne outbreaks [10,11,12,13,14]. Moreover, inadequate hygiene and waste disposal practices in food processing, coupled with substandard manufacturing methods, can strongly increase the risk of viral contamination in foods [15,16,17,18,19].
In the food industry, different sanitising technologies are applied such as the use of radiation, heat, different chemicals, gaseous ozone [20], gaseous chlorine dioxide [21], and high hydrostatic pressure [22]. Nevertheless, these solutions are often effective only for a limited time and present a lot of disadvantages, encouraging to explore innovative and alternative approaches to improve cleaning procedures and food safety. Indeed, in recent years several studies have been concentrated on less toxic sanitising solutions, involving the identification of new natural compounds to develop sustainable and efficient cleaning procedures for food processing, transformation and storage surfaces able to prevent the initial microbial attachment and eliminate the microorganisms from the food contact materials.
Recently, antimicrobial peptides (AMPs), also known as host-defence peptides, have attracted greatly attention [17]. AMPs are natural molecules present in the innate immune system of almost all living organisms, invertebrates, and vertebrates, identified as potential agents with therapeutic effects as they exhibit marked antibacterial, antiviral, antiparasitic and antifungal properties. Generally, the AMPs derived from different microorganisms, including plants, arthropods, reptiles, and amphibians [16,23,24,25] or are synthetically generated. AMPs represent a heterogeneous class, that shares several main physicochemical characteristics: (i) the presence of basic amino acids conferring a net positive charge; (ii) the presence of 50% hydrophobic amino acids; and (iii) the ability to assume alpha-helix or beta-sheet conformations during the interaction with the target cell membrane. To date, more than 3000 AMPs have been officially classified and registered in the AMP database with over 300 discovered in frog skin [26,27].
Previous studies have reported that antiviral AMPs target various stages in the viral life cycle, especially attachment, entry and fusion [28,29]. The attention has been directed towards identifying peptides targeting viral proteins [28]. AMPs can also neutralise both RNA and DNA viruses by incorporating into the viral envelope [30,31], causing membrane instability and disruption, and preventing the host cell infections [32,33]. AMPs such as defensins can also decrease the binding of viruses to host cells by interacting with viral glycoproteins and inhibiting the attachment of herpes simplex viruses (HSV) to the surface of host cells [34]. Additionally, some antiviral AMPs can prevent the entry of the viral particles into the host cells by binding to specific mammalian receptors [35,36], while others can cross the cell membrane, prompting changes in the gene expression profile of the host cells, and stimulating their defence system against viruses [37,38].
Temporins, derived from Rana temporaria, are peptides consisting of 10-13 amino acid residues with a slight cationic charge. In detail, their action is principally focused on enveloped viruses, including HSV-1 [39,40,41] and influenza A [17], although the mechanism of action is different: (i) some can act at the intracellular level by destroying the viral envelope and/or preventing the virus-membrane host cell fusion (influenza virus, HSV-1); or (ii) others can interfere in viral replication by preventing the release of new virions (parainfluenza viruses) [17,18].
Compared to the enveloped viruses, the absence of a lipid envelope in the naked variants may affect the antiviral mechanisms and the effectiveness of antiviral compounds. This may explain why there are few reports on the action of AMPs against non-enveloped viruses. In a previous study, the antiviral effects of cathelicidin-derived peptides were evaluated against non-enveloped Enterovirus 71 (EV71) with a single-stranded positive-sense RNA, belonging to Picornaviridae family [38]. Some of these compounds had already shown antiviral activity against naked viruses in vitro, such as human rhinovirus and adenovirus [38]. In this case, it was demonstrated that the mechanisms of action can be also more complex and can involve different steps of the infection.
We have recently demonstrated the antibacterial activity of a panel of designed AMPs against food pathogen contaminants, including Gram-positive and Gram-negative bacteria and fungi but not viruses [42,43,44,45,46]. The structural properties and mechanism of action of these peptides have been elucidated previously and these compounds were also found to be non-cytotoxic [42,43,44,45,46].
The objective of the present study was to evaluate the virucidal effect of the same antimicrobial peptides that were previously evaluated against bacteria, on two non-enveloped foodborne viruses, MNV-1 and HAV. Moreover, to expand the panel of peptides, two additional compounds have been included in the tests against the viruses, the newly designed RiLK30 and the already known HIV inhibitor AVP2 [47], which were structurally characterized. Attempts were also undertaken to elucidate the mechanism of action of two of these peptides.
Hence, the present work aims to consolidate the current knowledge and findings on the antiviral properties of antimicrobial peptides and their potential applications.
2. Materials and Methods
2.1. Virus Strains and Cell Lines
Murine norovirus strain 1 (MNV-1) was used as a surrogate for human NoroVirus (NoV). The virus was replicated in RAW 264.7 cells (murine macrophage cell line) cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 1% glutamine, 1% non-essential amino acids and 2% fetal bovine serum (FBS). Hepatitis A virus (HAV) strain HM-175 was replicated in Frp3 cells (non-human primate cell line derived from Fetal kidney) cultured in Minimum Essential Medium with Earle’s salts (MEM) supplemented with 1% glutamine, 2% non-essential amino acids and 2% FBS. Both the viruses were incubated in 5% CO2 at 37°C. All cell culture media were provided by EuroClone (Milan, Italy). The viral suspensions were prepared by freeze and thaw lysis (1 cycle for MNV-1 and 3 cycles for HAV) of infected monolayers, clarified using low speed centrifugation (800 ×g) to remove residual cell debris, then divided in aliquots and stored at -80 °C until use. The obtained HAV and MNV-1 stock suspensions had a final concentration of 4.7± 0.2 log TCID50/ml and 4.5± 0.2 log TCID50/ml, respectively, calculated by determining the 50% tissue culture infectious dose with the Reed and Muench method [48], using tenfold serial dilutions in 24-well plates.
2.2. Antimicrobial Peptides
The antimicrobial peptides (AMPs) tested in the study were: RiLK1, RiLK3, RiLK30, 1018-K6, MTP1, and AVP2. The peptides were resuspended in 5 mM DMSO to prepare the peptide stock solutions and stored at -20 °C until use. The following web server and software were used for determining the main physicochemical properties of peptides: PlifePred (PPred) [49], Antimicrobial Peptide Database3 (APD3) [50], and Meta-iAVP [51].
2.3. Circular Dichroism (CD) Spectroscopy
Circular Dichroism (CD) analysis was performed by Jasco J-810 spectropolarimeter. The samples were loaded into a quartz cuvette of 0.1 cm path length (Hellma Analytics) and the spectra were recorded in the 195 nm–250 nm range at a scan speed of 20 nm/min, by averaging 5 scans. CD experiments were carried out in 10 mM Tris-HCl buffer pH 7.0 as a function of SDS concentration (3, 50 and 150 mM) at a peptide concentration of 50 µM. The folding kinetic measurements of the peptides were performed after the addition of SDS (50 mM) to each sample (50 µM in 10 mM Tris-HCl, pH 7.0) up to 24 h incubation. For thermal stability analyses, the peptides were prepared to a final concentration of 50 μM in 10 mM Tris-HCl, pH 7.0 and then they were incubated at 4 and 25 °C up to 48 h before acquiring the CD spectra in the presence of 50 mM SDS. A blank spectrum of a sample containing all components except the peptide was acquired for the baseline-correction of the CD spectra of the peptide. The mean residue ellipticity ([θ], deg cm2 dmol−1) was obtained by the equation [θ]=100 θ/cnl, where θ is the ellipticity (mdeg), c is the peptide concentration (mM), n is the number of residues, and l is the path length (cm).
2.4. Fluorescence Spectroscopy
Tryptophan (Trp) fluorescence emission spectra were recorded at 25 °C on a Shimadzu RF-6000 spectrofluorometer (Kyoto, Japan) with both excitation and emission slit widths set at 5 nm. The intrinsic Trp was excited at a wavelength of 280 nm and the emission was monitored between 300 and 400 nm. Fluorescence measurements were carried out in 10 mM Tris-HCl buffer pH 7.0 as a function of SDS concentration (3, 50 and 150 mM) at a peptide concentration of 50 µM. The folding kinetic experiments of AVP2 and RiLK30 were performed after the addition of SDS (50 mM) to each sample (50 µM concentration in 10 mM Tris-HCl buffer pH 7.0) up to 24 h incubation. The effect of temperature on peptide folding was analyzed by dissolving the peptides at a final concentration of 50 µM in 10 mM Tris-HCl buffer pH 7.0 and incubating them at 4 and 25 °C up to 48 h. The spectra were recorded in the presence of 50 mM SDS.
2.5. Ionic Strength Stability
The effect of ionic strength on the peptide stability was examined by reverse-phase (RP) High-Performance Liquid Chromatography (HPLC) analysis. The antimicrobial molecules at a final concentration of 50 µM were incubated in a water solution containing 150 mM sodium chloride up to 96 hours at 25 °C. For the analyses, 200 µL of the sample solutions were recovered at different times and loaded onto a µBondapak C18 reverse-phase column (3.9 mm x 300 mm, Waters Corp., Milford, MA, USA) connected to a HPLC system (Shimadzu, Milan, Italy) using a linear gradient of 0.1% trifluoroacetic acid (TFA) in acetonitrile from 5 to 95%. A reference solution at time 0 (t=0) was prepared under the same reaction conditions and run in parallel. The peptide stability in saline solution was evaluated by comparing the peak area of the peptide at the different incubation times with that of the peptide at t=0.
2.6. Cytotoxicity Determination of Peptides on Cells
To identify the peptide concentrations that had no cytotoxic effect, preliminary tests were performed on cell cultures. Cell viability was evaluated by the 3-(4,5-dimethilthiazol-2-yl)-2,5-dipheniltetrazolium bromide (MTT) assay [52]. Peptide solutions were prepared in serum free MEM at different concentrations in the range from 10 to 100 µM (10, 20, 40, 80, 100 µM), treated overnight at 4 °C with antibiotic/antimycotic solution (Euroclone) and 100 µL were assayed on 24-48 h cell monolayers in a 24-well plate. The monolayers added with the peptides were incubated for 1 h at 37 °C in 5% CO2. Thereafter, cells were washed twice with Dulbecco’s Phosphate Buffer Solution (DPBS, EuroClone) and maintained in culture with DMEM/MEM supplemented with 2% of FBS for 48 h in 5% CO2 at 37 °C. After that, the medium was removed and 300 µl of MTT (Sigma Aldrich, Milan, Italy) solution (5 mg/ml) was added. The monolayers were incubated for 15-30 min at 37 °C in 5% CO2, the MTT was removed and 500 µl DMSO was added to each well to dissolve the purple formazan crystals. The absorbance was measured at 570 nm. DMSO and culture medium were used as controls.
2.7. Virucidal Effect of Peptides
Solutions 40 µM and 80 µM of each peptide of the panel (RiLK1, RiLK3, RiLK30, 1018-K6, MTP1, AVP2) were chosen for virucidal tests. HAV and MNV-1 were assayed at a concentration of 4.7 ± 0.2 log TCID50/mL and 4.5 ± 0.2 log TCID50/mL, respectively. The suspensions containing the peptides and viruses were incubated for 1 hour at room temperature (RT,20 ± 2°C) and at 4 °C. These temperatures have been chosen to simulate the use of sanitizing solutions in food production/processing environments. Residual viral infectivity was evaluated by titration (TCID50/ml) on Frp3 cells for HAV and on RAW 264.7 cells for MNV-1. Untreated HAV and MNV-1 suspensions and 40 µM and 80 µM AMPs solutions, incubated at same conditions, were used as controls. Each treatment condition was assayed in triplicate. For titration, 100 µl of serial tenfold dilutions of each sample were assayed in 24-well tissue culture plates containing 24-48 h cell monolayers, and incubated for 1 h in 5% CO2 at 37 °C. After that, the wells were washed twice with 200 µl of PBS and 500 µl of DMEM/MEM, supplemented with 2% of FBS, were added to each well; MNV-1 and HAV incubations were carried for up to 6 days and 14 days respectively in 5% CO2 at 37 °C with daily visual inspection to determine the cytopathic effect. Virucidal efficacy of AMPs was estimated by comparing the titres of treated to untreated viral suspensions.
2.8. Antibacterial Assays
The monophasic variant of Salmonella Typhimurium, Salmonella Napoli and Campylobacter jejuni were isolated by the Laboratory of Microbiological Food Control—Department of Food Microbiology of the Istituto Zooprofilattico Sperimentale del Mezzogiorno in Portici (Naples, Italy) in raw and processed foodstuffs of animal origin. Specifically, the monophasic variant of S. Typhimurium and S. Napoli were isolated from bivalve mollusks, while C. jejuni was isolated from poultry meat. Bacterial cells were cultured at 37 °C in the appropriate culture media (S. Typhimurium and S. Napoli in buffered peptone water (Thermo Fisher, Milan, Italy); C. jejuni in Bolton Broth (Oxoid, Madrid, Spain), until collection and then diluted in fresh broth to final concentration of 1.5 × 103 colony forming units (CFU)/mL. Thereafter, peptide stock solutions in DMSO were added to the bacterial suspension at the same concentrations used in the virucidal assays (40 and 80 µM) and incubated at 37 °C for 6 h. Samples containing only cell suspension and DMSO were used as controls. Therefore, 50 μL of each bacterial cell suspensions were transferred onto selective agar plates (S. Typhimurium and S. Napoli, Salmonella Chromogenic Agar - Oxoid, Madrid, Spain); C. jejuni, modified Charcoal Cefoperazone Deoxycholate Agar (Liofilchem, Roseto degli Abruzzi, Italy) in microaerophilic conditions and incubated for 24 h at 37 °C for S. Typhimurium and S. Napoli (ISO 6579-1:2020), and 48 h at 41 °C for C. jejuni (ISO 10272-2:2017). All values were obtained as the mean of three independent experiments conducted in triplicate.
2.9. Mechanism of Action of Rilk1 and Rilk3 Peptides
Four sets of experiments were performed to identify the step of viral infection [29] in which RiLK1 and RiLK3 exerted their activity on HAV and MNV-1. To this aim, 80 µM solutions of the two peptides, and HAV and MNV-1 suspensions at a final concentration of 4.7 ± 0.2 log TCID50/mL and 4.5 ± 0.2 log TCID50/mL, respectively, were used in the following experiments:
- i)
- virus pre-treatment assay, to assess the interaction between virus and peptide: incubation of peptide with viral suspension for 1 h at room temperature followed by incubation on cells for 1 h at 37 °C. Then, cell monolayers were washed twice with PBS and incubated with 500 µl DMEM/MEM supplemented with 2% of FBS to determine the TCID50/ml;
- ii)
- cell pre-treatment assay, to investigate peptide action on cells: cell monolayers were preincubated with 100 µl of peptide solution for 1 h at 37 °C followed by removal of the peptide, addition to cells of viral suspensions for 1 h at 37 °C, washing step and culture with 2% FBS DMEM/MEM;
- iii)
- attachment assay, to evaluate peptide effect on viral attachment to cells: cell monolayers were incubated with 100 µl of a suspension containing both peptide and virus, and were incubated for 1 h at 4 °C. Subsequently, the cell monolayers were washed with PBS and incubated for 1 h at 37 °C with a culture medium followed by a washing step and addition of culture with 2% FBS DMEM/MEM;
- iv)
- entry assay, to explore peptide activity on the internalization of viruses into cells: cell monolayers were incubated with 100 µl of the viral suspension for 1 h at 4 °C, followed by addition of the peptide and incubation for 1 h at 37 °C. Cells were then washed with PBS and 2% FBS DMEM/MEM was finally added.
Untreated viral suspensions and peptide solutions, incubated at the same conditions, were used as controls.
2.10. Statistical Analysis
For each treatment, the average and standard deviation of the triplicate analyses were calculated. Virucidal effects of AMPs was assessed by calculating the Log Reduction Value (LRV), i.e. the difference between the infectious titre (log transformed) of untreated versus treated samples. The statistical significance of differences between treated and untreated samples was determined through one-way analysis of variance (ANOVA) with Bonferroni post hoc comparisons, with a significance level of p < 0.05 (GraphPad Prism v9.5.0, software San Diego, CA, USA).
3. Results and Discussion
3.1. Peptide Design
Due to the lack of drugs for the treatment of many viral infections and the increase of multidrug-resistant viruses, there is a high urgency to develop new compounds to fight these pathogens [53]. Recently, different studies have shown that antimicrobial peptides (AMPs) can also exert antiviral effects in addition to the well-known anti-bacterial or anti-fungal activities [38,54,55], thus making them ideal candidates for the development of antiviral drugs.
In our previous studies, the structural characterization as well as the antibacterial and antifungal activities of the peptides 1018-K6, MTP1, RiLK1, and RiLK3 were demonstrated [42,43,44,45,46], thus leading to investigations of their possible antiviral effects that have not been studied so far, considering that the ultimate goal of research is to find agents that exhibit a broad-spectrum antimicrobial activity and are therefore attractive for manufacturing applications. Moreover, two further peptides were considered in our study to expand the panel of potential antiviral compounds (Table 1), the newly designed RiLK30 and the already-known HIV inhibitor AVP2 [47], which was used as a reference. Concerning RiLK30, it was selected among a series of RiLK1-derivatives that were rationally designed by following specific and systematic amino acid substitutions in the parental RiLK1 sequence that, based on our own experience, is crucial to preserve certain structural features, like the balance between the positive charges, hydro-phobicity, and content of lipophilic residues such as Trp, that promote the peptide-microbial membrane interactions and maintain the antimicrobial activity [56].
It is worth noting that RiLK1 was chosen as the lead compound among the other peptides available because of its broad spectrum of antimicrobial activity, its rapid production at low cost, being a smaller peptide than 1018-K6 and MTP-1, thus representing the best candidate to accelerate further studies and develop new molecules. The reliability of the new set of analogs in terms of antiviral activity was predicted using Meta-iAVP software [51], and based on this analysis, RiLK30 variant, where all the Arginine (R) residues within the RiLK1 sequence were exchanged with Lysine (K) and the single Lys at position 3 with Arg, was selected for the in vitro antimicrobial tests. The physico-chemical characteristics of all the peptides under investigation predicted using the available software PlifePred, APD3, and Meta-iAVP [49,50,51], are reported in Table 1.
3.2. Structural Characterization of RiLK30 and AVP2
As for the antibacterial activity, the interaction of AMPs with membrane components frequently correlates with antiviral effects, suggesting that the presence of a membrane could be one of the essential conditions in inducing the peptide conformation capable of inhibiting viral infection [57,58,59].
In this context, to acquire important information on how the conformational properties of peptides might influence the structure−function relationships and their distinct membrane-interaction profiles, the propensity of the new peptides, RiLK30 and AVP2, to adopt a defined secondary structure and to undergo conformational changes were examined by circular dichroism and fluorescence spectroscopy. These analyses were performed in presence of the negatively charged SDS that is commonly used as biomimetic membrane model to study the structural features of membranotropic molecules. In previous studies, the structure and the conformational stability of 1018-K6, MTP1, RiLK1 and RiLK3 have been already elucidated in detail [42,43,44,45,46]. Briefly, all these four peptides exhibited a high structural stability across a wide range of pH (2–11) and temperature (4–90 °C) but they assumed different conformations in mimicked membrane solutions such as mixed α-helix/β-sheet for MTP1, β-sheet for 1018-K6 and higher-ordered self-aggregates, assembling into bigger oligomeric species in equilibrium with partially structured monomers for RiLK1 and RiLK3.
Concerning RiLK30 and AVP2, in a first set of experiments the CD spectra were recorded at a fixed peptide concentration (50 μM) in 10 mM Tris-HCl buffer (pH 7.0) and in the absence or presence of SDS solutions, below and above the critical micelle concentration. As shown in Figure 1, both peptides present a dicroic spectrum in aqueous solution with the typical shape of a random coil structure characterized by a negative band at ~200 nm. Instead, the presence of SDS significantly affects the CD spectrum of peptides, that underwent a variation in shape when in contact with the oppositely charged amphiphile. Specifically, AVP2 adopts a typical α-helical conformation in SDS solutions at concentrations both lower (3 mM) and higher (50 and 150 mM) than the micellar one (Figure 1A). Conversely, RiLK30 showed a more complex conformation that was not correlated with the common secondary structure elements (α-helix, β-strand, or random coil) (Figure 1B), and could be associated with its high propensity to self-aggregate, as already observed for its analogues RiLK1 and RiLK3 [42,43,44].
These results were also confirmed by fluorescence spectroscopy analyses, which demonstrated the structuring of the two peptides induced by the microenvironment surrounding the tryptophan which becomes more hydrophobic following the addition of the detergent at different concentrations (Figure 2A,B).
Subsequently, to study the conformational rearrangements of RiLK30 and AVP2 over time, the molecules were incubated for 24 h at 37 °C and pH 7.0 after which the dichroic (Figure 3A,B) and fluorescence (Figure 3C,D) spectra were acquired in the presence of SDS 50 mM. These analyses highlighted that the peptides do not appear to undergo significant changes in the following 24 h, once their conformational distribution have been assumed (t=0).
It has been pointed out that the antimicrobial activity of some AMPs is greatly weakened by some physical parameters, such as temperature and salt. Therefore, the conformational and structural stability of RiLK30 (Figure 4) and AVP2 (Figure 5) in response to different temperature conditions were verified by incubating the peptides for 48 h at pH 7.0 and at 4 °C or 25 °C. The results obtained following spectroscopic analyses revealed that the thermal treatment is not able to induce conformational changes in both peptides, confirming their high thermal stability for at least 48 h in SDS micellar solution.
As far as the stability in saline solution is concerned, AVP2 and RiLK30 were incubated in the presence of NaCl (150 mM) up to 96 h and at each incubation time the solutions were quantitatively monitored by Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC). The analysis of the corresponding chromatographic profiles highlighted that AVP2 is structurally 'stable' in its α-helix structure up to 48 hours of incubation in salt, after which it undergoes conformational rearrangements following to aggregation phenomena, as demonstrated by the almost total disappearance of the main peak of the peptide together with the formation of new peaks (Figure 6A). The same behavior was evidenced by RiLK30 that exhibits comparable conformational transformations but already within the first 24 h of incubation, becoming more evident over the following 72 h (Figure 6B).
3.3. Antiviral Activity of Peptides
According to the Antimicrobial Peptide Database (APD), antiviral effects have been reported for only 185 out of more than 3000 registered AMPs, also due to the more difficult evaluation of antiviral effects [50]. Thus, considering that the issue of emerging and re-emerging infectious diseases, especially those related to viruses, has become an increasingly important area of concern in public health, the discovery of novel antiviral compounds deserves additional effort.
In this context, our peptides were tested against norovirus (NoV) and Hepatitis A virus (HAV), which are considered two of the principal enteric viruses causing food and waterborne outbreaks due to their ability to give infection with very low doses and to survive in the external environment for a long time. NoVs and HAV are small non-enveloped viruses having a positive-sense, single-stranded RNA genome. For NoV, the murine species (strain MNV-1) was used in the viability studies due to the lack of in vitro cultivation methods for human norovirus (HuNoV). Indeed, MNV-1 is currently recognized as the most widely accepted surrogate model being the most closely related virus and similar to HuNoV in its size, capsid structure, genomic organization, and replication cycle [60,61].
The first step of our analysis was devoted to discriminating between cytotoxicity and virucidal effect. Therefore, to exclude the possibility that our peptides could be cytotoxic on RAW 264.7 or Frp3 cells, the host cell lines used to assess the antiviral activity against MNV-1 and HAV, different concentrations of each peptide ranging from 10 to 100 μM were tested by MTT assays. The results demonstrated no cytotoxicity in all the tested range (data not shown). Subsequenlty, 40 and 80 µM were the concentrations chosen to perform the next experiments, and the potential virucidal activity of the peptides against HAV and MNV-1 was assessed at two different temperatures.
In a previous work, the antiviral effects of RiLK1 and RiLK3 were already investigated against the hepatitis A virus (HAV) following the treatment at room temperature [46] and the obtained results have been reported in Table 2. Briefly, RiLK1 was able to reduce the HAV infectivity more than 1 log in comparison with the untreated virus, displaying a Log Reduction Value (LRV) of 1.4 at 80 μM and of 1.1 at 40 μM concentration [46]. Conversely, RiLK3 was unable to inhibit HAV infection at the same concentrations tested. Therefore, based on these preliminary results, further insights into the virucidal activity of both RiLK1/RilK3 and the other peptides were provided in this study (Table 2 and Table 3). With respect to RiLK1 and RiLK3, the results obtained on HAV at 4 °C were comparable to those previously obtained at room temperature [46] (Table 2).
As regards MNV-1 (Table 3), a virucidal effect was observed only for RiLK1 after treatment at room temperature (LRV of 1.2 and 1.0 at 80 µM and 40 µM, respectively), while RiLK3 was not effective regardless of the concentration or temperature condition applied. RiLK30 showed no virucidal activity in any of the condition tested, neither toward HAV nor MNV-1. (Table 3).
On the other hands, 1018-K6 and MTP1 displayed to be effective on HAV at 40 µM and 80 µM, and at the two temperatures, causing an infectivity reduction of 1.3-1.4 log for 1018-K6 and 0.9 to 1.5 log for MTP-1. However, no significant effects were exhibited by these peptides toward MNV-1. Conversely, AVP2 was active against MNV-1 at both temperatures (LRV of 0.9 and 1.0 log), but provided a limited reduction of infectivity on HAV.
In terms of reduction of viral infectivity, HAV infectious titre was decreased up to 96.7% (1.4 LRV) by RiLK1 and 1018-K6, and up to 97.3% (1.5 LRV) by MTP1. Instead, MNV-1 infectivity was reduced up to 94.1% (1.2 LRV) by RiLK1, and up to 91.3% (1.0 LRV) by AVP2. Therefore, only RiLK1 showed effectiveness toward both viruses.
Based on these in vitro results, RiLK1 was chosen as a model molecule for an in-depth analysis to elucidate the mechanism of its antiviral action, using as control the analogue RiLK3, differing by only one amino acid, to understand the significant changes in virucidal activity observed between the two peptides. To do aim, four types of assays, varying in the timing of addition of the drug, were performed as described in paragraph 2.5 (virus pre-treatment assay, cell pre-treatment assay, attachment assay, entry assay). Results are reported in Table 4 for HAV and in Table 5 for MNV-1.
Consistently with the previous results, RiLK1 displayed a reduction of the infectious titre of 1.4 log for HAV and 1.2 log for MNV-1 in the virus pre-treatment assay through direct interaction with viral particles. Similarly, a LRV of 1.4 for both HAV and MNV-1 was shown in the attachment assay through the inhibition of virus attachment to the cell’s receptors.
Furthermore, RilLK1 also exhibited activity in preventing the entry of MNV-1 into the cells (entry assay, LRV 1.0). On the other hand, RiLK3 revealed a more pronounced antiviral action through the interaction with the cells (cell pre-treatment assay, LRV 0.9 and 1.0 for HAV and MNV-1, respectively), and not directly on the virus. Moreover, RiLK3 also affected the viral attachment to cells (LRV 1.0 for both HAV and MNV-1), and inhibited MNV-1 entry in the host cells (Table 5).
Considering that RiLK3 derives from RiLK1 by replacing a single lysine residue with arginine, it is important to underline how small variations in the peptide structure can determine significant changes in their antiviral properties, affecting the mechanism of action. In this case, the unique amino acid substitution makes RiLK3 no longer able to interact directly with the virus particles.
The obtained results are particularly interesting as the use of peptides against non-enveloped viruses is limited, as most AMPs act during the viral fusion to the cell membrane, exclusive of enveloped viruses. Therefore, there is still more work to be done in the design of inactivators against this class of pathogens because the structure and function of the receptor-binding domains in their capsid proteins have not been completely elucidated [62].
3.4. Antibacterial Activity of Peptides
The zoonoses that occur most frequently in the industrialised world today are food-borne infections caused by bacteria enzootic to food animals such as Campylobacter and Salmonella. As a result, campylobacteriosis and salmonellosis have emerged as a critical global health problem with a high socioeconomic burden. Hence, it is important to identify novel ‘weapons’ to reduce and/or prevent these infections.
In this study, all the designed peptides were tested against Campylobacter jejuni, the monophasic variant of S. Typhimurium (MVST) and Salmonella Napoli, three of the most relevant zoonotic pathogens reported to be strongly involved in human infections in the EU. As shown in Figure 7, all peptides exhibited a strong bactericidal activity towards both serovars of Salmonella, being able to completely inhibit the growth of bacteria already at 40 μM concentration excepted for MTP-1 that exerted no detectable killing effects against Salmonella Napoli even at the highest concentration tested (80 μM). Concerning Campylobacter jejuni strains, only AVP2 and 1018-K6 presented 100% inhibition already at a concentration of 40 µM while the RiLK-group resulted active only at 80 µM concentration with RiLK30 being the most ineffective. By contrast, MTP-1 did not display any inhibitory activity at both concentrations.
These findings open interesting future applications of these peptides in mixture exploiting the high potential of their synergistic action to treat various foodborne microbial infections.
4. Conclusions
The lack of therapeutic agents against many important viral infections along with the ever-growing problematic of drug resistance constitute a public health emergency of international concern, requiring the urgent discovery novel antimicrobial molecules, possibly with a low toxicity and limited costs. In this scenario, the natural compounds AMPs might provide a solution in preventing and controlling food-borne diseases. To date, few studies have been performed to investigate how AMPs interact with viruses and how such interaction affects viral infectivity [29,38,63].
In the present study, the antiviral activity of six synthetic peptides with different structural features was evaluated for the first time against two non-enveloped enteric viruses, HAV and MNV-1, a surrogate for human Norovirus, together with the mechanism of action of two of them. Our results demonstrated that RiLK1 is the most promising peptide among those tested, showing a remarkable virucidal effect against both viruses by interacting directly with the viral particles or inhibiting the viral bonding to the cell receptors. These findings together with those previously obtained against bacteria and fungi, could candidate RiLK1 as a new promising ‘multifaceted’ agent with an attractive applicative potential, due to its broad spectrum of antimicrobial activity. This peptide could be suitable as a natural surface and food sanitizer to prevent or reduce contamination by foodborne pathogens. However, further studies will be necessary to investigate the mechanism of action of all the other peptides to determine their antiviral activity and to evaluate a possible synergistic effect when used in a mixture.
Author Contributions
Conceptualization, L.C., G.P., Y.T.R.P., E.S, and M.G.; methodology, E.G., B.A., A.M., T.V., S.M., and S.D.P.; software, G.P., and M.G.; validation, M.G., G.P., Y.T.R.P., S.D.P., and L.C.; formal analysis, E.G., B.A., A.M., T.V., and S.M.; resources, G.P., Y.T.R.P., and L.C.; data curation, G.P., and M.G.; writing—original draft, M.G., G.P., L.C., Y.T.R.P., and E.S.; writing—review and editing, M.G., G.P., E.S., and L.C.; project administration, G.P., Y.T.R.P., and L.C.; funding acquisition, G.P., and Y.T.R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministero della Salute—"Dalle superfici agli alimenti: nuove soluzioni per alimenti più sicuri (NewSan)"—Ricerca Corrente 2020 project, grant number IZS ME 06/20 RC; project PNRR—PE00000003, PE10—Modelli per un’alimentazione sostenibile, “ON Foods” (Research and Innovation Network on Food and Nutrition Sustainability, Safety and Security—Working ON Foods).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in the study are available in this manuscript.
Acknowledgments
The authors would like to thank Dr. Chiara Nobile for administrative assistance and Sig. Antonio Suppa for the technical support.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on Major Food-Borne Zoonotic Bacterial Pathogens. J. Trop. Med 2020, 2020, 4674235. [Google Scholar] [CrossRef] [PubMed]
- Tohma, K.; Saito, M.; Pajuelo, M.J.; Mayta, H.; Zimic, M.; Lepore, C.J.; Ford-Siltz, L.A.; Gilman, R.H.; Parra, G.I. Viral intra-host evolution in immunocompetent children contributes to human norovirus diversification at the global scale. Emerg. Microbes Infect. 2021, 10, 1717–1730. [Google Scholar] [CrossRef] [PubMed]
- Refaat, A. Biodiesel production using solid metal oxide catalysts. Int. J. Environ. Sci. Te. 2011, 8, 203–221. [Google Scholar] [CrossRef]
- Pagán, R.; García-Gonzalo, D. Influence of environmental factors on bacterial biofilm formation in the food industry: A review. J.P. R. 2015, 3, 3–13. [Google Scholar]
- Rossi, C.; Chaves-López pez, C.; Serio, A.; Casaccia, M.; Maggio, F.; Paparella, A. Effectiveness and mechanisms of essential oils for biofilm control on food-contact surfaces: An updated review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2172–2191. [Google Scholar] [CrossRef]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Rowe, G.; Bolger, F. Final report on “the identification of food safety priorities using the Delphi technique”. EFSA Support. Publ. 2016, 13. [Google Scholar] [CrossRef]
- Wang, X.; Ren, J.; Gao, Q.; Hu, Z.; Sun, Y.; Li, X.; Rowlands, D.J.; Yin, W.; Wang, J.; Stuart, D.I.; et al. Hepatitis A virus and the origins of picornaviruses. Nature 2015, 517, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Ludwig-Begall, L.F.; Mauroy, A.; Thiry, E. Noroviruses-The State of the Art, Nearly Fifty Years after Their Initial Discovery. Viruses 2021, 13, 1541. [Google Scholar] [CrossRef]
- Payne, D.C.; Vinje, J.; Szilagyi, P.G.; Edwards, K.M.; Staat, M.A.; Weinberg, G.A.; Hall, C.B.; Chappell, J.; Bernstein, D.I.; Curns, A.T.; et al. Norovirus and medically attended gastroenteritis in US children. N. Engl. J. Med. 2013, 368, 1121–1130. [Google Scholar] [CrossRef]
- Yu, Z.; Shao, Q.; Xu, Z.; Chen, C.; Li, M.; Jiang, Y.; Cheng, D. Immunogenicity and Blocking Efficacy of Norovirus GII. Recombinant P Protein Vaccine. Vaccines 2023, 11, 1053. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, M.; van Beek, J.; Koopmans, M.P. Human norovirus transmission and evolution in a changing world. Nat. Rev. Microbiol. 2016, 14, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Ruscher, C.; Faber, M.; Werber, D.; Stark, K.; Bitzegeio, J.; Michaelis, K.; Sagebiel, D.; Wenzel, J.J.; Enkelmann, J. Resurgence of an international hepatitis A outbreak linked to imported frozen strawberries, Germany, 2018 to 2020. Euro Surveill. 2020, 25, 1900670–1900679. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.P.; Weng, M.K.; Hofmeister, M.G.; Moore, K.L.; Doshani, M.; Kamili, S.; Koneru, A.; Haber, P.; Hagan, L.; Romero, J.R.; et al. Prevention of Hepatitis A Virus Infection in the United States: Recommendations of the Advisory Committee on Immunization Practices, 2020. MMWR Recomm. Rep. 2020, 69, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.K.; Mishra, N.; Saxena, R. Advances in antiviral drug discovery and development: Part II: Advancements in antiviral drug development. Future Virol. 2009, 4, 209–215. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A venom-derived peptide with promising anti-viral properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Casciaro, B.; Genovese, A.; Brancaccio, D.; Marcocci, M.E.; Novellino, E.; Carotenuto, A.; Palamara, A.T.; Mangoni, M.L.; Nencioni, L. Temporin G, an amphibian antimicrobial peptide against influenza and parainfluenza respiratory viruses: Insights into biological activity and mechanism of action. FASEB J. 2021, 35, e21358. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.C.; Feng, H.; Lin, Y.C.; Guo, X.R. New strategies against drug resistance to herpes simplex virus. Int. J. Oral Sci. 2016, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The Amphibian Antimicrobial Peptide Temporin B Inhibits In Vitro Herpes Simplex Virus 1 Infection. Antimicrob. Agents Chemother. 2018, 62, e02367–17. [Google Scholar] [CrossRef]
- Brie, A.; Boudaud, N.; Mssihid, A.; Loutreul, J.; Bertrand, I.; Gantzer, C. Inactivation of murine norovirus and hepatitis a virus on fresh raspberries by gaseous ozone treatment. Food Microbiol. 2018, 70, 1–6. [Google Scholar] [CrossRef]
- Kingsley, D.H.; Pérez-Pérez, R.E.; Niemira, B.A.; Fan, X. Fan Evaluation of gaseous chlorine dioxide for the inactivation of Tulane virus on blueberries. Int. J. Food Microbiol. 2018, 273, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.W.; Hsu, C.P.; Wang, C.Y. Healthy expectations of high hydrostatic pressure treatment in food processing industry. J. Food Drug Anal. 2020, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.S.; Prodhan, Z.H.; Biswas, S.K.; Le, C.F.; Sekaran, S.D. Antimicrobial peptides from different plant sources: Isolation, characterisation, and purification. Phytochemistry 2018, 154, 94–105. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Pourmand, M.R. Venom-derived peptide Mastoparan-1 eradicates planktonic and biofilm embedded methicillin-resistant Staphylococcus aureus isolates. Microb. Pathog. 2018, 119, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Carvalho Ade, O.; Gomes, V.M. Plant defensins—Prospects for the biological functions and biotechnological properties. Peptides 2009, 30, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein Pept. Lett. 2020, 27, 4–16. [Google Scholar] [CrossRef]
- Urmi, U.L.; Vijay, A.K.; Kuppusamy, R.; Islam, S.; Willcox, M.D.P. A review of the antiviral activity of cationic antimicrobial peptides. Peptides 2023, 166, 171024. [Google Scholar] [CrossRef]
- Chianese, A.; Zannella, C.; Monti, A.; De Filippis, A.; Doti, N.; Franci, G.; Galdiero, M. The broad-spectrum antiviral potential of the amphibian peptide AR-23. Int J. Mol. Sci. 2022, 23, 883. [Google Scholar]
- Bastian, A.; Schafer, H. Human alpha-defensin 1 (hnp-1) inhibits adenoviral infection in vitro. Regul. Pept. 2001, 101, 157–161. [Google Scholar]
- Horne, W.S.; Wiethoff, C.M.; Cui, C.; Wilcoxen, K.M.; Amorin, M.; Ghadiri, M.R.; Nemerow, G.R. Antiviral cyclic D,L-α-peptides: Targeting a general biochemical pathway in virus infections. Bioorg. Med. Chem. 2005, 13, 5145–5153. [Google Scholar] [CrossRef] [PubMed]
- Robinson, W.E. Jr.; McDougall, B.; Tran, D.; Selsted, M.E. Anti-hiv-1 activity of indolicidin, an antimicrobial peptide from neutrophils. J. Leukoc. Biol. 1998, 63, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, N.; Nagaraj, R. Interaction of antimicrobial peptides with biological and model membranes: Structural and charge requirements for activity. Biochim. Biophys. Acta 1999, 1462, 29–54. [Google Scholar] [CrossRef] [PubMed]
- Belaid, A.; Aouni, M.; Khelifa, R.; Trabelsi, A.; Jemmali, M.; Hani, K. In vitro antiviral activity of dermaseptins against herpes simplex virus type 1. J. Med. Virol. 2002, 66, 229–234. [Google Scholar] [CrossRef]
- Tamamura, H.; Ishihara, T.; Otaka, A.; Murakami, T.; Ibuka, T.; Waki, M.; Matsumoto, A.; Yamamoto, N.; Fujii, N. Analysis of the interaction of an anti-hiv peptide, t22 ([tyr5, 12, lys7] polyphemusin ii), with gp120 and cd4 by surface plasmon resonance. Biochim. Biophys. Acta 1996, 1298, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Song, B.H.; Lee, G.C.; Moon, M.S.; Cho, Y.H.; Lee, C.H. Human cytomegalovirus binding to heparan sulfate proteoglycans on the cell surface and/or entry stimulates the expression of human leukocyte antigen class I. J. Gen. Virol. 2001, 82, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B. Lunatusin, a trypsin-stable antimicrobial peptide from lima beans (Phaseolus lunatus L.). Peptides 2005, 26, 2086–2092. [Google Scholar] [CrossRef]
- Yu, J.; Dai, Y.; Fu, Y.; Wang, K.; Yang, Y.; Li, M.; Xu, W.; Wei, L. Cathelicidin antimicrobial peptides suppress EV71 infection via regulating antiviral response and inhibiting viral binding. Antivir. Res. 2021, 187, 105021. [Google Scholar] [CrossRef]
- Stelitano, D.; Franci, G.; Chianese, A.; Galdiero, S.; Morelli, G.; Galdiero, M. HSV membrane glycoproteins, their function in viral entry and their use in vaccine studies. In Amino Acids, Peptides and Proteins; Royal Society of Chemistry: London, UK, 2019; pp. 14–43. [Google Scholar]
- Tulassay, T.; Rascher, W. Atrial natriuretic peptide. New aspects in the regulation of blood volume and electrolyte homeostasis. Monatsschr Kinderheilkd 1986, 134, 710–715. [Google Scholar]
- Roy, M.; Lebeau, L.; Chessa, C.; Damour, A.; Ladram, A.; Oury, B.; Boutolleau, D.; Bodet, C.; Leveque, N. Comparison of Anti-Viral Activity of Frog Skin Anti-Microbial Peptides Temporin-Sha and [K(3)]SHa to LL-37 and Temporin-Tb against Herpes Simplex Virus Type 1. Viruses 2019, 11, 77. [Google Scholar] [CrossRef]
- Palmieri, G.; Balestrieri, M.; Proroga, Y.T.; Falcigno, L.; Facchiano, A.; Riccio, A.; Capuano, F.; Marrone, R.; Neglia, G.; Anastasio, A. New antimicrobial peptides against foodborne pathogens: From in silico design to experimental evidence. Food Chem. 2016, 211, 546–54. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, G.; Balestrieri, M.; Capuano, F.; Proroga, Y.T.R.; Pomilio, F.; Centorame, P.; Riccio, A.; Marrone, R.; Anastasio, A. Bactericidal and antibiofilm activity of bactenecin-derivative peptides against the food-pathogen Listeria monocytogenes: New perspectives for food processing industry. Int. J. Food Microbiol. 2018, 279, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Agrillo, B.; Proroga, Y.T.R.; Gogliettino, M.; Balestrieri, M.; Tatè, R.; Nicolais, L.; Palmieri, G. A Safe and Multitasking Antimicrobial Decapeptide: The Road from De Novo Design to Structural and Functional Characterization. Int. J. Mol. Sci. 2020, 21, 6952. [Google Scholar] [CrossRef] [PubMed]
- Falcigno, L.; D'Auria, G.; Palmieri, G.; Gogliettino, M.; Agrillo, B.; Tatè, R.; Dardano, P.; Nicolais, L.; Balestrieri, M. Key Physicochemical Determinants in the Antimicrobial Peptide RiLK1 Promote Amphipathic Structures. Int. J. Mol. Sci. 2021, 22, 10011. [Google Scholar] [CrossRef] [PubMed]
- Agrillo, B.; Porritiello, A.; Gratino, L.; Balestrieri, M.; Proroga, Y.T.; Mancusi, A.; Cozzi, L.; Vicenza, T.; Dardano, P.; Miranda, B.; et al. Antimicrobial activity, membrane interaction and structural features of short arginine-rich antimicrobial peptides. Front Microbiol. 2023, 14, 1244325. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Watson, K.M.; Peterkofsky, A.; Buckheit, R.W. Jr. Identification of novel human immunodeficiency virus type 1-inhibitory peptides based on the antimicrobial peptide database. Antimicrob. Agents Chemother. 2010, 54, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoint. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Mathur, D.; Singh, S.; Mehta, A.; Agrawal, P.; Raghava, G.P.S. In silico approaches for predicting the half-life of natural and modified peptides in blood. PLoS ONE 2018, 13, e0196829. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Schaduangrat, N.; Nantasenamat, C.; Prachayasittikul, V.; Shoombuatong, W. Meta-iAVP: A Sequence-Based Meta-Predictor for Improving the Prediction of Antiviral Peptides Using Effective Feature Representation. Int. J. Mol. Sci. 2019, 20, 5743. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Sytar, O.; Smetanska, I. Special issue "Bioactive compounds from natural sources (2020, 2021)". Molecules 2022, 27, 6, 1929. [Google Scholar] [CrossRef]
- Barlow, P. G.; Findlay, E. G.; Currie, S. M.; Davidson, D. J. Antiviral potential of cathelicidins. Future Microbiol. 2014, 9, 55–73. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, J.; Qin, Z.; Dong, C.; Yang, H.; Sun, J.; Xu, W.; Wei, L. Endogenous cathelicidin is required for protection against ZIKV-caused testis damage via inactivating virions. Antivir. Res. 2022, 198, 105248. [Google Scholar] [CrossRef]
- Mikut, R.; Ruden, S.; Reischl, M.; Breitling, F.; Volkmer, R.; Hilpert, K. Improving short antimicrobial peptides despite elusive rules for activity. BBA Biomembr. 2016, 1858, 1024–1033. [Google Scholar] [CrossRef]
- Pasupuleti, M.; Walse, B.; Svensson, B.; Malmsten, M.; Schmidtchen, A. Rational design of antimicrobial C3a analogues with enhanced effects against Staphylococci using an integrated structure and function-based approach. Biochemistry 2008, 47, 9057–9070. [Google Scholar] [CrossRef]
- Galanth, C.; Abbassi, F.; Lequin, O.; Ayala-Sanmartin, J.; Ladram, A.; Nicolas, P.; Amiche, M. Mechanism of antibacterial action of dermaseptin B2: interplay between helix-hinge-helix structure and membrane curvature strain. Biochemistry 2009, 48, 313–327. [Google Scholar] [CrossRef]
- Kim, J. S.; Joeng, J. H.; Kim, Y. Design, Characterization, and Antimicrobial Activity of a Novel Antimicrobial Peptide Derived from Bovine Lactophoricin. J. Microbiol. Biotechnol. 2017, 27, 759–767. [Google Scholar] [CrossRef]
- Kamarasu, P.; Hsu, H.Y.; Moore, M.D. Research progress in viral inactivation utilizing human norovirus surrogates. Front. Sustain. Food Syst. 2018, 2, 89. [Google Scholar] [CrossRef]
- Wobus, C.E.; Thackray, L.B.; Virgin, H.W. , 4th. Murine norovirus: A model system to study norovirus biology and pathogenesis. J. Virol. 2006, 80, 5104–5112. [Google Scholar] [CrossRef]
- Day, P. M.; Schelhaas, M. Concepts of papillomavirus entry into host cells. Curr. Opin. Virol. 2014, 4, 24–31. [Google Scholar] [CrossRef]
- Findlay, E.G.; Currie, S.M.; Davidson, D.J. Cationic host defense peptides: potential as antiviral therapeutics. BioDrugs 2013, 27, 479–493. [Google Scholar] [CrossRef]
Figure 1.
Effect of SDS concentration on the secondary structure of AVP2 and RiLK30 monitored by circular dichroism. Far-UV CD spectra of (A) AVP2 and (B) RiLK30 at 50 µM concentration and 25 °C in 10 mM Tris-HCl buffer pH 7.0. in the absence (blue lines) or presence of SDS at different concentrations.
Figure 1.
Effect of SDS concentration on the secondary structure of AVP2 and RiLK30 monitored by circular dichroism. Far-UV CD spectra of (A) AVP2 and (B) RiLK30 at 50 µM concentration and 25 °C in 10 mM Tris-HCl buffer pH 7.0. in the absence (blue lines) or presence of SDS at different concentrations.
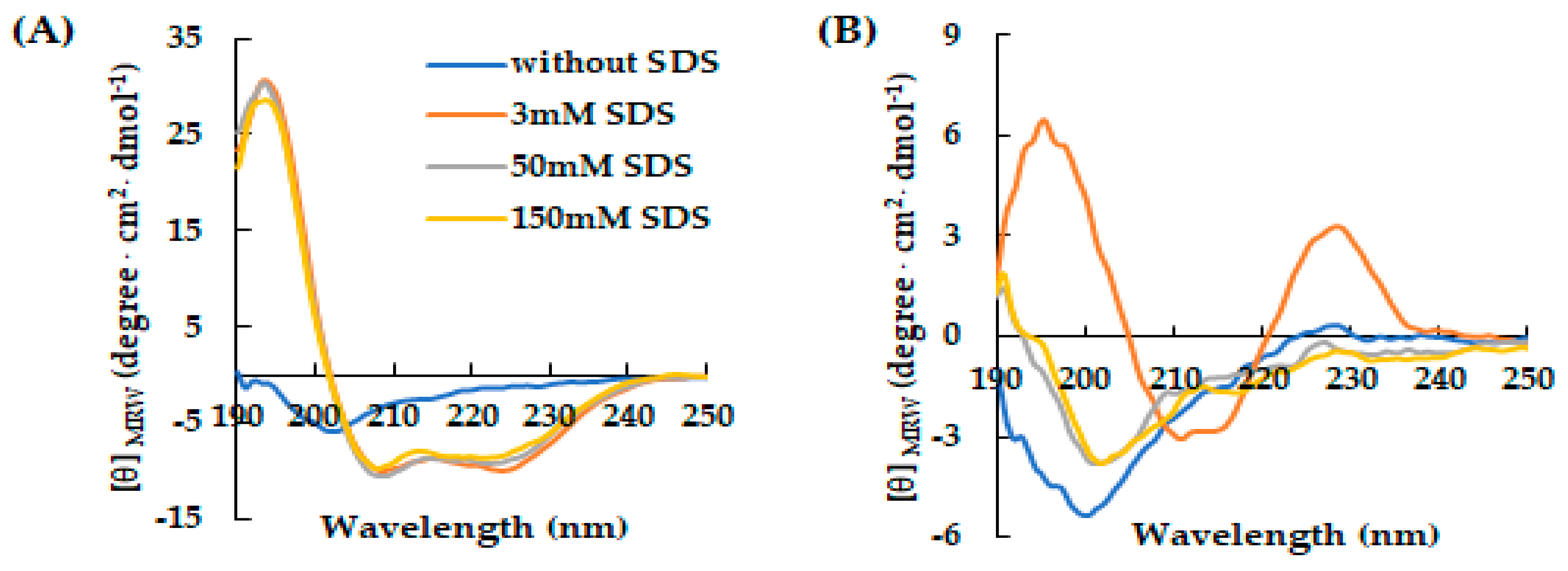
Figure 2.
Effect of SDS concentration on the secondary structure of AVP2 and RiLK30 monitored by fluorescence spectroscopy. Fluorescence spectra of (A) AVP2 and (B) RiLK30 at 50 µM concentration and 25 °C in 10 mM Tris-HCl buffer pH 7.0. in the absence (blue lines) or presence of SDS at different concentrations.
Figure 2.
Effect of SDS concentration on the secondary structure of AVP2 and RiLK30 monitored by fluorescence spectroscopy. Fluorescence spectra of (A) AVP2 and (B) RiLK30 at 50 µM concentration and 25 °C in 10 mM Tris-HCl buffer pH 7.0. in the absence (blue lines) or presence of SDS at different concentrations.
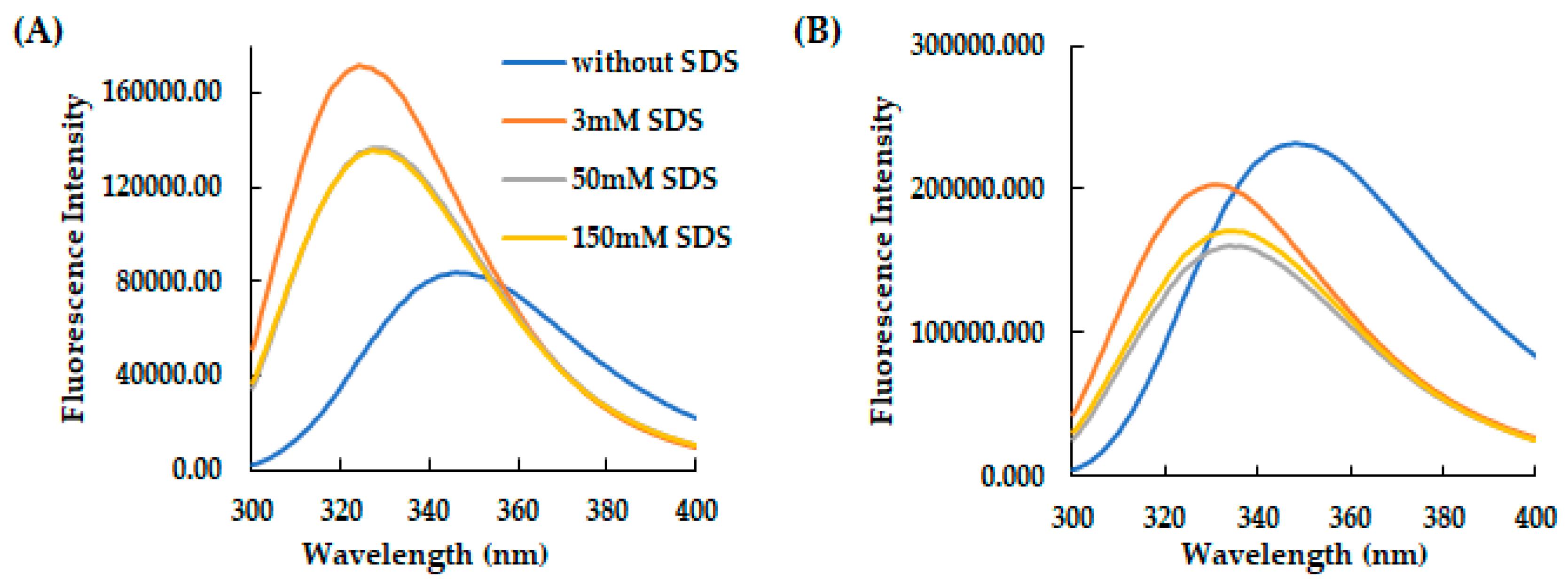
Figure 3.
Time-dependent effect of SDS on the secondary and tertiary structure of AVP2 and RiLK30 monitored by spectroscopic techniques. Far-UV circular dichroism spectra of (A) AVP2 and (B) RiLK30. Intrinsic fluorescence emission spectra of (C) AVP2 and (D) RiLK30. All spectra were recorded at a peptide concentration of 50 µM in 10 mM Tris-HCl. pH 7.0 in the presence or absence (blue lines) of SDS (50 mM) up to 24 h incubation at 25 °C.
Figure 3.
Time-dependent effect of SDS on the secondary and tertiary structure of AVP2 and RiLK30 monitored by spectroscopic techniques. Far-UV circular dichroism spectra of (A) AVP2 and (B) RiLK30. Intrinsic fluorescence emission spectra of (C) AVP2 and (D) RiLK30. All spectra were recorded at a peptide concentration of 50 µM in 10 mM Tris-HCl. pH 7.0 in the presence or absence (blue lines) of SDS (50 mM) up to 24 h incubation at 25 °C.
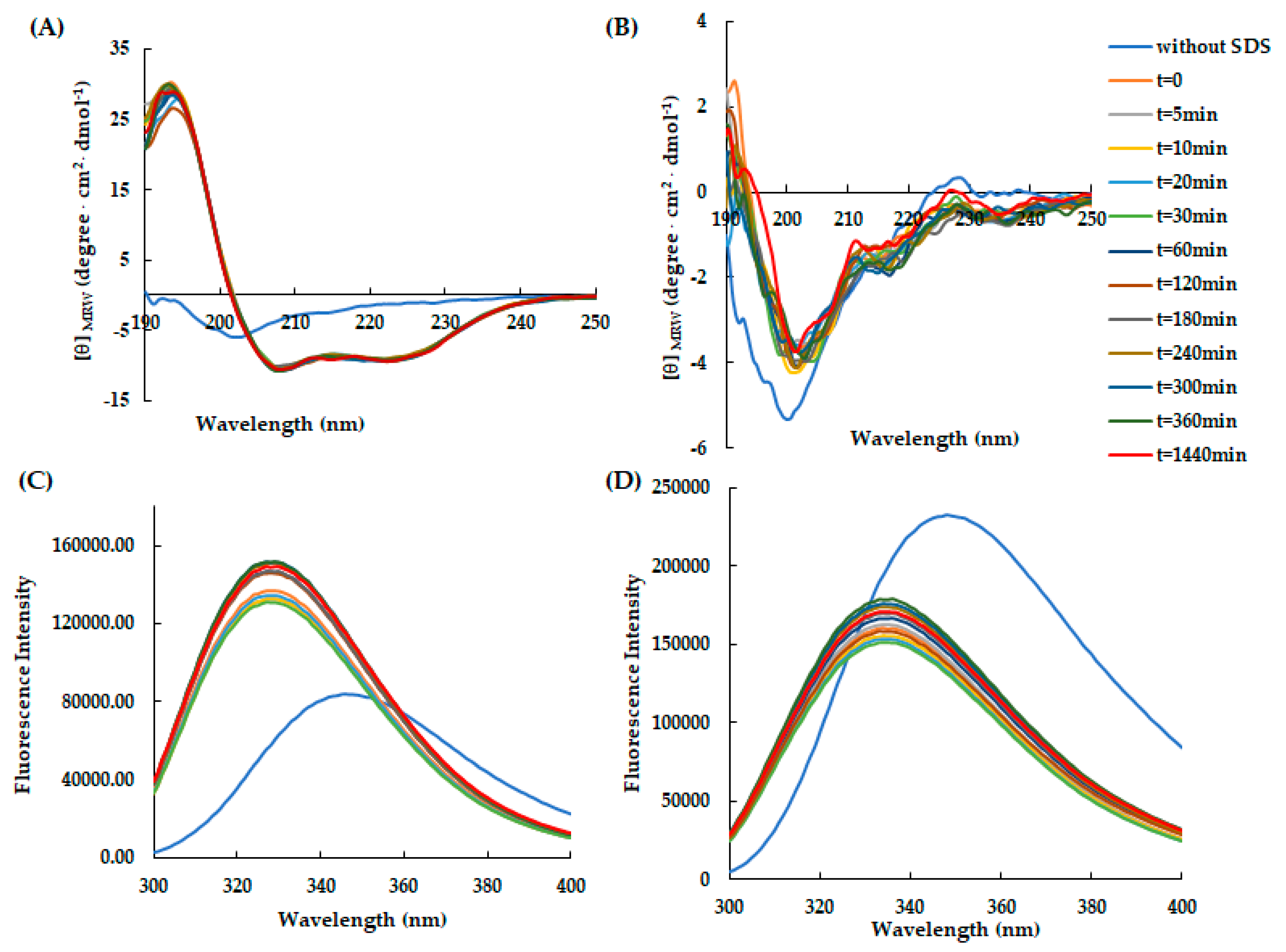
Figure 4.
Effect of temperature on the secondary and tertiary structure of RiLK30 monitored by spectroscopic techniques. Far-UV CD spectra of RiLK30 at (A) 4 °C and (B) 25 °C. Intrinsic fluorescence emission spectra of RiLK30 at (C) 4 °C and (D) 25 °C. All spectra were recorded at a peptide concentration of 50 µM in 10 mM Tris-HCl buffer pH 7.0 in the presence or absence (blue lines) of SDS (50 mM) up to 48 h incubation at 25 °C.
Figure 4.
Effect of temperature on the secondary and tertiary structure of RiLK30 monitored by spectroscopic techniques. Far-UV CD spectra of RiLK30 at (A) 4 °C and (B) 25 °C. Intrinsic fluorescence emission spectra of RiLK30 at (C) 4 °C and (D) 25 °C. All spectra were recorded at a peptide concentration of 50 µM in 10 mM Tris-HCl buffer pH 7.0 in the presence or absence (blue lines) of SDS (50 mM) up to 48 h incubation at 25 °C.
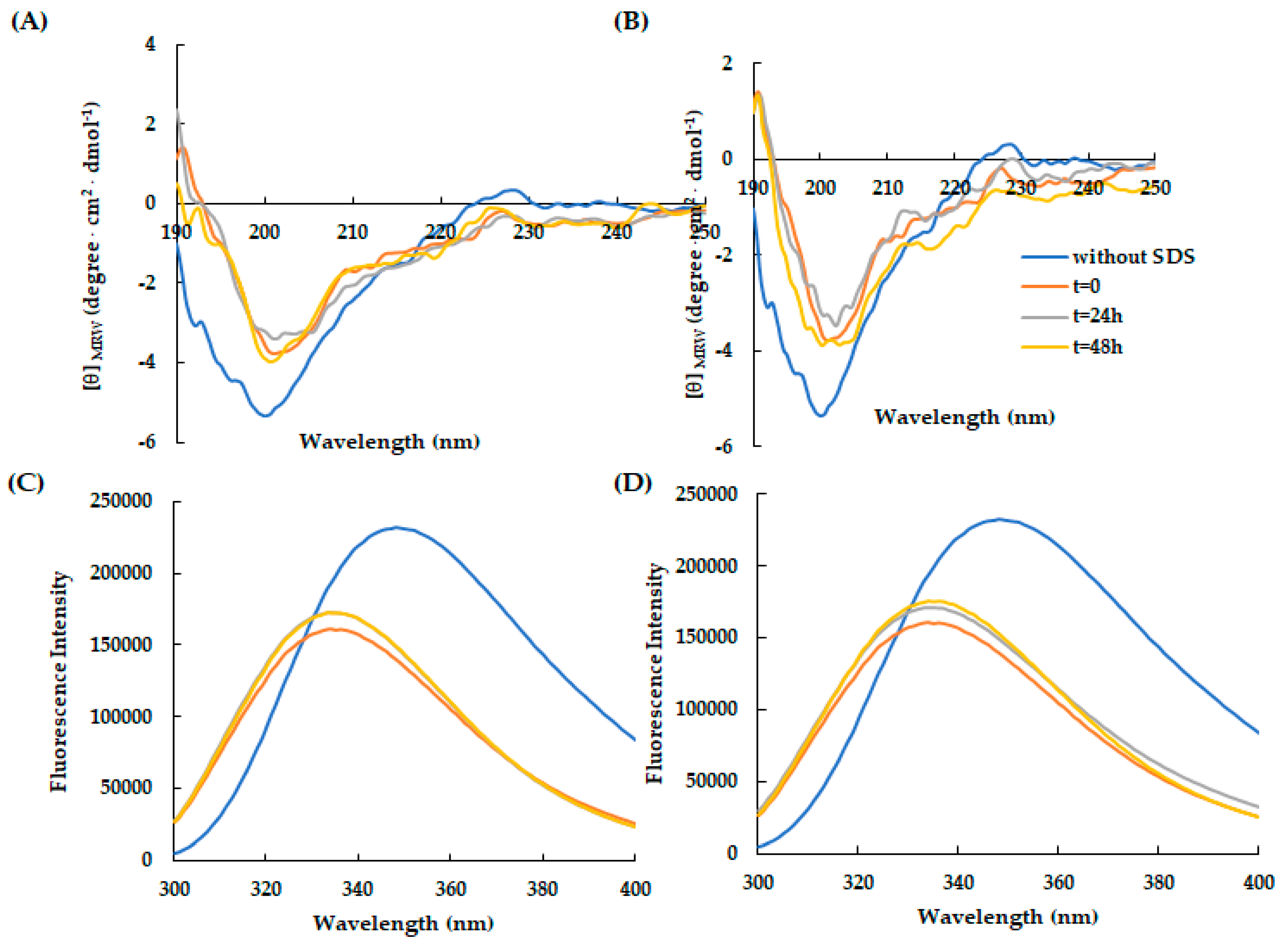
Figure 5.
Effect of temperature on the secondary and tertiary structure of AVP2 monitored by spectroscopic techniques. Far-UV CD spectra of AVP2 at (A) 4 °C and (B) 25 °C. Intrinsic fluorescence emission spectra of AVP2 at (C) 4 °C and (D) 25 °C. All spectra were recorded at a peptide concentration of 50 µM in 10 mM Tris-HCl buffer pH 7.0 in the presence or absence (blue lines) of SDS (50 mM) up to 48 h incubation at 25 °C.
Figure 5.
Effect of temperature on the secondary and tertiary structure of AVP2 monitored by spectroscopic techniques. Far-UV CD spectra of AVP2 at (A) 4 °C and (B) 25 °C. Intrinsic fluorescence emission spectra of AVP2 at (C) 4 °C and (D) 25 °C. All spectra were recorded at a peptide concentration of 50 µM in 10 mM Tris-HCl buffer pH 7.0 in the presence or absence (blue lines) of SDS (50 mM) up to 48 h incubation at 25 °C.
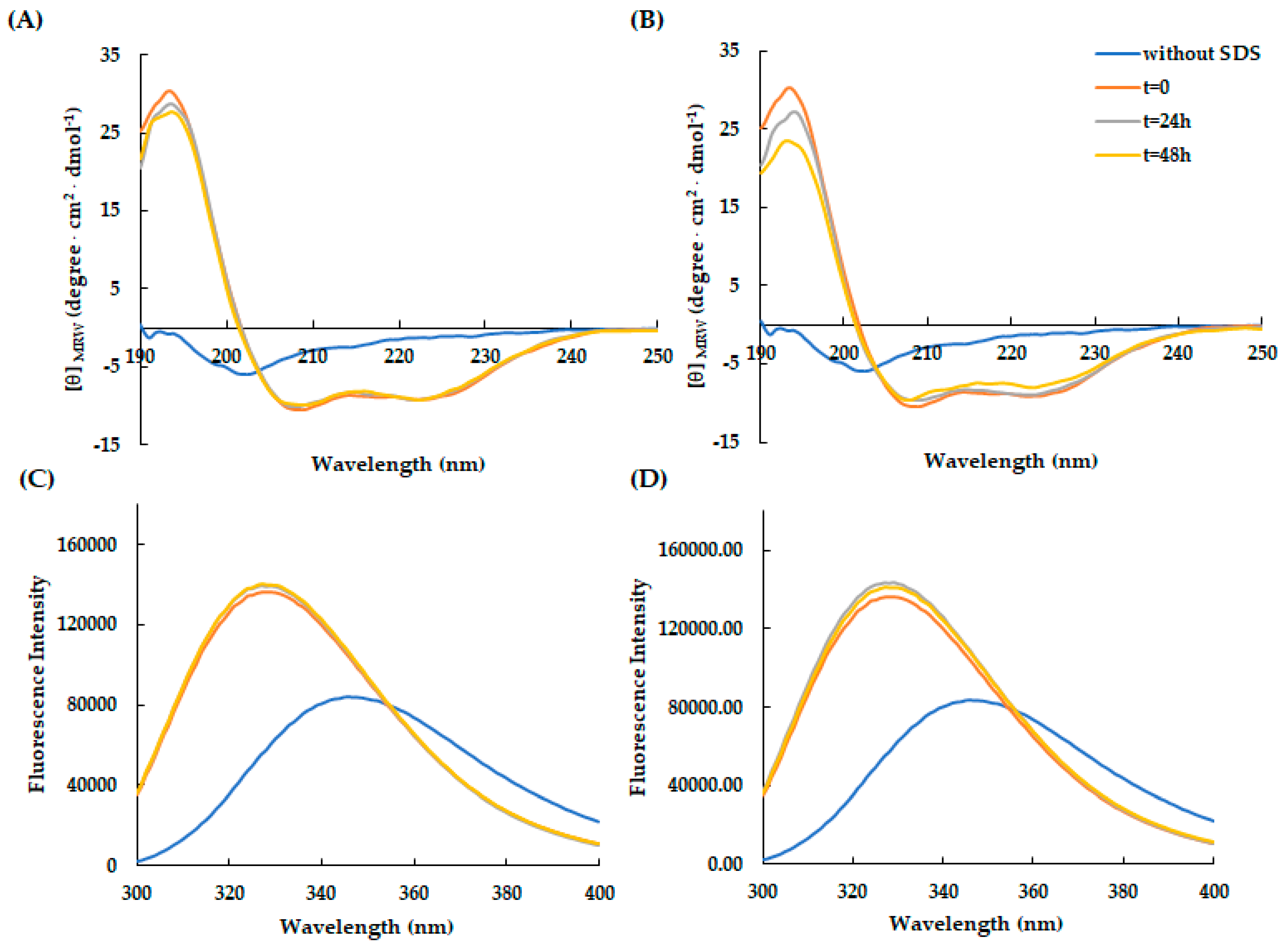
Figure 6.
Stability in saline solution of (A) AVP2 and (B) RiLK30 determined by Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC) on a C18 column. The peptides at final concentration of 50 μM were incubated in the presence of NaCl (150 mM) until 9h hours at 25 °C. At each incubation time, the peptide solutions were recovered and analyzed by RP-HPLC. The solutions at time 0 (t = 0) were used as control. The chromatograms are representative of three independent experiments.
Figure 6.
Stability in saline solution of (A) AVP2 and (B) RiLK30 determined by Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC) on a C18 column. The peptides at final concentration of 50 μM were incubated in the presence of NaCl (150 mM) until 9h hours at 25 °C. At each incubation time, the peptide solutions were recovered and analyzed by RP-HPLC. The solutions at time 0 (t = 0) were used as control. The chromatograms are representative of three independent experiments.
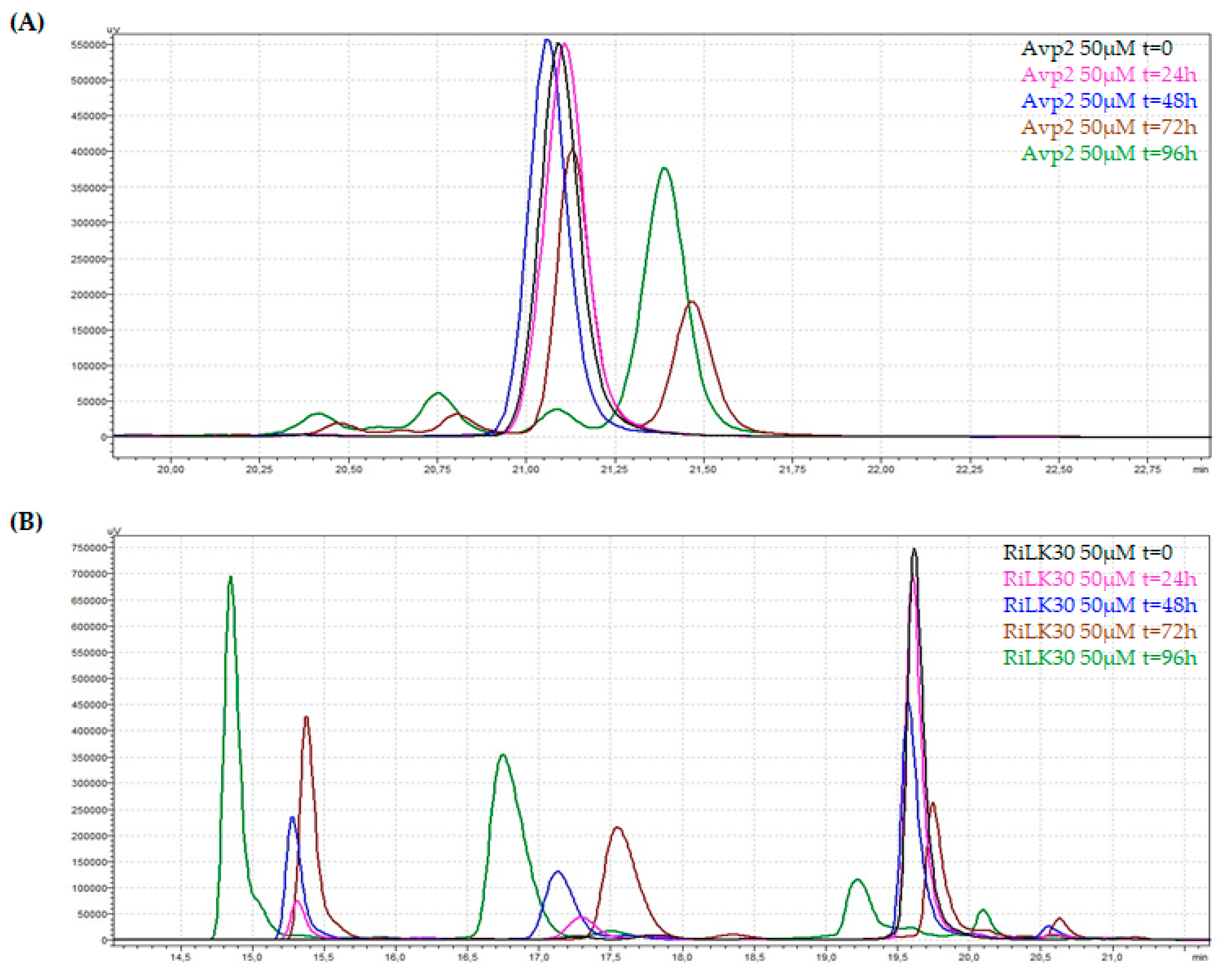
Figure 7.
Antibacterial activity of our peptides against the foodborne pathogens monofasic Salmonella Typhimurium, Salmonella Napoli, and Campylobacter jejuni. CTRL: microorganism without treatment. Bacterial cultures treated, or not, with peptides at two different concentrations were seeded on selective plates. The images are representative of three independent experiments performed in triplicate.
Figure 7.
Antibacterial activity of our peptides against the foodborne pathogens monofasic Salmonella Typhimurium, Salmonella Napoli, and Campylobacter jejuni. CTRL: microorganism without treatment. Bacterial cultures treated, or not, with peptides at two different concentrations were seeded on selective plates. The images are representative of three independent experiments performed in triplicate.
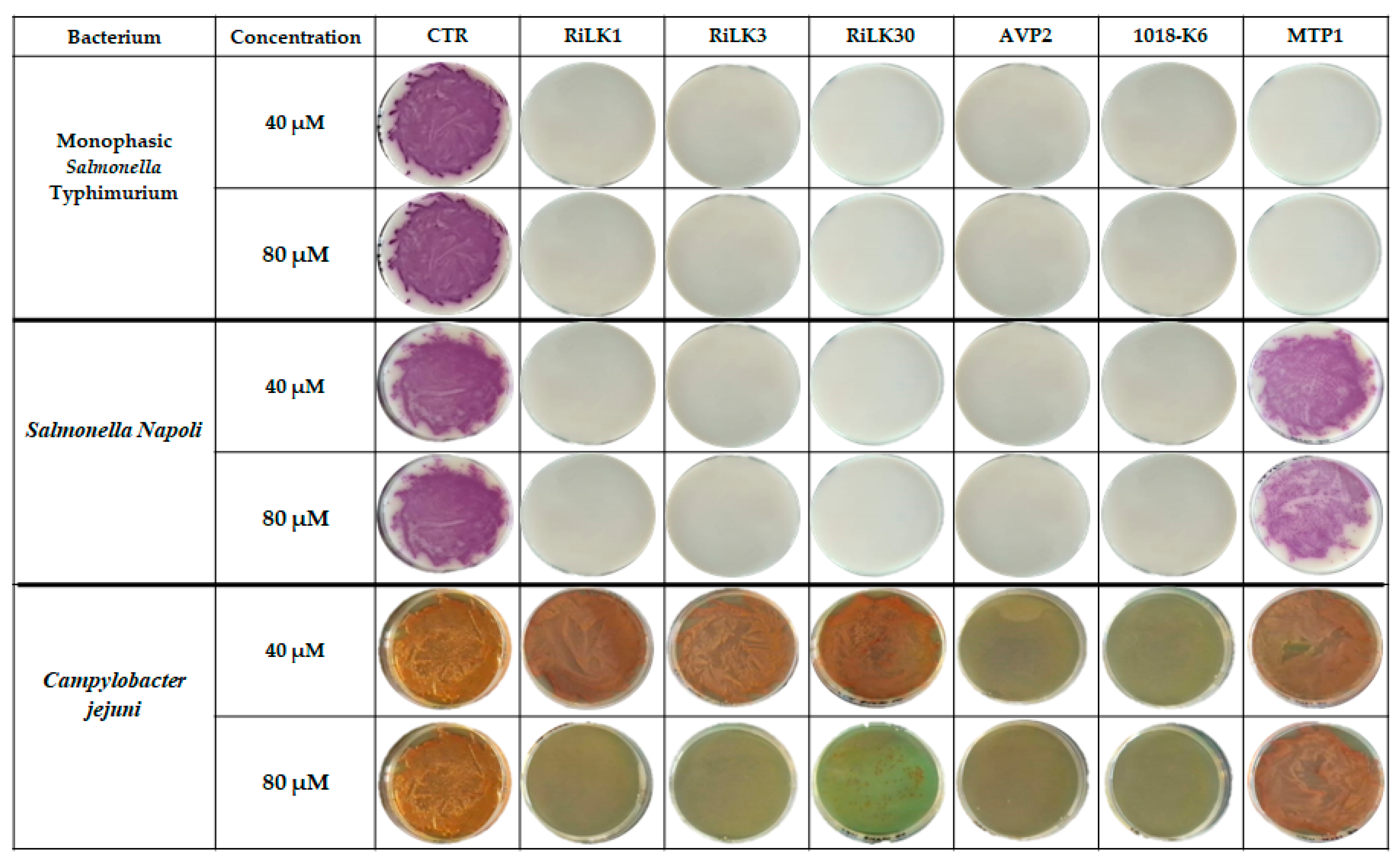

Table 1.
Characteristics of the selected peptides.
| Peptide | Sequence | MW (Da) |
Net Charge | BI (kcal/mol) |
Hydrophobicity | AVP§ |
|---|---|---|---|---|---|---|
| RiLK1a | RLKWVRIWRR (10 aa) | 1467.8 | +5 | 4.70 | -0.56 | 1 |
| RiLK3 b | RLRWVRIWRR (10 aa) | 1495.8 | +5 | 5.6 | -0.63 | 0.932 |
| RiLK30 | KLRWVKIWKK (10 aa) | 1383.8 | +5 | 1.85 | -0.36 | 0.994 |
| 1018-K6c | VRLIVKVRIWRR (12 aa) | 1594.0 | +5 | 3.00 | -0.35 | 0.558 |
| MTP1d | KVSGVLFGTGLWVAL (15 aa) | 1545.8 | +1 | -1.68 | 0.23 | 0.744 |
| AVP2e | GWFDVVKHIAKRF (13 aa) | 1601.9 | +3 | 1.18 | -0.10 | 0.926 |
Table 2.
In vitro effect of peptides on HAV.
| Viral titre after treatment (logTCID50/ml ± SD) |
LRV (logTCID50/ml ± SD) |
|||
|---|---|---|---|---|
| Treatment | 4°C | RT | 4°C | RT |
| Untreated HAV | 4.7±0.2 | 4.7±0.2a | n.r. | n.r. a |
| RiLK1 [80 µM] | 3.6±0.1 | 3.3±0.2 a | 1.1±0.3 | 1.4±0.4 a |
| RiLK1 [40 µM] | 3.9±0.2 | 3.6±0.1 a | 0.8±0.4 | 1.1±0.3 a |
| RiLK3 [80 µM] | 4.6±0.1 | 4.4±0.1 a | 0.1±0.3 | 0.3±0.3 a |
| RiLK3 [40 µM] | 4.6±0.3 | 4.4±0.1 a | 0.1±0.2 | 0.3±0.3 a |
| RiLK30 [80 µM] | 4.3±0.2 | 4.1±0.2 | 0.4±0.4 | 0.6±0.4 |
| RiLK30 [40 µM] | 4.3±0.2 | 4.1±0.2 | 0.4±0.4 | 0.6±0.4 |
| 1018-K6 [80 µM] | 3.3±0.2 | 3.3±0.2 | 1.4±0.4 | 1.4±0.4 |
| 1018-K6 [40 µM] | 3.4±0.1 | 3.3±0.2 | 1.3±0.3 | 1.4±0.4 |
| MTP1 [80 µM] | 3.8±0.2 | 3.5±0.1 | 0.9±0.4 | 1.2±0.3 |
| MTP1 [40 µM] | 3.6±0.3 | 3.2±0.2 | 1.1±0.2 | 1.5±0.4 |
| AVP2 [80 µM] | 4.3±0.2 | 4.0±0.3 | 0.4±0.4 | 0.7±0.5 |
| AVP2 [40 µM] | 4.3±0.2 | 4.3±0.2 | 0.4±0.4 | 0.4±0.4 |
n.r. = no reduction; LRV= log reduction value; RT= room temperature (20 ± 2°C); a Ref [46].
Table 3.
In vitro effect of peptides on MNV-1.
| Viral titre after treatment (logTCID50/ml ± SD) |
LRV (logTCID50/ml ± SD) |
|||
|---|---|---|---|---|
| Treatment | 4°C | RT | 4°C | RT |
| Untreated MNV-1 | 4.5±0.2 | 4.5±0.2 | n.r. | n.r. |
| RiLK1 [80 µM] | 4.3±0.2 | 3.3±0.2 | 0.2±0.4 | 1.2±0.4 |
| RiLK1 [40 µM] | 4.3±0.2 | 3.5±0.2 | 0.2±0.4 | 1.0±0.4 |
| RiLK3 [80 µM] | 4.5±0.2 | 4.5± 0.1 | 0±0.4 | 0±0.3 |
| RiLK3 [40 µM] | 4.5± 0.1 | 4.5±0.2 | 0±0.3 | 0±0.4 |
| RiLK30 [80 µM] | 3.9± 0.2 | 4.5± 0.1 | 0.6±0.4 | 0±0.3 |
| RiLK30 [40 µM] | 4.3±0.3 | 4.5±0.2 | 0.2±0.5 | 0±0.4 |
| 1018-K6 [80 µM] | 4.3±0.2 | 4.3±0.2 | 0.2±0.4 | 0.2±0.4 |
| 1018-K6 [40 µM] | 4.5±0.1 | 4.5±0.2 | 0±0.3 | 0±0.4 |
| MTP1 [80 µM] | 4.5±0.1 | 4.5±0.2 | 0±0.3 | 0±0.4 |
| MTP1 [40 µM] | 4.4±0.1 | 4.5±0.2 | 0.1±0.3 | 0±0.4 |
| AVP2 [80 µM] | 3.6±0.1 | 3.5±0.1 | 0.9±0.3 | 1.0±0.3 |
| AVP2 [40 µM] | 3.6±0.1 | 3.6±0.1 | 0.9±0.3 | 0.9±0.3 |
n.r. = no reduction; LRV= log reduction value; RT= room temperature (20 ± 2°C).
Table 4.
Mechanism of action of RiLK1 and RiLK3 peptides on HAV infectivity.
| RiLK1 | RiLK3 | |||
|---|---|---|---|---|
| Viral titre (logTCID50/ml ± SD) |
LRV (logTCID50/ml ± SD) |
Viral titre (logTCID50/ml ± SD) |
LRV (logTCID50/ml ± SD) |
|
| untreated virus | 4.7±0.2 | - | 4.7±0.2 | - |
| virus pre-treatment | 3.3±0.2 | 1.4±0.4 | 4.4±0.1 | 0.3±0.3 |
| cell pre-treatment | 4.6±0.1 | 0.1±0.3 | 3.8±0.2 | 0.9±0.4 |
| attachment | 3.3±0.2 | 1.4±0.4 | 3.7±0.2 | 1.0±0.4 |
| entry | 4.6±0.1 | 0.1±0.3 | 4.2±0.2 | 0.5±0.4 |
LRV= log reduction value.
Table 5.
Mechanism of action of RiLK1 and RiLK3 peptides on MNV-1 infectivity.
| RiLK1 | RiLK3 | |||
|---|---|---|---|---|
| Viral titre (logTCID50/ml ± SD) |
LRV (logTCID50/ml ± SD) |
Viral titre (logTCID50/ml ± SD) |
LRV (logTCID50/ml ± SD) |
|
| untreated virus | 4.5±0.2 | - | 4.5±0.2 | - |
| virus pre-treatment | 3.3±0.2 | 1.2±0.4 | 4.5±0.1 | 0±0.3 |
| cell pre-treatment | 4.2±0.2 | 0.3±0.4 | 3.5±0.1 | 1.0±0.3 |
| attachment | 3.1±0.2 | 1.4±0.4 | 3.5±0.1 | 1.0±0.3 |
| entry | 3.5±0.2 | 1.0±0.4 | 3.5±0.1 | 1.0±0.3 |
LRV= log reduction value.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



