
Submitted:
15 April 2024
Posted:
16 April 2024
You are already at the latest version
Abstract
Current microbiological methods for pneumonia diagnosis require invasive specimen collection and time-consuming analytical procedures. There is a need for less invasive and faster methods to detect lower respiratory tract infection. The analysis of volatile metabolites excreted by pathogenic microorganisms provides the basis for developing such a method. Given the synergistic role of Candida albicans in increasing the virulence of pathogenic bacteria causing pneumonia and the cross-kingdom metabolic interactions between microorganisms, we compare the emission of volatiles from Candida albicans yeasts and the bacteria Staphylococcus aureus using single and mixed co-cultures and apply that knowledge to human in vivo investigations. Gas-chromatography-mass spectrometry (GC-MS) analysis resulted in the identification of sixty-eight volatiles that were found to have significantly different levels in cultures compared to reference medium samples. Certain volatiles found in co-culture originated mainly from C. albicans metabolism (e.g., isobutyl acetate), whereas other volatiles primarily came from S. aureus (e.g., ethyl 2-methylbutyrate). Isopentyl valerate reflects synergic interactions of both microbes, as its level in co-cultures was found to be approximately three times higher than the sum of its amounts in monocultures. Hydrophilic-lipophilic balanced (HLB) coated meshes for thin-film microextraction (TFME) were used to pre-concentrate volatiles directly from bronchoalveolar lavage (BAL) specimens collected from patients suffering from ventilation-associated pneumonia (VAP) that was caused explicitly by C. albicans and S. aureus. GC-MS analyses confirmed the existence of in vitro-elucidated microbial VOCs in human specimens. Significant differences in BAL-extracted amounts respective to the pathogen causing pneumonia were found. The model in vitro experiments provided evidence that cross-kingdom interactions between pathogenic microorganisms affect the synthesis of the volatile compounds. The TFME meshes coated with HLB particles proved to be suitable for extracting VOCs from human material, enabling the translation of in vitro experiments on the microbial volatilome to the in vivo situation involving infected patients. This indicates the direction that should be taken for further clinical studies on VAP diagnosis based on volatile analysis.
Keywords:
Bacterial Metabolites
; Bronchoalveolar Lavage Sampling
; Cross-Kingdom Interactions
; Thin-Film Microextraction
; Ventilation-Associated Pneumonia Diagnosis
; Volatile Biomarkers
1. Introduction
According to the National Healthcare Safety Network, a tracking system from the Centers for Disease Control and Prevention, more than 15 thousand pathogens causing Ventilator-Associated Pneumonia (VAP) were reported from 1155 Acute Care Hospitals in the USA up to 2021, with Staphylococcus aureus being the most prevalent pathogen (29.6%), followed by Pseudomonas aeruginosa (13.4%) [1]. Notably, both pathogens interact with Candida spp., a yeast that readily colonizes the airways in more than 50% of patients with suspected VAP [2]. These fungal-bacterial interactions promote the growth of antibiotic-resistant bacteria [2-5] and enhance their virulence [6-8]. Hence, VAP patients colonized with Candida spp. are at risk of prolonged mechanical ventilation, extended ICU stay, and increased mortality [2, 3, 9-11]. As pointed out by Ricard and Roux, taking together all experimental and clinical clues, Candida spp. should no longer be considered a bystander [12]. Instead, it may constitute leads for new therapies and early non-invasive diagnosis of pneumonia, triggering an early workup and potentially providing methods for monitoring the response to treatment. Such a promising diagnostic technique utilizes Volatile Organic Compounds (VOCs) produced by microorganisms that can be identified and quantified in the exhaled breath gas of patients using mass spectrometry, either offline after sample preconcentration and chromatographic analysis in the laboratory [13-15] or real-time directly from mechanically ventilated patients [16]. Analysis of bacterial VOCs in exhaled breath has excellent applicability in VAP because it is entirely non-invasive and can be done repeatedly without any burden to the patient. Hence, the analysis of volatiles in exhaled breath could serve as a non-invasive and sensitive supportive method to the established invasive (tracheal aspirates or bronchoalveolar lavage sampling) and time-intensive procedure of microbiological testing. Ultimately, once the key volatiles have been correctly identified, an analytical analysis could be done using portable point-of-care devices such as sensors, leading to timely, effective, and personalized antibiotic use, so much needed to deescalate the problem of antimicrobial resistance[17,18].
The aim of the first part of this study is to identify the volatile metabolites excreted by C. albicans and S. aureus in single cultures and mixed co-cultures. Mixed co-cultures are investigated to determine whether the cross-kingdom interactions between these two species manifest in specific VOC profiles. For this purpose, separate bacterial, separate fungal and mixed cultures were established under strictly controlled conditions of temperature, humidity, and ventilation. Dynamic headspace sampling on multibed sorption tubes was performed at several time points to follow the changes in VOC profiles. The aim of the second part of our work is to reveal whether volatiles found under in vitro conditions (model study) are also present in vivo (clinical samples). Therefore, bronchoalveolar lavage (BAL) specimens were collected from mechanically ventilated patients coinfected explicitly with the two mentioned pathogens. Due to the small quantity of the biological material (BAL), a statical headspace (HS) sampling needs to be applied to preconcentrate VOCs before gas chromatographic-mass spectrometric (GC-MS) analysis. For this purpose, novel thin-film microextraction (TFME) devices with a mesh-like geometry are used which have a larger stationary phase area-to-volume ratio compared to the regular solid-phase microextraction (SPME) fibers. The resulting enhanced extraction efficiency is further improved for oxygenated VOCs (aldehydes, ketones, alcohols) by application of the hydrophilic-lipophilic balanced (HLB) polymer particles, enabling the trace analysis (down to the sub-ppb level) of the polar volatile metabolites [19] important also for microorganisms.
2. Materials and Methods
2.1. Chemicals
Liquid and gaseous chemicals (manufactured by Acros Organics, Alfa Aesar, ChemSampCo, Honeywell, Sigma-Aldrich, or Tokyo Chemical Industry) were purchased either from AlChem (Toruń, Poland) or from Merck KGaA (Darmstadt, Germany). Broth Medium (Caso Bouillon from Merck, Darmstadt, Germany) was prepared as recommended by Merck: 30 g of medium powder was dissolved in an Erlenmeyer flask with distilled water up to 1 L. The broth medium consists of casein peptone 17.0 g/L, peptone from soya flour 3.0 g/L, D + glucose 2.5 g/L, sodium chloride 5.0 g/L, and di-potassium hydrogen phosphate 2.5 g/L. The culture medium was decanted into four 100 mL cleaned culture flasks.
2.2. Pre-Culture Preparation and Colony-Forming Unit Counting
All in vitro experiments with fungal and bacterial cultures were performed at the Institute of Hygiene and Medical Microbiology, Medical University of Innsbruck. The collected samples were then analyzed at the Institute for Breath Research, Universität Innsbruck.
Stock cultures of Candida albicans (CBS 5982) and Staphylococcus aureus (ATCC 6538) were stored at 4 °C on Müller-Hinton agar plates (CM0337 Oxoid Ltd, Hampshire, Great Britain). Single colonies from these were inoculated into 5 mL of the medium (Caso Bouillon, Merck Darmstadt) and grown overnight at 37 °C (Memmert oven, Loading Modell 100-800, 91107 Schwabach Germany) to approximately 1 x 107 colony-forming units (CFU)/mL (C. albicans) and 3 x 109 CFU/mL (S. aureus). To investigate VOC production in single culture and co-culture, aliquots from the overnight culture of each microbe were inoculated in appropriate quantities into 100 mL of the medium to yield the required concentration (approximately 1 x 105 CFU/mL for C. albicans and 1 x 103 CFU/mL for S. aureus). A magnetic stirrer circulating the liquid culture at 50 rpm enabled the oxygen uptake and VOC release into the medium headspace.
At each considered time-point, 1 mL of medium suspension was collected to count microorganisms. Aliquots (50 µL per plate) of undiluted samples as well as of samples diluted with 0.9 % NaCl (10-2–10-6 -fold) were plated on Müller-Hinton agar plates with an automated spiral plater (model WASP2, Don Whitley Scientific, Shipley, UK). CFU counts were performed after incubation over 24 h at 37 °C. Morphological differences, primarily the color of C. albicans (white) and S. aureus (yellowish), allowed both species of co-cultures to be distinguished.
2.3. Experimental Setup for Cultivating and Headspace Sampling
An in-house system that has been previously described in detail was used [20]. Briefly, four glass bottles containing 100 mL of liquid sample each (either microbe or sterile reference medium) were kept within a water bath at 37 °C and gently stirred (50 rpm). The incubation setup was the following: flask No. 1 was the control medium (sterile, without microorganisms), flask No. 2 was a sole C. albicans culture, flask No. 3 was a sole S. aureus culture, and flask No. 4 contained co-cultured C. albicans and S. aureus. To sample the headspace above a liquid, synthetic air was flowed over the medium. This air was of purity 5.5, enriched with 5% CO2 (Linde, Stadl-Pradl, Austria) and additionally purified with a Supelcarb™ hydrocarbon trap (Supelco, Bellefonte, PA, USA) and an inline-catalyst (Parker Zero Air Generator, Balston®, model: 75-83-220, Parker Hannifin Corporation, Haverhill, Massachusetts, USA). To decrease the high relative humidity that could lead to undesired condensation in transfer lines or excessive water uptake on the adsorption tube, the entire system was placed in the incubator at 45 °C, and an additional flow of purified synthetic air was used (35 mL/min) to dilute the headspace sample (5 mL/min). All the flows in four cultures were precisely controlled and additionally checked at the outlet of a sorption tube using a flowmeter (Red-Y Compact GCR, Vögtlin Instruments AG, Aesch; Switzerland). For single sampling, 200 mL of the gas flow was adsorbed on a tube filled with Carbotrap B and Carbopack X within a 40-minute timeframe. The first gas samples were collected before the inoculation of the microorganisms to assess the volatile compounds present in the headspace of a sterile medium, after purging overnight with synthetic air (which considerably reduces background VOCs, as demonstrated in a previous study [21]). Time points at 2 h, 3.5 h, 5 h, 6.5 h, 8 h, 26 h, and 28 h were chosen for headspace sampling based on microorganisms growth rate. One mL of the culture suspension for the CFU counting was collected immediately after the headspace sampling using a slightly bent 12 cm needle (Sterican) introduced via a septum port (ensuring that the system remains gas-tight).
2.4. Collection and Preparation of Bronchoalveolar Lavage Specimens
All BAL samples were collected from mechanically ventilated patients hospitalized in the Anesthesiology and Intensive Care Unit of the 10th Military Research Hospital and Polyclinic in Bydgoszcz, Poland. Subsequent TFME-GC-MS analyses of BAL specimens were performed at the Department of Pharmacodynamics and Molecular Pharmacology, Nicolaus Copernicus University in Toruń, Poland. Ethical approval for this study was obtained from the local Bioethics Committee (KB-218/2018). The patients enrolled in this study had to fulfill the following inclusion criteria: (1) admission to ICU, (2) mechanically ventilated, (3) suspected of or confirmed for VAP (based on clinical and microbiological evidence). The exclusion criteria were: (1) age under 18, (2) pregnancy, (3) confirmed coexisting lung disease (traumatic lung injury or pulmonary cancer), (4) strict isolation at ICU, (5) increased intracranial pressure (ICP), (6) positive end-expiratory pressure (PEEP) > 10, and (7) extra corporal heart and lung assistance devices.
According to the routine clinical practice in the Anesthesiology and Intensive Care Unit of the 10th Military Research Hospital and Polyclinic in Bydgoszcz (Poland), BAL samples were taken from mechanically ventilated patients for microbiological testing upon suspicion of pneumonia based on clinical evidence and tailoring the antimicrobial therapy. In accordance with the applied standard microbiological procedure, BAL inoculation was performed with a calibrated loop with a volume of 10 microliter on a set of media plates. The purulent part of the material was selected for culture. The residual BAL specimen was immediately transported from the Hospital Microbiology Laboratory to the University Analytical Laboratory for GC-MS analysis (both in Bydgoszcz, Poland). Meanwhile, the plated cultures were kept overnight at 37 °C with the following quantitative read-out where the growth of at least 104 CFU/mL was considered diagnostic of the pathogen that was further identified and subjected to antimicrobial susceptibility testing.
Altogether, as many as 92 BAL specimens were analyzed with TFME-GC-MS. However, for consistency with the in vitro experiments, this study only included those BAL samples for which microbiological testing confirmed the presence of either sole C. albicans, sole S. aureus or both microbes and lack of any others (resulting in the number of BAL samples n = 6, n = 3, and n =4, respectively, collected from altogether 13 different patients).
2.5. Headspace Sampling from Bronchoalveolar Lavage Specimens
TFME meshes covered with the hydrophilic-lipophilic balanced (HLB) stationary phase were manufactured according to the protocol developed by Grandy et al. [22] and were kindly provided by Prof Pawliszyn (University of Waterloo, Waterloo, ON, Canada). Before each sampling, the membranes were placed in an empty glass sorption tube (Supelco, Merck, Darmstadt, Germany) and preconditioned by putting them into a TD Clean-Cube conditioning unit (Scientific Instruments Manufacturer GmbH, Oberhausen, Germany). They were heated up to a temperature of 250 °C for 30 min under a continuous flow of nitrogen 6.0 that had been further purified on a Carrier Gas Purifier (Agilent, Santa Clara, CA, USA).
Immediately after sample transport from the Hospital Microbiological Laboratory, 5 mL of each BAL specimen was placed into a 20 mL glass vial and warmed within an incubator up to 37 °C for 30 min. The preconcentration of VOCs secreted from the BAL sample was done according to the previously optimized protocol [19]. Briefly, TFME sheets were suspended on a metal clamp under the cap with silicon septum inside the 20 mL glass vial containing the sample. Extraction took place at 37 °C for 90 min. Afterward, the TFME mesh was removed from the vial and placed into an empty sorption glass tube tightly closed in a TDS 3TM Storage Container (Supelco, Merck, Darmstadt, Germany) until the GC-MS analysis (done on the day of headspace sampling).
2.6. Gas Chromatographic – Mass Spectrometric Analysis
For the in vitro experiments with microorganisms, the TD-GC-MS analyses were performed on a 6890N gas chromatograph with detection by the mass selective detector 5973N (both from Agilent Technologies, Waldbronn, Germany). Samples that were loaded on multi-bed sorption tubes were thermally desorbed at 300 °C over 10 min in a TDS3 unit equipped with a TDSA2 autosampler (both from Gerstel, Mulheim an der Ruhr, Germany). Cryofocusing during primary desorption was achieved at -90 °C in the CIS4 injector containing a trap filled with Carbotrap B (Gerstel, Mulheim an der Ruhr, Germany). The secondary desorption (i.e., injection into the GC column) was done in splitless mode at 320 °C over 2 min. To ensure the best possible separation of sample constituents, the Rt-Q-Bond capillary column 30 m x 0.32 mm x 10 µm (Restek, Bellefonte, PA, USA) was used with the following temperature program: initial 55 °C held for 6 min, then ramped 7 °C/min up to 97 °C (2 min), 2 °C/min to 110 °C (0 min), 5 °C/min to 130 °C (4 min), 5 °C/min to 160 °C (4 min), 4 °C/min to 230 °C (0 min) and 10 °C/min to 280 °C (4 min). For MS acquisition, the ionization energy was set to 70 eV, ion source and quadrupole temperatures were 230 °C and 105 °C, respectively, and scanning was done over a range of 20–200 m/z. Identification of detected analytes was carried out in two stages. A preliminary identification was obtained by matching the recorded mass spectrum to the NIST library. The identification was further confirmed through chromatographic parameters (retention time) of the respective standards. If particular standards were not available, the compound is accordingly marked. For quantitative analysis, a specialized software, BreathViewer 2.1, was used (ONCOTYROL, Center for Personalized Cancer Medicine GmbH, Innsbruck, Austria). This post-processing program enables evaluation of the complex GC-MS data utilizing signal deconvolution and baseline correction to identify and quantify detected analytes. The final statistical calculations and plotting were done with GraphPad Prism5 software.
For the analysis of BAL specimens, the TFME meshes with extracted analytes were thermally desorbed at 250 °C over 10 min in a TD-30R autosampler (Shimadzu, Shim-Pol, Warsaw, Poland). The cryofocusing during preliminary desorption was achieved at -20 °C on a cold trap filled with Carboxen. Rapid heating of the cold trap to 350 °C triggered injection into a Nexis 2030 Gas Chromatograph (Shimadzu, Shim-Pol, Warsaw, Poland) in splitless mode over 2 min. Sample constituents were separated on an Rt-Q-Bond capillary column 30 m x 0.25 mm x 8 µm (Restek, Bellefonte, PA, USA) using the oven temperature program developed for in vitro experiments (with only a slight modification of a final ramp). Data were acquired with a QP-2020-NX Mass Spectrometer (Shimadzu, Shim-Pol, Warsaw, Poland) operating in a SCAN mode within a range of 33–235 m/z. Analytes detected were identified using the same two-stage approach (NIST spectra library match and confirmation with standards) as mentioned above. Again, if a particular standard was not available, the compound is accordingly marked. For data processing, Shimadzu GCMS PostRun Analysis software was used for automatic peak detection and integration based on the previously established database of analytes, further manually corrected (if necessary) by an experienced GC-MS analyst. To avoid errors in data interpretation caused by background contributions, the “Target Ion” was assigned to each analyte to integrate correctly the area under the chromatographic peak. Those “Target Ions” were selected manually by the experienced analyst to ensure either the most selective ion (unique for only one compound in the case of coeluted peaks) or the most sensitive ion (for peaks resolved to the baseline). Further, characteristic reference ions were assigned to each analyte to verify the peak identity during its integration process. The final statistical calculations and plotting were done with Metaboanalyst 5.0 online software.
3. Results
3.1. Microbial Growth
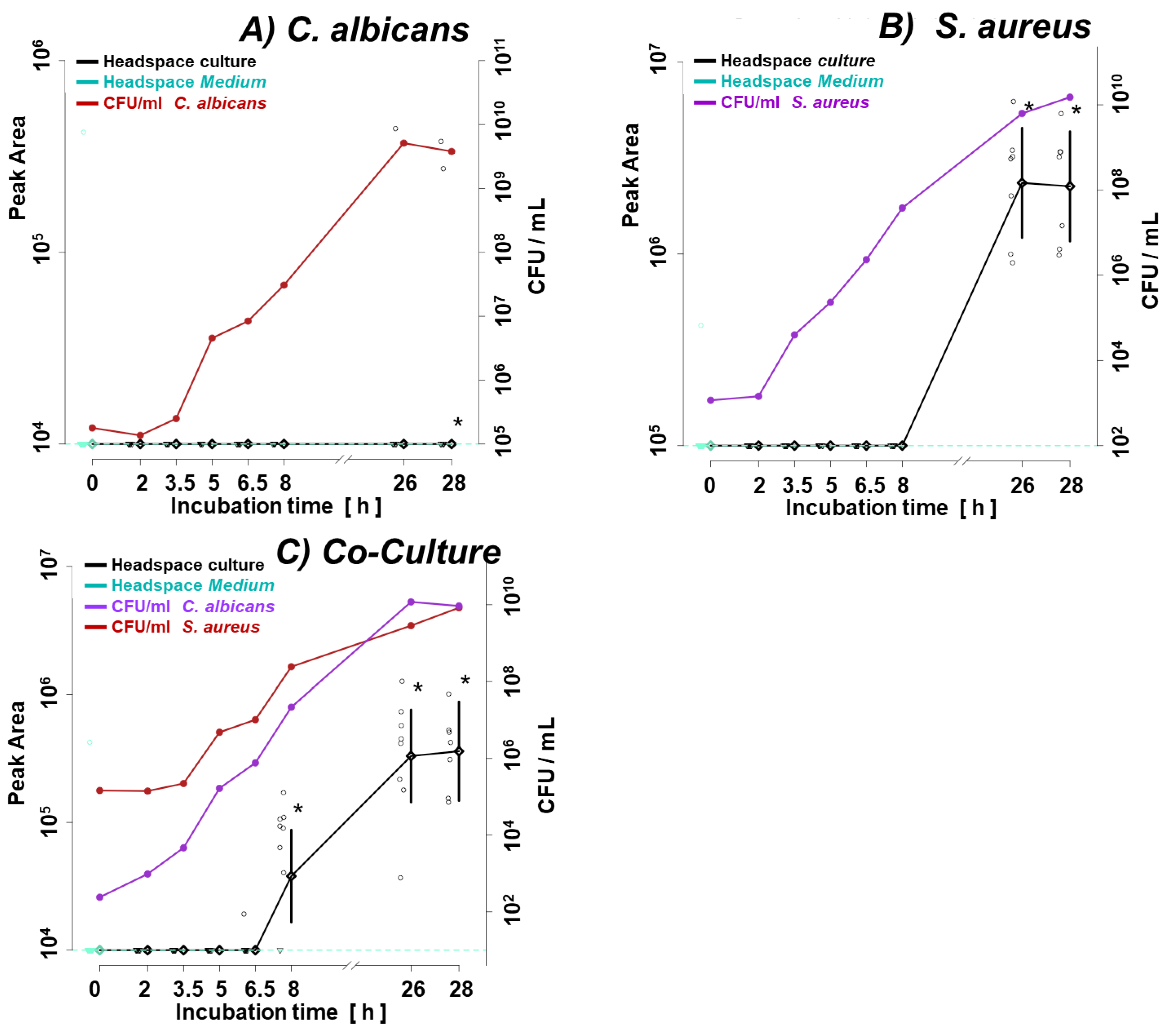
Figure 1a illustrates the growth curve of C. albicans and S. aureus in monocultures. Given the shorter lag phase and faster proliferation rate of the second, its initial inoculum was adjusted two orders of magnitude lower (ca. 1 × 103 CFU/mL for S. aureus vs. ca. 1 × 105 CFU/mL for C. albicans). The shift in microbes’ initial quantities resulted in similar amounts of both species in the late log phase (8 h) and steady phase (26 h, 28 h) (Figure 1a), enabling direct comparison of observed VOC profiles. Figure 1b and 1c demonstrate convergence in the growth rate of respective species in their single- and co-cultures. There was no difference in the course of CFU/mL from the start over the log to the stationary phase between single and co-cultures of both species.
3.2. VOCs of Microbial Origin
By comparing the peak areas for respective analytes measured at the same time points in the headspace of cultures and sterile reference medium, several volatiles were found at significantly decreased levels in cultures, implicating their consumption by the microbes. Other VOCs appeared unaffected by microbial cultures, and their profile did not change throughout the experiment. However, a third group of compounds present at an initially low level either showed an increased concentration (temporarily or continuously) or, after the initial absence in the culture, they appeared during microbial growth. All VOCs with significantly increased or decreased concentration in culture headspace compared to control medium headspace are given in Table 1, along with their parameters used for chromatographic identification (i.e., retention time, target ion, reference ions) and microbial synthesis or uptake status.
Since the VOC production in all cultures investigated in this study changed dynamically, they could be further split according to the microbial growth phase, distinguishing VOCs significantly higher in the log-phase (3.5–8 h of cultivation) or in the steady phase (24–26 h of cultivation). In this respect, C. albicans produced 28 VOCs and consumed 10 VOCs during the log phase (Supplementary Figure S1), while 20 volatiles were released and 14 were taken up during a steady phase (Supplementary Figure S2). In contrast, the metabolic activity of S. aureus was noticeably lower during the log phase, resulting in only 5 VOCs released and 1 VOC taken up (Supplementary Figure S3), but increased proportionally to the bacterial load, yielding 26 VOCs produced and 11 VOCs consumed during the steady phase (Supplementary Figure S4). The highest metabolic activity was observed for co-cultures of both species, where as many as 29 compounds were synthesized and 12 compounds catabolized during the log phase (Supplementary Figure S5). Similarly, 25 metabolites were produced, and 19 were degraded in the steady phase of co-culture growth (Supplementary Figure S6).
3.3. Co-Culture-Specific VOCs
To reveal the metabolic changes caused by interspecies’ interactions, the time-dependent profiles of VOC synthesis in co-cultures were compared to monocultures. For this purpose, VOCs with significantly different abundances in co-cultures and monocultures were normalized through log transformation and further subtracted. The positive value of such subtraction indicates a higher abundance of particular analytes in co-culture, while a negative value indicates a lower abundance in co-culture (and higher in a single culture). Similar to the previous considerations, these calculations were also performed separately for the log and steady phases of microbial growth. Figure 2 demonstrates differences in VOC profiles between C. albicans cultured alone and cultured with S. aureus. In the log phase, no single VOC could be found at a significantly higher level in the monoculture, whereas three volatiles showed a significantly higher production rate in the co-culture. An analogous comparison in the stationary phase confirmed the presence of four increasing and none significantly decreasing volatiles in the co-culture headspace.
A comparison of VOC metabolism in co-culture to the sole S. aureus during the log phase of microbial growth revealed 26 compounds significantly more abundant in the mixed culture headspace (those with positive values in Figure 3). Another 16 compounds were synthesized stronger in S. aureus monoculture during the log phase.
In the later phase of microbial growth, higher metabolic activity of the sole S. aureus was more pronounced since as many as 25 volatiles were found at higher levels during the steady phase of S. aureus compared to its co-culture with C. albicans (Figure 4). At this stage, only 16 metabolites were synthesized in higher quantities in co-cultures (those with positive values in Figure 4).
For a selection of volatile metabolites that could testify about the interactions between co-existing C. albicans and S. aureus, the following inclusion criteria were chosen: (i) significant change in the abundance of a particular compound relative to medium either in a log phase or in a steady phase, and (ii) significant change in abundance of this analyte in the headspace of co-culture relative to a single culture at the same stage of growth.
Accordingly, ethyl 2-methylbutyrate appeared as a co-culture specific metabolite in the log phase of microbial growth since it was absent in both single cultures but present in the co-culture. However, it discriminates only in the log phase, because it was produced by S. aureus in the steady phase in both the single and co-culture.
Figure 5.
Ethyl-2-methylbutyrate (butanoic acid, 2-methyl-, ethyl ester) synthesis in the monoculture of (a) C. albicans, (b) S. aureus, and (c) their co-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
Figure 5.
Ethyl-2-methylbutyrate (butanoic acid, 2-methyl-, ethyl ester) synthesis in the monoculture of (a) C. albicans, (b) S. aureus, and (c) their co-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
The analogous situation, where the metabolism of a particular VOC is dominated by one of the co-cultured species, concerns isobutyl acetate (Figure 6) and carbon disulfide (Figure 7), which both were strongly related to the activity of C. albicans in co-culture.
An intriguing profile was observed for methyl isobutyl ketone, which was significantly different from that observed in the reference medium only in the log phase of the co-culture (Figure 8).
The most prominent example of metabolites reflecting synergic interactions between tested bacteria and fungi was an ester Isopentyl valerate (another name: 3-methylbutyl pentanoate), whose abundance in the steady phase of co-culture was significantly higher from both single cultures (Figure 9).
3.4. Bacterial Metabolites in BAL Specimens
Application of the novel TFME extraction technique, optimized in our previous study [19], allows the detection of as many as 95 VOCs in the analyzed BAL specimens. Twenty-eight of these were also synthesized by the tested microorganisms under in vitro conditions. Some microbial metabolites detected in human specimens were found at significantly different amounts in relation to the pathogen causing pneumonia in mechanically ventilated patients. In this respect, the non-parametric ANOVA test revealed significantly different (p < 0.05) abundances of 9 metabolites between all three groups of BAL specimens (i.e., S. aureus, C. albicans and co-existence of both species) including 3-methylpyrrole, n-decane, 6-methyl-5-hepten-2-one, pyrrole, octanal, decanal, 2-cyclopenten-1-one, 4-nonene, n-nonane (Figure 10). A narrower comparison of only two groups of the analyzed specimens with a t-test indicated an additional set of compounds with statistical significance (p < 0.05). However, given the small number of BAL samples and their high natural biological heterogeneity, the result of statistical calculations in this section should be considered in terms of rough approximation rather than the rigid criteria for selecting biologically important VOC markers. Therefore, data given in the Table 2 comprises metabolites ordered according to the increasing p-value for the non-parametric ANOVA test of all three groups. The quantities of the first 35 VOCs with the lowest p-value (up to p = 0.25) averaged to each group of BAL specimens are given in the form of a heatmap as a Figure 11. The applied higher p-value limit offers a broader overview of VOC profiles observed in infectious specimens collected from the critically ill and their more relevant comparison to the in vitro experiments.
The most often observed profile of VOCs secretion from BAL specimens, in which both pathogens were identified, was related to the activity of S. aureus (as can be seen in Figure 10), but metabolites related to C. albicans were observed as well, e.g., 2-ethyl-1-hexanol, diethyl ether, 3-heptanone or cyclohexanone (Figure 12, panel A). The interactions between pathogens coexisting in BAL specimens could be observed as a synergistic effect on VOC synthesis, manifested with elevated abundances of acetic acid (panel B on Figure 12). Intriguingly, the suppressed production of volatile metabolites such as 2-butanone, 2-propanol, and ethyl acetate (panel C on Figure 12) was also found in co-cultures, although the growth rate of both microbes was regular. The overview of averaged abundances of particular VOCs for each type of BAL sample is given as a heatmap in Figure 11.
4. Discussion
Breath gas analysis has excellent applicability for VAP diagnosis and for monitoring treatments due to its non-invasiveness, ease of acquiring serial samples, and potential to characterize underlying pathogens, allowing targeted or preemptive actions in emerging pneumonia. Screening for volatile bacterial metabolites in exhaled air could support or even replace the need for the established invasive and time-intensive microbiological testing procedures. To achieve this ambitious goal, one needs a detailed knowledge of factors affecting the synthesis of microbial VOCs, particularly in the case of multi-pathogenic infections, since microorganisms can exhibit both competitive and synergistic behavior with each other and human host cells. It was shown by Filkins et al. [23] that the presence of P. aeruginosa induces a change in S. aureus metabolism and shifts its aerobic respiration to fermentation (hence the production of lactate instead of acetate) and ultimately reduces the viability of S. aureus. The suppression of S. aureus growth, especially in its early phase, was induced by E. coli, as Chen et al. [20] demonstrated by monitoring species-specific VOCs. Among other species, the synergistic behavior between P. aeruginosa and E. cloacae was observed by Lawal et al. [24], where the levels of VOCs in co-culture were significantly higher than the sum of both monocultures. The complexity of searching for volatile signatures of etiological factors (P. aeruginosa and respiratory syncytial virus, RSV) co-cultured with human cells (cystic fibrosis bronchial epithelial cells, CFBE) is well exemplified by Purcaro et al. [25], where a clear dominance of bacteria species over other constituents of the co-culture is shown, indisposing differentiation of RSV-infected from non-RSV infected CFBE cells, but enabling distinction of VOCs produced by P. aeruginosa in the absence and presence of CFBE cells.
The working hypothesis investigated here, namely that cross-kingdom interactions of S. aureus and C. albicans (frequently found opportunistic pathogen in VAP) influence the synthesis of microbial volatiles under in vitro conditions, is correct. Additionally, we tested the assumption of whether VOCs elucidated in vitro are convergent in the in vivo situation. To address adequately this complex problem, two different techniques were used for VOC sampling: (1) a dynamic adsorption on multibed tubes to follow the changing metabolism in growing cultures and (2) a static extraction on thin-film meshes to collect analytes ex-vivo in BAL specimens from VAP patients. The latter technique enables sensitive GC-MS analysis even from a small quantity of the sample (here: infectious human material) without the need for external carrier gas flow through the headspace. Compared to other static extraction techniques, it offers an enhanced adsorption efficiency and simultaneously low bleed and background due to a substantially increased stationary phase surface area. Another benefit of TFME meshes is the use of a new type of HLB adsorbent (unavailable for other commercial adsorptive techniques used in GC-MS) obtained by incorporating N-vinylpyrrolidone groups into the divinylbenzene polymer. Such a modification improves the extraction efficiency of oxygenated VOCs (i.e., aldehydes, ketones, and alcohols) due to stronger π-π interactions between adsorbent particles and electronegative elements constituting the functional groups of an analyte. Application of the TFME technique enabled precise and sensitive analysis of a wide range of VOCs in complex matrices, and the protocol applied for data evaluation allowed unambiguous identification of detected analytes.
4.1. In Vitro Experiments with Microbial Cultures
The results of the in vitro experiments testify that the proliferation rate of C. albicans and S. aureus under applied settings do not differ in monocultures compared to a co-cultivation of both species. The performed TD-GC-MS analyses suggest that each species activates partly different metabolic pathways in the subsequent growth phases resulting in a dynamically changing quantity and portfolio of VOCs secreted over time. It can be well exemplified by carbon disulfide (CS2), which C. albicans strain continuously takes up as an energy or substrate source during the entire log phase. Once the microbes enter the steady phase, their metabolism slows down, and demand for carbon disulfide reduces; hence its level in the culture headspace rises (Figure 7A). On the contrary, S. aureus cultures do not consume carbon disulfide in the logarithmic phase and even seem to release this metabolite, although the difference compared to its content in the TSB medium headspace was statistically not significant (p = 0.924). A decrease in CS2 level in S. aureus cultures was observed only at the latest times of cultivation (Figure 7B). Generally, it was frequently observed that the synthesis rate of certain metabolites was the highest when microbial division was at its biological maximum (i.e., the second part of the logarithmic phase) and was substantially reduced later. In this regard, twenty- one VOCs exhibited a temporary maximum in the log phase of C. albicans growth with diminished abundances afterward, while only fourteen VOCs reached the highest levels in their steady phase. On the contrary, the metabolism of S. aureus seems slower under applied conditions, as only 11 volatiles gained a temporary maximum in the log phase. Subsequently, the secretion of metabolites increased proportionally to the total burden of S. aureus, reaching 24 compounds with their highest amounts in a steady stage. In turn, the number of VOC produced in the co-cultures was nearly equally distributed between both proliferation phases, with 18 analytes higher in the log and 19 in the steady stage.
The most interesting metabolites in terms of bacterial-fungal interaction (in a setup used in this study) can be split into two groups. The first group comprises VOCs with profiles related to one microorganism but modified by the presence of another microbe. This is exemplified by carbon disulfide, which is clearly linked with the metabolic activity of the investigated fungi as it has the same profile in C. albicans cultured alone and mixed with S. aureus (compare Figures 7A and 7C). However, during the log phase, the abundance of carbon disulfide was significantly higher in co-culture compared to sole C. albicans (Figure 2), which can be only partly explained by the weak (statistically insignificant) production of this metabolite by S. aureus (Figure 7B). Apparently, the catabolism of carbon disulfide by C. albicans was temporarily weaker due to the presence of bacteria. The altered metabolic activity of C. albicans in co-culture seems to be confirmed also by isobutyl acetate, whose first release was observed after 6.5 h in monocultures of C. albicans but not until 25 h in co-culture. Intriguingly, despite the delayed synthesis, the ultimate abundance of isobutyl acetate was significantly higher in mixed compared to monoculture of C. albicans. Analogously, the existence of ethyl 2-methylbutyrate in co-culture is related to the metabolic activity of S. aureus, as it was never observed in a single culture of C. albicans in the present study (Figure 5A). However, the interspecies interactions lead to its much faster synthesis by S. aureus when it grows in co-culture compared to its monoculture (synthesis within 8 h instead of 25 h after inoculation). Nonetheless, there are no significant differences in the synthesis of this metabolite once the maximum density of bacteria is reached between sole and mixed cultures (Figures 5 B-C).
The second group comprises volatile metabolites whose quantities significantly increase in co-cultures due to the synergism in bacterial-fungal interactions. This is exemplified by isopentyl valerate, whose abundance increases 3-fold in the steady phase of co-culture compared to the sum of its amounts in both single cultures. Therefore, an increase in isopentyl valerate synthesis during the steady phase of co-culture cannot be explained by the cumulative effect summarizing its production by each species alone (Figures 9A-C). A similar situation concerns the release of methyl isobutyl ketone. This metabolite was detected only during the log phase in the co-culture (from 2h to 6.5 h), after which it was no longer synthesized. Noticeably, methyl isobutyl ketone was not observed in the headspace of single cultures of both species tested in this study.
4.2. Ex Vivo Analysis of Human Specimens
In the second part of our work, the bronchoalveolar lavage specimens were collected from mechanically ventilated patients known to be infected explicitly by the sole C. albicans, sole S. aureus, or both species together. Due to the unexpectedly low number of BAL specimens (available temporarily in the local hospital participating in our study) that fulfilled our inclusion criteria, the results gathered should be considered in terms of a rough comparison of volatiles synthesized by tested pathogens under in vitro (model study) and in vivo (clinical samples) conditions, and not in terms of selection of species-specific biomarkers for pathogen identification in clinical settings.
One of the most interesting observations in this comparison is the noticeably higher influence of S. aureus on the overall VOC portfolio in BAL samples, while C. albicans strain was determining metabolite range in the in vitro cultures. In this regard, the synthesis of all aldehydes and hydrocarbons was associated with the presence of S. aureus in the BAL specimens collected from critically ill patients, regardless of whether it was a sole pathogen in the sample or coexisting with C. albicans (Figure 10). These results are in agreement with the previous studies reporting aldehydes (with 3-methylbutanal being particularly abundant) as an important group of volatile metabolites synthesized by S. aureus [21, 26]. Another example is carbon disulfide, which was observed at elevated levels when S. aureus was present in both human specimens and co-cultures. On the other hand, the activity of Candida albicans in BAL specimens determined the excretion of certain alcohols and ketones, including 2-ethyl-1-hexanol, 2-methyl-2-butanol, 3-heptanone or cyclohexanone (Table 2). Importantly, none of these compounds was metabolized under in vitro conditions by the pathogens tested in this study. There is a curious case of 3-heptanone, as this ketone was observed only in BAL samples, whereas 4-heptanone was present in the in vitro cultures of these fungi (significantly released during a log phase of mono- and co-cultures). Since both the mass spectrum and the retention time from the chromatographic separation are different for these two compounds, there is no risk of mistaking them. Furthermore, 4-heptanone was previously reported in the pilot clinical study on pathogen identification using breath gas analysis in VAP patients infected with Candida albicans [13]. The reason for the discrepancy between 3-heptanone in biological material and 4-heptanone in cultures is unknown.
The interactions between pathogens co-existing in BAL specimens were also observed as their synergistic effect on VOC synthesis manifested with elevated abundances of acetic acid (panel B on Figure 12). This compound was synthesized under in vitro conditions in small quantities by both C. albicans in its log phase and by S. aureus in its steady growth phase. Still, the levels of acetic acid in co-cultures were lower than in both monocultures, whereas in BAL it reached the highest amounts in specimens containing both pathogens. In contrast, volatile metabolites such as 2-butanone, acrolein, and ethyl acetate were suppressed in BAL samples with mixed pathogens (panel C on Figure 12). Amongst them, the synthesis of ethyl acetate was directly proportional and acrolein was inversely proportional to the proliferation rate of C. albicans in mono- and co-cultures.
4.3. Comparison of the In Vitro and In Vivo Metabolome
Consistent and inconsistent results between in vitro and ex vivo / in vivo experiments are important because they demonstrate the complexity of volatilome studies and the need for further research, especially with human materials (BAL, blood, etc.).
Overall, the presented data show that the co-existence of both species under in vitro conditions does not affect their proliferation rate compared to monocultures, but that it does affect the synthesis of their volatile metabolites. Under optimal growth conditions, C. albicans dominates the volatilome in the co-cultures. In BAL material, S. aureus seems to have benefited from the co-existence with C. albicans (Figure 10) and adjusts the portfolio of VOCs to the new metabolic conditions in a far more complex matrix. S. aureus enhances the synthesis of new compounds, particularly hydrocarbons and aldehydes, while suppressing the production of esters (compared to in vitro conditions). Most of the produced VOCs are associated with S. aureus in this situation. The reason why some metabolites exhibit distinct profiles in culture and biological material remains unknown. Still, the co-existence of both tested species in the biological sample yields a particular volatile metabolic signature, with acetic acid, methyl methacrylate and carbon disulfide being the most prominent examples.
Additional studies with other relevant pathogens should be performed to investigate further the inter-species interactions and their impact on the synthesis of volatile metabolites. For clinical relevance in pneumonia diagnosis, the focus should be on the analysis of the most prevalent pathogens’ combinations with additional clinical impact on ICU patients. An example of this is an investigation of possible antibiotic treatment with its implied changes in pathogen metabolism. In addition to in vitro and ex vivo studies, breath gas analysis of ICU patients with confirmed coinfections (with the pathogens of interest) should be compared to the present in vitro study.
4.4. Study Limitation
Despite the newest available data from the NHSN report indicating S. aureus to be the most dominant pathogen in VAP patients (29,6% of cases in the USA) [1], its prevalence in the ICU ward participating in our study (Bydgoszcz, Poland) was considerably lower (9,8% amongst all 92 BAL samples analyzed with TFME-GC-MS). Therefore, the small number of BAL specimens analyzed, which fulfill the inclusion criteria - either sole C. albicans or S. aureus or the presence of both – together with the variability of clinical conditions of VAP patients, potentially affect the composition of the isolated BAL specimens, which can be considered a limitation of our work. Nevertheless, the key aims of this work have been achieved, namely that the volatile metabolites released by the tested microorganisms in single- and co-cultures were identified, their dynamically changing profiles were recognized, and part of them was further found in biological material isolated from patients infected with these pathogens, demonstrating the suitability of a novel microextraction technique for analysis of clinical samples.
4.5. Concluding Remarks
To determine whether putative interactions between S. aureus and C. albicans affect the secretion of VOCs under in vitro conditions, the solid phase extraction on multibed sorption tubes was used for sample collection and VOC preconcentration before GC-MS analysis. To collect analytes ex vivo in BAL specimens from VAP patients, the static extraction on thin-film meshes coated with a novel hydrophilic-lipophilic-balanced adsorbent was used. Both analytical protocols proved well suited for the chosen application, allowing the precise and reproducible analysis of the microbial volatilome.
The gathered data demonstrated the importance of considering the growth phase in which bacteria sampling is done when comparing the volatilomes of certain strains determined in unrelated studies. In this regard, one must pay particular attention to selecting “biomarkers” for pathogen identification, especially if applied to clinical settings with all putative consequences, such as treatment choice. It is noticeable that most of the VOCs in co-culture are associated with C. albicans metabolism, and some of them exclusively originate from this yeast (e.g., isobutyl acetate), while considerably fewer VOCs are released from S. aureus and then mainly in the latest phase of its growth (e.g., ethyl-2-methylbutyrate). Even more interesting are compounds such as methyl isobutyl ketone, which exhibit no difference from the control medium in a single culture but significantly differ in the co-culture compared to the control medium. In the BAL material collected from VAP patients, S. aureus seems to adjust the portfolio of VOCs to the new metabolic conditions and enhances the synthesis of new compounds, particularly hydrocarbons and aldehydes, while suppressing the production of esters and alcohols (compared to in vitro conditions). The coexistence of both tested species in the biological sample yields a volatile metabolic signature, including acetic acid, methyl methacrylate and carbon disulfide. Altogether, these observations give molecular evidence of the presence of the respective single microbe on the one hand and suspect potential multi-pathogenic infections on the other hand. The reported microbial metabolites should be used to pursue non-invasive VAP diagnosis in future clinical studies.
Author Contributions
Wojciech Filipiak: Conceptualization, Methodology, Validation, Formal analysis, Visualization, Investigation, Resources, Writing- Original draft preparation, Writing—Review and editing, Supervision, Project administration, Funding acquisition. Matthias Wenzel: Validation, Formal analysis, Investigation, Writing- Original draft preparation, Writing—Review and editing. Clemens Ager: Software, Validation, Formal Analysis, Visualization, Data curation. Chris A. Mayhew: Conceptualization, Resources, Writing- Original draft preparation, Writing—Review and editing, Funding acquisition. Tomasz Bogiel: Writing- Original draft preparation, Writing—Review and editing. Robert Włodarski: Investigation, Resources, Writing- Original draft preparation, Writing—Review and editing. Markus Nagl: Conceptualization, Investigation, Resources, Writing—Review and editing, Supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NATIONAL SCIENCE CENTRE, POLAND, grant number 2017/26/D/NZ6/00136.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Bioethics Committee of the Nicolaus Copernicus University in Toruń at the Ludwik Rydygier Collegium Medicum in Bydgoszcz under the number KB-218/2018.
Data Availability Statement
The dataset used and analyzed during the current study is available from the corresponding author upon reasonable request.
Acknowledgments
The authors are very thankful to Joanna Sierzputowska from the Department of Medical Analytics of the 10th Military Research Hospital and Polyclinic, Bydgoszcz, Poland, who kindly provided all clinical BAL specimens isolated from VAP patients and performed microbiological identification of detected strains. The authors are also very grateful to Janusz Pawliszyn (University of Waterloo, Canada), who kindly provided all TFME meshes. W.F. is grateful to the National Science Centre, Poland, for the financial support from Project No.: 2017/26/D/NZ6/00136.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Network, C. f. D. C. a. P.-N. H. S. HAI Pathogens and Antimicrobial Resistance Report, 2018 - 2021; Centers for Disease Control and Prevention: 2023.
- Hamet, M.; Pavon, A.; Dalle, F.; Pechinot, A.; Prin, S.; Quenot, J. P.; Charles, P. E. , Candida spp. airway colonization could promote antibiotic-resistant bacteria selection in patients with suspected ventilator-associated pneumonia. Intensive Care Med 2012, 38, (8), 1272-9.
- Azoulay, E.; Timsit, J. F.; Tafflet, M.; de Lassence, A.; Darmon, M.; Zahar, J. R.; Adrie, C.; Garrouste-Orgeas, M.; Cohen, Y.; Mourvillier, B.; Schlemmer, B.; Outcomerea Study, G. , Candida colonization of the respiratory tract and subsequent pseudomonas ventilator-associated pneumonia. Chest. 2006, 129, (1), 110-7.. [CrossRef] [PubMed]
- Harriott, M. M.; Noverr, M. C. , Candida albicans and Staphylococcus aureus form polymicrobial biofilms: Effects on antimicrobial resistance. Antimicrob Agents Chemother 2009, 53, 3914–22. [Google Scholar] [CrossRef]
- McAlester, G.; O'Gara, F.; Morrissey, J. P. , Signal-mediated interactions between Pseudomonas aeruginosa and Candida albicans. J Med Microbiol 2008, 57 Pt 5, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E. , Effect of strain of Staphylococcus aureus on synergism with Candida albicans resulting in mouse mortality and morbidity. Infect Immun 1983, 42, 285–92. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E. C. , Synergism of Candida albicans and delta toxin producing Staphylococcus aureus on mouse mortality and morbidity: Protection by indomethacin. Zentralbl Bakteriol Mikrobiol Hyg A 1988, 269, 377–86. [Google Scholar] [CrossRef] [PubMed]
- Roux, D.; Gaudry, S.; Dreyfuss, D.; El-Benna, J.; de Prost, N.; Denamur, E.; Saumon, G.; Ricard, J. D. , Candida albicans impairs macrophage function and facilitates Pseudomonas aeruginosa pneumonia in rat. Crit Care Med 2009, 37, 1062–7. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, M. P.; Pham, D.; Fein, M.; Kong, L.; Weisman, L. E. , Neonatal coinfection model of coagulase-negative Staphylococcus (Staphylococcus epidermidis) and Candida albicans: Fluconazole prophylaxis enhances survival and growth. Antimicrob Agents Chemother 2007, 51, 1240–5. [Google Scholar] [CrossRef] [PubMed]
- Delisle, M. S.; Williamson, D. R.; Perreault, M. M.; Albert, M.; Jiang, X.; Heyland, D. K. , The clinical significance of Candida colonization of respiratory tract secretions in critically ill patients. J Crit Care 2008, 23, 11–7. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Qi, M.; Hu, Y.; Yu, M.; Liang, Z. , The impact of Candida spp airway colonization on clinical outcomes in patients with ventilator-associated pneumonia: A systematic review and meta-analysis. Am J Infect Control 2020, 48, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Ricard, J. D.; Roux, D. , Candida colonization in ventilated ICU patients: No longer a bystander! Intensive Care Med 2012, 38, 1243–5. [Google Scholar] [CrossRef] [PubMed]
- Filipiak, W.; Beer, R.; Sponring, A.; Filipiak, A.; Ager, C.; Schiefecker, A.; Lanthaler, S.; Helbok, R.; Nagl, M.; Troppmair, J.; Amann, A. , Breath analysis for in vivo detection of pathogens related to ventilator-associated pneumonia in intensive care patients: A prospective pilot study. J Breath Res 2015, 9, 016004. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, R.; Fijten, R.; Smolinska, A.; Dallinga, J.; Boumans, M. L.; Stobberingh, E.; Boots, A.; Roekaerts, P.; Bergmans, D.; van Schooten, F. J. , Analysis of volatile organic compounds in exhaled breath to diagnose ventilator-associated pneumonia. Sci Rep 2015, 5, 17179. [Google Scholar] [CrossRef] [PubMed]
- van Oort, P. M.; de Bruin, S.; Weda, H.; Knobel, H. H.; Schultz, M. J.; Bos, L. D.; On Behalf Of The Mars, C. , Exhaled Breath Metabolomics for the Diagnosis of Pneumonia in Intubated and Mechanically-Ventilated Intensive Care Unit (ICU)-Patients. Int J Mol Sci 2017, 18, (2).
- Trefz, P.; Schmidt, M.; Oertel, P.; Obermeier, J.; Brock, B.; Kamysek, S.; Dunkl, J.; Zimmermann, R.; Schubert, J. K.; Miekisch, W. , Continuous real time breath gas monitoring in the clinical environment by proton-transfer-reaction-time-of-flight-mass spectrometry. Anal Chem 2013, 85, 10321–9. [Google Scholar] [CrossRef] [PubMed]
- Żuchowska, K.; Filipiak, W. , Modern approaches for detection of volatile organic compounds in metabolic studies focusing on pathogenic bacteria: Current state of the art. Journal of Pharmaceutical Analysis 2024, 14, 100898. [Google Scholar] [CrossRef] [PubMed]
- Filipiak, W.; Ager, C.; Troppmair, J. , Predicting the future from the past: Volatile markers for respiratory infections. Eur Respir J 2017, 49, (5).
- Filipiak, W.; Jaroch, K.; Szeliska, P.; Zuchowska, K.; Bojko, B. , Application of Thin-Film Microextraction to Analyze Volatile Metabolites in A549 Cancer Cells. Metabolites 2021, 11, (10).
- Filipiak, W.; Zuchowska, K.; Marszalek, M.; Depka, D.; Bogiel, T.; Warmuzinska, N.; Bojko, B. , GC-MS profiling of volatile metabolites produced by Klebsiella pneumoniae. Front Mol Biosci 2022, 9, 1019290. [Google Scholar] [CrossRef] [PubMed]
- Filipiak W. Sponring A., F. A., Baur M., Ager C., Wiesenhofer H., Margesin R., Nagl M., Troppmair J., Amann A., Volatile Organic Compounds (VOCs) Released by Pathogenic Microorganisms in vitro: Potential Breath Biomarkers for Early-Stage Diagnosis of Disease In Volatile Biomarkers: Non-Invasive Diagnosis in Physiology and Medicine, 1st ed.; Amann A., S. D., Ed. Elsevier: Amsterdam, The Netherlands, 2013; pp 463-512.
- Grandy, J. J.; Singh, V.; Lashgari, M.; Gauthier, M.; Pawliszyn, J. , Development of a Hydrophilic Lipophilic Balanced Thin Film Solid Phase Microextraction Device for Balanced Determination of Volatile Organic Compounds. Anal Chem 2018, 90, 14072–14080. [Google Scholar] [CrossRef] [PubMed]
- Filkins, L. M.; Graber, J. A.; Olson, D. G.; Dolben, E. L.; Lynd, L. R.; Bhuju, S.; O'Toole, G. A. , Coculture of Staphylococcus aureus with Pseudomonas aeruginosa Drives S. aureus towards Fermentative Metabolism and Reduced Viability in a Cystic Fibrosis Model. J Bacteriol 2015, 197, 2252–64. [Google Scholar]
- Lawal, O.; Knobel, H.; Weda, H.; Bos, L. D.; Nijsen, T. M. E.; Goodacre, R.; Fowler, S. J. , Volatile organic compound signature from co-culture of lung epithelial cell line with Pseudomonas aeruginosa. Analyst 2018, 143, 3148–3155. [Google Scholar] [CrossRef]
- Purcaro, G.; Rees, C. A.; Melvin, J. A.; Bomberger, J. M.; Hill, J. E. , Volatile fingerprinting of Pseudomonas aeruginosa and respiratory syncytial virus infection in an in vitro cystic fibrosis co-infection model. J Breath Res 2018, 12, 046001. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, J.; Shi, H.; Tang, C.; Zhang, R. , Characteristics of volatile organic compounds produced from five pathogenic bacteria by headspace-solid phase micro-extraction/gas chromatography-mass spectrometry. J Basic Microbiol 2017, 57, 228–237. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Growth curves of (a) single cultures of C. albicans and S. aureus, (b) C. albicans in single- and co-culture, (c) S. aureus in single- and co-culture. C.a. – Candida albicans; S.a. – Staphylococcus aureus.
Figure 1.
Growth curves of (a) single cultures of C. albicans and S. aureus, (b) C. albicans in single- and co-culture, (c) S. aureus in single- and co-culture. C.a. – Candida albicans; S.a. – Staphylococcus aureus.
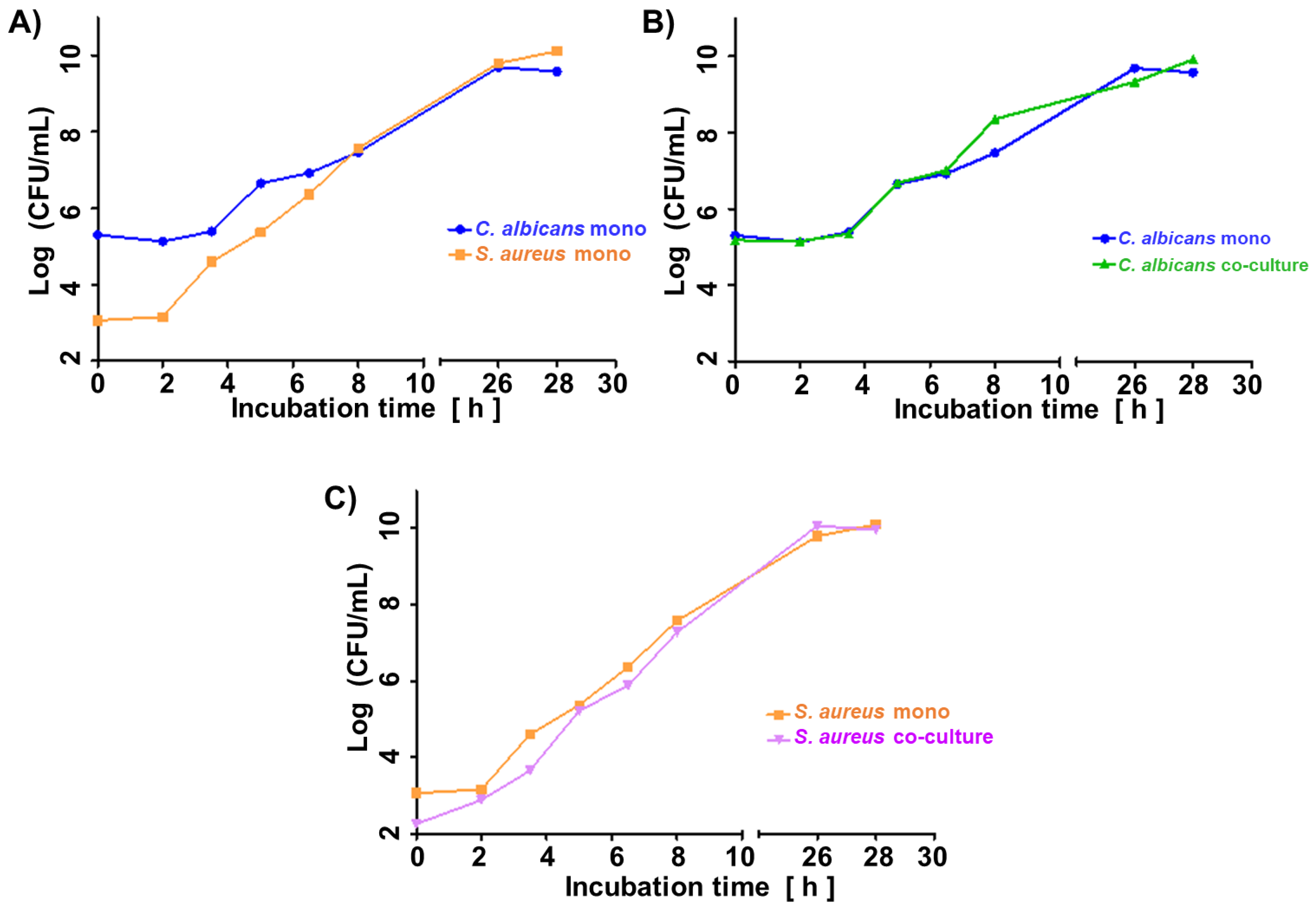
Figure 2.
Relative abundance of VOCs +/- standard error (horizontal whiskers) in co-culture compared to monoculture of Candida albicans given separately for log phase (A) and steady phase (B) of microbial growth. Positive values indicate a higher amount of respective VOC in co-culture than in a single culture.
Figure 2.
Relative abundance of VOCs +/- standard error (horizontal whiskers) in co-culture compared to monoculture of Candida albicans given separately for log phase (A) and steady phase (B) of microbial growth. Positive values indicate a higher amount of respective VOC in co-culture than in a single culture.
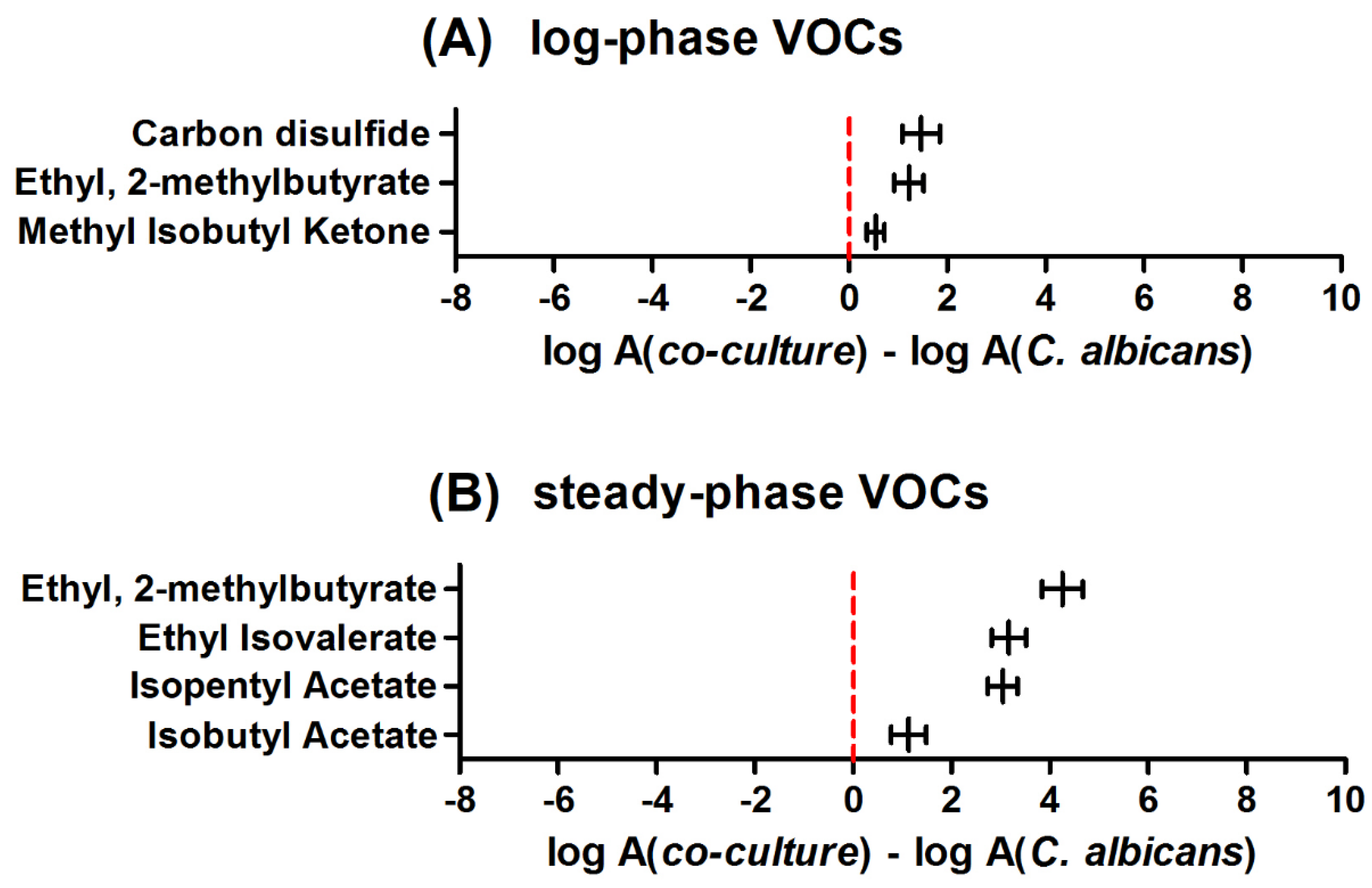
Figure 3.
Relative abundance of VOCs +/- standard error (horizontal whiskers) in co-culture compared to monoculture of Staphylococcus aureus in the log phase of growth. Positive values indicate a higher amount of respective VOC in a co-culture than in a single culture of S. aureus, and vice versa; negative values indicate metabolites more abundant in a monoculture compared to a co-culture.
Figure 3.
Relative abundance of VOCs +/- standard error (horizontal whiskers) in co-culture compared to monoculture of Staphylococcus aureus in the log phase of growth. Positive values indicate a higher amount of respective VOC in a co-culture than in a single culture of S. aureus, and vice versa; negative values indicate metabolites more abundant in a monoculture compared to a co-culture.
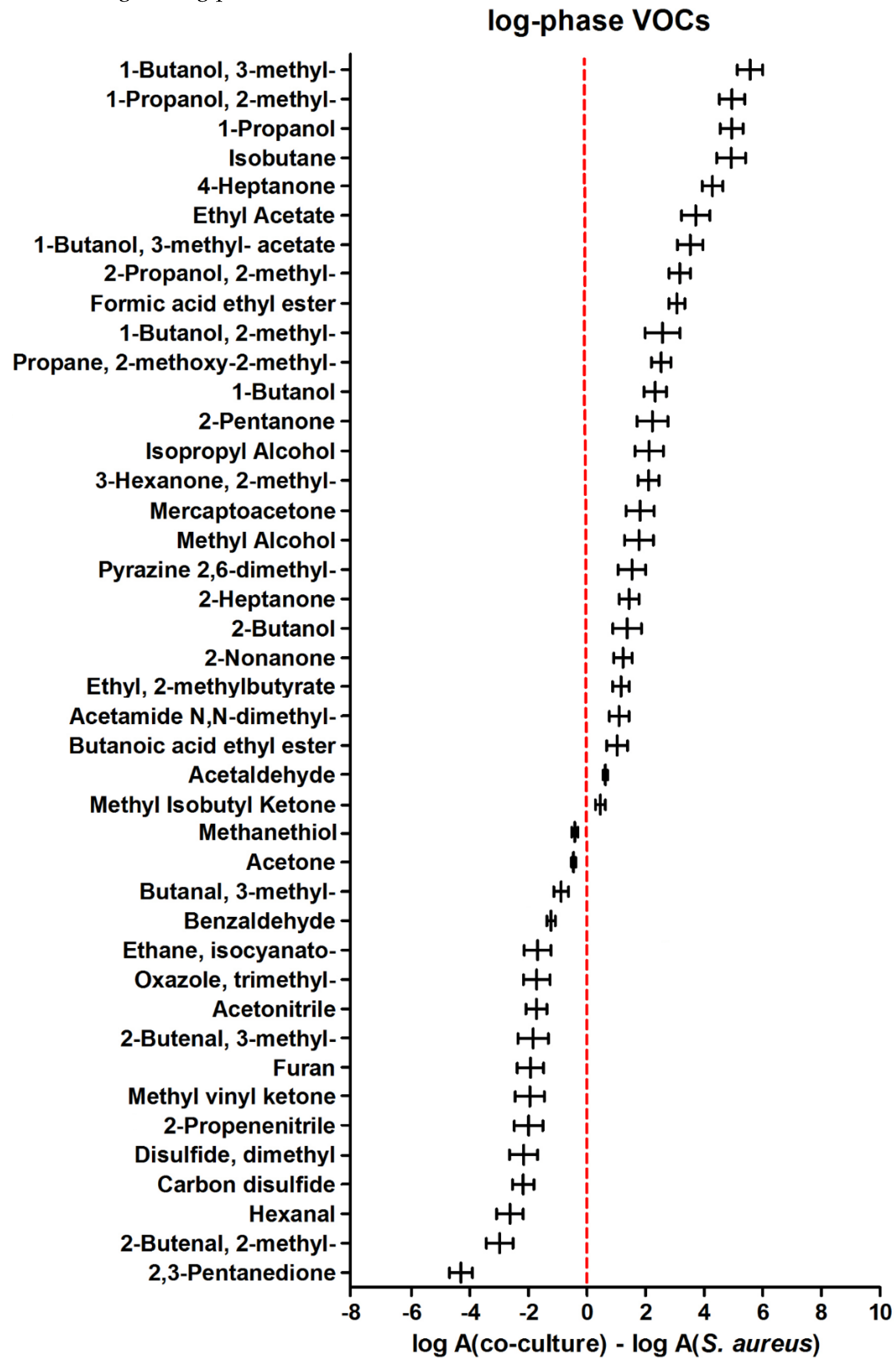
Figure 4.
Relative abundance of VOCs +/- standard error (horizontal whiskers) in co-culture compared to monoculture of Staphylococcus aureus in the steady phase of growth. Positive values indicate a higher amount of respective VOC in a co-culture than in a single culture, and vice versa; negative values indicate metabolites more abundant in a monoculture compared to a co-culture.
Figure 4.
Relative abundance of VOCs +/- standard error (horizontal whiskers) in co-culture compared to monoculture of Staphylococcus aureus in the steady phase of growth. Positive values indicate a higher amount of respective VOC in a co-culture than in a single culture, and vice versa; negative values indicate metabolites more abundant in a monoculture compared to a co-culture.
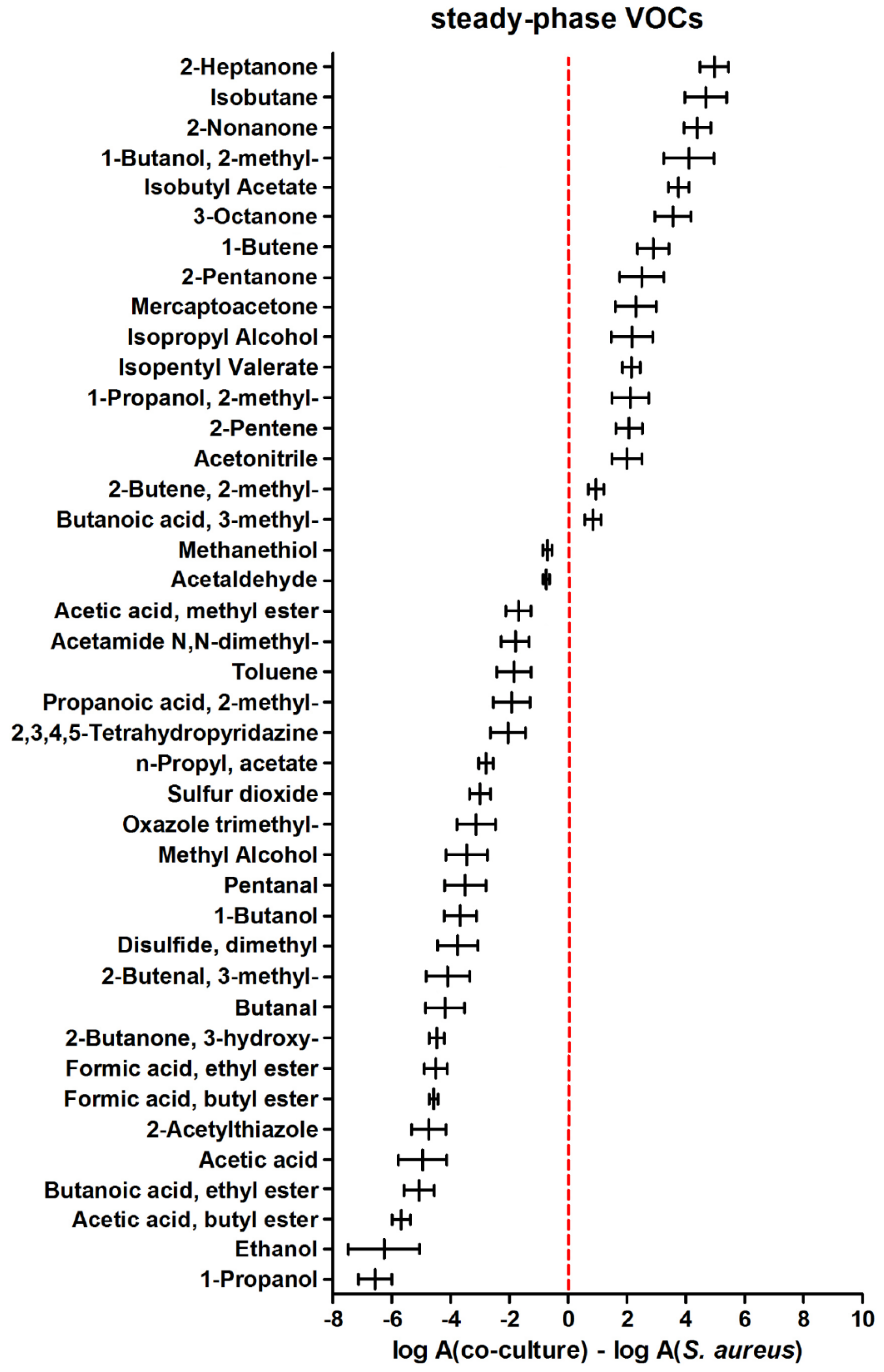
Figure 6.
Isobutyl acetate (Acetic acid, 2-methylpropyl ester) synthesis in the mono-culture of (a) C. albicans and (c) its co-culture with S. aureus. Isobutyl acetate was not detected in the S. aureus mono-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
Figure 6.
Isobutyl acetate (Acetic acid, 2-methylpropyl ester) synthesis in the mono-culture of (a) C. albicans and (c) its co-culture with S. aureus. Isobutyl acetate was not detected in the S. aureus mono-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
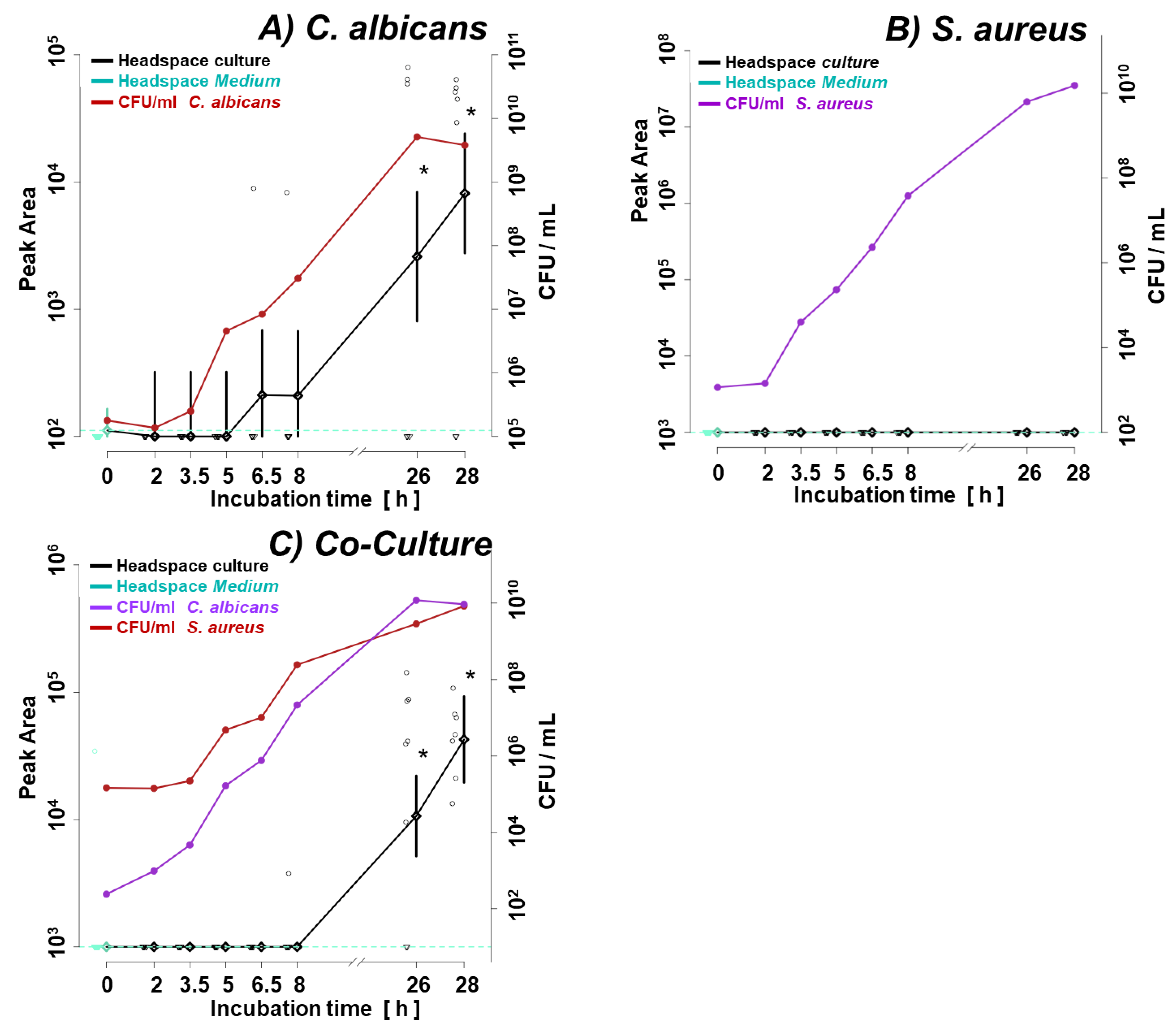
Figure 7.
Carbon disulfide (CS2) synthesis in the mono-culture of (a) C. albicans, (b) S. aureus, and (c) their co-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
Figure 7.
Carbon disulfide (CS2) synthesis in the mono-culture of (a) C. albicans, (b) S. aureus, and (c) their co-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
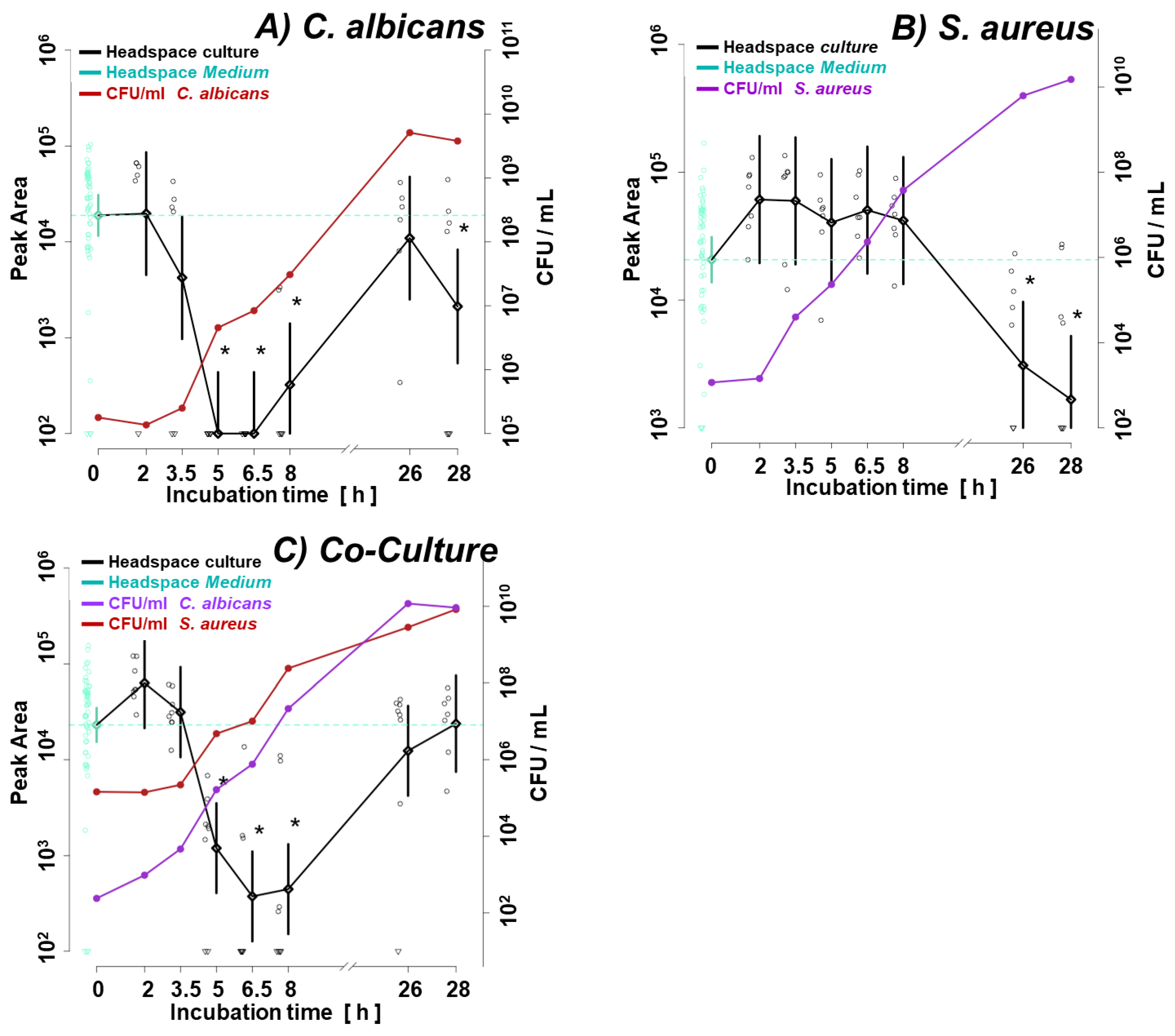
Figure 8.
Methyl Isobutyl Ketone synthesis in the co-culture with of C. albicans with S. aureus. Methyl Isobutyl Ketone was not detected in the mono-cultures of either microorganism. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
Figure 8.
Methyl Isobutyl Ketone synthesis in the co-culture with of C. albicans with S. aureus. Methyl Isobutyl Ketone was not detected in the mono-cultures of either microorganism. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
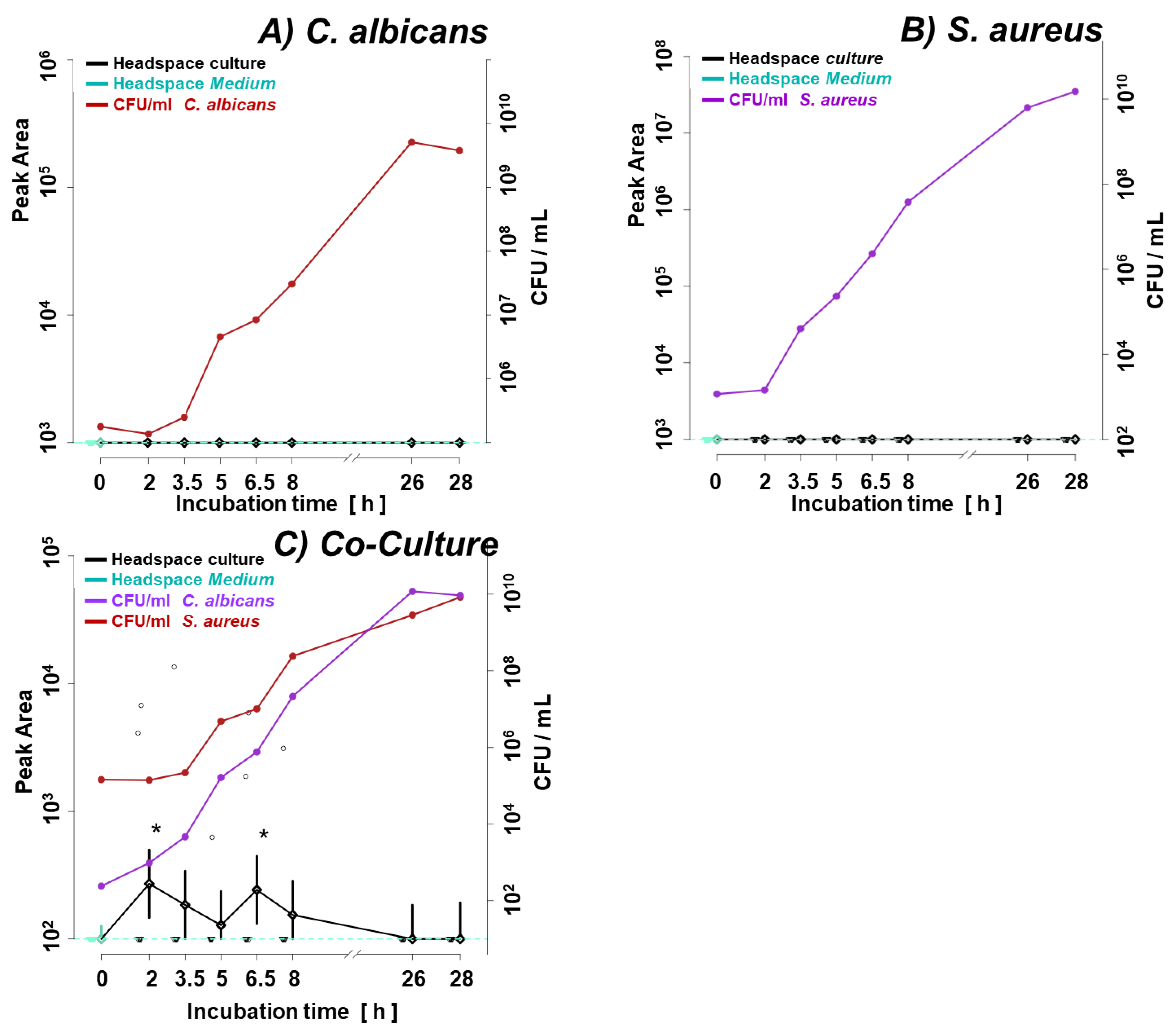
Figure 9.
Isopentyl valerate (pentanoic acid, 3-methylbutyl ester) synthesis in the mono-culture of (a) C. albicans, (b) S. aureus, and (c) their co-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
Figure 9.
Isopentyl valerate (pentanoic acid, 3-methylbutyl ester) synthesis in the mono-culture of (a) C. albicans, (b) S. aureus, and (c) their co-culture. Significant (p < 0.05) differences between VOC levels in the headspace of culture (black line) and medium control (dotted turquoise line) are marked with a black asterisk. Circles indicate the measured metabolite abundance in the headspace at a particular time point in an individual experiment. A black line connects the median values of peak areas corresponding to the particular time point (along with the whiskers indicating the 25th and 75th percentile), while a turquoise line denotes the cut-off value being the median of a given VOC in medium headspace at all time points within the experiments.
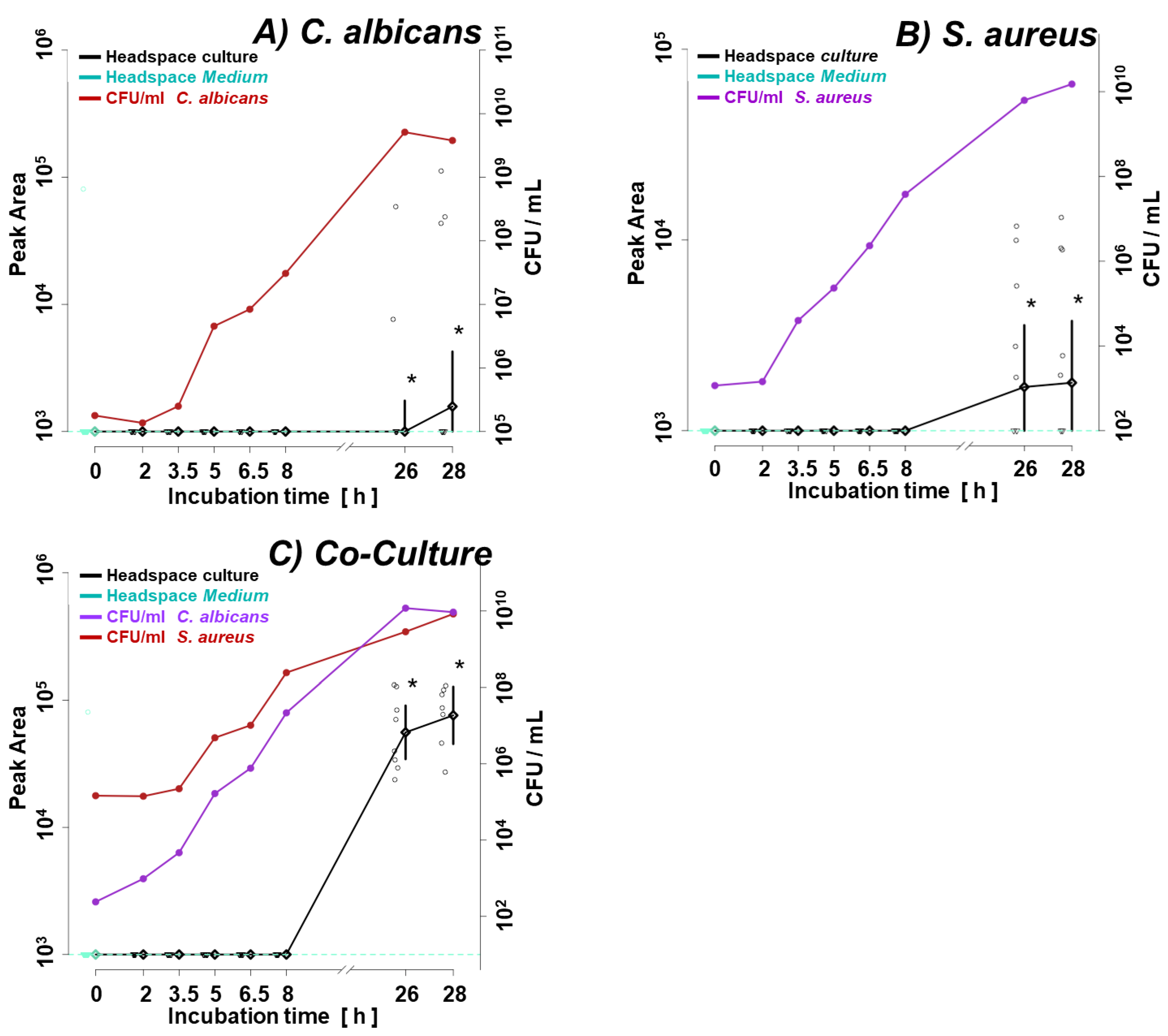
Figure 10.
Quantities of VOCs significant (ANOVA p < 0.05) for comparison between pathogens found in all BAL samples isolated from VAP patients, where SAU is for specimen with S. aureus (n = 3), CAL for C. albicans (n = 6), and SAU+CAL for BAL specimens with both species (n = 4). Additionally, p-values of the student t-test are given to compare two groups of interest, whereby significant differences from the non-parametric t-test (p < 0.05) are marked in blue. The black dots represent the abundance of the selected substance from all samples, whereby the whiskers indicates the 95% confidence interval around the median of each group and the yellow diamonds indicate the mean concentration of each group.
Figure 10.
Quantities of VOCs significant (ANOVA p < 0.05) for comparison between pathogens found in all BAL samples isolated from VAP patients, where SAU is for specimen with S. aureus (n = 3), CAL for C. albicans (n = 6), and SAU+CAL for BAL specimens with both species (n = 4). Additionally, p-values of the student t-test are given to compare two groups of interest, whereby significant differences from the non-parametric t-test (p < 0.05) are marked in blue. The black dots represent the abundance of the selected substance from all samples, whereby the whiskers indicates the 95% confidence interval around the median of each group and the yellow diamonds indicate the mean concentration of each group.
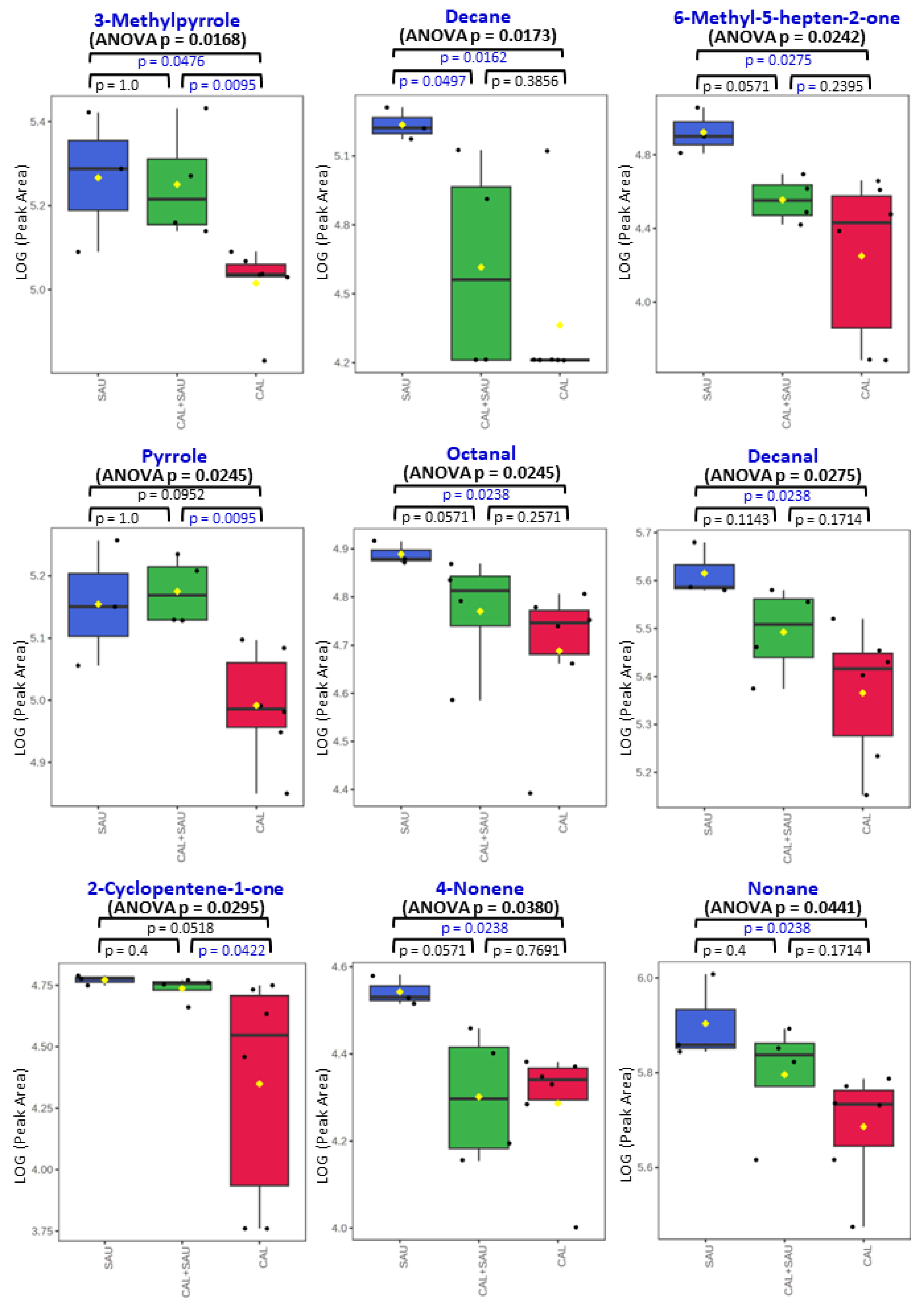
Figure 11.
Heatmap representing averaged abundances of 35 VOCs in BAL samples for which ANOVA p-values were up to 0.25 for comparison of all three groups of BAL specimens. Color contrast is autoscaled individually to each feature (substance) across all sample types (BAL specimens with different pathogens).
Figure 11.
Heatmap representing averaged abundances of 35 VOCs in BAL samples for which ANOVA p-values were up to 0.25 for comparison of all three groups of BAL specimens. Color contrast is autoscaled individually to each feature (substance) across all sample types (BAL specimens with different pathogens).
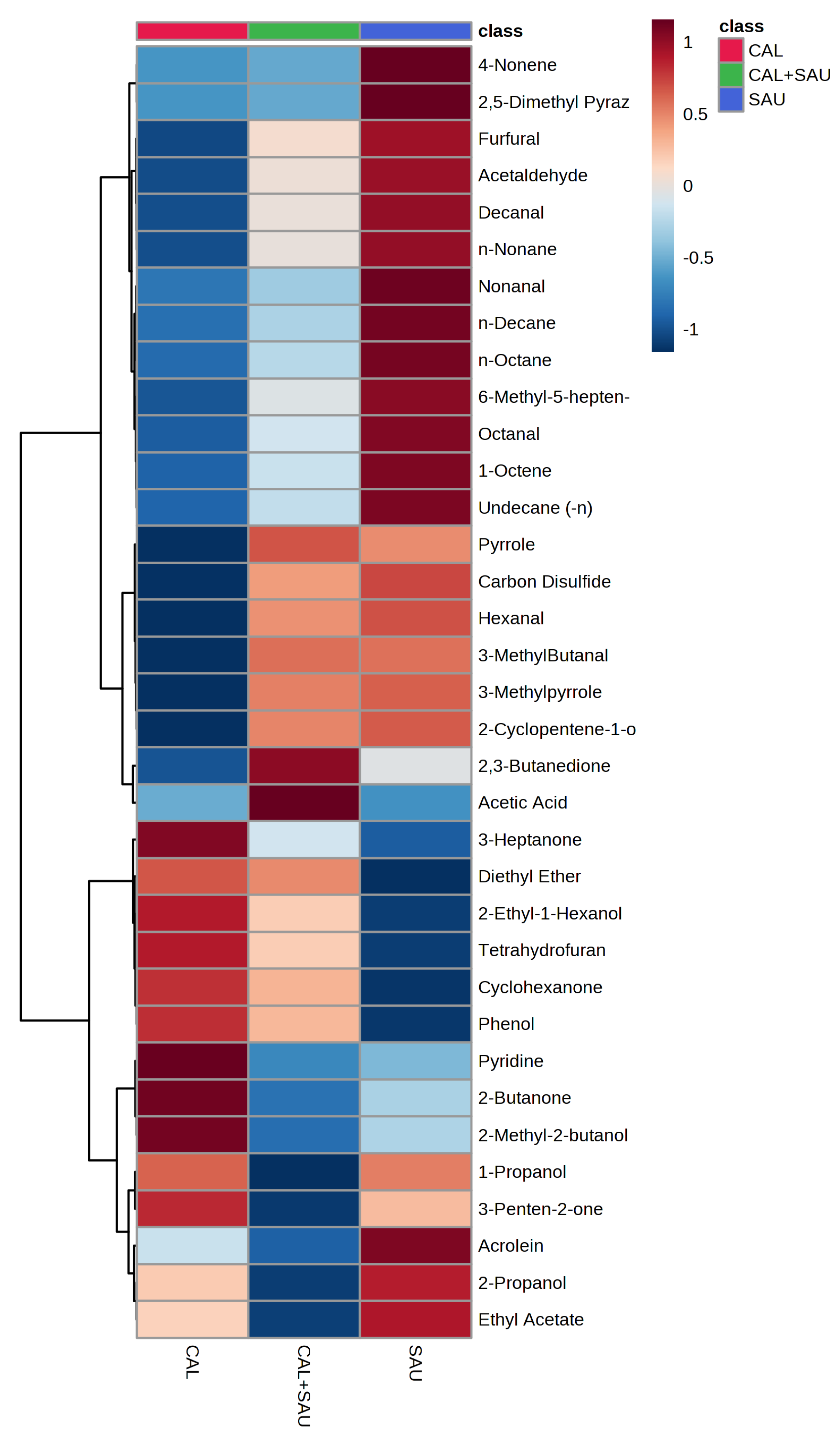
Figure 12.
Comparison of VOC profiles in BAL specimens collected from VAP patients: panel A represents metabolites related to the activity of C. albicans; panel B comprises VOCs elevated in BAL specimens with both pathogens investigated, while panel C demonstrates VOCs suppressed in BAL samples containing both studied species. ANOVA p-values (non-parametric) concern the comparison of all three BAL sample types together, and the remaining p-values concern the non-parametric t-test comparison of only two groups, indicated on plots with whiskers (statistical significance for p < 0.05 is marked in blue). The black dots represent the abundance of the selected substance from all samples, whereby the whiskers indicates the 95% confidence interval around the median of each group and the yellow diamonds indicate the mean concentration of each group.
Figure 12.
Comparison of VOC profiles in BAL specimens collected from VAP patients: panel A represents metabolites related to the activity of C. albicans; panel B comprises VOCs elevated in BAL specimens with both pathogens investigated, while panel C demonstrates VOCs suppressed in BAL samples containing both studied species. ANOVA p-values (non-parametric) concern the comparison of all three BAL sample types together, and the remaining p-values concern the non-parametric t-test comparison of only two groups, indicated on plots with whiskers (statistical significance for p < 0.05 is marked in blue). The black dots represent the abundance of the selected substance from all samples, whereby the whiskers indicates the 95% confidence interval around the median of each group and the yellow diamonds indicate the mean concentration of each group.
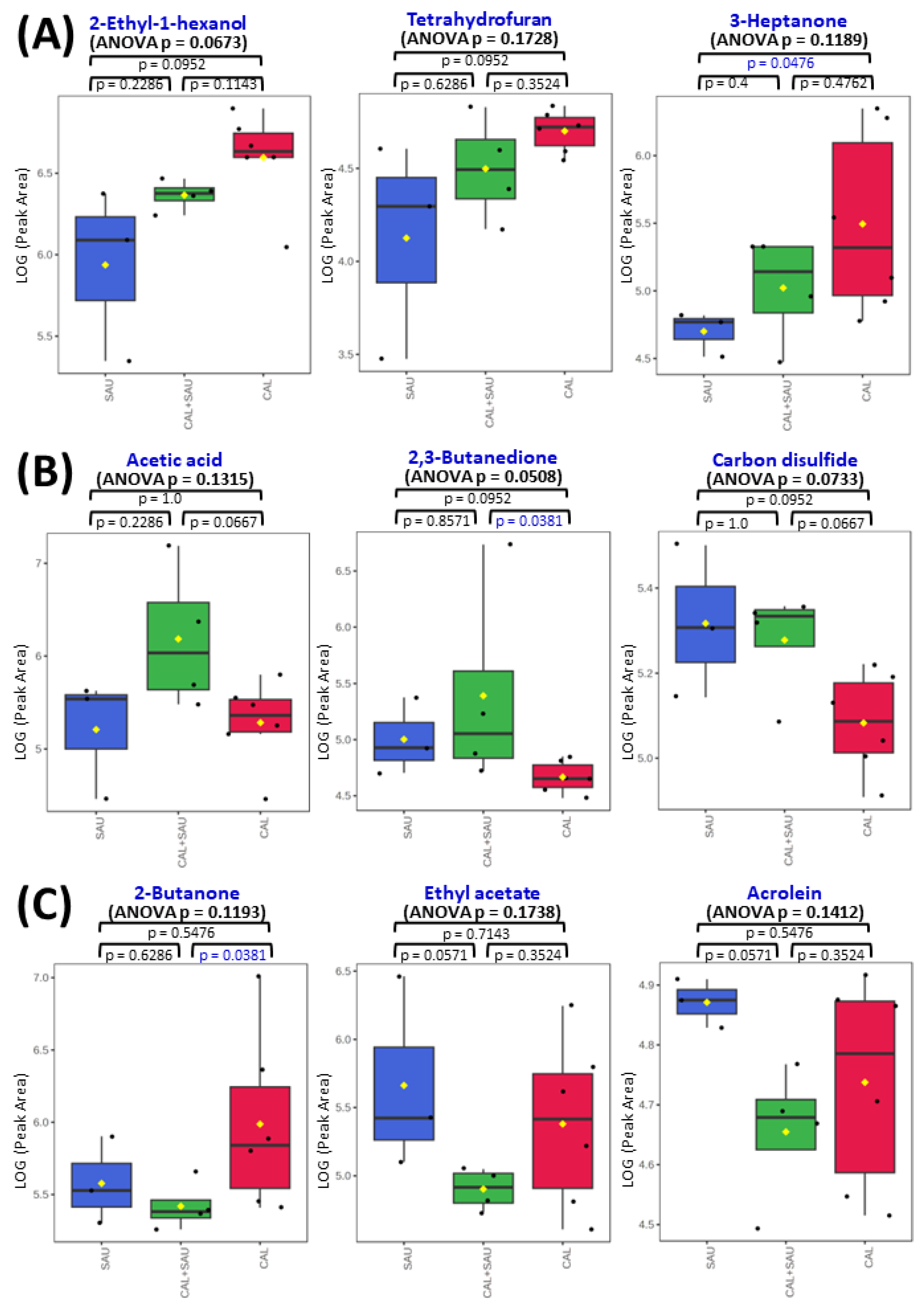

Table 1.
List of compounds identified in the investigated microbial cultures. Status “↑” indicates the significantly (p < 0.05) increased, respectively “↓“ decreased levels in culture compared to reference medium, period of growth phase during which significant change was observed is given in brackets. CAS is a substance-unique number in Chemical Abstract Service, tR is a retention time [min] of a substance in a given GC program, Target Ion is an extracted ion used for integration of a chromatographic peak area, Reference Ions were used to additionally confirm substance identification during integration, CAL stands for Candida albicans cultures, SAU for Staphylococcus aureus, Co-Culture is a mix of both microorganisms.
Table 1.
List of compounds identified in the investigated microbial cultures. Status “↑” indicates the significantly (p < 0.05) increased, respectively “↓“ decreased levels in culture compared to reference medium, period of growth phase during which significant change was observed is given in brackets. CAS is a substance-unique number in Chemical Abstract Service, tR is a retention time [min] of a substance in a given GC program, Target Ion is an extracted ion used for integration of a chromatographic peak area, Reference Ions were used to additionally confirm substance identification during integration, CAL stands for Candida albicans cultures, SAU for Staphylococcus aureus, Co-Culture is a mix of both microorganisms.
| Class | No. | Compound Name | Common Name | CAS | tR [min] | Target Ion [m/z] | Refererence Ions [m/z] | CAL | SAU | Co-Culture |
|---|---|---|---|---|---|---|---|---|---|---|
| VOCs with confirmed retention times of reference standards | ||||||||||
| Alcohols | 1 | Methyl alcohol | Methanol | 67-56-1 | 7.66 | 31 | 32, 29 | ↑ (log) | ↑ (steady) | ↑ (log) |
| 2 | Ethyl alcohol | Ethanol | 64-17-5 | 12.29 | 31 | 45, 46, 43 | ↑ (log) | ↑ (steady) | ↑ (log)↓ (steady) | |
| 3 | 2-Propanol | Isopropanol | 67-63-0 | 18.03 | 45 | 43, 41, 59 | ↑ (log, steady) | ↑ (log, steady) | ||
| 4 | n-Propyl alcohol | 1-Propanol | 71-23-8 | 22.30 | 31 | 60, 59, 27 | ↑ (log) | ↑ (steady) | ↑ (log) | |
| 5 | 2-Propanol, 2-methyl- | tert-Butanol | 75-65-0 | 25.61 | 59 | 57, 60, 51 | ↑ (log, steady) | ↑ (steady) | ↑ (log, steady) | |
| 6 | 1-Propanol, 2-methyl- | Isobutanol | 78-83-1 | 31.66 | 41 | 43, 74, 31 | ↑ (log, steady) | ↑ (steady) | ↑ (log, steady) | |
| 7 | n-Butyl alcohol | 1-Butanol | 71-36-3 | 43.76 | 56 | 31, 41, 55 | ↑ (log) | ↑ (steady) | ↑ (log) | |
| 8 | 1-Butanol, 3-methyl- | Isopentanol | 123-51-3 | 45.40 | 55 | 70, 42, 41 | ↑ (log, steady) | ↑ (steady) | ↑ (log, steady) | |
| Aldehydes | 9 | Acetic aldehyde | Acetaldehyde | 75-07-0 | 8.06 | 29 | 43, 44, 42 | ↑ (log) | ↑ (steady) | ↑ (log) ↓ (steady) |
| 10 | 2-Propenal | Acrolein | 107-02-8 | 13.95 | 27 | 56, 55, 53 | ↓ (log) | |||
| 11 | Propanal, 2-methyl- | Isobutanal | 78-84-2 | 24.69 | 72 | 41, 43, 39 | ↓ (steady) | |||
| 12 | Butyrylaldehyde | Butanal | 123-72-8 | 27.29 | 44 | 57, 72, 39 | (log, steady) | ↓ (log, steady) | ||
| 13 | Butanal, 3-methyl- | Isovaleraldehyde | 590-86-3 | 36.58 | 44 | 58, 71, 86 | ↓ (steady) | ↓ (steady) | ||
| 14 | 2-Butenal, 3-methyl- | 3-methylcrotonaldehyde | 107-86-8 | 47.04 | 84 | 83, 55, 39 | ↓ (steady) | ↓ (steady) | ||
| 15 | Benzoic aldehyde | Benzaldehyde | 100-52-7 | 64.48 | 106 | 105, 77, 51 | ↓ (log, steady) | ↓ (steady) | ↓ (log, steady) | |
| Acids | 16 | Acetic acid | 64-19-7 | 32.94 | 60 | 45, 43, 42 | ↑ (log) | ↑ (steady) | ↑ (log) | |
| 17 | Hexanoic acid | Caproic acid | 142-62-1 | 59.10 | 60 | 73, 87, 41 | ↑ (steady) | ↑ (steady) | ↑ (steady) | |
| Esters | 18 | Formic acid, ethyl ester | Ethyl formate | 109-94-4 | 17.51 | 74 | 45, 31, 43 | ↑ (log) | ↑ (log, steady) | ↑ (log) |
| 19 | Acetic acid, methyl ester | Methyl acetate | 79-20-9 | 18.19 | 43 | 74, 59 | ↑ (steady) | |||
| 20 | Acetic acid, ethyl ester | Ethyl Acetate | 141-78-6 | 29.47 | 43 | 70, 61, 45 | ↑ (log, steady) | ↑ (steady) | ↑ (log, steady) | |
| 21 | Acetic Acid, propyl ester | n-Propyl acetate | 109-60-4 | 42.09 | 61 | 73, 43, 27 | ↑ (steady) | |||
| 22 | Acetic acid, 2-methylpropyl ester | Isobutyl acetate | 110-19-0 | 50.85 | 73 | 56, 43, 86 | ↑ (steady) | ↑ (steady) | ||
| 23 | Acetic acid, butyl ester | n-Butyl acetate | 123-86-4 | 53.09 | 43 | 56, 73, 61 | ↑ (steady) | |||
| 24 | Ethyl, 2-methylbutyrate | 7452-79-1 | 57.63 | 57 | 102, 85, 74 | ↑ (steady) | ↑ (steady) | ↑ (log, steady) | ||
| 25 | Ethyl, 3-methylbutyrate | Ethyl isovalerate | 108-64-5 | 58.20 | 88 | 85, 60, 57 | ↑ (steady) | ↑ (steady) | ↑ (steady) | |
| 26 | 1-Butanol, 3-methyl-, acetate | Isoamyl acetate | 123-92-2 | 59.33 | 70 | 61, 55 | ↑ (log, steady) | ↑ (steady) | ↑ (log, steady) | |
| Ketones | 27 | Propanone | Acetone | 67-64-1 | 15.08 | 43 | 58, 57, 44 | ↓ (steady) | ↑ (log) | |
| 28 | 2-Pentanone | Methyl propyl ketone | 107-87-9 | 39.89 | 86 | 43, 57, 71 | ↑ (log, steady) | ↑ (log) | ↑ (log, steady) | |
| 29 | 4-heptanone | Dipropyl ketone | 123-19-3 | 58.60 | 114 | 71, 43, 41 | ↑ (log) | ↑ (log) | ||
| 30 | 2-Heptanone | Amyl methyl ketone | 110-43-0 | 59.87 | 58 | 43, 71, 114 | ↑ (log, steady) | ↑ (log, steady) | ||
| 31 | 3-Octanone | Amyl ethyl ketone | 106-68-3 | 67.09 | 99 | 128, 72, 57 | ↑ (log, steady) | ↑ (steady) | ||
| 32 | 2-Nonanone | Heptyl methyl ketone | 821-55-6 | 72.08 | 58 | 43, 71, 142 | ↑ (log, steady) | ↑ (log, steady) | ||
| VSCs | 33 | Carbon disulfide | CS2 | 75-15-0 | 15.79 | 76 | 44, 78, 38 | ↓ (log) | ↓ (steady) | ↓ (log) |
| 34 | Disulfide, dimethyl | DMDS | 624-92-0 | 42.02 | 94 | 43, 79, 61 | ↓ (steady) | ↓ (log, steady) | ||
| 35 | 2-Acetylthiazole | 24295-03-2 | 64.48 | 99 | 127, 112, 58 | ↓ (log, steady) | ↓ (steady) | |||
| VOCs identified with Mass Spectra Library Match (not confirmed with standards) | ||||||||||
| Alcohols | 36 | 2-Butanol | sec-Butanol | 78-92-2 | 31.31 | 45 | 59, 41 | ↑ (log, steady) | ↑ (log, steady) | |
| 37 | 1-Butanol, 2-methyl- | sec-butylcarbinol | 137-32-6 | 46.97 | 57 | 56, 29, 41 | ↑ (log, steady) | ↑ (log, steady) | ||
| Aldehydes | 38 | Pentanal | Valeraldehyde | 110-62-3 | 40.13 | 44 | 58, 86, 41 | ↓ (steady) | ↓ (steady) | |
| 39 | 2-Butenal, 2-methyl- | 2-Methylcrotonaldehyde | 1115-11-3 | 43.27 | 84 | 55, 39, 41 | ↓ (log, steady) | ↓ (steady) | ↓ (log, steady) | |
| 40 | 2-Furaldehyde | Furfural | 98-01-1 | 49.39 | 96 | 95, 67, 39 | ↓ (steady) | ↓ (steady) | ↓ (steady) | |
| 41 | Hexanal | Caproaldehyde | 66-25-1 | 52.23 | 56 | 72, 82, 44 | ↓ (log, steady) | |||
| Acid | 42 | Propionic acid, 2-methyl- | isobutyric acid | 79-31-2 | 52.75 | 43 | 73, 41, 27 | ↑ (log) | ↑ (steady) | ↑ (steady) |
| Esters | 43 | Formic acid, butyl ester | n-Butyl formate | 592-84-7 | 35.10 | 56 | 31, 41, 73 | ↑ (steady) | ||
| 44 | Butanoic acid, ethyl ester | Ethyl n-butyrate | 105-54-4 | 52.23 | 71 | 88, 43, 60 | ↑ (log) | ↑ (steady) | ↑ (log) | |
| 45 | Pentanoic acid, 3-methylbutyl ester | Isoamyl valerate | 2050-09-1 | 73.41 | 85 | 70, 55, 43 | ↑ (steady) | ↑ (steady) | ↑ (steady) | |
| Ketones | 46 | 2-Butanone | Ethyl methyl ketone | 78-93-3 | 27.66 | 72 | 43, 57, 42 | ↑ (log) | ↑ (log) | ↑ (log) |
| 47 | 2,3-Pentanedione | Acetylpropionyl | 600-14-6 | 41.10 | 29 | 43, 57, 100 | ↓ (log, steady) | ↑ (log)↓ (steady) | ↓ (log, steady) | |
| 48 | 2-Butanone, 3-hydroxy | Acetoin | 513-86-0 | 43.12 | 45 | 43, 88 | ↑ (steady) | |||
| 49 | 2-Pentanone, 4-methyl- | Isobutyl methyl ketone | 108-40-1 | 48.45 | 43 | 58, 85, 100 | ↑ (log) | |||
| 50 | 3-Hexanone, 2-methyl- | Isopropyl propyl ketone | 7379-12-6 | 58.8 | 71 | 114, 43, 41 | ↑ (log) | ↑ (log) | ||
| Hyrocarbons | 51 | Propane, 2-methyl- | Isobutane | 75-28-5 | 9.91 | 43 | 41, 27, 57 | ↑ (log, steady) | ↑ (log, steady) | |
| 52 | 1-Propene, 2-methyl- | Isobutene | 115-11-7 | 10.36 | 41 | 56, 39, 27 | ↓ (log) | |||
| 53 | 1-Butene | 106-98-9 | 11.39 | 41 | 56, 39, 27 | ↓ (log)↑ (steady) | ↓ (log)↑ (steady) | |||
| 54 | 2-Pentene | 2-Pentene | 109-68-2 | 19.63 | 55 | 70, 53, 56 | ↑ (steady) | |||
| 55 | 2-Butene, 2-methyl- | Amylene | 513-35-9 | 21.64 | 55 | 70, 41, 39 | ↑ (steady) | |||
| 56 | 1-Pentene, 3-methyl- | 3-methyl-1-pentene | 760-20-3 | 43.63 | 55 | 69, 84, 41 | ↑ (log) | |||
| VSCs | 57 | Sulfur dioxide | Sulfur dioxide | 2025884 | 7.90 | 64 | 48 | ↑ (steady) | ||
| 58 | Methanethiol | Mercaptomethane | 74-93-1 | 8.87 | 47 | 48, 45 | ↓ (steady) | |||
| 59 | 1-Mercapto-2-propanone | Mercaptoacetone | 24653-75-6 | 36.12 | 90 | 43, 45, 47 | ↑ (steady) | ↓ (log)↑ (steady) | ↑ (steady) | |
| VNCs | 60 | Methyl cyanide | Acetonitrile | 75-05-8 | 12.45 | 41 | 40, 39, 38 | ↓ (log) | ↓ (steady) | ↓ (log) |
| 61 | Propanenitrile, 2-methyl- | Isobutyronitrile | 78-82-0 | 30.53 | 42 | 68, 54, 41 | ↑ (log) | |||
| 62 | Butanenitrile, 3-methyl | Isoamylnitrile | 625-28-5 | 44.52 | 43 | 41, 68, 27 | ↓ (steady) | ↓ (steady) | ↓ (steady) | |
| 63 | Acetamide, N,N-dimethyl | DMA | 127-19-5 | 53.78 | 87 | 72, 44, 43 | ↑ (log) | ↑ (steady) | ↑ (log) | |
| 64 | Pyrazine, 2,6-dimethyl- | 108-50-9 | 57.61 | 42 | 108, 40, 67 | ↑ (log, steady) | ||||
| Others | 65 | Furan | Furan | 110-00-9 | 14.20 | 68 | 39, 38, 40 | ↓ (log) | ↓ (steady) | ↓ (log) |
| 66 | Propane, 2-methoxy-2-methyl- | tert-Butyl methyl ether | 1634-04-4 | 28.97 | 73 | 57, 41, 29 | ↑ (log) | ↑ (log) | ||
| 67 | Toluene | 108-88-3 | 47.44 | 91 | 92, 65, 39 | ↓ (steady) | ||||
| 68 | Oxazole, 2,4,5-trimethyl | Trimethyloxazole | 20662-84-4 | 53.50 | 111 | 68, 55, 82 | ↓ (steady) | ↑ (log) | ↓ (steady) | |
Table 2.
List of compounds identified in the BAL specimens isolated from VAP patients. Compounds identified based on only Mass Spectra Library match (NIST 2018), i.e., without confirmation of retention time of pure standard, are given in italics and labeled with an asterisk. Mean peak areas of respective substances within a given sample type are given along with the p-values for statistical comparison of VOC abundances, whereby the ANOVA concerns a non-parametric test to compare all three sample types together. The remaining p-values concern the T-test comparing only two groups of interest (significant p < 0.05 are given in blue bold font). CAS is a substance-unique number in Chemical Abstract Service, tR is the retention time [min] of a substance in a given GC program, Target Ion is an extracted ion used for integration of a chromatographic peak area, Reference Ions were used to additionally confirm substance identification during integration, CAL stands for BAL specimens with Candida albicans, SAU for BAL with Staphylococcus aureus, and MIX denotes BAL specimens with both pathogens present. Compounds are arranged according to the increasing p-value of the ANOVA test.
Table 2.
List of compounds identified in the BAL specimens isolated from VAP patients. Compounds identified based on only Mass Spectra Library match (NIST 2018), i.e., without confirmation of retention time of pure standard, are given in italics and labeled with an asterisk. Mean peak areas of respective substances within a given sample type are given along with the p-values for statistical comparison of VOC abundances, whereby the ANOVA concerns a non-parametric test to compare all three sample types together. The remaining p-values concern the T-test comparing only two groups of interest (significant p < 0.05 are given in blue bold font). CAS is a substance-unique number in Chemical Abstract Service, tR is the retention time [min] of a substance in a given GC program, Target Ion is an extracted ion used for integration of a chromatographic peak area, Reference Ions were used to additionally confirm substance identification during integration, CAL stands for BAL specimens with Candida albicans, SAU for BAL with Staphylococcus aureus, and MIX denotes BAL specimens with both pathogens present. Compounds are arranged according to the increasing p-value of the ANOVA test.
| Compound Name | CAS | tR [min] | Target Ion [m/z] | Reference Ion [m/z] | mean PA (MIX) | mean PA (SAU) | mean PA (CAL) | p-value (ANOVA) | p-Value (SAU-CAL) | p-Value (SAU-MIX) | p-Value (CAL-MIX) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3-Methylpyrrole | 616-43-4 | 54.360 | 80 | 44, 43, 42 | 1,85E+05 | 1,94E+05 | 1,05E+05 | 0.016774 | 0.047619 | 1 | 0.0095238 |
| n-Decane | 124-18-5 | 71.400 | 57 | 43, 71, 142 | 5,38E+04 | 1,73E+05 | 2,21E+04 | 0.017276 | 0.01621 | 0.049746 | 0.35857 |
| 6-Methyl-5-hepten-2-one | 110-93-0 | 69.140 | 108 | 69, 55, 93 | 3,70E+04 | 8,59E+04 | 2,35E+04 | 0.024258 | 0.027532 | 0.057143 | 0.23953 |
| Octanal | 124-13-0 | 70.320 | 84 | 69, 56, 100 | 6,07E+04 | 7,75E+04 | 5,10E+04 | 0.024507 | 0.02381 | 0.057143 | 0.25714 |
| Pyrrole | 109-97-7 | 44.650 | 67 | 39, 41, 40 | 1,50E+05 | 1,45E+05 | 9,99E+04 | 0.024507 | 0.095238 | 1 | 0.0095238 |
| Decanal | 112-31-2 | 76.010 | 57 | 43, 55, 82 | 3,16E+05 | 4,15E+05 | 2,42E+05 | 0.027504 | 0.02381 | 0.11429 | 0.17143 |
| 2-Cyclopentene-1-one | 930-30-3 | 53.085 | 82 | 39, 53, 54 | 5,48E+04 | 5,92E+04 | 3,03E+04 | 0.029498 | 0.051822 | 0.4 | 0.042197 |
| * 4-Nonene | 10405-85-3 | 66.105 | 126 | 69, 56, 41 | 2,09E+04 | 3,49E+04 | 2,01E+04 | 0.038036 | 0.02381 | 0.057143 | 0.7619 |
| n-Nonane | 111-84-2 | 66.600 | 57 | 43, 85, 99 | 6,43E+05 | 8,13E+05 | 5,00E+05 | 0.044118 | 0.02381 | 0.4 | 0.17143 |
| 2,3-Butanedione | 431-03-8 | 31.663 | 86 | 43, 42, 41 | 1,44E+06 | 1,24E+05 | 4,86E+04 | 0.050754 | 0.095238 | 0.85714 | 0.038095 |
| 2-Ethyl-1-Hexanol | 104-76-7 | 71.325 | 57 | 70, 83, 41 | 2,36E+06 | 1,20E+06 | 4,58E+06 | 0.067353 | 0.095238 | 0.22857 | 0.11429 |
| * Diethyl Ether | 60-29-7 | 24.505 | 74 | 45, 59, 41 | 5,61E+03 | 0,00E+00 | 4,63E+03 | 0.068751 | 0.025601 | 0.11867 | 0.91429 |
| Carbon Disulfide | 75-15-0 | 20.163 | 76 | 44, 78, 38 | 1,95E+05 | 2,19E+05 | 1,25E+05 | 0.073341 | 0.095238 | 1 | 0.066667 |
| 3-MethylButanal | 590-86-3 | 42.417 | 58 | 71, 41, 86 | 4,82E+06 | 4,98E+05 | 1,23E+05 | 0.080522 | 0.02381 | 0.4 | 0.47619 |
| n-Octane | 111-65-9 | 59.243 | 85 | 114, 43, 57 | 2,56E+05 | 3,24E+05 | 2,28E+05 | 0.11477 | 0.095238 | 0.11429 | 0.60952 |
| 3-Heptanone | 106-35-4 | 62.400 | 57 | 85, 72, 114 | 1,37E+05 | 5,25E+04 | 7,91E+05 | 0.11894 | 0.047619 | 0.4 | 0.47619 |
| 2-Butanone | 78-93-3 | 31.539 | 72 | 43, 57, 42 | 2,79E+05 | 4,46E+05 | 2,41E+06 | 0.11927 | 0.54762 | 0.62857 | 0.038095 |
| Acetic Acid | 64-19-7 | 36.000 | 60 | 45, 43, 42 | 4,66E+06 | 2,57E+05 | 2,69E+05 | 0.13148 | 1 | 0.22857 | 0.066667 |
| Cyclohexanone | 108-94-1 | 60.235 | 98 | 69, 42, 83 | 1,51E+06 | 6,74E+05 | 1,77E+06 | 0.13608 | 0.047619 | 0.4 | 0.60952 |
| 2-Propanol | 67-63-0 | 22.209 | 45 | 43, 41, 59 | 1,42E+07 | 1,40E+08 | 8,60E+07 | 0.13834 | 0.54762 | 0.11429 | 0.17143 |
| Acrolein | 107-02-8 | 17.158 | 56 | 55, 53, 37 | 4,63E+04 | 7,46E+04 | 5,83E+04 | 0.14818 | 0.54762 | 0.057143 | 0.35238 |
| Nonanal | 124-19-6 | 73.300 | 98 | 70, 43, 56 | 1,19E+05 | 1,66E+05 | 1,06E+05 | 0.15147 | 0.095238 | 0.22857 | 0.60952 |
| * 2-Methyl-2-butanol | 75-85-4 | 43.365 | 73 | 59, 55, 43 | 1,72E+05 | 3,00E+05 | 7,65E+05 | 0.16668 | 0.2619 | 0.85445 | 0.10874 |
| * 2,5-Dimethyl Pyrazine | 123-32-0 | 61.766 | 108 | 42, 81, 39 | 5,06E+04 | 2,49E+05 | 5,10E+04 | 0.1714 | 0.16667 | 0.11429 | 0.91429 |
| * Tetrahydrofuran | 109-99-9 | 30.900 | 71 | 72, 42, 41 | 3,67E+04 | 2,00E+04 | 5,16E+04 | 0.17282 | 0.095238 | 0.62857 | 0.35238 |
| * 1-Octene | 111-66-0 | 58.528 | 55 | 70, 41, 83 | 2,76E+05 | 3,66E+05 | 2,45E+05 | 0.17282 | 0.16667 | 0.11429 | 0.7619 |
| Furfural | 98-01-1 | 53.615 | 96 | 95, 67, 39 | 7,93E+04 | 9,43E+04 | 6,31E+04 | 0.17282 | 0.16667 | 0.4 | 0.25714 |
| Ethyl Acetate | 141-78-6 | 33.382 | 43 | 70, 61, 45 | 8,35E+04 | 1,10E+06 | 5,10E+05 | 0.17377 | 0.71429 | 0.057143 | 0.35238 |
| Pyridine | 110-86-1 | 46.200 | 79 | 52, 51, 50 | 9,96E+04 | 1,25E+05 | 3,18E+05 | 0.17764 | 0.2619 | 0.85714 | 0.11429 |
| 1-Propanol | 71-23-8 | 26.439 | 59 | 42, 60, 41 | 0,00E+00 | 1,26E+06 | 1,22E+07 | 0.20664 | 1 | 0.12274 | 0.1491 |
| Hexanal | 66-25-1 | 55.700 | 56 | 72, 82, 44 | 1,73E+05 | 1,73E+05 | 1,03E+05 | 0.22684 | 0.2619 | 0.85714 | 0.17143 |
| * 3-Penten-2-one | 3102-33-8 | 48.521 | 69 | 84, 55, 41 | 3,34E+03 | 1,77E+04 | 4,96E+04 | 0.22984 | 0.89553 | 0.24189 | 0.13933 |
| Acetaldehyde | 75-07-0 | 9.644 | 43 | 44, 42, 41 | 1,20E+06 | 1,42E+06 | 1,05E+06 | 0.23509 | 0.16667 | 0.4 | 0.47619 |
| * Phenol | 108-95-2 | 67.000 | 94 | 66, 65, 39 | 5,05E+05 | 3,71E+05 | 5,85E+05 | 0.23638 | 0.16667 | 0.22857 | 0.91429 |
| n-Undecane | 1120-21-4 | 73.800 | 57 | 43, 71, 85 | 3,60E+05 | 5,11E+05 | 3,49E+05 | 0.23638 | 0.2619 | 0.11429 | 1 |
| * 2-Heptene | 14686-13-6 | 49.945 | 69 | 98, 56, 41 | 3,29E+04 | 4,55E+04 | 5,41E+04 | 0.25555 | 0.71429 | 0.85714 | 0.087118 |
| * Methanethiol | 74-93-1 | 10.900 | 47 | 48, 45, 46 | 3,04E+05 | 2,60E+05 | 2,18E+05 | 0.27494 | 0.38095 | 0.85714 | 0.17143 |
| Methyl Methacrylate | 4655-34-9 | 45.175 | 69 | 100, 41, 39 | 1,31E+04 | 6,75E+03 | 1,79E+04 | 0.28948 | 0.89685 | 0.057143 | 0.45417 |
| * Propene | 115-07-1 | 7.250 | 41 | 39, 42, 44 | 2,77E+05 | 4,90E+05 | 3,92E+05 | 0.29287 | 0.54762 | 0.22857 | 0.35238 |
| Benzaldehyde | 100-52-7 | 65.584 | 106 | 105, 77, 51 | 3,57E+05 | 3,48E+05 | 2,62E+05 | 0.31369 | 0.16667 | 0.4 | 0.91429 |
| Acetone | 67-64-1 | 18.266 | 58 | 57, 44, 117 | 8,32E+06 | 2,18E+07 | 8,78E+06 | 0.31369 | 0.2619 | 0.22857 | 1 |
| Butyrolactone | 96-48-0 | 58.200 | 86 | 42, 56, 41 | 1,39E+04 | 2,60E+04 | 2,38E+04 | 0.32869 | 0.54762 | 0.4 | 0.25714 |
| 2-Methylfuran | 534-22-5 | 30.700 | 82 | 53, 81, 39 | 8,32E+04 | 5,96E+04 | 5,76E+04 | 0.35693 | 0.38095 | 0.85714 | 0.25714 |
| 2-Butenal | 4170-30-3 | 35.863 | 70 | 41, 39, 69 | 2,80E+04 | 4,68E+04 | 3,29E+04 | 0.3717 | 0.38095 | 0.22857 | 0.83066 |
| Butanal | 123-72-8 | 31.105 | 57 | 72, 44, 39 | 3,47E+04 | 2,89E+04 | 3,45E+04 | 0.37399 | 0.54762 | 0.62857 | 0.25714 |
| * 1-Methoxy-2-propanol | 107-98-2 | 43.910 | 45 | 75, 47, 57 | 1,11E+06 | 3,78E+05 | 0,00E+00 | 0.39161 | 0.23859 | 1 | 0.30743 |
| Pentanal | 110-62-3 | 45.598 | 44 | 58, 41, 57 | 6,08E+04 | 7,82E+04 | 4,56E+04 | 0.40058 | 0.2619 | 0.85714 | 0.47619 |
| n-Heptane | 142-82-5 | 50.100 | 71 | 57, 43, 100 | 9,93E+04 | 1,75E+05 | 1,46E+05 | 0.42671 | 0.71429 | 0.4 | 0.35238 |
| * 2-Propenenitrile | 107-13-1 | 21.062 | 53 | 52, 51, 50 | 2,53E+04 | 3,52E+04 | 3,11E+04 | 0.4533 | 0.90476 | 0.4 | 0.35238 |
| 2-(n)-PentylFuran | 3777-69-3 | 69.325 | 81 | 82, 53, 138 | 1,34E+05 | 1,34E+05 | 1,07E+05 | 0.45831 | 0.2619 | 0.85714 | 0.60952 |
| * N,N-Dimethyl-formamide | 68-12-2 | 48.577 | 73 | 44, 58 | 1,80E+05 | 6,15E+05 | 1,13E+06 | 0.46937 | 0.69364 | 0.47553 | 0.33142 |
| Furan | 110-00-9 | 17.916 | 68 | 39, 38, 40 | 3,82E+04 | 3,65E+04 | 2,88E+04 | 0.51719 | 0.38095 | 1 | 0.47619 |
| * iso-Butane | 75-28-5 | 12.000 | 43 | 41, 42, 39 | 1,78E+05 | 4,96E+04 | 3,92E+04 | 0.55244 | 0.54762 | 0.4 | 0.60952 |
| p-Xylene | 106-42-3 | 60.939 | 91 | 106, 105, 77 | 3,51E+05 | 8,26E+05 | 4,19E+05 | 0.57727 | 0.54762 | 0.4 | 0.91429 |
| 2-Methylpropanal | 78-84-2 | 28.588 | 72 | 41, 43, 39 | 8,41E+05 | 1,12E+05 | 6,02E+04 | 0.58204 | 0.38095 | 0.62857 | 0.91429 |
| 3-Methyl-1-Butanol | 123-51-3 | 50.395 | 55 | 70, 42, 41 | 7,55E+06 | 0,00E+00 | 1,18E+06 | 0.58778 | 0.37676 | 0.5637 | 1 |
| Isoprene | 78-79-5 | 24.927 | 67 | 68, 53, 39 | 1,09E+05 | 1,96E+05 | 1,11E+05 | 0.59498 | 0.38095 | 0.62857 | 0.91429 |
| * 1,3-Butadiene | 106-99-0 | 12.987 | 54 | 39, 53, 50 | 3,43E+04 | 4,19E+04 | 3,23E+04 | 0.59661 | 0.54762 | 0.4 | 0.91429 |
| * 2-Methyl-1,3-dioxolane | 497-26-7 | 35.455 | 73 | 58, 45, 87 | 3,29E+04 | 3,33E+04 | 3,93E+04 | 0.60653 | 0.54762 | 1 | 0.47619 |
| * 1-Butene | 106-98-9 | 13.659 | 56 | 55, 53, 50 | 2,33E+04 | 1,63E+04 | 1,37E+04 | 0.61831 | 0.38095 | 0.85714 | 0.7619 |
| 1-Heptene | 592-76-7 | 49.100 | 69 | 98, 70, 56 | 1,10E+05 | 8,79E+04 | 1,18E+05 | 0.61831 | 0.38095 | 0.85714 | 0.7619 |
| * Trichloromethane | 67-66-3 | 31.000 | 83 | 85, 47 | 4,80E+05 | 9,13E+04 | 5,50E+04 | 0.64079 | 1 | 0.85714 | 0.35238 |
| Propanal | 123-38-6 | 18.263 | 57 | 55, 39, 37 | 1,19E+05 | 2,72E+05 | 1,95E+05 | 0.66227 | 0.38095 | 0.85714 | 0.91429 |
| Benzene | 71-43-2 | 38.137 | 78 | 77, 51, 52 | 4,85E+05 | 5,44E+05 | 5,08E+05 | 0.66227 | 0.71429 | 0.4 | 0.91429 |
| (E)-2-Methyl-2-Butenal | 1115-11-3 | 47.964 | 84 | 55, 39, 41 | 6,55E+04 | 2,63E+04 | 9,62E+03 | 0.67383 | 0.51148 | 1 | 0.58949 |
| (E)-1,3-Pentadiene | 2004-70-8 | 27.082 | 67 | 68, 53, 39 | 2,63E+04 | 3,11E+04 | 2,48E+04 | 0.69013 | 0.54762 | 1 | 0.60952 |
| * (E)-2-Butene | 624-64-6 | 13.659 | 56 | 55, 53, 50 | 2,88E+04 | 2,19E+04 | 1,85E+04 | 0.69013 | 0.54762 | 0.62857 | 0.91429 |
| * n-Butane | 106-97-8 | 13.378 | 58 | 39, 57 | 1,32E+04 | 1,14E+04 | 1,08E+04 | 0.73716 | 0.90476 | 0.62857 | 0.60952 |
| * N-Methylpyrrole | 96-54-8 | 45.960 | 81 | 80, 39, 53 | 2,76E+04 | 2,54E+04 | 2,29E+04 | 0.76397 | 0.71429 | 0.85714 | 0.60952 |
| 1-Hexene | 592-41-6 | 36.000 | 41 | 84, 69, 56 | 1,31E+05 | 1,42E+05 | 1,34E+05 | 0.77453 | 0.71429 | 0.62857 | 0.91429 |
| * 1,2-Propanediol | 57-55-6 | 47.175 | 45 | 43, 61, 57 | 1,30E+07 | 1,48E+08 | 4,54E+07 | 0.78076 | 0.59588 | 0.8255 | 1 |
| Ethylbenzene | 100-41-4 | 60.000 | 91 | 106, 65, 77 | 3,90E+05 | 4,37E+05 | 3,98E+05 | 0.79174 | 0.71429 | 0.62857 | 0.91429 |
| 2-Pentanon | 107-87-9 | 44.911 | 43 | 86, 71, 58 | 1,55E+05 | 4,72E+05 | 1,48E+05 | 0.79174 | 0.71429 | 0.62857 | 0.91429 |
| (Z)-2-Pentene | 627-20-3 | 25.271 | 55 | 70, 71, 56 | 2,35E+04 | 2,31E+04 | 3,96E+04 | 0.79174 | 0.90476 | 0.4 | 0.7619 |
| Acetophenone | 98-86-2 | 71.213 | 105 | 77, 120, 51 | 4,52E+05 | 5,19E+05 | 6,43E+05 | 0.80998 | 0.79542 | 1 | 0.60952 |
| Ethanol | 64-17-5 | 14.000 | 45 | 46, 43 | 1,10E+07 | 5,69E+05 | 3,59E+07 | 0.82053 | 1 | 0.62857 | 0.7619 |
| Acetonitrile | 75-05-8 | 14.852 | 41 | 40, 39, 38 | 4,95E+05 | 5,01E+05 | 6,66E+05 | 0.82505 | 1 | 0.85714 | 0.60952 |
| Methacrolein | 78-85-3 | 28.261 | 70 | 41, 39, 42 | 6,86E+04 | 5,54E+04 | 5,92E+04 | 0.84107 | 1 | 0.4 | 1 |
| 2-Ethylacrolein | 922-63-4 | 41.552 | 55 | 84, 56, 39 | 6,40E+04 | 2,98E+04 | 3,16E+04 | 0.90335 | 0.90476 | 0.85714 | 0.7619 |
| n-Dodecane | 112-40-3 | 76.800 | 57 | 71, 43, 85 | 2,34E+05 | 2,04E+05 | 2,04E+05 | 0.91082 | 0.90476 | 1 | 0.7619 |
| n-Hexane | 110-54-3 | 37.800 | 86 | 57, 43, 41 | 9,79E+03 | 1,33E+04 | 9,36E+03 | 0.91082 | 0.90476 | 1 | 0.7619 |
| n-Pentane | 109-66-0 | 25.545 | 72 | 57, 43, 41 | 1,17E+04 | 1,07E+04 | 9,37E+03 | 0.92517 | 1 | 0.62857 | 1 |
| (E)-2-Pentene | 646-04-8 | 25.680 | 55 | 70, 53, 56 | 1,02E+05 | 1,14E+05 | 1,13E+05 | 0.92596 | 0.90476 | 0.85714 | 0.91429 |
| 1-Dodecene | 112-41-4 | 76.440 | 55 | 69, 83, 97 | 6,21E+04 | 4,61E+04 | 6,73E+04 | 0.93261 | 0.89274 | 0.85445 | 0.91406 |
| Heptanal | 111-71-7 | 63.800 | 81 | 70, 43, 55 | 2,40E+04 | 2,24E+04 | 2,20E+04 | 0.93619 | 0.71429 | 1 | 1 |
| * 2-Methyl-1-propene | 115-11-7 | 12.629 | 56 | 41, 39, 55 | 3,55E+05 | 2,94E+05 | 3,01E+05 | 0.93619 | 0.90476 | 0.85714 | 0.91429 |
| Dimethyl Disulfide | 624-92-0 | 47.136 | 94 | 43, 79, 61 | 7,54E+03 | 6,61E+03 | 6,14E+03 | 0.93816 | 0.89274 | 0.84536 | 1 |
| 4-Heptanone | 123-19-3 | 62.200 | 71 | 43, 114, 41 | 3,16E+04 | 3,34E+03 | 3,79E+04 | 0.94038 | 0.87809 | 1 | 0.89565 |
| 2-Methylpyrazine | 109-08-0 | 54.046 | 94 | 67, 53, 40 | 2,46E+04 | 1,83E+04 | 2,35E+04 | 0.94654 | 1 | 1 | 0.7619 |
| 3-Methyl-1-Butene | 563-45-1 | 24.148 | 55 | 70, 41, 42 | 2,01E+05 | 2,14E+05 | 2,43E+05 | 0.94654 | 0.90476 | 0.85714 | 0.91429 |
| * Isobutyronitrile | 78-82-0 | 34.940 | 68 | 42, 54, 52 | 4,14E+04 | 3,94E+04 | 4,23E+04 | 0.95175 | 1 | 0.85714 | 0.91429 |
| * (Z)-2-Butene | 590-18-1 | 12.766 | 41 | 56, 39, 55 | 2,01E+05 | 1,94E+05 | 1,68E+05 | 0.95175 | 0.90476 | 0.62857 | 1 |
| 2-Heptanone | 110-43-0 | 63.000 | 58 | 43, 71, 114 | 6,20E+04 | 5,34E+04 | 6,61E+04 | 0.95963 | 0.90476 | 1 | 0.91429 |
| Toluene | 108-88-3 | 51.942 | 91 | 92, 65, 39 | 1,18E+06 | 1,20E+06 | 1,06E+06 | 1 | 0.90476 | 0.85714 | 0.91429 |
| D-Limoinene | 5989-27-5 | 71.250 | 68 | 93, 67, 136 | 1,07E+05 | 5,58E+05 | 1,87E+05 | 1 | 1 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



