
Submitted:
15 April 2024
Posted:
17 April 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The replication of species A rotaviruses (RVAs) and other RNA viruses involves recruitment of and inter-action with the cellular organelles lipid droplets (LDs), both physically and functionally. Inhibition of en-zymes involved in the cellular fatty acid biosynthesis pathway or of cellular lipases that degrade LDs re-duces the functions of ‘viral factories’ (viroplasms for rotaviruses or replication compartments of other RNA viruses) and decreases the production of infectious progeny virus. Similarly, disturbance of the cellular lipid homeostasis in various ways blocks the replication of flaviviruses (hepatitis C viruses, Zikavirus, others), HIV-1, SARS-CoV-2, picornaviruses, noroviruses, influenza viruses and other negative-strand RNA viruses.
Keywords:
Lipid metabolism
; RNA virus replication
; Rotavirus
; hepatitis C virus
; flaviviruses
; picornaviruses
; no-roviruses
; HIV-1
; SARS-CoV-2
; influenza viruses
Introduction
While the replication cycles of many RNA and DNA viruses have been well researched, it has been recognized only relatively recently that various cellular lipids are involved in many of those steps (Herker, 2024; Qu et al, 2023; Farias et al, 2023; Madsen and Rossman, 2022; Li et al, 2022; Roingeard et al, 2022; Zhao et al, 2022; Theken et al, 2021; Husby and Stahelin, 2021; Crawford and Desselberger, 2016; Heaton and Randall, 2011) and also in the regulation of the host’s innate immune responses (Farias et al, 2023; Theken et al, 2021). Cellular lipids, summarizingly termed the ‘liposome’, are classified as membrane lipids (phospholipids, sphingolipids, glycolipids), cholesterol, steroids, triacylglycerol (TAG), fatty acids, and eicosanoids (Fahy et al, 2011). Viruses interact with host cell lipids in various ways, affecting their biosynthesis and metabolism, and gaining energy for their own replication (Heaton and Randall, 2011). In the following, the significance of cellular lipids for the replication of rotaviruses will be reviewed in some detail. Analogous data have been acquired for many other RNA viruses. The fact that disturbance of the cellular lipid metabolism decreases the yield of infectious viral progeny for many viruses, makes the search for lipid-disturbing compounds an attractive aim for the development of broadly active candidate antivirals (Cesar-Silva et al, 2023; Qu et al, 2023; Farias et al, 2023; Li et al, 2022; Roingeard et al, 2022; Theken et al, 2021).
Rotaviruses
Species A rotaviruses (RVA) are a major cause of severe gastroenteritis in infants and young children worldwide and in the young of many mammalian and avian host species (Crawford et al, 2017). RVA-associated disease leads to the deaths of approximately 130.000 children under the age of 5 y annually worldwide, mainly in low-income countries (Troeger et al, 2019). Two RVA vaccines were licensed since 2006 and are now used in expanded programs of immunization (EPIs) in over 100 countries (Bergman et al, 2021). In addition, alternative RV vaccines were licensed in India, Vietnam and China (Bergman et al, 2021). RVA EPIs have significantly decreased RVA-associated disease, although at different levels in different parts of the world (Desselberger, 2017; Parker et al, 2018). The molecular biology of RVA replication has been well studied (Crawford et al, 2017). During replication, ‘viroplasms’ form, which are intracytoplasmic inclusion bodies utilized for early RVA morphogenesis and viral RNA replication take place; they have been termed ‘viral factories’ and been recognized as essential (Crawford et al, 2017). The rotavirus-encoded components of viroplasms, NSP2 and NSP5, are subject to liquid-liquid phase separation (Geiger et al, 2021; Papa et al, 2020). In 2010 rotavirus viroplasms (containing VP1, VP2, VP3, VP6, viral RNA segments) were discovered to associate with components of the cellular organelles lipid droplets (LDs) (Cheung et al, 2010) [Figure 1]. LDs of various sizes (0.05-200 um in diameter) are the storage sites of sterol esters and neutral lipids which serve as the major cellular energy source and are in close contact with mitochondria and the endoplasmic reticulum (ER) from [PLINs], diacylglycerol acyl transferases 1 and 2 [DGAT1, DGAT2], Rab18 [transport], and CCT [CTP-phosphocholine cytidylyltransferase, the rate-limiting enzyme of phosphatidylcholine synthesis] (Farese and Walther, 2009). The LDs also contain acyl-CoA cholesterol acyltransferases 1 and 2 (ACAT1, ACAT2) which catalyse cholesterol esterification. A genome-wide, siRNA-based screen identified 550/18000 genes of human macrophages (THP-1) as being involved in modulating lipid storage according to number, size and cellular localization and functions such as proteasome activity, intracellular transport, transcription activity, E3 ligase activity and lipid modifying enzymes (Mejhert et al, 2021). In summary, LDs are central to the regulation of cellular lipid homeostasis (Mejhert et al, 2021). Fluorescence resonance energy transfer (FRET) has shown to occur between perilipin A and NSP2, proving the close spatial proximity of LDs and viroplasms (Cheung et al, 2010).
Iodixanol gradient velocity ultracentrifugation of RV-infected cell extracts resulted in co-sedimentation of viral dsRNA, NSP2 and perilipin A in low-density fractions (1.11-1.15 g/ml). By contrast, rotavirus dual-layered particles (DLPs; of density 1.38 g/ml), spiked into mock-infected lysates, sedimented to the bottom of the gradient (Cheung et al, 2010). Iodixanol gradient fractions of low density, containing peaks of the RV dsRNA genome and LD- and viroplasm-associated proteins, were analysed for 14 different classes of lipids by mass spectrometry and found to contain maximum amounts of lipids as typical components of LDs, confirming the close interaction of LDs with viroplasms (Gaunt et al, 2013b).
The complex formation of viroplasms and LDs has functional consequences. LD formation depends critically on the presence of long chain fatty acids. Their biosynthesis is catalysed by acetyl-CoA cocarboxylase (ACC-1), fatty acid synthase (FASN), long chain acyl-CoA synthetase (HCSL) and various glycerol-acyl transferases (GATs), the latter converting long chain fatty acids into triacyl esters (neutral fats) which are incorporated into the precursors of LDs in the endoplasmic reticulum (ER). The ER-localized enzymes DGAT1 and DGAT2, ACAT1 and ACAT2 synthesize TAGs from fatty acids and sterol esters from cholesterol, respectively. Lipid droplets bud from the lipid bilayer of the ER into the cytoplasm and acquire various lipid droplet-associated proteins (PLINs 1-5 and many others) (Crawford and Desselberger, 2016).
Key inhibitors of these pathways are: TOFA (inhibiting ACC-1), C75 (anti-FASN), triacsin C (anti-ACSL) and A922500 (anti-DGAT-1) (Kim et al, 2012; Crawford and Desselberger, 2016). Application of these inhibitors to RV-infected cells significantly reduce the size and number of viroplasms formed, the production of viral dsRNA and the titers of infectious viral progeny to various extent. Similarly, the treatment of cells with a combination of [isoproterenol + IBMX], which fragments LDs into smaller microdroplets (Marcinkiewicz et al, 2006) also reduces RV progeny production. Representative data for inhibition with TOFA, triacsin C and [Isoproterenol + IBMX) are presented in Table 1 (Cheung et al, 2010; Gaunt et al, 2013a).
Furthermore, TOFA-treated and RV-infected cells were analysed for the production of DLPs and TLPs (purified by CsCl gradient equilibrium ultracentrifugation). In the presence of TOFA the production of DLPs is decreased by a factor of 2, but that of TLPs by a factor of 20, compared to DLP and TLP production in untreated cells. These suggested that inhibition of fatty acid biosynthesis affects not only the recruitment of LDs by viroplasms but is also involved in interfering with the later step of RV maturation (Cheung et al, 2016).
The mechanism of interaction of viroplasms with LDs was explored further. Following cellular infection with a RVA mutant with delayed viroplasm formation (rRV NSP2 S313D, engineered by a RVA-specific plasmid only-based reverse genetics system; Kanai et al, 2017), an early interaction of viroplasm-bound NSP2 with phospho-perilipin 1 (leading to lipolysis; Tansey et al, 2003) was observed (Criglar et al, 2022), thus exploiting the lipid metabolism.
Recently, it was found that DGAT1, required for triacylglycerol and LD biosynthesis, is degraded in RV-infected MA104 cells and in human intestinal enteroids (HIEs) (Liu, Smith et al, 2023). In the uninfected cell, DGAT1 synthesizes TAGs from ER-resident DAGs and cytoplasmic acyl-CoAs, and TAGs are deposited in the ER lipid bilayer. Upon RV infection, NSP2 is expressed which interacts with DGAT1. The viroplasm-DGAT1 complexes are then tagged with ubiquitin and degraded in proteasomes. The loss of DGAT1 induces the budding of TAGs and LD formation by a still not fully understood mechanism (Liu, Smith et al, 2023). Suppression of DGAT1 by rotavirus infection leads to an increase in the number of viroplasms and of the infectivity titer of viral progeny in MA104 cells (Liu, Smith et al, 2023).
Other RNA Viruses
Besides for rotaviruses, the significance of cellular lipid metabolism has been recognized for the replication of many other RNA viruses, and some of the relevant information is summarized in Table 2. Members of the Flaviviridae (hepatitis C virus [HCV], Dengue virus (DENV], GB virus-B, Zika virus and others), Picornaviridae (poliovirus, enterovirus), Caliciviridae (norovirus), Coronaviridae (SARS-CoV-2), Retroviridae (HIV-1), Orthomyxoviridae (influenza virus) and Rhabdoviridae (lyssa virus) families utilize components and pathways of the cellular lipidome in various ways for their replication, such as in the formation of double membrane vesicles in contact with LDs (potentially acting as viral assembly platforms), the stimulation of cellular lipases, and the requirement of cellular lipids and membranes for viral envelope formation.
A kinetic study on lipidome changes in HCV (strain 96/JFH-1)-infected human hepatoma Huh-7 cells using mass spectrometry discovered a dynamic profile, with phosphatidyl phospholipids, triacylglycerol, phosphatidyl choline and ceramide derivatives being enriched early in infection, and free fatty acids, phosphatidylethanolamine peaking at late stages of the replication cycle. This profile could be correlated with different lipidome functions during viral replication (Islam et al, 2023; Qu et al, 2023).
For DENVs it was observed that a LD-associated membrane protein, the acyl transferase AUP1, associates with the viral NS4A protein to be relocalized to autophagosomes where the complexes are degraded; this triggers lipophagy and a decrease in virus production (Zhang et al, 2018).
For coronaviruses (mainly SARS-CoV-2), ceramide plays a central role during virus entry, and inhibitors of acid sphingomyelinase, the rate-limiting enzyme of ceramide biosynthesis, suppresses viral replication. Various drugs inhibiting enzymes of the sphingolipid metabolism, are under investigation as potential antivirals (Cesar-Silva et al, 2023, and text and Table 2 therein). Similarly, various drugs reducing cholesterol biosynthesis and esterification (and LD formation) are being explored (Cesar-Silva et al, 2023; D’Avila et al, 2024) or are already under clinical investigation (Theken et al, 2021, and Table 1 therein).
The double-membrane vesicles (DMVs), in which coronaviruses replicate, have similarities with analogous replication structures of HCV (Roingeard et al, 2021; Tabata et al, 2021).
Lipid rafts, located in proximity of cellular membranes, are rich in cholesterol and are involved in influenza A virus cell entry, particle maturation and budding (Madsen and Rossman, 2023; Li YJ et al, 2022). Influenza viruses and other negative-strand, non-segmented RNA viruses obtain their envelope during the budding step from the host cell’s plasma membrane where cholesterol forms microdomains (rafts) which act as scaffold for viral ‘envelope’ proteins (Husby and Stahelin, 2021; Madsen and Rossman, 2023). Statins which act as inhibitors of cholesterol biosynthesis, may have a role in blocking the replication of influenza viruses and of members of the Mononegavirales (Li et al, 2022).
[The significance of cellular lipid metabolism for DNA viruses has been reviewed by Farias et al, 2022].
Acknowledgements
For rotavirus research, the author gratefully acknowledges the collaboration with Oscar Burrone, Winsome Cheung, Serge Chwetzoff, Nathalie Couroussé, Sue E Crawford, Alessandro Esposito, Eleanor Gaunt, Michael Gill, Clemens F Kaminski, Nandita Keshavan, Andrew M L Lever, James E Richards, Germain Trugnan, Michael JO Wakelam, and Qifeng Zhang.
References
- Baek YB, Kwon HJ, Sharif M, Lim J, Lee IC, Ryu YB, Lee JI, Kim JS, Lee YS, Kim DH, Park SI, Kim DK, Kim JS, Choy HE, Lee S, Choi HS, Osborne TF, Jeon TI, Cho KO. Therapeutic strategy targeting host lipolysis limits infection by SARS-CoV-2 and influenza A virus. Signal Transduct Target Ther. 2022 Oct 17;7(1):367. [CrossRef] [PubMed] [PubMed Central]
- Belov GA, van Kuppeveld FJM. Lipid droplets grease enterovirus replication. Cell Host Microbe. 2019 Aug 14;26(2):149-151. [CrossRef] [PubMed] [PubMed Central]
- Bergman H, Henschke N, Hungerford D, Pitan F, Ndwandwe D, Cunliffe N, Soares-Weiser K. Vaccines for preventing rotavirus diarrhoea: vaccines in use. Cochrane Database Syst Rev. 2021 Nov 17;11(11):CD008521. [CrossRef] [PubMed] [PubMed Central]
- Boulant S, Targett-Adams P, McLauchlan J. Disrupting the association of hepatitis C virus core protein with lipid droplets correlates with a loss in production of infectious virus. J Gen Virol. 2007 Aug;88(Pt 8):2204-2213. [CrossRef] [PubMed]
- Cesar-Silva D, Pereira-Dutra FS, Giannini ALM, Maya-Monteiro CM, de Almeida CJG. Lipid compartments and lipid metabolism as therapeutic targets against coronavirus. Front Immunol. 2023 Dec 1;14:1268854. [CrossRef] [PubMed] [PubMed Central]
- Chatel-Chaix L, Bartenschlager R. Dengue virus- and hepatitis C virus-induced replication and assembly compartments: the enemy inside--caught in the web. J Virol. 2014 Jun;88(11):5907-11. [CrossRef] [PubMed] [PubMed Central]
- Chen Q, Gouilly J, Ferrat YJ, Espino A, Glaziou Q, Cartron G, El Costa H, Al-Daccak R, Jabrane-Ferrat N. Metabolic reprogramming by Zika virus provokes inflammation in human placenta. Nat Commun. 2020 Jun 11;11(1):2967. [CrossRef] [PubMed] [PubMed Central]
- Chen X, Wang S, Gan P, Zhang J, Tong G, Liu S. Comprehensive analysis of lipid metabolism in influenza virus infection. Microb Pathog. 2023 Feb;175:106002. [CrossRef] [PubMed]
- Cheung W, Gaunt E, Lever A, Desselberger U. Rotavirus replication: The role of lipid droplets. In: Viral Gastroenteritis, L Svensson, U Desselberger, HB Greenberg, MK Estes, eds, pp 175-187. Elsevier-Academic Press, Amsterdam, 2016.
- Cheung W, Gill M, Esposito A, Kaminski CF, Courousse N, Chwetzoff S, Trugnan G, Keshavan N, Lever A, Desselberger U. Rotaviruses associate with cellular lipid droplet components to replicate in viroplasms, and compounds disrupting or blocking lipid droplets inhibit viroplasm formation and viral replication. J Virol. 2010 Jul;84(13):6782-98. [CrossRef] [PubMed] [PubMed Central]
- Crawford SE, Desselberger U. Lipid droplets form complexes with viroplasms and are crucial for rotavirus replication. Curr Opin Virol. 2016 Aug;19:11-5. [CrossRef] [PubMed] [PubMed Central]
- Crawford SE, Ramani S, Tate JE, Parashar UD, Svensson L, Hagbom M, Franco MA, Greenberg HB, O'Ryan M, Kang G, Desselberger U, Estes MK. Rotavirus infection. Nat Rev Dis Primers. 2017 Nov 9;3:17083. [CrossRef] [PubMed] [PubMed Central]
- Criglar JM, Estes MK, Crawford SE. Rotavirus-induced lipid droplet biogenesis is critical for virus replication. Front Physiol. 2022 Mar 23;13:836870. [CrossRef] [PubMed] [PubMed Central]
- D'Avila H, Lima CNR, Rampinelli PG, Mateus LCO, Sousa Silva RV, Correa JR, Almeida PE. Lipid metabolism modulation during SARS-CoV-2 infection: A spotlight on extracellular vesicles and therapeutic prospects. Int J Mol Sci. 2024 Jan 4;25(1):640. [CrossRef] [PubMed] [PubMed Central]
- Desselberger U. Differences of rotavirus vaccine effectiveness by country: Likely causes and contributing factors. Pathogens. 2017 Dec 12;6(4):65. [CrossRef] [PubMed] [PubMed Central]
- Dias SSG, Soares VC, Ferreira AC, Sacramento CQ, Fintelman-Rodrigues N, Temerozo JR, Teixeira L, Nunes da Silva MA, Barreto E, Mattos M, de Freitas CS, Azevedo-Quintanilha IG, Manso PPA, Miranda MD, Siqueira MM, Hottz ED, Pão CRR, Bou-Habib DC, Barreto-Vieira DF, Bozza FA, Souza TML, Bozza PT. Lipid droplets fuel SARS-CoV-2 replication and production of inflammatory mediators. PLoS Pathog. 2020 Dec 16;16(12):e1009127. [CrossRef] [PubMed] [PubMed Central]
- Doerflinger SY, Cortese M, Romero-Brey I, Menne Z, Tubiana T, Schenk C, White PA, Bartenschlager R, Bressanelli S, Hansman GS, Lohmann V. Membrane alterations induced by nonstructural proteins of human norovirus. PLoS Pathog. 2017 Oct 27;13(10):e1006705. [CrossRef] [PubMed] [PubMed Central]
- Fahy E, Cotter D, Sud M, Subramaniam S. Lipid classification, structures and tools. Biochim Biophys Acta. 2011 Nov;1811(11):637-47. [CrossRef] [PubMed] [PubMed Central]
- Farese RV Jr, Walther TC. Lipid droplets finally get a little R-E-S-P-E-C-T. Cell. 2009 Nov 25;139(5):855-60. [CrossRef] [PubMed] [PubMed Central]
- Farías MA, Diethelm-Varela B, Kalergis AM, González PA. Interplay between lipid metabolism, lipid droplets and RNA virus replication. Crit Rev Microbiol. 2023 Jun 22:1-25. [CrossRef] [PubMed]
- Farías MA, Diethelm-Varela B, Navarro AJ, Kalergis AM, González PA. Interplay between Lipid Metabolism, Lipid Droplets, and DNA Virus Infections. Cells. 2022 Jul 17;11(14):2224. [CrossRef] [PubMed] [PubMed Central]
- Gaunt ER, Cheung W, Richards JE, Lever A, Desselberger U. Inhibition of rotavirus replication by downregulation of fatty acid synthesis. J Gen Virol. 2013a Jun;94(Pt 6):1310-1317. Epub 2013 Mar 13. Erratum in: J Gen Virol. 2013 Sep;94(Pt 9):2140. [CrossRef] [PubMed]
- Gaunt ER, Zhang Q, Cheung W, Wakelam MJO, Lever AML, Desselberger U. Lipidome analysis of rotavirus-infected cells confirms the close interaction of lipid droplets with viroplasms. J Gen Virol. 2013b Jul;94(Pt 7):1576-1586. [CrossRef] [PubMed] [PubMed Central]
- Geiger F, Acker J, Papa G, Wang X, Arter WE, Saar KL, Erkamp NA, Qi R, Bravo JP, Strauss S, Krainer G, Burrone OR, Jungmann R, Knowles TP, Engelke H, Borodavka A. Liquid-liquid phase separation underpins the formation of replication factories in rotaviruses. EMBO J. 2021 Nov 2;40(21):e107711. [CrossRef] [PubMed] [PubMed Central]
- Heaton NS, Randall G. Multifaceted roles for lipids in viral infection. Trends Microbiol. 2011 Jul;19(7):368-75. [CrossRef] [PubMed] [PubMed Central]
- Herker E. Lipid droplets in virus replication. FEBS Lett. 2024 Feb 13. [CrossRef] [PubMed]
- Hope RG, Murphy DJ, McLauchlan J. The domains required to direct core proteins of hepatitis C virus and GB virus-B to lipid droplets share common features with plant oleosin proteins. J Biol Chem. 2002 Feb 8;277(6):4261-70. [CrossRef] [PubMed]
- Husby ML, Stahelin RV. Negative-sense RNA viruses: An underexplored platform for examining virus-host lipid interactions. Mol Biol Cell. 2021 Oct 1;32(20):pe1. [CrossRef] [PubMed] [PubMed Central]
- Islam KU, Anwar S, Patel AA, Mirdad MT, Mirdad MT, Azmi MI, Ahmad T, Fatima Z, Iqbal J. Global lipidome profiling revealed multifaceted role of lipid species in hepatitis C virus replication, assembly, and host antiviral gesponse. Viruses. 2023 Feb 7;15(2):464. [CrossRef] [PubMed] [PubMed Central]
- Kanai Y, Komoto S, Kawagishi T, Nouda R, Nagasawa N, Onishi M, Matsuura Y, Taniguchi K, Kobayashi T. Entirely plasmid-based reverse genetics system for rotaviruses. Proc Natl Acad Sci U S A. 2017 Feb 28;114(9):2349-2354. [CrossRef] [PubMed] [PubMed Central]
- Kim Y, George D, Prior AM, Prasain K, Hao S, Le DD, Hua DH, Chang KO. Novel triacsin C analogs as potential antivirals against rotavirus infections. Eur J Med Chem. 2012 Apr;50:311-8. [CrossRef] [PubMed] [PubMed Central]
- Laufman O, Perrino J, Andino R. Viral generated inter-organelle contacts redirect lipid flux for genome replication. Cell. 2019 Jul 11;178(2):275-289.e16. [CrossRef] [PubMed] [PubMed Central]
- Li YJ, Chen CY, Yang JH, Chiu YF. Modulating cholesterol-rich lipid rafts to disrupt influenza A virus infection. Front Immunol. 2022 Sep 13;13:982264. [CrossRef] [PubMed] [PubMed Central]
- Liu Z, Smith H, Criglar JM, Valentin AJ, Karandikar U, Zeng XL, Estes MK, Crawford SE. Rotavirus-mediated DGAT1 degradation: A pathophysiological mechanism of viral-induced malabsorptive diarrhea. Proc Natl Acad Sci U S A. 2023 Dec 19;120(51):e2302161120. [CrossRef] [PubMed] [PubMed Central]
- Madsen JJ, Rossman JS. Cholesterol and M2 rendezvous in budding and scission of influenza A virus. Subcell Biochem. 2023;106:441-459. [CrossRef] [PubMed]
- Marcinkiewicz A, Gauthier D, Garcia A, Brasaemle DL. The phosphorylation of serine 492 of perilipin a directs lipid droplet fragmentation and dispersion. J Biol Chem. 2006 Apr 28;281(17):11901-9. [CrossRef] [PubMed]
- Mejhert N, Kuruvilla L, Gabriel KR, Elliott SD, Guie MA, Wang H, Lai ZW, Lane EA, Christiano R, Danial NN, Farese RV Jr, Walther TC. Partitioning of MLX-family transcription factors to lipid droplets regulates metabolic gene expression. Mol Cell. 2020 Mar 19;77(6):1251-1264.e9. [CrossRef] [PubMed] [PubMed Central]
- Miyanari Y, Atsuzawa K, Usuda N, Watashi K, Hishiki T, Zayas M, Bartenschlager R, Wakita T, Hijikata M, Shimotohno K. The lipid droplet is an important organelle for hepatitis C virus production. Nat Cell Biol. 2007 Sep;9(9):1089-97. Epub 2007 Aug 26. Erratum in: Nat Cell Biol. 2007 Oct;9(10):1216. [CrossRef] [PubMed]
- Nejmeddine M, Trugnan G, Sapin C, Kohli E, Svensson L, Lopez S, Cohen J. Rotavirus spike protein VP4 is present at the plasma membrane and is associated with microtubules in infected cells. J Virol. 2000 Apr;74(7):3313-20. [CrossRef] [PubMed] [PubMed Central]
- Papa G, Borodavka A, Desselberger U. Viroplasms: Assembly and functions of rotavirus replication factories. Viruses. 2021 Jul 12;13(7):1349. [CrossRef] [PubMed] [PubMed Central]
- Parker EP, Ramani S, Lopman BA, Church JA, Iturriza-Gómara M, Prendergast AJ, Grassly NC. Causes of impaired oral vaccine efficacy in developing countries. Future Microbiol. 2018 Jan;13(1):97-118. [CrossRef] [PubMed] [PubMed Central]
- Qu Y, Wang W, Xiao MZX, Zheng Y, Liang Q. The interplay between lipid droplets and virus infection. J Med Virol. 2023 Jul;95(7):e28967. [CrossRef] [PubMed]
- Roingeard P, Eymieux S, Burlaud-Gaillard J, Hourioux C, Patient R, Blanchard E. The double-membrane vesicle (DMV): a virus-induced organelle dedicated to the replication of SARS-CoV-2 and other positive-sense single-stranded RNA viruses. Cell Mol Life Sci. 2022 Jul 16;79(8):425. [CrossRef] [PubMed] [PubMed Central]
- Samsa MM, Mondotte JA, Iglesias NG, Assunção-Miranda I, Barbosa-Lima G, Da Poian AT, Bozza PT, Gamarnik AV. Dengue virus capsid protein usurps lipid droplets for viral particle formation. PLoS Pathog. 2009 Oct;5(10):e1000632. [CrossRef] [PubMed] [PubMed Central]
- Shavinskaya A, Boulant S, Penin F, McLauchlan J, Bartenschlager R. The lipid droplet binding domain of hepatitis C virus core protein is a major determinant for efficient virus assembly. J Biol Chem. 2007 Dec 21;282(51):37158-69. [CrossRef] [PubMed]
- Tabata K, Prasad V, Paul D, Lee JY, Pham MT, Twu WI, Neufeldt CJ, Cortese M, Cerikan B, Stahl Y, Joecks S, Tran CS, Lüchtenborg C, V'kovski P, Hörmann K, Müller AC, Zitzmann C, Haselmann U, Beneke J, Kaderali L, Erfle H, Thiel V, Lohmann V, Superti-Furga G, Brügger B, Bartenschlager R. Convergent use of phosphatidic acid for hepatitis C virus and SARS-CoV-2 replication organelle formation. Nat Commun. 2021 Dec 14;12(1):7276. [CrossRef] [PubMed] [PubMed Central]
- Tansey JT, Huml AM, Vogt R, Davis KE, Jones JM, Fraser KA, Brasaemle DL, Kimmel AR, Londos C. Functional studies on native and mutated forms of perilipins. A role in protein kinase A-mediated lipolysis of triacylglycerols. J Biol Chem. 2003 Mar 7;278(10):8401-6. [CrossRef] [PubMed]
- Theken KN, Tang SY, Sengupta S, FitzGerald GA. The roles of lipids in SARS-CoV-2 viral replication and the host immune response. J Lipid Res. 2021;62:100129. [CrossRef] [PubMed] [PubMed Central]
- Troeger C, Khalil IA, Rao PC, Cao S, Blacker BF, Ahmed T, Armah G, Bines JE, Brewer TG, Colombara DV, Kang G, Kirkpatrick BD, Kirkwood CD, Mwenda JM, Parashar UD, Petri WA Jr, Riddle MS, Steele AD, Thompson RL, Walson JL, Sanders JW, Mokdad AH, Murray CJL, Hay SI, Reiner RC Jr. Rotavirus vaccination and the burden of rotavirus diarrhea among children younger than 5 years. JAMA Pediatr. 2018 Oct 1;172(10):958-965. Erratum in: JAMA Pediatr. 2022 Feb 1;176(2):208. [CrossRef] [PubMed] [PubMed Central]
- Vieyres G, Pietschmann T. HCV pit stop at the lipid droplet: Refuel lipids and put on a lipoprotein coat before exit. Cells. 2019 Mar 12;8(3):233. [CrossRef] [PubMed] [PubMed Central]
- Viktorova EG, Nchoutmboube JA, Ford-Siltz LA, Iverson E, Belov GA. Phospholipid synthesis fueled by lipid droplets drives the structural development of poliovirus replication organelles. PLoS Pathog. 2018 Aug 27;14(8):e1007280. [CrossRef] [PubMed] [PubMed Central]
- Waheed AA, Zhu Y, Agostino E, Naing L, Hikichi Y, Soheilian F, Yoo SW, Song Y, Zhang P, Slusher BS, Haughey NJ, Freed EO. Neutral sphingomyelinase 2 is required for HIV-1 maturation. Proc Natl Acad Sci U S A. 2023 Jul 11;120(28):e2219475120. [CrossRef] [PubMed] [PubMed Central]
- Yoo SW, Waheed AA, Deme P, Tohumeken S, Rais R, Smith MD, DeMarino C, Calabresi PA, Kashanchi F, Freed EO, Slusher BS, Haughey NJ. Inhibition of neutral sphingomyelinase 2 impairs HIV-1 envelope formation and substantially delays or eliminates viral rebound. Proc Natl Acad Sci U S A. 2023 Jul 11;120(28):e2219543120. [CrossRef] [PubMed] [PubMed Central]
- Zhang J, Lan Y, Li MY, Lamers MM, Fusade-Boyer M, Klemm E, Thiele C, Ashour J, Sanyal S. Flaviviruses exploit the lipid droplet protein AUP1 to trigger lipophagy and drive virus production. Cell Host Microbe. 2018 Jun 13;23(6):819-831.e5. [CrossRef] [PubMed]
- Zhao J, Zeng Z, Chen Y, Liu W, Chen H, Fu ZF, Zhao L, Zhou M. Lipid droplets are beneficial for rabies virus replication by facilitating viral budding. J Virol. 2022 Jan 26;96(2):e0147321. [CrossRef] [PubMed] [PubMed Central]
Figure 1.
RV-infected cells grown on coverslips were processed for confocal microscopy (CM) as described by Nejmeddine et al, 2000. Cells were blocked with 1%BSA-0.1%triton X-100. Primary antibodies against perilipin A (rabbit polyclonal) were from Abcam, those against NSP2 (mouse monoclonal) were a kind gift of Dr Oscar Burrone, ICGEB, Trieste IT. Secondary antibodies were goat anti-rabbit IgG conjugated with Alexafluor 633 and goat anti-mouse IgG conjugated with Alexafluor 488, both from Invitrogen. Staining with secondary antibodies was in the presence of 2ug/ml of Hoechst33342 (Sigma). Coverslips were mounted on glass slides with Prolong bold Antifade mounting medium (Molecular Probes) and observed by CM using a Leica DM Libre TCS SP instrument. An individual viroplasm-LD complex is magnified in the inserts. From: Cheung W et al, 2010.
Figure 1.
RV-infected cells grown on coverslips were processed for confocal microscopy (CM) as described by Nejmeddine et al, 2000. Cells were blocked with 1%BSA-0.1%triton X-100. Primary antibodies against perilipin A (rabbit polyclonal) were from Abcam, those against NSP2 (mouse monoclonal) were a kind gift of Dr Oscar Burrone, ICGEB, Trieste IT. Secondary antibodies were goat anti-rabbit IgG conjugated with Alexafluor 633 and goat anti-mouse IgG conjugated with Alexafluor 488, both from Invitrogen. Staining with secondary antibodies was in the presence of 2ug/ml of Hoechst33342 (Sigma). Coverslips were mounted on glass slides with Prolong bold Antifade mounting medium (Molecular Probes) and observed by CM using a Leica DM Libre TCS SP instrument. An individual viroplasm-LD complex is magnified in the inserts. From: Cheung W et al, 2010.
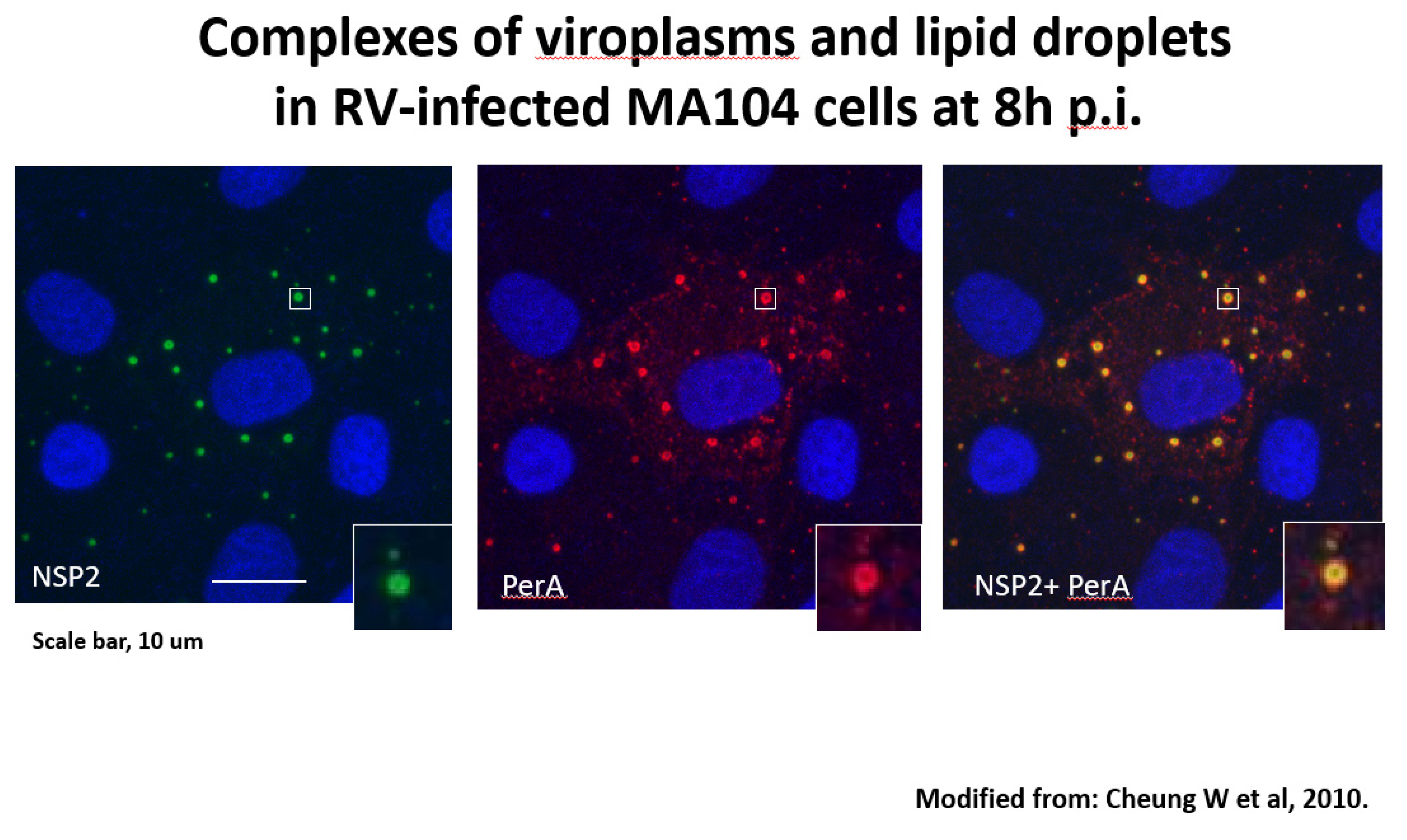

Table 1.
Comparison of inhibitory effects of different compounds affecting lipid droplet homeostasis on rotavirus replication.
Table 1.
Comparison of inhibitory effects of different compounds affecting lipid droplet homeostasis on rotavirus replication.
| Treatment of cells | Viral dsRNA | Infectivity of progeny | |||||
|---|---|---|---|---|---|---|---|
|
Relative Valuea |
Differenceb |
log TCID50/ml ± S.E. (n) |
Differenceb | ||||
| Isoproterenol+IBMXc | - | 1.00 | 8.2 ± 0.3 (3) | ||||
| + | 0.25 | 4.0-fold | 6.5 ± 0.1 (3) | 50-fold | |||
| Triacsin Cc | - | 1.00 | 7.5 ± 0.1 (3) | ||||
| + | 0.26 | 3.8-fold | 6.2 ± 0.2 (3) | 20-fold | |||
| TOFAd | - | 1.00 | 8.4 ± 0.5 (6) | ||||
| + | 0.17 | 5.9-fold | 6.7 ± 0.5 (6) | 50-fold | |||
a Calculated from densitometric values of RNA gels (Cheung et al, 2010). b Underlining indicates statistical difference (P<0.05). c From: Cheung et al, 2010. d From: Gaunt et al, 2013a.
Table 2.
Lipid metabolism involved in the replication of RNA viruses (other than rotaviruses).
| Virus | Mechanism | References |
|---|---|---|
| Hepatitis C virus (HCV) | Formation of double membrane vesicles (DMVs) for viral replication, interacting with LDs via viral core protein D2 domain and NS5A; role of apolipoproteins; dynamic profile of lipidome-DMV complexes during HCV infection | Miyanari et al, 2007; Shavinskaya et al, 2007; Boulant et al, 2007; Chatel-Chaix and Bartenschlager,2014; Vieyres and Pietschmann, 2019; Islam et al, 2023 |
| GB virus-B | similar to HCV | Hope et al, 2002 |
| Dengue virus | Viral capsid protein-LD interactions; similar to HCV | Samsa et al, 2009; Chatel-Chaix and Bartenschlager, 2014 |
| Zika virus | Disturbance of lipid homeostasis; induction of mitochondrial dysfunction | Chen Q et al, 2020 |
| Picornaviruses | Phosphatidylcholine synthesis to support membranous replication complexes; hydrolysis of TAGs in LDs, lipolysis | Victorova et al, 2018; Laufman et al, 2019; Belov and von Kuppeveld, 2019 |
| Norovirus | Multimembrane vesicle clusters replication organelles, as in picornaviruses, containing NoV NS proteins; | Doerflinger et al, 2017 |
| SARS-CoV-2 | DMVs as replication complexes; viral reprograming of lipid synthesis pathways; LDs as assembly platforms? | Dias et al, 2020; Roingeard et al, 2022; Cesar-Silva et al, 2023 |
| HIV-1 | Requirement of ceramide for viral capsid maturation and full infectivity | Waheed et al, 2023; Yoo et al, 2023 |
| Influenza virus | Upregulation of LDs upon infection; viral replication depending on cellular lipase activity; elevation of lipid metabolites in IV-infected cells | Baek et al, 2022; Chen X et al, 2023 |
Table 3.
Compounds interacting with the cellular lipidome as potential candidate antivirals.
| Virus | Compound |
| Rotavirus | TOFA; [isoproterenol + IBMX] |
| Hepatitis C virus | Lipid-specific compounds targeting the interaction of viral core proteins with LDs |
| Zikavirus | DGAT-1 inhibitor A922500 |
| Enterovirus | Adipose triglyceride lipase inhibitor atglistatin |
| SARS-CoV-2 | Inhibitors of sphingolipid metabolism; inhibitors of inflammatory compounds; lipase inhibitors; statins |
| HIV-1 | Neutral sphingomyelinase 2 inhibitor PDDC, preventing ceramide biosynthesis |
| Influenza A virus | methyl-beta-cytodextrin; sphingomyelinase inhibitors; lipase inhibitors; statins |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



