
Submitted:
15 April 2024
Posted:
17 April 2024
You are already at the latest version
Abstract
The predatory mirids Macrolophus pygmaeus Ramber and Nesidiocoris tenuis Reuter (Heteroptera: Miridae) are used for the biological control of Tuta absoluta Meyrick (Lep.: Gelechiidae) and other pests in tomato greenhouses. The functional response of 1 (young) and 10 (old) days old adult females and males of M. pygmaeus and N. tenuis on eggs of T. absoluta, was determined on tomato at two temperatures (20° and 25°C) and LD 16:8 and found to be Type II. M. pygmaeus females exhibited higher predation efficiency, than males at both tested temperatures. Young M. pygmaeus females had a higher efficiency than old ones, whereas males had a low efficiency irrespective of age. The predation efficiency of N. tenuis was high (but lower than M. pygmaeus) in both young females and males, although old females had a higher efficiency than the respective males. Our results show that the two predatory species have different functional response characteristics to their prey depending mainly on sex and age which may affect their role as biological control agents.
Keywords:
M. pygmaeus
; N. tenuis
; Miridae
; T.absoluta
; functional response
; tomato
; sex
; adult age
; temperature
; predator
1. Introduction
The dicyphine mirid bugs Macrolophus pygmaeus Ramber and Nesidiocoris tenuis Reuter (Heteroptera: Miridae), are endemically present in the Mediterranean basin and are commercialized as beneficial insects in biological pest control systems [1,2,3]. M. pygmaeus has been utilized for nearly three decades, predominantly in augmentative biological control in greenhouses, to regulate such small vegetable crop pests as spider mites, whiteflies, aphids, thrips, and lepidopteran pests (mainly eggs and young larvae) [2,3,4]. Despite being identified as an actor in biological control later, N. tenuis has gained significant attention, due to its great predatory capacity and performance at high temperatures [5,6]. The capacity of the two species as biocontrol agents against Tuta absoluta Meyrick (Lepidoptera: Gelechiidae), has been investigated and highlighted [7,8]. Both M. pygmaeus and N. tenuis exhibit zoophytophagy, a special type of omnivory, with the ability to feed on both plant tissue and a broad spectrum of arthropods [9]. The presence of both peptidase and amylase in the salivary glands has been associated with this polyphagous feeding capacity [10]. These two species have a proclivity for zoophagy, although they facultatively exhibit phytophagy, which may play a substantial role in their preservation and fitness [9,11,12,13,14,15,16,17,18,19]. The ability to utilize both trophic sources offers a significant advantage when it comes to the desired early establishment of the predator in the crop before pest infestation proliferates [20]. M. pygmaeus has the added ability to complete its life cycle without access to prey, feeding exclusively on the host plant tissue of tomato and a multitude of cultivated vegetable plants [21]. To contrast this, there are few recorded plant species, notably the native to tropical and subtropical regions Sesasum indicum Linnaeus (Pedaliaceae) and Cleome hassleriana Chod. (Brassicales: Cleomaceae) that are suitable banker plants for the preyless development and oviposition of N. tenuis [22,23]. N. tenuis is considered to possess a greater voracity than M. pygmaeus, and in cases of high population densities may cause economically important plant damage and yield loss, in the form of aborted flowers and fruit, and necrotic rings, due to phytophagy on vegetative and reproductive plant parts [24,25,26,27]. As such, the status of N. tenuis as pest or beneficial insect has been a point of contention [28,29]. However, in the Mediterranean region, the benefits emanating from its high generalist predation capacity, especially on more than one tomato pests, such as the invasive pest T. absoluta and whiteflies, have been argued to outweigh the danger of plant feeding-associated damage [9].
The microlepidodpteran pest, T. absoluta is considered one of the most devastating pests of tomato, along with the whitefly, Trialeurodes vaporariorum Westwood (Homoptera: Aleyrodidae). Native to South America, it has since extended its presence to Europe (beginning with Spain in 2006), Africa (starting with N.Africa, Tunisia and Morocco in 2008), and Asia (starting with Turkey in 2009) [30,31]. The larvae are voracious and feed on all aerial plant parts, in protected galleries within leaves, stems, and fruit. When appropriate control measures aren’t implemented, infestation can lead to complete crop failure [31]. Certain characteristics associated with the ecology and the biology of this pest also favour the development of resistance to a broad range of insecticides, rendering control ineffective [32,33,34,35,36]. The inadequacy of chemical control in delivering sufficient crop protection serves as a catalyst for endeavours aimed at developing and implementing alternative control methods, including the utilization of beneficial insects, as integral components of Integrated Pest Management (IPM).
Functional response experiments have been a longstanding tool for evaluating the effectiveness of biocontrol agents in controlling arthropod pests and defining the range of pest densities and conditions within which the biological control agent remains effective. The functional response of a predator to prey density describes the per capita feeding rate of a predator as a function of resource density and is classified according to the three types described by Holling, 1959 [37]. Both Type II and Type III functional responses of predators, are desired characteristics for beneficial arthropods utilized as biological control agents and have been reported for many mirid predators [38,39,40,41,42]. Two parameters, the attack rate coefficient (α) (ability to capture) and the handling time (Th) (ability to handle and consume prey), are used to provide insight into the functional response of the predator. Especially in the case of females, such foraging performance information is impactful, as it directly controls both the quantity and quality of offspring, thus influencing the establishment of predators, the dynamics of the populations, and the overall predation efficacy [43].
Various functional response experiments have been conducted to elucidate the efficacy of the two species when consuming a multitude of small arthropods, with the results depending on factors, such as host plant, spacial complexity, prey item, prey instar and temperature [40,41,42,44,45,46,47,48,49,50].
The primary objective of this study was to assess the predatory efficacy of the two species, M. pygmaeus and N. tenuis against eggs of T. absoluta. Functional response experiments were conducted at seven predefined densities of synchronous prey T.absoluta eggs under laboratory conditions, taking into account the impact of adult sex and predator aging on predation, at two experimental temperatures. Our findings indicate that both young and old males and females of the two predatory species exhibit a Type II functional response at the two experimental temperatures. However, the two species have different functional response characteristics to their prey depending mainly on their sex and age which may influence their role as biological control agents.
2. Materials and Methods
2.1. Insect Colonies
The stock colonies of M. pygmaeus and N. tenuis were maintained in the laboratory in cylindrical plastic cages (diameter, 25cm, height, 25cm), with fresh bean pods as oviposition substrate and dehydrated E. kuehniella Zeller (Lepidoptera: Pyralidae) eggs ad libitum, at 25°C and a photoperiod of 16:8 (L:D).
The stock colony of T. absoluta was established with adults emerged from heavily infested tomato leaves collected from a commercial plantation in northern Greece. The colony was maintained on tomato plants in tent cages, at 25°C and a photoperiod of 16:8 (L:D).
Predation experiments were conducted at two temperatures (20° and 25°C), a photoperiod of 16:8 (L:D) and 65 ±5% RH. One or 10-days old adult females and males of either M. pygmaeus or N. tenuis from the laboratory colonies were kept in an empty rearing cage, with access only to water for 24 hours, before their use in the functional response experiments.
2.2. Bioassay
For the functional response experiments, a large number of approximately 300 T. absoluta adults were transferred from the laboratory colony to an insect rearing cage (60cmx60cmx60cm) for egg laying. Tomato leaflets of comparable size (approximately 6cm, height and 3cm, width) were excised from tomato plants and their petioles enveloped in dampened cotton wool in plastic vials with water. The vials with the leaflets were maintained for 1 to 3 hours in the ovipositional cage for T. absoluta egg laying. Subsequently, the number of eggs laid on both the upper and under side of each leaflet was scored and the complete leaflet served as experimental arena. In functional response experiments the predator’s mobility is limited to the controlled experimental arena, whereas in natural agroecosystems, both predator and prey freely navigate. Our arenas were adopted in an attempt to align the experimental conditions more closely with field conditions, where the predator consumes T. absoluta eggs deposited on both upper and underside leaf surfaces. The egg densities were selected through preliminary tests, to ensure maximum levels of predation.
The tomato leaflets with the eggs were transferred into individual cages consisting of inverted plastic cups (volume, 500cm3) with dual openings on opposite sides of their cylindrical walls, covered with fine mesh, to facilitate air circulation. Each cup was sealed with a bottom lid and included a circular aperture at the top, allowing the introduction of the predator individual into the experimental arena before being sealed with a cork.
The experimental setup comprised an individual cage with one leaflet positioned in the vial, with a specific egg density, to which either a previously starved male or female of M. pygmaeus or N. tenuis was added. After a 24-hour period, the number of eggs consumed by the predator was scored.
2.3. Data Analysis
The mean number of prey consumed was plotted, as a function of prey density, and the data fitted to Rogers and Royamas’ random predator equation:
Ne is the number of prey/eggs consumed, N0 the initial prey density, Th the handling time, T the duration of the experiment (24 hours) and α the attack rate.
The values for the parameters α and Th were estimated through a non-linear fit of the data to the provided above equation in Python, accounting for the standard deviation of the data. The determination coefficient, R2, was calculated in addition to the parameters’ errors.
The statistical analysis concerning the effect of age, sex and predator species on the prey consumption at each temperature, was quantified through a three-way analysis of variance (ANOVA, type I), accounting for a generalized linear model (GLM) with quasipoisson distribution. The relevant analysis was performed in R (version 4.3.1; R core team 2023-06-16).
3. Results
3.1. Predation of M. pygmaeus on Eggs of T. absoluta
The functional response data of adult females and males of M. pygmaeus on eggs of T. absoluta was successfully fitted to Royama (1971) and Rogers (1972) equation and consequently exhibit a Type II functional response (Figure 1, Table 1).
As shown in Table 1, the estimated values of attack rate in 1-day old females were ~0.70 and ~0.51 and those of the handling time ~0.31 and ~0.27 hours, at 20° and 25°C respectively, whereas in males, the respective estimated values of attack rate were lower (~0.09 and ~0.10) and those of the handling time longer (~0.57 and ~0.33 hours).
In 10-day old females the estimated values of attack rate were ~0.33 and ~0.36 and handling time were ~0.28 and ~0.24 hours respectively, at 20° and 25°C, whereas in males the respective values of attack rate were ~0.19 and ~0.08 and those of handling times ~0.61 and ~0.42 hours respectively.
Our results show that females exhibited markedly higher attack rates and shorter handling times, than their male counterparts at both tested temperatures and age groups.
The lowest value of attack rate of females was ~0.33 and the highest ~0.70. The females’ shortest handling time was ~0.24 hours and the longest, ~0.31 hours. In comparison, the lowest attack rate value obtained for males was ~0.08 and the highest attack rate, ~0.19, much lower than the values for the females. The shortest handling time values for males were ~0.33 hours and the longest, ~0.61 hours, the latter being almost double the handling time of the females.
3.2. Predation of N. tenuis on Eggs of T. absoluta
As in M. pygmaeus, the functional response data of adult females and males of N. tenuis on eggs of T. absoluta was successfully fitted to the Royama (1971) and Rogers (1972) equation (Figure 2, Table 1). For N. tenuis the estimated values of the functional response parameters of the attack rate (a) and handling time (Th) are lower than the respective ones for M. pygmaeus (Table 1).
In 1-day old females of N. tenuis the estimated values of attack rate (a) were ~ 0.22 (compared to 0.70 for M. pygmaeus (M.p.)) and ~ 0.14 (0.51 for M.p.) and those of handling time ~0.18 (0.31 for M.p.) and ~0.10 (0.27 for M.p.) hours at 20° and 25°C respectively, whereas in males the respective estimated values of attack rate were ~0.20 (0.09 for M.p.) and ~0.10 (0.10 for M.p.) and those of the handling time ~0.24 (0.57 for M.p.) and ~ 0.19 (0.33 for M.p.) hours.
In 10 day old females the estimated values of attack rate were ~0.18 (compared to 0.33 for M. pygmaeus(M.p.)) and ~0.09 (0.36 for M.p.) and those of handling time were ~0.25 (0.28 for M.p.) and ~0.14 (0.24 for M.p.) hours respectively at 20° and 25°C, whereas in males the respective values of attack rate were ~0.09 (0.19 for M.p.) and ~ 0.04 (0.08 for M.p.) and those of handling time ~0.37 (0.61 for M.p.) and ~0.19 (0.42 for M.p.) hours.
Our results show that in N. tenuis, as in M. pygmaeus, females exhibited higher attack rates and shorter handling times, than their male counterparts at both tested temperatures and age groups. Based on these laboratory results we may assume that M. pygmaeus could be a more effective predator than N. tenuis for eggs of T. absoluta on tomato leaves.
3.3. Comparison of Predation Efficacy of M. pygmaeus and N. tenuis
Three-way ANOVA indicated that the effects of predator species, sex, age at both experimental temperatures and the interactions of predator*sex and predator*age at 20°C on the consumption of T. absoluta eggs were significant.

Table 2.
Three-way ANOVA (quasi-poisson distribution) on factors affecting consumption of eggs of T. absoluta by M. pygmaeus and N. tenuis, at 20 °C and 25 °C.
Table 2.
Three-way ANOVA (quasi-poisson distribution) on factors affecting consumption of eggs of T. absoluta by M. pygmaeus and N. tenuis, at 20 °C and 25 °C.
| Source | df | Deviance |
|---|---|---|
|
20 °C |
||
| Predator | 1 | 211.73a |
| Sex | 1 | 591.80a |
| Age | 1 | 104.83b |
| Predator*Sex | 1 | 88.28b |
| Predator*Age | 1 | 51.49c |
| Sex*Age | 1 | 5.82 |
| Predator*Sex*Age | 1 | 11.89 |
|
25 °C |
||
| Predator | 1 | 52.80c |
| Sex | 1 | 578.81a |
| Age | 1 | 136.81b |
| Predator*Sex | 1 | 24.74 |
| Predator*Age | 1 | 24.66 |
| Sex*Age | 1 | 23.53 |
| Predator*Sex*Age | 1 | 5.70 |
a: P<0.001, b: P<0.01, c: P<0.05.
4. Discussion
Functional response experiments illustrate the relationship between the initial prey density and the number of prey killed or parasitized and, based on this, the type of functional response with its parameters, the attack rate and handling time are estimated. From this determination, valuable practical conclusions can be derived of the suitability of a potential biological control agent against a pest of a crop. The dynamics of this predator-prey interactions are defined by a plethora of factors, such as predator age and sex, developmental instar and prey species [50]. In our experiments, we determined the functional responses, and estimated the values of attack rate and handling time of newly emerged (1 day old) and old (10 day old) M. pygmaeus and N. tenuis adult females and males feeding on different densities of T. absoluta eggs, laid on tomato leaves. We found that, in M. pygmaeus, the estimated values of attack rate were higher and handling time lower than the respective values for N. tenuis. The age and sex of both predator species affected the functional response parameters, i.e. handling time was lower and attack rate higher in female and young individuals than in males and old individuals.
The functional responses of M. pygmaeus and N. tenuis have been studied by other researchers following different experimental designs, prey densities and host plants [40,41,42,44,45,46,47,48,49]. Our findings are in accordance with the functional response type obtained in a study by Sharifian et al., 2015 [41], where 1-day old adults of unspecified sex of M. pygmaeus and N. tenuis fed on different densities of T. absoluta eggs, at 25°C. However, in this study, the values of attack rate and handling time did not differ between M. pygmaeus and N. tenuis, and were quite different to those obtained in our study. These observed differences between the obtained results of our and the aforementioned study, may be due to, among other factors, the different design of the experimental arenas, and the different sex of the experimental predators. It is known that the arena size and the varying spatial distribution of the prey affect functional response parameters, namely the attack rate [52,53]. The unique experimental design described in our experiments was adopted in an attempt to align the experimental conditions more closely with field conditions, where the predator consumes T. absoluta eggs laid by adult females freely on both sides of the leaf surface. By contrast, Sharifian et al., (2015) utilized an experimental arena consisting of three tomato leaflets in a petri dish, onto which the desired densities of eggs were added. These factors may provide possible explanations for the differential values of the functional response parameters.
In our functional response experiments the predators M. pygmaeus and N. tenuis preyed on T. absoluta eggs and showed a type II response. Prey type and even prey developmental stage may modify the functional response of predators. M. pygmaeus adult females have been shown to exhibit a Type II functional response when feeding on 2nd – 4th instar nymphs of Myzus persicae Sulzer (Hemiptera: Aphididae) [44,48], pupae of T. vaporariorum [54], eggs of E. kuenhiella and eggs of T. absoluta [41], the latter in accordance with the results of our experiments and Type III when feeding on eggs of T. vaporariorum [55]. Adult females of N. tenuis have been shown to exhibit a Type II response when feeding on T. vaporariorum nymphs [56], Tetranychus urticae Koch (Tetranychidae) adults [57], and eggs of T. absoluta [41], the latter in accordance with the results of our experiments and Type III when consuming eggs and nymphs of Bemisia tabaci Gennadius (Homoptera: Aleyrodidae) [49].
In certain predator insect and mite species, the functional response is affected by adult age. The functional response of the predator mite Amblyseius swirski Athias-Henriot (Acari: Phytoseiidae) feeding on T. urticae was of Type II for a major part of its life, with the exception of 12-day old females that exhibited a Type III functional response. This was attributed to the greater energy demands, due to the higher reproductive output of females at that age [58]. A Type III response was also obtained with 1-day old, mated females of the parasitoid Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae), when feeding on E. kuehniella eggs and a Type II response, with 2-9 day old females which may be due to the effect of age progression on searching efficacy (varied effect) and handling time (increase) [59]. For the mirid bugs Dicyphus bolivari and Dicyphus errans Wolff (Hemiptera: Miridae), both fifth instar nymphs and seven-day old females feeding on T. absoluta eggs exhibited a Type II response [60]. In the case of M. pygmaeus, the last nymphal instar and adult females, generally, have been shown to consume greater numbers of M. persicae Sulzer [61]. In our experiments, M. pymaeus and N. tenuis females and males maintained a Type II functional response as newly emerged (1 day old) and old (10 day old) adults.
We determined the functional responses of M. pygmaeus and N. tenuis, at two temperatures (20° and 25°C) and found a type II functional response with no substantial differences in the estimated values of attack rate and handling time between the two temperatures. It has been noted that changes in the ecological settings may be reflected in the functional response and associated parameters [52]. Investigations into the temperature thresholds of dicyphine species have determined that those vulnerable to low temperatures, are more resilient to high temperatures and vice versa. N. tenuis is considered the most thermophilous of the dicyphine species that have been investigated thus far, with optimum performance at temperatures between 20 and 30°C [62], and individuals reaching adulthood at a maximum temperature of 35°C [63]. Interestingly, in the context of the functional response, attack rate and handling time are expected to be optimized at intermediate temperatures [64,65]. As such, N. tenuis females have been indicated to exhibit a Type II response when consuming B. tabaci (Hemiptera: Aleyrodidae) at relatively low temperatures (15 and 25°C) and Type III response at a higher temperature of 35°C [66]. Conversely, M. pygmaeus maintained a Type II functional response over a range of temperatures [67].
We provide the first evidence of a M. pygmaeus population outperforming a population of N. tenuis. The previous history of an individual, such as previous host, prey that it was reared on, may influence its behaviour during the functional response experiment [43]. The two species we examined were reared in the lab on bean pods and E. kuehniella eggs. A preparatory period, where the predators were allowed to remain on tomato plants and feed on T. absoluta eggs, could have benefited and altered the outcome of the experiments.
In conclusion, our results provide evidence into the combined effect of sex and ageing on the functional response parameters of M. pygmaeus and N. tenuis at two temperatures and show that in both predator species, females have higher predation efficiency than males. In addition, newly emerged females have higher predation efficiency than 10-day old ones. Based on the estimated values of attack rate and handling time, we conclude that M. pygmaeus may have a better predation efficiency than N. tenuis for eggs of T. absoluta on tomato. However, further field experiments are required in order to verify our present laboratory results.
Author Contributions
Conceptualization, E.I.Y. and D.S.K; methodology, E.Y., D.S.K. and N.A.K.; software, S.D.K.; validation, D.S.K., N.A.K.; formal analysis, S.D.K.; investigation, E.I.Y.; resources, D.S.K., N.A.K.; data curation, D.S.K., E.Y.; writing—original draft preparation, E.I.Y.; writing—review and editing, D.S.K.; visualization, S.D.K.; supervision, D.S.K.; project administration, D.S.K.; funding acquisition, E.I.Y. All authors have read and agreed to the published version of the manuscript.
Funding
The research work was supported by the Hellenic Foundation for Research and Innovation (HFRI) under the 3rd Call for HFRI PhD Fellowships (Fellowship number: 6104).
Acknowledgments
Gratitude is extended to Stefanos Kovaios, PhD student in Informatics, AUTh for carrying out the statistical analysis of our results and providing constructive feedback on a previous draft of our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pérez-Hedo, M.; Urbaneja, A. Prospects for predatory mirid bugs as biocontrol agents of aphids in sweet peppers. J. Pest Sci. 2015, 88, 65–73. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Lopez-Gallego, E.; Pérez-Marcos, M.; Perera-Fernandez, L.G.; Ramirez-Soria, M.J.; 2018. How safe is it to rely on Macrolophus pygmaeus (Hemiptera: Miridae) as a biocontrol agent in tomato crops? Front. Ecol. Evol. 2018, 6, 132. [Google Scholar] [CrossRef]
- Arnó, J.; Castañé, C.; Alomar, O. ; et.al. Forty years of biological control in Mediterranean tomato greenhouses: The story of success. Isr. J. Entomol 2018, 48, 209–226. [Google Scholar]
- Moerkens, R.; Berckmoes, E.; Van Damme, V.; Wittemans, L.; Tirry, L.; Casteels, H. , et al. Inoculative release strategies of Macrolophus pygmaeus Rambur (Hemiptera: Miridae) in tomato crops: population dynamics and dispersal. J. Plant Dis. Prot. 2017, 124, 295–303. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Lacasa, A.; Arnó, J.; Castañé, C.; Alomar, O. Life history parameters for Nesidiocoris tenuis (Reuter) (Het., Miridae) under different temperature regimes. J. Appl. Entomol. 2009, 133, 125–32. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Pedroche, V.; Urbaneja, A. Temperature-driven selection of predatory Mirid bugs for improving aphid control in sweet pepper crops. Horticulturae 2023, 9, 572. [Google Scholar] [CrossRef]
- Urbaneja, A.; Montón, H.; Mollá, O. Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. J. Appl. Entomol. 2009, 133, 292–296. [Google Scholar] [CrossRef]
- Zappalà, L.; Biondi, A.; Alma, A.; Al-Jboory, I.J.; Arnò, J.; Bayram, A.; et al. Natural enemies of the South American moth, Tuta absoluta, in Europe, North Africa and Middle East, and their potential use in pest control strategies. J. Pest Sci. 2013, 86, 635–647. [Google Scholar] [CrossRef]
- Castañé, C.; Arnó, J.; Gabarra, R.; Alomar, O. Plant damage to vegetable crops by zoophytophagous mirid predators. Biol. Control 2011, 59, 22–29. [Google Scholar] [CrossRef]
- Torres, J.B.; Boyd, D.W. Zoophytophagy in predatory Hemiptera. Braz. Arch. Biol. Technol. 2009, 52, 1199–208. [Google Scholar] [CrossRef]
- Urbaneja, A.; Tapia, G.; Stansly, P. Influence of host plant and prey availability on developmental time and survivorship of Nesidiocoris tenius (Het.: Miridae). Biocontrol Sci. Technol. 2005, 15, 513–518. [Google Scholar] [CrossRef]
- Perdikis, D.C.; Lykouressis, D.P. Macrolophus pygmaeus (Hemiptera: Miridae) Population parameters and biological characteristics when feeding on eggplant and tomato without prey. J. Econ. Entomol. 2004, 97, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Maselou, D.A.; Perdikis, D.C.; Sabelis, M.W.; Fantinou, A.A. Use of plant resources by an omnivorous predator and the consequences for effective predation. Biol. Control 2014, 79, 92–100. [Google Scholar] [CrossRef]
- De Puysseleyr, V.; De Man, S.; Höfte, M.; De Clercq, P. Plantless rearing of the zoophytophagous bug Nesidiocoris tenuis. BioControl 2013, 58, 205–213. [Google Scholar] [CrossRef]
- Sanchez, J.A. Zoophytophagy in the plantbug Nesidiocoris tenuis. Agric. For. Entomol. 2008, 10, 75–80. [Google Scholar] [CrossRef]
- Lykouressis, D.; Perdikis, D.; Charalampous, P. Plant food effects on prey consumption by the omnivorous predator Macrolophus pygmaeus. Phytoparasitica 2014, 42, 303–309. [Google Scholar] [CrossRef]
- Ingegno, B.L.; Pansa, M.G.; Tavella, L. Plant preference in the zoophytophagous generalist predator Macrolophus pygmaeus (Heteroptera: Miridae). Biol. Control 2011, 58, 174–181. [Google Scholar] [CrossRef]
- Castañé, C; Zapata, R. Rearing the predatory bug Macrolophus caliginosus on a meat-based diet. Biol. Control 2005, 34, 66–72. [Google Scholar] [CrossRef]
- Lalonde, R. G.; McGregor, R. R.; Gillespie, D. R.; Roitberg, B. D. (1999). Plant-feeding by arthropod predators contributes to the stability of predator-prey population dynamics. Oikos 1999, 87, 603–608. [Google Scholar] [CrossRef]
- Castañé, C.; Arnó, J.; Gabarra, R.; Alomar, O. Plant damage to vegetable crops by zoophytophagous mirid predators. Biol. Control 2011, 59, 22–29. [Google Scholar] [CrossRef]
- Perdikis, D.; Lykouressis, D. Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Control 2000, 17, 55–60. [Google Scholar] [CrossRef]
- Biondi, A.; Zappalà, L.; Di Mauro, A.; Tropea Garzia, G.; Russo, A.; Desneux, N.; et al. Can alternative host plant and prey affect phytophagy and biological control by the zoophytophagous mirid Nesidiocoris tenuis? BioControl 2016, 61, 79–90. [Google Scholar] [CrossRef]
- Nakano, R.; Morita, T.; Okamoto, Y.; Fujiwara, A.; Yamanaka, T.; Adachi-Hagimori, T. Cleome hassleriana plants fully support the development and reproduction of Nesidiocoris tenuis. BioControl 2021, 66, 407–418. [Google Scholar] [CrossRef]
- Sánchez, J.A.; Lacasam, A. Impact of the zoophytophagous plant bug Nesidiocoris tenuis (Heteroptera: Miridae) on tomato yield. J. Econ. Entomol 2008, 101, 1864–1870. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.A. Density thresholds for Nesidiocoris tenuis (Heteroptera: Miridae) in tomato crops. Biol. Control 2009, 51, 493–498. [Google Scholar] [CrossRef]
- Calvo, J.; Bolckmansm, K.; Stansly, P.A.; Urbaneja, A. Predation by Nesidiocoris tenuis on Bemisia tabaci and injury to tomato. BioControl 2009, 54, 237–246. [Google Scholar] [CrossRef]
- Souto, P.; Abraços-Duarte, G.; da Silva, E.B.; Figueiredo, E. Half Friend, Half Enemy? Comparative phytophagy between two Dicyphini species (Hemiptera: Miridae). Insects 2022, 13, 175. [Google Scholar] [CrossRef] [PubMed]
- Moerkens, R.; Pekas, A.; Bellinkx, S.; Hanssen, I.; Huysmans, M.; Bosmans, L.; et al. Nesidiocoris tenuis as a pest in Northwest Europe: Intervention threshold and influence of Pepino mosaic virus. J. Appl. Entomol. 2020, 144, 566–77. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Urbaneja, A. The zoophytophagous predator Nesidiocoris tenuis: a successful but controversial biocontrol agent in tomato crops. In: Advances in Insect Control and Resistance Management, Horowitz, A., Ishaaya, I., Eds.; Springer, Cham, Switzerland, 2016, pp. 121–138.
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; et al. Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Biondi, A.; Guedes, R.N.C.; Wan, F.H.; Desneux, N. Ecology, worldwide spread, and management of the invasive South American tomato pinworm, Tuta absoluta: Past, Present, and Future. Annu. Rev. Entomol. 2018, 63, 239–58. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, H.Á.A.; Guedes, R.N.C.; Picanço, M.C. Insecticide resistance in populations of Tuta absoluta (Lepidoptera: Gelechiidae). Agric. For. Entomol. 2000, 2, 147–153. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; et al. Insecticide resistance in the tomato pinworm Tuta absoluta: patterns, spread, mechanisms, management and outlook. J. Pest Sci. 2019, 92, 1329–42. [Google Scholar] [CrossRef]
- Silva, G.A.; Picanço, M.C.; Bacci, L.; Crespo, A.L.B.; Rosado, J.F.; Guedes, R.N.C. Control failure likelihood and spatial dependence of insecticide resistance in the tomato pinworm, Tuta absoluta. Pest Manag. Sci. 2011, 67, 913–20. [Google Scholar] [CrossRef] [PubMed]
- Prasannakumar, N.R.; Jyothi, N.; Saroja, S.; Kumar, G.R. Relative toxicity and insecticide resistance of different field population of tomato leaf miner, Tuta absoluta (Meyrick). Int. J. Trop. Insect Sci. 2021, 41, 1397–405. [Google Scholar] [CrossRef]
- Roditakis, E.; Vasakis, E.; García-Vidal, L.; et al. A four-year survey on insecticide resistance and likelihood of chemical control failure for tomato leaf miner Tuta absoluta in the European/Asian region. J. Pest Sci. 2018, 91, 421–35. [Google Scholar] [CrossRef]
- Holling, C.S. The Components of predation as revealed by a study of small-mammal predation of the European pine sawfly. Can Entomol. 1959, 91, 293–320. [Google Scholar] [CrossRef]
- Hassell, M.P.; Lawton, J.H.; Beddington, J.R. Sigmoid functional responses by invertebrate predators and parasitoids. J. Anim. Ecol. 1977, 46, 249–62. [Google Scholar] [CrossRef]
- Holling, CS. The functional response of predators to prey density and its role in mimicry and population regulation. Mem. Ent. Soc. Can. 1965, 97, 5–60. [Google Scholar] [CrossRef]
- Hamdan, A.J.S. Functional and numerical responses of the predatory bug Macrolophus caliginosus Wagner fed on different densities of eggs of the greenhouse whitefly, Trialeurodes vaporariorum (Westwood). J. Biol. Res. 2006, 6, 147–54. [Google Scholar]
- Sharifian, I.; Sabahi, Q.; Khoshabi, J. Functional response of Macrolophus pygmaeus (Rambur) and Nesidiocoris tenuis (Reuter) feeding on two different prey species. Arch. Phytopathol. Plant Prot. 2015, 48, 910–20. [Google Scholar] [CrossRef]
- Michaelides, G.; Sfenthourakis, S.; Pitsillou, M.; Seraphides, N. Functional response and multiple predator effects of two generalist predators preying on Tuta absoluta eggs. Pest Manag. Sci. 2018, 74, 332–9. [Google Scholar] [CrossRef] [PubMed]
- van Alphen, J.J.M; Jervis, M.A. Foraging Behaviour. In: Insect Natural Enemies, Jervis, M., Kidd, N., Eds; Springer, Dordrecht, Netherlands, 1996, pp. 1–62.
- Foglar, H.; Malausa, C. The Functional response and preference of Macrolophus caliginosus [Heteroptera : Miridae] for two of its prey: Myzus persicae and Tetranychus urticae. Entomophaga 1990, 35, 465–74. [Google Scholar] [CrossRef]
- Montserrat, M.; Albajes, R.; Castane, C. Functional response of four Heteropteran predators preying on greenhouse whitefly (Homoptera: Aleyrodidae) and western flower thrips (Thysanoptera: Thripidae). Environ Entomol. 2000, 29, 1075–82. [Google Scholar] [CrossRef]
- Enkegaard, A.; Brødsgaard, H.F.; Hansen, D.L. ; Macrolophus caliginosus: Functional response to whiteflies and preference and switching capacity between whiteflies and spider mites. Entomol. Exp. Appl. 2001, 101, 81–8. [Google Scholar] [CrossRef]
- Panagakis, S.; Perdikis, D.; Fantinou, A. Functional response analysis of predation by an omnivore predator: effect of hunger level and sex. In Proceedings of the IOBC/WPRS Working Group "Integrated Control in Protected Crops", Crete, Greece, 6-11 September 2009. [Google Scholar]
- Maselou, D.; Perdikis, D.; Fantinou, A. Effect of hunger level on prey consumption and functional response of the predator Macrolophus pygmaeus. Bull. Insectology. 2015, 68, 211–8. [Google Scholar]
- Baños, H.L.; Ruiz Gil, T.; del Toro Benitez, M.; Miranda Cabrera, I. Consumption and functional response of Nesidiocoris tenuis Reuter (Hemiptera: Miridae) feeding on immature stages of Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae). Rev. Prot. Veg. 2016, 31, 217–23. [Google Scholar]
- Holling, C.S. Principles of insect predation. Annu. Rev. Entomol. 1961, 6, 163–182. [Google Scholar] [CrossRef]
- Rosenbaum, B.; Rall, B.C. Fitting functional responses: Direct parameter estimation by simulating differential equations. Methods Ecol. Evol. 2018, 9, 2076–2090. [Google Scholar] [CrossRef]
- Real, L.A. Ecological determinants of functional response. Ecology 1979, 60, 481–485. [Google Scholar] [CrossRef]
- Juliano, S. A.; Goughnour, J. A.; &Ower, G. D.; &Ower, G. D. Predation in many dimensions: spatial context is important for meaningful functional response experiments. Front. ecol. evol. 2022, 10, 845560. [Google Scholar] [CrossRef]
- Montserrat, M.; Albajes, R.; Castañé, C. Functional response of four heteropteran predators preying on greenhouse whitefly (Homoptera: Aleyrodidae) and western flower thrips (Thysanoptera: Thripidae). Environ. Entomol. 2000, 29, 1075–1082. [Google Scholar] [CrossRef]
- Enkegaard, A.; Brødsgaard, H.F.; Hansen, D.L. Macrolophus caliginosus: Functional response to whiteflies and preference and switching capacity between whiteflies and spider mites. Entomol. Exp. Appl. 2001, 101, 81–8. [Google Scholar] [CrossRef]
- Hassanpour, M.; Bagheri, M.; Golizadeh, A.; Farrokhi, S. Functional response of Nesidiocoris tenuis (Hemiptera: Miridae) to Trialeurodes vaporariorum (Hemiptera: Aleyrodidae): effect of different host plants. Biocontrol Sci. Technol. 2016, 26, 1489–503. [Google Scholar] [CrossRef]
- Gavkare, O.; Sharma, P.L.; Sanchez, J.A.; Shah, M.A. Functional response of Nesidiocoris tenuis (Hemiptera: Miridae) to the two-spotted spider mite, Tetranychus urticae. Biocontrol Sci. Technol. 2017, 27, 1118–22. [Google Scholar] [CrossRef]
- Fathipour, Y.; Karimi, M.; Farazmand, A.; Talebi, A.A. Age-specific functional response and predation rate of Amblyseius swirskii (Phytoseiidae) on two-spotted spider mite. Syst. Appl. Acarol. 2017, 22, 159–69. [Google Scholar] [CrossRef]
- Nikbin, R.; Sahragard, A.; Hosseini, M. Age-specific functional response of Trichogramma brassicae (Hymenoptera: Trichogrammatidae) parasitizing different egg densities of Ephestia kuehniella (Lepidoptera: Pyralidae). J. Agric. Sci. Technol. 2014, 16, 1217–27. [Google Scholar]
- Ingegno, B.L.; Messelink, G.J.; Bodino, N.; et al. Functional response of the mirid predators Dicyphus bolivari and Dicyphus errans and their efficacy as biological control agents of Tuta absoluta on tomato. J. Pest Sci. 2019, 92, 1457–1466. [Google Scholar] [CrossRef]
- Perdikis, D.C.; Lykouressis, D.P.; Economou, L.P. The influence of temperature, photoperiod and plant type on the predation rate of Macrolophus pygmaeus on Myzus persicae. BioControl 1999, 44, 281–289. [Google Scholar] [CrossRef]
- Hughes, G.E.; Alford, L.; Sterk, G.; Bale, J.S. Thermal activity thresholds of the predatory mirid Nesidiocoris tenuis: Implications for its efficacy as a biological control agent. BioControl 2010, 55, 493–501. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Lacasa, A.; Arnó, J.; Castañé, C.; Alomar, O. Life history parameters for Nesidiocoris tenuis (Reuter) (Het., Miridae) under different temperature regimes. J. Appl. Entomol. 2009, 133, 125–32. [Google Scholar] [CrossRef]
- Ingegno, B.L.; Messelink, G.J.; Leman, A.; Sacco, D.; Tavella, L. Development and thermal activity thresholds of European mirid predatory bugs. Biol. Control 2021, 152. [Google Scholar] [CrossRef]
- Uiterwaal, S.F.; DeLong, J.P. Functional responses are maximized at intermediate temperatures. Ecology 2020, 101. [Google Scholar] [CrossRef] [PubMed]
- Madbouni, M. A. Z.; Samih, M. A.; Namvar, P.; Biondi, A. Temperature-dependent functional response of Nesidiocoris tenuis (Hemiptera: Miridae) to different densities of pupae of cotton whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). Eur. J. Entomol. 2017, 114, 325. [Google Scholar] [CrossRef]
- Dorsaz, M.; Fischer, S.; Baroffio, C.A. Study of the temperature influence on the functional response of the biological control agent Macrolophus pygmaeus. In Proceedings of the Les Cochenilles: ravageur principal ou secondaire 9ème Conférence Internationale sur les Ravageurs en Agriculture, SupAgro, Montpellier, France, 25-27 October 2011. [Google Scholar]
Figure 1.
Functional response of 1 day and 10-day old females (open circles) and males (closed circles) of M. pygmaeus on eggs of T. absoluta at 20 and 25°C.
Figure 1.
Functional response of 1 day and 10-day old females (open circles) and males (closed circles) of M. pygmaeus on eggs of T. absoluta at 20 and 25°C.
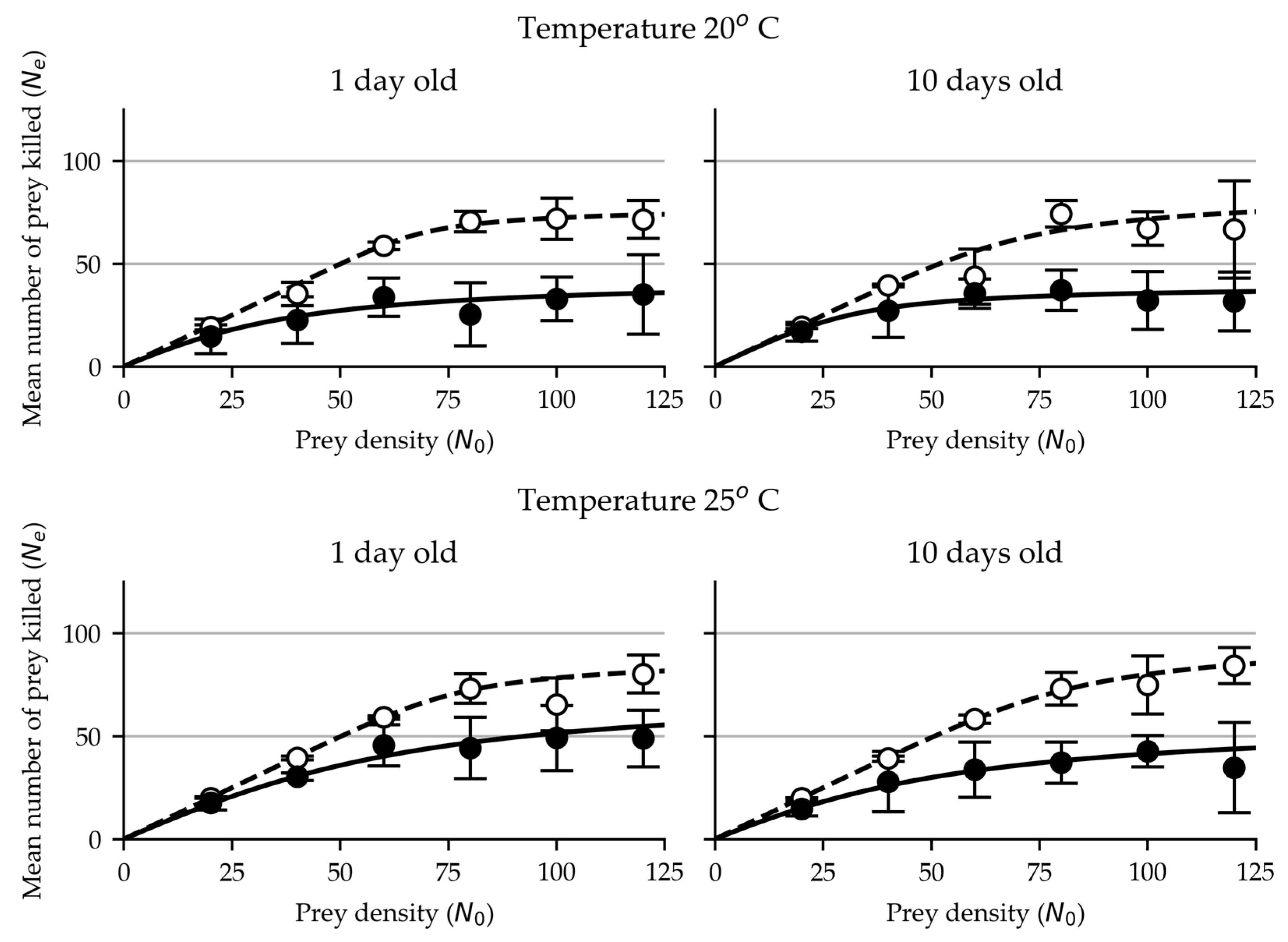
Figure 2.
Functional response of 1 day and 10-day old females (open circles) and males (closed circles) of N. tenuis on eggs of T. absoluta at 20 and 25°C.
Figure 2.
Functional response of 1 day and 10-day old females (open circles) and males (closed circles) of N. tenuis on eggs of T. absoluta at 20 and 25°C.
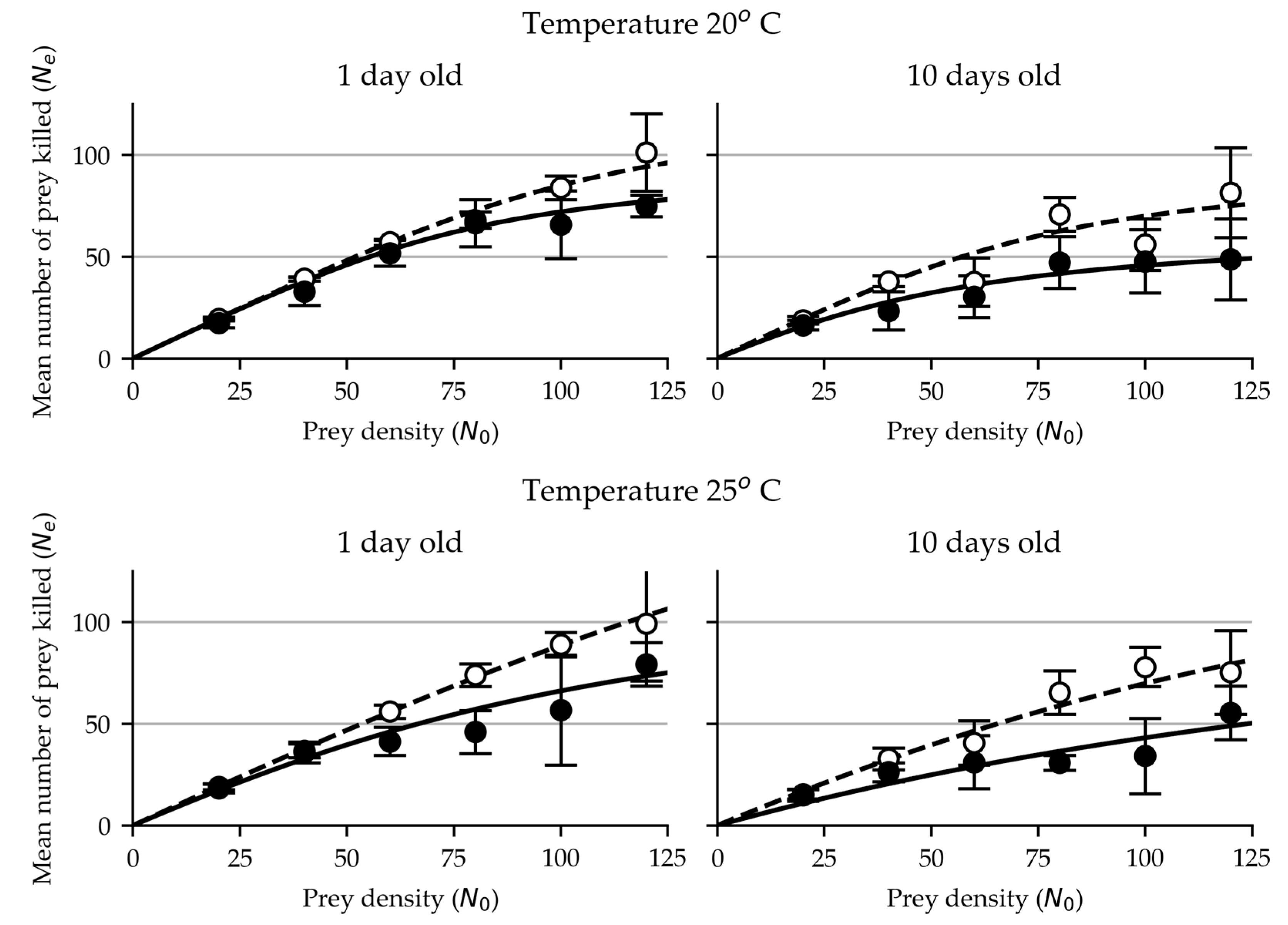
Table 1.
Effect of sex, adult age and temperature on the attack rate (a) and handling time (Th) of M. pygmaeus and N. tenuis adults feeding on eggs of T. absoluta on tomato leaflets.
Table 1.
Effect of sex, adult age and temperature on the attack rate (a) and handling time (Th) of M. pygmaeus and N. tenuis adults feeding on eggs of T. absoluta on tomato leaflets.
| Species | Sex* | Age (days) | Temperature (°C) |
a (±SE)(hrs-1) |
Th ( ±SE) (hrs) |
R2 |
|---|---|---|---|---|---|---|
| f | 1 | 20 | 0.70±0.26 | 0.31±0.02 | 0.99 | |
| f | 1 | 25 | 0.51±0.12 | 0.27±0.02 | 0.83 | |
| f | 10 | 20 | 0.33±0.09 | 0.28±0.03 | 0.92 | |
| f | 10 | 25 | 0.36±0.04 | 0.24±0.01 | 0.99 | |
| M. pygmaeus | m | 1 | 20 | 0.09±0.04 | 0.57±0.08 | 0.81 |
| m | 1 | 25 | 0.10±0.01 | 0.33±0.03 | 0.93 | |
| m | 10 | 20 | 0.19±0.08 | 0.61±0.05 | 0.71 | |
| m | 10 | 25 | 0.08±0.01 | 0.42±0.03 | 0.88 | |
| f | 1 | 20 | 0.22±0.02 | 0.18±0.02 | 0.99 | |
| f | 1 | 25 | 0.14±0.01 | 0.10±0.01 | 0.99 | |
| f | 10 | 20 | 0.18±0.07 | 0.25±0.06 | 0.87 | |
| N. tenuis | f | 10 | 25 | 0.09±0.01 | 0.14±0.03 | 0.96 |
| m | 1 | 20 | 0.20±0.07 | 0.24±0.03 | 0.98 | |
| m | 1 | 25 | 0.10±0.03 | 0.19±0.06 | 0.88 | |
| m | 10 | 20 | 0.09±0.01 | 0.37±0.05 | 0.93 | |
| m | 10 | 25 | 0.04±0.01 | 0.19±0.11 | 0.88 | |
*f=female, m=male.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



