
Submitted:
17 April 2024
Posted:
18 April 2024
You are already at the latest version
Abstract
Amylose content (AC) stands as a pivotal determinant of rice grain quality, primarily governed by the Waxy gene (Wx), with its allelic diversity significantly influencing AC variation. In the current culinary landscape, soft rice varieties, known for their low AC and superior palatability, are particularly sought after by consumers. Notably, many soft japonica rice varieties cultivated in southern China, including the Nangeng series, are characterized by the presence of the Wxmp allele, derived from the ancestral Wxmq allele. While both alleles are associated with reduced AC, there remains a paucity of detailed understanding regarding the interplay between specific functional single nucleotide polymorphisms (SNPs) within these alleles and the overarching rice grain quality. Employing site-directed mutagenesis, we engineered three distinct transgenic rice lines, each harboring either the Wxmp, Wxmq, or Wxb-5c alleles, against the background of the glutinous rice cultivar Nip(wx). This suite of transgenic rice showcased a gradation ingrain transparency inversely correlated to AC, delineating a sequence from Wxmq, signifying the lowest, through Wxmp, to Wxb-5c, indicating the highest. Consequently, this gradation in AC precipitated variations in other rice grain physicochemical attributes, such as the taste value of cooked rice, gel consistency (GC), and starch pasting properties. Furthermore, analyses of gene expression and enzyme activity revealed that the functional SNPs, Ex4-53G/A and Ex5-53T/C, lead to a decline in the GBSSI enzyme activity without affecting expression levels. These findings underscore the potential of harnessing these SNPs in breeding programs aimed at developing soft rice varieties endowed with enhanced culinary qualities.
Keywords:
soft rice
; low amylose
; grain transparency
; Wx
; eating and cooking quality
1. Introduction
The endosperm constitutes the edible portion of rice grain, with starch accounting for over 80% of its composition, thus rendering it the cornerstone of rice’s physicochemical properties critical to assessing eating quality [1]. Starch in rice is bifurcated into two primary types: branched amylopectin, which forms the bulk of the starch content, and amylose, characterized by its long-chain linear glucose polymers. In glutinous rice, amylopectin virtually constitutes the entirety of its starch content. Conversely, amylose emerges as a pivotal component in non-glutinous rice, with AC critically influencing the grain’s quality attributes [2]. Typically, rice with elevated AC yields a firmer texture upon cooking, whereas varieties with diminished AC are noted for their softness and stickiness [3]. Thus, varieties manifesting AC levels ranging from 8% to 13%, bridging the gap between glutinous and sticky rice, are often categorized as soft rice. These varieties are distinguished by their tender and cohesive texture, coupled with superior taste and palatability [4].
The Granule-Bound Starch Synthase I (GBSSI), encoded by the Wx gene, plays a crucial role in amylose synthesis within rice endosperm. The genetic diversity in AC across rice varieties is closely linked to allelic variations within the Wx gene, underscoring the gene’s critical influence on rice quality [5]. To date, researchers have identified at least nine Wx alleles, namely Wxlv, Wxa, Wxin, Wxb, Wxmw, Wxmp, Wxmq, Wxop, and wx, each contributing uniquely to the starch composition of rice [6]. The Wxmp allele, in particular, has gained prominence in the breeding of soft rice varieties in south China, leading to the popularization of japonica varieties such as Nangeng9108, and Huruan 1212. Originating from the low-AC japonica cultivar Milky Princess, the Wxmp allele is characterized by a G to A mutation in exon 4 (Ex4-53A), resulting in the substitution of Arg158 by His158, distinguishing it from the Wxb allele [7]. Milky Queen, a sibling line of Milky Princess, exhibits an additional mutation, featuring the Wxmq allele, a T to C mutation resulting in the substitution of Tyr191 by His191 in exon 5 (Ex5-52C), further diversifying the allelic variations from Wxb [8]. This nuanced genetic variation has, at times, led to confusion among breeders regarding the distinct contributions of the Wxmp and Wxmq alleles to rice grain quality. The dynamic interaction between these two functional alleles and their synergistic influence on rice grain phenotypes still requires thorough investigation for a clearer understanding.
In this study, we engineered complementary vectors encapsulating the complete genomic DNA sequences of the Wxmp (Ex4-53A), Wxmq (Ex4-53A+Ex5-52C), and Wxb-5c (Ex5-52C) alleles. These vectors were then introduced into the glutinous rice variety Nip(wx), creating a near-isogenic line (NIL) devoid of the wx allele in the japonica rice Nipponbare (NIP) background, which naturally carries the Wxb allele (Figures. 1A, 1B, 1S). Through a systematic evaluation of the grain quality attributes of the resulting homozygous transgenic rice lines, we elucidated the impacts of the two functional sites on amylose synthesis, grain morphology, the sensory qualities of cooked rice, and the physicochemical properties of starch. This research furnishes valuable genetic materials and lays a conceptual foundation for the development and enhancement of rice varieties characterized by moderate AC and superior culinary quality.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
The experimental materials used in this study comprised the japonica cultivar Nipponbare (Nip), which harbors the Wxb allele, and its near-isogenic line (NIL) within the Nip background, designated Nip(wx) and containing the null wx allele. Further, transgenic rice lines -Nip(wx)-Wxmp, Nip(wx)-Wxb-5c, and Nip(wx)-Wxmq- were generated against the Nip(wx) background. These specified rice lines were cultivated at the experimental farm of Yangzhou University (Jiangsu Province, China, 32°23′N). The experimental design included planting 20 seedlings for each strain, with the replication of three biological repeats. Consistent field management practices were implemented across all variants.
2.2. Plasmids and Rice Transformation
PCR amplification was conducted using genomic DNA from the Japonica cultivars Nip and NJ46 (carrying the Wxmp allele) as templates. This process employed primers QWx-1, QWx-2, QWx-3, and QWx-4 (Table S1), yielding two distinct fragments corresponding to the Wxb and Wxmp alleles, with lengths of 4 kb and 4.4 kb, respectively (Fig. S1). The PCR reactions were performed in a 50-µL volume using PrimeSTAR® HS DNA Polymerase (TaKaRa, Kyoto, Japan). The amplification conditions were as follows: an initial denaturation at 94 °C for 5 min, followed by 36 cycles of denaturation at 98 °C for 10 s and annealing/extension at 68 °C for 4 min, concluding with a final extension at 72 °C for 5 min. For the generation of the Wxb-5c and Wxmq expression vectors, genomic DNA from Nip and NJ46 was used as templates. Initially, the 1st DNA segment was amplified with primers QWx-1/QWx-2 from both Nip and NJ46 genomic DNA separately. Subsequently, a DNA segment bearing a T to C mutation in exon 5 was amplified using NJ46 genomic DNA and primers QWx-3/QWx-5C-R and QWx-5C-F/QWx-4 (Table S1), resulting in the 2nd DNA segment. This DNA segment was then ligated with the 1st DNA segment amplified from Nip to assemble the Wxb-5c segments. Similarly, the 2nd DNA segment was ligated with the 1st DNA segment amplified from NJ46 to construct the Wxmq allele. The resultant PCR products were cloned into the pCAMBIA1300 vector (Cambia, Canberra, Australia). The constructed plasmids, namely PC1300-Wxmp, PC1300-Wxb-5c, and PC1300-Wxmq (Fig. 1B), were introduced into Agrobacterium tumefaciens strain EHA105. This was followed by the transformation into Nip(wx) via agrobacterium-mediated transformation [9]. The identification of transgene copy number was performed according to the method established by Ding et al. (2004) [10], employing 10 ng of genomic DNA as a PCR template along with Wx-specific primers and an endogenous single-copy molecular marker (Table S1).
2.3. Observation of Grain Transparency
To evaluate grain transparency, photographs of various samples of white rice were captured under transmitted light conditions. These color images were subsequently converted into black and white for analysis. The transparency rate (%) was determined by calculating the ratio of the average grain grayscale value of the image background. The analysis was performed using the software ImageJ (available at https://imagej.net/), following the methodology described by Zhang et al. (2019) [11].
2.4. Examination of Cavities within Starch Granules
To observe the internal structure of starch granules, rice grains were subjected to rapid freezing in liquid nitrogen. Subsequently, these frozen samples were cut to produce cross-sections for microscopic examination. The prepared cross-sections were affixed to aluminum specimen stubs using dual adhesive tabs, followed by sputter-coating with gold to enhance electron conductivity and image clarity. The samples were then examined under an S-4800 environmental scanning electron microscopy (SEM) instrument (Hitachi, Tokyo, Japan) to visualize the microstructural details of the starch granules.
2.5. Grain Quality Analyses
The assessment of grain quality encompassed several critical parameters, including the apparent amylose content (AAC), gel consistency (GC) of rice flour, as described by Zhang et al. [12]. The evaluation of rice taste value was conducted using the Cooked Rice Taste Analyzer (CRTA) Model STA1B (Sake, Hiroshima, Japan). In this process, approximately 30 g of milled rice grains were submerged in water within an aluminum container for a duration of 30 min, subsequently rinsed three times with cold water. The rice grains were then subjected to steaming at a rice to water weight ratio (w/w) of 1:1.2 for 30 min, thereafter maintained in a warmed state for 10 min prior to being cooled to ambient temperature.
2.6. Measurement of the Gelatinization and Pasting Properties of Rice
The determination of gelatinization temperatures was executed employing a differential scanning calorimeter (DSC) model 200 F3 (Netzsch Instruments North America Llc, Burlington, MA, USA). The analysis yielded DSC curves that elucidated various parameters such as onset (To), peak (Tp), and conclusion (Tc) temperatures, along with the enthalpy of gelatinization (ΔH), which indicates the thermal energy required for the gelatinization process. Furthermore, the pasting properties of rice were determined using a Rapid Visco-Analyzer (RVA) (Techmaster, Newport Scientific, Warriewood, Australia) [12].
2.7. Gel Permeation Chromatograms
Isoamylase (EC 3.2.1.68, E-ISAMY) (Megazyme, Kilruddery, Ireland) was employed to debranch purified rice starch. Subsequently, the relative molecular weight distribution was assessed using gel permeation chromatography (GPC) on a PL-GPC 220 system (Polymer Laboratories Varian Inc., Amherst, MA, USA), following the method described by Zhu et al. [29]. The GPC data were analyzed using integral equations to construct molecular weight distribution curves, employing standard dextran samples with molecular weights of 2,800, 18,500, 111,900, 410,000, 1,050,000, 2,900,000, and 6,300,000. To facilitate comparison among amylopectin (AP), its short chains (AP1), long chains (AP2), and amylose (AM), two replicate measurements were performed.
2.8. Crystalline Structure Analysis
Crystalline structure analysis was performed to examine the supramolecular structure of rice starch. X-ray powder diffraction (XRD) measurements were conducted using a D8 ADVANCE X-ray diffractometer (Bruker AXS, Karlsruhe, Germany), and the relative crystallinity was quantified according to the methodology outlined by Cai et al. [13]. Additionally, to explore the short-range molecular order near the surface of starch granules, Fourier-transform infrared spectroscopy (FTIR) was employed using a Varian 7000 FTIR spectrometer (PerkinElmer, Wellesley, MA, USA), following the approach described by Zhang et al. [12]. Absorbance values at 1047 cm−1 and 1022 cm−1 were extracted and corrected from the FTIR spectra.
2.9. RNA Extraction and Quantitative Reverse Transcriptase PCR Analysis
Total RNAs were extracted from the developing seeds 15 days after flowering (DAF) using the RNAprep Pure Plant Kit (Tiangen, Beijing, China), in accordance with the instructions provided by the manufacturer. The total RNA was reverse transcribed using oligo (dT18) primers as per the protocol of the Vazyme reverse transcription kit (Vazyme, Nanjing, China). Quantitative reverse transcriptase PCR (RT-qPCR) analyses were performed using SYBR Green Real-Time PCR Master Mixes (ThermoFisher Scientific, Waltham, MA, USA). For each sample, biological triplicates were prepared, and each biological replicate was further subjected to technical triplicates to ensure precision. The expression level of each gene was quantified by averaging the value obtained from these replicates.
2.10. Enzyme Activity Assays
The enzyme activity of Granule-Bound Starch Synthase I (GBSSI) was assessed according to the methodology delineated by Liu et al. [14]. Seeds harvested at 15 DAF were deprived of hull, embryo, and pericarp and subsequently ground into a fine powder under liquid nitrogen. The activity of GBSSI was quantitatively defined as the amount of NADPH generated per minute per gram of the sample, with one unit of enzyme activity being equivalent to 1 nmol of NADPH.
2.11. Western Blot Assay
For the Western blot analysis, immature seeds collected at 15 DAF were pulverized in liquid nitrogen to obtain a fine powder. Protein extraction was achieved by homogenizing the seed powder with an extraction buffer consisting of 125 mmol/L Tris-HCl, pH 6.8, 4 mol/L urea, 4% SDS, and 5% 2-mercaptoethanol at a ratio of 1:15 (15 μL of buffer per 1 mg of powder), followed by incubation at 37 °C for 3 h. The extracted proteins were then separated via SDS−PAGE and electrotransferred onto polyvinylidene difluoride (PVDF) membranes. Subsequent incubation steps were performed using antibodies specifically targeting GBSSI and HSP to detect the respective proteins.
2.12. Statistical Analysis
Each test was performed independently, in triplicate, unless otherwise specified, and the data were reported as mean ± standard deviation. Statistical significance was analyzed with a one-way ANOVA followed by Duncan’s test at p < 0.05 (SPSS 21.0, SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Effects of Two Functional Single Nucleotide Polymorphisms on Seed Transparency
Agronomic trait evaluation revealed uniformity in plant architecture and grain dimensions across the transgenic lines, yet unveiled disparities in grain transparency (Figure 1C, S2). Intricately, Nip(wx)-Wxmq grains displayed a notably darker endosperm compared to Nip(wx)-Wxmp. The Wxb-5c line, while showing a subtly darker endosperm relative to Nip (harboring the Wxb allele), demonstrated a clear enhancement in grain transparency against Nip(wx)-Wxmp. A quantitative analysis of grain transparency mirrored this observation, unveiling a progressive transparency gradient following the sequence: Nip(wx) < Nip(wx)-Wxmq < Nip(wx)-Wxmp < Nip(wx)-Wxb-5c < Nip(Wxb) (Figure 1D). This gradation underscores the intricate link between grain transparency and the presence of microscopic voids within the endosperm’s starch granules [12].To further elucidate the microstructural characteristics of rice grains, the cross-sections of their kernels were further examined using a SEM. This exploration revealed a distinct architecture of starch granules in the Nip(Wxb) endosperm, which is compactly and symmetrically organized, devoid of any discernible voids. In stark contrast, the starch granules of the other four rice variants were marred by the presence of conspicuous, diminutive cavities, the extent and frequency of which varied markedly across the samples. Particularly, the cavities within the starch granules from Nip(wx)-Wxmp rice were notably larger compared to those in Nip(wx)-Wxb-5c. Meanwhile, the quantity and dimensions of the cavities within the starch granules of Nip(wx)-Wxmq bore a resemblance to those observed in Nip(wx) (Figure 1E). These intragranular voids have been substantiated to influence the transparency of the grain [15], aligning with our observations of the variance in grain transparency across these transgenic rice lines.
3.2. Effect of the Distinct Transgenic Rice Lines on Rice Eating Quality
Our investigation extended to the analysis of the principal constituents and physicochemical characteristics of rice grain. Notably, the glutinous variety, Nip(wx), exhibited an exceptionally low AC of approximately 3%. In comparison, the Nip(Wxb) and Nip(wx)-Wxb-5c variants demonstrated a modest AC of about 13%, whereas the Nip(wx)-Wxmp line presented an AC of approximately 10%. The Nip(wx)-Wxmq line among the trio of transgenic lines emerged with the lowest AC of about 6% (Figure 2A). Subsequent assessments of GC, an indicator of texture, revealed that Nip(wx) was the softest, which was sequentially followed by Nip(wx)-Wxmq, Nip(wx)-Wxmp, Nip(wx)-Wxb-5c, and Nip(Wxb) (Figure 2B). Intriguingly, these findings corroborate the established inverse relationship between GC and AC [12]. Moreover, the softer GC of the transgenic rice lines underscored a marked enhancement in their culinary appeal. To quantitatively gauge this enhancement, the taste value of the cooked rice was meticulously assessed. It emerged that all the transgenic variants transcended Nip(Wxb) in taste value, with Nip(wx)-Wxmq rice achieving a notably superior taste profile compared to both Nip(wx)-Wxmp and Nip(wx)-Wxb-5c rice (Figure 2C). This comprehensive analysis unveils the instrumental role of the functional mutations at Ex4-53A and Ex5-52C in modulating amylose synthesis, GC, and, ultimately, the taste value of cooked rice. The synergistic interplay between these mutations, particularly evident in the Wxmq allele, significantly accentuates the AC variation, enriching the culinary quality of these genotypes.
The viscosity of rice starch stands as another important indicator in the appraisal of rice’s culinary merit. To illuminate this aspect, we conducted an RVA analysis of flours derived from various transgenic rice lines. The RVA profiles, as shown in Figure 2D, delineated considerable diversity across the samples. Specifically, the Nip(wx) rice exhibited a prototypical glutinous RVA trajectory, characterized by an early onset of gelatinization and the lowest final viscosity among the rice lines examined. Intriguingly, the Nip(wx)-Wxmq rice mirrored the RVA pattern of Nip(wx) rice, albeit with a discernible shift towards the right, manifesting in elevated peak viscosity, hot paste viscosity, and final viscosity metrics. In addition, the RVA curves of Nip(wx)-Wxmp and Nip(wx)-Wxb-5c rice are situated between Nip(Wxb) and Nip(wx) rice. Despite the resemblance in their RVA curve contours, Nip(wx)-Wxmp rice distinguished itself with a marginally higher peak viscosity and breakdown values, albeit with a reduced final viscosity when compared with Nip(wx)-Wxb-5c rice (Table 1). These findings lend credence to the hypothesis that rice varieties with diminished AC display a greater susceptibility to gelatinization, as evidenced by leftward shifts in their pasting curves. This observation concurs with our preceding investigations [12], further substantiating the correlation between AC and starch viscosity’s role in defining the gastronomic quality of rice.
Additionally, the gelatinization properties of rice flours were characterized using the DSC. The gelatinization curves obtained, as depicted in Figure 2E, demonstrated a general uniformity across all samples, albeit with the glutinous Nip(wx) rice displaying a slight shift to the right in its curve. In terms of the gelatinization parameters, there was a consensus among the transgenic rice lines in terms of both onset temperature (To) and peak temperature (Tp); however, variations were observed in the enthalpy of gelatinization (ΔHgel) (Table 2). Prior research has indicated that the enthalpy of starch gelatinization reflects the thermal energy required to disrupt the semi-crystalline structure of starch molecules, predominantly constituted by amylopectin [16]. It is generally established that an increase in amylopectin content correlates with higher crystallinity in rice starch. It is therefore plausible that the glutinous Nip(wx) rice, presumably with a higher amylopectin content, exhibited the highest ΔHgel, followed in descending order by Nip(wx)-Wxmq, Nip(wx)-Wxmp, Nip(wx)-Wxb-5c, and Nip(Wxb). These findings are consistent with previously reported observations that there exists an inverse relationship between the enthalpy of gelatinization and the AC in rice grains [12].
3.3. Comparison of Starch Crystalline Structure in Transgenic Rice
Rice starch is a semi-crystalline complex comprising amylopectin and amylose, with its physicochemical properties intricately linked to its fine structure. Analysis of the fine structure is pivotal to understanding the variances in taste and quality among different rice varieties. The relative molecular weight distribution of starch in various transgenic lines was assessed using high-temperature GPC. As shown in Figure 3A, the starch from Nip(wx) exhibited only two fractions: the branched short chain component (AP1) and the branched long chain component (AP2). In contrast, starch from other samples showed, in addition to two amylopectin peaks, a distinct amylose peak (AM). It has been shown that the AP1 component of amylopectin consists of low-molecular-weight amylopectin A and short B chains (A+B1 chains), while the AP2 component contains high-molecular-weight long B chains [17]. Significant variations in the amylopectin to amylose ratios were observed across different transgenic lines, with notably lower AM values in Nip(wx)-Wxmq compared to Nip(wx)-Wxmp and Nip(wx)-Wxb-5c (Table 3). GPC analysis identified the AM components as indicative of the actual straight-chain starch content within the starch matrix. This finding implies that both single nucleotide polymorphisms (SNPs) Ex4-53G/A and Ex5-53T/C can differentially reduce the presence of straight-chain starch in rice, albeit with varying effects. Moreover, the combined impact of these two genetic loci significantly augments the observed AC disparity.
The degree of long-range order in starch granules from various transgenic lines was quantitatively assessed and evaluated using XRD. All examined samples exhibited pronounced diffraction peaks around diffraction angles of 15°, 17°, 18°, and 23°, characteristic of the A-type crystalline structure typically observed in cereal starches. Although the diffraction curves of samples from different transgenic lines were similar, crystallinity calculations indicated highly significant disparities (Figure 3B, Table 3). Specifically, the Nip(wx) sample demonstrated the highest crystallinity, followed by Nip(wx)-Wxmq, with Nip(Wxb) exhibiting the lowest. Previous studies have indicated a negative correlation between starch crystallinity and AC levels [13], and higher crystallinity levels necessitate more heat for the endothermic reaction [18], explaining the highest enthalpy value observed in Nip(wx) and the lowest in Nip(Wxb).
Further investigation into the short-range order of starches from the grains of different transgenic lines was conducted using attenuated total reflectance-Fourier transform infrared spectroscopy (ATR-FTIR). The results indicated that all samples yielded similar deconvoluted FTIR spectra. The analysis of parameters (1045/1022 cm–1) revealed that the degree of short-range order was the highest in the Nip(wx) sample, followed by Nip(wx)-Wxmq, with the lowest observed in Nip(Wxb) (Figure 3C, Table 3). These findings are consistent with the XRD analysis and corroborate our prior studies on the crystal structure of rice starch, which indicated variations in AAC resulting from alterations in GBSSI activity [12].\
3.4. Effects of Two Functional SNPs on Wx Gene Expression and GBSSI Enzyme Activity
To elucidate the impact of two functional mutations within the Wx gene on its expression and the activity of the GBSSI enzyme, we conducted a comparative analysis of Wx gene expression across three distinct transgenic lines. Gene expression was assessed in developing seeds at 15 DAF. The results revealed no significant variance in mRNA levels among the transgenic lines when compared to Nip(Wxb) rice. Notably, the glutinous Nip(wx) rice exhibited negligible expression (Figure 4A). These findings align with previous studies, suggesting that such SNP mutations do not substantially alter Wx gene expression, in agreement with some previous studies about the SNP mutation on Wx gene expression [7,19].
Subsequent assessments of GBSSI enzyme activity revealed that Nip(wx)-Wxmq exhibited the lowest activity, followed by Nip(wx)-Wxmp and Nip(wx)-Wxb-5c, in comparison to Nip(Wxb) rice (Figure 4B). Western blotting analyses of GBSSI corroborated these findings, showing a progressive decrease in GBSSI abundance in the order of Nip(Wxb) > Nip(wx)-Wxb-5c > Nip(wx)-Wxmp > Nip(wx)-Wxmq > Nip(wx), (Figure 4C) suggesting that the mutations not only reduce GBSSI enzyme activity but also impair its starch-binding capacity as previously documented [14]. Consequently, both the Ex4-53A and Ex5-52C mutations were observed to decrease GBSSI enzyme activity and abundance, leading to a reduction of AC in rice endosperm. Additionally, the results indicated that the Ex5-52C mutation exerts a minor effect on AC compared to the Ex4-53A mutation, suggesting its potential utility in rice breeding for optimizing grain transparency and culinary quality.
4. Discussion
In recent advancements, rice varieties with low AC, typically ranging from 8% to 12% and referred to as soft rice, have garnered popularity across China for their superior culinary quality. Notably, the soft rice varieties from the Nangeng series, which harbor the Wxmp allele, have achieved widespread cultivation in Jiangsu province, delivering substantial economic gains [4]. Beyond Wxmp and Wxmq alleles, Wxop/hp predominantly found in indica rice varieties, have been recognized for their contributions to lowering AC in rice [8,20]. Yet, in the context of japonica soft rice breeding programs within China, Wxmp has been the allele of choice. The Wxmq allele, despite its potential, has often been overlooked or confused with Wxmp, possibly due to its great impact on AC reduction. Prior research posited that the mutation at Ex4-53A could play a pivotal role in influencing AC in both Wxmp and Wxmq alleles, yet experimental validation has been scant [7]. Our research has substantiated that the Ex4-53A mutation exerts a more pronounced effect on AC and various grain quality attributes than the Ex5-52C mutation within the Wxb genetic background. Noteworthy are the findings that the Ex5-52C mutation, while having a marginal impact, does contribute to the attenuation of GBSSI enzyme activity. This modulation results in a moderately low AC and enhances grain appearance relative to the Wxmp allele. Additionally, the concurrent mutations at both Ex4-53A and Ex5-52C sites (Wxmq allele) manifest a more significant reduction in GBSSI activity, culminating in the lowest AC amongst the evaluated transgenic rice lines. Therefore, our study delineates the distinct impacts of the Ex4-53A and Ex5-52C mutations on the quality profile of rice grains, offering valuable insights for future breeding strategies aimed at optimizing AC and grain quality.
From the perspective of genetic regulation in rice cultivars, the control of the Wx gene, either directly or indirectly, can impact AC levels. Recent studies in rice have revealed the feasibility of creating novel Wx alleles to regulate AC. Genetic editing of cis-regulatory elements in the Wx promoter can be employed to fine-tune AC, thereby altering transparency [21,22]. Furthermore, nucleotide editing around the responsible sites for low AC alleles at the N-terminus of the GBSSI gene has resulted in a series of mutants with AC ranging from 1.4% to 11.9% [23], while editing nucleotides in the middle region of GBSSI has generated a range of new Wx alleles with AC levels from 0.3% to 29.43% [6]. The Ex4-53A and Ex5-52C in this study are both located in the region of low AC alleles at the N-terminus of GBSSI. Additionally, targeted deletion of the first intron of Wxb has increased AC levels, presenting a potential strategy to enhance transparency in low AC backgrounds [24].
In addition to mutations in the Wx gene sequence, the generation of AC in rice is influenced by multiple mechanisms: (i) the regulation of Wx gene expression; (ii) the efficiency of Wx pre-mRNA splicing; and (iii) changes in the expression or activity of other starch biosynthesis-related genes or proteins, such as soluble starch synthase (SSS) or starch branching enzyme (SBE), which alter the starch structure or the amylose/amylopectin ratio. Numerous studies have shown that transcription factors involved in regulating endosperm development often exhibit pleiotropy, concurrently influencing the expression of the Wx gene and other genes involved in starch synthesis. This interaction often leads to either downregulation or upregulation of these transcription factors, which can negatively impact other rice quality traits, such as chalkiness. Consequently, these genes are not directly utilizable for genetic modifications aimed at enhancing rice quality [6,25]. Furthermore, it has been observed that certain Dull variants in rice modify Wx expression and interact differently depending on the allelic genotype [26-28]. Typically, a reduced AC phenotype is only observed when the Wxb allelic gene is present, with AC generally falling below 12%, and the grains exhibiting either a semi-transparent or opaque appearance with a dull endosperm.
AC is a pivotal determinant in the transparency of rice grains. Varieties such as glutinous rice, which possess an extremely low AC, are characterized by an opaque waxy phenotype. Conversely, soft rice varieties with low AC often display a semi-transparent or opaque cloudy endosperm phenotype. Previous research has established a significant negative correlation between the number and size of pores within the starch granules of rice endosperm and the transparency of the rice [12,15]. This study corroborates these findings, observing a similar pattern in three distinct transgenic rice lines and the Nip(wx) endosperm, where large pores within the starch granules were particularly prominent in glutinous rice. Externally, the opaque endosperm phenotype constitutes a major defect in soft rice varieties with low AC, with lower amylose levels correlating with decreased transparency of the grains. Thus, the transparency of Nip(wx)-Wxmq is relatively lower compared to Nip(wx)-Wxmp and Nip(wx)-Wxb-c rice, aligning with its lower AC feature.
During the domestication of rice, there has been a noticeable global trend of decreasing AC from high to low. This shift is partially attributed to the natural Wx alleles that produce low to moderate AC, which often exhibit better eating and cooking quality (ECQ) [5]. Progress has been made in enhancing the ECQ of both indica and japonica rice varieties by incorporating Wxb (AC ~16%) and Wxin (AC ~20%) alleles into high AC (~25%) rice varieties with the Wxa allele. Additionally, the adoption of "soft rice" alleles, such as Wxop/hp and Wxmp, which impart even lower AC levels (5-12%), has further improved the ECQ. While reducing AC has positively impacted rice taste, it has concurrently introduced visual defects, particularly affecting grain transparency, a key aspect of rice visual quality. Consequently, a balanced approach to breeding that moderately adjusts AC could simultaneously optimize rice taste and visual appeal. In this context, the discovery of the mutation site Ex5-52C within the Wxmq allele heralds a significant breakthrough. This mutation offers a moderate AC level that could effectively reconcile the demands for both optimal grain transparency and desirable eating quality. Employing gene editing technologies to make precise alterations at specific genomic sites could resolve the transparency issues commonly observed in soft rice varieties. This strategy not only promises to enhance the sensory and aesthetic appeal of rice but also paves the way for breeding new rice varieties with fine-tuned AC levels.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: The schematic diagram of Wx gene structure and the primers used for plasmid construction, Figure S2. The phenotype of the transgenic rice plants, Table S1. Primers used in this study.
Author Contributions
Conceptualization, C.Z. and Q.L. (Qiaoquan Liu); methodology, Y.Y., L.Z., L.F. and J.J.; formal analysis, L.H. and Y.Z.; writing—original draft preparation, Y.Y. and Q.L. (Qing Liu); writing—review and editing, C.Z. and Q.L. (Qiaoquan Liu). All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Ministry of Science and Technology of China (2023YFD1200901), the National Natural Science Foundation of China (32230074 and 32161143004), the Research Programs from Jiangsu Government (BE2022336, and BM2022008-02), the Foreign Expert Project (G2023014064L), Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX21_3239), and the Yangzhou University high-end talent program.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jeon, J.S.; Ryoo, N.; Hahn, T.R.; Walia, H.; Nakamura, Y. Starch biosynthesis in cereal endosperm. Plant Physiol Biochem. 2010, 48, 383–392. [Google Scholar] [CrossRef]
- Zhou, L.H.; Zhang, C.Q.; Zhang, Y.D.; Wang, C.L.; Liu, Q.Q. Genetic manipulation of endosperm amylose for designing superior quality rice to meet the demands in the 21st century. Journal of Cereal Science. 2022, 105, 103481. [Google Scholar] [CrossRef]
- Li, H.Y.; Gilbert, R.G. Starch molecular structure: The basis for an improved understanding of cooked rice texture. Carbohydr Polym. 2018, 195, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Zhang, Y.D.; Zhu, Z.; Chen, T.; Zhao, Q.Y.; Zhong, W.G.; Yang, J.; Yao, S.; Zhou, L.H.; Zhao, L.; Li, Y.S. Research progress on the breeding of japonica super rice varieties in Jiangsu Province, China. J integr agr. 2017, 16, 992–999. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Zhu, J.H.; Chen, S.J.; Fan, X.L.; Li, Q.F.; Lu, Y.; Wang, M.; Yu, H.X.; Yi, C.D.; Tang, S.Z.; Gu, M.H.; Liu, Q.Q. Wxlv, the ancestral allele of rice Waxy gene. Mol Plant. 2019, 12, 1157–1166. [Google Scholar] [CrossRef]
- Huang, L.C.; Tan, H.Y.; Zhang, C.Q.; Li, Q.F.; Liu, Q.Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Communications. 2021, 2, 100237. [Google Scholar] [CrossRef]
- Yang, J.; Wang, J.; Fan, F.J.; Zhu, J.Y.; Chen, T.; Wang, C.L; Zheng, T.Q.; Zhang, J.; Zhong, W.G.; Xu, J.L. Development of AS-PCR marker based on a key mutation confirmed by resequencing of Wx-mp in Milky Princess and its application in japonica soft rice (Oryza sativa L.) breeding. Plant Breeding. 2013, 132, 595–603. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, Y.; Sakai, M.; Imbe, T. Molecular characterization of Wx-mq, a novel mutant gene for low-amylose content in endosperm of rice (Oryza sativa L.). Breed. Sci. 2002, 52, 131–135. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Zhang, J.L.; Wang, Z.Y.; Hong, M.M.; Gu, M.H. A highly efficient transformation system mediated by Agrobacterium tumefaciens in rice (Oryza sativa L.). Acta Phytophysiol. Sin. 1998, 24, 259–271. [Google Scholar]
- Ding, J.Y.; Jia, J.W.; Yang, L.T.; Wen, H.B.; Zhang, C.M.; Liu, W.X.; Zhang, D.B. Validation of a rice specific gene, sucrose phosphate synthase, used as the endogenous reference gene for qualitative and real-time quantitative PCR detection of transgenes. J Agric Food Chem. 2004, 52, 3372–3377. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, L.L.; Zhang, J.; Cai, X.L.; Liu, Q.Q.; Wei, C.X. Relationships between transparency, amylose content, starch cavity, and moisture of brown rice kernels. J Cereal Sci. 2019, 90, 102854. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Chen, S.J.; Ren, X.Y.; Lu, Y.; Liu, D.R.; Cai, X.L.; Li, Q.Q.; Gao, J.P.; Liu, Q.Q. Molecular structure and physicochemical properties of starches from rice with different amylose contents resulting from modification of OsGBSSI activity. J Agric Food Chem. 2017, 65, 2222–2232. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.W.; Man, J.M.; Huang, J.; Liu, Q.Q.; Wei, W.X.; Wei, C.X. Relationship between structure and functional properties of normal rice starches with different amylose contents. Carbohydr. Polym. 2015, 125, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.R.; Wang, W.; Cai, X.L. Modulation of amylose content by structure-based modification of OsGBSS1 activity in rice (Oryza sativa L.). Plant Biotech. J. 2014, 12, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Lv, D.J.; Zhou, L.; Yang, Y.; Hao, W.Z.; Huang, L.C.; Fan, X.L.; Zhao, D.S.; Li, Q.F.; Zhang, C.Q.; Liu, Q.Q. Combined effects of SSII-2RNAi and different Wx alleles on rice grain transparency and physicochemical properties. Carbohydr Polym. 2023, 15, 120651. [Google Scholar] [CrossRef]
- Vandeputte, G.E.; Vermeylen, R.; Geeroms, J.; Delcour., J.A. Rice starches. I. Structural aspects provide insight into crystallinity characteristics and gelatinization behavior of granular starch. J. Cereal Sci. 2003, 38, 43–52. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Zhou, L.H.; Zhu, Z.B.; Lu, H.W.; Zhou, X.Z.; Qian, Y.T.; Li, Q.F.; Lu, Y.; Gu, M.M.; Liu, Q.Q. Characterization of grain quality and starch fine structure of two japonica rice (Oryza Sativa) cultivars with good sensory properties at different temperatures during the filling stage. J. Agric. Food Chem. 2016, 64, 4048–57. [Google Scholar] [CrossRef] [PubMed]
- Cooke, D.; Gidley, M.J. Loss of crystalline and molecular order during starch gelatinisation: origin of the enthalpic transition. Carbohyd Polym. 1992, 227, 103–112. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Yang, Y.; Chen, S.J.; Liu, X.J.; Zhu, J.H.; Zhou, L.H.; Lu, Y.; Li, Q.F.; Fan, X.L.; Tang, S.Z.; Gu, M.H.; Liu, Q.Q. A rare Waxy allele coordinately improves rice eating and cooking quality and grain transparency. J Integr Plant Biol. 2021, 63, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Ma, X.D.; Liu, S.J.; Zhu, C.L.; Jiang, L.; Wang, Y.H.; Shen, Y.; Ren, Y.L.; Dong, H.; Chen, L.M.; Liu, X.; Zhao, Z.G.; Zhai, H.Q.; Wan, J.M. Identification and characterization of a novel Waxy allele from a Yunnan rice landrace. Plant Mol Biol, 2009, 71, 609–626. [Google Scholar] [CrossRef]
- Huang, L.C.; Li, Q.F.; Zhang, C.Q.; Chu, R.; Gu, Z.W.; Tan, H.Y.; Zhao, D.S.; Fan, X.L.; Liu, Q.Q. Creating novel Wx alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system. Plant Biotechnol. J. 2020, 18, 2164–2166. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.C.; Liu, T.L.; Ma, X.L.; Wang, B.; Zheng, Z.Y.; Zhang, Y.L.; Xie, X.R.; Yang, B.W.; Zhao, Z.; Zhu, Q.L.; Liu, Y.G. Quantitative regulation of Waxy expression by CRISPR/Cas9–based promoter and 5’UTR–intron editing improves grain quality in rice. Plant Biotechnol. J. 2020, 18, 2385–2387. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lin, Q.P.; Li, X.F.; Wang, F.Q.; Chen, Z.H.; Wang, J.; Li, W.Q.; Fan, F.J.; Tao, Y.J.; Jiang, Y.J.; Wei, X.D.; Zhang, R.; Zhu, Q.H.; Bu, Q.Y.; Yang, J.; Gao, C.X. Fine-tuning the amylose content of rice by precise base editing of the Wx gene. Plant Biotechnol. J. 2021, 19, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Ding, Q.; Wang, W.S.; Pan, Y.L.; Tan, C.; Qiu, Y.B.; Chen, Y.; Li, H.J.; Li, Y.L.; Ye, N.Z.; Xu, N.; Wu, X.; Ye, R.J.; Liu, J.F.; Ma, C.L. Targeted deletion of the first intron of the Wxb allele via CRISPR/Cas9 signifcantly increases grain amylose content in rice. Rice. 2022, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.S.; Zhang, C.Q.; Li, Q.F.; Liu, Q.Q. Genetic control of grain appearance quality in rice. Biotechnol Adv. 2022, 108014. [Google Scholar] [CrossRef] [PubMed]
- Isshiki, M.; Matsuda, Y.; Takasaki, A.; Wong, H.L.; Satoh, H.; Shimamoto, K. Du3, a mRNA cap-binding protein gene, regulates amylose content in Japonica rice seeds. Plant Biotechnol, 2008, 25, 483–487. [Google Scholar] [CrossRef]
- Igarashi, H.; Ito, H.; Shimada, T.; Kang, D.J.; Hamada, S. A novel rice dull gene, LowAC1, encodes an RNA recognition motif protein affecting Waxyb pre-mRNA splicing. Plant Physiol Biochem, 2021, 162, 100–109. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, W.W.; Fu, Y.S.; Shan, Z.Z.; Xu, J.H.; Wang, P.; Kong, F.; Jin, J.; Yan, H.G.; Ge, X.Y.; Wang, Y.X.; You, X.M.; Chen, J.; Li, X.; Chen, W.W.; Chen, X.G.; Ma, J.; Tang, X.J.; Zhang, J.; Bao, Y.Q.; Jiang, L.; Wang, H.Y.; Wan, J.M. Du13 encodes a C2 H2 zinc-finger protein that regulates Wxb pre-mRNA splicing and microRNA biogenesis in rice endosperm. Plant Biotechnol J. 2022, 20, 1387–1401. [Google Scholar] [CrossRef]
Figure 1.
Characterization of grain appearance quality in transgenic rice lines. (A) Depicts the structural organization and allelic diversity of Wx gene, highlighting the genetic variations that underpin starch synthesis and ultimately affect grain quality. (B) Illustrates the configurations of transgenic vectors, each incorporating genomic DNA segments specific to the Wxmp, Wxb-5c, and Wxmq alleles. (C) Presents a visual comparison of grain appearance under two lighting conditions: reflected light (upper panel) and transmitted light (down panel). (D) Quantifies grain transparency by analyzing the gray scale values derived from the image data. (E) Examines the starch morphology within transverse sections of rice grains at a microscopic level. Scale bar, 5 μm.
Figure 1.
Characterization of grain appearance quality in transgenic rice lines. (A) Depicts the structural organization and allelic diversity of Wx gene, highlighting the genetic variations that underpin starch synthesis and ultimately affect grain quality. (B) Illustrates the configurations of transgenic vectors, each incorporating genomic DNA segments specific to the Wxmp, Wxb-5c, and Wxmq alleles. (C) Presents a visual comparison of grain appearance under two lighting conditions: reflected light (upper panel) and transmitted light (down panel). (D) Quantifies grain transparency by analyzing the gray scale values derived from the image data. (E) Examines the starch morphology within transverse sections of rice grains at a microscopic level. Scale bar, 5 μm.
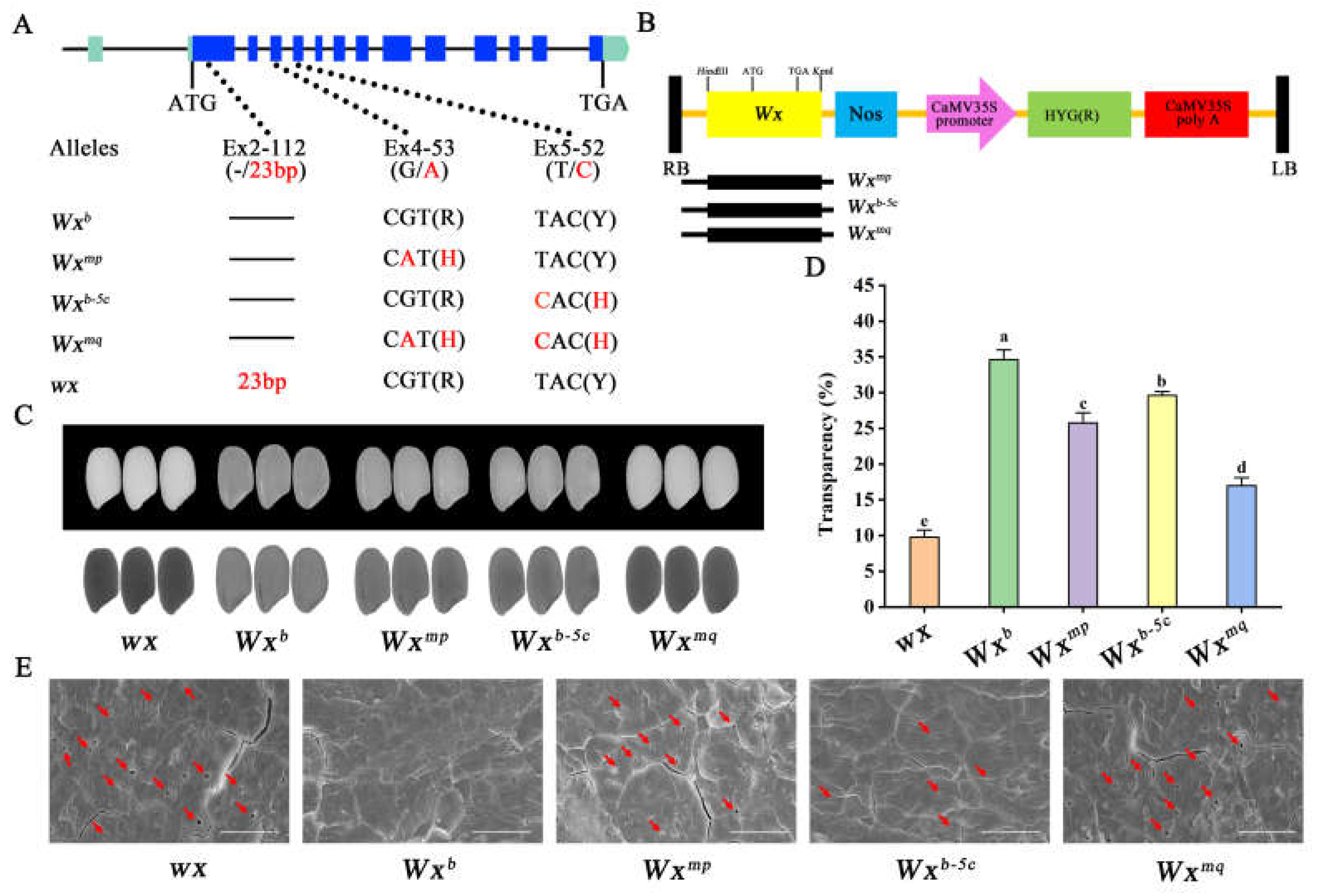
Figure 2.
Analysis of grain physicochemical properties in transgenic rice lines: (A) measures the amylose content (AC) across different transgenic lines. (B) evaluates the gel consistency (GC) of rice, a parameter that reflects the viscous properties of rice gel, indicative of the texture and palatability of cooked rice. (C) assess the taste value of cooked rice. (D) presents the Rapid Visco Analyzer (RVA) spectra of rice flours from the transgenic rice lines, offering insights into the pasting behavior and viscosity characteristics of the rice flour, which are pivotal for processing and culinary applications. (E) Details the gelatinization spectra of rice flours, which delineate the thermal transition properties of starch granules in rice flour.
Figure 2.
Analysis of grain physicochemical properties in transgenic rice lines: (A) measures the amylose content (AC) across different transgenic lines. (B) evaluates the gel consistency (GC) of rice, a parameter that reflects the viscous properties of rice gel, indicative of the texture and palatability of cooked rice. (C) assess the taste value of cooked rice. (D) presents the Rapid Visco Analyzer (RVA) spectra of rice flours from the transgenic rice lines, offering insights into the pasting behavior and viscosity characteristics of the rice flour, which are pivotal for processing and culinary applications. (E) Details the gelatinization spectra of rice flours, which delineate the thermal transition properties of starch granules in rice flour.
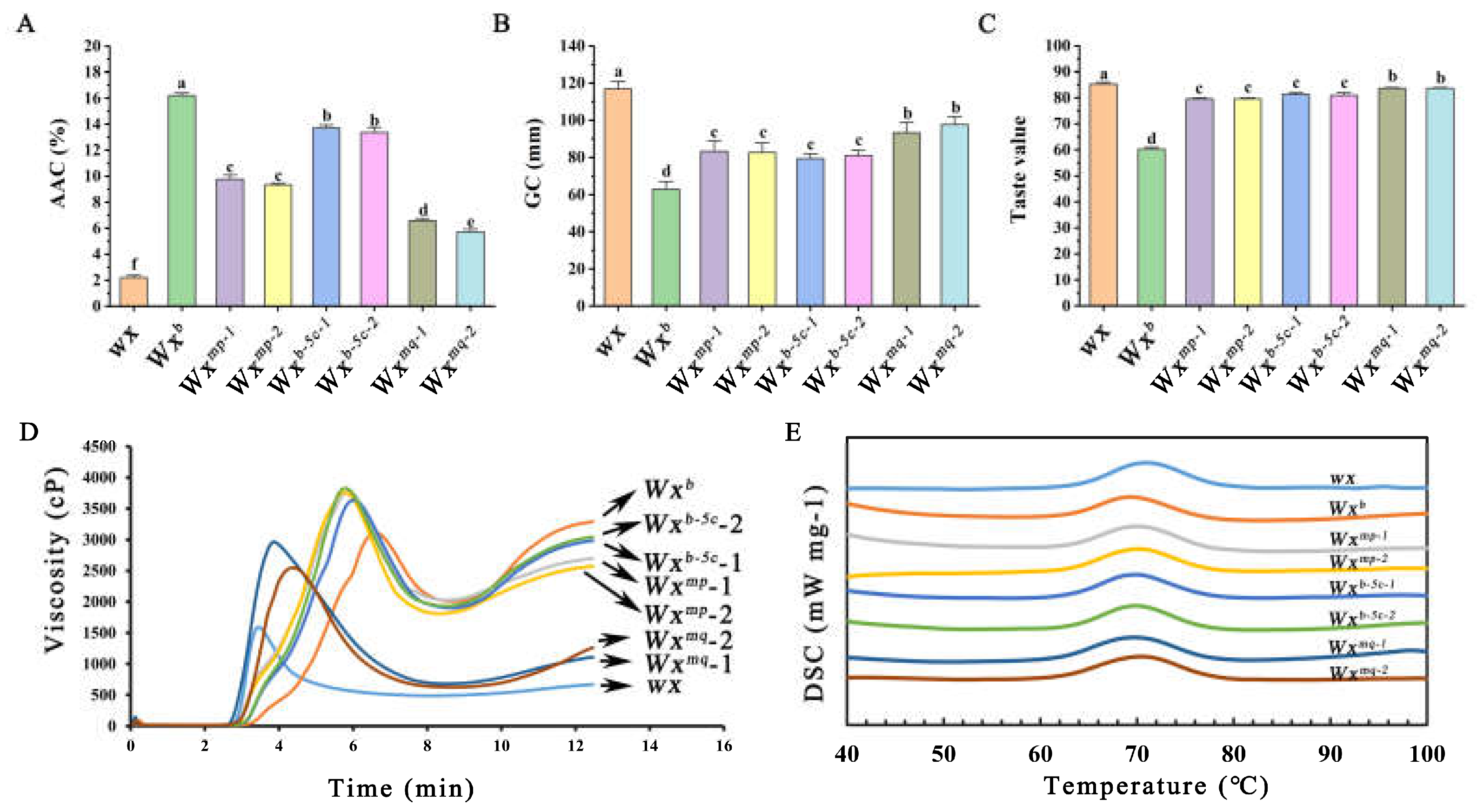
Figure 3.
Crystalline structures of starch granules from different transgenic rice lines:(A) Gel permeation chromatograms of isoamylase-debranched starches. AP1, AP2, and AM indicate amylopectin short-branched chains (AP1), long-branched chains (AP2), and amylose (AM), respectively. (B) XRD patterns and (C) ATR-FTIR spectra of rice starches from different transgenic rice lines.
Figure 3.
Crystalline structures of starch granules from different transgenic rice lines:(A) Gel permeation chromatograms of isoamylase-debranched starches. AP1, AP2, and AM indicate amylopectin short-branched chains (AP1), long-branched chains (AP2), and amylose (AM), respectively. (B) XRD patterns and (C) ATR-FTIR spectra of rice starches from different transgenic rice lines.
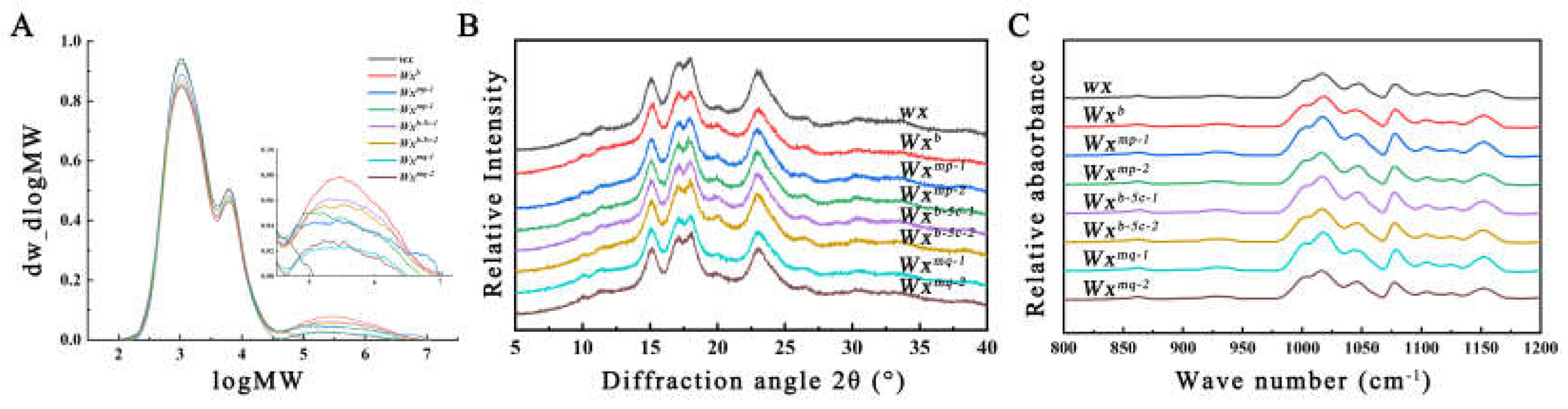
Figure 4.
Wx allele functionality in transgenic rice lines:(A) displays the relative expression levels of Wx gene in immature seeds. (B) reports on the activity of Granule Bound Starch Synthase I (GBSSI), the enzyme responsible for amylose synthesis in rice. (C) illustrates the results of Western blot analysis for the GBSSI protein, visualizing the presence and abundance of this key enzyme in the transgenic rice lines, further elucidating the molecular underpinnings of starch synthesis regulation.
Figure 4.
Wx allele functionality in transgenic rice lines:(A) displays the relative expression levels of Wx gene in immature seeds. (B) reports on the activity of Granule Bound Starch Synthase I (GBSSI), the enzyme responsible for amylose synthesis in rice. (C) illustrates the results of Western blot analysis for the GBSSI protein, visualizing the presence and abundance of this key enzyme in the transgenic rice lines, further elucidating the molecular underpinnings of starch synthesis regulation.
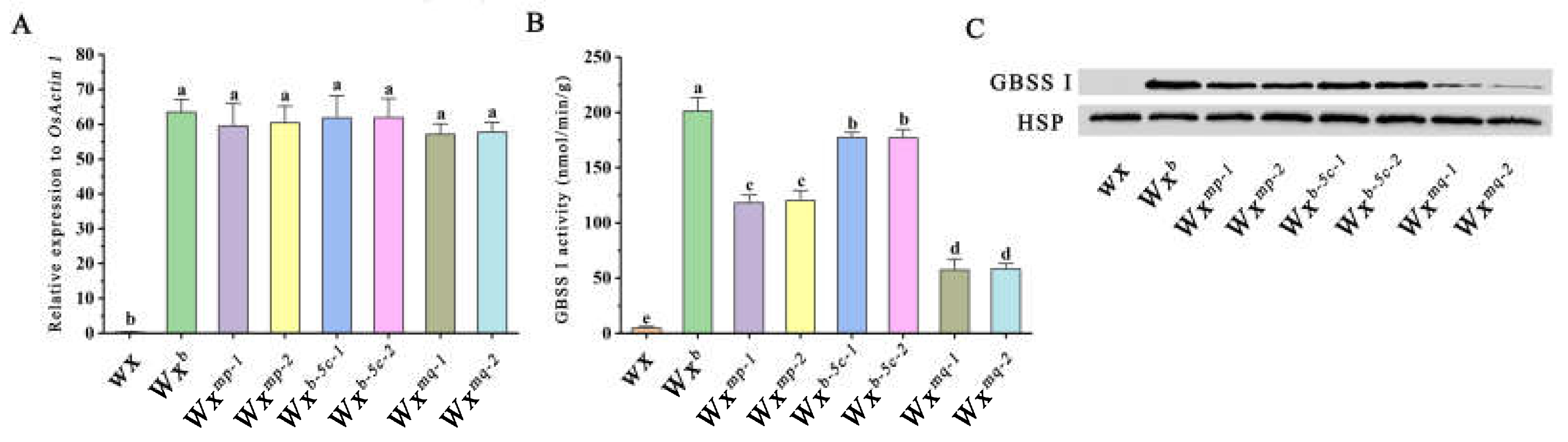

Table 1.
Pasting properties of rice flours from transgenic rice and its wild type.
| Line | Peak Visco (cP) |
Trough Visco (cP) |
Breakdown (cP) |
Final Visco (cP) |
Setback (cP) |
|---|---|---|---|---|---|
| Nip(wx) | 1575.50±17.68 g | 495.00±9.90 f | 1116.00±22.63 f | 660.00±9.90 g | -927.50±9.19 b |
| Nip(Wxb) | 3109.00±11.31 d | 1792.50±6.36 c | 1307.00±18.38 e | 3235.00±16.97 a | 135.50±7.78 a |
| Nip(wx)-Wxmp-1 | 3774.00±18.38 a | 1815.50±10.60 c | 1963.00±14.14 b | 2558.50±14.85 d | -1185.00±9.90 e |
| Nip(wx)-Wxmp-2 | 3762.50±10.61 a | 2014.50±19.09 a | 1954.00±16.97 bc | 2680.50±19.09 c | -1089.50±19.09 d |
| Nip(wx)-Wxb-5c-1 | 3568.00±18.19 c | 1672.00±18.38 d | 1904.00±15.56 cd | 2577.00±14.14 d | -1004.00±24.04 c |
| Nip(wx)-Wxb-5c-2 | 3654.00±24.49 b | 1974.50±19.09 b | 1886.50±20.51 d | 2796.00±24.04 b | -1032.50±20.51 c |
| Nip(wx)-Wxmq-1 | 2970.50±10.60 e | 667.00±19.80 e | 2262.50±27.58 a | 1124.00±26.87 f | -1845.00±18.38 g |
| Nip(wx)-Wxmq-2 | 2532.50±16.26 f | 636.50±16.26 e | 1935.50±23.33 bcd | 1251.50±16.26 e | -1295.00±19.80 f |
Data represent means ± standard deviations, n =2. For each column in the same flour or starch samples, values displaying different lowercase letters are significantly different by one-way ANOVA with multiple comparisons (p < 0.05).
Table 2.
Thermal properties of rice flours from transgenic rice and its wild type.
| Line | To (°C) | Tp (°C) | Tc (°C) | ΔH (J·g-1) |
|---|---|---|---|---|
| Nip(wx) | 61.55±0.21 c | 69.30±0.14 c | 77.15±0.14 c | 7.45±0.05 a |
| Nip(Wxb) | 63.85±0.35 a | 71.05±0.21 a | 78.65±0.21 a | 5.90±0.10 d |
| Nip(wx)-Wxmp-1 | 62.95±0.35 b | 70.05±0.07 b | 77.55±0.21 bc | 6.48±0.08 c |
| Nip(wx)-Wxmp-2 | 63.00±0.28 b | 70.15±0.07 b | 77.30±0.14 c | 6.50±0.06 c |
| Nip(wx)-Wxb-5c-1 | 62.80±0.14 b | 70.00±0.28 b | 77.45±0.07 bc | 6.48±0.06 c |
| Nip(wx)-Wxb-5c-2 | 62.95±0.35 b | 70.00±0.14 b | 77.40±0.28 bc | 6.49±0.03 c |
| Nip(wx)-Wxmq-1 | 62.65±0.35 b | 70.05±0.35 b | 77.65±0.21 bc | 7.25±0.03 b |
| Nip(wx)-Wxmq-2 | 62.92±0.07 b | 70.30±0.14 b | 77.85±0.21 b | 7.24±0.04 b |
The data represent means ± standard deviation, n = 3. To, Tp, Tc, and ΔH indicate onset temperature, peak temperature, conclusion temperature, and enthalpy of gelatinization, respectively. Means with different lowercase letters in each column for the same cultivar are significantly different by one-way ANOVA with multiple comparisons (p < 0.05).
Table 3.
GPC, Relative Crystallinities, and ATR-FTIR Parameters from transgenic rice and its wild type.
Table 3.
GPC, Relative Crystallinities, and ATR-FTIR Parameters from transgenic rice and its wild type.
| Line | AP1 | AP2 | AM | relative crystallinity (%) | 1045/1022 cm-1 |
|---|---|---|---|---|---|
| Nip(wx) | 74.22±0.29 a | 25.78±0.29 a | - | 29.6±0.13 a | 0.64±0.006 a |
| Nip | 63.52±0.3 e | 22.24±0.09 d | 14.24±0.21 a | 26.6±0.28 e | 0.55±0.004 e |
| Nip(wx)-Wxmp-1 | 69.42±0.23 c | 23.5±0.37 c | 7.08±0.13 c | 28.24±0.14 c | 0.6±0.002 c |
| Nip(wx)-Wxmp-2 | 69.22±0.28 c | 23.59±0.11 c | 7.19±0.17 c | 28.22±0.11 c | 0.6±0.004 c |
| Nip(wx)-Wxb-5c-1 | 66.39±0.01 d | 23.16±0.12 c | 10.46±0.11 b | 27.53±0.33 d | 0.57±0.003 d |
| Nip(wx)-Wxb-5c-2 | 65.82±0.4 d | 23.42±0.22 c | 10.76±0.18 b | 27.46±0.24 d | 0.58±0.004 d |
| Nip(wx)-Wxmq-1 | 71.82±0.35 b | 24.48±0.3 b | 3.7±0.05 d | 28.82±0.11 b | 0.63±0.004 b |
| Nip(wx)-Wxmq-1 | 72.17±0.42b | 24.52±0.33 b | 3.31±0.1 e | 28.82±0.2 b | 0.63±0.003 b |
Data represent means ± standard deviations, n = 2. For each column in the same flour or starch samples, values displaying different lowercase letters are significantly different by one-way ANOVA with multiple comparisons (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



