
Submitted:
17 April 2024
Posted:
18 April 2024
You are already at the latest version
Abstract
Small interfering RNA (siRNA) holds significant therapeutic potential by silencing target genes through RNA interference. Current clinical applications of siRNA have been primarily limited to liver diseases, while achievements in delivery methods are expanding its applications to various organs, including the lungs. Cholesterol-conjugated siRNA emerges as a promising delivery approach due to its low toxicity and high efficiency. This study focuses on developing a cholesterol-conjugated anti-Il6 siRNA and the evaluation of its potency for the potential treatment of inflammatory diseases using the example of acute lung injury (ALI). The biological activities of different Il6-targeted siRNAs containing chemical modifications were evaluated in J774 cells in vitro. The lead cholesterol-conjugated anti-Il6 siRNA after intranasal instillation demonstrated dose-dependent therapeutic effects in a mouse model of ALI induced by lipopolysaccharide (LPS). The treatment significantly reduced Il6 mRNA levels, inflammatory cell infiltration, and the severity of lung inflammation. IL6 silencing by cholesterol-conjugated siRNA proves to be a promising strategy for treating inflammatory diseases, with potential applications beyond the lungs.
Keywords:
acute lung injury
; inflammation
; chemically modified siRNA
; IL6
; LPS
1. Introduction
Small interfering RNAs (siRNAs) silence the expression of the target genes by inducing RNA interference (RNAi) [1]. The catalytic sequence-specific silencing activity of siRNA provides high efficiency and selectivity of their action, which allows them to be considered as a new class of drugs for the therapy of diseases that are not treatable by small molecules or monoclonal antibodies [2]. Currently, the clinical use of siRNA is limited to liver diseases; however, methods for delivering RNA to other organs, including the brain and spinal cord [3,4], kidneys [5,6], spleen [7], lungs [3,8] are currently under development. The employment of siRNA covalent conjugates with transport ligands is the most promising among siRNA delivery methods due to their low toxicity and high delivery efficiency [9]. 4 from 5 siRNA based drugs already approved for clinical use are siRNA conjugates, and they demonstrate safe toxicity and immunogenicity profiles [10,11,12,13,14]. The lungs are important target organs for siRNA therapy since they are involved in the development of life-threatening diseases. On the other hand, non-invasive local delivery to lungs could be used to reduce the incidence of systemic side effects and provide effective accumulation of the drug. The use of cholesterol-conjugated siRNA for this purpose seems particularly promising since it can retard in the organs after local delivery and excludes the use of delivery vehicles to avoid additional lung injury [15,16,17]. Therefore, in this work, we used intranasal administration of cholesterol-containing siRNA conjugates to block target gene expression in the lungs.
Inflammatory diseases affect, according to various estimates, 5%-9% of the world population [18,19], and their incidence increases every year [20]. Inflammation is associated with reduced life expectancy [19], as well as an elevated risk of cancer [21]. Existing drugs for the treatment of inflammation are characterized by side effects, including toxicity, immunogenicity, and an increased risk of developing infectious diseases [22,23]. In addition, not all patients respond to existing therapy, and drug resistance may develop over time [22,24].
Acute lung injury (ALI), or its more severe manifestation, acute respiratory distress syndrome (ARDS), is a clinical syndrome characterized by damage of the vascular endothelium and alveolar epithelium, which leads to interstitial and pulmonary edema and, ultimately, alveolar collapse [25]. An important role in increasing the permeability of the alveolar–capillary barrier is played by neutrophils that migrate into the lung, secreting pro-inflammatory and pro-apoptotic mediators that damage neighboring cells [26]. An estimated incidence of ALI/ARDS is 64.2 cases per 100,000 person-years, the mortality rate is 29-42% [26,27]. ALI treatment is not addressed adequately by available therapy and represents an unmet medical need.
We chose interleukin-6 (Il6) as a target gene, which is one of the master regulators of inflammatory processes [28], facilitating neutrophil recruitment to the lungs [29]. A decrease in its level is associated with a weakening of ALI symptoms [30]. Thus, in this work, we developed an anti-Il6 siRNA, conjugated it with cholesterol, validated their silencing activity in vitro, and examined its anti-inflammatory properties in the ALI mouse model.
2. Materials and Methods
Synthesis of siRNAs, their cholesterol-containing analogues, and duplex annealing
The anti-Il6 siRNA sequences are listed in Table 1, the control siRNA (siSCR) has no significant homology to any known mouse, rat, or human mRNA sequence. Oligoribonucleotides and their analogs were synthesized by the phosphoramidite method on an automatic ASM-800 synthesizer (Biosset, Novosibirsk, Russia). 2'-O-TBDMS-protected, 2'-F-, 2'-O-Me-ribophosphoramidites, and CPG polymeric carriers with an attached first nucleoside (Glen Research, Sterling, VA, USA) were used in the synthesis. Sulfurizing Reagent II (Glen Research, Sterling, VA, USA) was used to introduce phosphothioate linkages. For the synthesis of siRNA conjugates containing a cholesterol residue with a hexamethylene linker at the 5'-end, a solid-phase synthesis method was used based on the activation of the free 5'-hydroxyl group of a protected polymer-bound oligonucleotide with N,N'-disuccimidyl carbonate (Acros Organics, Geel, Belgium), followed by the interaction with cholesteryl-6-aminohexylcarbamate by analogy with [31]. After standard deprotaction the target products were isolated by preparative gel electrophoresis in 15% polyacrylamide gel (PAAG) under denaturing conditions, followed by elution of the products with a 0.3 M NaClO4 solution. The isolated products were desalted on a Sep-Pac C18 cartridge (Waters, Milford, MA, USA) or Amicon Ultra 3K (Millipore, Burlington, MA, USA) and precipitated with a 2% NaClO4 solution in acetone. To obtain duplexes, equimolar concentrations of the sense and antisense siRNA strands were incubated in 30 mM HEPES-KOH (pH 7.4), 100 mM potassium acetate, and 2 mM magnesium acetate at 90°C for 5 min. With a gradual decrease in temperature, the strands hybridized for 1 h, and the duplexes were stored at -20°C.
Cell Culture
The J774 macrophage cell line was obtained from the Russian Cell Culture Collection (Institute of Cytology, RAS, St. Petersburg, Russia). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Sigma-Aldrich Inc., St. Louis, MO, USA) supplemented with 10% heat-inactivated fetal bovine serum (BioloT, St. Petersburg, Russia) and an antibiotic-antimycotic solution (100 U/mL penicillin, 100 μg/mL streptomycin, 0.25 μg/mL amphotericin) and incubated at 37°C in a humidified 5% CO2-containing air atmosphere (hereafter standard conditions).
Mice
Female 6–8-week-old Balb/C mice (average weight 20–22 g) were obtained from the Vivarium of the Institute of Chemical Biology and Fundamental Medicine SB RAS (Novosibirsk, Russia). The mice were housed in plastic cages (3-6 animals per cage) under normal daylight conditions. Water and food were provided ad libitum. Experiments were carried out in accordance with the European Communities Council Directive 86/609/CEE. The experimental protocols were approved by the Committee on the Ethics of Animal Experiments of the Administration of the Siberian Branch of the Russian Academy of Sciences (Novosibirsk, Russia) (protocol No. 56 from August 10, 2019).
Transfection of siRNA
One day before the experiment, J774 cells in the exponential phase of growth were plated in 24-well plates at a density of 1.5×105 cells/well. After 24 h, the growth medium was replaced by fresh serum-free DMEM. The cells were transfected with siRNAs (1-100 nM) using Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) according to the manufacturer’s protocol (2 μL of Lipofectamine 2000 per well). Two days after transfection, cells were replated to prevent overgrowth. Then, 4 days after transfection, LPS (final concentration: 1 nM) was added to the cells. 6 hours after the addition of LPS, total RNA was isolated from the cells using a kit for RNA isolation, Namagp100 (Biolabmix, Novosibirsk, Russia), and an Auto-Pure 96 automatic nucleic acid isolation and purification system (Allsheng, Hangzhou, China), according to the manufacturer’s protocol. RT-qPCR was performed using M-MuLV-RH revertase and BioMaster HS-qPCR (Biolabmix, Novosibirsk, Russia). The amount of Il6 mRNA was normalized to the amount of Hprt mRNA used as an internal standard. To assess the mRNA level of the genes, the following primers and probes were used:
Il6_F 5'-AAACCGCTATGAAGTTCCTCTC-3'
IL6_Probe: 5'-((5,6)-FAM)-TTGTCACCAGCATCAGTCCCAAGA-3'-BHQ1
IL6_R: 5'-GTGGTATCCTCTGTGAAGTCTC-3'
Hprt_F: 5'-CCCCAAAATGGTTAAGGTTGC-3'
Hprt_Probe: 5'- ((5,6)-ROX)-CTTGCTGGTGAAAAGGACCT-3'-BHQ2
Hprt_R: 5'-AACAAAGTCTGGCCTGTATCC-3'
Data processing was carried out using Bio-Rad CFX Manager 3.1 software (Bio-Rad Laboratories Inc., Hercules, CA, USA).
LPS-induced acute lung injury (ALI)
Mice (n = 3-5 in each group) were challenged with LPS (10 µg per mouse, 055:B5, Sigma-Aldrich, USA) via intranasal (i.n.) instillations under isoflurane anesthesia. Ch-siIL6PS were administered i.n. 4 days before ALI induction. The mice were euthanized 6 h after the induction of lung inflammation; the lungs were lavaged with 1 mL of ice-cold saline buffer.
The 20 µL of collected bronchoalveolar lavage (BAL) fluids were processed for cell counting. 20 µl BAL fluids were incubated for 7 min after the addition of a 400 µL solution with 150 mM NH4Cl, 10 mM NaHCO3, and 0.1 mM EDTA (рН 7.5). Then the solution was centrifuged for 5 min at 500 g at room temperature, supernatant was removed, and cells were counted in 30 µl of saline solution. To determine the differential leukocyte counts, 50 µL of bronchoalveolar cell suspension were placed onto slides, stained with azur-eosin by Romanovsky-Giemsa method, and examined microscopically. The results were expressed as the number of total leukocytes (×105 cells/ml) and the percentages of subpopulations of granulocytes, lymphocytes, and monocytes (%).
The rest of the BAL fluids were centrifuged (1600 rpm, 10 min, 4 °C), the supernatant was removed, RNA isolation and RT-qPCR were carried out as described above.
Histology
For the histological study, lung specimens were fixed in 10% neutral-buffered formalin (BioVitrum, Moscow, Russia), dehydrated in ascending ethanols and xylols, and embedded in HISTOMIX paraffin (BioVitrum, Russia). Paraffin sections (up to 5 µm) were sliced on a Microm HM 355 S microtome (Thermo Fisher Scientific, Waltham, MA, USA) and stained with hematoxylin and eosin. All the images were examined and scanned using an Axiostar Plus microscope equipped with an Axiocam MRc5 digital camera (Zeiss, Oberkochen, Germany) at magnification of × 200.
The intensity of inflammatory infiltration in the lung tissue was assessed by a semi-quantitative method, where 0 is no inflammatory infiltration, 1 is low intensity of inflammatory infiltration, 2 is moderate intensity of inflammatory infiltration, and 3 is high intensity of inflammatory infiltration. Morphometric analysis of lung sections included evaluation of the volume densities (Vv, %) of alveolar septa outside the foci of inflammatory infiltration, reflecting interstitial edema, and was performed using a counting grid consisting of 100 testing points in a testing area equal to 3.2 × 106 μm2. The quantification was performed at a magnification of ×200 in 5 test fields for each lung sample; the number of samples studied was from two to four for each experimental group; thus, 10-20 random fields were analyzed in each experimental group.
Statistical analysis
The variables were expressed as the mean ± standard deviation (SD) or standard error of the mean (SEM). The data were analyzed with the Student’s t-test. The data obtained in vivo were statistically processed using a two-way ANOVA followed by Bonferroni’s post hoc test. The differences between the values are considered statistically significant at p < 0.05. The statistical package STATISTICA, version 10.0, used for analysis.
3. Results
3.1. Silencing activities of anti-Il6 siRNAs in vitro
We designed nine 2'OMe selectively modified siRNAs targeting at Il6 to silence its mRNA level (Table 1). After RNA synthesis and RNA duplex hybridization, we examined the silencing activity of the siRNAs in J774 cells using Lipofectamine 2000 as a transfection agent (Figure 1A).

Table 1.
Oligoribonucleotide sequences.
| Designation | Sequence1 |
|---|---|
| IL6.1.AS | GUUmAUmGCCUmAAGCmAUmAUCmAGUUU |
| IL6.1.S | ACUmGAUmAUmGCUUmAGGCmAUmAACGC |
| IL6.2.AS | UmAAGGACCmAAGACCmAUCCmA |
| IL6.2.S | UmGGAUmGGUCUUmGGUCCUUmAGC |
| IL6.3.AS | GUCmACUUmGAAAUmGUUmAUmAUmGU |
| IL6.3.S | AUmAUmAACmAUUUCmAAGUmGACmAC |
| IL6.4.AS | UUmGGGACmACUmAUUUUmAAUUmAU |
| IL6.4.S | AAUUmAAAAUmAGUmGUCCCmAACmA |
| IL6.5.AS | UmGCCUmAAGCmAUmAUCmAGUUUmGU |
| IL6.5_S | AAACUmGAUmAUmGCUUmAGGCmAUmA |
| IL6.6.AS | UmGCUmAAUUUmAAAUmAUmGUUUUU |
| IL6.6.S | AAACmAUmAUUUmAAAUUmAGCmAAU |
| IL6.7.AS | AGUCGGAGGCUUmAAUmUCmACmA |
| IL6.7.S | UmGUmAAUUmAAGCCUCCGACUUmG |
| IL6.8.AS | CUmACCmAAACUmGGAUmAUmAAUCmA |
| IL6.8.S | AUUmAUmAUCCmAGUUUmGGUmAGCmA |
| IL6.9.AS | CmAGGAAAUUUmGCCUmAUUmGAAA |
| IL6.9.S | UCmAAUmAGGCmAAAUUUCCUmGAU |
| IL6.1FM.AS | GmUmUmAmUmGmCfCmUfAfAfGmCmAmUmAmUmCmAmGmUmUmUm |
| IL6.1FM.S | AmCfUmGmAmUfAmUmGmCmUmUmAmGfGmCfAmUmAmAmCmGmCm |
| IL6.1PS.AS | Gm*Um*UmAmUmGmCfCmUfAfAfGmCmAmUmAmUmCmAmGmUm*Um*Um |
| IL6.1PS.S | Am*Cf*UmGmAmUfAmUmGmCmUmUmAmGfGmCfAmUmAmAmCm*Gm*Cm |
| Ch-IL6.1PS.AS | Ch-Gm*Um*UmAmUmGmCfCmUfAfAfGmCmAmUmAmUmCmAmGmUm*Um*Um |
| LucPS.AS | Cm*Gm*UmUmAmUmUfUmAfUfCfGmGmAmGmUmUmGmCm*Am*Gm |
| LucPS.S | Gm*Cf*AmAmCmUfCmCmGmAmUmAmAmAfUmAfAmCmGm*Cm*Gm |
| ScrFM.AS | AmAfUmAmUmCfUmGmCmUmCmUmUmCfAmUfGmCmGmGmGm |
| ScrFM.S | CmGmCmAmUmGmAfAmGfAfGfCmAmGmAmUmAmUmUmCmGm |
| ScrPS.AS | Am*Af*UmAmUmCfUmGmCmUmCmUmUmCfAmUfGmCmGm*Gm*Gm |
| Ch-ScrPS.S | Ch-Cm*Gm*CmAmUmGmAfAmGfAfGfCmAmGmAmUmAmUmUm*Cm*Gm |
1 Sequences are in the 5'–3' direction: Am, Cm, Gm, Um–2'-O-methyl modified; Af, Cf, Gf, Uf–2'-fluoro modified; *–phosphorothioate modification; AS-anti-sense strand; S-sense strand; Ch–cholesterol residue attached to the 5' end of the sense strand via a hexamethylenediamine linker.
The level of Il6 mRNA decreased significantly after transfection of 5 siRNAs with 50–66% efficacy; other siRNAs were less active (Figure 1B). The sequence of the most effective siRNA (siIL6.1) was used for the design of siRNA for in vivo experiments: 2'F, 2'OMe modifications were introduced according to [32] to improve nuclease resistance and silencing activity (siIL6.1FM). Further, siIL6FM was conjugated with cholesterol to provide carrier-free delivery of siRNA to target cells and equipped with additional phosphorothioate modifications (PS), resulting in Ch-siIL6.1PS, since nuclease resistance is more important to maintain the integrity of siRNA when delivered without a carrier in vivo than under transfection in vitro [33,34]. 100 nM siIL6.1FM and Ch-siIL6.1PS reduced Il6 mRNA levels after transfection with Lipofectamine 2000 with 69% and 68% efficacy, which is comparable with the action of selectively modified siIL6.1. However, siIL6.1FM was more active at lower concentrations than Ch-siIL6.1PS under transfection, with IC50 of 1 and 15 nM, respectively (Figure 1C). The decrease in siRNA activity upon the attachment of cholesterol is likely related to the steric block of siRNA interaction with RNAi machinery and has been observed previously [33]. Since Ch-siIL6.1PS efficiently decreased Il6 mRNA levels we used this conjugate in vivo.
3.2. Ch-siIL6.1PS silences Il6 mRNA level and reduces the severity of acute lung injury in mice
We used the acute lung injury (ALI) model in order to evaluate the significance of modulating Il6 mRNA level in alleviating lung tissue damage and determine the effectiveness of the obtained Ch-siIL6.1PS in vivo. Ch-siIL6.1PS was instilled intranasally (i.n.) 4 days before ALI induction by i.n. administration of LPS. The scheme including siRNA pretreatment was chosen because the dynamics of silencing caused by siRNA cholesterol conjugates develops more slowly than the activation of cytokine expression. The control and experimental animals were euthanized 6 h after ALI induction, and the Il6 mRNA level in bronchoalveolar lavage (BAL) fluid cells and pro-inflammatory parameters in BAL fluid and in lung tissue were analyzed (Figure 2A).
The time intervals were selected based on our previously obtained data on the dynamics of activation of IL6 synthesis during LPS-mediated ALI [35]. Data showed that the Il6 mRNA level increased by two orders of magnitude in LPS-challenged mice compared to the untreated group (Figure 2B). Instillation of Ch-siIL6.1PS at doses 2.1, 4.2, and 8.5μg/g decreased Il6 mRNA level in BAL fluid cells of LPS-challenged mice by 30, 71, and 78%, respectively (p < 0.001 for Ch-siIL6.1PS at 8.5 and 4.2 μg/g vs. LPS-challenged group; p < 0.05 and p < 0.01 for Ch-siIL6.1PS at 8.5 and 4.2 μg/g vs. Ch-siScrPS at dose 8.5 μg/g, respectively).
The analysis of inflammatory changes in the respiratory system showed that LPS instillation increased the number of total leukocytes in BAL fluid by 8-fold (Figure 2C), predominantly due to granulocytes, compared with healthy animals (Figure 2D). Histological analysis of lung tissue of LPS-challenged mice revealed inflammatory and destructive changes represented by granulocyte infiltration and exudation, as well as desquamation of the bronchial and alveolar epithelium (Figure 3 A,B). Moreover, LPS administration led to a 1.6-fold increase in the volume density of the alveolar septa, reflecting interstitial lung edema as one of the morphological components of ALI compared with healthy mice (Figure 3C).
The administration of Ch-siScrPS had no significant effect on the intensity of inflammatory changes in both BAL fluid and lung tissue of mice with ALI (Figure 2 C,D). Pretreatment of LPS-challenged mice with Ch-siIL6.1PS caused marked suppression of inflammatory changes in the respiratory system. As shown in Figure 2C, administration of Ch-siIL6.1PS at a dose of 2.1, 4.2, and 8.5 μg/g diminished the number of total leukocytes in BAL fluid 6 h after ALI induction 1.9, 2.7, and 5-fold compared to the non-treated control and 2.8, 4.1, and 7.6-fold compared to the Ch-siScrPS-treated group, respectively. Analysis of leukocyte subpopulations in the BAL fluid of Ch-siIL6.1PS-treated ALI mice demonstrated their ability to suppress inflammation-associated granulocyte recruitment, especially at a dose of 8.5 μg/g (Figure 2D).
Histological study of the lungs of ALI mice confirmed that administration of Ch-siIL6.1PS at a dose of 4.2 and 8.5 μg/g prevented the development of LPS-induced inflammatory changes in the respiratory system, with a 2.2- and 3.2-fold decrease in the intensity of inflammatory infiltration in the lung tissue compared to the control and a 1.7- and 2.5-fold decrease compared to Ch-siScrPS, respectively (Figure 3B). However, statistically significant differences were detected only for Ch-siIL6.1PS at a dose of 8.5 μg/g. Only residual inflammatory cells around the bronchi of experimental mice were detected. Assessing interstitial edema, it was also revealed that treatment of ALI mice with Ch-siIL6.1PS at a dose of 4.2 and 8.5 μg/g led to a 1.4- and 1.6-fold decrease in the volume density of alveolar septa compared to the control and a 1.3- and 1.5-fold decrease compared to Ch-siScrPS, respectively, causing almost normalization of this parameter (Figure 3C). All the identified differences were statistically significant. Thus, our findings clearly demonstrated that Ch-siIL6.1PS at a dose of 8.5 μg/g effectively prevents the development of LPS-induced ALI.
4. Discussion
Acute inflammation is a protective response to infection or tissue damage, but chronic inflammation, on the contrary, can lead to tissue damage and the development of pathological conditions such as fibrosis, autoimmune processes, malignant transformation, and metastasis [25,26]. Corticosteroids are widely used to treat chronic inflammatory diseases, but their effectiveness as an anti-inflammatory drug is limited by steroid drug resistance, which occurs or develops in a significant proportion of patients [36]. Long-term use of corticosteroids, which are required to treat chronic inflammation, can lead to serious side effects, causing immunosuppression, hypertension, diabetes, and adrenal dysfunction manifested by excess cortisol production [37]. The use of specific antibodies to combat inflammation has shown promising results, but the technology is not yet free from unwanted side effects [38]. So, for example, antidrug antibodies were detected in 53% of patients taking adalimumab (a fully humanized IgG1 anti-TNFa monoclonal antibody [22]), and high levels of antidrug antibodies were shown to lead to a reduced clinical response [39]. siRNA-based drugs are much less toxic compared to small molecule drugs, and they mostly do not cause an immune response to siRNA as antibodies do [11,12,13,14]. Antidrug antibodies were detected only in 0.9%, 6.0%, 1.7%, and 2.5% of patients receiving siRNA-based drugs givosiran, lumasiran, inclisiran, and vutrisiran, respectively, and they did not have a significant effect on PK, PD, efficacy, or safety [40,41,42]. Therefore, the development of drugs based on siRNA conjugates for the treatment of inflammatory diseases that are refractory to standard therapy is promising.
Previously, we demonstrated the possibility of reducing the severity of acute injury in the lungs after intranasal instillation of anti-Timp1 siRNA complexed with cationic liposomes [35]. We showed that the reduction of inflammation was accompanied by a reduced number of neutrophils in the BAL fluid. We also showed that silencing of Timp1 expression by siRNA led to a significant decrease in Il6 mRNA level, which can prevent leukocyte chemotaxis into lung tissue. IL6 is a multifunctional cytokine that plays an important role in a wide range of biological processes in various cell types, including tumor cells. There are evidences in the literature that deregulated IL6 expression is associated with tumor progression through inhibition of cancer cell apoptosis, stimulation of angiogenesis, and drug resistance [43]. In chronic inflammation, IL6 plays a detrimental role by promoting the accumulation of mononuclear cells at the site of injury through persistent MCP-1 secretion, angioproliferation, and inhibition of T cell apoptosis. Circulating levels of IL6 are elevated in a number of inflammatory diseases, including rheumatoid arthritis, systemic lupus erythematosus, and Crohn's disease [7,44]. Blocking IL6 signaling is a potential strategy for treating cancers characterized by pathological overproduction of IL6.
In this study, we directly silenced Il6 and found that this approach was effective in reducing the severity of ALI. We showed that cholesterol-conjugated anti-Il6 siRNA, selected based on the results of preliminary in vitro screening, has a dose-dependent therapeutic effect in LPS-induced ALI, reducing the number of cells in BAL fluid and the level of Il6 mRNA in them, as well as reducing the severity of inflammatory and edema changes in the lungs. The treatment of mice by anti-Il6 siRNA not only significantly changed the proportion of neutrophils in BAL but also reduced the absolute number of leukocytes compared with LPS-challenged mice, which was not achieved by the anti-Timp1 siRNA treatment. The relative change in inflammatory processes evaluated by histological scoring also showed positive trends: 2.5 and 3.2 folds for anti-Timp1 and anti-Il6 siRNAs, respectively [35]. However, these differences in therapy efficacy may also be due to the chosen targets as well as to different delivery methods and target cells. In a previous study [35], anti-Timp1 delivery to the target lung cells was performed using cationic lipids. In this study, we delivered anti-Il6 siRNA by covalent attachment to cholesterol in order to deliver it to alveolar macrophages. It should be noted that we also tried delivering anti-Il6 siRNA with cationic lipids; however, our preliminary data showed that delivery with a cholesterol conjugate was 44% more effective in reducing the level of Il6 mRNA in BAL fluid cells than siRNA /liposome complex [data not presented].
Cytokines, including IL6, are quite difficult targets for regulation by siRNAs because their expression increases rapidly and produces significant amount of mRNA, the translation products of which are secreted from the cell and have an effect on surrounding tissues or even the entire organism. In this work, we applied Ch-siIL6.1PS to prevent ALI in mice in pre-treatment mode; however, it is basically impossible to predict the rapid development of ALI except in cases of epidemic or pathogen exposure. Therefore, the question of whether the developed siRNA can be used in therapeutic regimens during the development of the disease remains open. This may be hampered by the relatively slow development of silencing effect by cholesterol conjugates of siRNA compared to siRNA/liposome complexes because the conjugates are entrapped in the endosomes [45,46]. On the other hand, the ability to avoid the use of lipid delivery systems, which themselves can have an immunostimulating effect and cause side effects, may increase the safety of the preparation.
Two main factors limit the length of time during which a single administration of siRNA can prevent the development of ALI: the nuclease resistance of siRNA and the lifetime of target cells in the lungs. Since siRNA is fully protected by 2’F or 2’OMe modifications, as well as PS at the ends of the duplex, it can be expected that its nuclease resistance will allow it to cause silencing within several months, as has been documented for a drug with similar modifications in the liver [32,47]. However, the second factor more strongly limits the duration of action of Ch-siIL6.1PS in the lungs after local application. The primary target of siRNA is resident alveolar macrophages, which, in response to LPS, attract macrophages and monocytes from the bloodstream into the lung tissue. Alveolar macrophages reside in the lungs for 1-7 weeks [48], therefore, we should not expect the effect of one instillation of Ch-siIL6.1PS to last more than a month. This purely theoretical assessment requires a more detailed experimental evaluation.
Thus, the designed cholesterol-conjugated anti-Il6 siRNA showed its high potential for preventing the development of ALI. The findings indicate that IL6 inhibition is a productive strategy to combat inflammatory diseases. It can be assumed that such a strategy could be applied to the treatment of inflammation-associated diseases beyond the lungs including inflammatory bowel disease [49], hepatic cirrhosis [50], sepsis [51], rheumatoid arthritis [52], psoriasis [53] and others. Moreover, further structural and chemical engineering of the anti-Il6 siRNA and fine tuning of the delivery systems may expand the scope of its application.
Author Contributions
Conceptualization, E.L.C. and I.V.C.; Funding acquisition, E.L.C.; Investigation, I.V.C., I.K.B. and A.V.S.; M.I.M. synthesized the siRNA and their derivatives; I.K.B. conducted the in vitro studies; I.V.C., I.K.B., A.V.S. and I.A.S. conducted the in vivo studies. Resources, M.A.Z. and V.V.V.; Writing—original draft, I.V.C., I.K.B. and A.V.S.; Writing—review and editing, E.L.C. and M.A.Z.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation (grant #19-74-30011) and project ICBFM SB RAS #121031300044-5 (synthesis of control siRNAs).
Institutional Review Board Statement
The animal study protocol was approved by the Committee on the Ethics of Animal Experiments of the Administration of the Siberian Branch of the Russian Academy of Sciences (Novosibirsk, Russia) (protocol No. 56 from August 10, 2019).
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors gratefully thank IIya S. Dovydenko for siRNA synthesis, Albina V. Vladimirova for cell maintenance, Alexandra G. Mozhnaya for animal handling and Marina A. Zenkova Laboratory members for their support (Institute of Chemical Biology and Fundamental Medicine SB RAS, Novosibirsk, Russia).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dana, H.; Mahmoodi Chalbatani, G.; Mahmoodzadeh, H.; Karimloo, R.; Rezaiean, O.; Moradzadeh, A.; Mehmandoost, N.; Moazzen, F.; Mazraeh, A.; Marmari, V.; et al. Molecular Mechanisms and Biological Functions of SiRNA. Int. J. Biomed. Sci. 2017, 13, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.J. Therapeutic SiRNA: State of the Art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.M.; Nair, J.K.; Janas, M.M.; Anglero-Rodriguez, Y.I.; Dang, L.T.H.; Peng, H.; Theile, C.S.; Castellanos-Rizaldos, E.; Brown, C.; Foster, D.; et al. Expanding RNAi Therapeutics to Extrahepatic Tissues with Lipophilic Conjugates. Nat. Biotechnol. 2022, 40, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Alterman, J.F.; Godinho, B.M.D.C.; Hassler, M.R.; Ferguson, C.M.; Echeverria, D.; Sapp, E.; Haraszti, R.A.; Coles, A.H.; Conroy, F.; Miller, R.; et al. A Divalent SiRNA Chemical Scaffold for Potent and Sustained Modulation of Gene Expression throughout the Central Nervous System. Nat. Biotechnol. 2019, 37, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, J.E.; Gale, A.; Wu, P.; Ma, R.; Davis, M.E. SiRNA Delivery to the Glomerular Mesangium Using Polycationic Cyclodextrin Nanoparticles Containing SiRNA. Nucleic Acid Ther. 2015, 25, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Thai, H.B.D.; Kim, K.R.; Hong, K.T.; Voitsitskyi, T.; Lee, J.S.; Mao, C.; Ahn, D.R. Kidney-Targeted Cytosolic Delivery of SiRNA Using a Small-Sized Mirror DNA Tetrahedron for Enhanced Potency. ACS Cent. Sci. 2020, 6, 2250–2258. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Hardie, J.; Liu, Y.; Ray, M.; Luo, X.; Das, R.; Landis, R.F.; Farkas, M.E.; Rotello, V.M. Nanocapsule-Mediated Cytosolic SiRNA Delivery for Anti-Inflammatory Treatment. J. Control. Release 2018, 283, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.B.; Kos, P.; Tieu, V.; Zhou, K.; Siegwart, D.J. Development of Cationic Quaternary Ammonium Sulfonamide Amino Lipids for Nucleic Acid Delivery. ACS Appl. Mater. Interfaces 2018, 10, 2302–2311. [Google Scholar] [CrossRef] [PubMed]
- Chernikov, I. V.; Vlassov, V. V.; Chernolovskaya, E.L. Current Development of SiRNA Bioconjugates: From Research to the Clinic. Front. Pharmacol. 2019, 10, 444. [Google Scholar] [CrossRef]
- Eberle, F.; Giessler, K.; Deck, C.; Heeg, K.; Peter, M.; Richert, C.; Dalpke, A.H. Modifications in Small Interfering RNA That Separate Immunostimulation from RNA Interference. J. Immunol. 2008, 180(5), 3229–3237. [Google Scholar] [CrossRef]
- Sioud, M.; Furset, G.; Cekaite, L. Suppression of Immunostimulatory SiRNA-Driven Innate Immune Activation by 2′-Modified RNAs. Biochem. Biophys. Res. Commun. 2007, 361, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, M.; Akinc, A.; Pandey, R.K.; Qin, J.; Hadwiger, P.; John, M.; Mills, K.; Charisse, K.; Maier, M.A.; Nechev, L.; et al. Unique Gene-Silencing and Structural Properties of 2′-Fluoro-Modified SiRNAs. Angew. Chemie Int. Ed. 2011, 50, 2284–2288. [Google Scholar] [CrossRef]
- Judge, A.D.; Bola, G.; Lee, A.C.H.; MacLachlan, I. Design of Noninflammatory Synthetic SiRNA Mediating Potent Gene Silencing in Vivo. Mol. Ther. 2006, 13, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Robbins, M.; Judge, A.; Liang, L.; McClintock, K.; Yaworski, E.; MacLachlan, I. 2′-O-Methyl-Modified RNAs Act as TLR7 Antagonists. Mol. Ther. 2007, 15, 1663–1669. [Google Scholar] [CrossRef]
- Chernikov, I. V.; Gladkikh, D. V.; Meschaninova, M.I.; Ven’yaminova, A.G.; Zenkova, M.A.; Vlassov, V. V.; Chernolovskaya, E.L. Cholesterol-Containing Nuclease-Resistant SiRNA Accumulates in Tumors in a Carrier-Free Mode and Silences MDR1 Gene. Mol. Ther. - Nucleic Acids 2017, 6, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Biscans, A.; Coles, A.; Haraszti, R.; Echeverria, Di.; Hassler, M.; Osborn, M.; Khvorova, A. Diverse Lipid Conjugates for Functional Extra-Hepatic SiRNA Delivery in Vivo. Nucleic Acids Res. 2019, 47, 1082–1096. [Google Scholar] [CrossRef] [PubMed]
- Chernikov, I. V.; Meschaninova, M.I.; Gladkikh, D. V.; Ven’yaminova, A.G.; Zenkova, M.A.; Vlassov, V. V.; Chernolovskaya, E.L. Interaction of Lipophilic Conjugates of Modified SiRNAs with Hematopoietic Cells In Vitro and In Vivo. Russ. J. Bioorganic Chem. 2021, 47, 399–410. [Google Scholar] [CrossRef]
- Cooper, G.S.; Bynum, M.L.K.; Somers, E.C. Recent Insights in the Epidemiology of Autoimmune Diseases: Improved Prevalence Estimates and Understanding of Clustering of Diseases. J. Autoimmun. 2009, 33, 197–207. [Google Scholar] [CrossRef] [PubMed]
- El-Gabalawy, H.; Guenther, L.C.; Bernstein, C.N. Epidemiology of Immune-Mediated Inflammatory Diseases: Incidence, Prevalence, Natural History, and Comorbidities. J. Rheumatol. 2010, 37, 2–10. [Google Scholar] [CrossRef]
- Lerner, A.; Jeremias, P.; Matthias, T. The World Incidence and Prevalence of Autoimmune Diseases Is Increasing. Int. J. Celiac Dis. 2015, 3, 151–155. [Google Scholar] [CrossRef]
- Yasunaga, M. Antibody Therapeutics and Immunoregulation in Cancer and Autoimmune Disease. Semin. Cancer Biol. 2020, 64, 1–12. [Google Scholar] [CrossRef]
- Li, P.; Zheng, Y.; Chen, X. Drugs for Autoimmune Inflammatory Diseases: From Small Molecule Compounds to Anti-TNF Biologics. Front. Pharmacol. 2017, 8, 460. [Google Scholar] [CrossRef]
- Connor, V. Anti-TNF Therapies: A Comprehensive Analysis of Adverse Effects Associated with Immunosuppression. Rheumatol. Int. 2011, 31, 327–337. [Google Scholar] [CrossRef]
- Yu, M.B.; Firek, A.; Langridge, W.H.R. Predicting Methotrexate Resistance in Rheumatoid Arthritis Patients. Inflammopharmacology 2018, 26, 699–708. [Google Scholar] [CrossRef]
- Ragaller, M.; Richter, T. Acute Lung Injury and Acute Respiratory Distress Syndrome. J. Emergencies, Trauma Shock 2010, 3, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Gotzev, R.; Kenarov, P. Acute Respiratory Distress Syndrome (ARDS). Anaesthesiol. Intensive Care 2013, 42, 43–49. [Google Scholar]
- Zoulikha, M.; Xiao, Q.; Boafo, G.F.; Sallam, M.A.; Chen, Z.; He, W. Pulmonary Delivery of SiRNA against Acute Lung Injury/Acute Respiratory Distress Syndrome. Acta Pharm. Sin. B 2022, 12, 600–620. [Google Scholar] [CrossRef]
- Trovato, M.; Sciacchitano, S.; Facciolà, A.; Valenti, A.; Visalli, G.; Di Pietro, A. Interleukin-6 Signalling as a Valuable Cornerstone for Molecular Medicine (Review). Int. J. Mol. Med. 2021, 47, 107. [Google Scholar] [CrossRef] [PubMed]
- Florentin, J.; Zhao, J.; Tai, Y.Y.; Vasamsetti, S.B.; O’Neil, S.P.; Kumar, R.; Arunkumar, A.; Watson, A.; Sembrat, J.; Bullock, G.C.; et al. Interleukin-6 Mediates Neutrophil Mobilization from Bone Marrow in Pulmonary Hypertension. Cell. Mol. Immunol. 2021, 18, 374–384. [Google Scholar] [CrossRef]
- Chen, I.C.; Wang, S.C.; Chen, Y.T.; Tseng, H.H.; Liu, P.L.; Lin, T.C.; Wu, H.E.; Chen, Y.R.; Tseng, Y.H.; Hsu, J.H.; et al. Corylin Ameliorates Lps-Induced Acute Lung Injury via Suppressing the Mapks and Il-6/Stat3 Signaling Pathways. Pharmaceuticals 2021, 14, 1046. [Google Scholar] [CrossRef]
- Meschaninova, M.I.; Novopashina, D.S.; Semikolenova, O.A.; Silnikov, V.N.; Venyaminova, A.G. Novel Convenient Approach to the Solid-Phase Synthesis of Oligonucleotide Conjugates. Molecules 2019, 24, 4266. [Google Scholar] [CrossRef] [PubMed]
- Foster, D.J.; Brown, C.R.; Shaikh, S.; Trapp, C.; Schlegel, M.K.; Qian, K.; Sehgal, A.; Rajeev, K.G.; Jadhav, V.; Manoharan, M.; et al. Advanced SiRNA Designs Further Improve In Vivo Performance of GalNAc-SiRNA Conjugates. Mol. Ther. 2018, 26, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Chernikov, I. V.; Ponomareva, U.A.; Meschaninova, M.I.; Bachkova, I.K.; Teterina, A.A.; Gladkikh, D. V.; Savin, I.A.; Vlassov, V. V.; Zenkova, M.A.; Chernolovskaya, E.L. Cholesterol-Conjugated Supramolecular Multimeric SiRNAs: Effect of SiRNA Length on Accumulation and Silencing In Vitro and In Vivo. Nucleic Acid Ther. 2023, 33, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Hassler, M.R.; Turanov, A.A.; Alterman, J.F.; Haraszti, R.A.; Coles, A.H.; Osborn, M.F.; Echeverria, D.; Nikan, M.; Salomon, W.E.; Roux, L.; et al. Comparison of Partially and Fully Chemically-Modified SiRNA in Conjugate-Mediated Delivery in Vivo. Nucleic Acids Res. 2018, 46, 2185–2196. [Google Scholar] [CrossRef] [PubMed]
- Chernikov, I. V.; Staroseletz, Y.Y.; Tatarnikova, I.S.; Sen’kova, A. V.; Savin, I.A.; Markov, A. V.; Logashenko, E.B.; Chernolovskaya, E.L.; Zenkova, M.A.; Vlassov, V. V. SiRNA-Mediated Timp1 Silencing Inhibited the Inflammatory Phenotype during Acute Lung Injury. Int. J. Mol. Sci. 2023, 24, 1641. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Dong, J.; Peh, H.Y.; Tan, L.H.; Lim, K.S.; Li, L.; Wong, W.S.F. Oligonucleotide Therapy for Obstructive and Restrictive Respiratory Diseases. Molecules 2017, 22, 139. [Google Scholar] [CrossRef]
- Gupta, R.; Fonacier, L.S. Adverse Effects of Nonsystemic Steroids (Inhaled, Intranasal, and Cutaneous): A Review of the Literature and Suggested Monitoring Tool. Curr. Allergy Asthma Rep. 2016, 16, 44. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Chandley, P.; Rohatgi, S. Recent Advances in the Development of Monoclonal Antibodies and Next-Generation Antibodies. ImmunoHorizons 2023, 7, 886–897. [Google Scholar] [CrossRef] [PubMed]
- van Schouwenburg, P.A.; Bartelds, G.M.; Hart, M.H.; Aarden, L.; Wolbink, G.J.; Wouters, D. A Novel Method for the Detection of Antibodies to Adalimumab in the Presence of Drug Reveals “Hidden” Immunogenicity in Rheumatoid Arthritis Patients. J. Immunol. Methods 2010, 362, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, J.L.S.; Chan, A.; Sehgal, A.; Butler, J.S.; Nair, J.K.; Racie, T.; Shulga-Morskaya, S.; Nguyen, T.; Qian, K.; Yucius, K.; et al. Evaluation of GalNAc-SiRNA Conjugate Activity in Pre-Clinical Animal Models with Reduced Asialoglycoprotein Receptor Expression. Mol. Ther. 2018, 26, 105–114. [Google Scholar] [CrossRef]
- Corydon, I.J.; Fabian-Jessing, B.K.; Jakobsen, T.S.; Jørgensen, A.C.; Jensen, E.G.; Askou, A.L.; Aagaard, L.; Corydon, T.J. 25 Years of Maturation: A Systematic Review of RNAi in the Clinic. Mol. Ther. - Nucleic Acids 2023, 33, 469–482. [Google Scholar] [CrossRef]
- An, G. Pharmacokinetics and Pharmacodynamics of GalNAc-Conjugated SiRNAs. J. Clin. Pharmacol. 2024, 64, 45–57. [Google Scholar] [CrossRef]
- Ene, C.V.; Nicolae, I.; Geavlete, B.; Geavlete, P.; Ene, C.D. IL-6 Signaling Link between Inflammatory Tumor Microenvironment and Prostatic Tumorigenesis. Anal. Cell. Pathol. 2022, 2022, 5980387. [Google Scholar] [CrossRef]
- Roda, G.; Chien Ng, S.; Kotze, P.G.; Argollo, M.; Panaccione, R.; Spinelli, A.; Kaser, A.; Peyrin-Biroulet, L.; Danese, S. Crohn’s Disease. Nat. Rev. Dis. Prim. 2020, 6, 23. [Google Scholar] [CrossRef]
- Petrova, N.S.; Chernikov, I. V.; Meschaninova, M.I.; Dovydenko, I.S.; Venyaminova, A.G.; Zenkova, M.A.; Vlassov, V. V.; Chernolovskaya, E.L. Carrier-Free Cellular Uptake and the Gene-Silencing Activity of the Lipophilic SiRNAs Is Strongly Affected by the Length of the Linker between SiRNA and Lipophilic Group. Nucleic Acids Res. 2012, 40, 2330–2344. [Google Scholar] [CrossRef]
- Gilleron, J.; Querbes, W.; Zeigerer, A.; Borodovsky, A.; Marsico, G.; Schubert, U.; Manygoats, K.; Seifert, S.; Andree, C.; Stöter, M.; et al. Image-Based Analysis of Lipid Nanoparticle-Mediated SiRNA Delivery, Intracellular Trafficking and Endosomal Escape. Nat. Biotechnol. 2013, 31, 638–646. [Google Scholar] [CrossRef]
- Nair, J.K.; Attarwala, H.; Sehgal, A.; Wang, Q.; Aluri, K.; Zhang, X.; Gao, M.; Liu, J.; Indrakanti, R.; Schofield, S.; et al. Impact of Enhanced Metabolic Stability on Pharmacokinetics and Pharmacodynamics of GalNAc-SiRNA Conjugates. Nucleic Acids Res. 2017, 45, 10969–10977. [Google Scholar] [CrossRef]
- van oud Alblas, A.B.; van Furth, R. Origin, Kinetics, and Characteristics in the Normal Macrophages Steady State. J Exp Med. 1979, 149, 1504–1518. [Google Scholar] [CrossRef]
- Veiga, N.; Goldsmith, M.; Diesendruck, Y.; Ramishetti, S.; Rosenblum, D.; Elinav, E.; Behlke, M.A.; Benhar, I.; Peer, D. Leukocyte-Specific SiRNA Delivery Revealing IRF8 as a Potential Anti-Inflammatory Target. J. Control. Release 2019, 313, 33–41. [Google Scholar] [CrossRef]
- Zhang, J.; Shen, H.; Xu, J.; Liu, L.; Tan, J.; Li, M.; Xu, N.; Luo, S.; Wang, J.; Yang, F.; et al. Liver-Targeted SiRNA Lipid Nanoparticles Treat Hepatic Cirrhosis by Dual Antifibrotic and Anti-Inflammatory Activities. ACS Nano 2020, 14, 6305–6322. [Google Scholar] [CrossRef]
- Lee, J.; Son, W.; Hong, J.; Song, Y.; Yang, C.S.; Kim, Y.H. Down-Regulation of TNF-α via Macrophage-Targeted RNAi System for the Treatment of Acute Inflammatory Sepsis. J. Control. Release 2021, 336, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, R.; Huo, S.; Chen, H.; Song, Q.; Jiang, G.; Yu, Y.; Huang, J.; Xie, S.; Gao, X.; et al. CaP-Based Anti-Inflammatory HIF-1α SiRNA-Encapsulating Nanoparticle for Rheumatoid Arthritis Therapy. J. Control. Release 2022, 343, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.; Kumbhojkar, N.; Reilly, C.; Dharamdasani, V.; Ukidve, A.; Ingber, D.E.; Mitragotri, S. Treatment of Psoriasis with NFKBIZ SiRNA Using Topical Ionic Liquid Formulations. Sci. Adv. 2020, 6, eabb6049. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Silencing activity of anti-Il6 siRNAs and their cholesterol derivatives in J774 cells. (A) Experimental setup. (B) Relative Il6 mRNA levels in J774 cells after LPS stimulation (1 μg/ml), siRNA transfection was carried out with Lipofectamine 2000. Mean values (±SD) and statistical significance of differences from LPS-treated cells (** - p < 0.01, *** p < 0.01), calculated from the results of three independent experiments, are shown in the figure. (C) Schematic structure of cholesterol conjugate.
Figure 1.
Silencing activity of anti-Il6 siRNAs and their cholesterol derivatives in J774 cells. (A) Experimental setup. (B) Relative Il6 mRNA levels in J774 cells after LPS stimulation (1 μg/ml), siRNA transfection was carried out with Lipofectamine 2000. Mean values (±SD) and statistical significance of differences from LPS-treated cells (** - p < 0.01, *** p < 0.01), calculated from the results of three independent experiments, are shown in the figure. (C) Schematic structure of cholesterol conjugate.
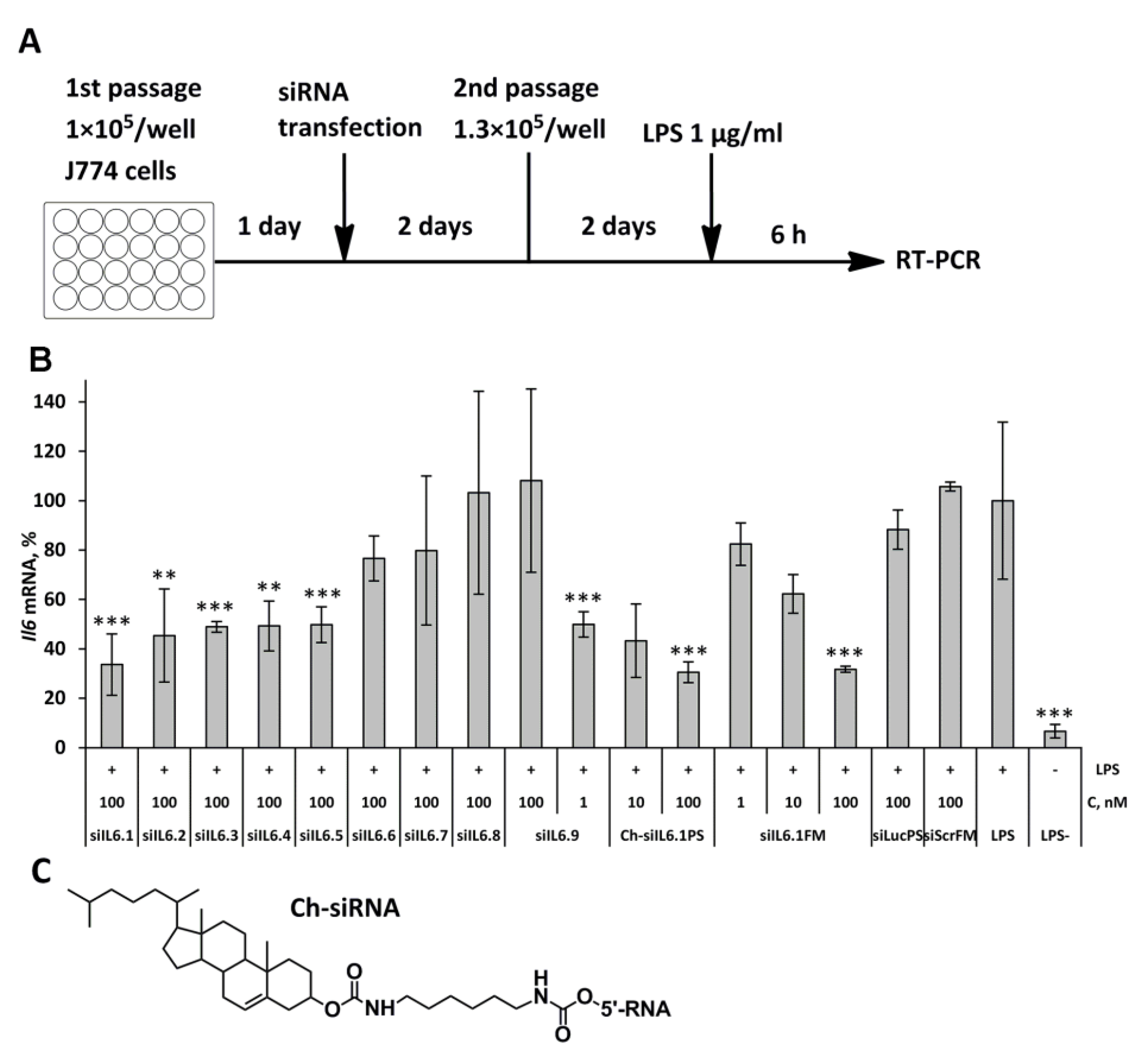
Figure 2.
Biological effects of cholesterol-modified anti-Il6 siRNA (Ch-siIL6.1PS) on the development of ALI. (A) Experimental setup. (B) Il6 mRNA levels in BAL fluid cells. (C) Total and (D) differential leukocyte counts in the BAL fluid of healthy (LPS-) and LPS-challenged mice after Ch-siIL6.1PS administration and without treatment. Data are presented as mean ± standard error of mean, n = 3-4, difference from LPS-challenged mice: * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 2.
Biological effects of cholesterol-modified anti-Il6 siRNA (Ch-siIL6.1PS) on the development of ALI. (A) Experimental setup. (B) Il6 mRNA levels in BAL fluid cells. (C) Total and (D) differential leukocyte counts in the BAL fluid of healthy (LPS-) and LPS-challenged mice after Ch-siIL6.1PS administration and without treatment. Data are presented as mean ± standard error of mean, n = 3-4, difference from LPS-challenged mice: * p < 0.05, ** p < 0.01, *** p < 0.001.
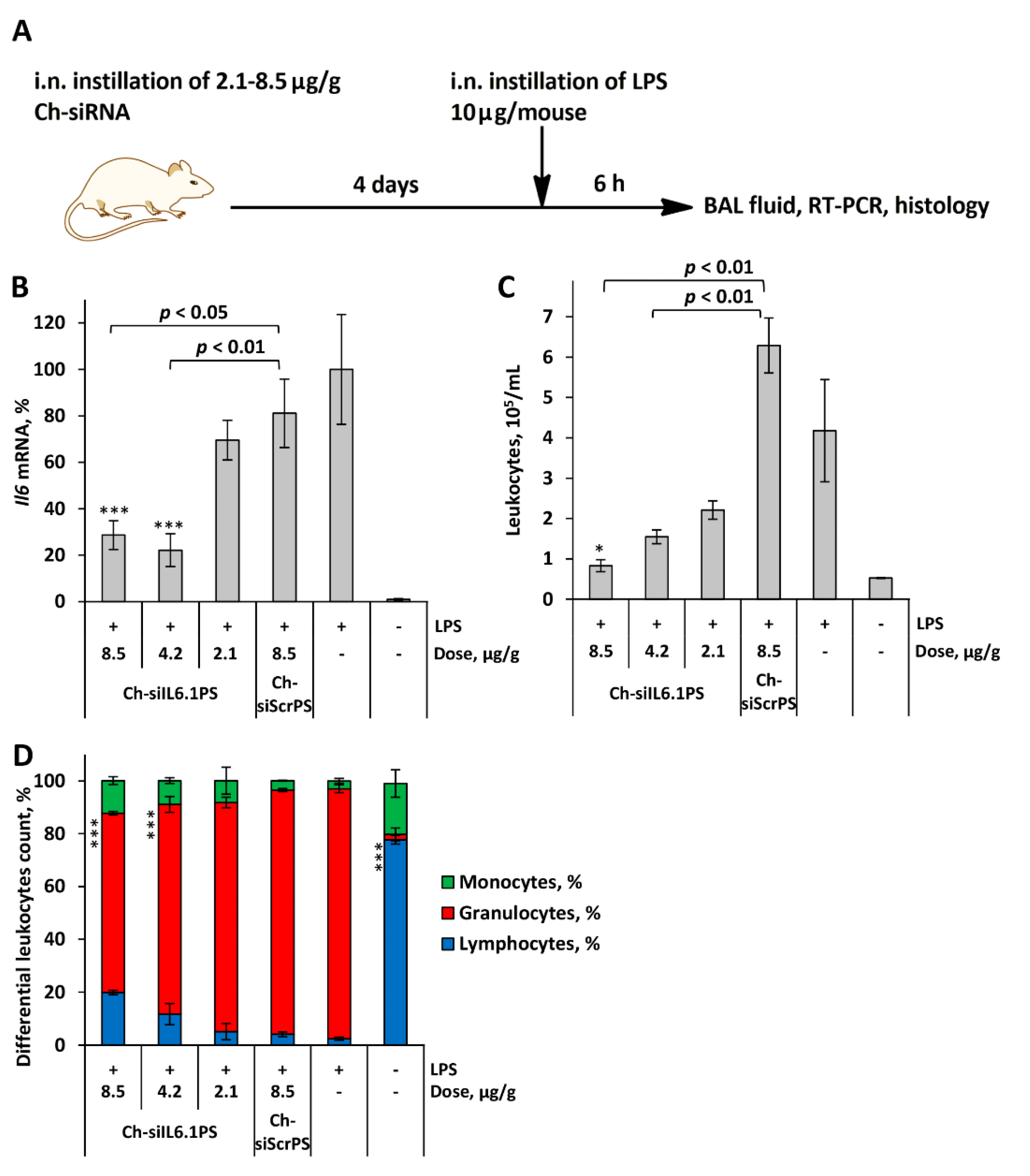
Figure 3.
Ch-siIL6.1PS effectively suppresses LPS-induced lung inflammation in vivo. (A) Representative histological images of lung tissue from ALI mice. Hematoxylin and eosin staining. Original magnification ×200. Black arrows indicate inflammatory infiltration in the lung tissue. (B) The intensity of inflammatory infiltration in the lung tissue of LPS-challenged mice calculated by the semi-quantitative histological scoring system, where 0 is no inflammatory infiltration, 1 is low intensity of inflammatory infiltration, 2 is moderate intensity of inflammatory infiltration, and 3 is high intensity of inflammatory infiltration. (С) The volume densities (Vv, %) of alveolar septa reflecting the intensity of interstitial edema in the lung tissue of LPS-challenged mice without treatment and after Ch-siIL6.1PS administration. All quantifications were performed in 5 random fields in each lung sample, forming 10-20 random fields from each experimental group. Data are presented as mean ± standard error of mean, n = 3-5, difference from LPS-challenged mice: * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 3.
Ch-siIL6.1PS effectively suppresses LPS-induced lung inflammation in vivo. (A) Representative histological images of lung tissue from ALI mice. Hematoxylin and eosin staining. Original magnification ×200. Black arrows indicate inflammatory infiltration in the lung tissue. (B) The intensity of inflammatory infiltration in the lung tissue of LPS-challenged mice calculated by the semi-quantitative histological scoring system, where 0 is no inflammatory infiltration, 1 is low intensity of inflammatory infiltration, 2 is moderate intensity of inflammatory infiltration, and 3 is high intensity of inflammatory infiltration. (С) The volume densities (Vv, %) of alveolar septa reflecting the intensity of interstitial edema in the lung tissue of LPS-challenged mice without treatment and after Ch-siIL6.1PS administration. All quantifications were performed in 5 random fields in each lung sample, forming 10-20 random fields from each experimental group. Data are presented as mean ± standard error of mean, n = 3-5, difference from LPS-challenged mice: * p < 0.05, ** p < 0.01, *** p < 0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



