
Submitted:
23 April 2024
Posted:
24 April 2024
You are already at the latest version
Abstract
Abiotic stress is responsible for significant reduction in crop plant productivity worldwide. Ultraviolet (UV) radiation is a natural component of sunlight and a permanent environmental stimulus. This study investigated the distinct responses of young wheat and einkorn plants to excessive UV-B radiation (180 min at λmax 312 nm) following foliar pretreatment with 1 µM synthetic cytokinin 4PU-30. Results demonstrated that UV radiation significantly amplified hydrogen peroxide levels in both wheat and einkorn, with einkorn exhibiting more pronounced increase compared to wheat. This elevation indicated the induction of oxidative stress by UV radiation in the two genotypes. Intensified antioxidant enzyme activities and increased accumulation of typical stress markers and non-enzyme protectants were evidenced. Transcriptional activity of genes encoding key antioxidant enzymes was also investigated aiming to elucidate the genetic regulation of the antioxidant systems in both wheat and einkorn seedlings. Our results suggested a role for POX1 and POX7 genes in the UV-B tolerance of the two wheat species as well as a cytokinin stimulated UV-B stress response in einkorn mediated through upregulation of the tau subfamily gene GSTU6. Based on these findings it could be concluded that 4PU-30 had the potential of alleviating oxidative stress by attenuating the symptoms of superfluous UV-B illumination in the two examined plant species.
Keywords:
antioxidant gene expression
; flavonoids
; oxidative stress
; phenolic compounds
; phenylurea-cytokinin
; proline
; remote sensing
; ROS
; spectral reflectance
1. Introduction
Wheat (Triticum aestivum, L.) is among the first domesticated plant species by mankind and has become a crop of key agronomical importance nowadays supplying the primary food sources for humans worldwide [1]. In recent years, young cereal seedlings are being used in healthy food industry as an innovative and fashionable alternative of tasty ready-to-eat vegetables rich in various bioactive compounds [2]. The increasing interest in healthful nutrition diets and higher quality of life determines the pursuit of specialized food products based on alternative cereal species like einkorn and emmer. Einkorn (Triticum monococcum, L.) is an ancient wheat species which possesses superior nutritional value and better technological characteristics of the grain compared to extensively grown modern wheat varieties [1,3,4]. Despite their lower seed productivity, ancient wheats have survived in adverse environmental conditions for many years and are presumably more resistant to stress factors compared to cultivated wheat which disclose possibilities for their implementation in organic farming [5].
Sudden changes in environmental factors have negative impact on plant growth and development. Abiotic stress could be developed in response to soil salinity, water deprivation, high temperatures or a combination of multiple factors. Although a natural component of sunlight and a portion of the non-ionizing region of the electromagnetic spectrum (comprising about 9 % of the emitted solar radiation), ultraviolet (UV) radiation is a strong environmental signal and a powerful abiotic stress factor [6]. While mostly attenuated by the stratospheric ozone layer, part of UV-B radiation (280-315 nm) penetrates deeper reaching the Earth’s surface and is more dangerous owing to its high energy and ability to interact with organic molecules. Exposure to excessive UV-B could be highly damaging for the plant at many organizational levels. At the genomic level, UV-B light generates DNA lesions, predominantly pyrimidine dimers, that can directly block DNA replication and RNA synthesis [7,8]. Additionally, UV-B induces the formation of reactive oxygen species (ROS) which are highly active and cause the oxidation of chief cellular components such as proteins and lipids eventually leading to oxidative stress. At a higher organizational level, UV-B impairs the structure and functioning of the photosynthetic apparatus, causes leaf wilting, yellowing and even death of the whole plant organism [9,10].
Plants have evolved a variety of mechanisms to limit UV-B induced lesions and promote acclimation to excessive radiation [6,11]. Defense resources for coping with the negative consequences of oxidative stress are aimed to detoxify ROS, reduce cellular damage, and provide DNA repair. They involve the biosynthesis of enzymatic and non-enzymatic protective molecules. Although the accumulation of low molecular solutes like proline, sugars and polyols is a common stress response, it could hardly minimize oxidative injury and so these substances are rightfully regarded as stress markers [12]. Another endogenous cellular constituent like malondialdehyde (MDA) is frequently used as stress reporter molecule representing a product of lipid peroxidation [13,14]. Non-enzymatic protection also comes from increasing levels of biological compounds such as ascorbic acid, phenolics, flavonoids, glutathione and proteins that carry redox active S -groups and performing like antioxidants. On the other hand, enzymes involved in antioxidant protection limit the consequences of oxidative stress by their ability to scavenge free radicals and toxic amounts of ROS either through direct decomposition and balancing cellular redox dynamics or prevent their formation by regulating gene expression and proteolysis [13].
Cytokinins (CKs) are phytohormones involved in various aspects of plant growth and development including cell division and differentiation, shoot and root morphogenesis, but were also shown to mediate plant stress responses [15,16,17,18,19,20]. The synthetic phenylurea type cytokinin 4PU-30 (N1-(2-chloro-4-pyridyl)-N2-phenylurea) was reported to reduce the negative effects of drought [21,22] and salinity [23,24], alleviate the action of the herbicide glyphosate [25,26] and provide protection against herbicides and biotic stress [25,27]. Application of growth regulators capable of activating the plant defense system might be regarded as an alternative approach to developing stress tolerance in important crop species [28,29].
Changes in gene expression related to antioxidant enzyme activities are regarded as a hallmark of plant oxidative stress tolerance. Modulated expression pattern of the genes encoding antioxidant enzymes such as catalase (CAT), peroxidase (POX), superoxide dismutase (SOD), and glutathione S-transferase (GST) in response to enhanced ROS production has been reported in several plants, including wheat, and after exposure to variety of stresses [30,31,32,33,34]. SOD genes were evidenced as highly responsive to abiotic stress and enhancing plant resistance. Their individual expression patterns, however, have revealed stress-specific differences and variations among species [35,36]. UV-B exposure triggered dynamic transcriptional activity of SOD genes with strong upregulation confirmed only for certain mRNAs [37] suggesting that the regulatory role at the gene expression level is crucial for establishing stress tolerance [38]. Increased transcript levels were found in einkorn plants subjected to heavy metal toxicity consistent with the stress-responsive cis-regulatory elements identified in the promoter region of TmMnSOD [39]. Accumulated experimental data indicated that at the transcriptional level, most of the antioxidant genes exhibited high plasticity and complex regulatory features allowing plants to properly respond and precisely activate their antioxidant defense system [40,41,42].
The aim of the present work was to compare physiological responses of young wheat and einkorn plants to excessive UV-B radiation in laboratory experiments and to explore the capacity of exogenously applied synthetic phenylurea type cytokinin 4PU-30 for alleviating oxidative stress damages. Furthermore, transcriptional activity of genes encoding certain antioxidant enzymes was measured to evaluate the genetic regulation of the antioxidant defense system. An additional focus of this paper was to use the possibilities of narrow-wavelength spectrometric measurements and to obtain spectral data in monitoring of wheat and einkorn plants under UV-B stress. For this purpose, remote sensing (RS) approach was applied which was previously tested in different experiments [43,44,45]. Here, spectral reflectance data acquired by a multichannel spectrometric system were used for establishing relation with unfavorable stress caused by UV-B radiation.
2. Results
2.1. Effect of Cytokinin Priming and Subsequent UV-B Treatment on Growth Parameters in Wheat and Einkorn Seedlings
UV-B treatment caused decrease in leaf length only in einkorn but not in wheat (Figure 1A,B) and reduced root length most notably in wheat plants (Figure 1C,D). 4PU-30 pretreatment led to increased leaf length in wheat and increased root length in einkorn plants (Figure 1A,D). Wheat plants treated with UV-B after cytokinin imposition showed slightly reduced root length compared to controls (Figure 1C) while in einkorn the combination of the two treatments led to increased root length (Figure 1D).
Pigment content in the leaves decreased after UV-B treatment in all variants (Table 1.). Chlorophyll a and b and carotenoids were more significantly reduced in wheat compared to einkorn leaves. Generally, 4PU-30 caused increase in chlorophylls and carotenoid amounts. The cytokinin also had positive effect on all pigments’ quantities after successive UV-B treatment compared to variants solely illuminated with UV-B radiation.
2.2. Effect of 4PU-30 and UV-B treatment on reflectance spectra of wheat and einkorn seedlings
To study the dynamics of wheat and einkorn control and treated plants at leaf level their reflectance spectra were obtained by remote sensing measurements. Intact, photosynthetically active plants usually exhibit a typical pattern of their spectral characteristics. Normally, a small peak in the green region of the visible spectrum (520-580 nm) is found that corresponds to the maximal chlorophyll reflectivity. After that, a decrease in the red range (640-680 nm) is seen, which is related to high chlorophyll absorption. Next well-manifested steep increase can be observed in the so-called “red edge” a range of electromagnetic spectrum (EMS) between 690-720 nm. In the near/shortwave infrared (NIR/SWIR) parts of EMS (720-2500 nm) the reflectance remains high up to 1200 nm and subsequently it starts to decrease. Both “red edge” and NIR/SWIR are considered as most informative regions of EMS for studying plants which was clearly expressed in wheat and einkorn averaged reflectance spectra (Figure 2A,B). It was seen that in wheat 4PU-30 did not affect the obtained reflectance spectra, while UV-B treatment and its combination with preliminary 4PU-30 application caused decrease in the reflectance response in the NIR region (Figure 2A). In einkorn, UV-B and its combination with 4PU-30 induced a significant increase in reflectance in the green spectral range (520-580 nm), but not in the “red edge” and NIR ranges (Figure 2B). In the NIR part of EMS, exposure to UV-B led to decrease in the reflectance while 4PU-30 treatment and its combination with UV-B resulted in visible increase of the registered reflectance (Figure 2B).
2.3. Effect of 4PU-30 and UV-B treatment on stress markers content in the leaves of wheat and einkorn seedlings
Analyses of oxidative damage and antioxidant status were performed on the first fully developed leaf of control, UV-B-treated and cytokinin-primed plants. Cytokinin priming had slightly supplementing effect in einkorn and no significant effect in wheat with respect to proline content. UV-B treatment caused increase in free proline levels in both plant genotypes, but it was greater in einkorn when compared to respective controls (Figure 3A,B). Following UV-B illumination proline content was increased with up to 35 % in wheat and 61 % in einkorn compared to the corresponding untreated plants. Proline values in einkorn seedlings subjected to a combination of 4PU-30 and consecutive UV-B were lower compared to UV-B treated variants and similar to controls and cytokinin-sprayed plants. In wheat, the combined treatment with cytokinin and UV-B stress led to slight increase in proline levels.
Cytokinin spraying did not significantly affect hydrogen peroxide levels in wheat and slightly reduced them in einkorn (Figure 3C,D). UV-B treatment led to increase in hydrogen peroxide values which were 30 % higher than controls in wheat and nearly 1,5-fold above einkorn controls (Figure 3C,D). The combination of cytokinin pretreatment followed by UV-B illumination decreased H2O2 in wheat but increased it in einkorn with respect to controls.
The amounts of MDA usually indicate the degree of membrane lipid peroxidation resulting from oxidative damage. MDA content only slightly increased in wheat after UV-B treatment but exhibited more than 2-fold growth in einkorn compared to respective controls (Figure 3E,F). Cytokinin spraying prior to the UV-B treatment reduced the amounts of this oxidation product in wheat compared to control and post-UV stress values but in respective einkorn variants MDA remained higher than controls.
2.4. Non-enzymatic antioxidants (free thiols, flavonoids, and total phenolic compounds)
The amount of free thiol group containing compounds, flavonoids, and total phenolics was measured in the first fully developed leaves of young wheat and einkorn plants. In wheat the levels of SH groups remained relatively unaffected by the applied treatments but a nearly 2-fold increase in free thiols compared to control was detected in einkorn plants after UV-B irradiation. However, they returned to controls after the combined action of 4PU-30 and illumination (Figure 4A,B).
Flavonoid content was only slightly affected by UV-B treatment in wheat. In einkorn, cytokinin spraying evoked 23 % decrease in flavonoids and the consecutive treatment of cytokinin and UV-B led to 20 % increase in these cellular compounds (Figure 4C,D).
Higher levels of phenolic compounds were measured in both genotypes following UV-B illumination, yet more pronounced rise was found in einkorn, 37.7 %, than in wheat, 18 %, compared to respective controls (Figure 4E,F). 4PU-30 application and subsequent UV-B illumination reverted the amounts of these substances to control values in wheat and left them slightly higher than controls in einkorn.
2.5. Free radical scavenging and antioxidant activity
In both genotypes free radical scavenging activity estimated from DPPH• neutralization was reduced in response to excessive UV-B radiation, but the reaction was more pronounced in wheat (Figure 5A,B). Leaf spraying with 4PU-30 prior to UV-B treatment had positive effect in both genotypes and resulted in preserved activity in wheat and slightly increased in einkorn compared to respective controls.
Antioxidant activity expressed as FRAP, was reduced in wheat, and enhanced in einkorn after UV-B radiation. Cytokinin spraying also reduced this parameter in wheat and had no significant impact in einkorn. The combination of treatments lessened antioxidant activity in wheat and improved it in einkorn as compared to respective controls (Figure 5C,D).
2.6. Activity of ROS-scavenging enzymes (SOD, CAT, POX, GST)
UV-B treatment caused a two-fold increase in the activity of SOD in wheat and 4.5-fold increase in einkorn. Cytokinin pretreatment significantly lowered the effect of UV stress in einkorn and had a visible influence on SOD activity in wheat as well (Figure 6A,B).
CAT activity in the leaves of wheat plants was only slightly reduced by UV-B treatment, cytokinin spraying and their mutual application. In einkorn CAT activity decreased to 57 % of controls due to UV-B illumination but cytokinin pre-treatment displayed a positive effect by almost reverting this enzyme activity to control levels (Figure 6C,D).
POX activity was greatly increased by UV-B treatment and was 12-times higher than controls in wheat and 35-times higher than einkorn controls. 4PU-30 had a certain recovering effect in both wheat and einkorn but could not bring POX activities to their respective control values in neither of the genotypes (Figure 6E,F).
GST exhibited elevated activity after UV-B treatment with nearly 3-fold rise above control levels in wheat and 3.5-fold increase in einkorn. 4PU-30 exerted an obvious recovery effect on GST enzyme activity when applied before the UV-B treatment and reverted GST to its control activities in both genotypes (Figure 6G,H).
2.7. Gene expression analysis
The results showing the effect of UV-B treatment and 4PU-30 application on the expression levels of antioxidant enzyme encoding genes are presented in Figure 7. The transcript abundance was assessed 72 h after UV-B irradiation of wheat and einkorn seedlings, pretreated or not with the cytokinin regulator 4PU-30 at a concentration of 1 μM. The relative expression levels of three genes belonging to the three groups of SODs (MnSOD, FeSOD, Cu/ZnSOD), one CAT gene (CAT1), two POX genes (POX1, POX7) and three GST genes (GSTL and two different GSTU6 genes designated as GSTU61 (localized in wheat chromosome 6A) and GSTU62 (residing in 7A and 7D, see Table 2) were analyzed. MnSOD gene was significantly upregulated in einkorn plants subjected to UV-B irradiation (Figure 7B) as well as in the 4PU-30-pretreated wheat and einkorn plants exposed to UV-B (Figure 7. A, B). On the contrary, UV-B irradiation downregulated the expression of FeSOD and Cu/ZnSOD in einkorn but had no effect on wheat seedlings (Figure 7 B). UV-B radiation and 4PU-30 application decreased the expression of CAT1 gene in both species, although this effect was more evident in einkorn (Figure 7A,B). The relative transcriptional activity of POX1 gene was increased in both wheat and einkorn in all studied groups compared to controls (Figure 7A,B). The increase in POX1 mRNA levels was more pronounced in einkorn than in wheat samples, especially after UV-B and combined treatment (Figure 7B). Similar pattern of overexpression was observed for the POX7 gene after UV-B irradiation in einkorn, but the plant regulator alone caused slight downregulation (Figure 7B). Both in einkorn and wheat POX7 expression increased after successive 4PU-30 and UV-B exposure (Figure 7A,B). The GST genes showed differential expression depending on the sample and gene analyzed. In einkorn, GSTU61 was upregulated in all studied groups compared to control, with most dramatic increase after cytokinin application (Figure 7. B). On the contrary, in wheat plants the expression of GSTU61 was downregulated in all samples (Figure 7A). The expression pattern of GSTU62 was similar in the two plants studied. Slight increase was observed after UV-B irradiation in einkorn (Figure 7B) and both in einkorn and wheat a marked increase of GSTU62 transcripts was detected after combined cytokinin and UV-B treatment (Figure 7A,B). The expression of GSTL gene differed between the two species with no substantial changes in the einkorn samples (Figure 7B) whereas in wheat plants GSTL was significantly downregulated in all variants compared to the control (Figure 7A).
3. Discussion
Unfavorable environmental conditions significantly reduce crop productivity thus challenging food security in a global scale. UV-B radiation is a natural abiotic factor with key role in plant life cycle but in higher dosages it has the potential for damaging and negatively affecting plant performance [46,47]. CKs are plant hormones which moderate growth and development under normal conditions but they are also acknowledged as active participants in plant responses to stress like heat, drought, cold, and excessive salinity [20,28,48,49]. It was revealed that CKs increased leaf chlorophyll content, chloroplast stability and net photosynthetic rate thus alleviating stress-induced damages [49,50]. Application of growth regulators for improving abiotic stress response is a promising strategy in contemporary agriculture and efforts are aimed at characterizing the molecular mechanisms underlying their mode of action [23,28,51].
In the present study we examined the capacity of exogenously applied synthetic cytokinin 4PU-30 for minimizing the consequences of oxidative stress in wheat and einkorn seedlings induced by excessive UV-B radiation. Our results showed that cytokinin priming stimulated root growth in einkorn and ameliorated leaf pigment content in UV-B treated plants of both genotypes diminishing the inhibitory effect of radiation on these parameters. Although cytokinin pre-treatment could not fully recuperate the reduction in leaf length caused by UV-B, it still had some supplementing effect on growth. These findings are consistent with previously described effect of CKs promoting root elongation [52,53] and recent studies revealing developmental and morphological adjustments in response to UV-B stress [54]. Exogenously applied growth regulator melatonin was reported to significantly alleviate the inhibitory effect of UV-B on root growth [55].
From the remote sensing measurements, the averaged reflectance spectra obtained in our study demonstrated a characteristic peak around 550 nm in the green range of EMS, which was higher for einkorn plants treated with UV-B and its combination with 4PU-30 and corresponded to the higher chlorophyll content in einkorn compared to the same wheat variants. Moreover, einkorn was affected to a higher degree by UV-B compared to wheat, but when treated with 4PU-30 einkorn plants recovered better than wheat plants. UV-B caused significant decrease in chlorophyll and carotenoid content while 4PU-30 treatment resulted in higher pigments’ values in both studied plants. Our results agree with previous findings showing that the reflectance measured in the visible region is generally affected by the concentration of leaf photosynthetic pigments (chlorophylls, carotenoids, and anthocyanins), while in the NIR region it is mostly due to light scattering inside the leaf and therefore related to anatomical traits like mesophyll thickness and stomata structure [56,57,58]. Therefore, the differences in the reflectance spectra between wheat and einkorn observed in the NIR region probably represent a genotypic response. Control wheat plants exhibited higher values in the NIR region of EMS compared to einkorn, while in the visible part of EMS their spectral characteristics were similar. This effect could be attributed to the denser structure of the wheat leaves seen under visual phenotypic observation.
The accumulation of certain cellular substances known as stress markers could give information about stress severity. Judging by the elevated levels of proline and MDA in the leaves of UV-B treated plants in our study it could be concluded that einkorn experienced higher degree of stress in response to the applied radiation. It should be noted, however, that 4PU-30 pretreatment had alleviating effect on MDA and proline accumulation. Oxidative stress is considered a harmful aspect of UV-B impact primarily by triggering the formation of highly reactive species. Imbalances between ROS production and antioxidant scavenging capacity could provoke non-specific damage to DNA, proteins and lipids [59]. The presence of elevated amounts of hydrogen peroxide is regarded as a symptom of impaired redox potential and increased oxidative strain [12]. On the other hand, ROS and membrane degradation products might as well play a role in mediating UV-B protection by modulating gene expression, proteolysis, and cellular redox dynamics [12,60,61]. Exogenously applied cytokinin 4PU-30 decreased hydrogen peroxide accumulation after UV-B treatment in wheat but failed to yield the same result in einkorn. Since einkorn maintained elevated hydrogen peroxide levels after 4PU-30 application and subsequent UV-B treatment, a limited effectiveness of 4PU-30 on reducing ROS formation could be assumed in this genotype.
Usually, abiotic stress increases the production of ROS, while CKs stimulate the antioxidant system to remove these toxic substances [17,60,61,62]. It is well documented that levels of endogenous CKs decline under stress and exogenous application of CKs could improve plant performance through amelioration of assimilate partitioning, retardation of senescence and moderating multiple metabolic processes [16,19,20]. Non-specific low molecular weight compounds like SH, phenols and flavonoids might improve oxidative stress defense primarily by acting as free radical scavengers [55,63,64,65]. Polyphenols and free soluble phenolic acids increased in einkorn in response to radiation and were reported to inhibit lipid peroxidation [2]. In addition, UV-B exposure caused fluctuation in the dynamic pool of redox interactions engaged in oxidative stress protection, which is maintained by multiple non-enzymatic antioxidants, including phenolics, carotenoids, cytochromes, ascorbate, glutathione, xanthophylls, α- tocopherol, polyamines, and proteins that carry redox active SH-groups [11,13,66]. In the present study we detected increase in total phenols in the leaves of wheat and einkorn plants subjected to excessive UV-B radiation and accumulation of thiols only in einkorn. Although flavonoid accumulation could serve as shield against UV damage, in our experiments no significant increase in the levels of these substances was observed. Our results suggested that 4PU-30 increased free radicals scavenging ability (neutralization of DPPH•) under UV-B stress in both genotypes which was correlated with the accumulation of SH compounds in wheat and flavonoids in einkorn. Thus, exogenous 4PU-30 contributed to the release of strain experienced by the plants. Similarly, kinetin pretreatment was found to improve cold stress performance of coffee plants through increase in non-enzymatic antioxidants and enhanced radical scavenging capacity [48]. The plant antioxidant system comprises enzymes that either remove free radicals or prevent their formation. The first line of defense against ROS accumulation is superoxide dismutase (SOD), which converts the O2− radicals to H2O2 [12]. We detected increased SOD activity in response to UV-B treatment in both studied genotypes, but this intensification was higher in einkorn compared to respective control. Two other groups of enzymes, catalase (CAT) and peroxidase (POX) scavenge H2O2 but have different affinities to this toxic substrate. CAT has low affinity whereas POX has higher affinity to H2O2 and might act in the fine regulation of ROS levels [60,61]. Guaiacol peroxidase can act as an effective extinguisher for reactive oxygen intermediates and peroxide radicals under extreme conditions and different environmental stresses induce its activity in plants [13]. From our results it was evidenced that a major role of hydrogen peroxide degradation was played by POX since its activity was substantially increased due to UV-B radiation exposure. However, most probably different isoforms of these enzymes could contribute to the detoxification effect [35]. Glutathione S-transferase (GST) is also regarded as “stress enzyme” due to its involvement in plant responses to drought, salt, and heavy metals [33,67,68]. It metabolizes toxic products of lipid peroxidation and enables the breakdown of xenobiotics (foreign compounds that are not naturally produced within the organism) in a coupling reaction with glutathione. We observed increased GST activity in both wheat and einkorn after exposure to UV-B and the higher SH-containing substances and MDA levels in einkorn corresponded to the more significant increase in GST activity in this genotype.
Variations in the expression patterns of different antioxidant genes, as well as a differential transcriptional regulation in the two Triticum species were observed. The most highly responsive genes found in our study were members of POX and GST families. Interestingly, POX1 and POX7 have been implicated in the biotic and abiotic stress response of einkorn plants although their individual transcription pattern was stress-specific; furthermore, only POX1 was up-regulated upon application of different signaling molecules playing important roles in regulating plant development and defense response [69]. In barley, POX1 transcript levels showed a steady increase after UV-B treatment, while GST regulation differed depending on the growth stage of the seedlings [70]. In this work, POX1 and POX7 showed expression changes that differed significantly in UV-B and 4PU-30 treated einkorn plants in comparison to the untreated controls. In wheat, POX1 and POX7 were also upregulated in seedlings exposed to combined UV-B and 4PU-30 treatment. These results were also in a very good agreement with the increased POX activity determined at the protein level in UV-B irradiated plants. A direct parallel between the two assays is not possible since we cannot exclude involvement of other POX isoforms, however such data imply a common response of the seedlings and suggests that POX1 and POX7 genes may be required for the UV-B tolerance of the two species studied.
Regarding the relative expression of the GST members assessed here, wheat and einkorn plants showed transcription patterns that were species- and gene-dependent. The GSTU61 was significantly downregulated in all sample variants in respect to the control wheat seedlings and almost identical was the transcriptional profile of the GSTL gene. On the other hand, the GSTU62 was not affected by the UV-B treatment alone, but increased its activity upon irradiation when the wheat seedlings were pretreated with the plant regulator 4PU-30. Einkorn plants responded in a similar way in respect to the abundance of their GSTU62 related transcript. On the other hand, a completely different gene regulation was observed for GSTU61 and GSTL in the einkorn seedlings. The drastic increase in GSTU61 mRNA levels in both UV and 4PU-30-treated samples implies that its respective gene product may play important role in the recovery of einkorn plants from UV-B induced oxidative damage. GST gene expression profiles did not correspond directly to the GST activity assessed here at the protein level, especially in wheat. However, it has to be pointed out that the GST family is very large and diverse, with hundreds of GST genes identified in the genomes of Triticum species that vary in their gene functions and transcriptome characteristics depending on the growth, development, hormone regulation as well as adaptation of plants to stress [67,68,71]. Thus, our data suggest an active contribution of certain GSTU6 and GSTL genes in the activation of UV-B induced stress response and oxidative tolerance of plants characterized here. Such a conclusion is in consonance with the established ability of GSTU7 overexpression to restore oxidative stress response in UV-B irradiated Arabidopsis plants [72].
In einkorn, the TmCAT1 gene was implicated in plant’s stress tolerance, its transcription was higher in leaves where an upregulation was observed three days after hormone application, however in response to abiotic stresses, including H2O2 treatment, TmCAT1 expression increased in roots, but not in leaves [73]. Up to now, ten CAT genes have been identified and molecularly characterized in wheat, and a complex transcription pattern involving up- and down-regulation of different CAT genes under various conditions and stress treatment was also observed [74,75]. In our experiments, CAT1 gene showed quite similar expression profiles in both wheat and einkorn plants where the transcript abundance decreased after UV-B treatment. Interestingly, a reduction in the CAT enzyme activity in the UV-B samples was also observed, particularly evident in einkorn, which is in full agreement with the UV-B response reported for field-grown wheat plants [76]. On the other hand, 4PU-30 application was associated with an increase in the enzyme activity in einkorn even in the UV-B irradiated plants. Studies have shown that plant priming can boost CAT gene expression, otherwise inhibited by UV-B, and thereby stimulate the radiation response of soybean seedlings [77]. Taken together, our results indicate that because CAT1 gene expression is negatively affected by both 4PU-30 and UV-B light, the other CAT genes may contribute to the UV-B tolerance of einkorn and wheat seedlings under our experimental conditions. The relative stability of CAT enzyme activity in wheat samples may also be related to the putatively higher numbers of CAT genes existing in the hexaploid wheat in comparison to einkorn thus ensuring an increased stress resistance of this species.
Three major groups of SODs are present in plants, classified based on their metal cofactor (MnSODs, FeSODs and Cu/ZnSODs) and cellular localization [36,78]. Recent studies have discovered more than twenty SOD genes in the hexaploid wheat genome, categorized as three MnSODs, six FeSODs and the rest forming the largest group of Cu/ZnSODs, with constitutive and/or stress modulated transcription patterns [38,79]. Therefore, in our work we selected for expression analysis three SOD genes, each representative of the different SOD classes. SOD genes showed diverse transcription profiles in wheat and einkorn dependent also on the gene analyzed. In wheat, all three SOD genes were transcriptionally stable, even after UV-B radiation; the only exception was MnSOD, whose transcript levels increased in 4PU-30-pretreated control and UV-B irradiated samples. In einkorn seedlings, on the other hand, they were specifically modulated i.e. MnSOD was upregulated and FeSOD and Cu/ZnSOD were down-regulated in seedlings exposed to UV-B. While 4PU-30 application was associated with relatively stable transcript levels of MnSOD and FeSOD, the Cu/ZnSOD showed reduction in these samples. At the protein level, SOD activity increased in both species, irradiated with UV-B, with a more pronounced effect in einkorn. Thus, under our experimental conditions, SOD enzyme activity improved UV-B tolerance of both species, and a particular role could be attributed to the mitochondria localized MnSOD. Notably, MnSOD gene has been implicated in the oxidative stress response of T. monococcum seedlings [39] and induction of specific MnSOD isoforms confers protection against UV-B in soybean [77].
In conclusion, 4PU-30 pretreatment exerted recuperating effect upon antioxidant enzymes triggered by UV-B stress as evidenced by the recovery of their activities to control levels. Although, most probably cytokinins alone could not provide adequate protection against UV damage, better performance of 4PU-30-pretreated wheat and einkorn plants under excessive irradiation was evidenced. Our results suggested that einkorn experienced a higher degree of stress in response to the applied UV-B radiation. These findings emphasize the differential responses of wheat and einkorn plants to excessive UV-B treatment, implying that the reaction towards this environmental stimulus could have a genotype-specific nature.
Stress damage could be crucial especially during seed germination and at early seedling stage. It was shown that 4PU-30 seed priming improved metabolic activity after cold storage thus contributing to enhanced seed viability [80]. Exogenous CK improved growth performance by attenuating the damaging effects of UV-B radiation and stabilizing the photosynthetic apparatus [15]. The results presented here emphasize the positive effect of cytokinin pretreatment and offer convincing evidence that priming with 4PU-30 could successfully be applied for improving stress tolerance in cereals. Furthermore, fundamental research on plant coping mechanisms and the pursuit of possibilities for alleviating the negative effects of stress could have substantial influence on future agricultural practices in view of the growing food demands of human population.
4. Materials and Methods
4.1. Plant material, growth conditions and experimental design
Wheat (Triticum aestivum, L.) from Bulgarian variety Enola and einkorn (Triticum monococcum, L.) seeds were grown for 10 days on half strength Hoagland-Arnon nutrient solution under controlled conditions with 25/21 °C day/night temperature, 16 h photoperiod, 250 µmol m-2 s-1 photon flux density at leaf level. Afterwards, sets of one control and three test variants were established. Controls remained untreated while the following test groups were formed as follows: 4PU-30-treated plants, UV-B-treated plants and plants subjected to a combination of 4PU-30 and subsequent 3 h UV-B illumination. Leaves of 10-days-old seedlings were sprayed with water solution of 1×10-6 (1 µM) cytokinin 4PU-30 containing 0.1 % Tween 80. Illumination with UV-B was imposed during the light period and 24 h after 4PU-treatment. Seedlings were irradiated with UV-B lamps (Philips TL 2X20W/12 RSSLV/25, λmax 312 nm) for 180 min. The distance between UV-B lamp and the top leaves of the treated plants was 0.12 m ± 0.04 m. Physiological and biometric parameters were measured 72 h after UV-B exposure. First fully developed leaves were sampled for analyses and were either used immediately or frozen in liquid nitrogen and stored at – 70 °C until measurement.
4.2. Plant biometry and estimation of leaf pigment content
Root length and length of the first fully developed leaf were measured from 10 plants of each variant. Results were given as means ± SD from two independent experiments (n = 20). For estimation of pigment content in the leaves fresh samples were homogenized in ice-cold 80 % (v/v) acetone and were then centrifuged at 5000×g for 5 min at 4 °C.
Chlorophyll (Chl a and Chl b) and total carotenoids (Car) absorption were measured spectrophotometrically at 663, 645 and 460 nm and pigment content was calculated by the equations of [81].
4.3. Remote sensing spectrometric data
The spectral reflectance characteristics of wheat and einkorn plants were measured using a remote sensing set-up based on the following devices - Ocean Optics USB4000 spectrometer (Figure 8A) in the electromagnetic range 400 nm to 900 nm, Ocean Optics durable fiber optic (Figure 8B) and certified reflectance standard Labsphere (Figure 8C). Remote sensing spectrometric measurements were performed with high spectral resolution (1-10 nm) allowing the registration of reflectance spectra in the highest possible detail due to better quality of the sensor USB4000 sensor. All data were acquired by means of specialized software controlling the spectrometric device [82]. The illumination source used in the experiments was diffused sunlight and the integration time was adjusted in each experiment for the best dynamic range of the reflectance measurements. The durable fiber was always pointed at nadir to the measured sample.
4.4. Biochemical analyses
Fresh leaf material collected from first fully developed leaves was used for analysis or stored in liquid nitrogen until usage. Extraction was performed at 4 °C by homogenization of 300 mg samples in 0.1 % (w/v) trichloroacetic acid followed by centrifugation at 15 000×g and collecting the supernatant for quantification of oxidative stress markers and non-enzymatic antioxidants.
4.4.1. Oxidative stress markers analysis (free proline, hydrogen peroxide, malondialdehyde)
Free proline was derivatized using acid ninhydrin reagent for 60 min at 100 °C, absorbance was read at 520 nm and the amounts were calculated from a previously plotted standard curve. Hydrogen peroxide (H2O2) content was evaluated spectrophotometrically based on its reaction with 1 M KI for 60 min in the dark as described in [83]. Absorbance was read at 390 nm and the amounts of H2O2 were calculated from a standard curve with known concentrations. Malondialdehyde (MDA) content was determined as thiobarbituric acid reagent product according to [84]. Absorbances were read at 532 and 600 nm and quantification performed using a 155 mM-1 cm-1 extinction coefficient.
4.4.2. Non-enzymatic antioxidants (total phenolic compounds, flavonoids and free thiol groups)
Total phenolic compounds contained in leaf extracts were measured using Folin-Ciocalteu reagent supplemented with sodium carbonate, absorbance was read at 725 nm and results were given as gallic acid equivalents (mg GA fresh weight g-1) according to [85].
Total flavonoid content was analyzed by the aluminum chloride colorimetric assay [64]. Absorbance was read at 510 nm and results expressed in rutin equivalents. Free thiol (SH) group containing compounds were measured after incubation of leaf extracts for 10 min at 20 °C in Ellman’s reagent according to [86]. Absorbance was measured at 412 nm.
4.4.3. Free radical scavenging activity and antioxidant capacity
Free radical scavenging activity was analyzed in leaf extracts based on the antioxidants ability to bleach purple methanol solution containing DPPH• (1,1-diphenyl-2-picrylhydrazyl) that can readily undergo reduction by the antioxidants in the sample [87].
where Acontrol is the absorbance of the control sample and Asample is the absorbance of the test sample measured at 517 nm.
Radical scavenging activity (DPPH inhibition %) = [(Acontrol − Asample)/Acontrol] × 100 %,
Ferric reducing antioxidant power (FRAP) assay was carried out according to [88]. The method is based on the rapid reduction of ferrous-tripyridyltriazine (FeIII-TPTZ) to ferric-tripyridyltriazine (FeII-TPTZ) blue product at low pH by antioxidants present in the samples. Absorbance was measured at 593 nm and results were given in μmol Fe2+ TPTZ g-1 FW expressing ferric reducing activity equivalent to 1 mM FeSO4.
Multiskan Spectrum spectrophotometer equipped with a microplate reader (Thermo Electron Corporation, Vantaa, Finland) was used in these measurements.
4.4.4. Preparation of crude extracts and assays for antioxidant (ROS-scavenging) enzymes (CAT, SOD, POX, GST) activities.
Antioxidant enzymes were extracted from leaf tissue. Fresh leaf material (500 mg) was homogenized at 4 °C in 5 ml of 50 mM potassium phosphate buffer (pH 7.8) supplemented with 10 mM ethylenediaminetetra-acetic acid (EDTA), 1 mM dithiotreitol, 0.1 mM phenylmethylsulfonyl fluoride (PMSF) and 1 % polyvinylpyrrolidone. After centrifugation at 12 000×g for 30 min supernatant aliquots were used for assaying of specific enzyme activities [89]. Total soluble protein content was determined according to [90] using bovine serum albumin as a standard.
Catalase (CAT, EC 1.11.1.6) activity was assayed as in [91] based on H2O2 decomposition measured spectrophotometrically by monitoring absorbance decrease at 240 nm and using molar extinction coefficient (39.4 mM-1 cm-1) in calculations. Results were presented as the amount of H2O2 decomposed per mg protein per min.
Total superoxide dismutase (SOD, EC 1.15.1.1) activity was estimated according to [92]. The method was based on the inhibition of photochemical reduction of nitroblue tetrazolium (NBT). The reaction product was measured spectrophotometrically following the absorbance increase at 560 nm. SOD activity was defined as mg protein causing 50% inhibition of NBT photoreduction rate.
Peroxidase (POX, EC 1.11.1.7) activity was assayed according to [93] using guaiacol as electron donor. Formation of tetraguaiacol (TG) was monitored through the increase in absorbance at 470 nm and calculated using molar extinction coefficient (26.6 mM-1 cm-1). Results were given in µmol TG mg−1 protein min−1.
Glutathione S-transferase (GST, E.C. 2.5.1.18) activity was determined by measuring the rate of conjugation of glutathione (GSH) with 1-chloro-2,4-dinitrobenzene (CDNB) according to [94]. The absorbance at 340 nm was monitored, a 9.6 nmol-1 cm-1 extinction coefficient was used for calculations and results were expressed as µmol reduced CDNB mg−1 protein min−1.
Enzyme activities were measured on a Shimadzu UV-1601 spectrophotometer (Kyoto, Japan).
4.5. Gene expression analysis
The experimental variants of einkorn and wheat seedlings subjected to gene expression analyses were as follows: Controls (without any treatment and pretreated with 1 µM 4PU-30) and UV-irradiated (UV-B only and 1 µM 4PU-30plus UV-B). The first leaf of 3-5 seedlings (equal to ~130 mg of tissue) was excised, wrapped in aluminum foil, and immediately frozen in liquid nitrogen, then stored at - 70 °C for further processing. Frozen material was initially ground in liquid nitrogen and pure RNA was isolated with the RNeasy plant mini kit (Qiagen, Germany) according to the manufacturer’s instructions with on-column DNase digestion. The RNA elution step was performed 3 times for each sample. RNA concentrations and purity were determined spectrophotometrically (BioSpec-nano, Shimadzu). Reverse transcription (RT) reactions for cDNA synthesis were performed with the FIREScript® RT cDNA synthesis KIT (Solis BioDyne, Tartu, Estonia) according to the recommended protocol utilizing random hexamer primers and 1μg of RNA template. Each RT reaction was diluted 1:5 with ultrapure water and 2 μl of the diluted cDNA was used as a template for qPCR analysis. The final PCR reaction mixture (10 μl) was prepared with the HOT FIREPol® EvaGreen® qPCR Supermix according to the manufacturer’s protocol (Solis BioDyne, Tartu, Estonia). The RT-qPCR was carried out in a PikoReal Real-Time PCR System (Thermo Scientific). The amplification conditions included initial denaturation for 15 min at 95 °C, 40 cycles of denaturation for 15 s at 95 °C, annealing for 30 s at 60 °C and extension for 30 s at 72 °C, followed by final elongation for 5 min at 72 °C and melting curve analysis (60-95 °C, temp. increment 0.2 °C). All data were analyzed using the PikoReal Software version 2.2 (Thermo Scientific). The fold change in the expression levels of the genes was estimated based on the Pfaffl’s method [95]. 18S ribosomal RNA fragment (118 bp) was used as a reference control amplified with the following primers: HV_18Ssh1F-CCTGCGGCTTAATTTGACTCA and HV_18SR- AACTAAGAACGGCCATGCAC [96]. The genes whose expression was characterized were SOD, CAT, POX, and GST. All of them are part of gene families, therefore a selection was made based on literature data for involvement in stress response and/or the availability of DNA sequences for both species in the NCBI database. Primers utilized in the current study were either newly designed or taken from literature. Detailed description of the primers, their target genes and the respective references are provided in Table 2. Gene-specific primers were designed utilizing the Primer Blast Tool of NCBI database (Primer designing tool (nih.gov))[97]. Primer pairs were designed to span exon-exon boundaries (where possible) to avoid amplification of any residual gDNA contamination during qPCR. The newly constructed primers were aimed to target transcripts of einkorn genome and all three wheat genomes (AABBDD) where possible. Primers were tested for specificity against wheat mRNA Seq NCBI Database and only those showing 100 % specificity and no other potential targets in wheat genome were selected. In addition, all primer pairs were tested for possible primer-dimers by the online tool “Multiple Primer Analyzer” set to maximum or optimal sensitivity (Multiple Primer Analyzer/Thermo Fisher Scientific)[98].

Table 2.
Primer pairs and their targets in wheat and einkorn.
| Primer name | Primer sequence | Reference | Amplicon length | Target gene Tm/Ta | Acc. No & Chromosome localization (in wheat) |
|---|---|---|---|---|---|
| MnSOD-F MnSOD-R |
TCCGCCGTCGTCCACCTC CCACCACCCTCGCTGATG CCACCACCCTCGCTAATA (this study) |
Karimi et al., 2017 [99] | 104 bp | TmMnSOD TaMnSOD3.1 |
MK091461.1 - Tm XM_044603645.1 – Ta 2A |
| TaFe-SOD-F TaFe-SOD-R |
CCTACTGGATGAGACGGAGAG GGACGAGGACAACGACGAA |
Luo et al., 2019 [100] | 124 bp | TaFeSOD | AK453889.1 – Ta 7D JX398977.1 – Ta 7D XM_044510132.1 – Ta 4A |
| TaCu/Zn-SOD-F TaCu/Zn-SOD-R |
TGGGAGAGCGTTTGTTGTTC GTCTTCCACCAGCATTTCCA |
Luo et al., 2019 [100] | 92 bp | TaCu/Zn-SOD, SOD1.2 |
XM_044573537.1 - Ta 7A, X1 XM_044573538.1 - Ta 7A, X2 XM_044573539.1 - Ta 7A, X3 XM_044578041.1 - Ta 7B, X1 XM_044578042.1 - Ta 7B, X2 XM_044587054.1 - Ta 7D |
| qTmCAT1-F qTmCAT1-R |
CGAGAAGATGGTGATCGAGAA TGTTGATGAATCGCTCTTGC |
Tounsi et al., 2019b [73] | 95 bp | TmCAT1 TaCAT1 |
MK091459.1 - Tm NM_001405704.1 – Ta 4B XM_044520955.1 – Ta 4D XM_044527958.1 – Ta 5A |
| TGST2-F TGST2-R |
TACGAGGACGTGGAGGAGAA TGTGGATGAGCACGGGTATC |
This study | 91 bp | TmGSTU TaGSTU61 |
Tm EF044232.1 XM_044552184.1 - 6A XM_044550330.1 – 6A |
| TaGSTU56-F TaGSTU56-R |
TTAAAGATCTCGTCGTTCCAC AACAGCTACTCACAAGGCAGA |
Wang et al., 2019 [71] | 92 bp | TaGSTU62 | XM_044570870.1 - Ta 7A XM_044581608.1 - Ta 7D |
| TaGSTL10-F TaGSTL10-R |
ATGTGCCATTTATCGAAAGGT TCCATGCTGCAGTAGTTCCC |
Wang et al., 2019 [71] | 234 bp | TaIN2-1 homolog B | XM_044507158.1 - Ta 4A XM_044514325.1 - Ta 4B XM_044519678.1 - Ta 4D, X1 XM_044519679.1 - Ta 4D, X2 |
| POX1u-F POX1u-R |
CTCCAGGGTGAACTCGTGAT GCCTTTGCATGAGAAAGTGGG |
This study | 219 bp | TmPOX1 TaPOX1 |
AY857755.1 - Tm XM_044599368.1 - Ta 2A XM_044466236.1 - Ta 2B XM_044474478.1 - Ta 2D |
| POX7u1-F POX7u1-R |
GTCGTGGACGAGGTCAAGAG TGGGTCCACCAGTCAGCA |
This study | 112 bp | TmPOX7 TaPOX17-like |
AY857761.1 – Tm XM_044525767.1 - Ta 5A XM_044533224.1 - Ta 5B XM_044541856.1 - Ta 5D |
4.6. Statistical analysis
Two independent experiments were carried out and parameters were measured in three technical replicates. Data are given as means ± SD. Significance of differences between treatments was analyzed by one-way ANOVA with post-hoc Duncan’s multiple range test at 0.05 level. Statistical analysis of gene expression data was completed using Student’s t-test at 0.05 probability level, based on the ΔCt values.
5. Conclusions
At an early developmental stage wheat and einkorn seedling responded to excessive UV-B radiation in a rather similar way by activation of specific defense mechanisms. However, einkorn experienced a higher degree of stress revealed by intensified enzyme activity, increased accumulation of non-enzyme components of the antioxidant system as well as activated upregulation of gene expression. Although our preliminary hypothesis that einkorn would perform better under increased irradiation was not confirmed, it was demonstrated that synthetic cytokinin 4PU-30 pretreatment had the potential to alleviate negative effects of oxidative stress by attenuating the symptoms of superfluous UV-B illumination in both studied genotypes. The use of remotely sensed spectral data represented a promising non-destructive technology for detection of UV-B stress impact and plant recovery caused by 4PU-30, these findings being supported by several independent types of data registration.
Author Contributions
I.M. – Conceptualization, coordination, funding acquisition, project administration; I.M., Z.T. and Y.K. – plant cultivation and treatment procedures, biometric measurements; I.M., E.K., I.S., Y.K. – biochemical analyses, data collection and discussion; E.K., I.S., K.K. – data interpretation, figure preparation; Y.K. and V.M. – gene expression experiment, data analysis, interpretation and presentation; H.N., D.B., V.D. performed spectrometric measurements and analysis of spectral data; D.B., H.N. – figure preparation; E.K., V.M., K.K. – prepared the original draft of the manuscript; I.S. – reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC was funded by Bulgarian National Science Fund (BNSF) grant KP-06 N56/15.
Data Availability Statement
Data in this research are contained within the manuscript tables and figures.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Feldman, M.; Levy, A.A. Wheat Evolution and Domestication; Springer: Switzerland, 2023; p. 673. [Google Scholar] [CrossRef]
- Benincasa, P.; Tosti, G.; Farneselli, M.; Maranghi, S.; Bravi, E.; Marconi, O.; Falcinelli, B.; Guidicci, M. Phenolic content and antioxidant activity of einkorn and emmer sprouts and wheatgrass obtained under different radiation wavelengths. Ann. Agric. Sci. 2020, 65, 68–76. [Google Scholar] [CrossRef]
- Keçeli, A. A review on the bioactive, antioxidant properties of einkorn (Triticum monococcum L. ssp. monococcum) populations and using in organic agriculture. Turk J. Agric. – Food Sci. Technol. 2019, 7, 2111–2120. [Google Scholar] [CrossRef]
- Desheva, G.; Valchinova, E.; Chipilski, R.; Uzundzhieva, K.; Kyosev, B. Morphophyziological and anatomical characteristics of leaves in accessions of wild einkorn (Triticum boeoticum Boiss.). Int. J. Env. Agric. Biotechnol. 2018, 3, 1391–1400. [Google Scholar] [CrossRef]
- Guzman, C.; Alvarez, J.B. Ancient wheats role in sustainable wheat cultivation. In Trends in Wheat and Bread Making. Galanakis, C.M., Ed.; Charlotte Cockle: Academic Press, 2021; pp. 29–66. [Google Scholar] [CrossRef]
- Chen, Z.; Dong, Y.; Huang, X. Plant responses to UV-B radiation: Signaling, acclimation and stress tolerance. Stress Biol. 2022, 2, 51. [Google Scholar] [CrossRef]
- Manova, V.; Gruszka, D. DNA damage and repair in plants–from models to crops. Front. Plant Sci. 2015, 6, 885. [Google Scholar] [CrossRef]
- Hideg, E.; Jansen, M.A.K.; Strid, Å. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Jankû, M.; Luhová, L.; Petřivalský, M. On the origin and fate of reactive oxygen species in plant cell compartments. Antioxidants 2019, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Song, S.; Abdulrashid, K.; Chai, Y.; Yue, M.; Liu, X. Separate and combined response to UV-B radiation and jasmonic acid on photosynthesis and growth characteristics of Scutellaria baicalensis. Int. J. Mol. Sci. 2018, 19, 1194. [Google Scholar] [CrossRef]
- Shi, C.; Liu, H. How plants protect themselves from ultraviolet-B radiation stress. Plant Physiol. 2021, 187, 1096–1103. [Google Scholar] [CrossRef]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, S.; Zulfiqar, F.; Alam, M.; Fujita, M. Regulation of ROS metabolism in plants under environmental stress: A review of recent experimental evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Kosakivska, I.V.; Vedenicheva, N.P.; Babenko, L.M.; Voyenko, L.V.; Romanenko, K.O.; Vasyuk, V.A. Exogenous phytohormones in the regulation of growth and development of cereals under abiotic stresses. Mol. Biol. Rep. 2022, 49, 617–628. [Google Scholar] [CrossRef]
- Veselov, D.S.; Kudoyarova, G.R.; Kudryakova, N.V.; Kusnetsov, V.V. Role of cytokinins in stress resistance of plants. Russ. J. Plant. Physiol. 2017, 64, 15–27. [Google Scholar] [CrossRef]
- Pavlů, J.; Novák, J.; Koukalová, V.; Luklová, M.; Brzobohatý, B.; Černý, M. Cytokinin at the crossroads of abiotic stress signalling pathways. Int. J. Mol. Sci. 2018, 19, 2450. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Cui, X.; Wang, K.; Wang, Y.; He, Y. Phytohormones regulate the abiotic stress: An overview of physiological, biochemical, and molecular responses in horticultural crops. Front. Plant Sci. 2023, 13, 1095363. [Google Scholar] [CrossRef] [PubMed]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell. Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, M.; Meng, Z.; Wang, B.; Chen, M. Research progress on the roles of cytokinin in plant response to stress. Int. J. Mol. Sci. 2020, 21, 6574. [Google Scholar] [CrossRef]
- Todorova, D.; Moskova, I.; Sergiev, I.; Alexieva, V.; Mapelli, S. Changes in endogenous polyamines and some stress markers content induced by drought, 4PU-30 and abscisic acid in wheat plants. In Abiotic Stress and Plant Responses; Khan, N., Singh, S.I.K., Eds.; International Publishing House Pvt. Ltd.: New Delhi, India, 2008; pp. 205–215. [Google Scholar]
- Yonova, P. Design, synthesis and properties of synthetic cytokinins. Recent advances on their application. Gen. App. Plant Physiol. 2010, 36, 124–147. [Google Scholar]
- Ellouzi, H.; Zorrig, W.; Amraoui, S.; Oueslati, S.; Abdelly, C.; Rabhi, M.; Siddique, K.H.M.; Hessini, K. Seed priming with salicylic acid alleviates salt stress toxicity in barley by suppressing ROS accumulation and improving antioxidant defense systems, compared to halo- and gibberellin priming. Antioxidants 2023, 12, 1779. [Google Scholar] [CrossRef]
- Moskova, I.; Kocheva, K. Phenylurea-type cytokinin ameliorates the performance of young pea plants under salt stress. Botanica 2021, 27, 141–148. [Google Scholar] [CrossRef]
- Sergiev, I.G.; Alexieva, V.S.; Ivanov, S.V.; Moskova, I.I.; Karanov, E.N. The phenylurea cytokinin 4PU-30 protects maize plants against glyphosate action. Pestic. Biochem. Physiol. 2006, 85, 139–146. [Google Scholar] [CrossRef]
- Shopova, E.; Katerova, Z.; Brankova, L.; Dimitrova, L.; Sergiev, I.; Todorova, D.; Talaat, N.B. Modulation of physiological stress response of Triticum aestivum L. to glyphosate by brassinosteroid application. Life 2021, 11, 1156. [Google Scholar] [CrossRef]
- Moskova, I.; Dikova, B.; Balacheva, E.; Sergiev, I. Protective effect of plant growth regulators MEIA and 4PU-30 against tomato spotted wilt virus (TSWV) on two tomato genotypes. Compt. Rend. Acad. Bulg. Sci. 2020, 73, 1538–1544. [Google Scholar] [CrossRef]
- Mandal, S.; Ghorai, M.; Anand, U.; Samanta, D.; Kant, N.; Mishra, T.; Rahman, M.H.; Jha, N.K.; Jha, S.K.; Lal, M.K.; Tiwari, R.K.; Kumar, M.; Radha; Prasanth, D. A.; Mane, A.B.; Gopalakrishnan, A.V.; Biswas, P.; Proćków, J.; Dey, A. Cytokinin and abiotic stress tolerance -What has been accomplished and the way forward? Front. Genet. 2022, 13, 943025. [Google Scholar] [CrossRef]
- Hudeček, M.; Nožková, V.; Plíhalová, L.; Plíhal, O. Plant hormone cytokinin at the crossroads of stress priming and control of photosynthesis. Front. Plant Sci. 2023, 13, 1103088. [Google Scholar] [CrossRef] [PubMed]
- Kolupaev, Y.E.; Karpets, Y.V.; Kabashnikova, L.F. Antioxidative system of plants: Cellular compartmentalization, protective and signaling functions, mechanisms of regulation. Appl. Biochem. Microbiol. 2019, 55, 441–459. [Google Scholar] [CrossRef]
- Cantarello, C.; Volpe, V.; Azzolin, C.; Bertea, C. Modulation of enzyme activities and expression of genes related to primary and secondary metabolism in response to UV-B stress in cucumber (Cucumis sativus L.). J. Plant Interact. 2005, 1, 151–161. [Google Scholar] [CrossRef]
- Gaafar, R.M.; Osman, M.E.-A.H.; Abo-Shady, A.M.; Almohisen, I.A.A.; Badawy, G.A.; El-Nagar, M.M.F.; Ismail, G.A. Role of antioxidant enzymes and glutathione S-transferase in bromoxynil herbicide stress tolerance in wheat plants. Plants 2022, 11, 2679. [Google Scholar] [CrossRef]
- Navabpour, S.; Yamchi, A.; Bagherikia, S.; Kafi, H. Lead-induced oxidative stress and role of antioxidant defense in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plant. 2020, 26, 793–802. [Google Scholar] [CrossRef]
- Nasirzadeh, L.; Sorkhilaleloo, B.; Majidi Hervan, E.; Fatehi, F. Changes in antioxidant enzyme activities and gene expression profiles under drought stress in tolerant, intermediate, and susceptible wheat genotypes. Cereal Res. Comm. 2021, 49, 83–89. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase—Mentor of abiotic stress tolerance in crop plants. Environ. Sci. Poll. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef]
- Zameer, R.; Fatima, K.; Azeem, F.; ALgwaiz, H.I.; Sadaqat, M.; Rasheed, A.; Batool, R.; Shah, A.N.; Zaynab, M.; Shah, A.A.; Attia, K.A. Genome-wide characterization of superoxide dismutase (SOD) genes in Daucus carota: Novel insights into structure, expression, and binding interaction with hydrogen peroxide (H2O2) under abiotic stress condition. Front. Plant Sci. 2022, 13, 870241. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J.; Monde, R.A.; Last, R.L. Superoxide dismutase in Arabidopsis: An eclectic enzyme family with disparate regulation and protein localization. Plant Physiol. 1998, 118, 637–650. [Google Scholar] [CrossRef]
- Jiang, W.; Yang, L.; He, Y.; Zhang, H.; Li, W.; Chen, H.; Ma, D.; Yin, J. Genome-wide identification and transcriptional expression analysis of superoxide dismutase (SOD) family in wheat (Triticum aestivum). Peer J 2019, 7, e8062. [Google Scholar] [CrossRef]
- Tounsi, S.; Feki, K.; Kamoun, Y.; Saïdi, M.N.; Jemli, S.; Ghorbel, M.; Alcon, C.; Brini, F. Highlight on the expression and the function of a novel MnSOD from diploid wheat (T. monococcum) in response to abiotic stress and heavy metal toxicity. Plant Physiol. Biochem. 2019, 142, 384–394. [Google Scholar] [CrossRef]
- Boldt, R.; Scandalios, J.G. Influence of UV-light on the expression of the Cat2 and Cat3 catalase genes in maize. Free Radic. Biol. Med. 1997, 23, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, M.; Zribi, I.; Haddaji, N.; Siddiqui, A.J.; Bouali, N.; Brini, F. Genome-wide identification and expression analysis of catalase gene families in Triticeae. Plants 2024, 13, 11. [Google Scholar] [CrossRef]
- Sharma, I.; Ahmad, P. Catalase: A versatile antioxidant in plants. In Oxidative Damage to Plants; Ahmad, P., Ed.; Elsevier, 2014; pp. 131–148. [Google Scholar] [CrossRef]
- Kancheva, R. Main principles in vegetation spectrometric studies. Annual of University of Mining and Geology 2003, 46, 351–353. [Google Scholar]
- Kancheva, R.; Iliev, I.; Borisova, D.; Chankova, S.; Kapchina, V. Detection of plant physiological stress using spectral data. Ecol. Engin. Environ. Prot. 2005, 1, 4–9. [Google Scholar]
- Kancheva, R.; Georgiev, G. Seasonal spectral response patterns of winter wheat canopy for crop performance monitoring. Proc. SPIE 8887, Remote Sens. Agric. Ecosyst. Hydrol. 2013, 15, 8887, 88871V. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Changing scenario in plant UV-B research: UV-B from a generic stressor to a specific regulator. J. Photochem. Photobiol. B: Biology 2015, 153, 334–343. [Google Scholar] [CrossRef]
- Lima, M.P.; Soares, A.M.; Loureiro, S. Responses of wheat (Triticum aestivum) and turnip (Brassica rapa) to the combined exposure of carbaryl and ultraviolet radiation. Environ. Toxicol. Chem. 2015, 34, 1665–1674. [Google Scholar] [CrossRef] [PubMed]
- Acidri, R.; Sawai, Y.; Sugimoto, Y.; Handa, T.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Exogenous kinetin promotes the nonenzymatic antioxidant system and photosynthetic activity of coffee (Coffea arabica L.) plants under cold stress conditions. Plants 2020, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Prerostova, S.; Dobrev, P.I.; Gaudinova, A.; Knirsch, V.; Korber, N.; Pieruschka, R.; Fiorani, F.; Brzobohaty, B.; Cerny, M.; Spichal, L.; Humplik, J.; Vanek, T.; Schurr, U.; Vankova, R. Cytokinins: Their impact on molecular and growth responses to drought stress and recovery in Arabidopsis. Front. Plant Sci. 2018, 9, 655. [Google Scholar] [CrossRef] [PubMed]
- Xiaotao, D.; Yuping, J.; Hong, W.; Haijun, J.; Hongmei, Z.; Chunhong, C.; Jizhu, Y. Effects of cytokinin on photosynthetic gas exchange, chlorophyll fluorescence parameters, antioxidative system and carbohydrate accumulation in cucumber (Cucumis sativus L.) under low light. Acta Physiol. Plant. 2013, 35, 1427–1438. [Google Scholar] [CrossRef]
- Katerova, Z.; Shopova, E.; Georgieva, N.; Nikolova, A.; Sergiev, I.; Todorova, D. MEIA acts as protector against UV-C irradiation in young wheat plants. Compt. Rend. Acad. Bulg. Sci. 2012, 65, 1373–1378. [Google Scholar]
- Ivanov, V.B.; Filin, A.N. Cytokinins regulate root growth through its action on meristematic cell proliferation but not on the transition to differentiation. Funct. Plant Biol. 2017, 45, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R. Cytokinin and its key role to enrich the plant nutrients and growth under adverse conditions - an update. Front. Genet. 2022, 13, 883924. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, D.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. Light signaling and UV-B-mediated plant growth regulation. J. Integr. Plant Biol. 2020, 62, 1270–1292. [Google Scholar] [CrossRef]
- Wei, Z.; Li, C.; Gao, T.; Zhang, Z.; Liang, B.; Lv, Z.; Zou, Y.; Ma, F. Melatonin increases the performance of Malus hupehensis after UV-B exposure. Plant Physiol. Biochem. 2019, 139, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Ustin, S.L.; Jacquemoud, S. How the optical properties of leaves modify the absorption and scattering of energy and enhance leaf functionality. In Remote Sensing of Plant Biodiversity; Cavender-Bares, J., Gamon, J.A., Townsend, P.A., Eds.; Springer: Cham, 2020; pp. 349–384. [Google Scholar]
- Mohd Asaari, M. S.; Mertens, S.; Verbraeken, L.; Dhondt, S.; Inzé, D.; Bikram, K.; Scheunders, P. Non-destructive analysis of plant physiological traits using hyperspectral imaging: A case study on drought stress. Comp. Electron. Agric. 2022, 195, e106806. [Google Scholar] [CrossRef]
- Williams, D.; Karley, A.; McCallum, S.; Graham, J. Raspberry plant stress detection using hyperspectral imaging. Plant Direct. 2023, 7, e490. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J. 2020, 105, 459–476. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Kumar, R.; Altaf, M.M.; Kumar, A.; Khan, L.U.; Saqib, M.; Nawaz, M.A.; Saddiq, B.; Bahadur, S.; Tiwari, R.K.; Lal, M.K; Naz, S. Phytohormones mediated modulation of abiotic stress tolerance and potential crosstalk in horticultural crops. J. Plant Growth Regul. 2023, 42, 4724–4750. [Google Scholar] [CrossRef]
- Zagorchev, L.; Seal, C.E.; Kranner, I.; Odjakova, M.A. Central role for thiols in plant tolerance to abiotic stress. Int. J. Mol. Sci. 2013, 14, 7405–7432. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Fut. J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Soares, C.; Carvalho, M.E.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges – A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Gallé, Á.; Csiszár, J.; Secenji, M.; Guóth, A.; Cseuz, L.; Tari, I.; Györgyey, J.; Erdei, L. Glutathione transferase activity and expression patterns during grain filling in flag leaves of wheat genotypes differing in drought tolerance: Response to water deficit. J. Plant Physiol. 2009, 166, 1878–1891. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Xu, S.; Lyu, Z.; Wang, H.; Kong, L.; Sun, S. Comparative analysis of the glutathione s-transferase gene family of four Triticeae species and transcriptome analysis of GST genes in common wheat responding to salt stress. Int. J. Genom. 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Sheng, X.; Greenshields, D.L.; Ogieglo, A.; Kaminskyj, S.; Selvaraj, G.; Wei, Y. Profiling of wheat class III peroxidase genes derived from powdery mildew-attacked epidermis reveals distinct sequence-associated expression patterns. Mol. Plant - Microbe Interact. 2005, 18, 730–741. [Google Scholar] [CrossRef]
- Wang, M.; Leng, C.; Zhu, Y.; Wang, P.; Gu, Z.; Yang, R. UV-B treatment enhances phenolic acids accumulation and antioxidant capacity of barley seedlings. LWT 2022, 153, 112445. [Google Scholar] [CrossRef]
- Wang, R.; Ma, J.; Zhang, Q.; Wu, C.; Zhao, H.; Wu, Y.; Yang, G.; He, G. Genome-wide identification and expression profiling of glutathione transferase gene family under multiple stresses and hormone treatments in wheat (Triticum aestivum L.). BMC Genom. 2019, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Vásquez, A.; Fonseca, A.; Ugalde, J.M.; Lamig, L.; Seguel, A.; Moyano, T.C.; Gutiérrez, R.A.; Salinas, P.; Vidal, E.A.; Holuigue, L. TGA class II transcription factors are essential to restrict oxidative stress in response to UV-B stress in Arabidopsis. J. Exp. Bot. 2021, 72, 1891–1905. [Google Scholar] [CrossRef] [PubMed]
- Tounsi, S.; Kamoun, Y.; Feki, K.; Jemli, S.; Saïdi, M.N.; Ziadi, H.; Alcon, C.; Brini, F. Localization and expression analysis of a novel catalase from Triticum monococcum TmCAT1 involved in response to different environmental stresses. Plant Physiol. Biochem. 2019, 139, 366–378. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Yun, L.; Ji, L.; Li, G.; Ji, M.; Shi, Y.; Zheng, X. Catalase (CAT) gene family in wheat (Triticum aestivum L.): Evolution, expression pattern and function analysis. Int. J.Mol. Sci. 2022, 23, 542. [Google Scholar] [CrossRef]
- Ermakov, A.; Bobrovskikh, A.; Zubairova, U.; Konstantinov, D.; Doroshkov, A. Stress-induced changes in the expression of antioxidant system genes for rice (Oryza sativa L.) and bread wheat (Triticum aestivum L.). Peer J 2019, 7, e7791. [Google Scholar] [CrossRef]
- Ambasht, N.K; Agrawal, M. Effects of enhanced UV-B radiation and tropospheric ozone on physiological and biochemical characteristics of field grown wheat. Biol. Plant. 2003, 47, 625–628. [Google Scholar] [CrossRef]
- Santa-Cruz, D.M. , Pacienza, N.A., Zilli, C.G., Tomaro, M.L., Balestrasse, K.B.; Yannarelli, G.G. Nitric oxide induces specific isoforms of antioxidant enzymes in soybean leaves subjected to enhanced ultraviolet-B radiation. J. Photochem. Photobiol. B: Biology 2014, 141, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.H.; Skinner, D.Z. Differential expression of manganese superoxide dismutase sequence variants in near isogenic lines of wheat during cold acclimation. Plant Cell Rep. 2006, 25, 223–230. [Google Scholar] [CrossRef]
- Tyagi, S.; Sharma, S.; Taneja, M.; Kumar, R.; Sembi, J.K.; Upadhyay, S.K. Superoxide dismutases in bread wheat (Triticum aestivum L.): Comprehensive characterization and expression analysis during development and, biotic and abiotic stresses. Agri Gene 2017, 6, 1–13. [Google Scholar] [CrossRef]
- Chipilski, R.; Moskova, I.; Pencheva, A.; Kocheva, K. Enhancement of maize seed viability after cold storage and induced senescence by priming with synthetic cytokinins. Zemdirbyste Agric. 2023, 110, 33–38. [Google Scholar] [CrossRef]
- Arnon, D. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Borisova, D.; Nikolov, H.; Dimitrov, V.; Bouzekova-Penkova, A. Framework concept of the project “Тhe ancient wheat - growth and physiological characteristics under unfavorable stress factors and possibilities to alleviate the negative effects”. Ninth International Conference on Remote Sensing and Geoinformation of Environment, 2023, 12786, SPIE, 27860A. [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Kramer, G.; Norman, H.; Krizek, D.; Mirecki, R. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochem. 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Swain, T.; Goldstein, L. Methods in Polyphenol Chemistry; Pridham, J.B., Ed.; Pergamon Press: Oxford, UK, 1964; pp. 131–146.
- Edreva, A.; Hadjiiska, E. About the determination of sulfhydril (thiol) group content in plant material. Bulg. J. Plant. Physiol. 1984, 10, 73–82. (in Bulgarian). [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.; Szeto, Y.T. Total antioxidant capacity of teas by ferric reducing antioxidant power assay. J. Agric. Food Chem. 1999, 47, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Hristozkova, M.; Geneva, M.; Stancheva, I.; Iliev, I.; Azcon-Aguilar, C. Symbiotic association between golden berry (Physalis peruviana) and arbuscular mycorrhizal fungi in heavy metal-contaminated soil. J. Plant Prot. Res. 2017, 57, 173–184. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebigo, M. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Dias, I.; Costa, M. Effect of low salt concentration on nitrate reductase and peroxidase of sugar beet leaves. J. Exp. Bot. 1983, 34, 537–543. [Google Scholar] [CrossRef]
- Gronwald, J.W.; Fuerst, E.P.; Eberlein, C.V.; Egli, M.A. Effect of herbicide antidotes on glutathione content and glutathione S-transferase activity of sorghum shoots. Pestic. Biochem. Physiol. 1987, 29, 66–76. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nuc. Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Manova, V.; Georgieva, R.; Borisov, B.; Stoilov, L. Efficient removal of cyclobutane pyrimidine dimers in barley: Differential contribution of light-dependent and dark DNA repair pathwys. Physiol. Plant. 2016, 158, 236–253. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 1–11. [Google Scholar] [CrossRef]
- Multiple Primer Analyzer | Thermo Fisher Scientific.
- Karimi, J.; Mohsenzadeh, S.; Niazi, A.; Moghadam, A. Differential expression of mitochondrial manganese superoxide dismutase (SOD) in Triticum aestivum exposed to silver nitrate and silver nanoparticles. Iran. J. Biotechnol. 2017, 15, 284. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Pang, D.; Jin, M.; Chen, J.; Kong, X.; Li, W.; Chang, Y.; Li, Y.; Wang, Z. Identification of plant hormones and candidate hub genes regulating flag leaf senescence in wheat response to water deficit stress at the grain-filling stage. Plant Direct. 2019, 3, e00152. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Leaf (A, B) and root length (C, D) of wheat and einkorn seedlings treated with cytokinin 4PU-30 and subjected to subsequent UV-B radiation. Results are given as means ± SD (n = 20). Different letters represent statistical significance at p < 0.05.
Figure 1.
Leaf (A, B) and root length (C, D) of wheat and einkorn seedlings treated with cytokinin 4PU-30 and subjected to subsequent UV-B radiation. Results are given as means ± SD (n = 20). Different letters represent statistical significance at p < 0.05.
Figure 2.
Averaged reflectance spectra of wheat (A) and einkorn (B) plants treated with excessive UV-B radiation and sprayed with synthetic cytokinin 4PU-30 separately and in combination.
Figure 2.
Averaged reflectance spectra of wheat (A) and einkorn (B) plants treated with excessive UV-B radiation and sprayed with synthetic cytokinin 4PU-30 separately and in combination.
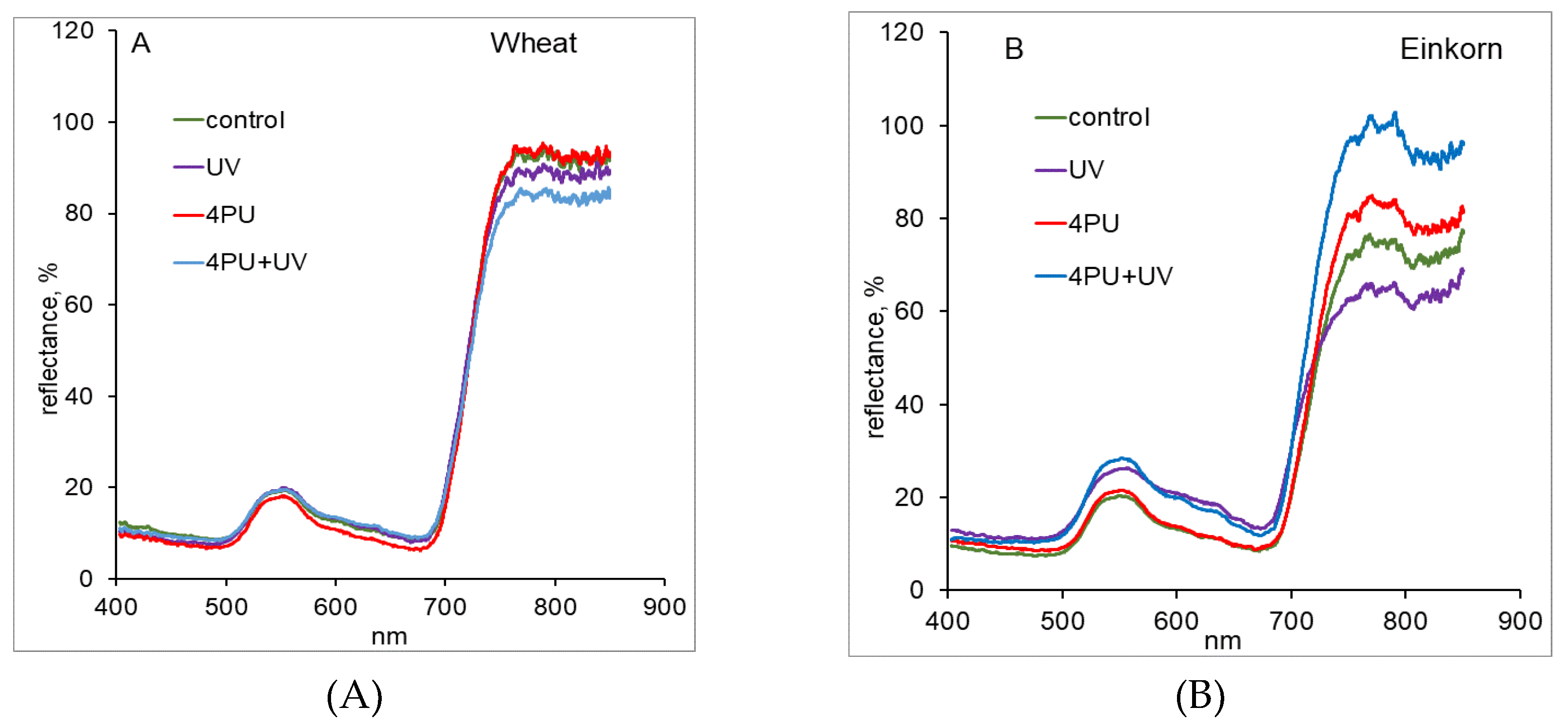
Figure 3.
Oxidative stress markers in the leaves of wheat and einkorn plants treated with excessive UV-B radiation and sprayed with synthetic cytokinin 4PU-30. Amounts of free proline (A, B); Hydrogen peroxide, H2O2 (C, D); Malondialdehyde, MDA (E, F) given as means ± SD (n = 6), different letters represent statistical significance at p < 0.05.
Figure 3.
Oxidative stress markers in the leaves of wheat and einkorn plants treated with excessive UV-B radiation and sprayed with synthetic cytokinin 4PU-30. Amounts of free proline (A, B); Hydrogen peroxide, H2O2 (C, D); Malondialdehyde, MDA (E, F) given as means ± SD (n = 6), different letters represent statistical significance at p < 0.05.
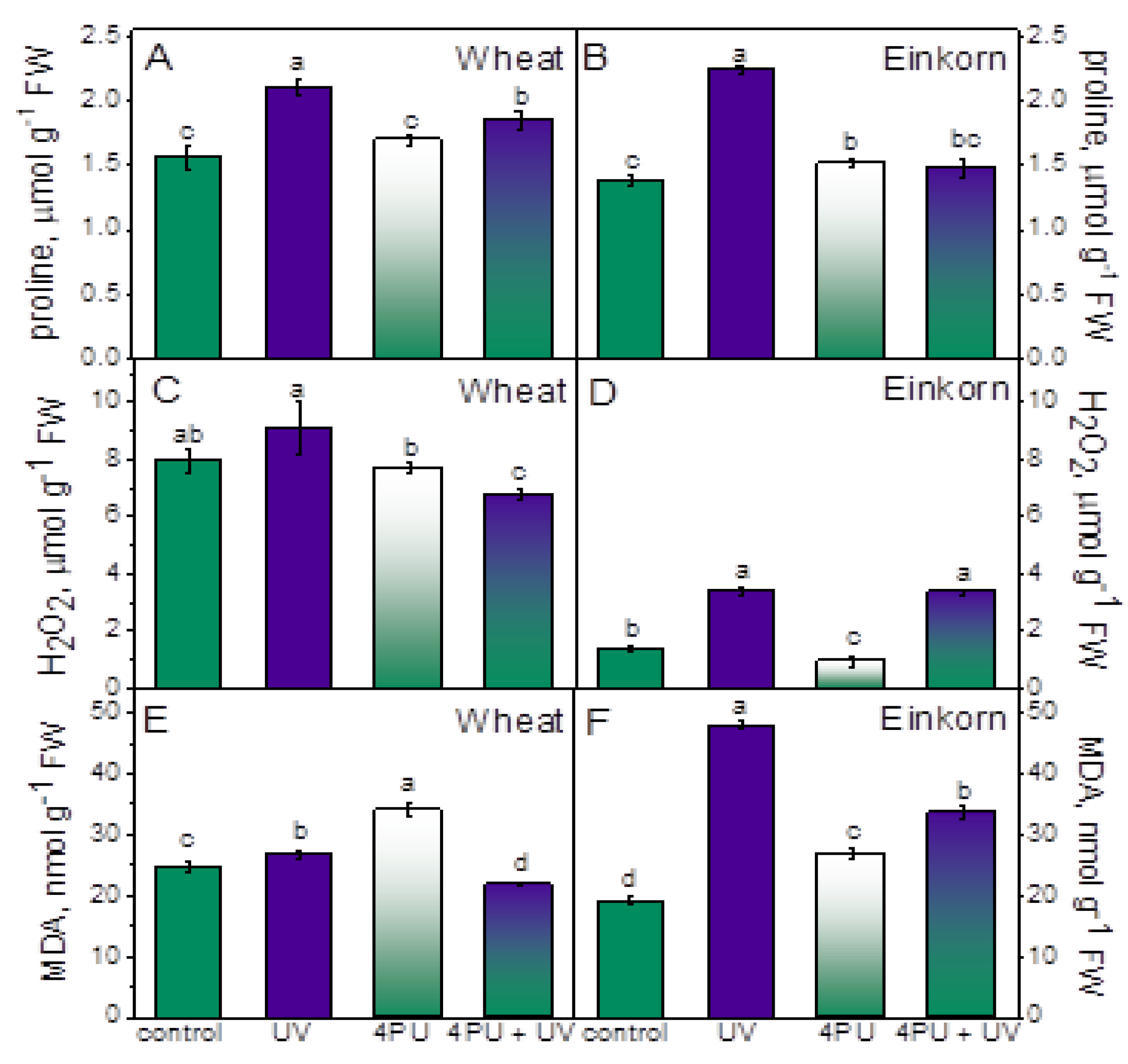
Figure 4.
Non-enzyme antioxidants in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Amounts of free thiol compounds, SH groups (A,B); Flavonoids (C,D) and total phenolic compounds (E, F) in wheat and einkorn respectively given as means ± SD (n = 6). Different letters represent statistical significance at p < 0.05.
Figure 4.
Non-enzyme antioxidants in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Amounts of free thiol compounds, SH groups (A,B); Flavonoids (C,D) and total phenolic compounds (E, F) in wheat and einkorn respectively given as means ± SD (n = 6). Different letters represent statistical significance at p < 0.05.
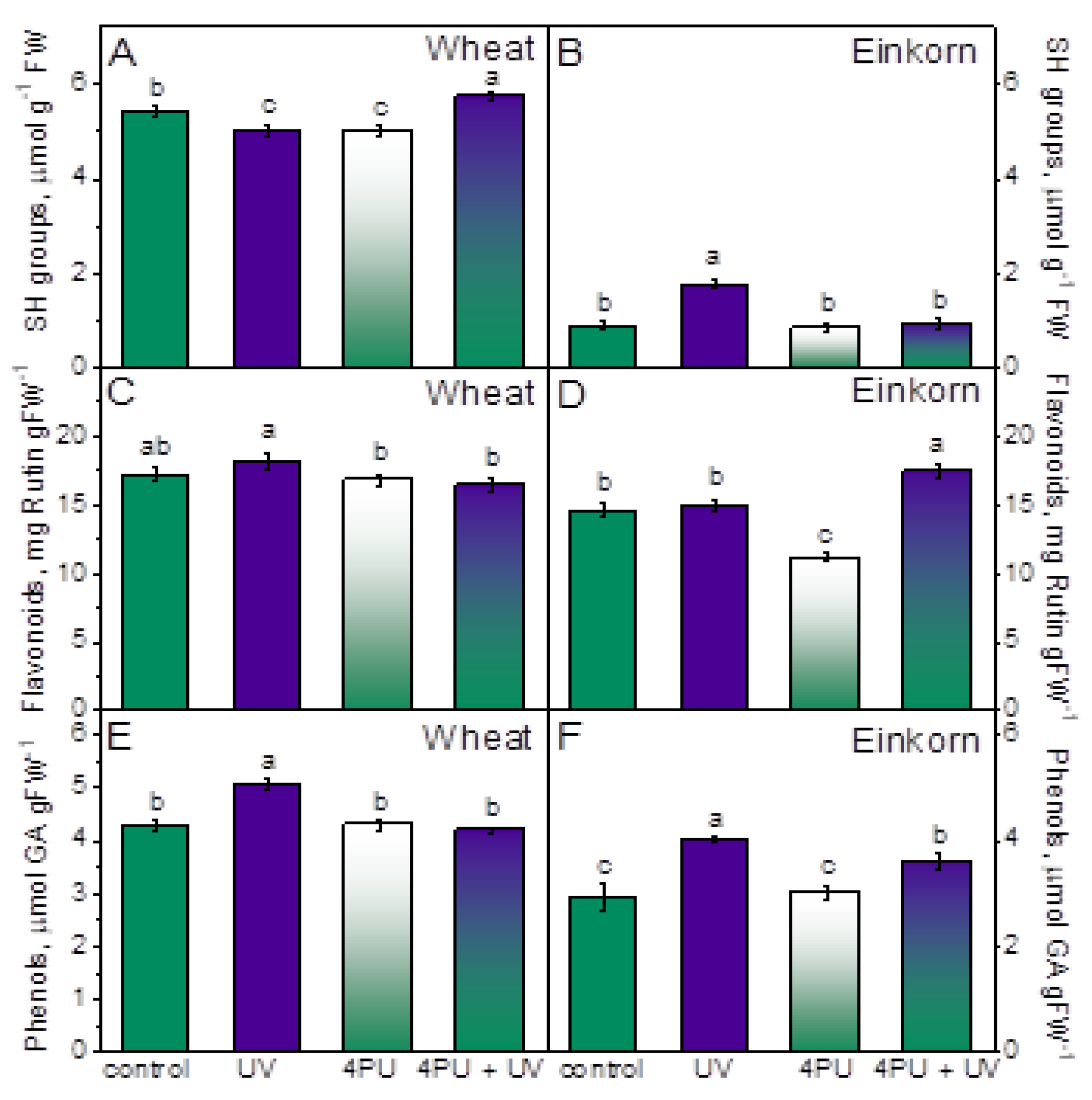
Figure 5.
Free radical scavenging activity measured by the level of DPPH reduction, in % (A, B) and antiradical activity expressed as ferric reducing antioxidant power (FRAP) (C, D) in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Data is given as means ± SD (n = 6). Different letters represent statistical significance at p < 0.05.
Figure 5.
Free radical scavenging activity measured by the level of DPPH reduction, in % (A, B) and antiradical activity expressed as ferric reducing antioxidant power (FRAP) (C, D) in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Data is given as means ± SD (n = 6). Different letters represent statistical significance at p < 0.05.
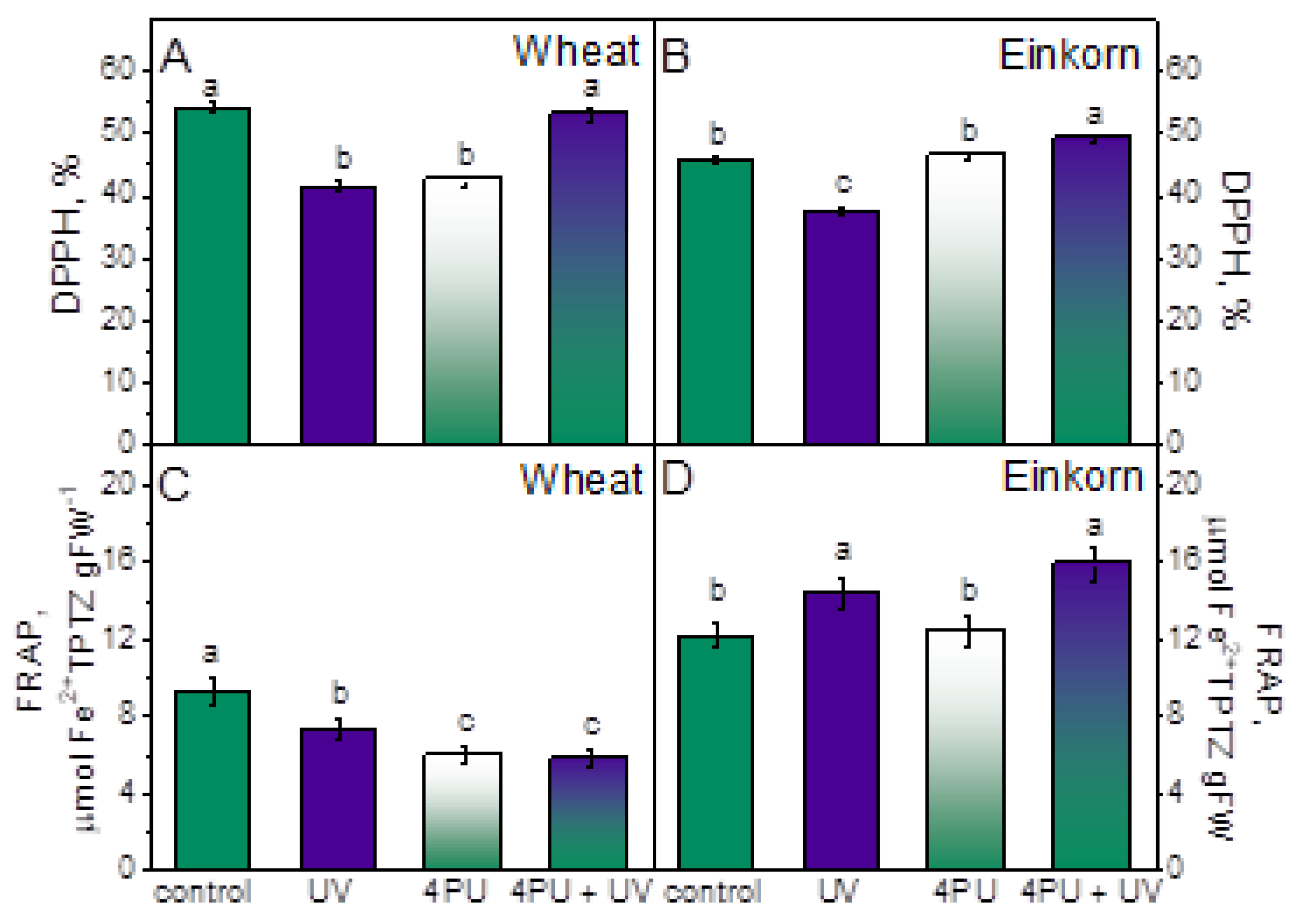
Figure 6.
Activity of antioxidant enzymes in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Superoxide dismutase, SOD (A, B); Catalase, CAT (C, D); Guaiacol peroxidase, POX (E, F); Glutathione S-transferase, GST (G, H). Data is given as means ± SD (n = 6). Different letters represent statistical significance at p < 0.05.
Figure 6.
Activity of antioxidant enzymes in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Superoxide dismutase, SOD (A, B); Catalase, CAT (C, D); Guaiacol peroxidase, POX (E, F); Glutathione S-transferase, GST (G, H). Data is given as means ± SD (n = 6). Different letters represent statistical significance at p < 0.05.
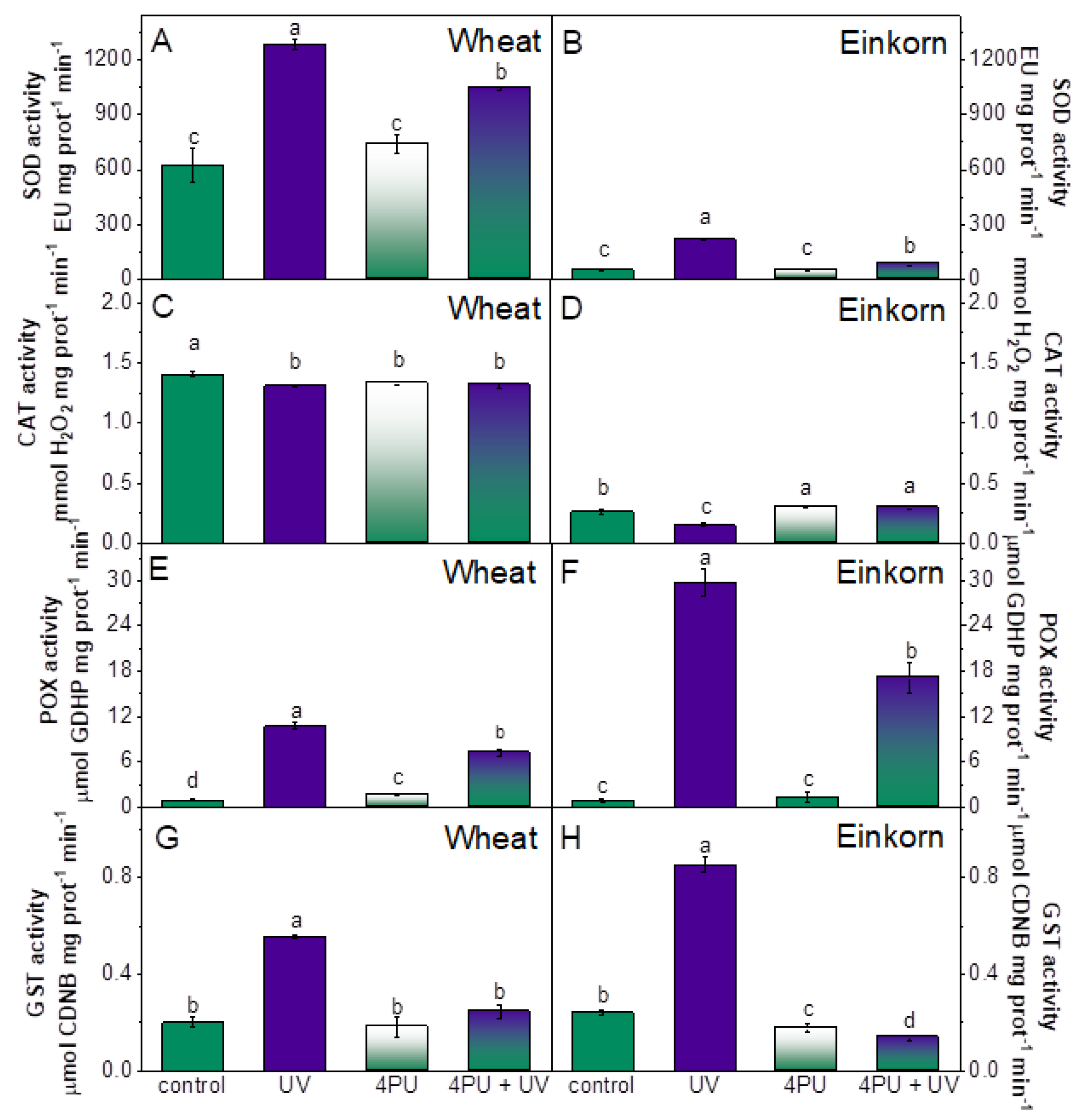
Figure 7.
Relative expression levels of antioxidant enzyme coding genes MnSOD, FeSOD, Cu/ZnSOD, CAT1, POX1, POX7, GSTU61, GSTU62, GSTL analyzed in wheat (A) and einkorn (B). Control (C), UV-B exposure (UV), application of 1 µM 4PU-30 (4PU) and combined treatment with UV-B and 1 µM 4PU-30 (4PU+UV). Results are presented as mean ± SE, n = 3. Asterisks (*) indicate significant difference compared to controls (p≤ 0.05).
Figure 7.
Relative expression levels of antioxidant enzyme coding genes MnSOD, FeSOD, Cu/ZnSOD, CAT1, POX1, POX7, GSTU61, GSTU62, GSTL analyzed in wheat (A) and einkorn (B). Control (C), UV-B exposure (UV), application of 1 µM 4PU-30 (4PU) and combined treatment with UV-B and 1 µM 4PU-30 (4PU+UV). Results are presented as mean ± SE, n = 3. Asterisks (*) indicate significant difference compared to controls (p≤ 0.05).
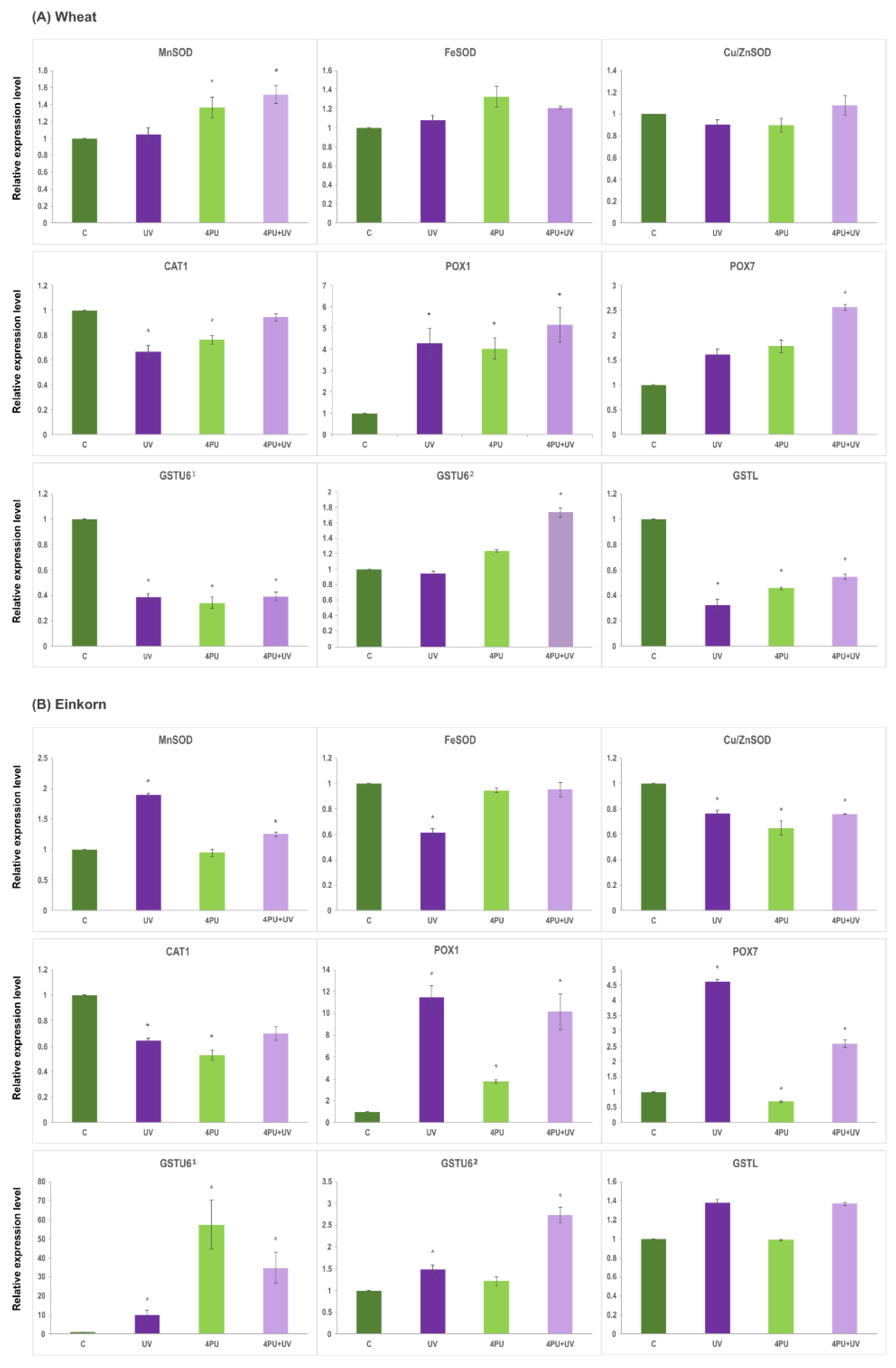
Figure 8.
Hardware components included in the experimental set-up: Ocean Optics USB4000 spectrometer (A), durable fiber optic (B) and certified reflectance standard Labsphere (C).
Figure 8.
Hardware components included in the experimental set-up: Ocean Optics USB4000 spectrometer (A), durable fiber optic (B) and certified reflectance standard Labsphere (C).

Table 1.
Pigment content in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Data are means ± SD (n = 10). Different letters represent statistical significance at p < 0.05.
Table 1.
Pigment content in the leaves of young wheat and einkorn plants illuminated with UV-B, sprayed with synthetic cytokinin 4PU-30 and a combination of the two treatments. Data are means ± SD (n = 10). Different letters represent statistical significance at p < 0.05.
| Variants | Chlorophyll a (mg/g FW) |
Chlorophyll b (mg/g FW) |
Carotenoids (mg/g FW) |
|
|---|---|---|---|---|
| Wheat | ||||
| Control | 1.307 ± 0.036 b | 0.452 ± 0.017 a | 0.799 ± 0.026 a | |
| UV stress | 0.845 ± 0.010 d | 0.270 ± 0.003 c | 0.510 ± 0.006 c | |
| 4PU-30 | 1.419 ± 0.025 a | 0.480 ± 0.011 a | 0.848 ± 0.023 a | |
| 4PU + UV | 1.186 ± 0.015 c | 0.394 ± 0.005 b | 0.670 ± 0.010 b | |
| Einkorn | ||||
| Control | 1.158 ± 0.038 b | 0.392 ± 0.024 b | 0.679 ± 0.024 b | |
| UV stress | 0.756 ± 0.025 c | 0.285 ± 0.011 c | 0.559 ± 0.020 c | |
| 4PU-30 | 1.281 ± 0.036 a | 0.449 ± 0.015 a | 0.750 ± 0.018 a | |
| 4PU + UV | 1.139 ± 0.036 b | 0.395 ± 0.014 b | 0.686 ± 0.023 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



