
Submitted:
25 April 2024
Posted:
25 April 2024
You are already at the latest version
Abstract
Dietary fibers regulate intestinal barrier function; however, the precise mechanisms remain unclear. This study investigated the effects of psyllium fibers on antimicrobial protein expression, focusing on the type II immunity and tuft cell-group 2 innate lymphoid cell (ILC2) circuit in the small intestine of the mouse. Supplemental psyllium fiber upregulated antimicrobial proteins, such as small proline-rich protein 2A (SPRR2A) and resistin-like beta (RELMβ), in mouse small intestine, evidently affecting cecal microbiota composition. Psyllium fibers also increased the RNA and protein expression of molecules related to ILC2 and tuft cells, such as IL-13, IL-25, DCLK1, Gfi-1b, SH2 domain-containing protein 3C, and Spi-B. In addition, ILC2 inhibitor (disulfiram) and bitter taste receptor blocker administration reduced psyllium-induced SPRR2A and RELMβ expression. Collectively, psyllium supplementation upregulates antimicrobial proteins such as SPRR2A and RELMß via the type II immune response and tuft cell-ILC2 circuit in the mouse small intestine.
Keywords:
Antimicrobial protein
; Intestinal microbiota
; Mice
; Psyllium fiber
; Type II immunity
; Tuft cells
1. Introduction
Substantial evidence demonstrates that supplemental intake of dietary fiber provides various health benefits and reduces the risk of developing coronary heart disease, stroke, hypertension, diabetes, and certain intestinal diseases. As a dietary fiber is resistant to digestive enzyme activity, it passes through the small intestine structurally intact and reaches the colon. In the colon, dietary fibers affect the composition and metabolism of intestinal microbiota [1]. Accordingly, dietary fiber supplementation often has physiological effects in a microbiota-dependent manner [2], while dietary fibers directly interact with the intestinal epithelium to affect cellular functions. For example, fructans, including inulin and fructooligosaccharides, protect the intestinal barrier via the protein kinase C pathway [3]. Guar gum fibers reduce the inflammatory reaction via toll-like receptor-2 (TLR-2) and dectin-1 activation [4]. These observations suggest that certain dietary fibers play a role in maintaining intestinal homeostasis in an intestinal microbiota-independent manner; however, the precise mechanisms remain unclear.
Psyllium fiber is produced from the seed husk of Plantago ovata and contains soluble and insoluble fibers with a high quantity of hemicellulose, which consists of a xylan backbone associated with rhamnose, arabinose, and galacturonic acid units (arabinoxylan). Highly branched arabinoxylan is characterized by high water holding and gel formation abilities, which are associated with the physiological effects of psyllium fibers, such as the reduction of sugar and lipid absorption and anti-constipation action [5]. Psyllium supplementation reduces intestinal inflammation induced by a high-fat diet and dextran sulfate sodium in mice [6,7]. In addition, Hino et al., showed that psyllium supplementation increases the number of mucin-producing goblet cells and mucin secretion in the small intestine of the rat, where intestinal bacterial activity is relatively low [8]. Although different mechanisms underlying psyllium-mediated actions may exist [9], psyllium appears to directly interact with intestinal epithelial cells to regulate intestinal function.
Intestinal epithelial cells are responsible for the digestion and absorption of nutrients whilst serving as a barrier to eliminate the permeation of luminal inflammatory substances, including bacteria, helminths, and viruses, into the internal environment of the body. The intestinal barrier is generally composed of chemical (antimicrobial proteins and immunoglobulin A) and physical (intercellular tight junctions and mucus layers) barriers [10]. Several antimicrobial proteins such as defensins, regenerating islet-derived protein 3 (REG3), small proline-rich protein 2A (SPRR2A), resistin-like beta (RELMβ) are produced in the intestine. Antimicrobial proteins protect against pathogenic bacteria to prevent infections [1] and are key determinants of intestinal microbiota composition. Accumulating evidence indicates that the regulation of intestinal antimicrobial protein production is associated with several diseases such as infection, inflammatory bowel diseases, and obesity. Antimicrobial proteins exhibit a site-specific spectrum of antimicrobial activity and expression in the small and large intestines [11]. SPRR2A and RELMβ are primarily produced by Paneth and goblet cells in response to type II immunity [12]. SPRR2A is a bactericidal protein that targets gram-positive bacteria [13]. Its intestinal expression increases during helminth infections and commensal bacteria colonization, suggesting that it plays a role in both helminth clearance and microbial composition. In contrast, RELMβ targets gram-negative bacteria and promotes spatial segregation of the microbiota and intestinal epithelium [14]. Type II cytokines, IL-4 and IL-13, stimulate the production of SPRR2A and RELMβ in the intestines [13,15]. Thus, regulating intestinal antimicrobial proteins may be a novel approach to promote the human health.
The intestinal epithelium is composed of several cell types such as enterocytes, goblet cells, enteroendocrine cells, Paneth cells, and tuft cells, with varying abundances. Tuft cells, accounting for <1% of the total intestinal epithelial cells [16], are relatively rare. Nevertheless, tuft cells play an important role in regulating intestinal homeostasis and serve as key epithelial sentinels that initiate type II mucosal immunity during parasitic and bacterial infections [17]. During helminth infection, tuft cells produce IL-25 to activate group II innate lymphoid cells (ILC2) in the lamina propria, which produce IL-13 [18,19]. In response, epithelial cells produce the antimicrobial proteins such as SPRR2A and RELMβ. Tuft cells express several G-protein-coupled receptors (GPCRs) [20], including bitter taste receptors (type 2 taste receptors, T2Rs), and serve as chemosensory taste cells. Luo et al., demonstrated that T2Rs in tuft cells sense luminal helminths and release IL-25 [21]. However, the involvement of tuft cells and bitter taste receptors in dietary fiber-mediated regulation of intestinal homeostasis has not been elucidated.
The present study investigated the novel effects of psyllium fibers in regulating intestinal homeostasis via direct interaction with the intestinal epithelium in mice. In particular, we focused on the production of antimicrobial proteins via tuft cell-mediated type II immune response.
2. Materials and Methods
2.1. Chemicals
Psyllium fibers were provided by Bizen Chemical Co., Ltd. (Akaiwa, Japan). Probenecid and 6-methoxyflavone were purchased from Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan), and γ-aminobutyric acid was purchased from Nacalai Tesque (Kyoto, Japan). All the other chemicals were purchased from Nacalai Tesque and FUJIFILM Wako Pure Chemicals (Osaka, Japan).
2.2. Mice
All animal experiments were approved by the Animal Care Committee of the Hiroshima University (approval no. C22-44-2), and mice were maintained in accordance with the Guidelines for the Care and Use of Laboratory Animals at Hiroshima University, the National Research Council’s Guide for the Care and Use of Laboratory Animals, and ARRIVE guidelines. Female, 7-week-old C57BL/6J mice, weighing approximately 20 g, were purchased from Charles River Laboratories Japan (Yokohama, Japan). They were housed under controlled temperature (22 ± 2 ºC), relative humidity (40–60 %), and lighting (lights on 08:00–20:00) conditions throughout the study. The mice were allowed to acclimate to the laboratory environment with free access to a fiber-free AIN-93G formula diet (control diet, Supplemental File – Table 1) and tap water for at least 1 week. Mice were euthanized via exsanguination under isoflurane anesthesia.
2.3. Experimental Design
Five animal studies (Experiments 1–5) were conducted to investigate the effects of psyllium fibers on intestinal antimicrobial proteins in mice.
In Experiment 1, the effect of psyllium fibers on gene expression in the small intestine was examined using whole-transcriptome analysis. Mice (n = 16) were randomly allocated to two groups, control and 10% psyllium (n=8/group), and fed the test diets for 5 d (Supplementary File – Figure S1 and Supplementary File – Table S1). Psyllium fiber was added to the control diet by substituting an equal amount of starch. Jejunal samples (1 cm distal to the ligament of Treitz) were subjected to RNA sequencing as described below. Jejunal and ileal samples (2 cm proximal to the ileocecal junction) were subjected to quantitative reverse transcription-polymerase chain reaction (qRT-PCR), immunoblotting, and immunofluorescence analysis. The cecal contents were subjected to intestinal microbiota analysis as described below.
In Experiment 2, the effect of psyllium fiber was examined at different dietary doses in mice. Mice (n = 24) were randomly allocated to four groups: control, 2.5%, 5%, and 10% psyllium (n = 6/group), and mice were fed the corresponding test diets for 5 d (Supplementary File – Figure S1 and Supplementary File – Table S1). Segments of the jejunum, ileum, and proximal colon were subjected to qRT-PCR and immunoblot analyses. Jejunal samples were embedded in OCT compound (Sakura Finetek, Chuoku, Tokyo, Japan) for immunofluorescence analysis, as described below.
In Experiment 3, the effects of psyllium fiber were examined at different feeding periods. Mice (n = 24) were randomly allocated to four groups: control, 5, 10, and 15 d groups (n = 6/group). Mice in the 5, 10, and 15 d groups were fed a 10% psyllium diet for 5, 10, and 15 d, respectively (Supplementary File – Figure S1 and Supplementary File – Table S1). The control mice were fed a control diet for 15 d. Jejunal samples were subjected to qRT-PCR and immunoblot analysis as described below.
In Experiment 4, the role that ILC2 played in the psyllium fiber-mediated effect was examined using the ILC2 inhibitor, disulfiram [22]. Mice (n = 28) were randomly allocated to four groups: control, 5% psyllium, 5% psyllium + low disulfiram, and 5% psyllium + high disulfiram (Supplementary File – Figure S1 and Supplementary File – Table S1). The 5% psyllium + low disulfiram and 5% psyllium + high disulfiram groups were orally administered with disulfiram at 1.2 and 2.4 mg/g body weight (180 and 360 mg/mL ultrapure water), respectively, once daily for 7 d, whereas the control and 5% psyllium groups received ultrapure water only. The mice were fed each test diet for 5 d, 2 d after the start of disulfiram administration. Jejunal samples were subjected to qRT-PCR, immunoblot, and immunofluorescence analysis as described below.
In Experiment 5, the involvement of bitter taste receptors in psyllium fiber-mediated effects was examined using blockers for several T2R families. Mice (n = 28) were randomly allocated to four groups: control, 5% psyllium, 5% psyllium + low blocker, and 5% psyllium + high blocker (Supplementary File – Figure S1 and Supplementary File – Table S2). Three T2R blockers, probenecid, 6-methoxyflavanone, and γ-aminobutyric acid, were added to the diets at 0.25 and 0.5 g/100 g powder for the low and high blocker groups, respectively. Probenecid inhibits T2R16, 38, and 43, 6-methoxyflavone targets T2R1, 7, 14, and 39, while γ-aminobutyric acid acts on T2R4, respectively [23,24,25,26]. The 5% psyllium + low blocker and 5% psyllium + high blocker groups were fed a control diet supplemented with the blockers for 2 d, followed by a 5% psyllium diet supplemented with the blockers for 5 d (Supplementary File – Figure S1). The control and 5% psyllium groups were fed the control and 5% psyllium diets without blockers for 5 d after feeding the control diet for 2 d. Jejunal samples were subjected to qRT-PCR, immunoblotting, and immunofluorescence as described below.
2.4. Whole Transcriptome Analysis of Mouse Small Intestine via RNA Sequencing
Total RNA was isolated from mouse jejunal tissues using NucleoSpin RNA (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions. Eight RNA samples from the control and 10% psyllium groups were combined and submitted to BGI Japan (Kobe, Japan) for eukaryotic transcriptome sequencing analysis with DNBSEQ™ (40 M clean reads (PE100)). The RNA-seq profiles were mapped to the Mus musculus reference genome GCF_000001635.26_GRCm38.p6. The mapped read counts were normalized to transcripts per million (TPM). Data analysis was performed using Dr. Tom software (BGI Japan).
2.5. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Analysis
Total RNA was extracted from the jejunal, ileal, and colonic tissues of mice using NucleoSpin RNA and Sepasol® RNA I Super G (Nacalai Tesque). Total RNA (500 ng) was reverse-transcribed into cDNA using the ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan) according to the manufacturer’s instructions. qRT-PCR was performed using 2× Brilliant III Ultra-Fast SYBR Green QPCR Master Mix (Agilent Technologies, Santa Clara, CA, USA) and a StepOne Real-Time PCR System (Thermo Fisher Scientific). The primer sequences are listed in Supplementary File – Table S3. mRNA expression was calculated using the 2–ΔΔCt method with ribosomal protein S28 (Rps28) as the reference gene.
2.6. Immunoblot Analysis
Mouse jejunal, ileal, and colonic tissues (20 mg) were homogenized in 500 µL lysis buffer [1% (w/v) SDS, 1% (v/v) TritonX-100, and 1% (w/v) sodium deoxycholate in 30 mM Tris with protease and phosphatase inhibitors (pH 7.4)] using a Polytron homogenizer. The protein concentrations were measured using a bicinchoninic acid assay kit (FUJIFILM Wako Pure Chemical). Tissue extracts were mixed with one-half volume of Laemmli’s sample buffer (3× concentrated) containing 6% (w/v) SDS, 30% (v/v) glycerol, 15% (v/v) β-mercaptoethanol, and 0.02% (w/v) bromophenol blue in 188 mM Tris (pH 6.8). Thereafter, the mixture was heated at 95 °C for 10 min. Proteins (20 μg) were separated using SDS-PAGE and transferred to polyvinylidene fluoride membranes. After visualizing total protein using Ponceau S staining, the membranes were incubated with the corresponding primary antibodies and horseradish peroxidase-conjugated anti-rabbit and anti-mouse IgG, and the blots were developed using an enhanced chemiluminescence method (Western Lightning Plus-ECL, PerkinElmer, Waltham, MA, USA). Densitometric analysis of specific bands was performed using the ImageJ software (version 1.53c, NIH, Bethesda, MD, USA). Protein expression was normalized to the total protein level on the membrane, as visualized by Ponceau S staining. Detailed information on the antibodies used is provided in Supplementary File – Table S4.
2.7. Immunofluorescence Analysis
Cryosections of mouse jejunum and ileum were fixed with 4% paraformaldehyde for 10 min, permeabilized with 0.2 % TritonX-100 for 10 min, and blocked with 5 % normal goat or donkey serum for 30 min. The sections were incubated with corresponding primary antibodies at 4 ºC overnight followed by secondary antibodies and DAPI for 1 h. The sections were preserved in anti-fade fluorescence mounting fluid and immunofluorescence was visualized using a Leica FW4000 fluorescence microscope (Leica Microsystems, Wetzlar, Germany). Detailed information on the antibodies used is provided in Supplementary File – Table S4.
2.8. Metagenomic Analysis of 16S rRNA Genes in Intestinal Microbiota
Bacterial DNA present in the cecal contents of mice was extracted using the QuickGene DNA Tissue Kit (Kurabo Industries, Osaka, Japan) according to the manufacturer’s instructions. Library preparation for analysis using an Illumina MiSeq Next Generation Sequencer (Illumina, San Diego, CA, USA) was performed as previously described [27]. PCR amplification of the 16S rRNA V3-V4 regions (460 bp) was performed using primers 314F and 805R. Sequence data were analyzed using the Quantitative Insights into Microbial Ecology (QIIME) 2 (ver. 2021.11) [28], with denoising by the DADA2 plugin. Amplicon sequence variants (ASVs) was defined and their taxonomy was classified by the sklearn-classifier against the SILVA database (138; 99% Operational Taxonomic Units (OTUs) full-length sequences). In this study, we removed singletons and ASVs assigned to mitochondria and chloroplasts. Subsequently, a phylogenetic tree was generated using SATé-enabled phylogenetic placement (SEPP) [29].
2.9. Statistical Analyses
Data were analyzed using JMP software (version 16; SAS Institute Inc., Cary, NC, USA) and presented as the mean ± SEM. Two-tailed unpaired Student’s t-test was used for two-group comparisons. For multiple group comparisons, Tukey–Kramer post-hoc test and Steel–Dwass test were used. P <0.05 was considered statistically significant.
3. Results
3.1. Psyllium Supplementation Upregulates the Production of Antimicrobial Proteins in the Mouse Small Intestine
Whole transcriptome analysis was performed to identify the genes affected by psyllium supplementation in the small intestine (jejunum) of mice. Mice were fed the control or 10% psyllium diets for 5 d (Supplementary File – Figure S1). Psyllium supplementation led to a more than 2-fold upregulation of 503 genes and downregulation of 218 genes compared with the control group (Figure 1A). Of the 503 genes, several encoding antimicrobial proteins, such as small proline-rich protein 2a (Sprr2a), resistin-like beta (Retnlb), and defensins (Defa), were upregulated by psyllium supplementation (Figure 1B). We focused on the regulation of Sprr2A and Retnlb by psyllium fiber owing to the recent discovery of the antimicrobial activity of SPRR2A [13], Sprr2a having relatively high TPM among these antimicrobial proteins, and the greater upregulation of Retnlb by psyllium fibers than others. qRT-PCR analysis confirmed that Sprr2a and Retnlb mRNA expression levels were approximately 6-fold and 200-fold higher than those in control mice, respectively (Figure 1C,D). Immunoblot and immunofluorescence analysis also showed that the protein expression of SPRR2A and RELMß in the psyllium group was higher than that in the control mice (Figure 1E–H). Consistent with previous studies, SPRR2A and RELMβ were predominantly expressed in goblet and Paneth cells [15]. These results showed that the psyllium supplementation upregulates the production of antimicrobial proteins such as SPRR2A and RELMβ in the mouse small intestine.
3.2. Psyllium Supplementation Alters Cecal Microbiota Composition
The cecal microbiota, which is more relevant to the small intestinal microbiota than fecal microbiota, was analyzed in mice fed control and 10% psyllium diets for 5 d. Psyllium supplementation reduced the diversity of cecal microbiota, as indicated by the lower observed species number and Chao1 index in the 10% psyllium group compared to those in the control mice (Figure 2A,B). The Shannon index, which indicates the evenness of the microbiota composition, in the 10% psyllium group was also lower than that in the control group (Figure 2C). The principal coordinates analysis (PCoA) based on the Weighted UniFrac, Unweighted UniFrac, and Bray–Curtis distance revealed that psyllium supplementation had an impact on the β-diversity of the microbiota (Figure 2D–F). Clustering in these analyses differed between the two groups. At the phylum level, the relative abundances of Firmicutes, Desulfobacterota, Patescibacteria, and Compilobacterota in the 10% psyllium group were lower than those in the control group (Figure 2G). Conversely, the Verrucomicrobiota, Proteobacteria, and Cyanobacteria phyla in the 10% psyllium group were higher than those in the control group. The bacterial genera affected by the psyllium fibers are shown in Figure 2H,I, and, Supplementary File – Figure S2. Figure 2H,I show the genera representing the relative abundances >5% and 1% in either group, respectively. All gram-positive genera presented in Figure 2H,I, such as Desulfovibrionaceae_uncultured, Lachnospiraceae_unclassified, Blautia, Lachnospiraceae_NK4A136_group, Oscillospiraceae_unclassified, Oscillibacter, and Oscillospiraceae_uncultured in the 10% psyllium group were lower than those in the control group. While some gram-negative genera such as Akkermansia, Bacteroides, Escherichia-Shigella, Rikenellaceae_RC9_gut_group, Gastranaerophilales, Parabacteroides, and Parasutterella in the 10% psyllium group were higher than those in the control group, other gram-negative genera in the 10% psyllium group were lower than those in the control group. The genera >0.5 % in both groups are shown in Supplementary File – Figure S2A. Psyllium supplementation increased the abundance of gram-positive genera, such as Enterococcus, GCA-900066575, Marvinbryantia, and [Eubacterium]_coprostanoligenes_group, and the gram-negative genus Helicobacter. Other gram-positive and -negative genera were reduced by psyllium fiber feeding. Several genera >0.1% in both groups, shown in Supplementary File – Figure S2B, were also affected by psyllium fibers.
3.3. Psyllium Supplementation Upregulates Antimicrobial Proteins, SPRR2A and RELMβ, in a Dose-Dependent Manner
To investigate the effects of different doses of psyllium fiber on the production of SPRR2A and RELMβ, mice were fed diets containing psyllium at 2.5, 5, and 10% for 5 d (Supplementary File – Figure S1). Psyllium supplementation increased SPRR2A and RELMβ at the mRNA and protein levels in the jejunum in a dose-dependent manner (Figure 3A–D), whereas the increases in SPRR2A and RELMβ expression were similar with 2.5, 5, and 10% psyllium diets, except for RELMβ protein expression in the ileum (Figure 3E–H). The results also showed that the psyllium-induced increases in the mRNA and protein expression of SPRR2A and RLMMβ in the jejunum were greater than those in the ileum. In contrast, SPRR2A expression in the colon did not differ at either the mRNA or protein level (Figure S3A,C). The Retnlb mRNA expression, but not its protein expression, in the colon of mice fed the 5 and 10% psyllium diets was higher than that in the control mice (Supplementary File – Figure S3B,D). Based on the observations mentioned above, we focused on the effects of psyllium in the mouse jejunum.
3.4. Psyllium Fiber Sustains Upregulation of Antimicrobial Proteins for 15 d
To investigate whether the upregulation of antimicrobial proteins by psyllium fibers was sustained longer than 5 d, mice were fed 10% psyllium for 0, 5, 10, and 15 d (Supplementary File – Figure S1). Consistent with previous observations, psyllium supplementation for 5 d increased SPRR2A (p<0.001) and RELMß (p<0.001) at the mRNA and protein levels in the jejunum (Supplementary File – Figure S4A–D). Significant increases were also observed in the 10 and 15 d psyllium supplementation groups compared to the control group. These results showed that the upregulation of SPRR2A and RELMß by psyllium fibers is sustained for at least 15 d.
3.5. Psyllium Supplementation Increases Tuft Cell Proliferation via IL-13 Signaling
Given that previous studies demonstrated type Ⅱ immunity via the tuft cells-ILC2 circuit was involved in the production of SPRR2A and RELMβ the small intestine [12,30], we examined whether psyllium supplementation affected tuft cell proliferation and the type Ⅱ immune response in the mouse jejunum. qRT-PCR analysis showed that psyllium supplementation dose-dependently increased the mRNA expression of tuft cell markers (Pou2f3, Dclk1, and Gfi1b); however, the increases of the 2.5 and 5% psyllium diets were not statistically significant (Figure 4A–C). Sh2d6 and Tppp3 are markers of tuft-2 and tuft-1 cells, respectively, in mouse small intestine [31]. Sh2d6 mRNA expression in the 10% psyllium group was higher than that in the control group. However, there was no difference in Tppp3 mRNA expression among the groups, indicating the involvement of tuft-2 cells in psyllium-mediated effects (Figure 4D,E). In addition, Spib mRNA encoding the SpiB transcription factor, which plays a role in tuft-2 development, in the 10% psyllium group was higher than that in the control group [31]. The number of DCLK1+ cells indicating tuft cells per crypt in the 10 % psyllium group was higher than that in the control group, as determined by immunofluorescence staining (Figure 4G). Tuft cells are the primary source of IL-25 [32], which induces IL13 secretion in ILC2 to promote STAT6 phosphorylation in the intestinal epithelium [30]. Psyllium supplementation dose-dependently increased Il25 and Il13 mRNA expression in the small intestine, with significant increases in 5 and 10% psyllium diets (Figure 4H,I). No difference was observed for the Il4 or Il33 mRNA expression among groups (Figure 4J and Supplementary File – Figure S5). The phosphorylation of STAT6, a transcription factor that regulates Sprr2a and Retnlb transcription, in the 5 and 10 % psyllium groups was higher than that in the control group (Figure 4K). These results suggested that the production of antimicrobial proteins (SPRR2A and RELMβ) by psyllium fibers is induced by the type Ⅱ cytokine IL-13 via the tuft cells-ILC2 circuit.
3.6. ILC2 is Involved in the Psyllium-Induced Antimicrobial Protein Production
To investigate the role of ILC2 in psyllium-induced production of antimicrobial proteins, an ILC2 inhibitor, disulfiram, was orally administered to mice in two doses (Supplementary File – Figure S1). Consistent with previous observations, the mRNA and protein expression of antimicrobial proteins, SPRR2A and RELMβ, in the mouse small intestine was increased by the 5% psyllium diet (Figure 5A–F). Disulfiram administration at low and high doses suppressed the psyllium-induced SPRR2A and RELMβ expression. Disulfiram administration also reduced psyllium-induced upregulation of Il13 mRNA and pSTAT6 protein expression (Figure 5G–I). These results indicated the involvement of ILC2 in antimicrobial protein production by psyllium fibers. Similarly, disulfiram administration inhibited the psyllium-induced mRNA expression of tuft cell-associated molecules Pou2f3 and Il25 (Figure 5J–L). These results suggested the presence of a positive feedback loop from the type II immune response to the tuft cells.
3.7. Bitter Taste Receptors of Tuft Cells are Involved in Psyllium-Induced Antimicrobial Protein Production
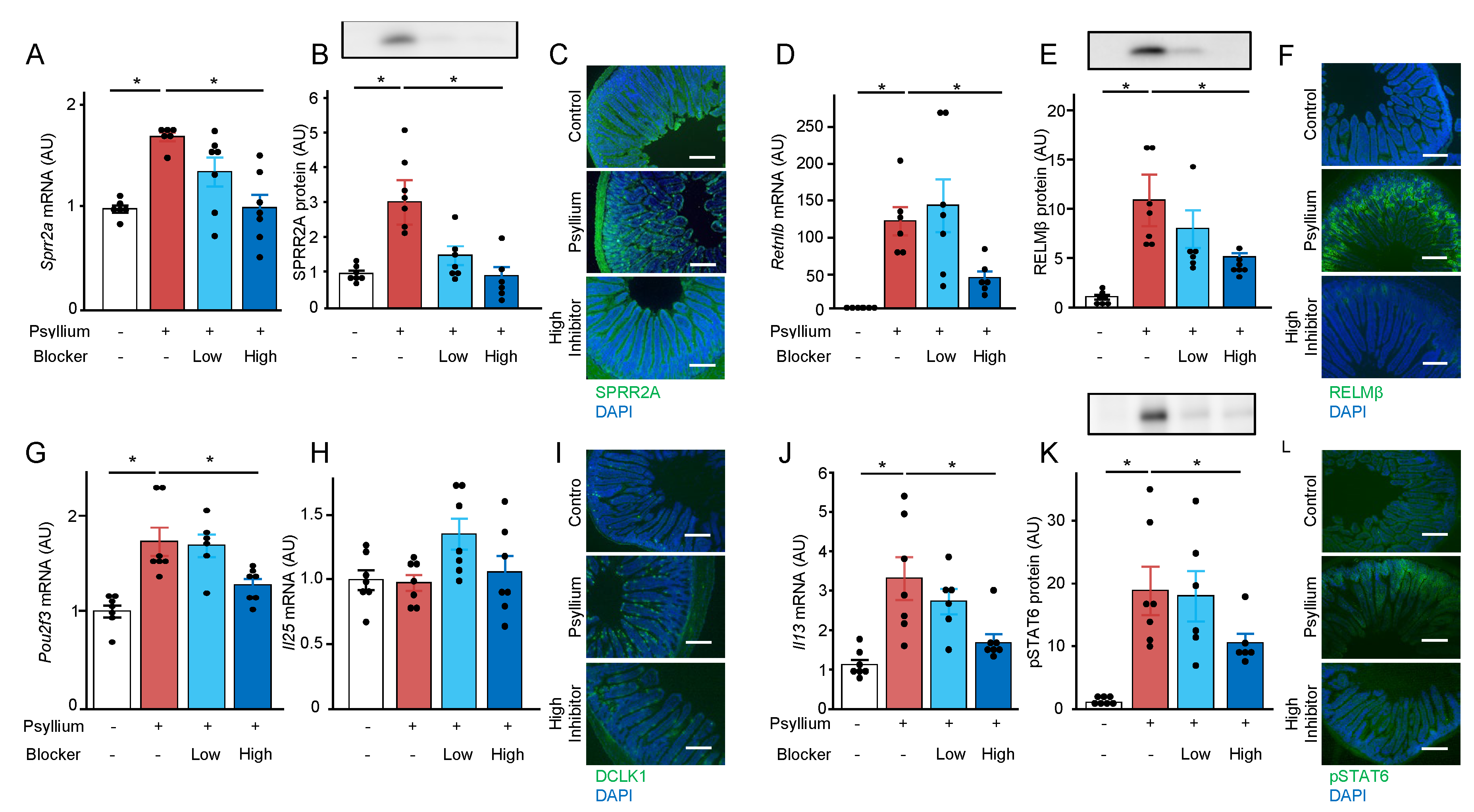
Bitter taste receptors expressed in tuft cells are involved in the activation of tuft cell-ILC2 circuits to induce antimicrobial protein production in the mouse small intestine [21]. To investigate whether bitter taste receptors are involved in psyllium-mediated SPRR2A and RELMβ expression, mice were simultaneously administered three blockers of bitter taste receptors (probenecid, 6-methoxyflavanone, and γ-aminobutyric acid) in two doses. As observed in previous findings, protein and mRNA expression of SPRR2A and RELMβ was upregulated by 5% psyllium diets. However, this upregulation was suppressed by the administration of bitter taste receptor blockers at high doses (Figure 6A–F). Similarly, the administration of blockers at high doses reduced psyllium-induced Pou2f3 mRNA, Il13 mRNA, and pSTAT6 protein expression (Figure 6G–L). These results suggested that the bitter taste receptors expressed in tuft cells play a role in sensing luminal psyllium fibers to induce SPRR2A and RELMβ expression.
4. Discussion
Although the physiological effects of dietary fibers are often exerted through metabolism by the intestinal microbiota, direct interaction of dietary fibers with intestinal cells also regulates intestinal functions [4,33]. This study demonstrated that supplemental feeding of psyllium fiber upregulates antimicrobial proteins including SPRR2A and RELMβ in the small intestine of the mouse, an area where microbial activity is relatively lower than that in the colon. Our results suggested that the psyllium-mediated effect is involved in type II immunity through the tuft cell-ILC2 circuit. Bitter taste receptors expressed in tuft cells likely sense luminal psyllium fibers, eliciting an intestinal response. Recently, the regulation of type II immunity by helminths has been proposed as a therapeutic option for some immune disorders such as inflammatory bowel diseases, however, the prolonged helminth infection can cause complications [34]. In addition, the intestinal barrier and microbiota are involved in the pathogenesis of not only intestinal diseases, but also extraintestinal diseases such as obesity and diabetes [35]. Accordingly, the supplemental psyllium fiber may lead to the development of safe, effective, and novel therapeutic approaches for these disorders.
Psyllium supplementation increases the production of SPRR2A and RELMβ in the small intestine, indicating that psyllium fiber directly stimulates the intestinal epithelial cells in a microbiota-independent manner. Antimicrobial proteins that rapidly kill or inactivate microorganisms are among the most evolutionarily ancient immune defense mechanisms [36]. The upregulation of antimicrobial proteins can reduce bacterial infections and alter the intestinal microbiota composition. Psyllium supplementation drastically altered cecal microflora composition in mice. Each antimicrobial protein has a specific antibacterial spectrum. SPRR2A and RELMβ selectively kill gram-positive and -negative bacteria, respectively [13,14]. Hu et al. demonstrated that SPRR2A-deficient mice exhibited increased levels of gram-positive bacteria in the small intestinal lumen and mucus layer [13], and were more susceptible to infection by Listeria monocytogenes, a pathogenic gram-positive bacterium. In contrast, RELMβ-deficient mice showed an increase in the entry of Gram-negative commensal bacteria, including Proteobacteria, into the colon inner mucus layer [14]. Upon infection by Citrobacter rodentium, the colonic tissue-associated bacteria in RELMβ-deficient mice were higher than that in wild type mice. This evidence suggested that the psyllium-mediated production of antimicrobial proteins such as SPRR2A and RELMβ reduces the susceptibility to bacterial infection in both physiological and pathological conditions. However, the effect of psyllium fiber on cecal microbiota composition was complex, possibly due to upregulation of other antimicrobial proteins, not just SPRR2A and RELMβ. For example, psyllium supplementation increased Muribaculaceae and decreased Bacteroides, although both belong to gram-negative genera. In the gram-positive genera, psyllium fiber supplementation decreased several genera, whereas some genera, such as Enterococcus, Marvinbryantia, and [Eubacterium]_coprostanoligenes_group increased. Further research is required to determine the physiological and pathological implications of psyllium-mediated regulation of the intestinal microbiota.
Our results showed that the psyllium-mediated upregulation of SPRR2A and RELMβ is involved in the type Ⅱ immune response via the tuft cell-ILC2 circuit. Tuft cells are the primary source of IL-25 in the intestine, rapidly producing IL-25 to initiate the type Ⅱ immune response during helminth infections [18]. The tuft cell-derived IL-25 activates ILC2 to induce the IL-13 production [17], consequently stimulating Paneth and goblet cells to produce the antimicrobial proteins such as SPRR2A and RELMβ via the phosphorylation of a transcription factor, STAT6 [17,37]. In our study, psyllium supplementation increased the levels of type II immunity-related molecules such as IL-25 and IL-13 and antimicrobial proteins. In addition, an ILC2 inhibitor, disulfiram, reduced the psyllium-mediated upregulation of SPRR2A, RELMβ, STAT6 phosphorylation, IL-25, and IL-13.
Administration of bitter taste receptor blockers (probenecid, 6-methoxyflavanone, and γ-aminobutyric acid) reduced the psyllium-mediated upregulation of SPRR2A and RELMβ in mice, suggesting that the bitter taste receptors expressed in tuft cells are involved in sensing the luminal psyllium fiber. Tuft cells express several GPCRs, including bitter taste receptors (T2Rs) [38]. Luo et al., demonstrated that T2R detects parasitic helminths, initiating type II immunity [21]. In mice, there are at least 35 Tas2r genes, and the blockers used in the study (probenecid, 6-methoxyflavanone, and γ-aminobutyric acid) inhibit eight T2Rs viz., T2R1, 4, 7, 14, 16, 38, 39, and 47 [23,24,25,26], suggesting that tuft cells sense the luminal psyllium fiber via one or several of these TR2s. Whereas, succinate produced by helminth has been reported to activate the type II immunity through the succinate receptor (Sucnr1) of tuft cells [38]. However, our previous study demonstrated that supplemental psyllium fiber did not influence the succinate production in the mouse cecum [6]. Recent single-cell RNA sequencing analysis has categorized intestinal tuft cells into two subsets: tuft-1 and tuft-2. Tuft-2 cells are specified by Sh2d6 expression and three transcription factors: Pou2f3, Gfi1b, and Spib, which play a role in the development of tuft-2 cells [31]. Psyllium supplementation increased these tuft-2 cells-related molecules, and not the tuft-1 cell marker Tppp3. This suggested that tuft-2 cells sense luminal psyllium as a chemosensor to mediate type II immune responses and the production of antimicrobial proteins in the small intestine. Similarly, [31] demonstrated that tuft-2 cells sense the bacterial metabolite, N-undecanoyl glycine, via another GPCR, Vmn2r26, to exert antimicrobial immunity.
Although this study revealed some mechanisms underlying psyllium fiber-induced antimicrobial proteins in the small intestine, some limitations are to be addressed in future studies. First, this study was conducted exclusively in mice, necessitating the investigation of psyllium-mediated effects in humans. Second, the specific bitter taste receptors that sense psyllium fibers remain unclear, and understanding the molecular interactions between psyllium fibers and these receptors is essential.
5. Conclusions
Supplemental feeding of psyllium fibers upregulates antimicrobial protein production in the small intestine of mice. Intestinal tuft-2 cells may sense luminal psyllium fibers via bitter taste receptors and secrete IL-25. The tuft-2 cell-derived IL-25 stimulates the ILC2 to produce the IL-13, which consequently induces production of antimicrobial proteins such as SPRR2A and RELMβ in Paneth and goblet cells. Intestinal antimicrobial proteins exhibit broad-spectrum and high-efficiency antibacterial activities and are potent and versatile effector molecules in the innate immune system. Psyllium-mediated antimicrobial proteins may help prevent bacterial infections and influence intestinal microbiota. Although further studies should address some limitations such as translating the results from mice to humans and identifying the bitter taste receptor for psyllium fibers, our findings highlight the role of dietary fibers in regulating intestinal homeostasis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Experimental design and schedule; Figure S2. Psyllium supplementation alters the cecal microbiome profile in mice; Figure S3. Effect of psyllium fibers on antimicrobial protein expression, SPRR2A and RELMβ, in mouse colon; Figure S4. Psyllium fiber sustains upregulation of antimicrobial proteins in the mouse small intestine for 15 d; Figure S5. Psyllium supplementation did not affect the Il33 mRNA expression in the mouse small intestine; Table S1. Diet compositions in Experiment 1, 2, 3, and 4; Table S2. Diet compositions in Experiment 5; Table S3. Primers for qRT-PCR analysis used in this study; Table S4. The details of antibodies used in this study.
Author Contributions
Conceptualization, Y.I. and T.S.; methodology, Y.I. and T.S.; validation, T.S. and Y.Y.; investigation, Y.I., T.M., T.Y., DM.R. and R.I.; writing - original draft preparation, Y.I.; writing- reviewing and editing, T.S.; visualization, Y.I.; supervision, T.S.; project administration, T.S.; funding acquisition, T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by JSPS Kakenhi (Grant Number 22H03512) and the Tojuro Iijima Foundation for Food Science and Technology.
Institutional Review Board Statement
All animal experiments were approved by the Animal Care Committee of the Hiroshima University (approval no. C22-44-2), and mice were maintained in accordance with the Guidelines for the Care and Use of Laboratory Animals at Hiroshima University, the National Research Council’s Guide for the Care and Use of Laboratory Animals, and ARRIVE guidelines.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available on request.
Acknowledgments
We would like to thank Editage (www.editage.jp) for their English language editing services.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Blyth, G.A.D.; Connors, L.; Fodor, C.; Cobo, E.R. The Network of Colonic Host Defense Peptides as an Innate Immune Defense Against Enteropathogenic Bacteria. Front Immunol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.Y.; Abdullah, M.; Määttänen, P.; Pilar, A.V.C.; Scruten, E.; Johnson-Henry, K.C.; Napper, S.; O’Brien, C.; Jones, N.L.; Sherman, P.M. Protein Kinase Cσ Signaling Is Required for Dietary Prebiotic-Induced Strengthening of Intestinal Epithelial Barrier Function. Sci Rep 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, T.; Suzuki, T. Guar Gum Fiber Increases Suppressor of Cytokine Signaling-1 Expression via Toll-like Receptor 2 and Dectin-1 Pathways, Regulating Inflammatory Response in Small Intestinal Epithelial Cells. Mol Nutr Food Res 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Van Der Schoot, A.; Drysdale, C.; Whelan, K.; Dimidi, E. The Effect of Fiber Supplementation on Chronic Constipation in Adults: An Updated Systematic Review and Meta-Analysis of Randomized Controlled Trials. American Journal of Clinical Nutrition 2022, 116, 953–969. [Google Scholar] [CrossRef]
- Ogata, M.; Van Hung, T.; Tari, H.; Arakawa, T.; Suzuki, T. Dietary Psyllium Fiber Increases Intestinal Heat Shock Protein 25 Expression in Mice. Nutrition Research 2017, 39, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Bretin, A.; Yeoh, B.S.; Ngo, V.L.; Reddivari, L.; Pellizzon, M.; Vijay-Kumar, M.; Gewirtz, A.T. Psyllium Fiber Protects Mice against Western Diet-Induced Metabolic Syndrome via the Gut Microbiota-Dependent Mechanism. Gut Microbes 2023, 15, 2221095. [Google Scholar] [CrossRef] [PubMed]
- Hino, S.; Takemura, N.; Sonoyama, K.; Morita, A.; Kawagishi, H.; Aoe, S.; Morita, T. Small Intestinal Goblet Cell Proliferation Induced by Ingestion of Soluble and Insoluble Dietary Fiber Is Characterized by an Increase in Sialylated Mucins in Rats. Journal of Nutrition 2012, 142, 1429–1436. [Google Scholar] [CrossRef]
- Bretin, A.; Zou, J.; San Yeoh, B.; Ngo, V.L.; Winer, S.; Winer, D.A.; Reddivari, L.; Pellizzon, M.; Walters, W.A.; Patterson, A.D.; et al. Psyllium Fiber Protects Against Colitis Via Activation of Bile Acid Sensor Farnesoid X Receptor. CMGH 2023, 15, 1421–1442. [Google Scholar] [CrossRef]
- France, M.M.; Turner, J.R. The Mucosal Barrier at a Glance. J Cell Sci 2017, 130, 307–314. [Google Scholar] [CrossRef]
- Byndloss, M.X.; Pernitzsch, S.R.; Bäumler, A.J. Healthy Hosts Rule within: Ecological Forces Shaping the Gut Microbiota. Mucosal Immunol 2018, 11, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Fung, C.; Fraser, L.M.; Barrón, G.M.; Gologorsky, M.B.; Atkinson, S.N.; Gerrick, E.R.; Hayward, M.; Ziegelbauer, J.; Li, J.A.; Nico, K.F.; et al. Tuft Cells Mediate Commensal Remodeling of the Small Intestinal Antimicrobial Landscape. Proc Natl Acad Sci U S A 2023, 120. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhang, C.; Sifuentes-Dominguez, L.; Zarek, C.M.; Propheter, D.C.; Kuang, Z.; Wang, Y.; Pendse, M.; Ruhn, K.A.; Hassell, B.; et al. Small Proline-Rich Protein 2A Is a Gut Bactericidal Protein Deployed during Helminth Infection. Science (1979) 2021, 374. [Google Scholar] [CrossRef] [PubMed]
- Propheter, D.C.; Chara, A.L.; Harris, T.A.; Ruhn, K.A.; Hooper, L. V. Resistin-like Molecule β Is a Bactericidal Protein That Promotes Spatial Segregation of the Microbiota and the Colonic Epithelium. Proc Natl Acad Sci U S A 2017, 114, 11027–11033. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, K.S.B.; Morampudi, V.; Chan, J.M.; Bhinder, G.; Lau, J.; Yang, H.; Ma, C.; Huang, T.; Ryz, N.; Sham, H.P.; et al. Goblet Cell Derived RELM-β Recruits CD4+ T Cells during Infectious Colitis to Promote Protective Intestinal Epithelial Cell Proliferation. PLoS Pathog 2015, 11. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; O’Leary, C.E.; Locksley, R.M. Regulation of Immune Responses by Tuft Cells. Nat Rev Immunol 2019, 19, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Howitt, M.R.; Lavoie, S.; Michaud, M.; Blum, A.M.; Tran, S. V.; Weinstock, J. V.; Gallini, C.A.; Redding, K.; Margolskee, R.F.; Osborne, L.C.; et al. Tuft Cells, Taste-Chemosensory Cells, Orchestrate Parasite Type 2 Immunity in the Gut. Science 2016, 351, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Von Moltke, J.; Ji, M.; Liang, H.E.; Locksley, R.M. Tuft-Cell-Derived IL-25 Regulates an Intestinal ILC2-Epithelial Response Circuit. Nature 2016, 529, 221–225. [Google Scholar] [CrossRef]
- Gerbe, F.; Sidot, E.; Smyth, D.J.; Ohmoto, M.; Matsumoto, I.; Dardalhon, V.; Cesses, P.; Garnier, L.; Pouzolles, M.; Brulin, B.; et al. Intestinal Epithelial Tuft Cells Initiate Type 2 Mucosal Immunity to Helminth Parasites. Nature 2016, 529, 226–230. [Google Scholar] [CrossRef]
- McGinty, J.W.; Ting, H.A.; Billipp, T.E.; Nadjsombati, M.S.; Khan, D.M.; Barrett, N.A.; Liang, H.E.; Matsumoto, I.; von Moltke, J. Tuft-Cell-Derived Leukotrienes Drive Rapid Anti-Helminth Immunity in the Small Intestine but Are Dispensable for Anti-Protist Immunity. Immunity 2020, 52, 528–541.e7. [Google Scholar] [CrossRef]
- Luo, X.C.; Chen, Z.H.; Xue, J.B.; Zhao, D.X.; Lu, C.; Li, Y.H.; Li, S.M.; Du, Y.W.; Liu, Q.; Wang, P.; et al. Infection by the Parasitic Helminth Trichinella Spiralis Activates a Tas2r-Mediated Signaling Pathway in Intestinal Tuft Cells. Proc Natl Acad Sci U S A 2019, 116, 5564–5569. [Google Scholar] [CrossRef] [PubMed]
- Mucke, H.A.M. Drug Repurposing Patent Applications January-March 2020. Assay Drug Dev Technol 2020, 18, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Davaatseren, M.; Hwang, J.T.; Park, J.H.; Kim, M.S.; Wang, S.; Sung, M.J. Allyl Isothiocyanate Ameliorates Angiogenesis and Inflammation in Dextran Sulfate Sodium-Induced Acute Colitis. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, F.; Dey, B.; Yoshida, Y.; Nishimura, S.; Tabata, S. Bitter Taste Receptor Antagonists Inhibit the Bitter Taste of Canola Meal Extract in Chickens. Journal of Poultry Science 2020, 57, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Prandi, S.; Bromke, M.; Hübner, S.; Voigt, A.; Boehm, U.; Meyerhof, W.; Behrens, M. A Subset of Mouse Colonic Goblet Cells Expresses the Bitter Taste Receptor Tas2r131. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Roland, W.S.U.; Gouka, R.J.; Gruppen, H.; Driesse, M.; Van Buren, L.; Smit, G.; Vincken, J.P. 6-Methoxyflavanones as Bitter Taste Receptor Blockers for HTAS2R39. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, A.; Mikami, Y.; Miyamoto, K.; Kamada, N.; Sato, T.; Mizuno, S.; Naganuma, M.; Teratani, T.; Aoki, R.; Fukuda, S.; et al. Intestinal Dysbiosis and Biotin Deprivation Induce Alopecia through Overgrowth of Lactobacillus Murinus in Mice. Cell Rep 2017, 20, 1513–1524. [Google Scholar] [CrossRef]
- Gao, L.; Huang, Y.; Liu, Y.; Mohamed, O.A.A.; Fan, X.; Wang, L.; Li, L.; Ma, J. Bacterial Community Structure and Potential Microbial Coexistence Mechanism Associated with Three Halophytes Adapting to the Extremely Hypersaline Environment. Microorganisms 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Janssen, S.; McDonald, D.; Gonzalez, A.; Navas-Molina, J.A.; Jiang, L.; Xu, Z.Z.; Winker, K.; Kado, D.M.; Orwoll, E.; Manary, M.; et al. Phylogenetic Placement of Exact Amplicon Sequences Improves Associations with Clinical Information. mSystems 2018, 3. [Google Scholar] [CrossRef]
- Stockinger, S.; Albers, T.; Duerr, C.U.; Ménard, S.; Pütsep, K.; Andersson, M.; Hornef, M.W. Interleukin-13-Mediated Paneth Cell Degranulation and Antimicrobial Peptide Release. J Innate Immun 2014, 6, 530–541. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhu, X.; Geng, J.; Xu, Y.; Wu, R.; Li, C.; Fan, D.; Qin, X.; Du, Y.; Tian, Y.; et al. Intestinal Tuft-2 Cells Exert Antimicrobial Immunity via Sensing Bacterial Metabolite N-Undecanoylglycine. Immunity 2022, 55, 686–700.e7. [Google Scholar] [CrossRef] [PubMed]
- Gerbe, F.; Legraverend, C.; Jay, P. The Intestinal Epithelium Tuft Cells: Specification and Function. Cellular and Molecular Life Sciences 2012, 69, 2907–2917. [Google Scholar] [CrossRef] [PubMed]
- Moerings, B.G.J.; van Bergenhenegouwen, J.; Furber, M.; Abbring, S.; Schols, H.A.; Witkamp, R.F.; Govers, C.; Mes, J.J. Dectin-1b Activation by Arabinoxylans Induces Trained Immunity in Human Monocyte-Derived Macrophages. Int J Biol Macromol 2022, 209, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Atagozli, T.; Elliott, D.E.; Ince, M.N. Helminth Lessons in Inflammatory Bowel Diseases (IBD). Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. The Intestinal Epithelial Barrier: A Therapeutic Target? Nat Rev Gastroenterol Hepatol 2017, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Hooper, L. V. Antimicrobial Defense of the Intestine. Immunity 2015, 42, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; O’Leary, C.E.; von Moltke, J.; Liang, H.E.; Ang, Q.Y.; Turnbaugh, P.J.; Radhakrishnan, S.; Pellizzon, M.; Ma, A.; Locksley, R.M. A Metabolite-Triggered Tuft Cell-ILC2 Circuit Drives Small Intestinal Remodeling. Cell 2018, 174, 271–284.e14. [Google Scholar] [CrossRef]
- Nadjsombati, M.S.; McGinty, J.W.; Lyons-Cohen, M.R.; Jaffe, J.B.; DiPeso, L.; Schneider, C.; Miller, C.N.; Pollack, J.L.; Nagana Gowda, G.A.; Fontana, M.F.; et al. Detection of Succinate by Intestinal Tuft Cells Triggers a Type 2 Innate Immune Circuit. Immunity 2018, 49, 33–41.e7. [Google Scholar] [CrossRef]
Figure 1.
Psyllium supplementation increases antimicrobial protein expression in mouse small intestine. Mice were fed the control and a 10% psyllium diet for 5 d, and jejunal samples were collected. Total RNA from these samples was subjected to RNA sequencing and qRT-PCR. Data (transcripts per million, TPM) were log 2 transformed after the addition of 1 for TPM, and the MA plot is shown (A). “Average” represents the mean values of the control and 10% psyllium groups. “Minus” represents the differences between values of the control and 10% psyllium groups. Each dot indicates a specific gene, and red and blue dots indicate genes >2-fold higher or lower in the 10% psyllium group than those in the control group, respectively. Heatmap shows the Z-scores of the genes encoding antimicrobial proteins (B). Srpp2a and Retnlb mRNA expression levels were determined using qRT-PCR (C, D). Protein expression of levels of SPRR2A and RELMβ were examined using immunoblot (E, F) and immunofluorescence (G, H) analyses. Results are shown as the mean ± SEM; n = 8. Statistical analysis was performed using unpaired Student’s t-test, *p <0.05.
Figure 1.
Psyllium supplementation increases antimicrobial protein expression in mouse small intestine. Mice were fed the control and a 10% psyllium diet for 5 d, and jejunal samples were collected. Total RNA from these samples was subjected to RNA sequencing and qRT-PCR. Data (transcripts per million, TPM) were log 2 transformed after the addition of 1 for TPM, and the MA plot is shown (A). “Average” represents the mean values of the control and 10% psyllium groups. “Minus” represents the differences between values of the control and 10% psyllium groups. Each dot indicates a specific gene, and red and blue dots indicate genes >2-fold higher or lower in the 10% psyllium group than those in the control group, respectively. Heatmap shows the Z-scores of the genes encoding antimicrobial proteins (B). Srpp2a and Retnlb mRNA expression levels were determined using qRT-PCR (C, D). Protein expression of levels of SPRR2A and RELMβ were examined using immunoblot (E, F) and immunofluorescence (G, H) analyses. Results are shown as the mean ± SEM; n = 8. Statistical analysis was performed using unpaired Student’s t-test, *p <0.05.
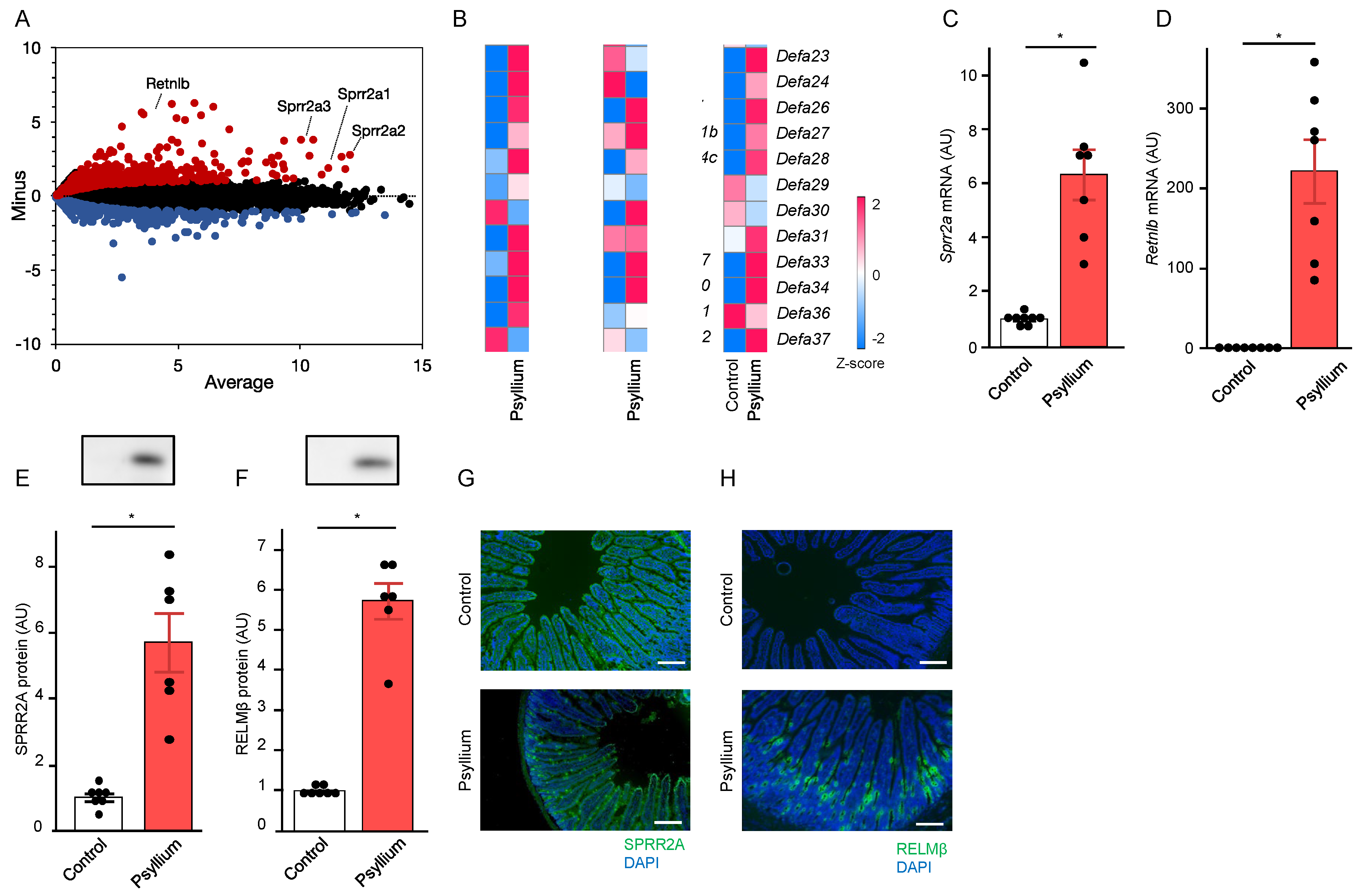
Figure 2.
Psyllium supplementation alters the microbiome profile in the mouse cecum. Mice were fed the control and a 10% psyllium diet for 5 d, and fresh cecal samples were subjected to 16S rRNA sequencing. The number of observed species (A), Chao1 index (B), and Shannon index (C) of the cecal microbiota were calculated. Principal coordinate analysis (PCoA) plots of intestinal microbiota were generated based on weighted UniFrac (D), unweighted UniFrac (E), and Bray–Curtis metrics (F) using QIIME. (G) Relative abundance of cecal microbiota at the phylum level. Genera with statistically significant differences observed between groups are shown (H, I). The genera representing relative abundances >5% (H) and 1% (I) in either group are shown. Results are shown as the mean ± SEM, n = 8. Statistical analysis was performed using unpaired Student’s t-test, *p <0.05.
Figure 2.
Psyllium supplementation alters the microbiome profile in the mouse cecum. Mice were fed the control and a 10% psyllium diet for 5 d, and fresh cecal samples were subjected to 16S rRNA sequencing. The number of observed species (A), Chao1 index (B), and Shannon index (C) of the cecal microbiota were calculated. Principal coordinate analysis (PCoA) plots of intestinal microbiota were generated based on weighted UniFrac (D), unweighted UniFrac (E), and Bray–Curtis metrics (F) using QIIME. (G) Relative abundance of cecal microbiota at the phylum level. Genera with statistically significant differences observed between groups are shown (H, I). The genera representing relative abundances >5% (H) and 1% (I) in either group are shown. Results are shown as the mean ± SEM, n = 8. Statistical analysis was performed using unpaired Student’s t-test, *p <0.05.
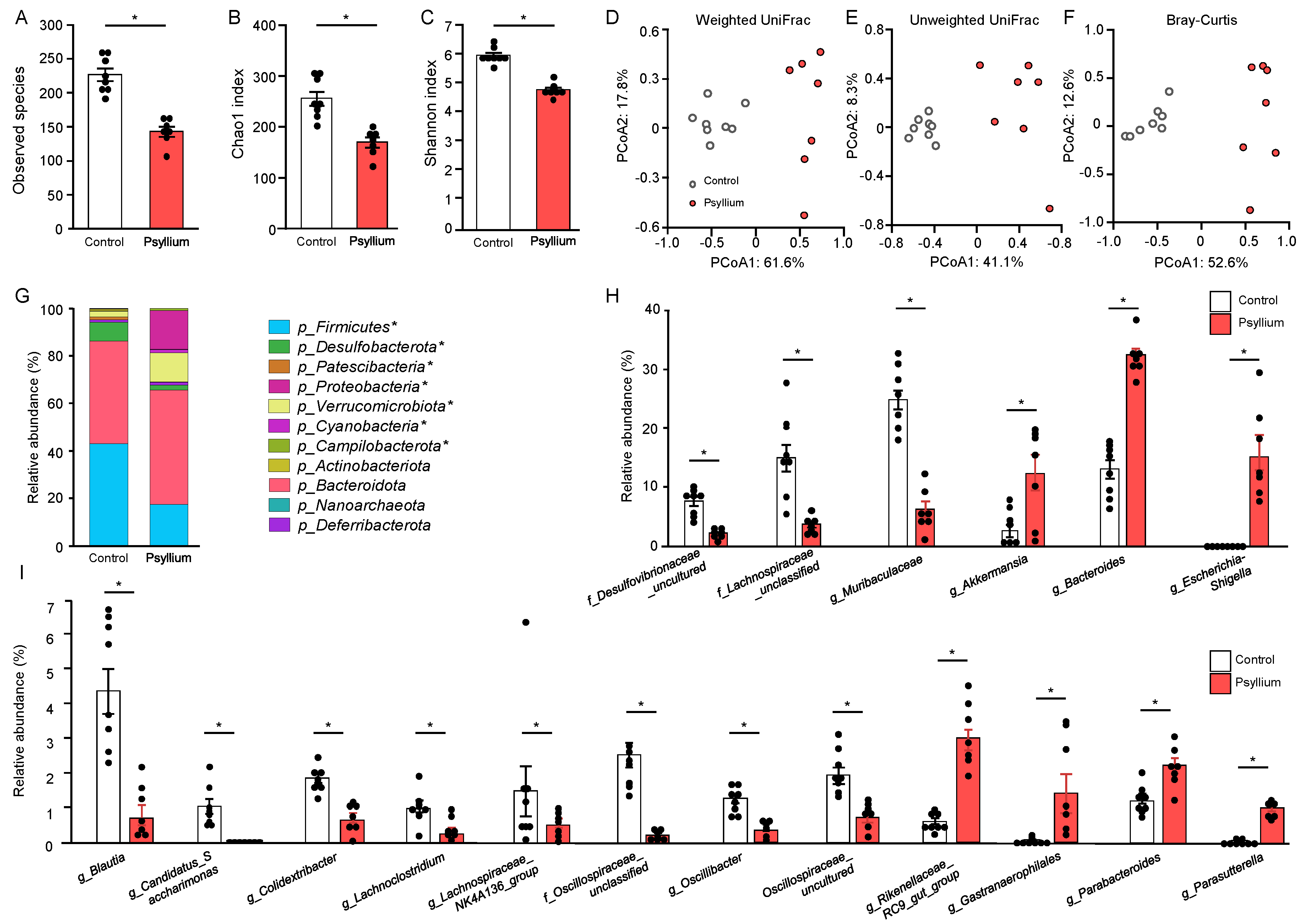
Figure 3.
Psyllium supplementation upregulates antimicrobial proteins, SPRR2A and RELMβ in mouse jejunum and ileum. Mice were fed diets with and without psyllium fiber (2.5, 5, 10%) for 5 d. Jejunal and ileal samples were subjected to qRT-PCR and immunoblotting. mRNA expression levels of Sprr2a (A, E) and Retnlb (C, G) levels were determined in the mouse jejunum (A, C) and ileum (E, G) using qRT-PCR. Protein expression levels of SPRR2A (B, F) and RELMß (D, H) were determined in the mouse jejunum (B, D) and ileum (F, H) by immunoblot analysis. Results are shown as the mean ± SEM; n = 6. Statistical analysis was performed using the Tukey–Kramer post-hoc test and Steel–Dwass test, *p <0.05.
Figure 3.
Psyllium supplementation upregulates antimicrobial proteins, SPRR2A and RELMβ in mouse jejunum and ileum. Mice were fed diets with and without psyllium fiber (2.5, 5, 10%) for 5 d. Jejunal and ileal samples were subjected to qRT-PCR and immunoblotting. mRNA expression levels of Sprr2a (A, E) and Retnlb (C, G) levels were determined in the mouse jejunum (A, C) and ileum (E, G) using qRT-PCR. Protein expression levels of SPRR2A (B, F) and RELMß (D, H) were determined in the mouse jejunum (B, D) and ileum (F, H) by immunoblot analysis. Results are shown as the mean ± SEM; n = 6. Statistical analysis was performed using the Tukey–Kramer post-hoc test and Steel–Dwass test, *p <0.05.
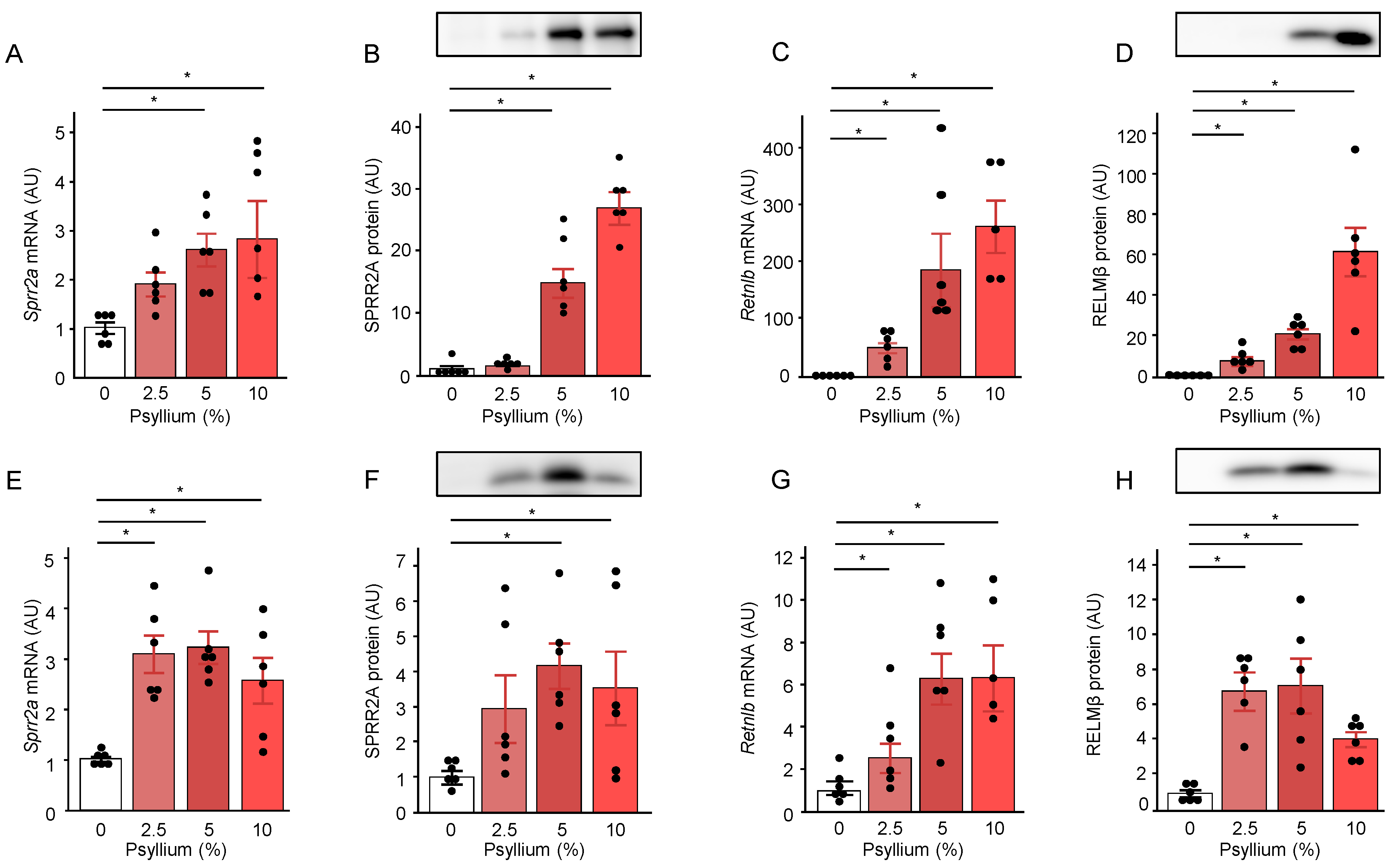
Figure 4.
Psyllium supplementation promotes tuft cell proliferation and the ILC2 immune response. Mice were fed diets with and without psyllium fiber (2.5, 5, 10%) for 5 d, and jejunal samples were subjected to qRT-PCR, immunoblot, and immunofluorescence analyses. mRNA expression levels of Pou2f3 (A), Dclk1 (B), Gfi1b (C), Sh2d6 (D), Tppp3 (E), Spib (F), Il25 (H), Il13 (I), and Il4 (J) were determined using qRT-PCR. A tuft cell marker, DCLK1, in the mouse jejunum was visualized via immunofluorescence (Scale bar: 200 µm), and the number of DCLK1+ tuft cells in a villus was quantified (G). pSTAT6 protein expression in mouse jejunum was determined using immunoblot analysis (K). Results are shown as the mean ± SEM; n = 6. Statistical analyses were performed using the Tukey–Kramer post-hoc test and unpaired Student’s t-test; *p <0.05.
Figure 4.
Psyllium supplementation promotes tuft cell proliferation and the ILC2 immune response. Mice were fed diets with and without psyllium fiber (2.5, 5, 10%) for 5 d, and jejunal samples were subjected to qRT-PCR, immunoblot, and immunofluorescence analyses. mRNA expression levels of Pou2f3 (A), Dclk1 (B), Gfi1b (C), Sh2d6 (D), Tppp3 (E), Spib (F), Il25 (H), Il13 (I), and Il4 (J) were determined using qRT-PCR. A tuft cell marker, DCLK1, in the mouse jejunum was visualized via immunofluorescence (Scale bar: 200 µm), and the number of DCLK1+ tuft cells in a villus was quantified (G). pSTAT6 protein expression in mouse jejunum was determined using immunoblot analysis (K). Results are shown as the mean ± SEM; n = 6. Statistical analyses were performed using the Tukey–Kramer post-hoc test and unpaired Student’s t-test; *p <0.05.
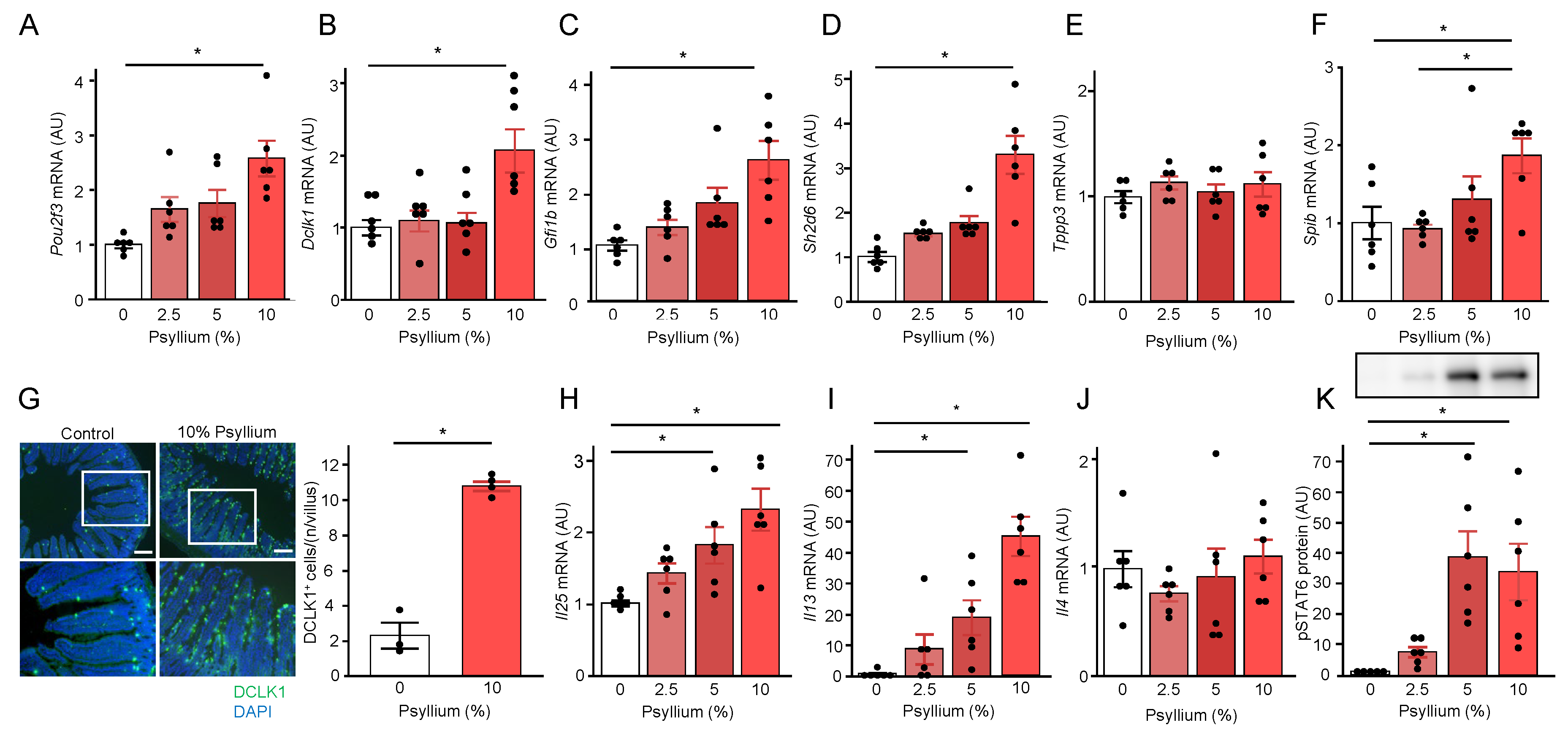
Figure 5.
ILC2 is involved in psyllium-induced antimicrobial protein expression. Mice were fed control and 5% psyllium diets for 5 d. An ILC2 inhibitor, disulfiram, was administered orally at 1.2 and 2.4 mg/g body weight 2 d before the start of the test diets. Jejunal samples were subjected to qRT-PCR, immunoblot, and immunofluorescence analyses. mRNA expression levels of Sprr2a (A), Retnlb (D), Pou2f3 (G), Il25 (H), and Il13 (J) in the mouse jejunum was determined using qRT-PCR analysis. Protein expression levels of SPRR2A (B), RELMβ (E), and pSTAT6 (K) were determined by immunoblot analysis. Expression of SPRR2A (C), RELMβ (F), DCLK1 (I), and pSTAT6 (L) was visualized using immunofluorescent analysis (Scale bar: 200 µm). Results are shown as the mean ± SEM; n = 7. Statistical analysis was performed using the Tukey–Kramer post-hoc test, *p <0.05.
Figure 5.
ILC2 is involved in psyllium-induced antimicrobial protein expression. Mice were fed control and 5% psyllium diets for 5 d. An ILC2 inhibitor, disulfiram, was administered orally at 1.2 and 2.4 mg/g body weight 2 d before the start of the test diets. Jejunal samples were subjected to qRT-PCR, immunoblot, and immunofluorescence analyses. mRNA expression levels of Sprr2a (A), Retnlb (D), Pou2f3 (G), Il25 (H), and Il13 (J) in the mouse jejunum was determined using qRT-PCR analysis. Protein expression levels of SPRR2A (B), RELMβ (E), and pSTAT6 (K) were determined by immunoblot analysis. Expression of SPRR2A (C), RELMβ (F), DCLK1 (I), and pSTAT6 (L) was visualized using immunofluorescent analysis (Scale bar: 200 µm). Results are shown as the mean ± SEM; n = 7. Statistical analysis was performed using the Tukey–Kramer post-hoc test, *p <0.05.
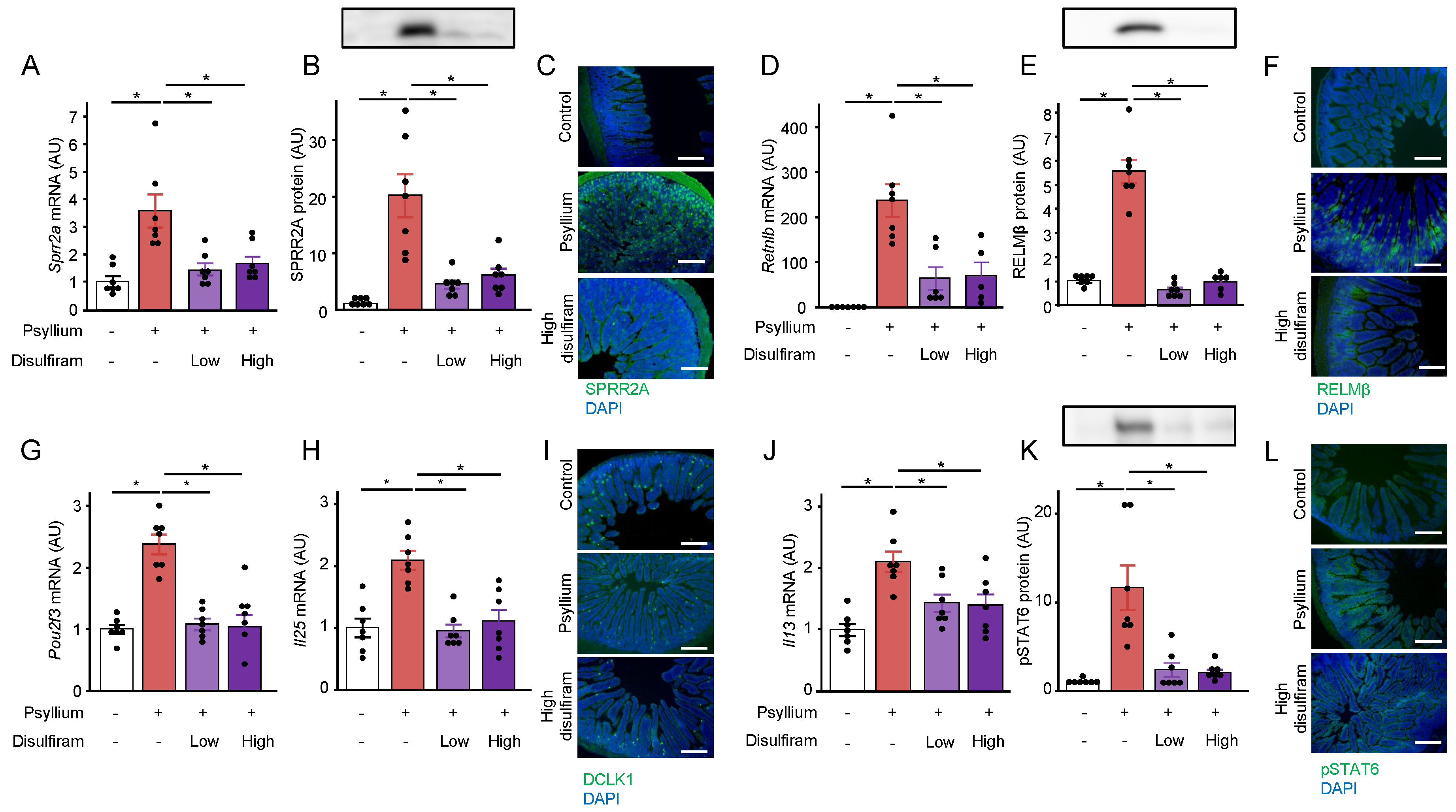
Figure 6.
Bitter taste receptors are involved in the psyllium-induced antimicrobial protein expression. Mice were fed control and 5% psyllium diets with and without bitter taste receptor blocker cocktail (0.25 and 0.5 %) for 5 d. Jejunal samples were subjected to qRT-PCR, immunoblot, and immunofluorescence analyses. mRNA expression levels of Sprr2a (A), Retnlb (D), Pou2f3 (G), Il25 (H), and Il13 (J) in the mouse jejunum was determined using qRT-PCR analysis. Protein expression levels of SPRR2A (B), RELMβ (E), and pSTAT6 (K) were determined using immunoblot analysis. Expression of SPRR2A (C), RELMβ (F), DCLK1 (I), and pSTAT6 (L) was visualized using immunofluorescent analysis (Scale bar:200 µm). Results are shown as the mean ± SEM; n = 7. Statistical analysis was performed using the Tukey–Kramer post-hoc test, *p <0.05.
Figure 6.
Bitter taste receptors are involved in the psyllium-induced antimicrobial protein expression. Mice were fed control and 5% psyllium diets with and without bitter taste receptor blocker cocktail (0.25 and 0.5 %) for 5 d. Jejunal samples were subjected to qRT-PCR, immunoblot, and immunofluorescence analyses. mRNA expression levels of Sprr2a (A), Retnlb (D), Pou2f3 (G), Il25 (H), and Il13 (J) in the mouse jejunum was determined using qRT-PCR analysis. Protein expression levels of SPRR2A (B), RELMβ (E), and pSTAT6 (K) were determined using immunoblot analysis. Expression of SPRR2A (C), RELMβ (F), DCLK1 (I), and pSTAT6 (L) was visualized using immunofluorescent analysis (Scale bar:200 µm). Results are shown as the mean ± SEM; n = 7. Statistical analysis was performed using the Tukey–Kramer post-hoc test, *p <0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



