
Submitted:
25 April 2024
Posted:
26 April 2024
You are already at the latest version
Abstract
West Nile virus (WNV) is a re-emerging flavivirus, primarily circulating among avian hosts and mosquito vectors, causing periodic outbreaks in humans and horses, often leading to neuroinvasive disease and mortality. Spain has reported several outbreaks, most notably in 2020 with seventy-seven human cases and eight fatalities. WNV has been serologically detected in horses in the Community of Madrid, but to our knowledge, it has never been reported from wild birds in this region. To estimate the seroprevalence of WNV in wild birds and horses in the Community of Madrid, 159 wild birds at a wildlife rescue center and 25 privately owned equines were sampled. Serum from thirteen birds (8.2%) and one equine (4.0%) tested positive with a WNV competitive ELISA designed for WNV antibodies detection, but sensitive to cross-reacting antibodies to other flaviviruses. VNT confirmed WNV antibodies in four bird samples (2.5%), and antibodies to undetermined flavivirus in four additional samples. One equine sample (4.0%) tested positive for WNV, although this horse previously resided in a WN-endemic area. ELISA positive birds included both migratory and resident species, juveniles and adults. Two seropositive juvenile birds suggest local flavivirus transmission within the Community of Madrid, while WNV seropositive adult birds may have been infected outside Madrid. The potential circulation of flaviviruses, including WNV, in birds in Madrid Community raises concerns, although further surveillance in mosquitos, wild birds and horses in Madrid is necessary to establish the extent of transmission, and the principal species involved.
Keywords:
Flavivirus
; West Nile virus
; Birds
; Equines
; Seroprevalence
; Central Spain
1. Introduction
West Nile virus (WNV, Orthoflavivirus nilense) is a small positive-stranded, enveloped, RNA virus [1] belonging to genus Orthoflavivirus in the viral family Flaviviridae. It is a re-emerging zoonotic virus and an arthropod-borne virus, typically transmitted by Culicine mosquitoes, as part of an enzootic cycle in which many bird species can act as reservoir, or amplifying host, and other species, including humans and horses are incidental (dead-end) hosts [2]. Typically, the virus undergoes a closed cycle between wild birds and mosquitoes, with birds potentially carrying the virus during their movements, which range from local to long distance [3].
Genus Orthoflavivirus comprises more than 70 taxa of viruses, all transmitted by arthropods, with two major clades vectored by mosquitoes or ticks, respectively, a clade with no-known arthropod vector, and a small number of tentative species. In most cases, the primary hosts of genus Orthoflavivirus are mammals or birds. Infections range from asymptomatic to severe or fatal hemorrhagic fever or neurological disease. The most significant human pathogens in this group include yellow fever virus, dengue virus, Japanese encephalitis virus (JEV), tick-borne encephalitis virus (TBEV), WNV and Zika virus. WNV belongs to the well-supported JE group, which includes 10 taxa [4,5].
WNV was discovered in Uganda in 1937 in a febrile woman [6], and was known only from Africa, Eurasia and Australia [4] prior to emerging in New York in 1999 [2]. Currently, WNV is present in numerous countries worldwide, including all countries neighboring Spain: France [7], Portugal [8] and Morocco [9], where cases have been reported since the 1960’s. In Spain, cases of WNV have been detected in humans, horses and wild birds. The first human case of WNV in Spain was retrospectively diagnosed from a 2004 sample [10]. Since 2010, human and horse WNV cases have been recovered from Andalusia and Extremadura, Catalonia, and the Valencian Community. The largest outbreak, from Andalusia and Extremadura, in 2020, led to 77 infections and eight deaths [11]. Human cases have been recovered annually since then, with six from Andalusia in 2021, five cases from Andalusia and Catalonia in 2022, and 17 cases from Andalusia, Extremadura, Catalonia, Castilla-La Mancha and the Valencian Community in 2023 [12]. The first equine outbreak in Spain was reported in 2010 and new cases have occurred annually since then [13]. Moreover, seropositive samples from asymptomatic horses were recovered in Central Spain in 2012, including Madrid, where one IgM-positive case was reported, which implied recent infection [14].
WNV has been found to infect over 392 distinct bird species [15]. The first, indirect, evidence of WNV transmission in wild birds in Spain, in 2003, was provided by a serosurvey of Common Coot (Fulica atra), a partially migratory aquatic bird, in Doñana National Park (Huelva, Southern Spain) [11]. WNV has since been detected in wild [16,17] and captive birds [18,19] in Spain by both direct and indirect surveillance techniques.
Phylogenetic studies of WNV have shown that there are at least eight distinct lineages [5,20], with different biogeographical patterns. Among them, lineages 1 (L1) and 2 (L2) exhibited the broadest geographical distribution and have been implicated in major outbreaks affecting both humans and animals. Both lineages circulate in Europe [21,22], with L2 increasingly detected in Central and Eastern Europe since 2004, in humans, birds and horses [21] and where most current human cases of WNV were caused by L2 [20].
In Spain, WNV L1 is responsible for the majority of WNV cases in humans, horses, birds, and mosquitoes [23]. It is the only lineage detected in the southwest of Spain [24], particularly in Andalusia, with additional cases reported in Extremadura. Rare cases have been notified in Madrid and neighboring regions, such as Ávila (Castile and León), Ciudad Real and Toledo (Castilla-La Mancha). A secondary, smaller focus of WNV activity has been described in the north-east Spain, in Catalonia, and more recently, in the neighboring Valencian Community [12]. In contrast to the rest of Spain, where L1 has been detected, L2 has been found in Catalonia, specifically in birds since 2017 [25,26], and in mosquitoes in 2021 [27]. WNV disease cases in horses and humans, lineage unknown, have been detected in the northeast since 2022 [28]. The emergence of L2 in the northeast of Spain aligns with a westward dispersion from central Europe [26].
Other phylogenetically related Flavivirus taxa have been detected in Spain, showing serological cross-reactivity with WNV [29]. These taxa include another potential zoonosis, Usutu virus (USUV), a JEV serogroup virus which has an overlapping distribution with WNV in south-west and north-east Spain, and its circulation has also been detected in birds and mosquitoes [30,31,32], and Bagaza virus (BAGV), a virus in the Ntaya virus serocomplex, known from gamebirds in south-west Spain [33]. Circulation in Spain of other more distantly related flaviviruses, such as Meaban virus [34], TBEV, and Spanish sheep/goat encephalitis virus, have been described, but expected cross-reactivity with common WNV serological tests is lower as they belong to distant serocomplexes [32,35].
Although WNV has shown to be spreading through the south, and north-east of the country, to the best of our knowledge the only evidence of WNV circulation in the Community of Madrid was the detection of specific antibodies in horses in 2012 [14]. The aim of the present study was to verify the possible circulation of WNV in wild birds and equines in Madrid between 2020 and 2022. Furthermore, several factors that could provide insights into the extent of virus exposure among these animals in this area were examined.
2. Materials and Methods
This study consisted of a retrospective study, conducted on wild birds, and a prospective study on equines, in order to verify the possible circulation of WNV in these animal species in the Community of Madrid. Serum samples were used in all cases. Institutional ethics clearance was obtained to conduct this study (Decision 2022_1/117).
2.1. Samples
2.1.1. Birds
Serum samples were corrected from 159 birds in different areas of the Community of Madrid by GREFA wildlife rescue center (Grupo de Rehabilitación de la Fauna Autóctona y su Hábitat) between August 2020 and August 2021. The field sampling areas were divided into the following regions within the Madrid Community: Metropolitan area and Henares corridor, Upper Manzanares Basin, Sierra Oeste, Henares Basin, Jarama Basin, Upper Guadarrama Basin, Guadarrama Basin, Sierra Norte, Comarca Sur and Comarca de Las Vegas (Figure 1). Blood was extracted from the cubital vein or the medial metatarsal vein using 23G needles. Serum was obtained by centrifugation of the samples, and then stored at -80ºC for further analysis.
Different variables were collected from each bird, including species, migratory status, gender, age, size, area of origin, cause of admission to the center, subsequent situation/evolution, and presence of pathology or signs of disease at the time of admission.
Seventeen bird species were sampled, predominantly white storks (Ciconia ciconia) with 95 samples, followed by 17 red kites (Milvus milvus), 10 Bonelli's eagles (Aquila fasciata), nine griffon vultures (Gyps fulvus), five black kites (Milvus migrans), four peregrine falcons (Falco peregrinus), four Eurasian eagle owls (Bubo bubo), three Spanish imperial eagles (Aquila adalberti), three black storks (Ciconia nigra), two golden eagles (Aquila chrysaetos), one great cormorant (Phalacrocorax carbo), one grey heron (Ardea cinerea), one lesser black-backed gull (Larus fuscus), one European honey buzzard (Pernis apivorus), one short-toed snake eagle (Circaetus gallicus), one great bustard (Otis tarda) and one common kestrel (Falco tinnunculus).
Birds were routinely sampled by GREFA personnel as part of standard activities for a variety of purposes, including relocation of nestlings, transfers, breeding programs for captive-born animals, population monitoring and treating cases of trauma, malnutrition, natural disease, poisoning, or ingestion of foreign objects.
Birds without sexual dimorphism were either sexed by DNA in an external laboratory if necessary for a specific project or left as "undetermined" if the sex was not relevant in the clinical management or destiny of the animal. The cost and invasiveness of additional tests were taken into consideration when deciding whether to determine the sex.
Birds were classified as juveniles (including the subcategories nestling, fledgling, juvenile) or adults based on their feather type. Birds without down, neossoptilus, pre-down, or pre-feather at the time of sample collection were considered adults [36]. Additionally, physical-anatomical characteristics were used to determine age [37].
The size of the bird was determined based on pre-established criteria, considering the bird's wingspan, and categorized as either large or very large [38].
2.1.2. Equines
Twenty-five serum samples were collected from clinically healthy equids, consisting of 23 horses and 2 donkeys, from different equestrian centers and private properties in the Community of Madrid, between January and April 2022 (Figure 1). Sample inclusion criteria were equids that: (1) did not present clinical symptoms; (2) resided in the Community of Madrid and had not traveled outside the Community at least three months prior to sample collection and (3) were not vaccinated at any time against WNV. In all cases, informed consent was obtained and signed by the owners of the equids included in the study.
A minimum of 2 ml of blood was obtained in tubes without anticoagulant and subsequently centrifuged to obtain the serum and transported to the Biological Research Unit of the UAX Veterinary Hospital (UIB-UAX). Serum samples were stored at -20ºC until analyzed.
Different data were collected on the origin, sex, age, and species or breed of the animals.
2.2. Serological Testing
Serological testing for the detection of WNV antibodies was conducted using the commercially available competition Enzyme Linked ImmunoSorbent-Assay kit (Ingezim WNV Compac enzymatic assay kit, Ingenasa, Spain). Each ELISA plate used for testing included duplicate samples and was read at a wavelength of 450 nm. Results were classified as positive when the inhibition percentage was equal to or greater than 40%. Conversely, results were classified as negative when the inhibition percentage was equal to or less than 30%. Results falling within these two intervals were considered doubtful. Positive cELISA samples from horses were forwarded to the regional reference laboratory (Laboratorio Central de Veterinaria, Algete) for IgM detection (IDVET, France and Ingenasa, Spain).
To assess specificity of antibodies present in the samples, the virus microneutralization test (VNT) was performed on cELISA positive samples containing sufficient serum (12 birds and one horse) and doubtful samples (15 birds and one horse), at the Animal Health Research Centre (CISA-INIA), CSIC, following Llorente, et al. 29. Due to the possibility of cross-reactivity with antigenically related flaviviruses documented to be present in Spain, samples were tested in parallel against WNV (strain E101), USUV (SAAR-1776), and BAGV (Spain/RLP-Hcc1/2010) (GenBank accession nos. AF260968, AY453412 and KR108244, respectively). Samples yielding positive neutralization (complete absence of cytopathic effects at dilutions equal or higher than 1:10) were scored as positive. Presence of specific antibodies for a virus was assigned when titers obtained for that virus were at least 4-fold higher than the others. If the titer differences did not reach this threshold the sample was scored as “undetermined flavivirus.
2.3. Statistical Analysis
A descriptive statistical study was conducted, considering all variables collected from each animal. The qualitative variables are presented with their frequency distribution. Positive WNV antibodies in birds, identified using cELISA, were calculated as a percentage, and their respective 95% confidence intervals (95% CI) were determined using the exact binomial method. The statistical analyses were performed using Stata version 13.
3. Results
3.1. Birds
In the birds examined from Madrid, the percentage of serum samples yielding positive results by WNV cELISA was 8.2% (13/159; 95% CI: 4.4%-13.6%). Fifteen avian samples that produced weak inhibition, and were considered doubtful, were submitted for VNT testing alongside the cELISA positive samples. Of the 27 bird serum samples analyzed by VNT, four that were positive for cELISA exhibited specific antibodies for WNV (14.8% of the samples challenged by VNT), while four (14.8%), three of them positive and one considered doubtful by cELISA, were categorized as undetermined flavivirus (Table 1). None of the samples were classified as USUV or BAGV positive (presence of specific antibodies). cELISA and VNT titers of the birds that tested positive by VNT are included in Table 2. Municipalities with positive results for WNV are highlighted in Figure 2.
The four WNV-infected birds in Madrid included Bonelli's eagle (Aquila fasciata), black kite (Milvus migrans), European honey buzzard (Pernis apivorus), and short-toed snake eagle (Circaetus gallicus), all belonging to family Accipitridae (Table 3). These birds were all sampled in different regions of the Madrid Community, namely the Sierra Oeste, Metropolitan area, Sierra Norte, and Guadarrama Basin, respectively. Except for Bonelli's eagle, all were migratory species. All four WNV seropositive birds were at least 12 months old, though the Milvus migrans had not yet attained definitive adult plumage. The primary reason for admission to the rescue center for the majority of the individuals tested was trauma, except for Bonelli’s eagle, which was captured, marked, and released in the field without requiring passage through the facilities of the rescue center. The four birds exposed to undetermined flaviviruses consisted of three species: one white stork (Ciconia ciconia), family Ciconiidae, two Bonelli's eagles (Aquila fasciata), and one griffon vulture (Gyps fulvus), both family Accipitridae. Two of these flavivirus-positive birds were juveniles tested in Madrid on 28th September 2020 and 3rd June 2021.
To the best of our knowledge, WNV exposure has been detected in at least 67 avian species in Spain so far, including all the species that tested positive in this study (Table S1).
3.2. Equids
One of 25 equid samples collected in Madrid tested positive for WNV antibodies using cELISA screening, showing a prevalence of 4.0% (95% CI: 0.1%-20.3%). This horse also tested positive by VNT for WNV. However, IgM detection by the reference laboratory yielded negative results for acute infection. This was a male Anglo-Arab horse whose sample was obtained in the Guadarrama Sierra. A second sample which produced weak inhibition, and was considered doubtful for cELISA, was VNT negative.
4. Discussion
The main objective of this study was to determine the possible circulation of WNV in birds and horses in the Community of Madrid by detecting antibodies against this virus. Additionally, we aimed to analyze variables that could help us understand the exposure of these animals to the virus in central Spain. 8.2% of the birds and 4% of the equines tested yielded positive results for a cELISA test, which is noted for its high sensitivity, but is cross-reactive with other related flaviviruses [29]. Seroprevalence findings in birds from this study fall within the range of previous research in Spain using the cELISA tests, indicating seroprevalence rates ranging from 1.96% to 31.25% [39]. Serological results in wild birds from around the world using various cELISA protocols also reveal a broad range of seroprevalence rates: 1.3% in Cyprus [40], 5.5% in Senegal [41], 6.6% in the United States [42], 11.9% in Bangladesh [43], 13.3 % in Poland [44], 27.33% in Iran [45], and 32.1% in Romania [46].
The WNV seroprevalence of 2.5% recovered by VNT (with a further 2.5% positive for undetermined flavivirus) falls in the lower bound of the range observed for VNT results in birds in Spain of other VNT results in Spain, ranging from 0.66% of the birds originally screened by cELISA to 18.23% [39]. Furthermore, several wild bird studies in Spain show seroprevalence values of 4.1% [30], 4.2% [39], 7.4% [47] and 9.4% [17] for undetermined flaviviruses using VNT. This reflects the challenge of the specific determination of flavivirus antibodies caused by cross-reactions between different assayed viruses. Additionally, we cannot rule out the presence of additional flavivirus taxa.
Similarly, a wide range of seroprevalence values have been reported from equines by VNT, spanning 2.47% in Lebanon [48], 9.8% in Spain [32], 31% in Morocco [49], and 35.7% in Nigeria [50].
The birds that showed specific WNV antibodies by VNT were all adults. Three of them (short-toed snake eagle, European honey buzzard, black kite) were long distance migrants, and may have been infected in Spain, or equally plausibly, in the sub-Saharan wintering areas to which they migrate. One individual, a Bonelli’s eagle, belongs to a species resident in Spain. However, daily movements of this species are large, especially for juveniles, for instance, one juvenile male tagged with a GPS was recorded to travel 350 km in 48 hours [51]. This individual was first captured as an adult in Madrid Province in 2019, and tagged with a GPS locator, and was then recaptured twice in 2020. This individual did not leave Madrid after it was tagged. Unfortunately, no sample was collected at the first time of capture, so it was no possible to determine if the individual was infected in Madrid. In this case, WNV transmission may have occurred in Madrid or in contiguous areas with very similar environmental characteristics to this region. Two hatch year birds were seropositive for undetermined flavivirus, and WNV infection was neither ruled out, nor confirmed, for these two individuals. One was a nestling white stork, removed to GREFA as part of a nest relocation scheme, prior to first flight. This seropositive bird must have been infected in its nest in Alcobendas, a conurbation with nearly 120,000 inhabitants [52]. The second bird seropositive for an undetermined flavivirus, a griffon vulture, was captured as a fledgling, on the 28th of September. This individual could barely fly and could not have travelled beyond the confines of the Madrid Province. It must have been infected in Madrid. The two adult birds, one resident and one migrant, seropositive for an undetermined flavivirus are not informative in terms of transmission.
The presence of maternal antibodies to West Nile virus has been reported in wild birds typically persisting for less than 30 days [53,54,55]. One study found that maternal antibodies usually persist for 28 days post hatching in domestic chickens, but exceptionally lasted for 56 days [56]. The white stork sampled in early June 2021 in Madrid was judged to be considerably older than 30 days due to its large size and weight, and thus is unlikely to possess maternal antibodies. Griffon vultures have a very long period before fledging from their nest of over 120-140 days [57]. This individual was sampled in late September 2020 and is likely to have been at least 150 days-old, which is too long for the continued maintenance of maternal antibodies. Similarly, the presence of seroconversion to flavivirus in these birds that have not left Madrid and are too old to maintain maternal antibodies implies circulation within Madrid in 2020 and 2021.
All the bird species seropositive for WNV or undetermined flavivirus in this study are classified as “Least Concern” by the International Union for the Conservation of Nature (IUCN). This means none of these species are evaluated as being at risk of extinction at a global level. However, the global population of Bonelli’s eagle is less than 50,000 individuals and is considered to be declining [58]. In Spain, the species is classified as “Vulnerable”. All four adult Bonelli’s eagles tested were seropositive by cELISA, and one and two individuals, respectively, were positive for specific WNV or undetermined flavivirus antibodies. This species may be especially at risk from flavivirus infection. However, it is unclear if West Nile or other flavivirus are mortal to this species, or impact it in other ways, for instance, in breeding success. In addition, European honey buzzards are classified as “Near-threatened” in Spain, with an estimated population of about 1850 individuals, and a low population in Madrid [59]. Only one individual of this species was tested, so data is sparse for drawing any conclusions.
All seropositive birds in this study, and indeed all birds tested, were classified as large or very large birds. It has been observed that prevalence of antibodies is directly related to bird size, with higher prevalence found in larger birds [60]. Since we did not test small birds, we cannot directly comment on this observation. It is probable that larger birds are highly attractive to vectors [60].
All the four birds with specific antibodies to WNV were raptors from the family Accipitridae, supporting the previous observation of the high susceptibility to the infection by the virus in raptors, indicating their important role as reservoirs and spreaders of the virus [61].
Regarding the seropositive result found in a horse in this study, the confirmation from the reference laboratory indicated a negative IgM result. IgM antibodies, indicative of recent infection, typically persist in horses for approximately 1-2 months [62], while IgG antibodies can remain detectable for over a year [13,63]. Considering the sampling period of this horse (January 2022), it is plausible that the horse might have been infected during the preceding months, coinciding with the activity of the transmitting vector (from May to October) [64]. However, this animal resided in a WN-endemic area of South of Spain two years before its relocation to the Madrid Community, therefore we could not discard that it was an imported case in Madrid Community, especially given the absence of outbreaks reported in horses in the region.
Most WNV cases from humans, horses, and birds in Spain were reported from the southwest, chiefly from Andalusia and Extremadura, with rare cases notified from Madrid neighboring regions. A second, smaller, focus of activity has been described more recently in the northeast of Spain, in Catalonia, later in the neighboring Community of Valencia [12]. Short WNV sequences were isolated from two golden eagles and a Bonelli´s eagle at a rehabilitation and breeding center in Castilla-La-Mancha, south-central Spain, 2007 [16]. WNV cases has been detected in Toledo, Ciudad Real and Avila [14,65]. These three locations are 80-180 km from Madrid, and the environmental conditions are similar to those found in Madrid Province. There is thus no obvious barrier to the transmission of WNV in Madrid Province.
This work provides the first evidence for the transmission of flavivirus in Class Aves in Madrid Province in 2020 and 2021. Transmission of this undetermined flavivirus must have occurred by early June, the date of first detection in a hatch-year bird in the Alcobendas municipality. Furthermore, this study underlines that wild birds have been exposed to WNV within travelling distance of Madrid, probably in areas which share similar environmental characteristics. This is concerning, since it has been observed that WNV frequently re-occurs in the same area in subsequent years after a first outbreak is reported [26]. The conditions for WNV maintenance and circulation, known to exist in certain areas of the Iberian Peninsula, especially in the south-west, seem to be increasing, consistent with the observation that climate change may favor the establishment of competent mosquito populations in a broader area [13]. Increased surveillance of wild birds, sentinel horses and insect vectors in Madrid Province, and throughout Spain is of utmost importance to better characterize the virus, its behavior and spread, in order to implement appropriate measures to protect public health in Madrid, the third largest conurbation in western Europe. Moreover, characterizing the West Nile virus lineage or lineages potentially present in the Community of Madrid would be essential for risk planning, and is especially intriguing as both lineages 1 and 2 are known from Spain..
Author Contributions
Conceptualization, T.A.S., F.G.G., I.L.M., N.B.; methodology, H.A.C.C., I.L.M, F.L., M.A.J.-C, M.M.B; software, G.O.-D.; validation, F. L., M.A.J.-C., T.A.S.; formal analysis, G.O.-D.; investigation, H.A.C.C., I.L.M., F. L., .M.A.J.-C., M.M.B., T.A.S.; resources, T.A.S.; data curation, G.O.-D.; writing—original draft preparation, R.A.J.W., T.A.S.; writing—review and editing, R.A.J.W., I.L.M, F.L., M.A.J.-C, N.B, M.M.B., T.A.S.; visualization, R.A.J.W., T.A.S.; supervision, T.A.S., F.G.G., I.L.M.; project administration, T.A.S.; funding acquisition, T.A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was part of a project funded during the XII call for Research Projects Santander-UAX (Project code: 1.012.017). It is important to note that the funder was not involved in the study design, data collection, analysis, interpretation, writing of the article, or the decision to submit it for publication. This work was also partially funded by AEI project PID2020-116768RR-C21.
Institutional Review Board Statement
This study was approved by the Ethics Committee of Alfonso X El Sabio University, Spain (Protocol code: 2022_1/117) on January 15, 2022.
Informed Consent Statement
Written informed consent was obtained from the owners of the equines prior to blood collection.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
The authors herein express their gratitude to the horse and donkey owners for their outstanding cooperation and support throughout this study and the veterinarians participating in blood collection and sample handling. Moreover, we would like to thank the technicians from the Biomedical Research Unit of Alfonso X El Sabio University (UIB-UAX) for their support in serological analyses, and the Central Reference Laboratory of Algete for the IgM testing for confirmation of horse’ results. We also thank GREFA volunteers for their commitment and dedication in attending to GREFA animals.
Conflicts of Interest
The authors declare no conflict of interest.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: WNV detection records recovered from class Aves, using PCR and sequencing (PCR), serum neutralization (VNT) or competitive Enzyme Linked ImmunoSorbent-Assay (cELISA) in the following autonomous communities: Andalusia (A), Canary Islands (CI), Castilla La Mancha (CLM), Catalonia (C) and Extremadura (E). References in Supplementary Materials are cited in main text [11,16,17,18,19,26,30,32,39,47], and in Supplementary Materials [66,67,68,69,70,71,72,73,74,75].
References
- Brinton, M.A. Replication Cycle and Molecular Biology of the West Nile Virus. Viruses 2014, 6, 13–53, . [CrossRef]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental Infection of North American Birds with the New York 1999 Strain of West Nile Virus. Emerg. Infect. Dis. 2003, 9, 311–322, . [CrossRef]
- Malkinson, M.; Banet, C.; Weisman, Y.; Pokamunski, S.; King, R.; Drouet, M.T.; Deubel, V. Introduction of West Nile virus in the Middle East by migrating white storks. Emerg Infect Dis 2002, 8, 392-397.
- Scherret, J.H.; Poidinger, M.; Mackenzie, J.S.; Broom, A.K.; Deubel, V.; Lipkin, W.I.; Briese, T.; Gould, E.A.; Hall, R.A. The relationships between West Nile and Kunjin viruses. Emerg Infect Dis 2001, 7, 697-705.
- Fall, G.; Di Paola, N.; Faye, M.; Dia, M.; de Melo Freire, C.C.; Loucoubar, C.; de Andrade Zanotto, P.M.; Faye, O.; Sall, A.A. Biological and phylogenetic characteristics of West African lineages of West Nile virus. PLOS Negl. Trop. Dis. 2017, 11, e0006078, doi:10.1371/journal.pntd.0006078.
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A Neurotropic Virus Isolated from the Blood of a Native of Uganda 1. Am. J. Trop. Med. Hyg. 1940, s1-20, 471–492, . [CrossRef]
- Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J.; Panthier, R. Epidémiologie du virus West Nile: étude d’un foyer en Camargue. IV. La méningo-encéphalomyélite du cheval. Ann Inst Pasteur (Paris) 1970, 118, 239-247.
- Filipe, A.R.; De Andrade, H.R. Arboviruses in the Iberian Peninsula.. 1990, 34, 582–91.
- El Harrack, M.; Le Guenno, B.; Gounon, P. Isolement du virus West Nile au Maroc. Virologie 1997, 1, 248-249.
- Kaptoul, D.; Viladrich, P.F.; Domingo, C.; Niubó, J.; Martínez-Yélamos, S.; De Ory, F.; Tenorio, A. West Nile virus in Spain: Report of the first diagnosed case (in Spain) in a human with aseptic meningitis. Scand. J. Infect. Dis. 2007, 39, 70–71, . [CrossRef]
- Figuerola, J.; Soriguer, R.; Rojo, G.; Tejedor, C.G.; Jimenez-Clavero, M.A. Seroconversion in Wild Birds and Local Circulation of West Nile Virus, Spain. Emerg. Infect. Dis. 2007, 13, 1915–1917, . [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). West Nile virus - infections among humans and outbreaks among equids and/or birds, 13 December 2023. Available online: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/disease-data-ecdc (accessed on 22nd January 2024).
- Magallanes, S.; Llorente, F.; Ruiz-López, M.J.; la Puente, J.M.-D.; Soriguer, R.; Calderon, J.; Jímenez-Clavero, M..; Aguilera-Sepúlveda, P.; Figuerola, J. Long-term serological surveillance for West Nile and Usutu virus in horses in south-West Spain. One Heal. 2023, 17, 100578, . [CrossRef]
- Abad-Cobo, A.; Llorente, F.; Barbero, M.d.C.; Cruz-López, F.; Forés, P.; Jiménez-Clavero, M.. Serosurvey Reveals Exposure to West Nile Virus in Asymptomatic Horse Populations in Central Spain Prior to Recent Disease Foci. Transbound. Emerg. Dis. 2017, 64, 1387–1392, . [CrossRef]
- Williams, R.A.J.; Sánchez-Llatas, C.J.; Doménech, A.; Madrid, R.; Fandiño, S.; Cea-Callejo, P.; Gomez-Lucia, E.; Benítez, L. Emerging and Novel Viruses in Passerine Birds. Microorganisms 2023, 11, 2355, . [CrossRef]
- Jiménez-Clavero, M.A.; Sotelo, E.; Fernandez-Pinero, J.; Llorente, F.; Blanco, J.M.; Rodriguez-Ramos, J.; Perez-Ramirez, E.; Höfle, U. West Nile Virus in Golden Eagles, Spain, 2007. Emerg. Infect. Dis. 2008, 14, 1489–1491, . [CrossRef]
- Marzal, A.; Ferraguti, M.; Muriel, J.; Magallanes, S.; Ortiz, J.; García-Longoria, L.; Bravo-Barriga, D.; Guerrero-Carvajal, F.; Aguilera-Sepúlveda, P.; Llorente, F.; et al. Circulation of zoonotic flaviviruses in wild passerine birds in Western Spain. Veter- Microbiol. 2022, 268, 109399, . [CrossRef]
- López, G.; Jiménez-Clavero, M..; Vázquez, A.; Soriguer, R.; Gómez-Tejedor, C.; Tenorio, A.; Figuerola, J. Incidence of West Nile Virus in Birds Arriving in Wildlife Rehabilitation Centers in Southern Spain. Vector-Borne Zoonotic Dis. 2011, 11, 285–290, . [CrossRef]
- Llorente, F.; Pérez-Ramírez, E.; Fernández-Pinero, J.; Soriguer, R.; Figuerola, J.; Jiménez-Clavero, M.. Flaviviruses in Game Birds, Southern Spain, 2011–2012. Emerg. Infect. Dis. 2013, 19, 1023–1025, . [CrossRef]
- Zehender, G.; Veo, C.; Ebranati, E.; Carta, V.; Rovida, F.; Percivalle, E.; Moreno, A.; Lelli, D.; Calzolari, M.; Lavazza, A.; et al. Reconstructing the recent West Nile virus lineage 2 epidemic in Europe and Italy using discrete and continuous phylogeography. PLOS ONE 2017, 12, e0179679–e0179679, . [CrossRef]
- Lu, L.; Zhang, F.; Munnink, B.B.O.; Munger, E.; Sikkema, R.S.; Pappa, S.; Tsioka, K.; Sinigaglia, A.; Molin, E.D.; Shih, B.B.; et al. West Nile virus spread in Europe: Phylogeographic pattern analysis and key drivers. PLOS Pathog. 2024, 20, e1011880, . [CrossRef]
- Sotelo, E.; Fernández-Pinero, J.; Llorente, F.; Vázquez, A.; Moreno, A.; Agüero, M.; Cordioli, P.; Tenorio, A.; Jiménez-Clavero, M.Á. Phylogenetic relationships of Western Mediterranean West Nile virus strains (1996–2010) using full-length genome sequences: single or multiple introductions? J Gen Virol 2011, 92, 2512-2522.
- Rodríguez-Alarcón, L.G.S.M.; Fernández-Martínez, B.; Moros, M.J.S.; Vázquez, A.; Pachés, P.J.; Villacieros, E.G.; Martín, M.B.G.; Borras, J.F.; Lorusso, N.; Aceitero, J.M.R.; et al. Unprecedented increase of West Nile virus neuroinvasive disease, Spain, summer 2020. Eurosurveillance 2021, 26, 2002010, . [CrossRef]
- Ruiz-López, M.J.; Aguilera-Sepúlveda, P.; Cebrián-Camisón, S.; Figuerola, J.; Magallanes, S.; Varona, S.; Cuesta, I.; Cano-Gómez, C.; Sánchez-Mora, P.; Camacho, J.; et al. Re-Emergence of a West Nile Virus (WNV) Variant in South Spain with Rapid Spread Capacity. Viruses 2023, 15, 2372, . [CrossRef]
- Busquets, N.; Laranjo-González, M.; Soler, M.; Nicolás, O.; Rivas, R.; Talavera, S.; Villalba, R.; Miguel, E.S.; Torner, N.; Aranda, C.; et al. Detection of West Nile virus lineage 2 in North-Eastern Spain (Catalonia). Transbound. Emerg. Dis. 2019, 66, 617–621, . [CrossRef]
- Aguilera-Sepúlveda, P.; Napp, S.; Llorente, F.; Solano-Manrique, C.; Molina-López, R.; Obón, E.; Solé, A.; Jiménez-Clavero, M..; Fernández-Pinero, J.; Busquets, N. West Nile Virus Lineage 2 Spreads Westwards in Europe and Overwinters in North-Eastern Spain (2017–2020). Viruses 2022, 14, 569, . [CrossRef]
- Ministerio de Sanidad. Dirección General de Salud Pública. Centro de Coordinación de Alertas y Emergencias Sanitarias. Meningoencefalitis por el virus del Nilo occidental en España 2021. Available online: https://www.sanidad.gob.es/profesionales/saludPublica/ccayes/alertasActual/docs/20210902_ERR_Nilo_Occidental.pdf (accessed on 5th April 2024).
- European Centre for Disease Prevention and Control (ECDC). Historical data by year - West Nile virus seasonal surveillance. 2023. Available online: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/historical (accessed on 22nd January 2024).
- Llorente, F.; García-Irazábal, A.; Pérez-Ramírez, E.; Cano-Gómez, C.; Sarasa, M.; Vázquez, A.; Jiménez-Clavero, M.Á. Influence of flavivirus co-circulation in serological diagnostics and surveillance: a model of study using West Nile, Usutu and Bagaza viruses. Transbound Emerg Dis 2019, 66, 2100-2106.
- Jurado-Tarifa, E.; Napp, S.; Lecollinet, S.; Arenas, A.; Beck, C.; Cerdà-Cuéllar, M.; Fernández-Morente, M.; García-Bocanegra, I. Monitoring of West Nile virus, Usutu virus and Meaban virus in waterfowl used as decoys and wild raptors in southern Spain. Comp. Immunol. Microbiol. Infect. Dis. 2016, 49, 58–64, . [CrossRef]
- Höfle, U.; Gamino, V.; de Mera, I.G.F.; Mangold, A.J.; Ortíz, J.-A.; de la Fuente, J. Usutu Virus in Migratory Song Thrushes, Spain. Emerg. Infect. Dis. 2013, 19, 1173–1175, . [CrossRef]
- Napp, S.; Llorente, F.; Beck, C.; Jose-Cunilleras, E.; Soler, M.; Pailler-García, L.; Amaral, R.; Aguilera-Sepúlveda, P.; Pifarré, M.; Molina-López, R.; et al. Widespread Circulation of Flaviviruses in Horses and Birds in Northeastern Spain (Catalonia) between 2010 and 2019. Viruses 2021, 13, 2404, . [CrossRef]
- Agüero, M.; Fernandez-Pinero, J.; Buitrago, D.; Sanchez, A.; Elizalde, M.; Miguel, E.S.; Villalba, R.; Llorente, F.; Jimenez-Clavero, M.A. Bagaza virus in partridges and pheasants, Spain, 2010. Emerg Infect Dis 2011, 17, 1498-1501.
- Arnal, A.; Gómez-Díaz, E.; Cerdà-Cuéllar, M.; Lecollinet, S.; Pearce-Duvet, J.; Busquets, N.; García-Bocanegra, I.; Pagès, N.; Vittecoq, M.; Hammouda, A.; et al. Circulation of a Meaban-Like Virus in Yellow-Legged Gulls and Seabird Ticks in the Western Mediterranean Basin. PLOS ONE 2014, 9, e89601, . [CrossRef]
- Mansfield, K.L.; de la Fuente, J.; Ayllón, N.; de Mera, I.G.F.; Johnson, N.; Gortázar, C.; Alberdi, P.; Marín, J.F.G.; Morales, A.B.; Fooks, A.R.; et al. Identification and characterization of a novel tick-borne flavivirus subtype in goats (Capra hircus) in Spain. J. Gen. Virol. 2015, 96, 1676–1681, . [CrossRef]
- López-Albors, O.; Gil, F.; Vázquez, J.M.; Latorre, R.; Ramíre-Zarzosa, G.; Orenes, M.; Moreno, F. Revisión: nomenclatura e iconografía de las partes de la pluma y sus diferentes tipos. Anal Vet Murcia 1999, 15, 3-16.
- Svensson, L.; Mullarney, K. Guía de Aves de España y Europa; Ediciones Omega, 2023.
- Gómez-Catasús, J.; Carrascal, L.M.; Moraleda, V.; Colsa, J.; Garcés, F.; Schuster, C. Factors Affecting Differential Underestimates of Bird Collision Fatalities at Electric Lines: A Case Study in the Canary Islands. Ardeola 2020, 68, 71–94, . [CrossRef]
- Bravo-Barriga, D.; Aguilera-Sepúlveda, P.; Guerrero-Carvajal, F.; Llorente, F.; Reina, D.; Pérez-Martín, J.E.; Jiménez-Clavero, M..; Frontera, E. West Nile and Usutu virus infections in wild birds admitted to rehabilitation centres in Extremadura, western Spain, 2017–2019. Veter- Microbiol. 2021, 255, 109020, . [CrossRef]
- Pallari, C.T.; Efstathiou, A.; Moysi, M.; Papanikolas, N.; Christodoulou, V.; Mazeris, A.; Koliou, M.; Kirschel, A.N. Evidence of West Nile virus seropositivity in wild birds on the island of Cyprus. Comp. Immunol. Microbiol. Infect. Dis. 2021, 74, 101592, . [CrossRef]
- Chevalier, V.; Reynaud, P.; Lefrançois, T.; Durand, B.; Baillon, F.; Balança, G.; Gaidet, N.; Mondet, B.; Lancelot, R. Predicting West Nile Virus Seroprevalence in Wild Birds in Senegal. Vector-Borne Zoonotic Dis. 2009, 9, 589–596, . [CrossRef]
- Beveroth, T.A.; Ringia, A.M.; Lampman, R.L.; Novak, R.J.; Ward, M.P. CHANGES IN SEROPREVALENCE OF WEST NILE VIRUS ACROSS ILLINOIS IN FREE-RANGING BIRDS FROM 2001 THROUGH 2004. Am. J. Trop. Med. Hyg. 2006, 74, 174–179, . [CrossRef]
- Islam, A.; Islam, S.; Hossain, M.E.; Ferdous, J.; Abedin, J.; Rahman, M.Z.; Rahman, K.; Hoque, A.; Hassan, M.M. Serological Evidence of West Nile Virus in Wild Birds in Bangladesh. Veter- Sci. 2020, 7, 164, . [CrossRef]
- Niczyporuk, J.S.; Samorek-Salamonowicz, E.; Lecollinet, S.; Pancewicz, S.A.; Kozdruń, W.; Czekaj, H. Occurrence of West Nile Virus Antibodies in Wild Birds, Horses, and Humans in Poland. BioMed Res. Int. 2015, 2015, 1–5, . [CrossRef]
- Amin, M.; Zaim, M.; Edalat, H.; Basseri, H.R.; Yaghoobi-Ershadi, M.R.; Rezaei, F.; Azizi, K.; SalehiVaziri, M.; Ghane, M.; Yousefi, S.; et al. Seroprevalence Study on West Nile Virus (WNV) Infection, a Hidden Viral Disease in Fars Province, Southern Iran. J. Arthropod-Borne Dis. 2020, 14, 173–184, . [CrossRef]
- Paştiu, A.I.; Pap, P.L.; Vágási, C.I.; Niculae, M.; Páll, E.; Domşa, C.; Brudaşcă, F.G.; Spînu, M. Wild Birds in Romania Are More Exposed to West Nile Virus Than to Newcastle Disease Virus. Vector-Borne Zoonotic Dis. 2016, 16, 176–180, . [CrossRef]
- Ferraguti, M.; LA Puente, J.M.-D.; Soriguer, R.; Llorente, F.; Jiménez-Clavero, M..; Figuerola, J. West Nile virus-neutralizing antibodies in wild birds from southern Spain. Epidemiology Infect. 2016, 144, 1907–1911, . [CrossRef]
- Zakhia, R.; Dupuis, A.P.; Khodr, F.; Fadel, M.; Kramer, L.D.; Haddad, N. Evidence of West Nile Virus Circulation in Lebanon. Viruses 2021, 13, 994, . [CrossRef]
- Benjelloun, A.; El Harrak, M.; Calistri, P.; Loutfi, C.; Kabbaj, H.; Conte, A.; Ippoliti, C.; Danzetta, M.L.; Belkadi, B. Seroprevalence of West Nile virus in horses in different Moroccan regions. Vet Med Sci 2017, 3, 198-207.
- Sule, W.F.; Oluwayelu, D.O.; Adedokun, R.A.M.; Rufai, N.; McCracken, F.; Mansfield, K.L.; Johnson, N. High Seroprevelance of West Nile Virus Antibodies Observed in Horses from Southwestern Nigeria. Vector-Borne Zoonotic Dis. 2015, 15, 218–220, . [CrossRef]
- Aquila a-Life. Informe anual de seguimiento de aves liberadas en Madrid 2022. Available online: https://aquila-a-life.org/index.php/es/avances/acciones/monitorizacion/62-acciones-d/476-accion-d1-informe-anual-de-seguimiento-de-aves-liberadas-en-madrid-2022 (accessed on 10 November 2023).
- Instituto Nacional de Estadística. List of place name: Population of the Continuous Municipal Register by Population Unit: Alcobendas. 2024. Available online: https://www.ine.es/en/index.htm (accessed on 25 January 2024).
- Baitchman, E.J.; Tlusty, M.F.; Murphy, H.W. PASSIVE TRANSFER OF MATERNAL ANTIBODIES TO WEST NILE VIRUS IN FLAMINGO CHICKS (PHOENICOPTERUS CHILENSIS AND PHOENICOPTERUS RUBER RUBER). J. Zoo Wildl. Med. 2007, 38, 337–340, . [CrossRef]
- Nemeth, N.M.; Hahn, D.C.; Gould, D.H.; Bowen, R.A. Experimental West Nile Virus Infection in Eastern Screech Owls (Megascops asio). Avian Dis. 2006, 50, 252–258, . [CrossRef]
- Millins, C.; Reid, A.; Curry, P.; Drebot, M.A.; Andonova, M.; Buck, P.; Leighton, F.A. Evaluating the Use of House Sparrow Nestlings as Sentinels for West Nile Virus in Saskatchewan. Vector-Borne Zoonotic Dis. 2011, 11, 53–58, . [CrossRef]
- Nemeth, N.M.; Bowen, R.A. DYNAMICS OF PASSIVE IMMUNITY TO WEST NILE VIRUS IN DOMESTIC CHICKENS (GALLUS GALLUS DOMESTICUS). Am. J. Trop. Med. Hyg. 2007, 76, 310–317, . [CrossRef]
- Xirouchakis, S.M. Breeding biology and reproductive performance of Griffon Vultures Gyps fulvus on the island of Crete (Greece). Bird Study 2010, 57, 213–225, . [CrossRef]
- BirdLife International. IUCN Red List for birds. 2023. Available online: http://datazone.birdlife.org (accessed on 27th October 2023).
- SEO/Birdlife. La Guía de Aves de España. 2023. Available online: https://seo.org/guia-de-aves/ (accessed on 15 November 2023).
- Figuerola, J.; Jiménez-Clavero, M.A.; López, G.; Rubio, C.; Soriguer, R.; Gómez-Tejedor, C.; Tenorio, A. Size matters: West Nile Virus neutralizing antibodies in resident and migratory birds in Spain. Veter- Microbiol. 2008, 132, 39–46, . [CrossRef]
- Vidaña, B.; Busquets, N.; Napp, S.; Pérez-Ramírez, E.; Jiménez-Clavero, M..; Johnson, N. The Role of Birds of Prey in West Nile Virus Epidemiology. Vaccines 2020, 8, 550, . [CrossRef]
- Castillo-Olivares, J.; Mansfield, K.L.; Phipps, L.P.; Johnson, N.; Tearle, J.; Fooks, A.R. Antibody Response in Horses Following Experimental Infection with West Nile Virus Lineages 1 and 2. Transbound. Emerg. Dis. 2011, 58, 206–212, . [CrossRef]
- Jimenez-Clavero, M.A.; Llorente, F.; Sotelo, E.; Soriguer, R.; Gomez-Tejedor, C.; Figuerola, J. West Nile virus serosurveillance in horses in Doñana, Spain, 2005 to 2008. Vet Rec 2010, 167, 379-380.
- European Centre for Disease Prevention and Control. Culex pipiens - Factsheet for experts. 2020. Available online: https://www.ecdc.europa.eu/en/infectious-disease-topics/related-public-health-topics/disease-vectors/facts/mosquito-factsheets/culex-pipiens (accessed on 22nd January 2024).
- Höfle, U.; Blanco, J.M.; Crespo, E.; Naranjo, V.; Jiménez-Clavero, M.A.; Sanchez, A.; de la Fuente, J.; Gortazar, C. West Nile virus in the endangered Spanish imperial eagle. Veter- Microbiol. 2008, 129, 171–178, . [CrossRef]
- Aguilera-Sepúlveda, P.; Gómez-Martín, B.; Agüero, M.; Jiménez-Clavero, M..; Fernández-Pinero, J. A new cluster of West Nile virus lineage 1 isolated from a northern goshawk in Spain. Transbound. Emerg. Dis. 2021, 69, 3121–3127, . [CrossRef]
- Alba, A.; Allepuz, A.; Napp, S.; Soler, M.; Selga, I.; Aranda, C.; Casal, J.; Pages, N.; Hayes, E.B.; Busquets, N. Ecological Surveillance for West Nile in Catalonia (Spain), Learning from a Five-Year Period of Follow-up. Zoonoses Public Heal. 2013, 61, 181–191, . [CrossRef]
- Cano-Terriza, D.; Guerra, R.; Lecollinet, S.; Cerdà-Cuéllar, M.; Cabezón, O.; Almería, S.; García-Bocanegra, I. Epidemiological survey of zoonotic pathogens in feral pigeons (Columba livia var. domestica) and sympatric zoo species in Southern Spain. Comp. Immunol. Microbiol. Infect. Dis. 2015, 43, 22–27, . [CrossRef]
- Figuerola, J.; Jiménez-Clavero, M.A.; Rojo, G.; Gómez-Tejedor, C.; Soriguer, R. Prevalence of West Nile virus neutralizing antibodies in colonial aquatic birds in southern Spain. Avian Pathol. 2007, 36, 209–212, . [CrossRef]
- Gangoso, L.; Grande, J.M.; Llorente, F.; Jiménez-Clavero, M.Á.; Pérez, J.M.; Figuerola, J. Prevalence of neutralizing antibodies to West Nile virus in Eleonora's Falcons in the Canary Islands. J Wildlife Dis 2010, 46, 1321-1324.
- García-Bocanegra, I.; Busquets, N.; Napp, S.; Alba, A.; Zorrilla, I.; Villalba, R.; Arenas, A. Serosurvey of West Nile Virus and Other Flaviviruses of the Japanese Encephalitis Antigenic Complex in Birds from Andalusia, Southern Spain. Vector-Borne Zoonotic Dis. 2011, 11, 1107–1113, . [CrossRef]
- García-Bocanegra, I.; Franco, J.J.; León, C.I.; Barbero-Moyano, J.; García-Miña, M.V.; Fernández-Molera, V.; Gómez, M.B.; Cano-Terriza, D.; Gonzálvez, M. High exposure of West Nile virus in equid and wild bird populations in Spain following the epidemic outbreak in 2020. Transbound. Emerg. Dis. 2022, 69, 3624–3636, . [CrossRef]
- López, G.; Jiménez-Clavero, M..; Tejedor, C.G.; Soriguer, R.; Figuerola, J. Prevalence of West Nile Virus Neutralizing Antibodies in Spain Is Related to the Behavior of Migratory Birds. Vector-Borne Zoonotic Dis. 2008, 8, 615–622, . [CrossRef]
- la Puente, J.M.-D.; Ferraguti, M.; Ruiz, S.; Roiz, D.; Llorente, F.; Pérez-Ramírez, E.; Jiménez-Clavero, M..; Soriguer, R.; Figuerola, J. Mosquito community influences West Nile virus seroprevalence in wild birds: implications for the risk of spillover into human populations. Sci. Rep. 2018, 8, 1–7, . [CrossRef]
- Napp, S.; Montalvo, T.; Piñol-Baena, C.; Gómez-Martín, M.B.; Nicolás-Francisco, O.; Soler, M.; Busquets, N. Usefulness of Eurasian Magpies (Pica pica) for West Nile virus Surveillance in Non-Endemic and Endemic Situations. Viruses 2019, 11, 716, . [CrossRef]
Figure 1.
Geographic distribution of bird and horse sampling sites across the Community of Madrid municipalities. (Adapted from: mapasdeespana.com using Procreate®, Australia).
Figure 1.
Geographic distribution of bird and horse sampling sites across the Community of Madrid municipalities. (Adapted from: mapasdeespana.com using Procreate®, Australia).
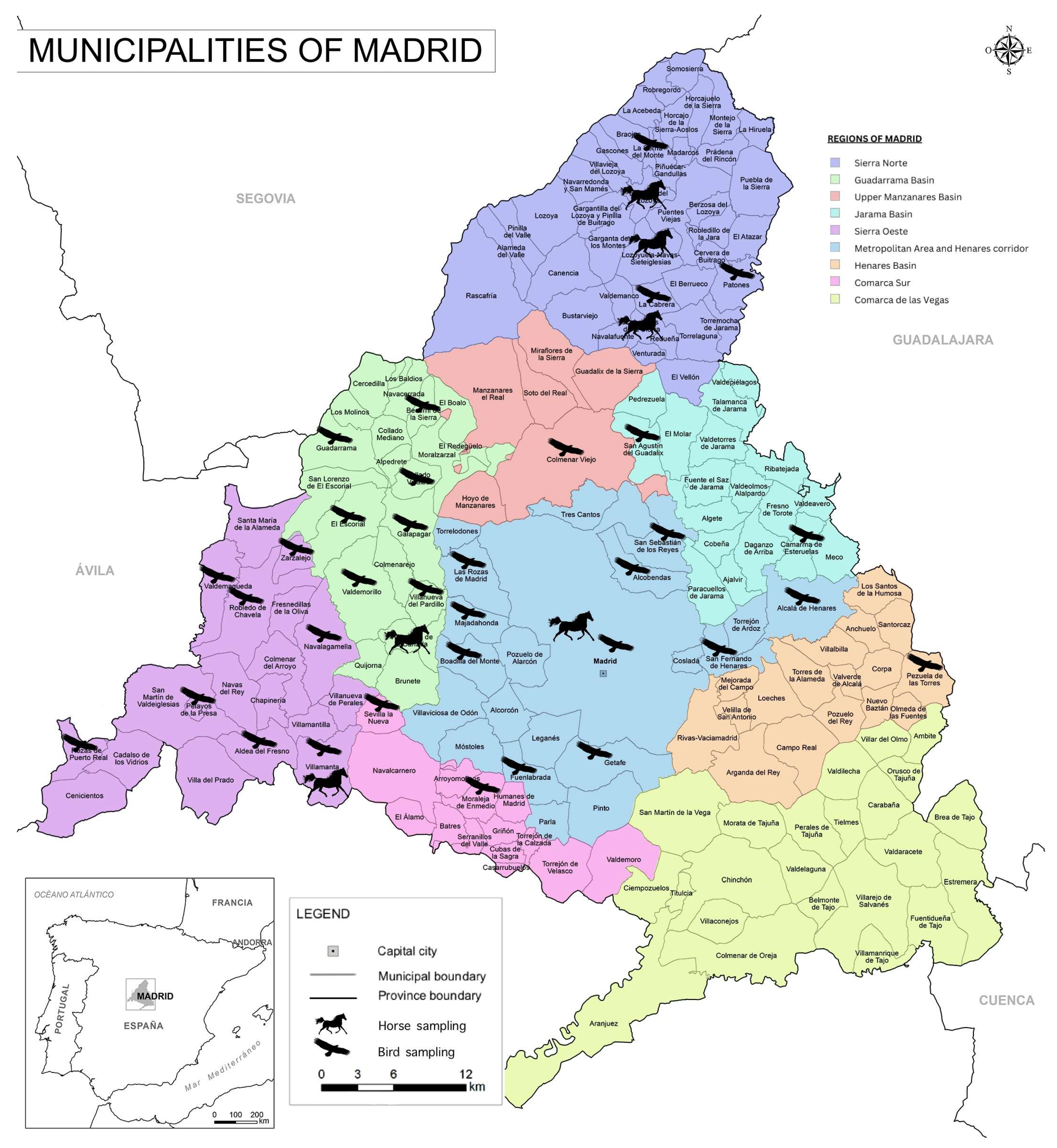
Figure 2.
Municipalities in the Community of Madrid, highlighting the locations where birds and equids tested positive by VNT for West Nile virus (WNV) or for undetermined flavivirus antibodies. (Adapted from: mapasdeespana.com using Procreate®, Australia).
Figure 2.
Municipalities in the Community of Madrid, highlighting the locations where birds and equids tested positive by VNT for West Nile virus (WNV) or for undetermined flavivirus antibodies. (Adapted from: mapasdeespana.com using Procreate®, Australia).
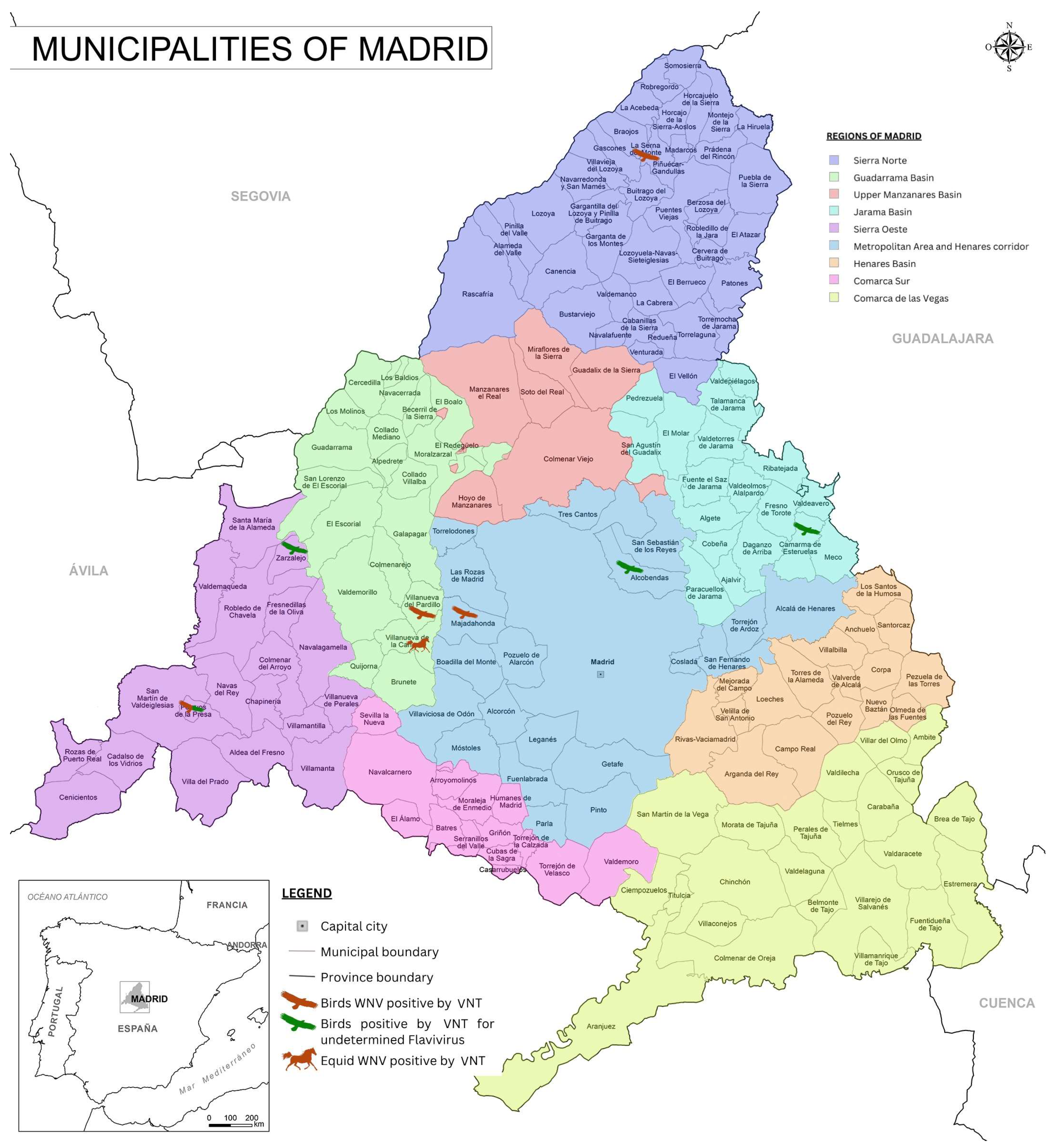

Table 1.
Species of birds and equines tested for flavivirus antibodies against WNV by capture competitive Enzyme-linked Immunosorbent Assay (cELISA) and neutralization tests (VNT).
Table 1.
Species of birds and equines tested for flavivirus antibodies against WNV by capture competitive Enzyme-linked Immunosorbent Assay (cELISA) and neutralization tests (VNT).
| cELISA | VNT | ||||||||||
| Order | Family | Species | Common name | N | Pos | Doubt. | Neg | N | Undet. Flavivirus | WNV | Neg |
| BIRDS | |||||||||||
| Accipitriformes | Accipitridae | Aquila adalberti | Spanish imperial eagle | 3 | 3 | ||||||
| Aquila chrysaetos | Golden eagle | 2 | 2 | ||||||||
| Aquila fasciata | Bonelli's eagle | 10 | 4 | 1 | 5 | 5 | 2 | 1 | 2 | ||
| Gyps fulvus | Griffon vulture | 9 | 1 | 5 | 3 | 6 | 1 | 5 | |||
| Circaetus gallicus | Short-toed snake eagle | 1 | 1 | 0 | 1 | 1 | 0 | ||||
| Milvus migrans | Black kite | 5 | 3 | 1 | 1 | 3 | 1 | 2 | |||
| Milvus milvus | Red kite | 17 | 3 | 14 | 3 | 3 | |||||
| Pernis apivorus | European honey buzzard | 1 | 1 | 0 | 1 | 1 | 0 | ||||
| Charadriiformes | Laridae | Larus fuscus | Lesser black-backed gull | 1 | 1 | ||||||
| Ciconiiformes | Ciconiidae | Ciconia ciconia | White stork | 95 | 3 | 3 | 89 | 6 | 1 | 5 | |
| Ciconia nigra | Black stork | 3 | 3 | ||||||||
| Falconiformes | Falconidae | Falco peregrinus | Peregrine falcon | 4 | 4 | ||||||
| Falco tinnunculus | Common kestrel | 1 | 1 | ||||||||
| Otidiformes | Otididae | Otis tarda | Great Bustard | 1 | 1 | ||||||
| Pelecaniformes | Ardeidae | Ardea cinea | Grey heron | 1 | 1 | ||||||
| Strigiformes | Strigidae | Bubo bubo | Eurasian eagle owl | 4 | 1 | 3 | 1 | 1 | |||
| Suliformes | Phalacrocoracidae | Phalacrocorax carbo | Great cormorant | 1 | 1 | 0 | 1 | 1 | |||
| 158 | 13 | 15 | 131 | 27 | 4 | 4 | 19 | ||||
| EQUINES | |||||||||||
| Perissodactyla | Equidae | Equus caballus | Horse | 23 | 1 | 1 | 21 | 2 | 1 | 0 | |
| Equus asinus | Donkey | 2 | 2 | ||||||||
Abbreviations: cELISA: competitive Enzyme Linked ImmunoSorbent-Assay; VNT: virus microneutralization test; N: total number; Pos: positive; Neg: negative; Doubt: doubtful; Undet: undetermined; WNV: West Nile Virus.
Table 2.
Titers for all VNT positive birds and equines for virus-neutralization Tests (VNT), showing the neutralization against West Nile (WNV), Usutu (USUV) and Bagaza (BAGV) antigen.
Table 2.
Titers for all VNT positive birds and equines for virus-neutralization Tests (VNT), showing the neutralization against West Nile (WNV), Usutu (USUV) and Bagaza (BAGV) antigen.
| cELISA | VNT | ||||||
| Common name | Competition % | Result | WNV | USUV | BAGV | Result | |
| BIRDS | |||||||
| Bonelli's eagle | 48.54 | Positive | 1:10 | 1:20 | <1:10 | Undetermined flavivirus | |
| 73.25 | Positive | 1:10 | <1:10 | <1:10 | Undetermined flavivirus | ||
| 87.2 | Positive | 1:20 | 1:5 | 1.5 | WNV | ||
| Griffon vulture | 35.17 | Doubtful | 1:10 | <1:10 | <1:20 | Undetermined flavivirus | |
| European honey buzzard | 90.98 | Positive | 1:40 | 1:10 | <1:10 | WNV | |
| Black kite | 91.27 | Positive | 1:160 | 1:40 | <1:10 | WNV | |
| White stork | 50.54 | Positive | 1:10 | <1:10 | <1:10 | Undetermined flavivirus | |
| Short-toed snake eagle | No data | Positive | 1:80 | 1:10 | <1:10 | WNV | |
| EQUINES | |||||||
| Horse | 94.74 | Positive | 1:320 | 1:20 | <1:10 | WNV | |
Abbreviations: cELISA: competitive Enzyme Linked ImmunoSorbent-Assay; VNT: virus microneutralization test; %: percentage; WNV: West Nile Virus; USUV: Usutu virus; BAGV: Bagaza Virus.
Table 3.
Details on the age, migratory status, sex, location and zone of all birds found to be VNT positive in this study. Migratory status is shown in parenthesis for one nestling and one fledgling bird, since the former never left its nest, and the latter had only flown very short distances.
Table 3.
Details on the age, migratory status, sex, location and zone of all birds found to be VNT positive in this study. Migratory status is shown in parenthesis for one nestling and one fledgling bird, since the former never left its nest, and the latter had only flown very short distances.
| Common name | VNT Result | Age | Migratory status | Sex | Location | Zone |
|---|---|---|---|---|---|---|
| Bonelli's eagle | Undet. flavivirus | Adult | R | Male | Zarzalejo | Sierra Oeste |
| Undet. flavivirus | Adult | R | Male | Pelayos de la Presa | Sierra Oeste | |
| WNV | Adult | R | Female | Pelayos de la Presa | Sierra Oeste | |
| Griffon vulture | Undet. flavivirus | Fledgling | (R) | Indetermined | Camarma de Esteruelas | Jarama Basin |
| European honey buzzard | WNV | Adult | M | Indetermined | La Serna del Monte | Sierra Norte |
| Black kite | WNV | Adult | M | Female | Majadahonda | Metropolitan area |
| White stork | Undet. flavivirus | Nestling | (M) | Indetermined | Alcobendas | Metropolitan area |
| Short-toed snake eagle | WNV | Adult | M | Male | Villanueva del Pardillo | Guadarrama Basin |
Abbreviations: Under: undetermined; R: resident; M: migrant; ( ): juveniles with limited movement.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



