
Submitted:
27 April 2024
Posted:
28 April 2024
You are already at the latest version
Abstract
Background In this study we investigated in detail the role of cannabidiol (CBD), Beta-caryophyllene (BC), or their combinations in diabetic peripheral neuropathy (DN). The key factors that contribute to DN include mitochondrial dysfunction, inflammation, and oxidative stress. Methods Briefly, Streptozotocin (STZ) (55 mg/kg) was injected intraperitoneally to induce DN in Sprague Dawley rats and performed procedures including Randall Sellito Calipers, Von Frey Aesthesiometer, hot plate, and cold plate methods to determine mechanical and thermal hyperalgesia in vivo. The blood flow to the nerves was assessed by using the Laser Doppler device. Schwann cells were exposed to high glucose (HG) at a dose of 30 mM to induce hyperglycemia and DCFDA, JC1 & Mitosox staining were done to determine mitochondrial membrane potential, reactive oxygen species and mitochondrial superoxides in-vitro. The rats were administered i.p with BC (30mg/kg), CBD (15mg/kg), or combination while Schwann cells were treated with 3.65 µM CBD, 75 µM BC, or combination to assess their role in DN amelioration. Results Our results revealed that exposure to BC and CBD diminished HG-induced hyperglycemia in Schwann cells, in part, through reducing mitochondrial membrane potential, reactive oxygen species and mitochondrial superoxides. Further, BC and CBD combination treatment in-vivo could prevent the deterioration of mitochondrial quality control system by promoting autophagy and mitochondrial biogenesis while improving blood flow. CBD and BC treatments also reduced pain hypersensitivity to hyperalgesia and allodynia with increased antioxidant and anti-inflammatory action in diabetic rats. These in vivo effects were attributed to significant upregulation of AMPK, SIRT3, Nrf2, PINK1, PARKIN, LC3B, Beclin1 and TFAM functions while downregulation of NLRP3 inflammasome, NFκB, COX2 and p62 activity was noted as determined by western blotting. Conclusion the present studies demonstrated that STZ & HG induced oxidative and nitrosative stress play a crucial role in the pathogenesis of diabetic neuropathy. We find, for the first time, that CBD and BC combination ameliorate DN via modulating the mitochondrial quality control system.
Keywords:
Beta-Caryophyllene
; Cannabidiol
; AMPK
; SIRT3
; NLRP3
; mitochondrial biogenesis
; autophagy
Introduction
A common microvascular complication of chronic diabetes, diabetic peripheral neuropathy (DN), impairs the blood supply to peripheral nerves, leading to loss in the nerve conduction and other neuropathic manifestations [1,2]. The number of people suffering from diabetes is anticipated to rise from the current 536.6 million cases to 783.2 million by 2045 [3]. Two drugs frequently prescribed for neuropathic pain are: pregabalin and duloxetine, which function to provide symptomatic relief [4,5]. A general lack of treatment for DN and the incomplete understanding of its pathobiology need further research for development of innovative and effective therapies to overcome DN symptoms.
According to recent studies, decreased biogenesis and mitochondrial function are the main factors driving DN progression [6,7]. The changes in cellular NAD+/NADH ratio are mostly caused by a metabolic protein known as Sirtuin-1 [8,9]. By deacetylating transcription factors, Sirtuin-1 (SIRT1) stimulates downstream targets involved in biogenesis of mitochondria and antioxidant Défense [10,11]. SIRT1 signaling is essential for DN because it regulates mitochondrial function and antioxidant enzymes [11,12,13]. The Nrf2 activation significantly improves mitochondrial function in both preclinical and clinical experiments [14,15]. Reactive Oxygen species (ROS), which oxidize cellular proteins, are produced when hyperglycemia causes electron leakage from mitochondria [16,17]. This reduces the antioxidant defense systems such as heme oxygenase-1, superoxide dismutase and NADPH dehydrogenase quinone 1 [16,18]. Numerous studies have demonstrated that SIRT1 suppresses inflammatory responses by inhibiting the NLRP3 inflammasome in vascular endothelial cells [19,20]. Multiple investigations have also shown that SIRT1 activation is neuroprotective in DN in part by enhancing mitochondrial bioenergetics and autophagy [21,22]. Additional studies have also demonstrated that AMPK and SIRT1 help to reduce DN by improving mitochondrial function through the PGC-1α and Nrf2 axis [23,24,25].
The animal models are utilized to replicate human clinical problems swiftly and accurately, since the disease progression in animal models often follows a pattern similar that noted in human pathologies. As a result, evaluation of potential novel targets for their expected therapeutic benefits becomes straightforward and feasible [26,27,28,29]. Sprague Dawley (SD) rats show a human-like genetic background in addition to having clear genetic tractability [30]. The use of STZ in generation of type 1 diabetes mellitus (T1DM)-induced DN has been validated by numerous published reports [31,32]. The peripheral nerves in these rats undergo pathological changes after induction of hyperglycemia, including axonal atrophy, and demyelination, which eventually leads to decreased sensory and motor nerve conduction [33,34,35]. Schwann cells have been shown to be sensitive to insulin and glucose which are involved in diabetic peripheral neuropathy (DPN) pathogenesis. It is also reported that high glucose causes Schwann cell apoptosis in the development of DN [36,37,38,39].
Cannabinoids, terpenoids, and flavonoids are just a few of the more than 500 unique compounds found in cannabis. Some studies have suggested that the components of the cannabis plant may cooperate to produce superior therapeutic results in pathologies such as hyperglycemia induced DN [40,41]. However, thus far only a few of these cannabinoids have had their benefits investigated. Furthermore, components of the cannabis plant when used in certain combinations may produce superior therapeutic results possibly due to the entourage effect [42,43,44,45,46].
The goal of this study was to ascertain whether the interaction between a terpene (β-caryophyllene, BC) and a minor cannabinoid (cannabidiol, CBD) have therapeutic potential for treatment of hyperglycemia induced DN. CBD is known to be an isomer of Δ9- tetrahydrocannabinol, the main psychoactive substance found in Cannabis sativa [47]. Cannabidiol pharmacology is intricate and is yet to be comprehensively investigated. It should be noted that CBD has a low affinity for cannabinoid receptors CB1 and CB2 and is thought to act on a number of different targets (e.g. 5-Hydroxytrypyamine (5HT1A) instead [48]. β-caryophyllene (BC), a sesquiterpene which is additionally present in clove & black pepper, is one of the terpenes that are also most prevalent in cannabis. It was shown that BC is a naturally occurring, selective CB2 receptor agonist with such positive effects that include analgesia, antioxidant protection, anti-inflammation, and neuroprotection [49,50]. Therefore, there is a need to determine the beneficial and undesirable (negative) consequences of the medical use of cannabinoids given the fast-changing legislation affecting access to cannabinoids and promote interest in the potential medical applications of cannabis (especially for DN) [51,52]. In particular, the combination of cannabinoid compounds could result in superior benefits compared with the benefits noted from their separate use. Here we tested a hypothesis that a combination of a terpene (BC) and a minor cannabinoid (CBD) has superior therapeutic potential in treatment of DN [45,53,54,55,56].
We conducted studies to determine the protective effects of BC and CBD combination on HG induced Schwann cells neurotoxicity in vitro and STZ-induced DN in rats by performing behavioural, functional and antioxidant analyses. Further, ROS generation, changes in mitochondrial membrane potential, and mitochondrial superoxides were evaluated by western blotting, immunohistochemistry and intraepidermal nerve fiber (IENF) density to determine the impact of BC and CBD on mitochondrial dysfunction and mitochondrial biogenesis.
Material and Methods
- Experimental design
Human Schwann cells (obtained from ATCC) were cultivated at 37 °C in a humidified environment of 95% air and 5% CO2 and cultured in DMEM (5.5 mM glucose) enriched with 10% FBS, glutamine (2 mM), and streptomycin/penicillin (1%). Cell culture condition with high glucose was created by adding 24.5 mM glucose to the solution that resulted in a final glucose concentration of 30 mM. In the cell culture system, diabetic condition is defined as glucose levels that exceed 10 mM [57,58] This study used a 30 mM glucose concentration that was previously used to promote in vitro diabetic conditions in many cellular models of diabetes [59,60,61,62,63,64]. The Schwann cells were plated into a 6-well plate in DMEM medium containing 10% FBS. The treatment groups consisted of a) normal control cells (NC) b) high glucose (HG)-treated cells (30 mM) c) HG induced cells treated with 75 µM of BC (HG+BC) d) HG induced cells treated with 8 µM of CBD (HG+CBD) and e) HG induced cells treated with 75 & 3.64 µM of BC and CBD (HG+BC+CBD), respectively. At the conclusion of 24 h treatment, various biochemical and molecular parameters were measured.
- Cell viability determination
With a density of roughly 5000–8000 cells per well, the Schwann cells were plated in 96-well plates with DMEM medium and maintained for 24 hours. After 24 hours of incubation, cells were exposed to various concentrations of CBD (100-3.125 µM), BC (500-15.625 µM) & BC+CBD (8-0.25 µM) respectively. Cell viability was determined by MTT assay. Briefly, 100 µl MTT (5 mg/10 ml) was added to plates after 24 hours of treatments, and the plates were then maintained in an incubator at 37 °C for 4 hours. The media was removed and 100 µl of DMSO was added to 96-well plates for 10 min and the absorbance value was recorded at 570 nm using a TECAN infinite M200 plate reader [65].
- DCFDA
- FDA staining to measure intracellular ROS
Schwann cells were seeded at a density of 5×104 cells per well in a six well plate and treated with HG (30 mM) as well as with BC (75 µM), CBD (8 µM) & BC +CBD (75 µM+ 3.64 µM) for 12 hours. After 12 hours 10 µM of DCFDA dye (Cat no. D399; Thermo Fisher Scientific) were added in each well and cells were incubated for additional 30 minutes followed by PBS washing. Cells were then imaged using OLYMPUS IX73 microscope. Images were analysed by image j software [66,67,68].
- MitoSOX staining to measure mitochondrial superoxides
Schwann cells were seeded in 6 well plates as above and treated with HG (30 mM) as well as BC (75 µM), CBD (8 µM) & BC +CBD (75 µM+ 3.64 µM) for 12 hours to measure the amount of mitochondrial superoxide anion produced. After treatment period of 12 hours, 5 µM of MitoSOX dye (Cat no. M36008; Thermo Fisher Scientific) was added, cells were incubated for 15 min followed by PBS washing. Fluorescence images were captured using the OLYMPUS IX73 microscope. Images were analysed by image j software [69,70].
- JC1
- JC1 staining for mitochondrial membrane potential
Hyperglycemia was induced with HG (30 mM) exposure to the Schwann cells followed by treatments with BC (75 µM), CBD (8 µM) & BC+CBD (75 µM+ 3.64 µM) for 12 hours. After 12 hours, the cells were incubated for 15 min with 5 µM of the JC-1 dye (Cat no. T3168; Thermo Fisher Scientific). After three PBS washes, cells were photographed using an OLYMPUS IX73 microscope. Images were analysed by image j software [69,71,72].
- Western blotting
Schwann cells were seeded at a concentration of 2×106 in a 6 well plates with DMEM media. Hyperglycemia was induced by HG (30 mM) followed by BC (75 µM), CBD (8 µM) & BC +CBD (75 µM+ 3.64 µM) treatment for 24 hours. After performing a phosphate buffer wash, the cell lysate was prepared by using RIPA buffer with phosphatase inhibitor and protease inhibitor (1:100) and incubated for 30 minutes on ice. The supernatant was then collected by centrifuging the lysates at 12000 rpm for 15 minutes. Bradford reagent was used to estimate the amount of protein in the samples. A 40–60 µg of equivalent proteins was added with loading buffer, heated at 98 °C for 10 min and then electrophoresed on SDS–PAGE. After the electrophoresis, the proteins were transferred to nitrocellulose membranes and the membranes incubated in 5% BSA in TBST. The membranes were then incubated with primary antibodies prepared at dilution of 1:1000 to 1:3000 in the TBST at 40C overnight on a shaker. Various primary antibodies were purchased either from Cell Signalling Technology or Santa Cruz biotechnology. These antibodies included AMPK (Cat no.2532S), SIRT1 (Cat no. 9475S), Sirt3 (Cat no. 5490S), PGC-1α (Cat no. 2178S), NRF1 (Cat no. 46743S), PINK1 (Cat no. SC517353), PARKIN (Cat no. SC32282), p62 (Cat no. 5114S), TFAM (Cat no. 8076S), SOD2 (Cat no. SC30080), Nrf2 (Cat no. 12721S), KEAP1 (Cat no. 8047S), HO-1 (Cat no. SC390991), NQO1 (cat no. SC376023), phosphomTOR (cat no. 2971S), NLRP3 (cat no. 15101S), ASC (Cat no. 67824S), Caspase-1 (Cat no. 3866S), IL-1 β (Cat no. 12242S), IL-18 (Cat no. 54943S), PHB2 (Cat no. 14085S), BAX (Cat no. 2772S), BCL2 (Cat no. 3498S), COX2 (Cat no. 12282S), Phospho-NF-κB p65 (Ser536) (93H1) (Cat no. 3033S), FOXO3a (Cat no.2497S), Beclin1 (Cat no. 3495S), atg3 (Cat no. 3414S), atg7 (Cat no. 8558S), LC3B (Cat no. 2775S) (CST, USA), and β-Actin (Cat no. 4970L). After incubation with primary antibodies, membranes were washed three times for 10 min each. Finally, the secondary antibodies tagged with HRP anti-rabbit (Cat no. 7074S), or anti-mouse (Cat no. 7076S) were added onto the membrane and kept at room temperature for 1.30 hours on a shaker followed by washing with TBST. The signals for presence of respective proteins on the NC membrane were captured by using a ChemiDocTM XRS+ Imaging system (BIO-RAD Molecular Imager) and the relative band intensities were quantified by using densitometry Image-J software [73,74,75,76,77].
- In vivo Experiments
32 healthy male Sprague-Dawley rats, aged three months, weighing 200-280 g, were kept in plastic cages with a 12-hour light/dark cycle and were supplied a regular diet and water as needed. The American Association for Accreditation of Laboratory Animal Care (AAALAC) has given FAMU approval for the facilities where animals are kept and cared for in accordance with strict pathogen-free standards. Animals were given a week of acclimatization prior to the commencement of studies following Florida A&M University’s Institutional Animal Care and Use Committee (IACUC) standards (protocol number: 021-04 dated 05/10/2021) and Animal Research Reporting of in-vivo studies (ARRIVE) guidelines. Efforts were made to reduce the number of animals used and their distress [78].
- Induction of diabetic neuropathy and the experimental designs
SD male rats were selected and fasted for 2 h. STZ at 55 mg/kg, dissolved in the citrate buffer, was given through i.p. route to develop diabetes. Rats with their plasma glucose levels higher than 250 mg/dl were taken as diabetic for the study [31,79,80,81]. Animals with diabetes were randomized into groups consisting of diabetic control animal group (STZ, n=8), diabetic rats treated with 30 mg/kg BC, i.p. (STZ+BC30, n=6) and 15 mg/kg CBD, i.p. (STZ+CBD15, n=6), diabetic rats treated with 30 mg/kg BC+15 mg/kg CBD, i.p. (STZ+BC30+CBD15, n=6) after 5th week of DN induction. The dosing was done thrice a week for three weeks. Age matched control rats were included as normal control animals (NC, n=6). The treatment scheme is depicted in Figure 1. After last dose of drug administration behavioural and functional parameters were performed followed by animal sacrifice with sciatic nerve, spinal cord, DRG and hind paw collection and tissue were stored in -80oC for western blotting and in formalin for histology. Both, CBD and BC were dissolved in a vehicle comprised of 5% DMSO, 5% ethanol, 5% Tween 80, and 85% saline [82].
- Behavioural parameters
- Mechanical and thermal hyperalgesia
A cutoff time of 15 seconds was used to record the time duration to flick the rat tail during hot plate tests at a temp 45°C and cold plate tests at a temp 10°C, and in the 30 second interval between each reading. At least three readings were taken to calculate the paw withdrawal time in seconds [32]. Von Frey Aesthesiometer test & Randall Sellito Calipers were used to determine mechanical hyperalgesia. The animal paw withdrawal threshold in grams is determined by the force at which it was observed. A minimum of 5 readings were taken, with a 5 minute gap between each [70].
- Nerve functional studies
- Nerve blood flow
According to our earlier findings, a Laser Doppler Oxymeter (Moor Instruments, UK) was used to measure the blood flow in the animals’ sciatic nerves. The sciatic nerve of sedated animals was exposed on the left flank, and by utilizing moor instrument software, the flux was monitored for 10 minutes. The arbitrary perfusion units for the animals were recorded [83].
- Western blotting
The sciatic nerve was taken from -800C and lysate were prepared by tissue extraction reagent (TPER, Sigma) containing protease and phosphatase inhibitors. Remaining all of the procedure was followed as mentioned above in Schwann cell line WB experiments.
- Immunohistochemistry (IHC)
Briefly, deparaffinized rehydrated 5 µm microsections of sciatic nerves were heated in citrate buffer at pH: 6.0 for antigen retrieval. The sections were treated with 3% H2O2 to prevent endogenous peroxidase activity, followed by a 60-minute blocking step with 3% BSA in PBS. The nerve microsections were then incubated with SOD2 antibody at 1:200 dilution (Cat no. SC30080), LC3B antibody at a dilution of 1:400 (Cat no. 2775S), or NF-κB p65 (D14E12) antibody at 1:200 dilution (Cat no.8242S) (CST, USA) for two hours at room temperature. The sections were then washed with tris buffered saline (pH 7.4) before being incubated for 30 minutes at room temperature with anti-rabbit secondary antibodies (1:200) dilution each; CST, USA). Chromogenic staining was performed using the VECTASTAIN® Elite ABC Reagent kit (Vector Labs, CA, USA) according to the manufacturer’s instructions, and the emergence of a brown color was noted. The sections were counterstained with hematoxylin, dehydrated, and mounted with DPX. To ascertain the immunopositivity, each segment was examined with an OLYMPUS IX73 microscope. Image analysis were done by image J software [83].
- Intra
- tra epidermal nerve fibre density in the hind paw of diabetic rats
8 µm sections of a diabetic rat hind paw was treated in accordance with the above IHC procedure. The nerve fibers were marked with the PGP 9.5 antibody (Invitrogen, Thermo Fisher Scientific; Cat no. 381000) at 1:200 dilutions. Photographs were captured at a 20x magnification and numbers of nerve fibers to the epidermal junction were calculated in relation to the length of the tissue under examination. In each group at least five sections were counted, and the average number of nerve fibers/mm was derived. Images were analysed by image J software [69,83].
- Statistical analysis
Data were expressed as Mean± standard error mean (SEM) and were compared by one way analysis of variance (ANOVA) with the help of Graph Pad Prism followed by ‘’Benferroni’s multiple comparison post hoc test”. p<0.05 was considered statistically significant.
Results
- BC, CBD, and their combination effect on the cell viability
MTT results revealed that the dose of BC, CBD & BC+CBD at 75 µM, 8 µM & 3.65 µM, respectively, was suitable to evaluate their protective potential in high glucose exposed Schwann cells (Figure 2).
- Effect of BC, CBD, and their combination on the generation of ROS, mitochondrial superoxide, and the mitochondrial potential in high glucose exposed Schwann cells
DCFDA and Mito Sox labelling were used to evaluate the ROS and the mitochondrial superoxide levels in Schwann cells. As compared to cells in a normoglycemic environment, we observed that hyperglycemia induced a significant (p < 0.001) rise in ROS and the mitochondrial superoxide (Figure 3A, B and 3E, F). BC and CBD treatments at 75 and 3.65 µM, respectively, resulted in a significant (p < 0.001) reduction in the ROS and superoxide levels (Figure 3A & 3E).
JC1 labelling showed two separate cellular populations, a green fluorescence that indicates JC1 in its monomeric state and an orange fluorescence that signals the formation of JC1 aggregates which means negatively polarized mitochondria. When compared to high glucose conditions, BC, and CBD treatment at 75 & 3.65 µM doses, respectively, restored negative MMP (mitochondrial membrane potential) (p < 0.001) (Figure 3C, D).
- BC, CBD and their combination effect on mitochondrial biogenesis and antioxidant effect in HG induced Schwann cells
AMPK, SIRT1 & PGC-1α have significant roles in mitochondrial quality functions especially in the mitochondriogenesis and autophagy. Figure 4A show expression of AMPK, SIRT1, PGC-1α, Nrf2, HO-1, SOD2, and NQO1 in Schwann cells that were either untreated or treated with HG, HG+BC, HG+CBD, or HG+BC+CBD. The mitochondrial proteins expression was significantly decreased for AMPK (p < 0.001), SIRT1 (p < 0.001), PGC-1α (p < 0.001), Nrf2 (p < 0.001), HO-1 (p < 0.001), SOD2 (p < 0.001) and NQO1 (p < 0.001) in HG-induced Schwann cells when compared to the untreated cells (Figure 4B). The quantitation analyses in Figure 4B also show that BC and CBD combination treatment significantly increased the AMPK (p < 0.01), SIRT1 (p < 0.001), PGC-1α (p < 0.001), HO-1 (p < 0.001), SOD2 (p < 0.001) and NQO1 (p < 0.001) expression levels relative to their HG induced counterparts. The results suggest that BC and CBD combination provide a protective mitochondriogenesis through promoting increased expression of AMPK, PGC-1α & SIRT1 in HG-induced cells that likely function to restore normal mitochondrial function (Figure 4A & 4B).
- BC, CBD, and their combination effects on neuroinflammation in HG induced Schwann cells
NLRP3, ASC, IL-1β, IL18, pNFkB, and COX2 are well known pro-inflammatory signalling molecules. Figure 5A shows expression of NLRP3, ASC, IL-1β, IL18, pNFkB, and COX2 in Schwann cells that were either untreated or treated with HG, HG+BC, HG+CBD, or HG+BC+CBD. Expression and/or phosphorylation (activation) of NLRP3 (p < 0.001), ASC (p < 0.001), IL-1β (p < 0.001), IL18 (p < 0.001), was significantly elevated in HG-treated Schwann cells as compared to normal, untreated cells (Figure 5B). The quantitation analyses in Figure 5B also show that BC and CBD combination significantly decreased NLRP3 (p < 0.001), ASC (p < 0.001), IL-1β (p < 0.001), and IL18 (p < 0.001) expression relative to their HG induced counterparts, indicating that HG promotes inflammasome activation that could enhance the neuroinflammation which was reduced by the BC and CBD combination treatment. Additionally, pNFkB (p < 0.001) and COX2 (p < 0.001) were also significantly elevated in the HG-treated Schwann cells while there was significant reduction in the levels of pNFkB (p < 0.001) and COX2 (p < 0.001) in cells treated with HG+BC, HG+CBD, or HG+BC+CBD relative to HG-treated cells (Figure 5B). Together these data suggest that BC and CBD combination has anti-inflammatory action in HG-treated Schwann cells (Figure 5A & 5B).
- BC and CBD combination effect on mitochondrial quality control and autophagy in high glucose induced Schwann cells
Expression of the mitochondrial proteins SirT3, PHB2, PINK1, PARKIN play roles in mitochondrial quality control and regulation of autophagy. Figure 6A shows expression of Sirt3, PHB2, PINK1, PARKIN, phospho-mTOR, p62, and LC3B in Schwann cells that were either untreated or treated with HG, HG+BC, HG+CBD, or HG+BC+CBD. The quantitation analyses in Figure 6B further show that Schwann cells when exposed to HG (30 mM) resulted in significant decrease in SirT3 (p < 0.001), PHB2 (p < 0.001), PINK1 (p < 0.001), PARKIN (p < 0.001), and LC3B (p<0.001) while expression of phospho-mTOR (p < 0.001), p62 (p < 0.001) was significantly increased when compared with their normal, untreated cells indicating that HG treatment likely impact mitochondrial quality and promote autophagy. CBD and HG+CBD+BC treatment significantly enhanced SirT3 (p < 0.001), PHB2 (p < 0.001), PINK1 (p < 0.001), and PARKIN (p < 0.001) expression relative to their respective levels in HG-treated cells, suggesting a positive role of these proteins in maintaining mitochondrial quality control (Figure 6A & 6B). Additionally, the expression of phospho-mTOR (p < 0.001), p62 (p < 0.001) significantly increased and LC3B (p<0.001) significantly decreased in the HG-treated Schwann cells as compared to normal Schwann cells, while the BC and CBD combination caused significant decrease in phospho-mTOR (p < 0.001), p62 (p < 0.001) and increase in LC3B (p < 0.001) expression, suggesting that BC and CBD combination has significant protective role in mitochondrial autophagy role relative to the HG treatments.
- BC, CBD, and their combination effect on nerve function in SD rats
Effects of BC, CBD, or their combination were investigated in the SD rat model as described in methods. Animals were administered STZ to induce diabetes. The diabetic animals showed significantly reduced paw withdrawal latency responses in Cold plate (p < 0.001) (Figure 7A), hot plate (p < 0.001) (Figure 7B) and Hargreaves test (p < 0.001) (Figure 7C). Similar reduction of such responses was also noted by mechanical Randall-Selitto (p < 0.001) (Figure 8B), Von Frey (p < 0.001) (Figure 8A) studies. BC and CBD treatments at a dose of 30 and 15 mg/kg, respectively, for 3 weeks or a combination of both the agents significantly improved the paw withdrawal latency responses in thermal Cold plate (p < 0.001) (Figure 7A), hot plate (p < 0.001) (Figure 7B), and Hargreaves test (p < 0.001) (Fig 7C) as well as the mechanical hyperalgesia, Randall-Selitto (p < 0.001) (Figure 8B),Von Frey (p < 0.001) (Fig 8A) readings in the diabetic rats. The STZ-induced diabetes over the 8-week period also impaired the nerve blood flow significantly (p < 0.001) (Figure 8C) as compared to normal control rats. Three-week treatment with BC and CBD (p < 0.001), significantly restored the nerve blood flow (p < 0.001) impairments associated with DN (Figure 8C).
- BC, CBD, and their combination effect on mitochondrial biogenesis in diabetic SD rats
Diabetic animals were treated with BC, CBD, or a combination as noted above. Following the treatments, the sciatic nerves were analysed first by Immunohistochemistry for levels of SDO2 (Figure 9A). The diabetic SD rat sciatic nerve showed a visible decline in SOD2 staining when compared with the control group rats. The BC and CBD treated rats showed increased SOD2 as noted by brown staining in Figure 9A. Histogram in Figure 9B shows relative levels of SOD2 in stained sciatic nerves in Figure 9A. In SD diabetic rat sciatic nerves SOD2 (p < 0.001) protein declined significantly, while treatments with BC, CBD or a combination resulted in significant restoration in SOD2 levels in sciatic nerves (9B). Similar to our observations in the Schwann cell culture assays, western blot analyses of the sciatic nerves in diabetic rats also showed a significant decrease in AMPK (p < 0.001), SIRT1 (p < 0.001) expression when compared to their levels in the sciatic nerves of normal control group animals. The decline in the expression of SIRT1 resulted in significantly decreased expression of PGC-1α (p < 0.001) in sciatic nerves of diabetic SD rats relative to the PGC-1α levels in the sciatic nerves of normal control group rats (Figure 9C, D). Upon BC and CBD combination treatments, there was a significant increase in the levels of AMPK (p < 0.001), SIRT1 (p < 0.01) & PGC-1α (p < 0.01) in the sciatic nerves of diabetic SD rats relative to the levels of these proteins in the sciatic nerves of their untreated counterparts (Figure 9C, D). As also noted in the in-vitro studies, STZ-treated diabetic rats had reduced levels of NRF1 (p < 0.001), TFAM (p < 0.001), SOD2 (p < 0.01), NQO1 (p < 0.001), and HO1 (p < 0.001) proteins as compared to their respective levels in the control group rats. BC and CBD treatment with 30 and 15 mg/kg body weight dose, respectively, resulted in a significantly increased levels of NRF1 (p < 0.001), TFAM (p < 0.001), SOD2 (p < 0.01), NQO1 (p < 0.001), and HO1 (p < 0.001) proteins relative to their levels in the sciatic nerves of the diabetic animals (Figure 9C, D).
- BC, CBD and their combination effect on inflammasome and Nrf2 linked antioxidant effect in diabetic SD rats
The sciatic nerves from the control, untreated, diabetic animals, and the diabetic animals that were treated with BC, CBD, or a combination were analysed first by Immunohistochemistry for levels of pNFkB as above (Figure 10A). The diabetic SD rat sciatic nerve showed a visible increase in pNFkB staining when compared with the pNFkB staining in the control group rats. The BC and CBD treated rats showed decreased pNFkB as noted by brown staining in Figure 10A. Histogram in Figure 10B shows relative levels of pNFkB in stained sciatic nerves in Figure 10A. In SD diabetic rat sciatic nerves pNFkB (p < 0.001) protein increased significantly, while treatments with BC, CBD or a combination resulted in significant decline in pNFkB levels in sciatic nerves (Figure 10B). Similar to our data in the Schwann cell culture assays, western blot analyses of the sciatic nerves in diabetic rats also showed a significant increase in pro-inflammatory genes NLRP3 (p < 0.001), IL18 (p < 0.001), COX2 (p < 0.001), pNFkB (p < 0.001), and Keap1 (p < 0.001), while decrease in the level of Nrf2 (p < 0.001), and FOXO3a (p < 0.001) occurred relative to their respective levels in the sciatic nerves of untreated control animals (Figure 10C, D). BC and CBD at a dose of 30 and 15 mg/kg, respectively, for 3 weeks significantly decreased the level of NLRP3 (p < 0.001), IL18 (p < 0.001), COX2 (p < 0.01), pNFkB (p < 0.05), and Keap1 (p < 0.001) while inducing expression of Nrf2 (p < 0.01), FOXO3a (p < 0.001) relative to their respective levels in the sciatic nerves of the diabetic animals (Figure 10C & 10D). Collectively, our data in Figure 10 suggest that BC and CBD treatments lower the inflammatory responses while promoting antioxidant signalling in the sciatic nerves of the diabetic animals.
- BC, CBD, and their combination effect on autophagy in the diabetic SD rats
Several mitochondrial proteins play important roles in mitochondrial quality control and regulation of autophagy. On the basis of our in vitro studies in Schwann cells, we determined whether induction of diabetes and subsequent interventions by administration of BC and CBD altered levels of autophagy-regulating proteins in the sciatic nerves of SD rats. First, the sciatic nerves from the control, untreated, diabetic animals, and the diabetic animals that were treated with BC, CBD, or a combination were analysed by Immunohistochemistry for levels of LC3B as above (Figure 11A). The diabetic SD rat sciatic nerve showed a visible decline in LC3B staining when compared with the LC3B staining in the sciatic nerve from the control group rats. The BC and CBD treated rats showed an elevated level of LC3B in their sciatic nerves relative to the LC3B levels in the sciatic nerve from the diabetic SD rats (Figure 11A). Western blot analyses of the proteins from peripheral sciatic nerves in diabetic rats also showed a significant decrease in the expression levels of SirT3 (p < 0.001), PINK1 (p < 0.001), PARKIN (p < 0.001), LC3B (p < 0.001), Beclin1 (p < 0.001), atg3 (p < 0.001), and atg7 (p < 0.001) proteins while there was significant increase in the expression of p62 (p < 0.001) when compared to the levels of the respective proteins in the peripheral sciatic nerves of control group rats (Figure 11C, D). BC and CBD at a dose of 30 and 15 mg/kg, respectively, for 3 weeks significantly increased the sirt3 (p < 0.001), PINK1 (p < 0.001), PARKIN (p < 0.01), LC3B (p < 0.01), Beclin1 (p < 0.001), atg3 (p < 0.001), atg7 (p < 0.001) while there was significant decrease in the expression of p62 (p < 0.05) protein relative to their respective levels in the sciatic nerves of the diabetic animals (Fig 11C & 11D). Additionally, the level of apoptotic marker BCL2 (p < 0.001) was significantly reduced while BAX (p < 0.001) was significantly increased in sciatic nerves from diabetic rats as compared to its levels in sciatic nerves from the control group rats. BC and CBD treatments caused significant increase and decrease in the BCL2 (p < 0.001) and BAX (p < 0.001) protein levels, respectively, suggesting that BC and CBD combination interventions serve to inhibit apoptosis in the DN (Figure 11C, D). The data in Figure 11 further support our hypothesis that BC and CBD play significant roles in the treatment of diabetic neuropathy in part through reduction of autophagy and nerve apoptosis.
- BC, CBD and their combination effect on the loss of Intraepidermal nerve fiber (IENF) in the STZ induced neuropathic SD rats
As the DN pathology is known to cause degeneration of IENFs, we conducted histological staining of the nerve fibers obtained from the control, untreated, diabetic animals, and the diabetic animals that were treated with BC, CBD, or a combination. The diabetic SD rat showed a decline in IENF staining when compared with the IENF levels noted in the control group rat (Figure 12A). The BC and CBD treated rat showed an elevated level of IENFs relative to that noted in diabetic SD rats (Figure 12A). The quantitation analyses in Figure 12B further show that the DN rat had a significant IENF degeneration (p < 0.001) as compared to the IENF in the control group rats while BC and CBD combination treatments resulted in significant restoration (p < 0.001) of IENF levels relative to that noted in their diabetic counterparts (Figure 12A).
Discussion
Our present study focused on the evaluation of the antioxidant, anti-inflammatory and neuroprotective potential of BC and CBD in HG-promoted toxicity and STZ-induced diabetic neuropathy through alterations in AMPK and SIRT1 activation which may also be connected to altered, PGC-1α mediated, mitochondrial biogenesis, PARKIN-PINK1-p62 mediated autophagy, antioxidant effect through Nrf2 overactivation and anti-inflammatory effect through NFkB and NLRP3 inflammasome. Though many studies have reported the role of CBD and BC in neuropathy, this will be the first study to evaluate the role of their combination in diabetic peripheral neuropathy [84,85,86].
Increased amounts of ROS, mitochondrial superoxides, and mitochondrial membrane depolarization of mitochondrial dysfunction were observed in Schwann cells after high glucose exposure. Reduced superoxide generation following the BC and CBD therapy prevented high glucose-dependent ROS formation, superoxide generation & mitochondrial membrane depolarization. These effects of BC and CBD have demonstrated their mito-protective properties, thus providing a rationale for further assessment of the molecular underpinnings. These results also unambiguously imply that BC and CBD may protect against high glucose-induced neurotoxicity by limiting the cellular homeostasis and balancing oxidative stress and mitochondrial function [87]. The study by Hashish HM & Ni B et al [49] observed that BC and CBD treatment prevented HG-induced mitochondrial dysfunction and ROS production, respectively, by impacting, AMPK and SIRT1 induced mitochondrial biogenesis and Nrf2-mediated antioxidant signalling [40,88]. These observations demonstrate that mitochondrial biogenesis deficits play a critical role in the pathophysiology of DN, and activation of AMPK and SIRT1 function to restore this condition by promoting mitochondrial biogenesis. These findings are consistent with other studies that demonstrate that activation of SIRT1 alleviates oxidative damage and restores mitochondrial biogenesis [80,89,90]. The current findings point to the involvement of autophagy and mitochondrial biogenesis in the pathophysiology of DN triggered by Type 1 DM and provide a proof-of-concept rationale for the use of CBD and BC as pharmacological moderators of mitochondrial dysfunction through SIRT1 and AMPK [88]. The characteristics of STZ-induced DN in rats include lower nerve conduction and blood supply to the peripheral nerves, and increased pain hypersensitivity to hyperalgesia and allodynia. In diabetic rats, abnormal nerve conduction in the peripheral nerve was connected to a variety of abnormal sensations, such as tingling and discomfort [91]. The advancement of peripheral neuropathy in rats was confirmed by the development of tactile allodynia, mechanical hyperalgesia, and lower NBF levels in rats with chronic hyperglycaemia. In this context, the study by Wang G et al demonstrated that treatment with cannabidiol induces autophagy and improves neuronal health [92].
In diabetic rats, decreased AMPK Thr 172 phosphorylation impairs the development of autophagosomes and mitochondrial biogenesis. Our experiments also provided evidence that, in hyperglycemic settings, AMPK activation increases PGC-1α mediated mitochondrial biogenesis and reduces the activation of NFκB induced neuroinflammation. It is also known that NFκB mediated neuroinflammation affects vascular impairments seen in DN via inducing COX-2. BC and CBD combination treatments increased peripheral blood flow to the neurons in diabetic mice, which may be because AMPK activation of NFκB inhibition [93,94,95,96]. By enhancing mitochondrial activity, SIRT1 activation has also been associated to a decrease in aging and other chronic disorders [97]. Activating SIRT1 has been shown to lower oxidative stress, have anti-inflammatory effects, and improve mitochondrial function, according to earlier reports [98,99,100]. By restricting mTOR phosphorylation (ser 2884), AMPK activation aids autophagy induction and prevents the depolarized and ROS producing mitochondria, while concurrently promoting the production of new functional mitochondria in peripheral nerves under hyperglycaemic conditions [101,102]. Through the stimulation of FOXO3a, SIRT1 and AMPK activation have also been shown to have the capacity to reverse metabolic memory by reversing the epigenetic modifications caused by high glucose in stem cells. Previously, it was observed that SIRT1 expression decreased oxidative stress in disease-related experimental models via deacetylating FOXO3a activity [103,104]. In our studies, increased IHC expression of NFκB, COX-2 in the sciatic nerves of diabetic SD rats and increased expression levels of NFκB, COX-2 in Schwann cells exposed to high glucose were evidence of the neuroinflammation caused by hyperglycaemia. Both in STZ induced diabetic rats and in Schwann cells exposed to hyperglycaemia, AMPK activation by BC and CBD decreased the neuroinflammation linked to NFκB signalling, which is consistent with past studies on AMPK directed NFκB suppression [105,106].
The capacity of BC and CBD to reverse the impairments of nerve blood flow in diabetic rats may be partly due to the reduction of oxidative damage to the peripheral nerves. Through involvement in the Nrf2/NFκB crosstalk, BC and CBD combination likely affect cellular inflammation, redox state, and the response to oxidative stress [40,107]. Our hypothesis that BC and CBD combination mediated neuroprotection in DN may be due to their antioxidant and anti-inflammatory activity is supported by increased SIRT3, SOD2, and Nrf2 in diabetic rats treated with BC and CBD combination and decreased NLRP3, NFκB, and COX2 levels which is also supported by other prior findings [108,109,110].
Activation of NLRP3 has been linked to a variety of diseases from cancer to infectious disorders, [111]. NLRP3 role in diabetic neuropathy have yet to be fully determined. Even though NLRP3 is the inflammasome that has been most thoroughly studied, mechanisms regulating its targets are not completely elucidated. Discovering and developing treatment candidates that target NLRP3 in many inflammatory disease situations, including DN, therefore will depend on understanding the cellular and molecular features of NLRP3 inflammasome activation and inhibition. A few studies have recently clarified the potential function of autophagy and mitochondrial quality control in the regulation of NLRP3 activation [112,113]. Evidence is mounting that NLRP3 activation is crucial to the development of diabetic neuropathy after nerve damage [114,115]. Thus, elevating Nrf2 activity may induce autophagy, which may lessen NLRP3-induced neuroimmune abnormalities while also enhancing the antioxidant status of the neurons.
Under typical conditions, Nrf2 is sequestered in the cytoplasm by the Kelch-like ECH-associated protein 1 (Keap1)-Nrf2 complex, where it is susceptible to ubiquitin-mediated destruction. Oxidative stress promotes the Keap1-Nrf2 complex separation. Next, Nrf2 enters the nucleus and activates ARE to regulate the transcription of antioxidant genes. The primary regulator which is involved in the endogenous antioxidant défense system is Nrf2 [99,116,117,118,119]. In rats with STZ-induced diabetes, the loss of Nrf2-mediated antioxidant defense due to T1DM increased oxidative burden has previously been linked to nerve damage [120]. We also found decreased Nrf2 signalling and the expression of the enzyme NQO1 under diabetes circumstances, further corroborating the earlier findings [121,122]. Diabetic rats treated with BC and CBD combination had significantly (p<0.001) increased Nrf2 antioxidant response element transcription and promoted production of antioxidant enzymes such NQO1 and HO-1. In addition to the in vivo evidence, it was observed that BC and CBD combination significantly (p<0.001) increased Nrf2 mediated signalling in Schwann cells that had been damaged by HG [123]. In order to prevent the protein aggregates and damaged cellular components in neurological diseases, autophagy serves as a neuroprotective strategy [124,125]. Due to the decreased expression of LC3B and Beclin1, that we observed in our studies underscore that the production of autophagosomes was inhibited in hyperglycemic circumstances. Beclin1 aids in the creation of the phagophore and the recruitment of other autophagy-related proteins like LC3B to the phagophore, which results in the formation of autophagosomes, with the aid of the class III PI3K enzyme Vsp34 [126]. Treatment with BC and CBD combination significantly (p<0.001) increased the cleavage of LC3B from LC3B-I and the expression of Beclin1, which is a sign that autophagy was increased. By clearing away the damaged mitochondria, this enhanced autophagosome production further improves mitochondrial function by BC and CBD [87,94,123].
Diabetes and the accompanying hyperglycemia have varied effects on different organ metabolic processes. When a damage sensing mechanism is activated, PINK1, a resident protein of the inner membrane of mitochondria (IMM), begins to build up at the outer mitochondrial membrane (OMM), allowing the removal of unhealthy and dysfunctional mitochondria. In our studies, we observed that BC and CBD combination have shown a significant potential role in autophagy by increasing the PINK1 & PARKIN levels [87,127]. Overall, the results of our current studies have successfully demonstrated the neuroprotective potential of BC and CBD combination in HG induced neurotoxicity and experimental hyperglycemia in rats. By balancing the two opposed processes of mitochondrial biogenesis and autophagy, SIRT1, AMPK and Nrf2 activation by BC and CBD results in effective metabolic sensing related to mitochondrial function (Figure 13). To provide the required bioenergetic needs for enhanced impulse conduction and neurotransmission, the mitochondrial function must be optimized. Additionally, by making nociceptors more sensitive, activating SIRT1 and AMPK lowers oxidative damage, neuroinflammation, and nociceptive firing brought on by inflammatory cytokines. Because SIRT1 and AMPK have a protective impact against the neurodegenerative alterations associated with DN through modification of mitochondrial quality control, they can be taken into consideration while creating cutting-edge strategies for the management of DN.
Conclusion
In summary, the present studies demonstrated that STZ and HG induced oxidative and nitrosative stress play a crucial role in the pathogenesis of diabetic neuropathy. The functional, behavioural, and molecular deficits were due to oxidant- induced damage, neuroinflammation and bioenergetic deficit. These pathological consequences of nerve injury have been attenuated by the combination of CBD and BC in vitro and in vivo (Figure 13). Our findings suggest that enhanced neuroprotective effects of combination therapy may be attributable to simultaneous inhibition of oxidative stress, neuroinflammation and NLRP3 and activation of Nrf2. Hence, the combination therapy could be suggested as a potential strategy that can be further pursued for the management of STZ and HG induced diabetic neuropathy.
Author Contributions
Dr. IK performed studies, evaluated data, prepared figures, and wrote manuscript; Ms. SDK performed studies, evaluated data, prepared figures, and reviewed manuscript, Dr. AKR evaluated data, reviewed manuscript, and wrote manuscript; Ms. BB & MA performed studies, evaluated data, and reviewed manuscript; Dr. MS performed studies, evaluated data, prepared figures and wrote manuscript.
Funding
The National Institute on Minority Health and Health Disparities of National Institutes of Health, Grant/Award Number: U54 MD007582 and Consortium for Medical Marijuana Clinical Outcomes Research (Reference Award number: SUB00002097) provided the funding for carrying out this study. Also Supported, in part, by a MERIT Review Award and Research Career Scientist Award from the US Department of Veterans Affairs (AKR).
Data Availability Statements
The data that support the findings of this study is available in the manuscript as well as supplementary information file. In addition, data can be available from the corresponding author upon reasonable request.
Conflict of Interest Statement
The authors declare no competing interests.
Abbreviations
AMPK- AMP-activated protein kinase
SIRT3- Sirtuin-3
Nrf2- Nuclear factor erythroid 2 [NFE2]- related factor 2
TFAM- Mitochondrial transcription factor A
NFκB- Nuclear factor kappa light chain enhancer of B cells
COX2 - Cyclooxygenase2
p62- Ubiquitin-binding protein p62
PGC-1α- Peroxisome proliferator activated receptor gamma coactivator 1-alpha
DMEM- Dulbecco’s Modified Eagle Medium
DCFDA-2′,7′-dichlorodihydrofluorescein diacetate
Keap1- Kelch-like ECH-associated protein 1
SIRT1- Silent information regulator T1
NQO1- NAD(P)H dehydrogenase [quinone] 1
HO-1- Heme oxygenase-1
Foxo3a- Forkhead Transcription Factor O Subfamily Member 3a
References
- H. Yang, G. Sloan, Y. Ye, S. Wang, B. Duan, S. Tesfaye, L. Gao, New Perspective in Diabetic Neuropathy: From the Periphery to the Brain, a Call for Early Detection, and Precision Medicine, Front. Endocrinol. (Lausanne). 10 (2020) 503723. [CrossRef]
- E.L. Feldman, B.C. Callaghan, R. Pop-Busui, D.W. Zochodne, D.E. Wright, D.L. Bennett, V. Bril, J.W. Russell, V. Viswanathan, Diabetic neuropathy, Nat. Rev. Dis. Prim. 5 (2019) 42. [CrossRef]
- H. Sun, P. Saeedi, S. Karuranga, M. Pinkepank, K. Ogurtsova, B.B. Duncan, C. Stein, A. Basit, J.C.N. Chan, J.C. Mbanya, M.E. Pavkov, A. Ramachandaran, S.H. Wild, S. James, W.H. Herman, P. Zhang, C. Bommer, S. Kuo, E.J. Boyko, D.J. Magliano, IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045, Diabetes Res. Clin. Pract. 183 (2022) 109119. [CrossRef]
- V.K. Arruri, C. Gundu, I. Khan, D.K. Khatri, S.B. Singh, PARP overactivation in neurological disorders, Mol. Biol. Rep. 48 (2021) 2833–2841. [CrossRef]
- M. Rakusa, I. Marolt, Z. Stevic, S.V. Rebrina, T. Milenkovic, A. Stepien, Efficacy of Pregabalin and Duloxetine in Patients with Painful Diabetic Peripheral Neuropathy (PDPN): A Multi-Centre Phase IV Clinical Trial—BLOSSOM, Pharmaceuticals 16 (2023) 1017. [CrossRef]
- P. Fernyhough, Mitochondrial dysfunction in diabetic neuropathy: a series of unfortunate metabolic events, Curr. Diab. Rep. 15 (2015). [CrossRef]
- R. V. Intine, M.P. Sarras, Metabolic Memory and Chronic Diabetes Complications: Potential Role for Epigenetic Mechanisms, Curr. Diab. Rep. 12 (2012) 551. [CrossRef]
- I. Khan, K. Preeti, V. Fernandes, D.K. Khatri, S.B. Singh, Role of MicroRNAs, Aptamers in Neuroinflammation and Neurodegenerative Disorders, Cell. Mol. Neurobiol. 2021 427 42 (2021) 2075–2095. [CrossRef]
- Y. Yang, Y. Liu, Y. Wang, Y. Chao, J. Zhang, Y. Jia, J. Tie, D. Hu, Regulation of SIRT1 and Its Roles in Inflammation, Front. Immunol. 13 (2022) 831168. [CrossRef]
- K. Vargas-Ortiz, V. Pérez-Vázquez, M.H. Macías-Cervantes, Exercise and Sirtuins: A Way to Mitochondrial Health in Skeletal Muscle, Int. J. Mol. Sci. 20 (2019). [CrossRef]
- Y. Majeed, N. Halabi, A.Y. Madani, R. Engelke, A.M. Bhagwat, H. Abdesselem, M. V. Agha, M. Vakayil, R. Courjaret, N. Goswami, H. Ben Hamidane, M.A. Elrayess, A. Rafii, J. Graumann, F. Schmidt, N.A. Mazloum, SIRT1 promotes lipid metabolism and mitochondrial biogenesis in adipocytes and coordinates adipogenesis by targeting key enzymatic pathways, Sci. Reports 2021 111 11 (2021) 1–19. [CrossRef]
- I. Khan, K. Preeti, R. Kumar, D. Kumar Khatri, S. Bala Singh, Piceatannol promotes neuroprotection by inducing mitophagy and mitobiogenesis in the experimental diabetic peripheral neuropathy and hyperglycemia-induced neurotoxicity, Int. Immunopharmacol. 116 (2023) 109793. [CrossRef]
- Y.C. Chuang, S. Der Chen, S. Bin Jou, T.K. Lin, S.F. Chen, N.C. Chen, C.Y. Hsu, Sirtuin 1 Regulates Mitochondrial Biogenesis and Provides an Endogenous Neuroprotective Mechanism Against Seizure-Induced Neuronal Cell Death in the Hippocampus Following Status Epilepticus, Int. J. Mol. Sci. 20 (2019). [CrossRef]
- F. Villavicencio Tejo, R.A. Quintanilla, Contribution of the Nrf2 Pathway on Oxidative Damage and Mitochondrial Failure in Parkinson and Alzheimer’s Disease, Antioxidants 10 (2021). [CrossRef]
- H.-R. Li, Q. Liu, C.-L. Zhu, X.-Y. Sun, C.-Y. Sun, C.-M. Yu, P. Li, X.-M. Deng, J.-F. Wang, Î2-Nicotinamide mononucleotide activates NAD+/SIRT1 pathway and attenuates inflammatory and oxidative responses in the hippocampus regions of septic mice, Redox Biol. 63 (2023) 102745. [CrossRef]
- A.P. Rolo, C.M. Palmeira, Diabetes and mitochondrial function: role of hyperglycemia and oxidative stress, Toxicol. Appl. Pharmacol. 212 (2006) 167–178. [CrossRef]
- J.W. Russell, D. Golovoy, A.M. Vincent, P. Mahendru, J.A. Olzmann, A. Mentzer, E.L. Feldman, High glucose-induced oxidative stress and mitochondrial dysfunction in neurons, FASEB J. 16 (2002) 1738–1748. [CrossRef]
- A. Loboda, M. Damulewicz, E. Pyza, A. Jozkowicz, J. Dulak, Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: an evolutionarily conserved mechanism, Cell. Mol. Life Sci. 73 (2016) 3221. [CrossRef]
- H. Chen, J. Deng, H. Gao, Y. Song, Y. Zhang, J. Sun, J. Zhai, Involvement of the SIRT1-NLRP3 pathway in the inflammatory response, Cell Commun. Signal. 21 (2023) 185. [CrossRef]
- Y. Li, P. Wang, X. Yang, W. Wang, J. Zhang, Y. He, W. Zhang, T. Jing, B. Wang, R. Lin, SIRT1 inhibits inflammatory response partly through regulation of NLRP3 inflammasome in vascular endothelial cells, Mol. Immunol. 77 (2016) 148–156. [CrossRef]
- G.E.S. Batiha, H.M. Al-kuraishy, A.I. Al-Gareeb, E. Elekhnawy, SIRT1 pathway in Parkinson’s disease: a faraway snapshot but so close, Inflammopharmacology 31 (2023) 37. [CrossRef]
- X. Li, Y. Feng, X.X. Wang, D. Truong, Y.C. Wu, The Critical Role of SIRT1 in Parkinson’s Disease: Mechanism and Therapeutic Considerations, Aging Dis. 11 (2020) 1608. [CrossRef]
- M. Entezari, D. Hashemi, A. Taheriazam, A. Zabolian, S. Mohammadi, F. Fakhri, M. Hashemi, K. Hushmandi, M. Ashrafizadeh, A. Zarrabi, Y.N. Ertas, S. Mirzaei, S. Samarghandian, AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: A pre-clinical and clinical investigation, Biomed. Pharmacother. 146 (2022) 112563. [CrossRef]
- N.B. Ruderman, X.J. Xu, L. Nelson, J.M. Cacicedo, A.K. Saha, F. Lan, Y. Ido, AMPK and SIRT1: A long-standing partnership?, Am. J. Physiol. - Endocrinol. Metab. 298 (2010) 751–760. [CrossRef]
- K. Huang, X. Luo, Y. Zhong, L. Deng, J. Feng, New insights into the role of melatonin in diabetic cardiomyopathy, Pharmacol. Res. Perspect. 10 (2022) e00904. [CrossRef]
- M.I. Martić-Kehl, R. Schibli, P.A. Schubiger, Can animal data predict human outcome? Problems and pitfalls of translational animal research, Eur. J. Nucl. Med. Mol. Imaging 39 (2012) 1492–1496. [CrossRef]
- N. Shanks, R. Greek, J. Greek, Are animal models predictive for humans?, Philos. Ethics. Humanit. Med. 4 (2009). [CrossRef]
- M.B. Bracken, Why animal studies are often poor predictors of human reactions to exposure, J. R. Soc. Med. 102 (2009) 120. [CrossRef]
- D.E. Ingber, Human organs-on-chips for disease modelling, drug development and personalized medicine, Nat. Rev. Genet. 2022 238 23 (2022) 467–491. [CrossRef]
- A.F. Gileta, C.J. Fitzpatrick, A.S. Chitre, C.L. St. Pierre, E. V. Joyce, R.J. Maguire, A.M. McLeod, N.M. Gonzales, A.E. Williams, J.D. Morrow, T.E. Robinson, S.B. Flagel, A.A. Palmer, Genetic characterization of outbred Sprague Dawley rats and utility for genome-wide association studies, PLoS Genet. 18 (2022). [CrossRef]
- V.G. Yerra, A. Kumar, Adenosine Monophosphate-Activated Protein Kinase Abates Hyperglycaemia-Induced Neuronal Injury in Experimental Models of Diabetic Neuropathy: Effects on Mitochondrial Biogenesis, Autophagy and Neuroinflammation, Mol. Neurobiol. 54 (2017) 2301–2312. [CrossRef]
- V.G. Yerra, A.K. Kalvala, A. Kumar, Isoliquiritigenin reduces oxidative damage and alleviates mitochondrial impairment by SIRT1 activation in experimental diabetic neuropathy, J. Nutr. Biochem. 47 (2017) 41–52. [CrossRef]
- A. Höke, Animal Models of Peripheral Neuropathies, Neurotherapeutics 9 (2012) 262. [CrossRef]
- R. Singh, S.A. Farooq, A. Mannan, T.G. Singh, A. Najda, Z. Grażyna, G.M. Albadrani, A.A. Sayed, M.M. Abdel-Daim, Animal models of diabetic microvascular complications: Relevance to clinical features, Biomed. Pharmacother. 145 (2022). [CrossRef]
- S. Green, M.R. Dietrich, S. Leonelli, R.A. Ankeny, “Extreme” organisms and the problem of generalization: interpreting the Krogh principle, Hist. Philos. Life Sci. 40 (2018). [CrossRef]
- Y.C. Cheng, L.W. Chu, J.Y. Chen, S.L. Hsieh, Y.C. Chang, Z.K. Dai, B.N. Wu, Loganin Attenuates High Glucose-Induced Schwann Cells Pyroptosis by Inhibiting ROS Generation and NLRP3 Inflammasome Activation, Cells 9 (2020) 1–18. [CrossRef]
- J. Li, R. Guan, L. Pan, Mechanism of Schwann cells in diabetic peripheral neuropathy: A review, Medicine (Baltimore). 102 (2023) E32653. [CrossRef]
- A.P. Mizisin, Mechanisms of diabetic neuropathy: Schwann cells, Handb. Clin. Neurol. 126 (2014) 401–428. [CrossRef]
- N.P. Gonçalves, C.B. Vægter, H. Andersen, L. Østergaard, N.A. Calcutt, T.S. Jensen, Schwann cell interactions with axons and microvessels in diabetic neuropathy, Nat. Rev. Neurol. 2017 133 13 (2017) 135–147. [CrossRef]
- H.M. Hashiesh, M.F. Nagoor Meeran, C. Sharma, B. Sadek, J. Al Kaabi, S.K. Ojha, Therapeutic Potential of β-Caryophyllene: A Dietary Cannabinoid in Diabetes and Associated Complications, Nutr. 2020, Vol. 12, Page 2963 12 (2020) 2963. [CrossRef]
- B. Horvth, P. Mukhopadhyay, G. Hask, P. Pacher, The Endocannabinoid System and Plant-Derived Cannabinoids in Diabetes and Diabetic Complications, Am. J. Pathol. 180 (2012) 432. [CrossRef]
- Z. Atakan, Cannabis, a complex plant: different compounds and different effects on individuals, Ther. Adv. Psychopharmacol. 2 (2012) 241. [CrossRef]
- V. Desaulniers Brousseau, B. Sen Wu, S. MacPherson, V. Morello, M. Lefsrud, Cannabinoids and Terpenes: How Production of Photo-Protectants Can Be Manipulated to Enhance Cannabis sativa L. Phytochemistry, Front. Plant Sci. 12 (2021) 620021. [CrossRef]
- H. Blanton, L. Yin, J. Duong, K. Benamar, Cannabidiol and Beta-Caryophyllene in Combination: A Therapeutic Functional Interaction, Int. J. Mol. Sci. 23 (2022) 15470. [CrossRef]
- B. Donertas, C. Cengelli Unel, K. Erol, Cannabinoids and agmatine as potential therapeutic alternatives for cisplatin-induced peripheral neuropathy, J. Exp. Pharmacol. 10 (2018) 19–28. [CrossRef]
- J. Zhang, C. Lin, S. Jin, H. Wang, Y. Wang, X. Du, M.R. Hutchinson, H. Zhao, L. Fang, X. Wang, The pharmacology and therapeutic role of cannabidiol in diabetes, Exploration 3 (2023). [CrossRef]
- R. Verma, F. Hoda, M. Arshad, A. Iqubal, A.N. Siddiqui, M.A. Khan, S.E. Haque, M. Akhtar, A.K. Najmi, Cannabis, a Miracle Drug with Polyvalent Therapeutic Utility: Preclinical and Clinical-Based Evidence, Med. Cannabis Cannabinoids 4 (2021) 43. [CrossRef]
- D. An, S. Peigneur, L.A. Hendrickx, J. Tytgat, Targeting Cannabinoid Receptors: Current Status and Prospects of Natural Products, Int. J. Mol. Sci. 21 (2020) 1–33. [CrossRef]
- H.M. Hashiesh, M.F. Nagoor Meeran, C. Sharma, B. Sadek, J. Al Kaabi, S.K. Ojha, Therapeutic Potential of β-Caryophyllene: A Dietary Cannabinoid in Diabetes and Associated Complications, Nutrients 12 (2020) 1–30. [CrossRef]
- E.P. Baron, P. Lucas, J. Eades, O. Hogue, Patterns of medicinal cannabis use, strain analysis, and substitution effect among patients with migraine, headache, arthritis, and chronic pain in a medicinal cannabis cohort, J. Headache Pain 2018 191 19 (2018) 1–28. [CrossRef]
- M.S. Wallace, T.D. Marcotte, A. Umlauf, B. Gouaux, J.H. Atkinson, Efficacy of Inhaled Cannabis on Painful Diabetic Neuropathy, J. Pain 16 (2015) 616–627. [CrossRef]
- J. Zhang, C. Lin, S. Jin, H. Wang, Y. Wang, X. Du, M.R. Hutchinson, H. Zhao, L. Fang, X. Wang, The pharmacology and therapeutic role of cannabidiol in diabetes, Exploration 3 (2023) 20230047. [CrossRef]
- D.S. Aguilar-Ávila, M.E. Flores-Soto, C. Tapia-Vázquez, O.A. Pastor-Zarandona, R.I. López-Roa, J.M. Viveros-Paredes, β-Caryophyllene, a Natural Sesquiterpene, Attenuates Neuropathic Pain and Depressive-Like Behavior in Experimental Diabetic Mice, J. Med. Food 22 (2019) 460–468. [CrossRef]
- H. Li, D. Wang, Y. Chen, M. Yang, β-Caryophyllene inhibits high glucose-induced oxidative stress, inflammation and extracellular matrix accumulation in mesangial cells, Int. Immunopharmacol. 84 (2020) 106556. [CrossRef]
- H. Blanton, L. Yin, J. Duong, K. Benamar, Cannabidiol and Beta-Caryophyllene in Combination: A Therapeutic Functional Interaction, Int. J. Mol. Sci. 23 (2022). [CrossRef]
- K.M. King, A.M. Myers, A.J. Soroka-Monzo, R.F. Tuma, R.J. Tallarida, E.A. Walker, S.J. Ward, Single and combined effects of Δ9-tetrahydrocannabinol and cannabidiol in a mouse model of chemotherapy-induced neuropathic pain, Br. J. Pharmacol. 174 (2017) 2832. [CrossRef]
- Glucose in Cell Culture, (n.d.). https://www.sigmaaldrich.com/US/en/technical-documents/technical-article/cell-culture-and-cell-culture-analysis/mammalian-cell-culture/glucose (accessed February 22, 2024).
- S. Clement, S.S. Braithwaite, M.F. Magee, A. Ahmann, E.P. Smith, R.G. Schafer, I.B. Hirsh, Management of Diabetes and Hyperglycemia in Hospitals, Diabetes Care 27 (2004) 553–591. [CrossRef]
- S. Yadranji Aghdam, Z. Gurel, A. Ghaffarieh, C.M. Sorenson, N. Sheibani, High glucose and diabetes modulate cellular proteasome function: Implications in the pathogenesis of diabetes complications, Biochem. Biophys. Res. Commun. 432 (2013) 339–344. [CrossRef]
- W. Wang, Z. Chai, M.E. Cooper, P.Z. Zimmet, H. Guo, J. Ding, F. Yang, X. Chen, X. Lin, K. Zhang, Q. Zhong, Z. Li, P. Zhang, Z. Wu, X. Guan, L. Zhang, K. He, High Fasting Blood Glucose Level With Unknown Prior History of Diabetes Is Associated With High Risk of Severe Adverse COVID-19 Outcome, Front. Endocrinol. (Lausanne). 12 (2021). [CrossRef]
- Y. pu Liu, S. jin Shao, H. dong Guo, Schwann cells apoptosis is induced by high glucose in diabetic peripheral neuropathy, Life Sci. 248 (2020) 117459. [CrossRef]
- L.Q. Sun, Y.Y. Chen, X. Wang, X.J. Li, B. Xue, L. Qu, T.T. Zhang, Y.M. Mu, J.M. Lu, The protective effect of Alpha lipoic acid on Schwann cells exposed to constant or intermittent high glucose, Biochem. Pharmacol. 84 (2012) 961–973. [CrossRef]
- Q. Li, Y. Jiao, Y. Yu, G. Wang, Y. Yu, Hydrogen-rich medium alleviates high glucose-induced oxidative stress and parthanatos in rat Schwann cells in vitro, Mol. Med. Rep. 19 (2019) 338. [CrossRef]
- W.D. Pallas, E.S. Pak, J.L. Hannan, In vitro high glucose increases apoptosis, decreases nerve outgrowth, and promotes survival of sympathetic pelvic neurons, Sex. Med. 11 (2023) 1–7. [CrossRef]
- A.K. Kalvala, R. Nimma, A. Bagde, S.K. Surapaneni, N. Patel, P. Arthur, L. Sun, R. Singh, N. Kommineni, A. Nathani, Y. Li, M. Singh, The role of Cannabidiol and tetrahydrocannabivarin to overcome doxorubicin resistance in MDA-MB-231 xenografts in athymic nude mice, Biochimie 208 (2023) 19–30. [CrossRef]
- A. Sood, V. Fernandes, K. Preeti, M. Khot, D.K. Khatri, S.B. Singh, Fingolimod Alleviates Cognitive Deficit in Type 2 Diabetes by Promoting Microglial M2 Polarization via the pSTAT3-jmjd3 Axis, Mol. Neurobiol. 60 (2023) 901–922. [CrossRef]
- H. Kim, X. Xue, Detection of Total Reactive Oxygen Species in Adherent Cells by 2′,7′-Dichlorodihydrofluorescein Diacetate Staining, J. Vis. Exp. 2020 (2020) 1–5. [CrossRef]
- J. Luo, Y. Xiang, X. Xu, D. Fang, D. Li, F. Ni, X. Zhu, B. Chen, M. Zhou, High Glucose-Induced ROS Production Stimulates Proliferation of Pancreatic Cancer via Inactivating the JNK Pathway, Oxid. Med. Cell. Longev. 2018 (2018). [CrossRef]
- A. Kumar Kalvala, A. Bagde, P. Arthur, S. Kumar Surapaneni, N. Ramesh, A. Nathani, M. Singh, Role of Cannabidiol and Tetrahydrocannabivarin on Paclitaxel-induced neuropathic pain in rodents, Int. Immunopharmacol. 107 (2022). [CrossRef]
- P. Bheereddy, V.G. Yerra, A.K. Kalvala, B. Sherkhane, A. Kumar, SIRT1 Activation by Polydatin Alleviates Oxidative Damage and Elevates Mitochondrial Biogenesis in Experimental Diabetic Neuropathy, Cell. Mol. Neurobiol. (2020). [CrossRef]
- S. Gunes, Z. He, R. Malone, P.J. Cullen, J.F. Curtin, J. Curtin, Platinum nanoparticles inhibit intracellular ROS generation and protect against Cold Atmospheric Plasma-induced cytotoxicity, BioRxiv (2021) 2021.02.18.431888. [CrossRef]
- K. Preeti, V. Fernandes, A. Sood, I. Khan, D.K. Khatri, S.B. Singh, Necrostatin-1S mitigates type-2 diabetes-associated cognitive decrement and lipotoxicity-induced neuro-microglia changes through p-RIPK-RIPK3-p-MLKL axis, Metab. Brain Dis. 38 (2023) 1581–1612. [CrossRef]
- S.K. Surapaneni, N. Patel, L. Sun, N. Kommineni, A.K. Kalvala, A. Gebeyehu, P. Arthur, L.C. Duke, R. Nimma, D. G Meckes, M. Singh, Anticancer and chemosensitization effects of cannabidiol in 2D and 3D cultures of TNBC: involvement of GADD45α, integrin-α5, -β5, -β1, and autophagy, Drug Deliv. Transl. Res. 12 (2022) 2762. [CrossRef]
- I. Khan, K. Preeti, R. Kumar, D.K. Khatri, S.B. Singh, Activation of SIRT1 by silibinin improved mitochondrial health and alleviated the oxidative damage in experimental diabetic neuropathy and high glucose-mediated neurotoxicity, Arch. Physiol. Biochem. 0 (2020) 1–17. [CrossRef]
- A.K. Kalvala, A. Bagde, P. Arthur, T. Kulkarni, S. Bhattacharya, S. Surapaneni, N.K. Patel, R. Nimma, A. Gebeyehu, N. Kommineni, D.G. Meckes, L. Sun, B. Banjara, K. Mosley-Kellum, T.C. Dinh, M. Singh, Cannabidiol-Loaded Extracellular Vesicles from Human Umbilical Cord Mesenchymal Stem Cells Alleviate Paclitaxel-Induced Peripheral Neuropathy, Pharmaceutics 15 (2023). [CrossRef]
- K. Preeti, A. Sood, V. Fernandes, I. Khan, D.K. Khatri, S.B. Singh, Experimental Type 2 diabetes and lipotoxicity-associated neuroinflammation involve mitochondrial DNA-mediated cGAS/STING axis: implication of Type-1 interferon response in cognitive impairment, Mol. Neurobiol. (2024) 1–28. [CrossRef]
- A. Nathani, L. Sun, I. Khan, M. Aare, A. Bagde, Y. Li, M. Singh, Combined Role of Interleukin-15 Stimulated Natural Killer Cell-Derived Extracellular Vesicles and Carboplatin in Osimertinib-Resistant H1975 Lung Cancer Cells with EGFR Mutations, Pharm. 2024, Vol. 16, Page 83 16 (2024) 83. [CrossRef]
- N.P. du Sert, V. Hurst, A. Ahluwalia, S. Alam, M.T. Avey, M. Baker, W.J. Browne, A. Clark, I.C. Cuthill, U. Dirnagl, M. Emerson, P. Garner, S.T. Holgate, D.W. Howells, N.A. Karp, S.E. Lazic, K. Lidster, C.J. MacCallum, M. Macleod, E.J. Pearl, O.H. Petersen, F. Rawle, P. Reynolds, K. Rooney, E.S. Sena, S.D. Silberberg, T. Steckler, H. Würbel, The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research, PLoS Biol. 18 (2020). [CrossRef]
- A. Kumar, G. Negi, S.S. Sharma, Suppression of NF-κB and NF-κB regulated oxidative stress and neuroinflammation by BAY 11-7082 (IκB phosphorylation inhibitor) in experimental diabetic neuropathy, Biochimie 94 (2012) 1158–1165. [CrossRef]
- V.G. Yerra, A.K. Kalvala, A. Kumar, Isoliquiritigenin reduces oxidative damage and alleviates mitochondrial impairment by SIRT1 activation in experimental diabetic neuropathy, J. Nutr. Biochem. 47 (2017) 41–52. [CrossRef]
- A.K. Kalvala, V.G. Yerra, B. Sherkhane, C. Gundu, V. Arruri, R. Kumar, A. Kumar, Chronic hyperglycemia impairs mitochondrial unfolded protein response and precipitates proteotoxicity in experimental diabetic neuropathy: focus on LonP1 mediated mitochondrial regulation, Pharmacol. Reports 2020 726 72 (2020) 1627–1644. [CrossRef]
- H. Blanton, L. Yin, J. Duong, K. Benamar, Cannabidiol and Beta-Caryophyllene in Combination: A Therapeutic Functional Interaction, Int. J. Mol. Sci. 23 (2022). [CrossRef]
- A. Areti, P. Komirishetty, M. Akuthota, R.A. Malik, A. Kumar, Melatonin prevents mitochondrial dysfunction and promotes neuroprotection by inducing autophagy during oxaliplatin-evoked peripheral neuropathy, J. Pineal Res. 62 (2017) e12393. [CrossRef]
- R.M.P. Campos, A.F.L. Aguiar, Y. Paes-Colli, P.M.P. Trindade, B.K. Ferreira, R.A. de Melo Reis, L.S. Sampaio, Cannabinoid Therapeutics in Chronic Neuropathic Pain: From Animal Research to Human Treatment, Front. Physiol. 12 (2021) 785176. [CrossRef]
- J.M. Quintero, G. Pulido, L.F. Giraldo, M.X. Leon, L.E. Diaz, R.H. Bustos, A Systematic Review on Cannabinoids for Neuropathic Pain Administered by Routes Other than Oral or Inhalation, Plants 11 (2022). [CrossRef]
- J. Mlost, M. Bryk, K. Starowicz, Cannabidiol for Pain Treatment: Focus on Pharmacology and Mechanism of Action, Int. J. Mol. Sci. 21 (2020) 1–22. [CrossRef]
- Z. Wang, P. Zheng, X. Chen, Y. Xie, K. Weston-Green, N. Solowij, Y.L. Chew, X.F. Huang, Cannabidiol induces autophagy and improves neuronal health associated with SIRT1 mediated longevity, GeroScience 44 (2022) 1505–1524. [CrossRef]
- B. Ni, Y. Liu, M. Dai, J. Zhao, Y. Liang, X. Yang, B. Han, M. Jiang, The role of cannabidiol in aging, Biomed. Pharmacother. 165 (2023) 115074. [CrossRef]
- P. Bheereddy, V.G. Yerra, A.K. Kalvala, B. Sherkhane, A. Kumar, SIRT1 Activation by Polydatin Alleviates Oxidative Damage and Elevates Mitochondrial Biogenesis in Experimental Diabetic Neuropathy, Cell. Mol. Neurobiol. 41 (2021) 1563–1577. [CrossRef]
- A.K. Kalvala, I. Khan, C. Gundu, A. Kumar, An Overview on ATP Dependent and Independent Proteases Including an Anterograde to Retrograde Control on Mitochondrial Function; Focus on Diabetes and Diabetic Complications, Curr. Pharm. Des. 25 (2019) 2584–2594. [CrossRef]
- G.J. Biessels, V. Bril, N.A. Calcutt, N.E. Cameron, M.A. Cotter, R. Dobrowsky, E.L. Feldman, P. Fernyhough, J. Jakobsen, R.A. Malik, A.P. Mizisin, P.J. Oates, I.G. Obrosova, R. Pop-Busui, J.W. Russell, A.A. Sima, M.J. Stevens, R.E. Schmidt, S. Tesfaye, A. Veves, A.I. Vinik, D.E. Wright, S. Yagihashi, M.A. Yorek, D. Ziegler, D.W. Zochodne, Phenotyping animal models of diabetic neuropathy: a consensus statement of the diabetic neuropathy study group of the EASD (Neurodiab), J. Peripher. Nerv. Syst. 19 (2014) 77. [CrossRef]
- D.H. Xu, B.D. Cullen, M. Tang, Y. Fang, The Effectiveness of Topical Cannabidiol Oil in Symptomatic Relief of Peripheral Neuropathy of the Lower Extremities, Curr. Pharm. Biotechnol. 21 (2020) 390–402. [CrossRef]
- E. Schulz, S. Schuhmacher, T. Münzel, When metabolism rules perfusion: AMPK-mediated endothelial nitric oxide synthase activation, Circ. Res. 104 (2009) 422–424. [CrossRef]
- V.G. Yerra, A. Kumar, Adenosine Monophosphate-Activated Protein Kinase Abates Hyperglycaemia-Induced Neuronal Injury in Experimental Models of Diabetic Neuropathy: Effects on Mitochondrial Biogenesis, Autophagy and Neuroinflammation, Mol. Neurobiol. 54 (2017) 2301–2312. [CrossRef]
- K.A. Weikel, N.B. Ruderman, J.M. Cacicedo, Unraveling the Actions of AMP-activated Protein Kinase in Metabolic Diseases: Systemic to Molecular Insights, Metabolism. 65 (2016) 634. [CrossRef]
- C. Rodríguez, M. Muñoz, C. Contreras, D. Prieto, AMPK, metabolism, and vascular function, FEBS J. 288 (2021) 3746–3771. [CrossRef]
- M.C.N. Sack, T. Finkel, Mitochondrial metabolism, sirtuins, and aging, Cold Spring Harb. Perspect. Biol. 4 (2012). [CrossRef]
- Y. Li, L. Wang, G. Zhang, X. Qiao, M. Zhang, SIRT1 Mediates Neuropathic Pain Induced by Sciatic Nerve Chronic Constrictive Injury in the VTA-NAc Pathway, Pain Res. Manag. 2020 (2020). [CrossRef]
- Y.Q. Zhou, W. Mei, X.B. Tian, Y.K. Tian, D.Q. Liu, D.W. Ye, The therapeutic potential of Nrf2 inducers in chronic pain: Evidence from preclinical studies, Pharmacol. Ther. 225 (2021). [CrossRef]
- F.H. Song, D.Q. Liu, Y.Q. Zhou, W. Mei, SIRT1: A promising therapeutic target for chronic pain, CNS Neurosci. Ther. 28 (2022) 818–828. [CrossRef]
- K.C. Chang, P.F. Liu, C.H. Chang, Y.C. Lin, Y.J. Chen, C.W. Shu, The interplay of autophagy and oxidative stress in the pathogenesis and therapy of retinal degenerative diseases, Cell Biosci. 12 (2022) 1–20. [CrossRef]
- W. Ornatowski, Q. Lu, M. Yegambaram, A.E. Garcia, E.A. Zemskov, E. Maltepe, J.R. Fineman, T. Wang, S.M. Black, Complex interplay between autophagy and oxidative stress in the development of pulmonary disease, Redox Biol. 36 (2020). [CrossRef]
- Y.L. Wu, Z.J. Lin, C.C. Li, X. Lin, S.K. Shan, B. Guo, M.H. Zheng, F. Li, L.Q. Yuan, Z. hong Li, Epigenetic regulation in metabolic diseases: mechanisms and advances in clinical study, Signal Transduct. Target. Ther. 2023 81 8 (2023) 1–27. [CrossRef]
- G.F.S. Fernandes, G.D.B. Silva, A.R. Pavan, D.E. Chiba, C.M. Chin, J.L. Dos Santos, Epigenetic Regulatory Mechanisms Induced by Resveratrol, Nutrients 9 (2017). [CrossRef]
- A. Salminen, J.M.T. Hyttinen, K. Kaarniranta, AMP-activated protein kinase inhibits NF-κB signaling and inflammation: impact on healthspan and lifespan, J. Mol. Med. (Berl). 89 (2011) 667–676. [CrossRef]
- A.K. Kalvala, A. Bagde, P. Arthur, T. Kulkarni, S. Bhattacharya, S. Surapaneni, N.K. Patel, R. Nimma, A. Gebeyehu, N. Kommineni, D.G. Meckes, L. Sun, B. Banjara, K. Mosley-Kellum, T.C. Dinh, M. Singh, Cannabidiol-Loaded Extracellular Vesicles from Human Umbilical Cord Mesenchymal Stem Cells Alleviate Paclitaxel-Induced Peripheral Neuropathy, Pharm. 2023, Vol. 15, Page 554 15 (2023) 554. [CrossRef]
- S. Atalay Ekiner, A. Gęgotek, E. Skrzydlewska, The molecular activity of cannabidiol in the regulation of Nrf2 system interacting with NF-κB pathway under oxidative stress, Redox Biol. 57 (2022) 102489. [CrossRef]
- R. Prakash, N. Kumari, A.J. Siddiqui, A.Q. Khan, M.A. Khan, R. Khan, R. Haque, A.A. Robertson, J. Boltze, S.S. Raza, MCC950 Regulates Stem Cells Destiny Through Modulating SIRT3-NLRP3 Inflammasome Dynamics During Oxygen Glucose Deprivation, Stem Cell Rev. Reports 19 (2023) 1415–1426. [CrossRef]
- T. Zhang, Z. Fang, K.G. Linghu, J. Liu, L. Gan, L. Lin, Small molecule-driven SIRT3-autophagy-mediated NLRP3 inflammasome inhibition ameliorates inflammatory crosstalk between macrophages and adipocytes, Br. J. Pharmacol. 177 (2020) 4645–4665. [CrossRef]
- M.L. Chen, X.H. Zhu, L. Ran, H.D. Lang, L. Yi, M.T. Mi, Trimethylamine-N-Oxide Induces Vascular Inflammation by Activating the NLRP3 Inflammasome Through the SIRT3-SOD2-mtROS Signaling Pathway, J. Am. Heart Assoc. 6 (2017). [CrossRef]
- K. Schroder, J. Tschopp, The Inflammasomes, Cell 140 (2010) 821–832. [CrossRef]
- R. Zhou, A.S. Yazdi, P. Menu, J. Tschopp, A role for mitochondria in NLRP3 inflammasome activation, Nat. 2010 4697329 469 (2010) 221–225. [CrossRef]
- F. Marín-Aguilar, B. Castejón-Vega, E. Alcocer-Gómez, D. Lendines-Cordero, M.A. Cooper, P. De La Cruz, E. Andújar-Pulido, M. Pérez-Alegre, J. Muntané, A.J. Pérez-Pulido, B. Ryffel, A.A.B. Robertson, J. Ruiz-Cabello, P. Bullón, M.D. Cordero, NLRP3 Inflammasome Inhibition by MCC950 in Aged Mice Improves Health via Enhanced Autophagy and PPARα Activity, J. Gerontol. A. Biol. Sci. Med. Sci. 75 (2020) 1457–1464. [CrossRef]
- W. Chen, X. Wang, Q. Sun, Y. Zhang, J. Liu, T. Hu, W. Wu, C. Wei, M. Liu, Y. Ding, D. Liu, Y. Chong, P. Wang, H. Zhu, W. Cui, J. Zhang, Q. Li, F. Yang, The upregulation of NLRP3 inflammasome in dorsal root ganglion by ten-eleven translocation methylcytosine dioxygenase 2 (TET2) contributed to diabetic neuropathic pain in mice, J. Neuroinflammation 19 (2022) 1–19. [CrossRef]
- Y. Bai, Q. Mu, X. Bao, J. Zuo, X. Fang, J. Hua, D. Zhang, G. Jiang, P. Li, S. Gao, D. Zhao, Targeting NLRP3 Inflammasome in the Treatment Of Diabetes and Diabetic Complications: Role of Natural Compounds from Herbal Medicine, Aging Dis. 12 (2021) 1587. [CrossRef]
- Y. qun Zhou, D. qiang Liu, S. ping Chen, N. Chen, J. Sun, X. mei Wang, F. Cao, Y. ke Tian, D. wei Ye, Nrf2 activation ameliorates mechanical allodynia in paclitaxel-induced neuropathic pain, Acta Pharmacol. Sin. 41 (2020) 1041–1048. [CrossRef]
- N. Chen, M.M. Ge, D.Y. Li, X.M. Wang, D.Q. Liu, D.W. Ye, Y.K. Tian, Y.Q. Zhou, J.P. Chen, β2-adrenoreceptor agonist ameliorates mechanical allodynia in paclitaxel-induced neuropathic pain via induction of mitochondrial biogenesis, Biomed. Pharmacother. 144 (2021). [CrossRef]
- J. Sun, J.Y. Li, L.Q. Zhang, D.Y. Li, J.Y. Wu, S.J. Gao, D.Q. Liu, Y.Q. Zhou, W. Mei, Nrf2 Activation Attenuates Chronic Constriction Injury-Induced Neuropathic Pain via Induction of PGC-1 α-Mediated Mitochondrial Biogenesis in the Spinal Cord, Oxid. Med. Cell. Longev. 2021 (2021). [CrossRef]
- J. Sun, F.H. Song, J.Y. Wu, L.Q. Zhang, D.Y. Li, S.J. Gao, D.Q. Liu, Y.Q. Zhou, W. Mei, Sestrin2 overexpression attenuates osteoarthritis pain via induction of AMPK/PGC-1α-mediated mitochondrial biogenesis and suppression of neuroinflammation, Brain. Behav. Immun. 102 (2022) 53–70. [CrossRef]
- G. Negi, A. Kumar, R.P. Joshi, S.S. Sharma, Oxidative stress and Nrf2 in the pathophysiology of diabetic neuropathy: old perspective with a new angle, Biochem. Biophys. Res. Commun. 408 (2011) 1–5. [CrossRef]
- G. Negi, A. Kumar, S. S. Sharma, Nrf2 and NF-κB modulation by sulforaphane counteracts multiple manifestations of diabetic neuropathy in rats and high glucose-induced changes, Curr. Neurovasc. Res. 8 (2011) 294–304. [CrossRef]
- G. Negi, A. Kumar, S.S. Sharma, Melatonin modulates neuroinflammation and oxidative stress in experimental diabetic neuropathy: effects on NF-κB and Nrf2 cascades, Undefined 50 (2011) 124–131. [CrossRef]
- L. Yang, R. Rozenfeld, D. Wu, L.A. Devi, Z. Zhang, A. Cederbaum, Cannabidiol protects liver from binge alcohol-induced steatosis by mechanisms including inhibition of oxidative stress and increase in autophagy, Free Radic. Biol. Med. 68 (2014) 260–267. [CrossRef]
- W. Xu, U. Ocak, L. Gao, S. Tu, C.J. Lenahan, J. Zhang, A. Shao, Selective autophagy as a therapeutic target for neurological diseases, Cell. Mol. Life Sci. 78 (2021) 1369. [CrossRef]
- O. Corti, K. Blomgren, A. Poletti, P.M. Beart, Autophagy in neurodegeneration: New insights underpinning therapy for neurological diseases, J. Neurochem. 154 (2020) 354–371. [CrossRef]
- R. Kang, H.J. Zeh, M.T. Lotze, D. Tang, The Beclin 1 network regulates autophagy and apoptosis, Cell Death Differ. 18 (2011) 571. [CrossRef]
- G. Jîtcă, B.E. Ősz, C.E. Vari, C.M. Rusz, A. Tero-Vescan, A. Pușcaș, Cannabidiol: Bridge between Antioxidant Effect, Cellular Protection, and Cognitive and Physical Performance, Antioxidants 2023, Vol. 12, Page 485 12 (2023) 485. [CrossRef]
Figure 1.
Experimental design. There were 5 groups of healthy male SD rats. Single dose of STZ 55 mg/kg, i.p. was given to the SD rats and BC (30 mg/kg, i.p), CBD (15 mg/kg, i.p.) & BC+CBD (30 mg/kg, i.p&15 mg/kg, i.p. respectively) were given for last 3 weeks. Normal control SD rats (NC, n=6), STZ induced SD rats (STZ, n=8), STZ induced SD rats with BC (STZ+BC, n=6) and STZ induced SD rats with CBD (STZ+CBD, n=6) and STZ induced SD rats with BC & CBD (STZ+BC+CBD, n=6), and after that functional and behavioural parameters were recorded.
Figure 1.
Experimental design. There were 5 groups of healthy male SD rats. Single dose of STZ 55 mg/kg, i.p. was given to the SD rats and BC (30 mg/kg, i.p), CBD (15 mg/kg, i.p.) & BC+CBD (30 mg/kg, i.p&15 mg/kg, i.p. respectively) were given for last 3 weeks. Normal control SD rats (NC, n=6), STZ induced SD rats (STZ, n=8), STZ induced SD rats with BC (STZ+BC, n=6) and STZ induced SD rats with CBD (STZ+CBD, n=6) and STZ induced SD rats with BC & CBD (STZ+BC+CBD, n=6), and after that functional and behavioural parameters were recorded.
Figure 2.
Effect of BC, CBD & their combination on cell viability. BC and CBD has shown significant toxicity at above 200 µM and 20 µM concentrations respectively. Schwann cells were given treatment at different concentrations of CBD (100-3.125 µM) and BC (500-15.625 µM) & BC+CBD (8-0.25 µM) respectively. Data values are represented as Mean±SEM (n=3). ***p < 0.001, **p < 0.01, Vs Normal control.
Figure 2.
Effect of BC, CBD & their combination on cell viability. BC and CBD has shown significant toxicity at above 200 µM and 20 µM concentrations respectively. Schwann cells were given treatment at different concentrations of CBD (100-3.125 µM) and BC (500-15.625 µM) & BC+CBD (8-0.25 µM) respectively. Data values are represented as Mean±SEM (n=3). ***p < 0.001, **p < 0.01, Vs Normal control.
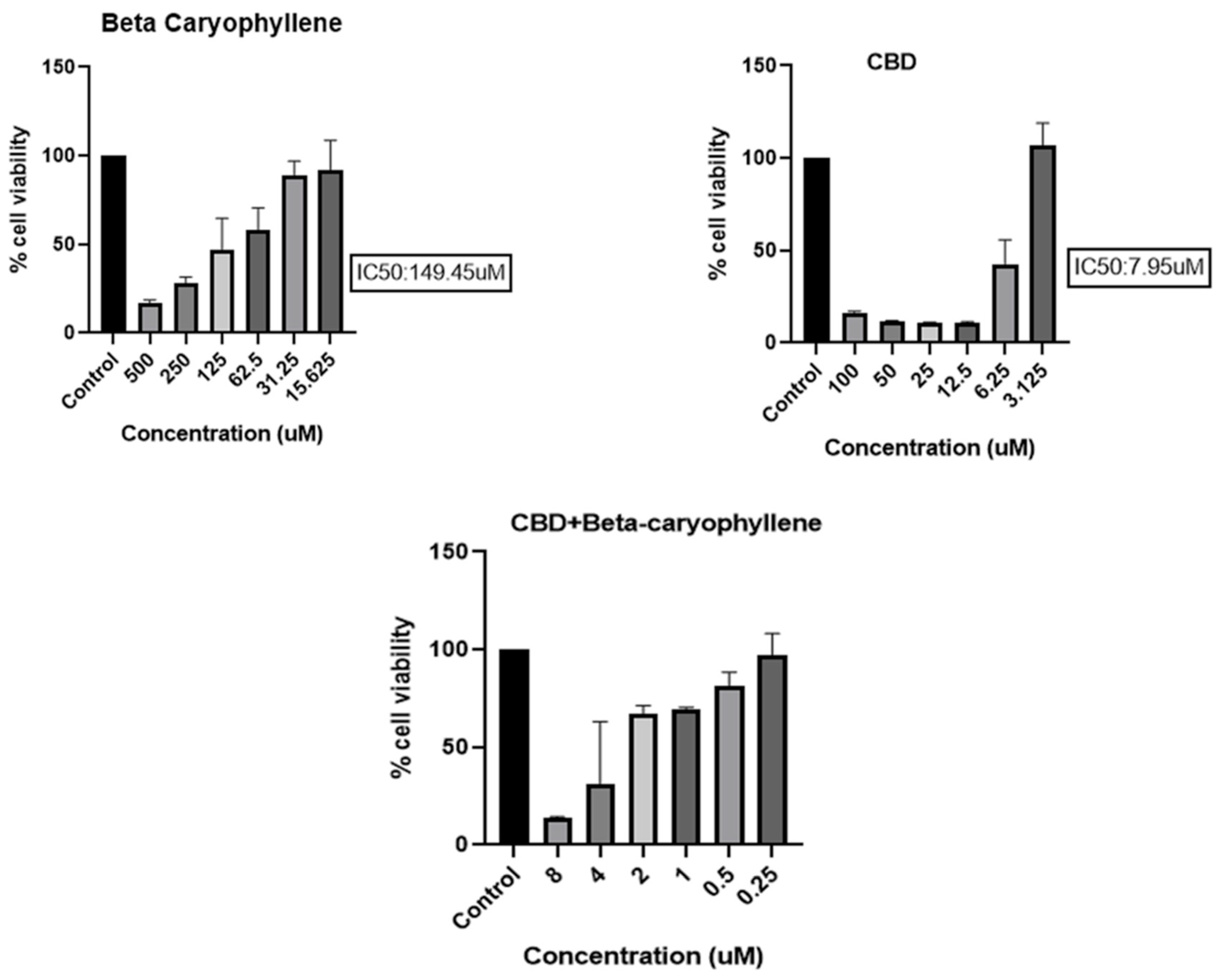
Figure 3.
Effect of BC, CBD and their combination on ROS, superoxide, and mitochondrial membrane potential in HG induced Schwann cells. (A, B), Fluorescent microscopic images and bar graph of Schwann cells showing the reactive oxygen species. (C, D) Fluorescence microscopic images and bar graph of Schwann cells indicating mitochondrial membrane potential. (E, F) Fluorescence microscopic images and bar graph of Schwann cells indicating mitochondrial superoxide generation. Photographs were captured at 40x magnification. All the data values are represented as Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01 Vs HG, ***p < 0.001 Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.
Figure 3.
Effect of BC, CBD and their combination on ROS, superoxide, and mitochondrial membrane potential in HG induced Schwann cells. (A, B), Fluorescent microscopic images and bar graph of Schwann cells showing the reactive oxygen species. (C, D) Fluorescence microscopic images and bar graph of Schwann cells indicating mitochondrial membrane potential. (E, F) Fluorescence microscopic images and bar graph of Schwann cells indicating mitochondrial superoxide generation. Photographs were captured at 40x magnification. All the data values are represented as Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01 Vs HG, ***p < 0.001 Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.

Figure 4.
BC, CBD and their combination effect on mitochondrial biogenesis and antioxidant effect through the expression of AMPK, SIRT1 and NRF2 in HG induced Schwann cells. (A) Western blots of respective proteins AMPK, SIRT1, NRF2, PGC-1α, HO1, SOD2, NQO1 and β-actin. (B) The densitometric analysis of respective blots. All the data values are represented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs HG, ***p < 0.001 Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.
Figure 4.
BC, CBD and their combination effect on mitochondrial biogenesis and antioxidant effect through the expression of AMPK, SIRT1 and NRF2 in HG induced Schwann cells. (A) Western blots of respective proteins AMPK, SIRT1, NRF2, PGC-1α, HO1, SOD2, NQO1 and β-actin. (B) The densitometric analysis of respective blots. All the data values are represented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs HG, ***p < 0.001 Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.
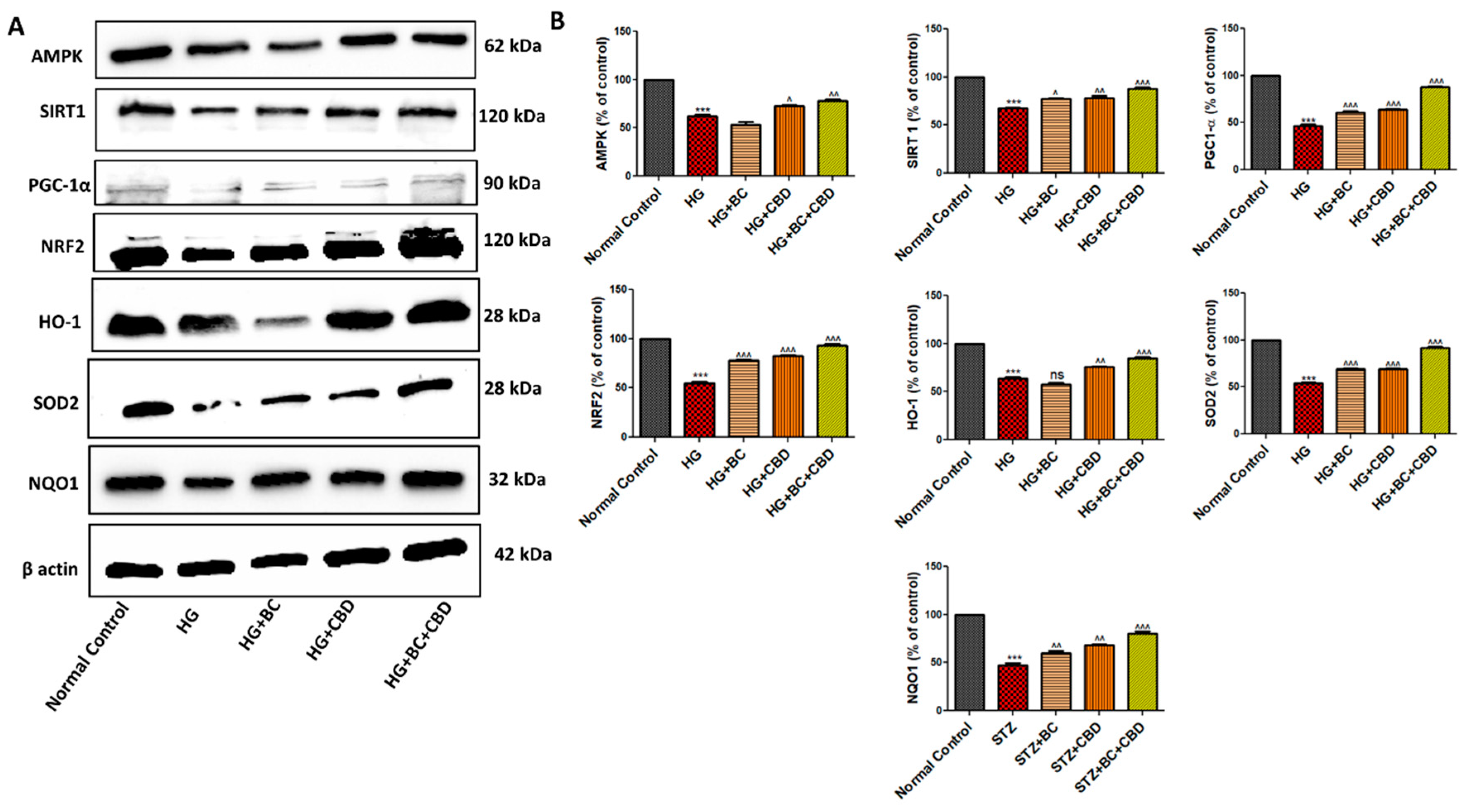
Figure 5.
BC, CBD and their combination effect on mitochondrial function and autophagy in HG induced Schwann cells. (A) The western blotting of SIRT3, PHB2, PARKIN, PINK1, pmTOR, p62, LC3B and β-actin (B) The densitometric analysis of SIRT3, PHB2, PARKIN, PINK1, pmTOR, p62, LC3B level in Schwann cells. All the data values were presented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05, Vs HG, ***p < 0.001, **p < 0.01 Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.
Figure 5.
BC, CBD and their combination effect on mitochondrial function and autophagy in HG induced Schwann cells. (A) The western blotting of SIRT3, PHB2, PARKIN, PINK1, pmTOR, p62, LC3B and β-actin (B) The densitometric analysis of SIRT3, PHB2, PARKIN, PINK1, pmTOR, p62, LC3B level in Schwann cells. All the data values were presented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05, Vs HG, ***p < 0.001, **p < 0.01 Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.
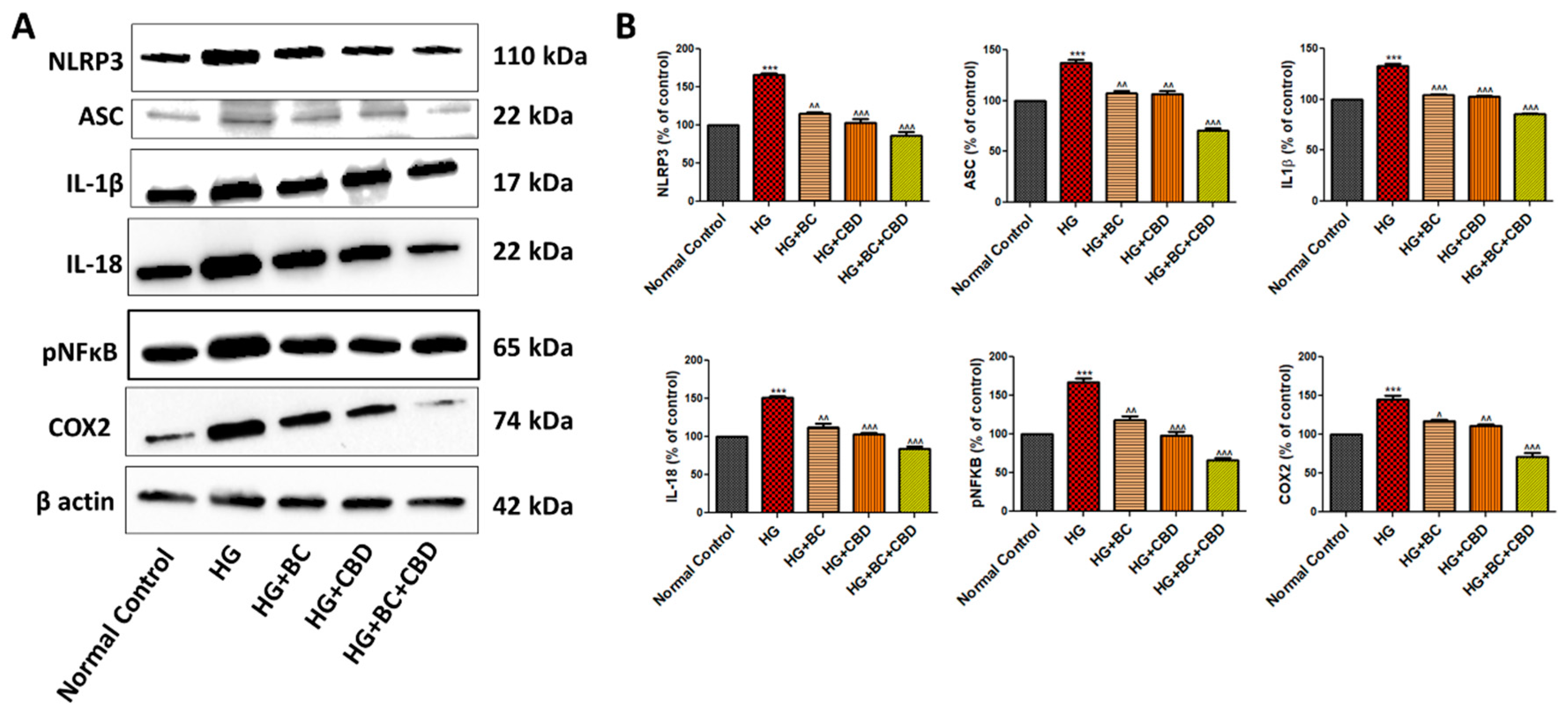
Figure 6.
BC, CBD, and their combination effect on neuroinflammation in HG induced Schwann cells. (A) the western blotting of NLRP3, ASC, IL1β, IL18, NFKB, COX2 and β-actin (B) the densitometric analysis of NLRP3, ASC, IL-1β, IL18, NFkB, COX2 expression in Schwann cells. All the data values were expressed in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs HG, ***p < 0.001, Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.
Figure 6.
BC, CBD, and their combination effect on neuroinflammation in HG induced Schwann cells. (A) the western blotting of NLRP3, ASC, IL1β, IL18, NFKB, COX2 and β-actin (B) the densitometric analysis of NLRP3, ASC, IL-1β, IL18, NFkB, COX2 expression in Schwann cells. All the data values were expressed in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs HG, ***p < 0.001, Vs NC. NC: Normal Schwann cells, HG: Schwann cells exposed to high glucose (30 mM), HG+BC: HG induced cells treated with BC 75 µM, HG+CBD: HG induced cells treated with CBD 8 µM and HG+BC+CBD: High glucose induced cells treated BC & CBD with 75µM and 3.64 µM respectively.
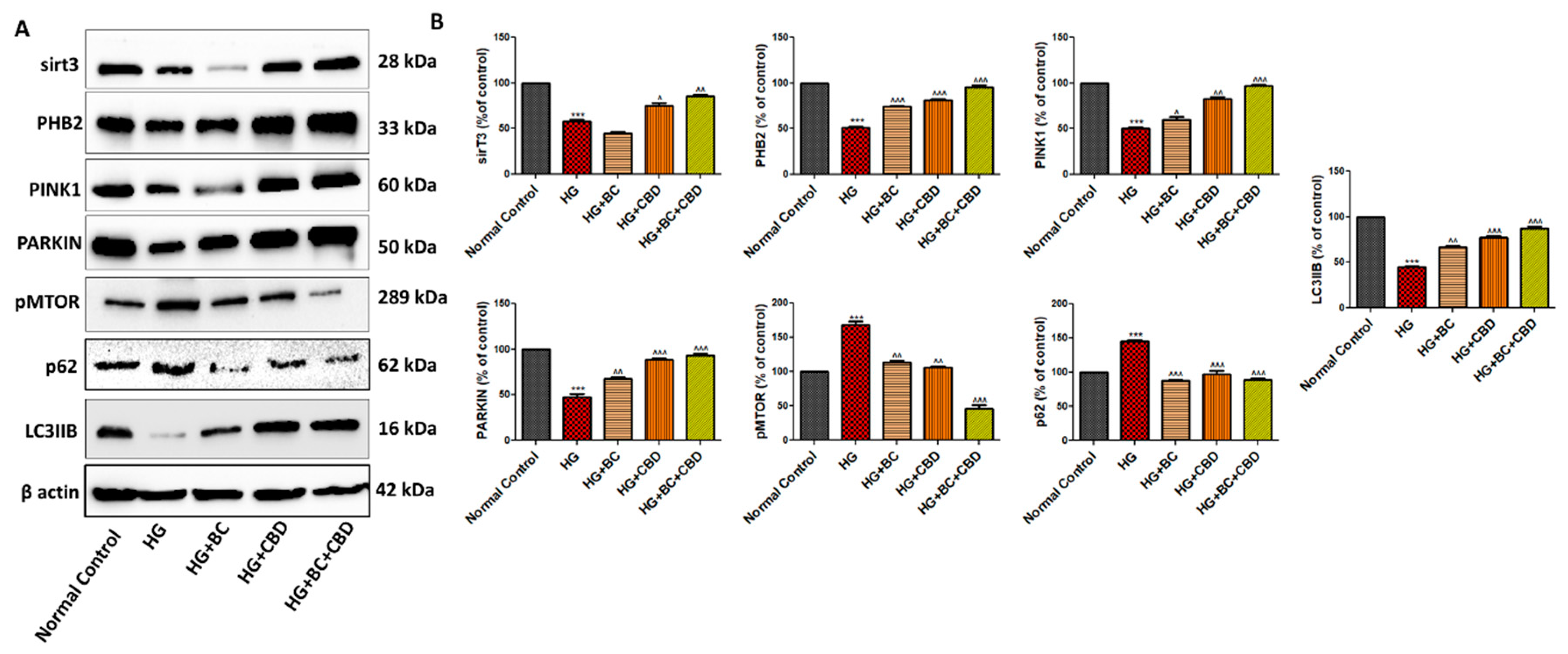
Figure 7.
BC, CBD, and their combination effect on thermal stimuli in diabetic rats. The thermal stimuli (A) cold immersion test (B) hot immersion test and (C) Hargreaves test, (n=6). All the data values are presented in the form of Mean±SEM. ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001 Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
Figure 7.
BC, CBD, and their combination effect on thermal stimuli in diabetic rats. The thermal stimuli (A) cold immersion test (B) hot immersion test and (C) Hargreaves test, (n=6). All the data values are presented in the form of Mean±SEM. ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001 Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
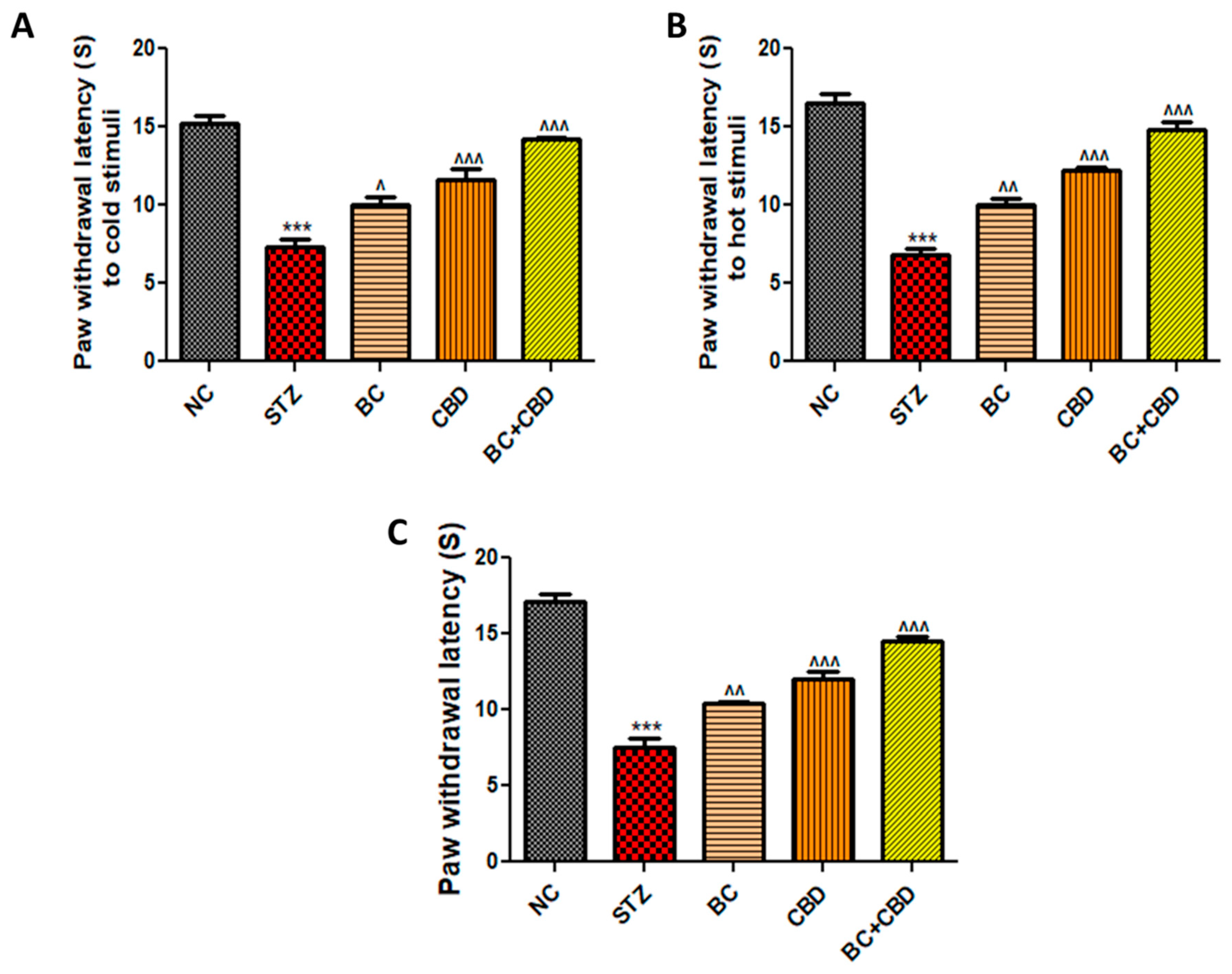
Figure 8.
BC, CBD and their combination effect on mechanical stimuli and nerve function in diabetic rats. The mechanical stimuli, (A) Von Frey filaments test (B) Randall- Selitto apparatus test (n=6) (C) Nerve blood flow test (NBF), (n=3). All the data values are presented in the form of Mean±SEM. ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001 Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
Figure 8.
BC, CBD and their combination effect on mechanical stimuli and nerve function in diabetic rats. The mechanical stimuli, (A) Von Frey filaments test (B) Randall- Selitto apparatus test (n=6) (C) Nerve blood flow test (NBF), (n=3). All the data values are presented in the form of Mean±SEM. ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001 Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
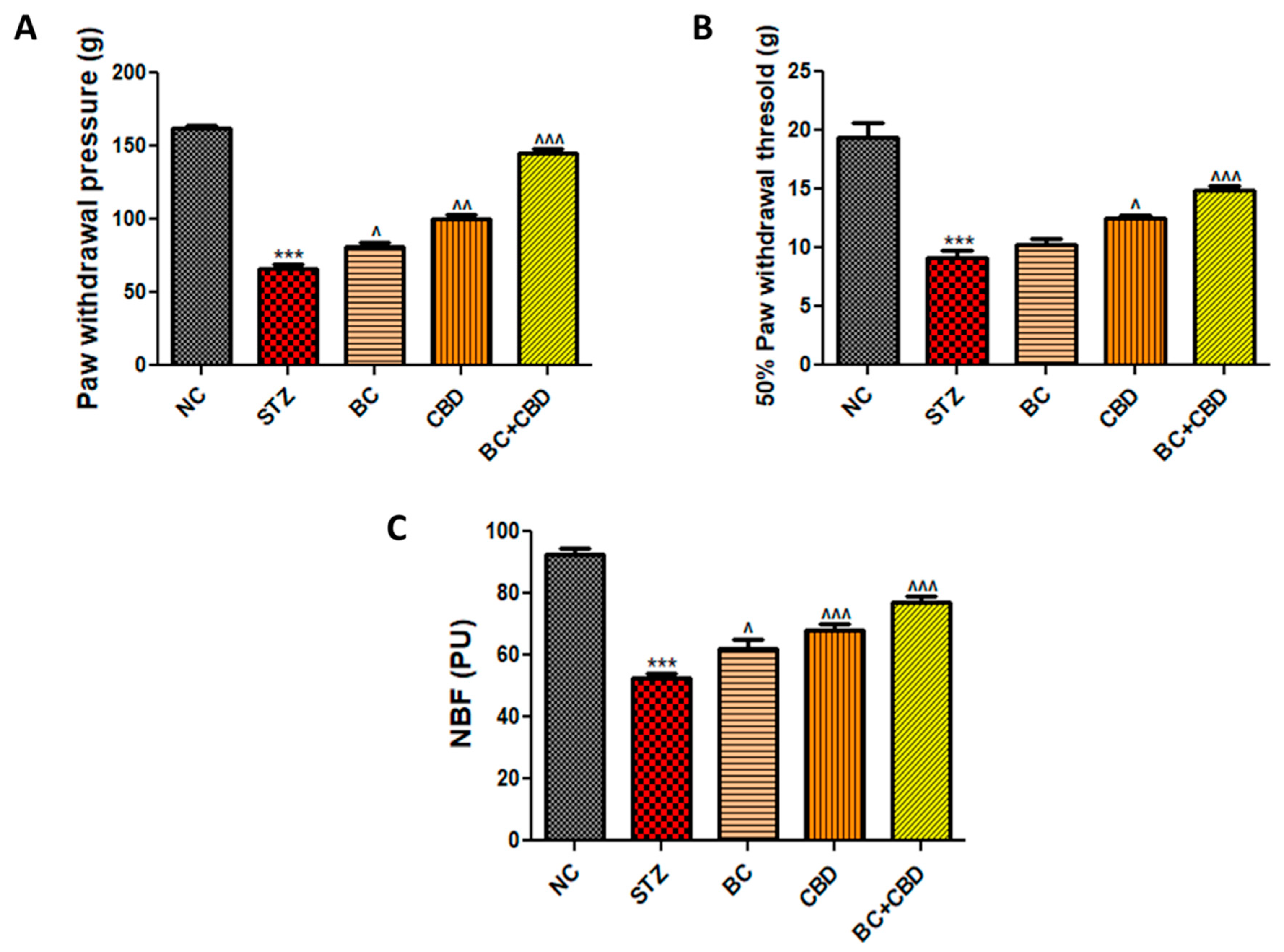
Figure 9.
BC, CBD and their combination effect on AMPK, SIRT1-mediated mitochondrial biogenesis in SD rats. (A) The expression of SOD2 immunopositivity in the SD rat’s sciatic nerve. (B) The bar graph of respective images. Photographs were taken at 40X magnification. (C) The western blot of AMPK, SIRT1, NRF1, PGC-1α, TFAM, NQO1, SOD2, HO1 and β-actin in the sciatic nerve of diabetic rats. (D) The corresponding graphical representation of blot by densitometric analysis. All the data values are represented in the form of Mean±SEM (N=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
Figure 9.
BC, CBD and their combination effect on AMPK, SIRT1-mediated mitochondrial biogenesis in SD rats. (A) The expression of SOD2 immunopositivity in the SD rat’s sciatic nerve. (B) The bar graph of respective images. Photographs were taken at 40X magnification. (C) The western blot of AMPK, SIRT1, NRF1, PGC-1α, TFAM, NQO1, SOD2, HO1 and β-actin in the sciatic nerve of diabetic rats. (D) The corresponding graphical representation of blot by densitometric analysis. All the data values are represented in the form of Mean±SEM (N=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
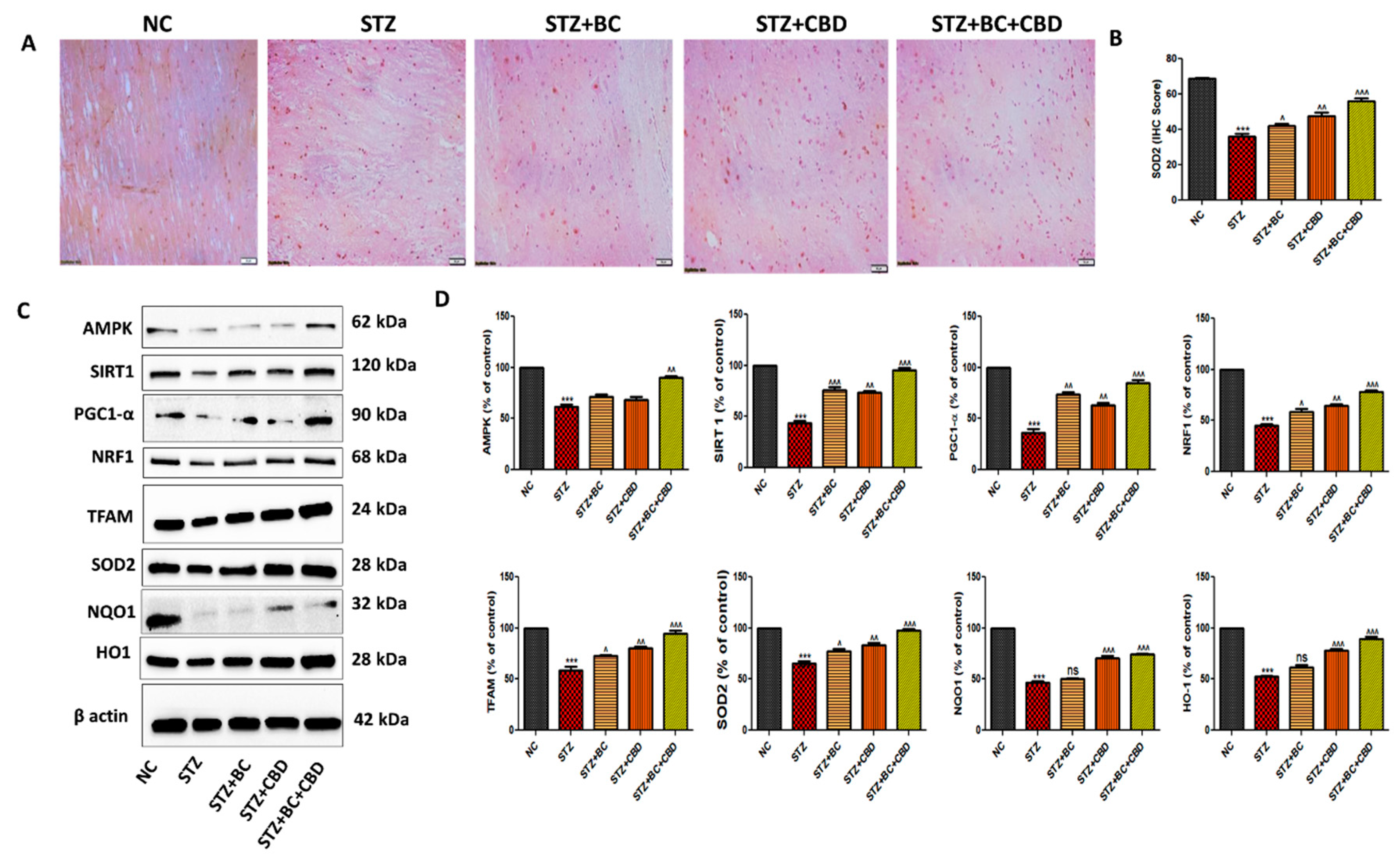
Figure 10.
BC, CBD and their combination effect on inflammasome and Nrf2 linked antioxidant defense in SD rats. (A) The representative immunohistochemical representation of NFkB immunopositivity in sciatic nerve of rats. (B) The bar graph of respective images. Photographs were captured at 40X magnification. (C) The representative western blot of NLRP3, IL-18, COX2, NFkB, Nrf2, keap1, Foxo3a & β-actin in the diabetic rats. (D) The corresponding graphical representation of blot by densitometric analysis. All the data values are represented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
Figure 10.
BC, CBD and their combination effect on inflammasome and Nrf2 linked antioxidant defense in SD rats. (A) The representative immunohistochemical representation of NFkB immunopositivity in sciatic nerve of rats. (B) The bar graph of respective images. Photographs were captured at 40X magnification. (C) The representative western blot of NLRP3, IL-18, COX2, NFkB, Nrf2, keap1, Foxo3a & β-actin in the diabetic rats. (D) The corresponding graphical representation of blot by densitometric analysis. All the data values are represented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
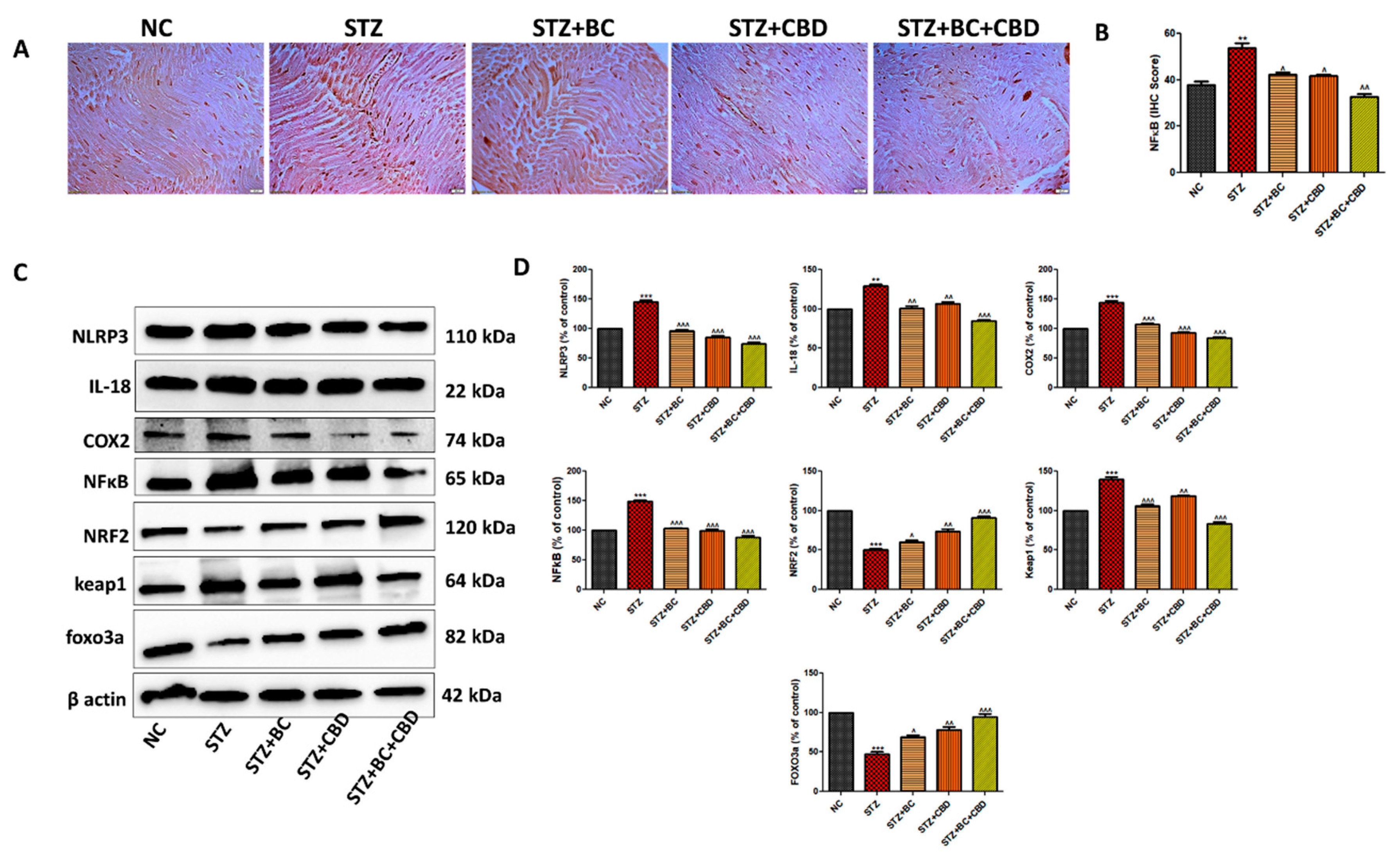
Figure 11.
BC, CBD, and their combination effect on autophagy in SD rats. (A) The expression of LC3B immunopositivity in sciatic nerve of rats. (B) The bar graph for respective photographs. Photographs were taken at 40X magnification. (C) The representative western blot of SirT3, PINK1, PARKIN, p62, LC3B, pMTOR, atg3, atg7, BCL2 and β-actin in diabetic rats. (D) The corresponding graphical representation of blot by densitometric analysis. All the data values are represented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
Figure 11.
BC, CBD, and their combination effect on autophagy in SD rats. (A) The expression of LC3B immunopositivity in sciatic nerve of rats. (B) The bar graph for respective photographs. Photographs were taken at 40X magnification. (C) The representative western blot of SirT3, PINK1, PARKIN, p62, LC3B, pMTOR, atg3, atg7, BCL2 and β-actin in diabetic rats. (D) The corresponding graphical representation of blot by densitometric analysis. All the data values are represented in the form of Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
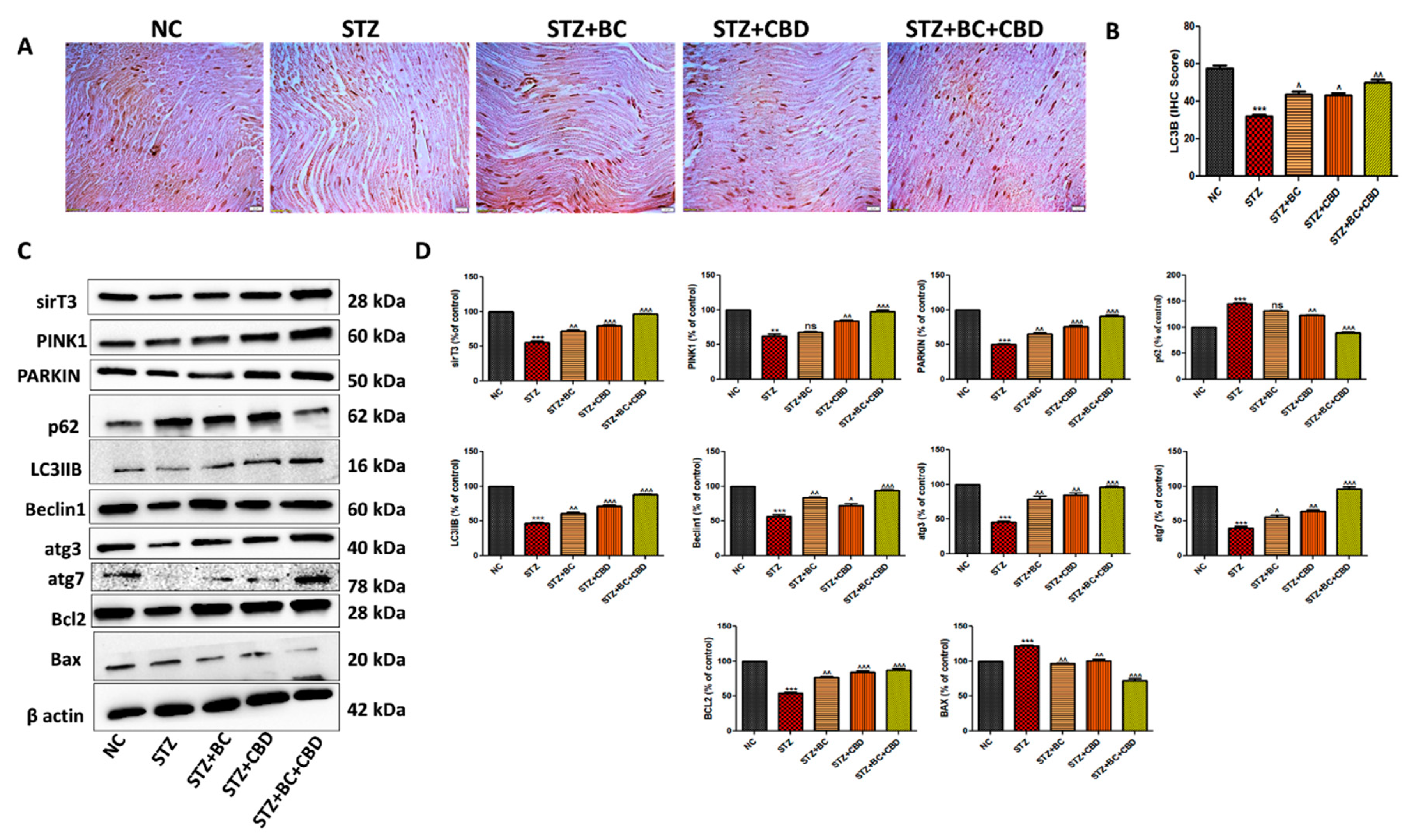
Figure 12.
BC, CBD, and their combination effect on IENF degeneration in SD rats. (A) The representative immunohistochemistry images of PGP 9.5 in the diabetic rat planter skin. The photographs were captured at 40x magnification. (B) The Bar graph shows IENF/mm. The results are represented as Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
Figure 12.
BC, CBD, and their combination effect on IENF degeneration in SD rats. (A) The representative immunohistochemistry images of PGP 9.5 in the diabetic rat planter skin. The photographs were captured at 40x magnification. (B) The Bar graph shows IENF/mm. The results are represented as Mean±SEM (n=3). ^^^p < 0.001, ^^p < 0.01, ^p < 0.05 Vs STZ, ***p < 0.001, Vs NC. NC: Normal control, STZ: Diabetic control, STZ+BC: Diabetic control treated with BC 30 mg/kg, STZ+CBD: Diabetic control treated with CBD 15 mg/kg, and STZ+BC+CBD: Diabetic control treated with BC & CBD at 30 mg/kg and 15 mg/kg body wt, respectively.
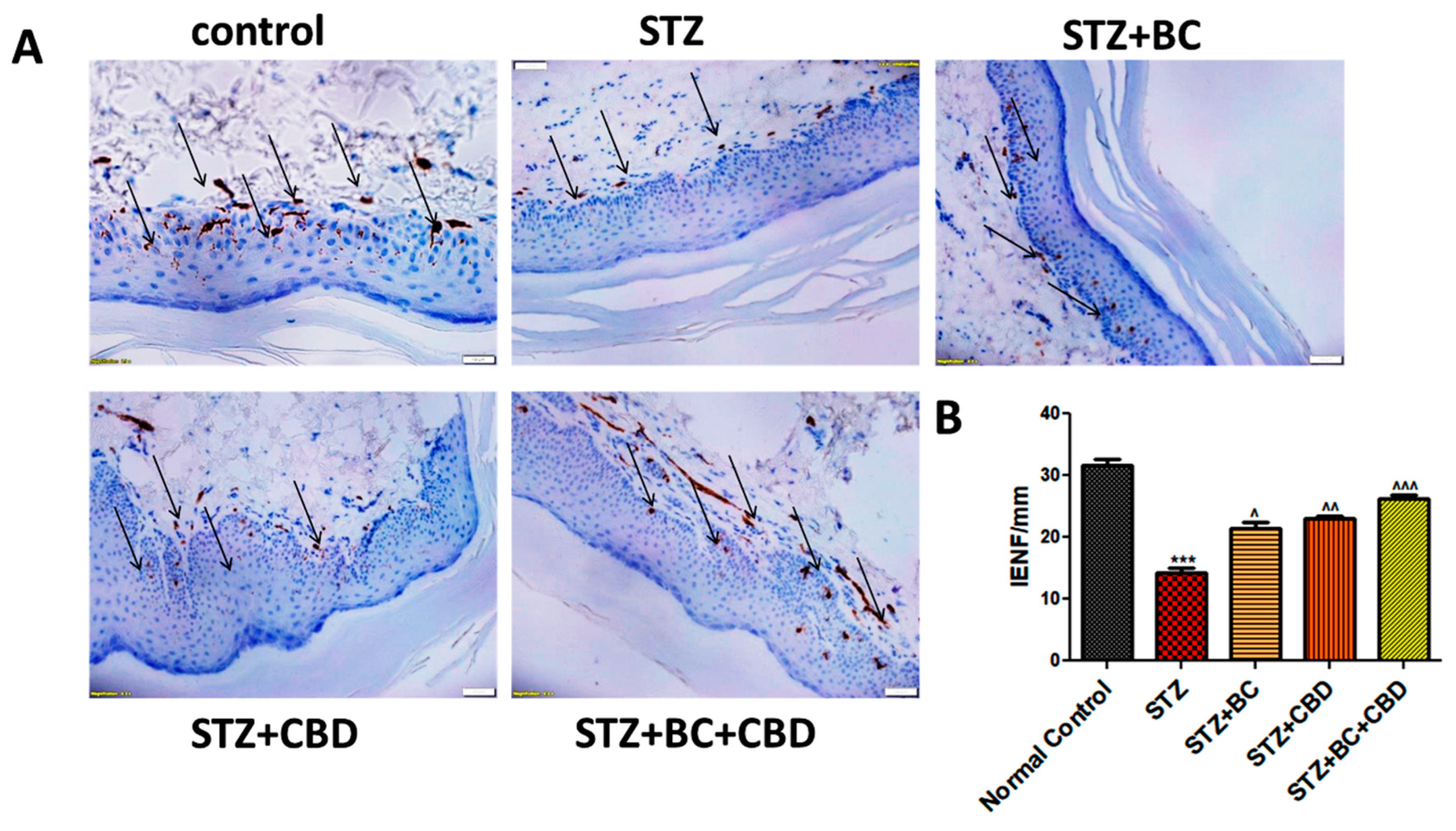
Figure 13.
Hypothesized mechanism of BC and CBD via AMPK/SIRT1/Nrf2 activation and NLRP3 Inactivation. Peripheral nerve may accumulate more glucose and ROS because of diabetes associated oxidative stress/nitrosative stress and dysfunctional mitochondria, which act as priming and activation signals for the activation of NLRP3 and release of IL-1β. On the other side, NLRP3 activation comes from abnormal clearance of damaged mitochondria caused by defective autophagy, which failed to balance mitochondrial homeostasis. To combat the NLRP3 linked neuroinflammation that is thought to be responsible for the onset and maintenance of diabetic neuropathy, BC and CBD, which are antioxidants, enhance Nrf2 mediated p62 induction and improve both autophagy and antioxidant effect. On the other activation of AMPK, SIRT1 Nrf2 and the inhibition of mTOR, by BC and CBD promote autophagy. PGC-1α deacetylation and associated transcriptional activation of NRF1 mediated mitochondriogenesis are both outcomes of BC and CBD facilitating SIRT1. Additionally, ROS linked neuroinflammation and oxidative stress can be prevented by activating AMPK and SIRT1. Under hyperglycemic conditions of DN, these AMPK & SIRT1-mediated impacts on redox balance and mitochondrial homeostasis can avoid neuronal damage. Overall, the ROS directed mitochondrial dysfunction and oxidative stress within the cellular system are reduced by the newly synthesized healthy mitochondria and via autophagy. (AMPK: AMP-activated protein kinase, SIRT1: Silent mating type information regulation 2 homolog, PGC-1α: Peroxisome proliferator-activated receptor-γ coactivator, Nrf2: Nuclear respiratory factor 2, p62: Sequestome1, TFAM: Transcription factor A, mTOR: Mammalian target of rapamycin).
Figure 13.
Hypothesized mechanism of BC and CBD via AMPK/SIRT1/Nrf2 activation and NLRP3 Inactivation. Peripheral nerve may accumulate more glucose and ROS because of diabetes associated oxidative stress/nitrosative stress and dysfunctional mitochondria, which act as priming and activation signals for the activation of NLRP3 and release of IL-1β. On the other side, NLRP3 activation comes from abnormal clearance of damaged mitochondria caused by defective autophagy, which failed to balance mitochondrial homeostasis. To combat the NLRP3 linked neuroinflammation that is thought to be responsible for the onset and maintenance of diabetic neuropathy, BC and CBD, which are antioxidants, enhance Nrf2 mediated p62 induction and improve both autophagy and antioxidant effect. On the other activation of AMPK, SIRT1 Nrf2 and the inhibition of mTOR, by BC and CBD promote autophagy. PGC-1α deacetylation and associated transcriptional activation of NRF1 mediated mitochondriogenesis are both outcomes of BC and CBD facilitating SIRT1. Additionally, ROS linked neuroinflammation and oxidative stress can be prevented by activating AMPK and SIRT1. Under hyperglycemic conditions of DN, these AMPK & SIRT1-mediated impacts on redox balance and mitochondrial homeostasis can avoid neuronal damage. Overall, the ROS directed mitochondrial dysfunction and oxidative stress within the cellular system are reduced by the newly synthesized healthy mitochondria and via autophagy. (AMPK: AMP-activated protein kinase, SIRT1: Silent mating type information regulation 2 homolog, PGC-1α: Peroxisome proliferator-activated receptor-γ coactivator, Nrf2: Nuclear respiratory factor 2, p62: Sequestome1, TFAM: Transcription factor A, mTOR: Mammalian target of rapamycin).
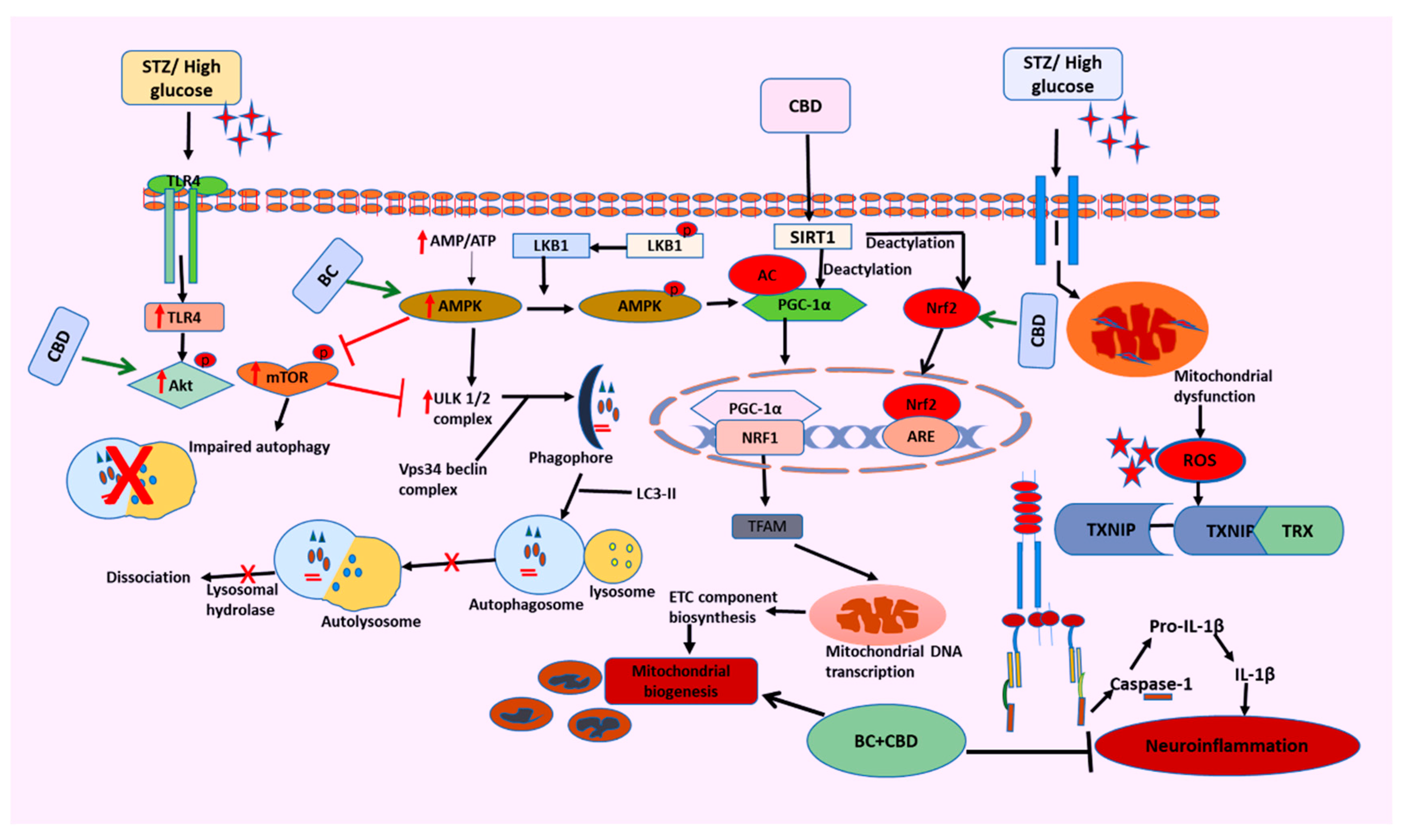
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



