
Submitted:
28 April 2024
Posted:
29 April 2024
You are already at the latest version
Abstract
Marshes are an important ecosystem, acting as biodiversity hotspot, a carbon sink as well as a bioremediation site, breaking down anthropogenic waste such as antibiotics, metals and fertilizers. For participating in these metabolic activities and their capability to contribute to primary productivity, microorganisms in such habitats have become of interest to investigate. Since Proteobacteria were previously found to be abundant, and the waters are well-aerated and organic-rich, this study on the presence of anoxygenic phototrophic bacteria, purple non-sulfur bacteria or aerobic anoxygenic phototrophs in marshes was initiated. Samples were collected at 7 Manitoban sites and anoxygenic phototrophs were cultivated and preliminary enumerated. A group of 14 strains, which represented the phylogenetic diversity of the isolates was further physiologically investigated. Aerobic anoxygenic phototrophs and purple bacteria were present at each location and they belonged to the α- and β-Proteobacteria subphyla. Some were closely related to known heavy metal reducers (Brevundimonas) and xenobiotic decomposers (Novosphingobium, Sphingomonas). All were able to synthesize the photosynthetic complexes aerobically. This research highlights the diversity and potential contribution’s anoxygenic phototrophs make to the essential functions taking place in wetlands.
Keywords:
wetlands
; marshes
; aerobic anoxygenic phototrophs
; purple non-sulfur bacteria
; anoxygenic phototrophs
; Proteobacteria
1. Introduction
Wetlands form a significant proportion of North America’s ecosystems [1] and are defined as sites adapted to a wet environment as a result of having a water table situated near or above surface level long term [2]. They fulfill many systemic roles which provide stability to surrounding locale and are an invaluable habitat for a variety of plants, fish and waterfowl [2,3]. Examples are functioning as initial reservoirs for water during flooding, limiting damage to nearby locations [2,4]. In addition, their ecological capacity to act as a carbon sink, potentially makes them a key player in combatting global warming and climate change [2]. The filter, breakdown of anthropogenic waste and pollutants like metals, fertilizers, antibiotics and sewage has been shown to occur [2,4,5]. In fact, the many beneficial contributions have led to development of constructed wetlands as wastewater treatment systems, important biodiversity hot zones, educational sites or for recreation [2].
In Manitoba, Canada, there are many wetlands, comprising approximately 43% of the total land area, accredited to the generally flat topography [1]. This includes swamps, bogs, fens, marshes and prairie potholes. Marshes do not form peat, have large seasonal fluctuations, are affected by ground and surface waters, usually do not have tree or shrubs and tend to be found near shallow open waters [1]. They are the minority, comprising about 2.5% of total provincial terrestrial land [1]. Prairie potholes also known as sloughs are shallow ponds containing marsh-like features abundant in Southern Manitoba [6,7].
Growing interest in understanding these ecosystems, has coincided with greater research into the microbes occupying the habitats. Previously, microorganisms were identified to contribute significantly to primary productivity in biofilms [8] and shown to impact carbon uptake and accumulation [9], both critical activities which researchers have primarily attributed to plants [2]. In some studies of wetland communities, eDNA sequencing of the V4 region of the 16S rRNA gene, found Proteobacteria comprised a major group [10,11]. The analysis of their environmental contributions have mainly centered around the capability of some to participate in biogeochemical cycling of sulfur, nitrogen and phosphorus [10,12]. However, there are also Proteobacteria that participate in carbon cycling and photosynthesis, undoubtedly playing a part in primary organic productivity and sequestration in marshes. This includes two groups which primarily reside in α- and β-Proteobacteria, purple non-sulfur bacteria (PNSB) and aerobic anoxygenic phototrophs (AAP). They utilize light energy through a cyclic pathway, where it is converted to chemical energy (ATP) through photophosphorylation with the involvement of pigment bacteriochlorophyll a (Bchl a). The process of anoxygenic photosynthesis is nearly identical in both groups with a few exceptions: AAP only perform it aerobically, and PNSB anaerobically; AAP use it as a supplemental source of energy (to cellular respiration) and PNSB have the metabolic capability to grow exclusively by photosynthesis. Furthermore, AAP are unable to fix carbon while PNSB can [13]. AAP and PNSB inhabit a variety of environments [13,14,15], however there are limited studies on their presence in wetlands [16,17,18]. Additionally, anoxygenic phototrophic bacteria’s (AnPB) contribution was not considered in previous work, as Bchl a measurements were not done [9]. Therefore, it is important to fill these gaps of knowledge to get a clear picture of the AnPB impact to the critical roles’ marshes serve, especially carbon sequestration and bioremediation.
As marsh waters are rich in organics and well-aerated due to enhanced algal and cyanobacterial oxygenic photosynthetic productivity, it was expected that AAP would be present in abundance and the metabolically versatile PNSB may be as well. To gain insight into the composition of the culturable aerobic AnPB in wetlands, three Manitoban locations were sampled. This included a slough in King’s Park, constructed marshes FortWhyte Alive, both within Winnipeg, and Oak Hammock Marsh in South-central Manitoba. These are the initial results of a longer term study on marsh water microbiology, of which is the first to focus on AnPB.
2. Materials and Methods
2.1. Sample Collection and Strain Cultivation
Water was obtained on July 10th 2023 from 8 different sites located in constructed Manitoban marshes: King’s Park, FortWhyte Alive in Winnipeg, and Oak Hammock Marsh near Stonewall. The pH was taken using a broad range pH (2.0 – 10.0) paper strip. Once collected, specimens were immediately placed on ice in the dark and kept there until return to the laboratory. Next, 10-fold serial dilutions to 10-8 were prepared for each sample with the following solution (g/L): MgCl2, 0.5; KH2PO4, 0.3; NH4Cl, 0.3; and CaCl2, 0.1, adjusted to pH 7.0 after autoclaving [19]. All samples (100 – 10-8) were plated on three different media: rich organic (RO), oligotrophic (OM) and potato-broth (PM) as previously described [20]. Incubation of plates occurred at 28 °C in the dark for two weeks, then grown at room temperature for two additional weeks. Throughout this period, they were monitored for colored colonies of interest to be streaked to obtain pure cultures for subsequent analysis. Once a strain was pure, it was cryopreserved at -75 °C using OM and 30% glycerol.
2.2. Identification of AnPB
Colonies of interest were restreaked on their respective isolation media. Once confirmed to be pure, whole cell absorption spectra in the range of 300 – 1100 nm was taken for detection of carotenoids, Bchl a and light harvesting (LH) complexes, by identifying corresponding peak(s). Plate-grown cells were resuspended in 0.3 ml of 20 mM TRIS-HCl buffer (pH 7.8) and 0.7 ml glycerol to minimize light scattering [21]. Subsequent testing (described below) of phototrophic anaerobic and photoautotrophic aerobic growth alongside 16S rRNA gene sequencing confirmed identities of AnPB isolates. 14 strains which represented the phenotypic and phylogenetic diversity were subjected to further analysis.
2.3. Morphology and Physiology of Isolates
Shape and size of cells were assessed with phase contrast microscopy (Zeiss Axioskop 2) after 4 days of growth at 28 °C in the dark on isolation medium. Motility was evaluated in a similar manner, except after 2 days with a hanging drop slide. A Gram stain [22] as well as a KOH test [23] was checked for all strains.
Physiological experiments were conducted at 28 °C, pH 7.0 in RO for a week in the dark, aerobically, in a shaker incubator unless stated otherwise. Temperature growth range and optimum were determined at the following (approximate values, °C): 7, 12, 16, 20, 25, 28, 32, 37 and 41. For pH, a range of 4.0 to 11.0 at 1.0 increments was studied. Utilization of individual carbon sources were evaluated in RO medium initially prepared without organics and then before inoculation supplemented with 0.5% of the following: Na-acetate, Na-butyrate, Na-citrate, ethanol, Na-formate, fructose, glucose, Na-glutamate, lactose, malic acid, Na-pyruvate and Na-succinate. Photoheterotrophic anaerobic growth was assessed in purple non-sulfur medium (PNSM) [24]. Liquid cultures in PNSM as well as a modified version which substituted L-cysteine and L-methionine for 1.0 mM of Na2S, were incubated in filled screw-capped tubes, in constant light. Aerobic photoautotrophy was evaluated using liquid basal, organic-free RO supplemented with 1.5 g/L NaHCO3 as a carbon source and 0.5 g/L Na2S2O3 as an electron donor [25] and grown in constant illumination provided by an incandescent light bulb. To account for organics contained within the inoculum liquid culture, 2 additional transfers of cells to fresh basal RO were conducted to ensure growth was truly photoautotrophic. The ability to ferment glucose, fructose and sucrose were evaluated as described [19]. Oxidase, catalase, aerobic nitrate reduction as well as hydrolysis of tween 20, 40, 60 and 80, starch, gelatin and agar, were determined [19,26]. Antibiotic susceptibility assessed with diffusion disks of the following (μg): ampicillin (10), chloramphenicol (30), erythromycin (15), imipenem (10), kanamycin (30), penicillin G (10 IU), polymyxin B (300 IU), streptomycin (10), and tetracycline (30).
2.4. 16S rRNA Sequencing and Phylogenetic Study
DNA was extracted and partial 16S rRNA gene was Sanger sequenced with universal primers [27]. Chromatograms processed using DNA Baser Assembler v4.36.0 (Heracle BioSoft SRL, Romania) [27]. Standard nucleotide BLAST determined the most related type species for each strain [28]. Sequences of OHM48, OHM176, OHM172, OHM24, OHM16, OHM14, KP164, FW250, KP4, FW199, FW159, FW36, FW5 and FW153 were deposited and are available in GenBank, accession numbers PP726887-PP726900, respectively.
A phylogenetic tree based on the 16S rRNA gene was constructed with MEGA X software [29] through neighbour joining alignment with 1000 bootstrap replicates. The evolutionary history was inferred using the Maximum Likelihood method. The model chosen for the tree, Tamura-3 parameter [30] was selected with the MEGA X ‘find best DNA model’ tool. A total of 62 nucleotide sequences and 1582 positions were in the final dataset.
3. Results and Discussion
3.1. Site Description, Isolation, and Detection of AnPB
Sampling sites are shown in Figure 1. King’s Park, Site 1, is a recreational area in its South part with one slough, located right beside the Red River. As marshes are typically affected by ground and surface waters [1], it is very likely that the activity along this portion of the river also impacts the slough. Site 1 was at the edge of a pond with plentiful aquatic vegetation. The water was clear, and sample was obtained just below the surface to avoid collecting plant matter. Sites 2-4 were at FortWhyte Alive. This is a protected environment comprised of forests, prairie grassland, constructed lakes and marshes, serving as a recreational and educational area [31]. In recent years, the waters have had rising phosphorus and nitrogen levels, causing eutrophication [32]. Site 2 was abundant in greenery and had floating algal-cyanobacterial mats. Site 3 was copious in macrophytes, nearly covering the entire surface. Samples were taken just below a large algal bloom and hydrophytes, if present, at each respective location. At Site 4, a thin olive-green bacterial mat from the top layer of the sediment was collected. It had rocks, and more turbid water around. Sites 5-7 were at Oak Hammock Marsh in South-central Manitoba. Formerly an extensive wetland called St. Andrew’s bog, this 36 km2 area is the reconstructed remnants of considerable drainage of the fertile land for agriculture [33]. Located directly beside a walkway, Site 5 had abundant reeds and floating plants at the surface. The water was slightly brown and turbid. Site 6 had an abundance of dense grass-like macrophytes. A brownish, purple sulfur bacterial mat layer on the subaqueous soil was Site 7. This was identified based on the smell of sulfide coming from the sample. As sulfide reacts with oxygen, it was presumed this site was anaerobic. It was agitated during collection and as such, water directly above was acquired as well. This sample came from the deepest zone below the surface of all sites. In general, prairie marshes are shallow [8] with anaerobic sediment at the bottom [2], therefore they will usually have a relatively steep oxygen gradient. As such, there is potential to find AnPB’s which display aerotolerant pigment production and have the capacity to conduct anoxygenic photosynthesis both aerobically and anaerobically, like the transitional Charonomicrobium ambiphototrophicum EG17 [25].
Surprisingly, in the 3 studied habitats, there was quite poor microbial mat development, suggesting bacterial communities within the water remained mostly suspended, attached to sediments or surrounding floating plants. Sampling took place during the afternoon on a sunny day, however, the vegetation covering the surface of some Sites (1, 3 and 6) may have affected the amount of light that penetrated and was available for photosynthesis. The pH was taken and for each site was approximately as follows, in order from 1 to 7: 6.0-7.0, 9.0-10.0, 7.0, 7.0, 8.0, 9.0-10.0, 8.0. As these were pH paper estimations, the values reflect the range of the sites and are not exact. However, most fall within the expected, as marshes are known to be relatively neutral [2]. Future experimentation will require an accurate pH meter to get precise values.
Colored colonies were present in each media tested from all sites, with more appearing throughout the duration of incubation. AnPB were found by the identification of Bchl a peak in whole cell absorption spectra. In total, 102 or 43.4% of pigmented isolates from the 235 tested were AnPB (Table 1): 62% were in RO, followed by OM (19.6 %) and PM (18.3 %). Majority of selected colonies had orange or yellow hues. From the total, 14 were selected to represent the diversity at sites (Table 2). Four of the strains were PNSB and all isolated on PM. This does not necessarily mean colonies did not develop on RO and OM, but were likely not isolated from these plates, because PNSB usually do not actively synthesize photosynthetic pigments aerobically and produce pale colors due to limited carotenoids [14]. However, on PM, the PNSB colonies were colored, and produced pigment-protein complexes (Figure 2). When these strains were plated on RO, the complexion was muted or non-colored (not shown), although they also displayed their photosynthetic apparatus (Figure 2). Therefore, based on appearance, such colonies on RO and OM were not chosen. The other 10 strains studied were AAP, confirmed through physiological activity. In general, the group constituted the majority of the AnPB obtained. AAP were present in all samples regardless of depth, indicating the waters were well-aerated. Support comes from the fact that the places have plentiful vegetation, algae and cyanobacteria, and as such, a significant amount of oxygen was produced from their oxygenic photosynthetic activity [2]. It is especially interesting considering a purple sulfur bacteria mat was observed, which should be anoxic due to the sulfide presence, identified by scent, as it is reacting with the surrounding oxygen and that the majority in this community were probably anaerobes [14]. The possible explanation for the isolation of AAP from this site, is that they were situated in the aerobic water just above the mat. In such a case, the presence of AAP nearby is proof of the presence of a steep oxygen gradient.
The presence of AnPB in the marshes aligns with previous works that identified their residence in wetland-like environments with infrared epifluorescence microscopy [16,17,34] or sampling of PNSB from soils [18]. Nonetheless our paper is the first describing the isolation of PNSB and AAP from constructed marshes and sloughs. Obtaining pure cultures is especially important as it allows the roles attributed to microbes to be studied directly and may help to indicate other activities the bacterial community contributes to. Here, it was used to accurately identify AnPB and distinguish PNSB and AAP, a task difficult to perform through environmental sequencing or microscopy. Typically, sequencing of the pufM gene is used to indicate AAP presence in aerobic environments [35], however this gene is also present in PNSB and if they are growing aerobically in the areas measured, they will also be counted. Epifluorescence microscopy [36] uses infrared lighting to infer AAP cells from others, but the issues still remain as PNSB can also be detected in this approach. As a result, both of these methods do not precisely differentiate the two.
3.2. Spectral Analysis
All of PNSB cultures (KP4, FW5, OHM24 and FW36) produced photosynthetic pigment-protein complexes anaerobically as well as aerobically (Figure 2). Bchl a and carotenoids were higher while grown anoxically in PNSM as expected, since light harvesting is usually conducted photoheterotrophically or photoautotrophically where oxygen is absent [14]. Interestingly, for each isolate, the relative level of expression of LHI, LHII and carotenoids varied in aerobic dark and anaerobic light conditions, as well as between the two media (RO and PM) in the presence of oxygen. This characteristic corresponded well with visual difference of pigmentation in all strains investigated, supporting the conclusion that there was varied expression of carotenoids (400-600 nm) and Bchl a (LH peak(s) at 850-880 nm). In general, on PM, PNSB had greater primary: accessory pigment concentrations as reported earlier [13] and LHII in comparison to RO (Figure 2).
Both aerobic and anaerobic photosynthetic complex expression by the same species has been seen before, although it is not common [13]. Mentioned above strain EG17, a γ-Proteobacterium isolated from a hypersaline spring East German Creek, Manitoba is to date the sole known which synthesizes Bchl a regardless of oxygen presence [25]. However, there are a few AAP closely related to the PNSB Rhodobacter [37,38] suggesting the possibility of such expression of pigments may occur in some not yet taxonomically defined strains. These absorption spectra do not show definitively PNSB were using light energy aerobically, but it would be worthwhile to investigate in KP4 and FW5 (related to a species currently or formerly placed in the Rhodobacter genus [39]). FW36, interestingly showed significantly more LHI in oxic conditions than the others investigated. A close relative Rubrivivax gelatinosus, has been previously shown to produce photosynthetic pigments in semi-aerobic conditions [40] and the observations in FW36 could potentially be explained by the center of colonies having less oxygen, allowing greater expression. OHM24 synthesizing its pigments aerobically and anaerobically, was the least surprising, considering its relation to Rhodopseudomonas sulfidophila, which also displayed this characteristic [41]. As these strains, which reside in different subphyla of Proteobacteria showed aerobic photosynthetic pigment-protein complex expression, it would be worthwhile to investigate if that is the case for some other PNSB.
The AAP isolates in general displayed typical spectral features also found in the most closely related genera (Figure 3). They all had an abundance of carotenoids relative to low Bchl a, which is a common attribute of the group. The majority of accessory pigments have been found to protect cells from photooxidation and only a few support light absorbance when directly incorporated in LH complexes [13]. This was similar to the PNSB strains when grown aerobically, but was vastly different to anoxic, where carotenoids and Bchl a were expressed in relatively equal proportions (Figure 2). FW153 displayed the most red-shifted Bchl a peak (872 nm) of the strains, but was still within the known range [42,43]. FW159 and FW176 had the most defined LHI peak among isolates. FW250 and OHM48 were the only AAP with an LHII produced. This has been seen previously in Polymorphobacter [44] and not in Erythrobacter, their respective closely related genera.
With a wide phylogenetic diversity of anoxygenic phototrophs isolated comprising part of the microbial community in marshes, they probably occupy an important ecological niche, although this has not yet been well investigated. Unfortunately, no studies show AnPB’s contribution to overall photosynthesis in wetlands as they tend to focus on Cyanobacteria and chlorophyll a measurement [9]. Some approaches utilize pufM gene, which codes for a part of the reaction center in both groups of AnPB, as a genetic indicator of AAP in aerobic environments [35]. This is an issue as it may also capture PNSB in the oxygenated portion of the water. Therefore, it is likely such numbers were overestimates. Another strategy of AAP detection and enumeration using epifluorescence microscopy and infrared lighting [36] also has a chance to not be fool-proof. We identified a set of PNSB occupying a wide breadth of phylogenetic diversity, including different subphyla, that express their photosynthetic pigments aerobically. There is a high probability, especially in nutrient-rich locations such as marshes and high-peat content wetlands, some PNSB will express Bchl a aerobically, leading them to also be detected with infrared light. Again, it would misrepresent the pervasiveness of AAP. A method that can effectively differentiate the two with the utmost accuracy has not yet been designed, and as such, these possibilities should at the very least, be acknowledged. Studies on the Bchl a prevalence and photosynthetic activity of AnPB communities would be insightful for understanding their contribution to the overall primary productivity in wetlands.
3.3. Phenotypic Features of Strains
All isolates have gram-negative cell wall. They grew at and near neutral pH (Table 3) as expected, since most sites were near pH 7.0-8.0. No strain could survive at pH 5.0 or lower and OHM14 was the only one to not grow at pH 6.0. The cultures from Site 3 and 7 (FW199, OHM172, FW153, OHM176) which were alkaline (pH 9.0 to 10.0), grew at 9.0 except for OHM176. This is likely because paper tests are not very accurate, so the actual pH could be different than what was paper-estimated as marshes are typically neutral [2] making Sites 3 and 7 unusual. The optimal growth for the group was either at pH 6.0 or 7.0. Interestingly, FW199, FW36 and FW5 were able to grow at significantly alkaline conditions. Aside from these instances, their growth pH range is a reflection of the sites and what was expected from isolates’ phylotype.
In general, the AnPB had broad temperature growth ranges, although the best for each one was at 32 or 37 °C (Table 3). FW5 and OHM16 grew in all temperatures tested. The thermotolerance in all representatives may be attributed to the climate of Manitoba experiencing some of the coldest and hottest temperatures in Canada annually. It is credited to the flat topography and lack of mountains which usually act as temperature stabilizers.
As expected of PNSB, FW5, KP4, FW36 and OHM24 grew anaerobically as photoheterotrophs. The AAP could not. All strains were incapable of aerobic phototautotrophic growth. These two components in conjunction with the production of Bchl a (Figure 3), brought the conclusion that the other 10 AnPB were indeed AAP.
Most of isolates were not motile after 2 days of growth (Table 3). The strains’ morphology (Figure 4) was coccoid (FW5, OHM14), ovoid (FW250, KP4) or rod (FW199, OHM172, KP164, OHM176, FW153, OHM48, OHM16, FW159, OHM24, FW36) with FW153 having tapered ends (Figure 4F). FW5 was coccoidal, although it’s close relative Cereibacter azotoformans was characterized as ovoid to rod-shaped. KP164 had light-refractuile circular globules inside, varying from 1-5 per cell (Figure 4E). This could possibly be an accumulation of polyhydroxyalkanoate, which have been previously shown in some AAP, depending on the conditions [45].
Most strains could use at least one carbon source (Table 4), with the exception of OHM14 and FW250. Both were able to grow on RO and OM, suggesting there are essential for growth components in the complex media. They were all incapable of growing with ethanol or Na-formate. Phylogenetically close groups showed similar trends. Erythrobacteraceae members (OHM16, OHM48, FW159, FW172) all used Na-butyrate, Na-glutamate but not Na-citrate, fructose, lactose and malic acid. FW199, KP164 and FW153 of Sphingomonadaceae did not grow with Na-citrate, malic acid and Na-succinate as sole carbon sources but could use glucose. Paracoccaceae KP4 and FW5 were most versatile, utilizing: Na-acetate, Na-butyrate, fructose, glucose, Na-glutamate, Na-pyruvate and Na-succinate, but not lactose or malic acid. Interestingly this also applied to FW36, but not the remaining PNSB, OHM24. FW199 was the only one able to assimilate lactose and OHM24 was the sole strain that metabolized malic acid. Fermentation did not occur with the sugars tested. KP4 produced an acid, when grown with fructose. This is typical, as only a few AnPB can ferment and some make acids as a result of metabolizing sugars. As for enzymes, all strains were oxidase positive. FW199, OHN172, KP164, OHM176, OHM48 and OHM16 were catalase positive; FW199, KP164, OHM48, OHM16, FW159, and FW36 could break down starch; KP164, FW250, OHM176, OHM16, FW159, OHM14, FW36 and KP4 hydrolyzed gelatin; FW153, FW36, FW5 reduced nitrate to nitrite aerobically. None hydrolyzed agar. Strong lipolytic activity was found in Erythrobacteraceae members, FW199, KP164, OHM172 and FW36. The rich variety of organic carbon types used or broken down by the AnPB (amino acids, mono-, di- and polysaccharides, organic acids, lipids) indicates their great contribution to carbon cycling and important role in sequestering accumulated organics in marshes. Further physiological examination of pure cultures could lead to the discovery of additional influences AnPB made within wetland communities.
The AnPB had varying degrees of antibiotic resistance (Table 5), however some general trends existed among the group. All AnPB were susceptible to imipenem and resistant to nalidixic acid. Most were susceptible to kanamycin as well with the exception of FW250. PNSB isolates FW5, KP4 and FW36 had sensitivity to all tested, except nalidixic acid. OHM24 and FW250 resisted the highest number of antibiotics. In marshes, this is of interest as they are recreational areas with increased human activity, and therefore have greater exposure to anthropogenic waste potentially including antibiotics. Some wetlands have also been constructed as wastewater treatment locations [2] and have been shown to breakdown antibiotics, primarily through the function of microbes, like some Proteobacteria, which use them as carbon sources [46,47,48]. Sequencing has indicated a decrease in resistance genes [46,49]. Additionally, pathogenic bacteria have been known to be dismantled through processes such as antibiotic secretion from macrophytes [50]. In contrast, it was revealed that resistance can accumulate in such environments because of selective pressure, through gene transfer and by bacteria settling in sediments and waters, which varies by season and is dependent on wetland type [46,48,49]. As such, the microorganisms’ performance in removing excess nitrogen and phosphorus in the habitats could be affected as antibiotics may selectively target participators in the biogeochemical cycles [46,48]. This could potentially change the community structure to more antibiotic-resistant bacteria, decreasing total biodiversity and possibly heightening eutrophication overtime [46]. Recently, concerns have been raised with the rise of nitrogen and phosphorus levels in marsh waters at FortWhyte [32]. Whether it was caused by antibiotics or not, the eutrophication may worsen the marsh capability to self-sustain, especially if important community members are out-competed. Furthermore, when used as wastewater treatment centers, wetlands act as a point of waste release into habitats close by. If mechanisms of inhibition are not proficient, antibiotic resistance could spread to those areas. As most of our isolates were sensitive, such persistence may also affect the relative abundance of AnPB’s. Even if previous studies showed increase in Proteobacteria [48], it will not affect all within the phylum the same way, as observed in our strains. Antibiotics could limit their ability to contribute to carbon cycling and photosynthetic activity. Continual monitoring of antibiotic resistance genes in the water and among microbes within wetland wastewater treatment sites would provide a greater insight on the potential impacts it may have on the habitat’s primary productivity.
3.4. 16S rRNA Gene-Based Phylogenetics
Results from 16S rRNA partial gene sequencing (1300-1400 bp) show most strains are related to AAP or PNSB (Table 2). All isolates belong to α-Proteobacteria, with the exception of a β-proteobacterium, FW36. This matches the phylogenetic placement of most known AAP and PSNB [14,51].
Majority of isolates (KP164, FW250, OHM176, OHM48, OHM16, FW159, OHM24, FW36, FW5, and KP4) likely represent new strains within most related species, because of their very high 16S rRNA gene similarity. FW199, OHM172, FW153, and OHM14 may potentially be new species, however they have yet to be taxonomically described as such and DNA-DNA hybridization of complete genome would be required to support such conclusion as well as finding of other phenotypic distinguishing features could help. Four of the isolates belong to the Erythrobacteraceae family. OHM16, OHM48 and FW159 are all members of Erythrobacter and OHM172 is from Novosphingobium, known AAP genera [52]. KP164, FW199 and FW153 are from the family, Sphingomonadaceae, genera Blastomonas, Sphingomonas and Rhizorhabdus, respectively. Novosphingobium and Sphingomonas representatives have been shown to degrade aromatic compounds and other xenobiotics, as well as the ability to tolerate and accumulate heavy metals [53,54,55,56]. Therefore, they may play a significant role in bioremediation and degradation of anthropogenic waste in marshes. FW153’s most closely related genus has no AAP and none have been shown to produce Bchl a. Prior reports indicated Rhizorhabdus did not synthesize carotenoids [57], however the newest member described, Rhizorhabdus phycosphaerae, FW153’s closest relative, was proven otherwise [58]. Identification of the photosynthetic gene cluster in Rhizorhabdus spp. may aid in evaluating if this genus has other AAP members or if its features are unique to FW153. All other strains were related to published AnPB. FW250 is the sole representative of Polymorphobacter a tentative genus in Sphingosinicellaceae [59]. FW5 and KP4 in Paracoccaceae, are closely related to Ceribacter azotoformans (previously known as Rhodobacter azotoformans [39]) and Rhodobacter capsulatus, respectively. Both species have denitrification capabilities that were investigated for wastewater treatment [60,61]. OHM176 is associated with the Brevundimonas genus, found in Caulobacteraceae. They are resistant to high levels of heavy metals [19,62,63] possibly contributing to filtering and treating metal waste known to occur in wetlands [4]. Nitrobacteraceae is represented with Rhodopseudomonas relative, OHM24. The most distant (based on 16S rRNA gene phylogeny, Figure 5) from all other α-Proteobacteria is OHM14 connected to Roseomonas (synonym, Falsiroseomonas) in Acidobacteraceae. FW36, is the sole strain found from the β-Proteobacteria subphylum. It is closely related to Rubrivivax gelatinosus of the Comamonadaceae family. Alongside contributing to photosynthetic productivity and carbon cycling, AnPB also participate in other activities such as degrading anthropogenic pollutants and heavy metal oxides as mentioned in the specific examples. These have been broadly advertised as important roles AAP and PNSB play in bioremediation [13,64].
4. Conclusions
Cultivated isolates revealed a diverse, and readily available community of AnPB, indicating they are important microbial contributors to the life in marsh ecosystems. This is likely because such places are highly enriched in organics, have a neutral pH, and are aerobic due to oxygenic photosynthetic activity. Furthermore, waters there are relatively shallow and have limited peat accumulation, making sunlight accessible in excess. Although the conditions better support the growth of AAP then PNSB, the metabolic flexibility of the latter has also made it possible to culture. It is especially important to consider as many AAP studies do not factor PNSB as the error contributors to their relative abundance detected through infrared epifluorescence microscopy and pufM sequencing. However, they are indeed present, can synthesize photosynthetic pigments aerobically, and therefore must be accounted as influencing such measurements. This is one example of how cultivation is important for precise analysis and comprehension of microbial contributions, as proof of activity can be directly assessed, and it can appropriately compliment sequencing and microscopy techniques. There is a need for a better understanding of AnPB’s participation in the total primary productivity in the habitats to accurately evaluate their ecological role. Although AnPB here represent a wide breadth of the potential community in Manitoba marshes, there are many more, which simply remain unculturable. It could include AnPB that express the photosynthetic apparatus regardless of oxygen availability. Since there is a steep oxygen gradient in marshes, going from well aerated shallow waters to anaerobic bottom sediments, such flexibility in using anoxygenic PS would be advantageous and could exist here. Nonetheless, the phylogenetic diversity of the isolates and the known physiology provide some context to functions they possibly perform. While some insights into the AnPB community have been enriched, more work is necessary to better elucidate their contribution to the essential activity’s wetlands perform for the biosphere. This section is not mandatory but can be added to the manuscript if the discussion is unusually long or complex.
Author Contributions
Conceptualization, Vladimir Yurkov and Katia Messner; Data curation, Vladimir Yurkov and Katia Messner; Formal analysis, Vladimir Yurkov and Katia Messner; Funding acquisition, Vladimir Yurkov; Investigation, Vladimir Yurkov and Katia Messner; Methodology, Vladimir Yurkov and Katia Messner; Project administration, Vladimir Yurkov; Resources, Vladimir Yurkov; Supervision, Vladimir Yurkov; Validation, Vladimir Yurkov and Katia Messner; Visualization, Vladimir Yurkov and Katia Messner; Writing – original draft, Katia Messner; Writing – review & editing, Vladimir Yurkov. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant 1501 and a University of Manitoba GETS grant, both held by V. Yurkov.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Halsey, L.; Vitt, D.; Zoltai, S. Climatic and Physiographic Controls on Wetland Type and Distribution in Manitoba, Canada. Wetlands 1997, 17, 243–262. [Google Scholar] [CrossRef]
- Kennedy, G.; Mayer, T. Natural and Constructed Wetlands in Canada: An Overview. Water Qual. Res. J. Can. 2002, 37, 295–325. [Google Scholar] [CrossRef]
- Wrubleski, D.; Badiou, P.; Goldsborough, G. Coastal Wetlands of Manitoba’s Great Lakes (Canada). In The Wetland Book II: Distribution, Description, and Conservation; Finlayson, C., Milton, G., Prentice, R., Davidson, N., Eds.; Springer: Dordrecht, Netherlands, 2018; Volume 1, pp. 591–604. [Google Scholar]
- Watchorn, K.E.; Goldsborough, L.G.; Wrubleski, D.A.; Mooney, B.G. A Hydrogeomorphic Inventory of Coastal Wetlands of the Manitoba Great Lakes: Lakes Winnipeg, Manitoba, and Winnipegosis. J Great Lakes Res 2012, 38, 115–122. [Google Scholar] [CrossRef]
- Reddy, K.R.; Gale, P.M. Wetland Processes and Water Quality: A Symposium Overview. J. Environ. Qual. 1994, 23, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Brunet, N.N.; Westbrook, C.J. Wetland Drainage in the Canadian Prairies: Nutrient, Salt and Bacteria Characteristics. Agric. Ecosyst. Environ. 2012, 146, 1–12. [Google Scholar] [CrossRef]
- Ducks Unlimited Canada Prairie Pothole Region. Available online: https://www.ducks.ca/places/prairie-pothole-region/ (accessed on 9 February 2024).
- Robarts, R.D.; Waiser, M.J. Effects of Atmospheric Change and Agriculture on the Biogeochemistry and Microbial Ecology of Prairie Wetlands. Great Plains Research 1998, 8, 113–136. [Google Scholar]
- Hamard, S.; Céréghino, R.; Barret, M.; Sytiuk, A.; Lara, E.; Dorrepaal, E.; Kardol, P.; Küttim, M.; Lamentowicz, M.; Leflaive, J.; et al. Contribution of Microbial Photosynthesis to Peatland Carbon Uptake along a Latitudinal Gradient. J. Ecol. 2021, 109, 3424–3441. [Google Scholar] [CrossRef]
- Mellado, M.; Vera, J. Microorganisms That Participate in Biochemical Cycles in Wetlands. Can. J. Microbiol. 2021, 67, 771–788. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ji, Z.; Pei, Y. Nutrient Removal and Microbial Community Structure in an Artificial-Natural Coupled Wetland System. Process Saf. Environ. 2021, 147, 1160–1170. [Google Scholar] [CrossRef]
- Gutknecht, J.L.M.; Goodman, R.M.; Balser, T.C. Linking Soil Process and Microbial Ecology in Freshwater Wetland Ecosystems. Plant Soil. 2006, 289, 17–34. [Google Scholar] [CrossRef]
- Yurkov, V. V; Beatty, J.T. Aerobic Anoxygenic Phototrophic Bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 695–724. [Google Scholar] [CrossRef]
- Madigan, M.T.; Jung, D.O. An Overview of Purple Bacteria: Systematics, Physiology, and Habitats. In The Purple Phototrophic Bacteria; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, Netherlands 2009; Volume 3, pp. 1–15. [Google Scholar]
- Yurkov, V.; Csotonyi, J.T. New Light on Aerobic Anoxygenic Phototrophs. In The Purple Phototrophic Bacteria; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, Netherlands 2009; Volume 3, pp. 31–55. [Google Scholar]
- Szabó-Tugyi, N.; Vörös, L.; Balogh, K.; Botta-Dukát, Z.; Bernát, G.; Schmera, D.; Somogyi, B. Aerobic Anoxygenic Phototrophs Are Highly Abundant in Hypertrophic and Polyhumic Waters. FEMS Microbiol. Ecol. 2019, 95, fiz104. [Google Scholar] [CrossRef] [PubMed]
- Lew, S.; Koblížek, M.; Lew, M.; Medová, H.; Glińska-Lewczuk, K.; Owsianny, P.M. Seasonal Changes of Microbial Communities in Two Shallow Peat Bog Lakes. Folia Microbiol. 2015, 60, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.E.; Gorham, E.; Pratt, D.C. Distribution of Purple Photosynthetic Bacteria in Wetland and Woodland Habitats of Central and Northern Minnesota. J. Bacteriol. 1974, 117, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Hughes, E.; Head, B.; Maltman, C.; Piercey-Normore, M.; Yurkov, V. Aerobic Anoxygenic Phototrophs in Gold Mine Tailings in Nopiming Provincial Park, Manitoba, Canada. Can. J. Microbiol. 2017, 63, 212–218. [Google Scholar] [CrossRef]
- Kuzyk, S.B.; Ma, X.; Yurkov, V. Seasonal Dynamics of Lake Winnipeg’s Microbial Communities Reveal Aerobic Anoxygenic Phototrophic Populations Coincide with Sunlight Availability. Microorganisms 2022, 10, 1690. [Google Scholar] [CrossRef]
- Rathgeber, C.; Yurkova, N.; Stackebrandt, E.; Schumann, P.; Beatty, J.T.; Yurkov, V. Roseicyclus mahoneyensis gen. nov., sp. nov., an Aerobic Phototrophic Bacterium Isolated from a Meromictic Lake. Int. J. Syst. Evol. Microbiol. 2005, 55, 1597–1603. [Google Scholar] [CrossRef]
- Beveridge, T.J. Use of the Gram Stain in Microbiology. Biotech. Histochem. 2001, 76, 111–118. [Google Scholar] [CrossRef]
- Gregersen, T. Rapid Method for Distinction of Gram-Negative from Gram-Positive Bacteria. Eur. J. Appl. Microbiol. Biotech. 1978, 5, 123–127. [Google Scholar] [CrossRef]
- Bilyj, M.; Lepitzki, D.; Hughes, E.; Swiderski, J.; Stackebrandt, E.; Pacas, C.; Yurkov, V. V. Abundance and Diversity of the Phototrophic Microbial Mat Communities of Sulphur Mountain Banff Springs and Their Significance to the Endangered Snail, Physella Johnsoni. Open J. Ecol. 2014, 4, 488–516. [Google Scholar] [CrossRef]
- Csotonyi, J.T.; Stackebrandt, E.; Swiderski, J.; Schumann, P.; Yurkov, V. An Alphaproteobacterium Capable of Both Aerobic and Anaerobic Anoxygenic Photosynthesis but Incapable of Photoautotrophy: Charonomicrobium ambiphototrophicum, gen. nov., sp. nov. Photosynth. Res. 2011, 107, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Yurkov, V. V.; Krieger, S.; Stackebrandt, E.; Beatty, J.T. Citromicrobium bathyomarinum, a Novel Aerobic Bacterium Isolated from Deep-Sea Hydrothermal Vent Plume Waters That Contains Photosynthetic Pigment- Protein Complexes. J. Bacteriol. 1999, 181, 4517–4525. [Google Scholar] [CrossRef]
- Kuzyk, S.B.; Jafri, M.; Humphrey, E.; Maltman, C.; Kyndt, J.A.; Yurkov, V. Prosthecate Aerobic Anoxygenic Phototrophs Photocaulis sulfatitolerans gen. nov. sp. nov. and Photocaulis rubescens sp. nov. Isolated from Alpine Meromictic Lakes in British Columbia, Canada. Arch. Microbiol. 2022, 204, 1–15. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the Number of Nucleotide Substitutions When There Are Strong Transition-Transversion and G+C-Content Biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed]
- FortWhyte Alive Our Story. Available online: https://www.fortwhyte.org/about/our-story/ (accessed on 1 February 2024).
- Ayumu, B.B. Water Budget Investigation for FortWhyte Alive; University of Manitoba: Winnipeg, Manitoba, Canada, 2020. [Google Scholar]
- Oak Hammock Marsh History. Available online: https://www.oakhammockmarsh.ca/about/history-of-oak-hammock/ (accessed on 1 February 2024).
- Lew, S.; Lew, M.; Koblížek, M. Influence of Selected Environmental Factors on the Abundance of Aerobic Anoxygenic Phototrophs in Peat-Bog Lakes. Environ. Sci. Pollut. Res. 2016, 23, 13853–13863. [Google Scholar] [CrossRef] [PubMed]
- Gazulla, C.R.; Cabello, A.M.; Sánchez, P.; Gasol, J.M.; Sánchez, O.; Ferrera, I. A Metagenomic and Amplicon Sequencing Combined Approach Reveals the Best Primers to Study Marine Aerobic Anoxygenic Phototrophs. Microb. Ecol. 2023, 86, 2161–2172. [Google Scholar] [CrossRef] [PubMed]
- Mašín, M.; Nedoma, J.; Pechar, L.; Koblížek, M. Distribution of Aerobic Anoxygenic Phototrophs in Temperate Freshwater Systems. Environ. Microbiol. 2008, 10, 1988–1996. [Google Scholar] [CrossRef]
- Pérez, V.; Dorador, C.; Molina, V.; Yáñez, C.; Hengst, M. Rhodobacter sp. Rb3, an Aerobic Anoxygenic Phototroph Which Thrives in the Polyextreme Ecosystem of the Salar de Huasco, in the Chilean Altiplano. Anton. Leeuw. 2018, 111, 1449–1465. [Google Scholar] [CrossRef]
- Kyndt, J.A.; Robertson, S.; Shoffstall, I.B.; Ramaley, R.F.; Meyer, T.E. Genome Sequence and Characterization of a Xanthorhodopsin-Containing, Aerobic Anoxygenic Phototrophic Rhodobacter Species, Isolated from Mesophilic Conditions at Yellowstone National Park. Microorganisms 2022, 10, 1169. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Muramatsu, K.; Ueda, Y. Molecular Genetic Analyses of Rhodobacter azotoformans sp. nov. and Related Species of Phototrophic Bacteria. Syst. Appl. Microbiol. 1996, 19, 168–177. [Google Scholar] [CrossRef]
- Takaichi, S.; Shimada, K. Pigment Composition of Two Pigment-Protein Complexes Derived from Anaerobically and Semi-Aerobically Grown Rubrivivax gelatinosus, and Identification of a New Keto-Carotenoid, 2-Ketospirilloxanthin. Plant Cell Physiol. 1999, 40, 613–617. [Google Scholar] [CrossRef]
- Hansen, T.A.; Veldkamp, H. Rhodopseudomonas sulfidophila, nov. spec., a New Species of the Purple Nonsulfur Bacteria. Arch. Mikrobiol. 1973, 92, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Csotonyi, J.T.; Swiderski, J.; Stackebrandt, E.; Yurkov, V. A New Environment for Aerobic Anoxygenic Phototrophic Bacteria: Biological Soil Crusts. Environ. Microbiol. Rep. 2010, 2, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Csotonyi, J.T.; Swiderski, J.; Stackebrandt, E.; Yurkov, V. V. Novel Halophilic Aerobic Anoxygenic Phototrophs from a Canadian Hypersaline Spring System. Extremophiles 2008, 12, 529–539. [Google Scholar] [CrossRef]
- Jia, L.; Feng, X.; Zheng, Z.; Han, L.; Hou, X.; Lu, Z.; Lv, J. Polymorphobacter fuscus sp. nov., Isolated from Permafrost Soil, and Emended Description of the Genus Polymorphobacter. Int. J. Syst. Evol. Microbiol. 2015, 65, 3920–3925. [Google Scholar] [CrossRef]
- Xiao, N.; Jiao, N. Formation of Polyhydroxyalkanoate in Aerobic Anoxygenic Phototrophic Bacteria and Its Relationship to Carbon Source and Light Availability. Appl. Environ. Microbiol. 2011, 77, 7445–7450. [Google Scholar] [CrossRef]
- Liu, X.; Guo, X.; Liu, Y.; Lu, S.; Xi, B.; Zhang, J.; Wang, Z.; Bi, B. A Review on Removing Antibiotics and Antibiotic Resistance Genes from Wastewater by Constructed Wetlands: Performance and Microbial Response. Environ. Pollut. 2019, 254, 112996. [Google Scholar] [CrossRef]
- Ma, J.; Cui, Y.; Li, A.; Zou, X.; Ma, C.; Chen, Z. Antibiotics and Antibiotic Resistance Genes from Wastewater Treated in Constructed Wetlands. Ecol. Eng. 2022, 177, 106548. [Google Scholar] [CrossRef]
- Ohore, O.E.; Qin, Z.; Sanganyado, E.; Wang, Y.; Jiao, X.; Liu, W.; Wang, Z. Ecological Impact of Antibiotics on Bioremediation Performance of Constructed Wetlands: Microbial and Plant Dynamics, and Potential Antibiotic Resistance Genes Hotspots. J. Hazard. Mater. 2022, 424, 127495. [Google Scholar] [CrossRef]
- Sabri, N.A.; Schmitt, H.; van der Zaan, B.M.; Gerritsen, H.W.; Rijnaarts, H.H.M.; Langenhoff, A.A.M. Performance of Full Scale Constructed Wetlands in Removing Antibiotics and Antibiotic Resistance Genes. Sci. Total Environ. 2021, 786, 147368. [Google Scholar] [CrossRef] [PubMed]
- Berezowsky, M. Constructed Wetlands for Remediation of Urban Waste Waters. Geosci. Can. 1995, 22, 129–141. [Google Scholar]
- Yurkov, V.; Hughes, E. Aerobic Anoxygenic Phototrophs: Four Decades of Mystery. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Springer International Publishing: Switzerland, 2017; pp. 193–214. [Google Scholar]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. Photosynthesis is Widely Distributed among Proteobacteria as Demonstrated by the Phylogeny of PufLM Reaction Center Proteins. Front. Microbiol. 2018, 8, 2679. [Google Scholar] [CrossRef]
- Gupta, S.K.; Lal, D.; Lal, R. Novosphingobium panipatense sp. nov. and Novosphingobium mathurense sp. nov., from Oil-Contaminated Soil. Int. J. Syst. Evol. Microbiol. 2009, 59, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.P.; Wang, B.J.; Liu, Y.H.; Liu, S.J. Novosphingobium taihuense sp. nov., a Novel Aromatic-Compound-Degrading Bacterium Isolated from Taihu Lake, China. Int. J. Syst. Evol. Microbiol. 2005, 55, 1229–1232. [Google Scholar] [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From Diversity and Genomics to Functional Role in Environmental Remediation and Plant Growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Chettri, B.; Singh, A.K. Kinetics of Hydrocarbon Degradation by a Newly Isolated Heavy Metal Tolerant Bacterium Novosphingobium panipatense P5:ABC. Bioresour. Technol. 2019, 294, 122190. [Google Scholar] [CrossRef]
- Francis, I.M.; Jochimsen, K.N.; De Vos, P.; van Bruggen, A.H.C. Reclassification of Rhizosphere Bacteria Including Strains Causing Corky Root of Lettuce and Proposal of Rhizorhapis suberifaciens gen. nov., comb. nov., Sphingobium mellinum sp. nov., Sphingobium xanthum sp. nov. and Rhizorhabdus argentea gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 1340–1350. [Google Scholar]
- Kim, M.; Bae, J.; Park, W. Rhizorhabdus phycosphaerae sp. nov., Isolated from the Phycosphere of Microcystis aeruginosa. Int. J. Syst. Evol. Microbiol. 2022, 72, 005324. [Google Scholar] [CrossRef]
- Hördt, A.; López, M.G.; Meier-Kolthoff, J.P.; Schleuning, M.; Weinhold, L.M.; Tindall, B.J.; Gronow, S.; Kyrpides, N.C.; Woyke, T.; Göker, M. Analysis of 1,000+ Type-Strain Genomes Substantially Improves Taxonomic Classification of Alphaproteobacteria. Front. Microbiol. 2020, 11, 468. [Google Scholar]
- Li, Z.; Li, L.; Sun, H.; Wang, W.; Yang, Y.; Qi, Z.; Liu, X. Ammonia Assimilation: A Double-Edged Sword Influencing Denitrification of Rhodobacter azotoformans and for Nitrogen Removal of Aquaculture Wastewater. Bioresour. Technol. 2022, 345, 126495. [Google Scholar] [CrossRef]
- Costa, S.; Ganzerli, S.; Rugiero, I.; Pellizzari, S.; Pedrini, P.; Tamburini, E. Potential of Rhodobacter capsulatus Grown in Anaerobic-Light or Aerobic-Dark Conditions as Bioremediation Agent for Biological Wastewater Treatments. Water-SUI. 2017, 9, 108. [Google Scholar] [CrossRef]
- Ali, A.; Li, M.; Su, J.; Li, Y.; Wang, Z.; Bai, Y.; Ali, E.F.; Shaheen, S.M. Brevundimonas diminuta Isolated from Mines Polluted Soil Immobilized Cadmium (Cd2+) and Zinc (Zn2+) through Calcium Carbonate Precipitation: Microscopic and Spectroscopic Investigations. Sci. Total Environ. 2022, 813, 152668. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Chaturvedi, P.; Chandra, R.; Kumar, S. Identification of Heavy Metals Tolerant Brevundimonas sp. from Rhizospheric Zone of Saccharum Munja L. and Their Efficacy in in-Situ Phytoremediation. Chemosphere 2022, 295, 133823. [Google Scholar] [CrossRef] [PubMed]
- Dhar, K.; Venkateswarlu, K.; Megharaj, M. Anoxygenic Phototrophic Purple Non-Sulfur Bacteria: Tool for Bioremediation of Hazardous Environmental Pollutants. World J. Microbiol. Biotechnol. 2023, 39, 283. [Google Scholar] [CrossRef]
Figure 1.
Locations sampled at Manitoba marshes. Site 1, King’s Park (A); Site 2-4 FortWhyte (B-D); Site 5-7 Oak Hammock Marsh (E-G).
Figure 1.
Locations sampled at Manitoba marshes. Site 1, King’s Park (A); Site 2-4 FortWhyte (B-D); Site 5-7 Oak Hammock Marsh (E-G).

Figure 2.
Whole cell absorption spectra of PNSB grown at various conditions. OHM24 (A), KP4 (B), FW5 (C), FW36 (D). Spectra were taken after 4 days of growth at 28 °C in the following conditions: anaerobically in PNSM illuminated (purple), aerobically in the dark on PM (dark blue) and RO (green). Bchl a peaks are indicated.
Figure 2.
Whole cell absorption spectra of PNSB grown at various conditions. OHM24 (A), KP4 (B), FW5 (C), FW36 (D). Spectra were taken after 4 days of growth at 28 °C in the following conditions: anaerobically in PNSM illuminated (purple), aerobically in the dark on PM (dark blue) and RO (green). Bchl a peaks are indicated.
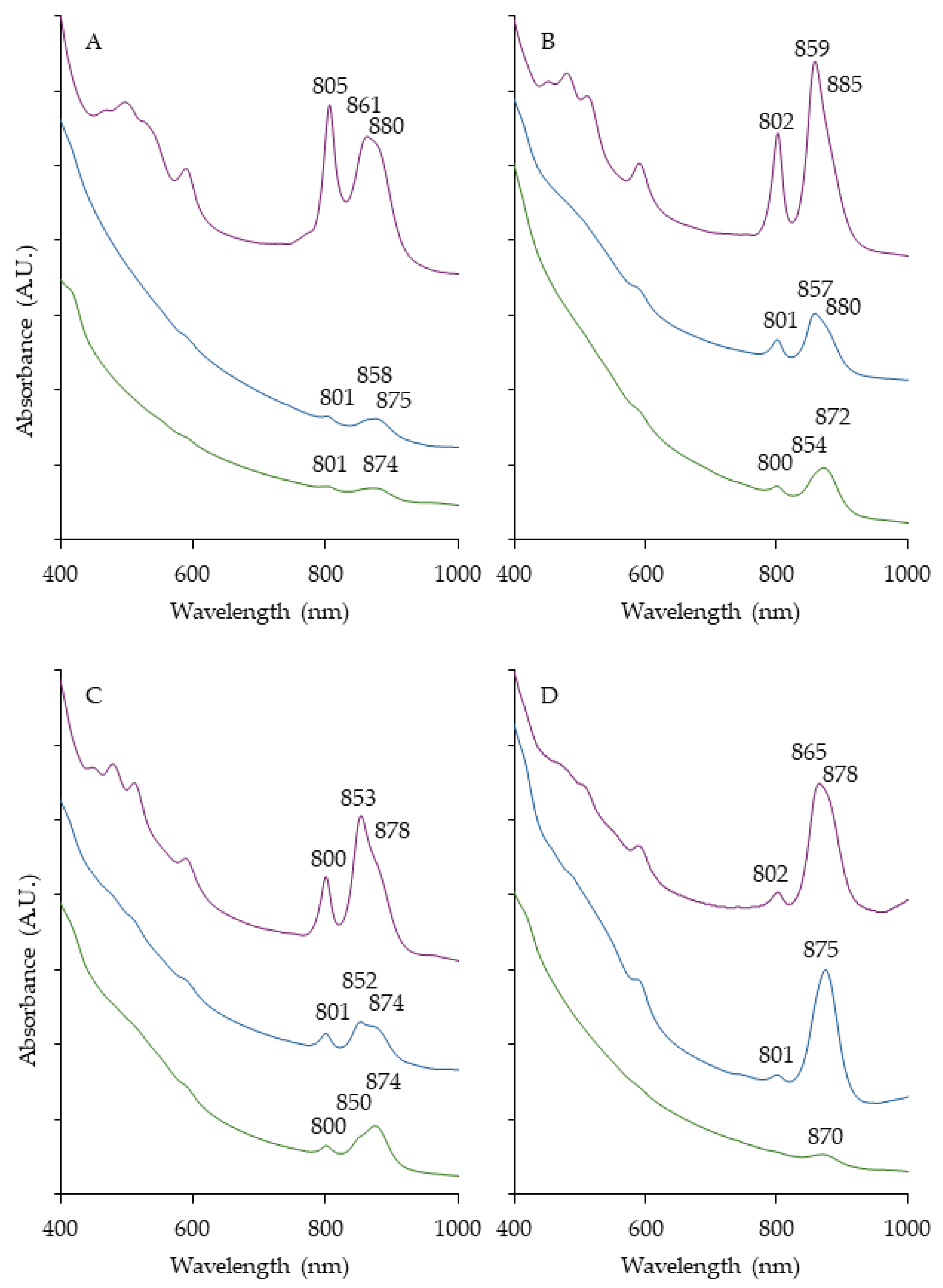
Figure 3.
Whole cell absorption spectra of AAP. Taken after 7 days of growth at 28 °C on their respective isolation media. Bchl a peaks marked. OHM14 (A), OHM16 (B), OHM48 (C), FW153 (D), FW159 (E), KP164 (F), FW172 (G), FW176 (H), FW199 (I), FW250 (J).
Figure 3.
Whole cell absorption spectra of AAP. Taken after 7 days of growth at 28 °C on their respective isolation media. Bchl a peaks marked. OHM14 (A), OHM16 (B), OHM48 (C), FW153 (D), FW159 (E), KP164 (F), FW172 (G), FW176 (H), FW199 (I), FW250 (J).
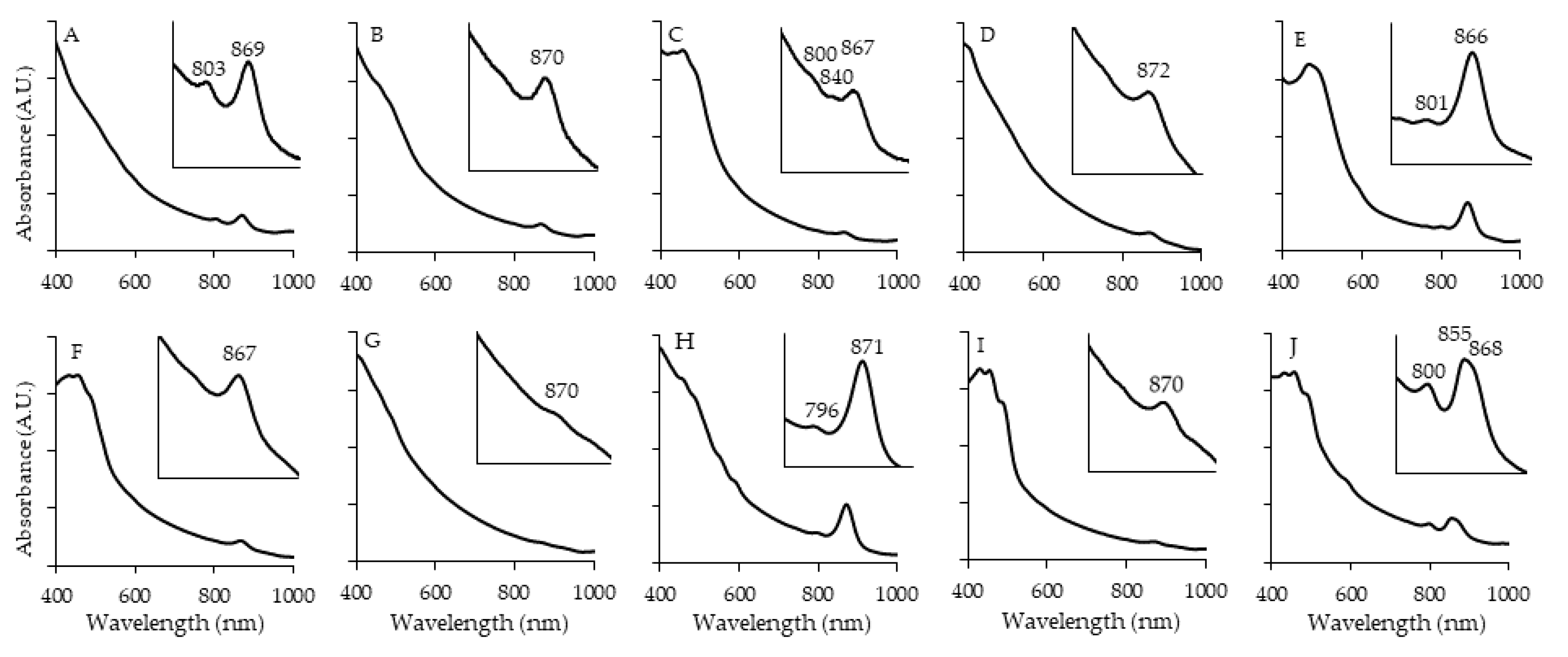
Figure 4.
Morphology of selected strains. Included are OHM14 (A), FW5 (B), FW250(C), FW36 (D), KP164 (E, insert of cell included), FW153 (F). Scale bar is 5 μm.
Figure 4.
Morphology of selected strains. Included are OHM14 (A), FW5 (B), FW250(C), FW36 (D), KP164 (E, insert of cell included), FW153 (F). Scale bar is 5 μm.
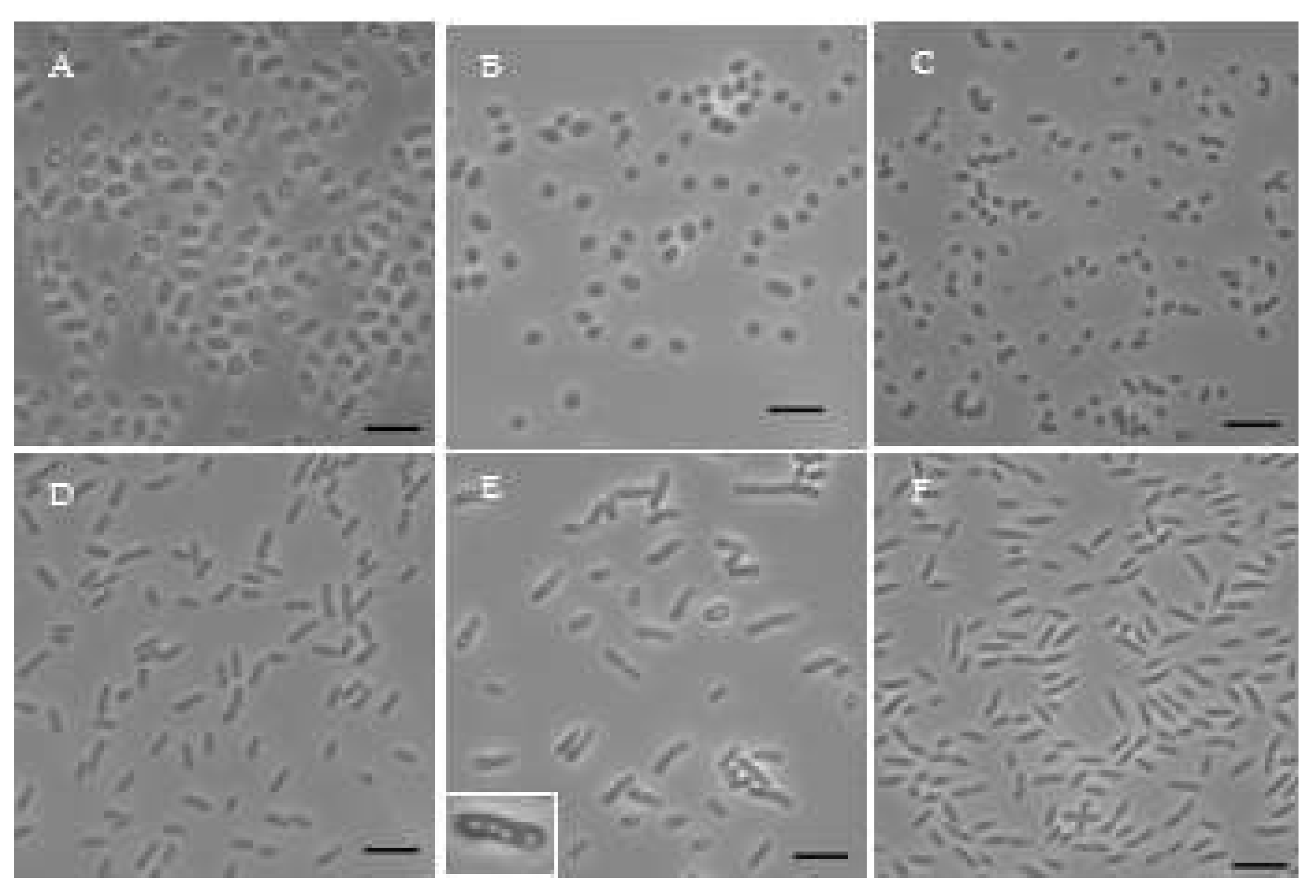
Figure 5.
Phylogenetic tree of representative strains from Manitoba marshes and most related based on 16S rRNA gene sequences. Version with highest log likelihood (-11688.47) is presented. Branch lengths measured in the number of substitutions per site. Percentage of trees in which the associated taxa clustered together is shown next to the branches. Accession number for sequences used included in parentheses.
Figure 5.
Phylogenetic tree of representative strains from Manitoba marshes and most related based on 16S rRNA gene sequences. Version with highest log likelihood (-11688.47) is presented. Branch lengths measured in the number of substitutions per site. Percentage of trees in which the associated taxa clustered together is shown next to the branches. Accession number for sequences used included in parentheses.
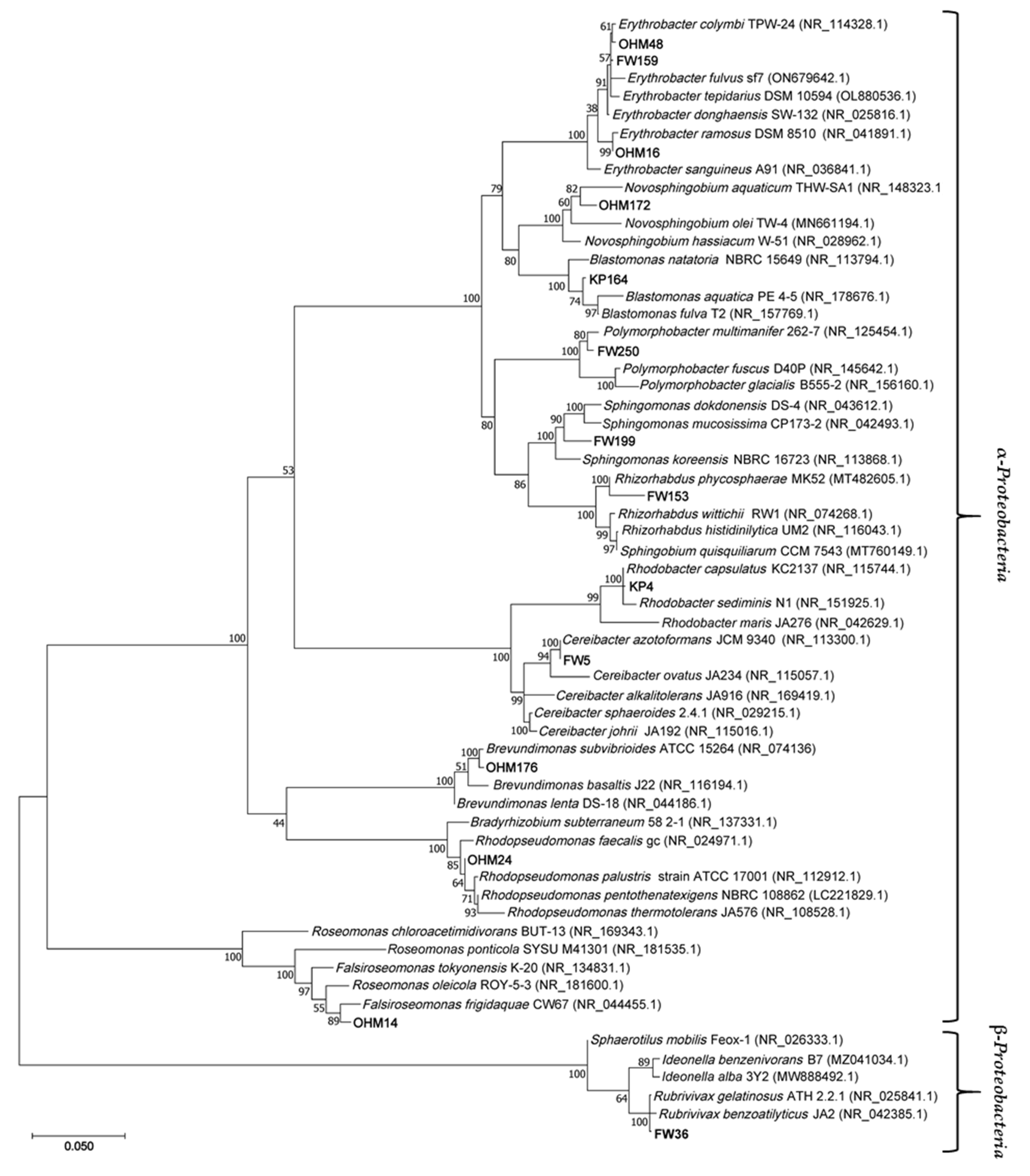

Table 1.
Total number of isolates from Manitoba marshes.
| Site | Colonies | Media | Colony Pigmentation | AnPB | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PM | RO | OM | Yellow | Orange | Red | Pink | Brown | |||
| King’s Park | ||||||||||
| 1 | 38 | 15 | 18 | 5 | 8 | 11 | 10 | 3 | 1 | 17 |
| FortWhyte Alive | ||||||||||
| 2 | 47 | 10 | 32 | 5 | 12 | 17 | 8 | 3 | 3 | 27 |
| 3 | 19 | 5 | 8 | 6 | 4 | 5 | 3 | 1 | 1 | 4 |
| 4 | 46 | 2 | 27 | 17 | 8 | 9 | 4 | 11 | 5 | 18 |
| Oak Hammock Marsh | ||||||||||
| 5 | 31 | 5 | 20 | 6 | 6 | 17 | 3 | 1 | 1 | 18 |
| 6 | 41 | 4 | 36 | 1 | 10 | 16 | 4 | 4 | 2 | 14 |
| 7 | 13 | 2 | 5 | 6 | 1 | 1 | 1 | 9 | 0 | 4 |
| Total Colonies | 235 | 43 | 146 | 46 | 49 | 76 | 33 | 32 | 13 | 102 |
| Overall Percentage (%) | - | 18.3 | 62.1 | 19.6 | 20.9 | 32.3 | 14.0 | 13.6 | 5.5 | 43.4 |
Table 1.
Representative AnPB used in this study.
| Strain | Sample Site | Dilution | Media | Most related species | 16S rRNA similarity (%) |
|---|---|---|---|---|---|
| FW199 | 3 | 10-3 | PM | Sphingomonas dokdonensis DS-4 | 97.69 |
| OHM172 | 7 | 10-2 | RO | Novosphingobium hassiacum W-51 | 98.65 |
| KP164 | 1 | 10-1 | RO | Blastomonas aquatica PE 4-5 | 99.47 |
| FW250 | 5 | 10-4 | RO | Polymorphobacter multimanifer 262-7 | 99.32 |
| FW153 | 3 | 10-3 | RO | Rhizorhabdus phycosphaerae MK52 | 97.11 |
| OHM176 | 7 | 10-2 | RO | Brevundimonas subvibrioides ATCC 15264 | 99.77 |
| OHM48 | 6 | 10-1 | RO | Erythrobacter colymbi TPW-24 | 99.70 |
| OHM16 | 8 | 10-2 | OM | Erythrobacter ramosus DSM 8510 | 99.77 |
| FW159 | 5 | 10-5 | RO | Erythrobacter donghaensis SW-132 | 99.77 |
| OHM14 | 8 | 100 | OM | Falsiroseomonas frigidaquae CW67 | 98.87 |
| OHM24 | 8 | 100 | PM | Rhodopseudomonas pentothenatexigens NBRC 108862 | 99.54 |
| FW36 | 4 | 10-3 | PM | Rubrivivax gelatinosus ATH 2.2.1 | 99.93 |
| FW5 | 4 | 10-3 | PM | Cereibacter azotoformans JCM 9340 | 100.00 |
| KP4 | 1 | 100 | PM | Rhodobacter capsulatus KC2137 | 99.92 |
Table 3.
Phenotypic features of representative AnPB.1.
| Strain | FW199 | OHM172 | KP164 | FW250 | FW153 | OHM176 | OHM48 | OHM16 | FW159 | OHM14 | OHM24 | FW36 | FW5 | KP4 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Colony colour | Yellow | Yellow | Yellow | Yellow-Brown | Light Orange | Orange | Orange | Orange | Red | Pink | Pink | Purple-Brown | Red | Red |
| Cell shape | Rod | Rod | Rod | Ovoid | Rod | Rod | Rod | Rod | Rod | Coccoid | Rod | Rod | Coccoid | Ovoid-rod |
| Cell size (µm) | 2.29±0.41 x 0.63±0.02 | 4.16±0.88 x 0.72±0.07 | 3.85±0.36 x 0.81±0.04 | 1.40±0.11 x 0.78±0.03 | 3.08±0.22 x 0.68±0.05 | 3.09±0.44 x 0.60±0.03 | 2.76±0.34 x 0.65±0.03 | 1.66±0.26 x 0.87±0.05 | 2.29±0.32 x 0.58±0.08 | 1.69±0.16 x 1.18±0.08 | 2.48±0.18 x 0.82±0.07 | 3.02±0.30 x 0.72±0.06 | 1.65±0.13 x 1.31±0.08 | 2.46±0.71 x 0.84±0.05 |
| Motility | + | - | - | - | - | + | - | - | - | + | + | + | + | - |
| Temperature range/optimum (°C) | 20-41/32 | 12-37/32 | 12-37/28 | 16-37/32 | 12-41/32 | 12-32/32 | 12-37/37 | 7-41/32 | 16-37/32 | 12-37/32 | 12-41/32 | 12-41/37 | 7-41/37 | 12-41/32 |
| pH range/optimum | 6-10/6 | 6-9/8 | 6-9/6 | 6-9/7 | 6-9/6 | 6-8/6 | 6-9/6 | 6-9/6 | 6-9/7 | 7-9/7 | 6-9/6 | 6-10/6 | 6-11/6 | 6-9/6 |
| Enzyme Activity: | ||||||||||||||
| Catalase | + | + | + | - | - | + | + | + | - | - | - | - | - | - |
| Oxidase | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Nitrate reductase | - | - | - | - | + | - | - | - | - | - | - | + | + | - |
| Hydrolysis of: | ||||||||||||||
| Gelatin | - | - | + | + | - | + | - | + | + | + | - | + | - | + |
| Starch | + | - | + | - | - | - | + | + | + | - | - | + | - | - |
| Tween 20 | + | - | + | - | - | - | - | + | - | - | - | + | - | - |
| Tween 40 | - | + | + | - | - | - | + | + | + | - | - | + | - | - |
| Tween 60 | + | + | + | + | - | + | + | + | + | - | - | + | - | - |
| Tween 80 | + | + | + | - | - | + | + | + | + | - | - | + | - | - |
1 +, Growth; -, No Growth.
Table 4.
Organic carbon sources utilized by representative strains.1.
| Strain | FW199 | OHM172 | KP164 | FW250 | FW153 | OHM176 | OHM48 | OHM16 | FW159 | OHM14 | OHM24 | FW36 | FW5 | KP4 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Utilization of: | ||||||||||||||
| Acetate | - | + | + | - | + | - | + | + | - | - | + | + | + | + |
| Butyrate | - | + | + | - | + | - | + | + | + | - | + | + | + | + |
| Citrate | - | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Ethanol | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Formate | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Fructose | + | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Glucose | + | + | + | - | + | + | + | + | - | - | - | + | + | + |
| Glutamate | + | + | - | - | + | + | + | + | + | - | - | + | + | + |
| Lactose | + | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Malate | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| Pyruvate | - | + | + | - | + | + | + | + | - | - | + | + | + | + |
| Succinate | - | - | - | - | - | + | + | + | - | - | + | + | + | + |
| Fermentation: | ||||||||||||||
| Fructose | - | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Glucose | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Sucrose | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
1 +, Growth; -, No Growth.
Table 5.
Organic carbon sources utilized by representative strains.1.
| Strain | FW199 | OHM172 | KP164 | FW250 | FW153 | OHM176 | OHM48 | OHM16 | FW159 | OHM14 | OHM24 | FW36 | FW5 | KP4 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antibiotics: | ||||||||||||||
| Ampicillin | R | R | S | R | R | R | R | R | R | S | R | S | S | S |
| Chloramphenicol | R | S | S | R | S | S | S | S | S | S | R | S | S | S |
| Erytromycin | S | S | S | R | R | S | S | S | S | S | R | S | S | S |
| Imipenem | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Kanamycin | S | S | S | R | S | S | S | S | S | S | S | S | S | S |
| Naladixic Acid | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| Penicilin G | R | R | S | R | R | R | R | R | R | S | R | S | S | S |
| Polymixin B | S | S | S | R | S | S | S | S | S | R | R | S | S | S |
| Streptomycin | R | R | R | S | R | S | R | R | R | S | R | S | S | S |
| Tetracycline | S | S | S | R | S | S | S | S | S | S | R | S | S | S |
1 R, resistant; -, S, Susceptible.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



