
Submitted:
29 April 2024
Posted:
30 April 2024
You are already at the latest version
Abstract
Broflanilide is a novel active ingredient from a unique insecticide class (IRAC Group 30) and was investigated as a control option against modern insecticide resistant strains of the common bed bug, Cimex lectularius. The susceptibility of technical grade broflanilide and efficacy of a broflanilide-based aerosol formulation (Earth Corp., Japan) were evaluated against five C. lectularius strains (MONH, SYD, DARL, PARRA, RIPPO). Results demonstrated that bed bugs had high susceptibility to broflanilide (RR50: SYD=1.22-fold, DARL=1.03-fold, PARRA=3.65-fold, RIPPO=2.77-fold), compared to the susceptible MONH strain. The aerosol showed high efficacy to adult bed bugs (97.5–100 % mortality to five strains after 24 hrs post-spray) and produced a notable decrease in egg hatch rates (MONH=9.7–23.4 %, PARRA=14.9–22.6 %). The aerosol also demonstrated high residual activity with 97.5–100% mortality in all five bed bug strains within 7 days exposure. The impact of different surface types upon aerosol residual efficacy was limited, although residues on a non-porous surface (tile) killed bed bugs slightly quicker than that on a porous surface (filter paper). The aerosol provided long residual activity to bed bugs, as 6-month-old residues were as equally efficacious to that of fresh residues. Bed bugs did not avoid surfaces treated with the aerosol. The high efficacy (including ovicidal efficacy), long residual activity, and non-avoidance of bed bugs on treated surfaces, demonstrated that broflanilide is highly suitable for modern bed bug management.
Keywords:
bed bug
; broflanilide
; susceptibility
; efficacy
; surface substrates
; aged residues
; ovicidal efficacy
1. Introduction
Over the past 25 years, worldwide there has been a dramatic increase in the number of bed bug infestations, including both the common (Cimex lectularius L.) and tropical (Cimex hemipterus [F.]) species [1,2]. Numerous suggestions have been offered to account for the return of this public health pest [3]. However, it is now recognised that insecticide resistance has been the main contributing cause [2,4]. Resistance in bed bugs has been widely reported to multiple insecticide classes [1,5,6,7]. Unfortunately, most of the insecticides registered for bed bug control around the globe today belong to those groups that have demonstrated resistance [1,6]. This has meant that control is a major challenge and treatment failures have been common. Nevertheless, insecticide application is still a universally mentioned method in modern bed bug management [8]. Therefore, there is an urgent need for new insecticides to be available for the control of resistant bed bug strains.
Broflanilide [N-[2-bromo-4-(1,1,1,2,3,3,3-heptafluoropropan-2-yl)-6-(trifluoromethyl)phenyl]-2-fluoro-3-(N-methylbenzamido)benzamide] is a novel insecticide recently discovered by Mitsui Chemicals Agro, Inc., Japan [9]. Broflanilide has a unique chemical structure characterized as a meta-diamide targeting the gamma-aminobutyric acid (GABA) gated chloride channel in the insect nervous system, resulting in a block of inhibitory neurotransmissions, convulsions, and death [9]. Broflanilide has been classified by the Insecticide Resistance Action Committee (IRAC) as a GABA-gated chloride channel allosteric modulator (IRAC Group 30)[10]. Broflanilide has been reported to have excellent efficacy against a range of insect pests including multiple-insecticide resistant strains, such as against the malaria mosquito (Anopheles gambiae Giles 1902) [11], wireworm (Agriotes obscurus L. 1758) [12], cotton bollworm (Helicoverpa armigera Hübner 1808) and beet armyworm (Spodoptera exigua Hübner 1808) [13]. Multiple formulations of the insecticide have been produced for the control of urban pests, including a wettable powder (e.g., VECTRON™ T500) for mosquito control, aerosols (e.g., PT® VediraTM Pressurized Insecticide and Iyana Mushi ZEROeKNIGHT One Push Spray for 60 times use [IMZOPS 60]), a fumigant for bed bug control (e.g., Iyana Mushi ZEROeKNIGHT for 10–13 m2), as well as various formulations to control agricultural pests (Cimegra® SC and TeraxxaTM F4 seed treatment [Liquid]).
Broflanilide has been registered for bed bug control for some time (e.g., Australia since 2019) [14], however no publication has appeared on the efficacy of the chemical against insecticide susceptible and resistant bed bug strains. Therefore, in this study we investigated: 1) the susceptibility of technical grade broflanilide and efficacy of a broflanilide-based aerosol formulation (IMZOPS 60) against insecticide susceptible and resistant bed bug C. lectularius strains; 2) the effect of surface substrate (porous versus non-porous) and aging upon the efficacy of IMZOPS 60 against C. lectularius, and; 3) the behavioral response of C. lectularius to areas treated with IMZOPS 60.
2. Materials and Methods
2.1. Bed Bug Populations
The bed bug species used in this study was the common bed bug, C. lectularius, and included five strains (Table 1). Bed bug strains were maintained separately in plastic jars (250 mL flat bottom container, Techno Plas Pty Ltd, St Marys, SA Australia, Item Code No.: S10065SL) and provided with folded filter paper (Whatman #1, Cat. No. 1001931) as a harborage. The insects were held in an insectary with conditions of 25±1 °C and 75±10 % RH, with a photoperiod of 12:12 (L:D) hrs. All the insects were maintained via a blood meal provided once per week on anesthetized rats. Blood-feeding was conducted on the rodents as approved by the Westmead Hospital Animal Ethics Committee (WHAEC Protocol No. 2004.03.24) and in accordance with NSW Animal Research Review Panel (ARRP). No bed bug strain had undergone insecticide selection. The species of all the bed bug strains were confirmed using the taxonomic keys of Usinger (1966) [15]. The bed bugs were blood fed 5 d prior to experimental usage. All assays were carried out in a room under laboratory conditions of 23±1 °C, 40±10 % RH, with a photoperiod of 10:14 (L:D) hrs.
2.2. Chemicals
Both technical grade broflanilide and the broflanilide-based aerosol formulation were supplied by Earth Corporation, Japan. The technical grade active ingredient had a purity of 99% and was manufactured by Mitsui Chemicals Agro, Inc., Tokyo, Japan. The broflanilide-based aerosol formulation (Iyana Mushi ZEROeKNIGHT One Push Spray for 60 times use [IMZOPS 60]) has a broflanilide concentration of 1% (w/v). All the chemicals were applied by an Australian pest management licensed operator (Dang, K. License No. 5088512). The technical grade broflanilide was dissolved in acetone (Sigma-Aldrich Australia, CAS No.: 67-64-1) prior to the assays.
2.3. Topical Assays of Technical Grade Broflanilide
Based on a pilot study, the concentration of the technical grade broflanilide was adjusted to 0.5–50 ng/µL for testing against the susceptible Monheim strain (MONH), and 0.5–100 ng/µL for testing against the insecticide resistant bed bug strains of Sydney (SYD), Darlinghurst (DARL), Parramatta (PARRA), and Ripponlea (RIPPO) strains. In the topical assays, ten adult bed bugs (mixed sex and age) were placed in a clean glass petri dish (90 mm diameter and 15 mm height, unbranded), and anesthetized with CO2 gas for 5 seconda prior to topical application of the diluted insecticide. The diluted insecticide-acetone solution (1 µL) was applied onto the dorsal surface of the abdomen of each bed bug using a micro-pipette (FINNPIPETTE 4500 0.5–10 µL single channel pipette, Thermo Fisher Scientific, Macquarie Park, NSW Australia, Item Code No.: U56359) equipped with an aerosol barrier 20-µL pipette tip (Interpath Services, Somerton, VIC Australia, Cat. No.: 24500). The treated bed bugs were held in the glass Petri dish until the applied insecticide solution had dried. Then, the bed bugs were gently moved to a clean plastic Petri dish (60 mm diameter and 14 mm height, Techno Plas Pty Ltd, ST Marys, SA Australia, Item Code No.: S6014S10) lined with one filter paper disc (55 mm diameter, Whatman #1, Cat No. 1001-055). The filter paper (including in the below assays) was fixed to the bottom of the plastic Petri dish with glue (COS®Glue Stick, non-toxic, COS, Australia, Item Code No.: ADHE4101) to prevent bed bugs from crawling under the paper. The number of bed bugs from each strain used in the assays are shown in Table 2. For the control, each bed bug (corresponding replicates of 10 insects of each strain, mixed sex, and age) was treated with 1 µL of acetone only. Due to the slow mode of action of broflanilide [17], mortality was assessed after 72 hrs post-treatment by gently nudging each individual insect with soft entomological forceps. The moribund individuals that were unable to maintain balance and showed uncoordinated twitching were recorded as dead in these assessments, and none subsequently recovered.
2.4. Efficacy of the Broflanilide-Based Aerosol Formulation (IMZOPS 60)
2.4.1. Direct Spray Assays on Adult Bed Bugs
Adult bed bugs (mixed sex and age, four replicates of 10 insects) were placed into plastic Petri dishes (90 mm diameter and 14 mm height, Techno Plas Pty Ltd, ST Marys, SA Australia, Item Code No.: TPG9014G) lined with a filter paper disc (90 mm diameter, Whatman #1, Cat. No. 1001-090). The broflanilide-based aerosol formulation, IMZOPS 60, was sprayed directly at the bed bugs from a distance of 15–20 cm. It is important to note that this aerosol is not a constant flow spray, with the insecticide being released only during the depressing of the trigger. Once fully depressed, the insecticide flow ceases. To ensure each insect was fully treated, two presses of the aerosol were applied to each replicate of 10 insects in a fume cupboard (Dynamic Fume Exhaust Systems, Australia). Untreated bed bugs (mixed sex and age, four replicates of 10 insects) placed in plastic Petri dishes lined with filter paper served as the control. Mortality was recorded 24 hrs post-spray. The assays were carried out against all five bed bug strains (e.g., MONH, SYD, DARL, PARRA, RIPPO).
2.4.2. Direct Spray Assays on Bed Bug Eggs
Black cardstock paper (COS® A4 300 gsm Leathergrain Binding Report Covers, COS, Australia, order code: BIND9120) was cut into 90 mm diameter round discs, which then were glued on the bottom of plastic Petri dishes (14 mm height x 90 mm diameter). Twenty-five adult bed bugs (20 females and 5 males, mixed ages) were introduced into each Petri dish and then held in the insectary. Fifteen replicates of 25 insects were set up for each strain of MONH and PARRA. After 48 hr, all adult bed bugs were removed from the Petri dishes. Eggs were checked and counted under a stereo microscope (Leica M80, Danaher Corporation, USA) and non-viable eggs (e.g., deflated eggs) were removed. The fifteen Petri dishes with eggs were divided into three groups; 1) GROUP ONE, the control, 2) GROUP TWO, eggs were directly sprayed with IMZOPS 60 (two pushes), of which, the age of eggs ranged 1–2 days old at the time of treatment, and 3) GROUP THREE, eggs were held for 3 days, and then directly sprayed by IMZOPS 60 (two pushes), of which, the age of eggs ranged 4–5 days old at the time of treatment. All spray applications were conducted in a fume cupboard. Egg hatch rate (hatch status: egg capsules were completely opened, and the first instars completely emerged from the egg) was recorded daily after the eggs were 6 days old, until no eggs hatched for 3 days.
2.4.3. Residual Assays against Adult Bed Bugs
Two surface substrates were evaluated in the residual assays: a smooth non-porous surface (ceramic tiles) and a porous surface (filter paper). The ceramic tile (100 x 100 mm, unbranded), and filter paper discs (90 mm diameter, Whatman #1) lined within plastic Petri dishes (14 mm height x 90 mm diameter), were directly sprayed with IMZOPS 60 in a fume cupboard. They were then left to air dry over 24 hrs prior to the assays. To ensure the whole surface were fully treated, two presses of the IMZOPS 60 were applied to each tile and filter paper. Untreated tiles and filter paper served as controls. Adult bed bugs (mixed sex and age, four replicates of 10 insects) were placed directly onto, and continually held, on the treated surfaces. Plastic cups (150 mL, 75mm diameter x 61mm height, Genfac Plastic, Dandenong South, VIC Australia, Code No.: RB150) with the base removed, were held in place with tape on the treated tiles. The cup ensures that bed bugs were in continuous contact with the tiles. The cup base was removed to allow the introduction of the insects onto tiles [18]. A total of 160 bed bugs (40 insects for each of tile-treated, tile-control, filter paper-treated, and filter paper-control) were used in the assays with each strain. Due to the slow mode of action of broflanilide, mortality was recorded daily for up to 7 days post exposure. The residual assays were carried out against all five bed bug strains.
2.4.4. Residual Assays with First Instar Bed Bugs
First instars (mixed age, four replicates of 10 insects) were placed onto treated tiles (40 insects) and filter paper (40 insects) as above. Another four replicates of 10 insects were placed onto untreated tiles (40 insects) and untreated filter paper (40 insects), to serve as controls. The assays were carried against the susceptible MONH and resistant PARRA strains only. Mortality was recorded daily until all insects were dead on the treated surfaces.
2.4.5. Aged Residual Assays against Adult Bed Bugs
The two substrate types as used above, were evaluated in the aged residual assays. The ceramic tiles (10 cm x 10 cm), and filter paper discs (90 mm diameter) lined within plastic Petri dishes (14 mm height x 90 mm diameter), were sprayed with IMZOPS 60 in a fume cupboard, and then left to air dry over 24 hrs prior to the assays. Two presses of IMZOPS 60 were applied to the tiles and filter paper as above. Untreated tiles and filter paper served as controls. Treated tiles and filter paper aged for 1 DAY served as the fresh residues. Treated surfaces and controls were aged for 1 WEEK, 2 WEEKS, 4 WEEKS, 3 MONTHS, and 6 MONTHS, which were then compared against the results of the fresh residual assays. Adult bed bugs (mixed sex and age, four replicates of 10 insects) were placed onto the tiles and filter paper after the gaining period. The plastic cups mentioned above were used for confining bed bugs on the tiles. The assays tested the MONH and PARRA strains only. A total of 160 bed bugs for each strain and each aged residue were used in the assays. In total, 800 bed bugs of each strain were used. Mortality was recorded daily up to 7 days.
2.5. Behavioral Responses to Harborages Treated with IMZOPS 60
2.5.1. One-Choice Harborage Assays
To establish if bed bugs are repelled on surfaces treated with the formulated broflanilide-based aerosol, choice tests were conducted with insecticide treated and untreated harborages. Firstly, a One-Choice test was conducted with a single treated harborage. This was carried out in plastic Petri dishes (14 mm height x 90 mm diameter), which had a filter paper disc (90 mm diameter) glued to the base and is referred to as an arena. A line was drawn down the center of each filter paper disc, dividing each into two semicircles (zones I and II, Figure 1A). A filter paper disc (55 mm diameter) was treated by spraying directly IMZOPS 60 (one push for each side of the paper). After drying for 3 hrs, the treated filter paper disc was folded four times as a harborage (around 1 cm tent height), and then fixed onto zone II of the filter paper lined Petri dish with double-sided adhesive tape (Scotch® Double-Sided Tape 137, 12.7 mm x 11 m, 3M, Australia, ID No.: 70005266401) (Figure 1A). Ten adult insects (mixed sex and age) were acclimated to zone I for 15 min by containing them in a glass vial (20 mm diameter × 80 mm height, unbranded) [19]. Insects were released by raising the vial. The control consisted of a harborage of untreated filter paper (55 mm diameter) placed in zone B of the arena, in separate Petri dishes. The behavioral response assays were carried out with both the MONH and PARRA strains. In total, 80 insects (four replicates of 10 insects for treated harborage and another four replicates for the control) were evaluated with each strain. The assays were set up at 4 pm. The number of bed bugs resting on and/or under harborages, wandering, or dead in each arena were recorded daily at 4 pm for 7 days.
2.5.2. Two-Choice Harborage Assays
The second choice test involved the use of two harborages. In these trials, the tent of the filter paper discs (55 mm diameter) were treated by spraying directly with IMZOPS 60 (one push for each side of the paper), and then evenly cut into two semicircles. Untreated filter paper discs (55 mm diameter) were evenly cut into two semicircles and served as blank harborages. Both blank and treated semicircles were folded two times and fixed onto zones I (blank) and II (treated) (Figure 1B) respectively, with double-sided adhesive tape (Scotch® Double-Sided Tape 137, 12.7 mm x 11 m, 3M, Australia, ID No.: 70005266401). Both folded harborages were placed away from middle area of the Petri dishes (Figure 1B). Ten adult insects (mixed sexes and ages) were acclimated to middle of the arena for 15 min by containing them in a glass vial as above. Four replicates from the MONH and PARRA strains were evaluated. The assays were set up at 4 pm. The number of bed bugs resting on and/or under both harborages (treated and blank), wandering, or dead in the arenas, were recorded daily at 4 pm for 7 days.
2.6. Data Analysis
In the topical assays evaluating the technical grade broflanilide, mortality over-dose was fitted to a natural log-probit regression model to estimate the dose required to kill either 50% (LD50) or 95% (LD95) of the exposed insects using Polo Plus [20,21]. LD50s were considered significantly different if the 95% confidence intervals (95% CI) did not overlap [22,23]. The Resistance Ratio at the LD50 (RR50) was obtained by dividing the LD50 of resistant strain by that of the susceptible MONH strain.
In the direct spray and residual assays evaluating the broflanilide-based aerosol formulation, the mean accumulative mortality of insects was examined for statistical significance (P < 0.05) using a one-way ANOVA (Tukey’s test) in GraphPad Prism 7.00 (GraphPad Software Inc., San Diego, CA, USA).
In the behavioral response assays, the daily recorded number of bed bugs assembling on/under harborages and wandering in arenas were examined for statistical significance (P < 0.05) using a two-way ANOVA (either Tukey’s or Šídák’s test) in GraphPad Prism 7.00. The survival time data from the residual assays were analyzed using the Survival Analysis in GraphPad Prism 7.00. Kaplan–Meier survival curves from the residual assays were generated and the survival curves were pairwise compared using the log-rank (Mantel-Cox) test. The Bonferroni correction was applied in multiple pairwise comparisons of survival curves in a group [24]. P values from the log-rank test less than α (Bonferroni correction, α = 0.05/K, K is the total number of pairwise comparisons of survival curves in a group) indicate a statistical significance. The values of median survival (MS) were generated from Survival Analysis.
3. Results
3.1. Topical Assays of Technical Grade Broflanilide
Both susceptible and resistant C. lectularius strains showed high susceptibility to broflanilide. The SYD, DARL, and RIPPO strains were susceptible to broflanilide with only 1.22-, 1.03- and 2.77-fold (RR50) difference in the LD50, respectively, when compared to the reference susceptible MONH strain (Table 2). The PARRA strain was less susceptible to broflanilide, with a 3.65- fold (RR50) difference in the LD50, when compared to the reference susceptible MONH strain (Table 2).
3.2. Efficacy of the Broflanilide-Based Aerosol Formulation (IMZOPS 60)
3.2.1. Direct Spray on Adult Bed Bugs
Direct spray of IMZOPS 60 against adult bed bugs of the five C. lectularius strains resulted in high mortality (Figure 2). The MONH, SYD, DARL and RIPPO strains produced 100% mortality after 24 hrs post-treatment, while the PARRA strain had a mean mortality of 97.5±5 % (mean±SD). There was no statistical difference between the susceptible and resistant strains. After 48 hrs post-treatment, the test bed bugs of the PARRA strain were all dead.
3.2.2. Direct Spray on Bed Bug Eggs
Without the treatment of IMZOPS 60 against the eggs (control), hatch rates of both MONH and PARRA strains were high (Figure 3), with rates of 93.0±4.5 % (mean±SD, MONH-CONTROL) and 98.2±1.1 % (PARRA-CONTROL) respectively. With the treatment of IMZOPS 60, the egg hatch rates of both MONH and PARRA strains were significantly lower, declining to 9.7 – 23.4 % (Figure 3). After treatment, 9.7±12 % of MONH eggs (1–2 days old) hatched, while 23.4±29.9 % of eggs (4–5 days old) hatched. For PARRA, 14.9±11.7 % of eggs (1–2 days old) hatched, while 22.6±5.2 % of eggs (4–5 days old) hatched. Statistical analyses showed that hatch rates of the PARRA strain were similar to the reference susceptible MONH strains following treatment with IMZOPS 60 (Figure 3).
3.2.3. Residues against Adult Bed Bugs
The survival time of the SYD, DARL, and RIPPO strains on the residually treated tiles with IMZOPS 60 was longer than the MONH strain, whereas the survival time of PARRA strain was similar to the MONH strain (Figure 4A). Nevertheless, the residues resulted in high mortality to all five strains after 3 days constant exposure, without any statistic differences between the susceptible and resistance strains (Figure 5A). All test bed bugs from the five strains completely died within 6 days post exposure (MONH=2 days, PARRA=3 days, DARL=4 days, SYD=5 days, RIPPO=6 days) (Figure 4A).
For the filter paper (Figure 4B), the survival times of the SYD, DARL, and PARRA strains on the residual IMZOPS 60 was longer than the MONH strain, whereas the survival time of the RIPPO strain was similar to the MONH strain (Figure 4B). Nevertheless, the residues resulted in high mortality to MONH, DARL, PARRA, and RIPPO strains after 3 days exposure (Figure 5B). The mortality of 3 days exposure in the SYD strain was lower than the MONH strain (Figure 5B). Excepting for the PARRA strain, all test bed bugs of four strains completely died within 7 days exposure (MONH = 3 days, SYD = 6 days, DARL = 6 days, RIPPO = 7 days) (Figure 4B), whereas the mortality of the PARRA strain after 7 days exposure was 97.5±5.0 %.
The residues of IMZOPS 60 on tiles killed all the bed bugs slightly quicker than that on filter paper. However, a statistical difference (P <0.05) in the survival time between tiles and paper was only found in the MONH and PARRA strains.
3.2.4. Residues against First Instar Bed Bugs
The residual IMZOPS 60 on the surface of both tiles and filter paper resulted in a similar lethal rate between the MONH and PARRA strains (survival time: MONH-TILE = PARRA-TILE, MONH-PAPER = PARRA-PAPER) (Figure 6). On the treated tiles, all test bed bugs of both the MONH and PARRA strains died after 2 days exposure. On the treated filter paper, all test bed bugs of the MONH strains died after 2 days exposure, while the PARRA strain died after 3 days exposure.
As per the adults, the first instars of both the MONH and PARRA strains died more quickly on the residually treated tiles with IMZOPS 60 than on filter paper (Survival times: MONH-TILE < MONH-PAPER, PARRA-TILE < PARRA-PAPER) (Figure 6). Nevertheless, there were no statistical differences (one-way ANOVA, Turkey’s test, P < 0.05) in mortality between the MONH and PARRA strains after 2 days exposure to residues on the tiles and filter paper (e.g., MONH-TILE = 100 %, MONH-PAPER = 100 %, PARRA-TILE = 100 %, PARRA-PAPER = 92.5±5 %).
3.2.5. Aged Residues against Adult Bed Bugs
The residual efficacy of IMZOPS 60 on tiles was not affected by aging over six months. Survival analysis showed that the survival time of MONH bed bugs on different aged residues of tiles (e.g., 1 WEEK, 2 WEEKS, 4 WEEKS, 3 MONTHS and 6 MONTHS old) was either similar to, or shorter than, that of the fresh residuals (1 DAY old) (e.g., 1 WEEK = 1 DAY, 2 WEEKS = 1 DAY, 4 WEEKS = 1 DAY, 3 MONTHS < 1 DAY, and 6 MONTHS = 1 DAY) (Figure 7A). The different aged residues against the MONH strain resulted in 100% mortality after 2 days exposure, excluding the residues aged for 6 months that resulted in 100 % mortality after 3 days exposure. For the PARRA strain, survival analysis showed that the insect’s survival time on the different aged residues was less than or equal that of the fresh residues (Figure 7B) (e.g., 1 WEEK = 1 DAY, 2 WEEKS <1 DAY, 4 WEEKS = 1 DAY, 3 MONTHS < 1 DAY, and 6 MONTHS = 1 DAY). The different aged residues against the PARRA strain resulted in 100% mortality within 3 days exposure, excluding the residues aged for 6 months that resulted in 100% mortality after 6 days exposure.
Similar to the tiles, the residual efficacy of IMZOPS 60 on filter paper was not affected by aging. Survival analysis showed that the survival time of MONH and PARRA bed bugs on the different aged residues of filter paper was comparable to that of fresh residues (Figure 8). The different aged residues achieved high mortality against the MONH strain after 3 days exposure (mortality [mean±SD]: 1 DAY = 100 %, 1 WEEK = 100 %, 2 WEEKS = 97.5±5 %, 4 WEEKS = 97.5±5 %, 3 MONTHS = 95±5.8 %, and 6 MONHS = 92.5±9.6 %) (one-way ANOVA, Turkey’s test, P > 0.05), and to PARRA strain after 7 days exposure (mortality [mean±SD]: 1 DAY = 97.5±5 %, 1 WEEK = 90±8.2 %, 2 WEEKS =97.5±5 %, 4 WEEKS = 92.5±9.6 %, 3 MONTHS = 90.2±8.5 %, and 6 MONHS = 92.5±5 %) (one-way ANOVA, Turkey’s test, P > 0.05).
3.3. Behavioral Responses to Harborages Treated with IMZOPS 60
3.3.1. One-Choice Harborage Assays
Bed bugs from both the MONH and PARRA strains did not avoid resting on and/or under harborages treated with IMZOPS 60 (Figure 9 and Figure 10). For the MONH strain (Figure 9), the daily recorded number of bed bugs assembling on and/or under harborages treated with the IMZOPS 60 was similar to that of wandering bed bugs in DAY 1, 2, 6, and 7, but more than that of wandering bed bugs in DAY 3, 4, and 5. In the control, the number of bed bugs assembling on and/or under untreated harborages was similar to that of wandering bed bugs in DAY 1 to 6, but more than that of wandering bed bugs in DAY 7. The number of bed bugs assembling on or under treated harborages were comparable to that on or under untreated harborages in each individual day.
For the PARRA strain (Figure 10), the daily recorded number of bed bugs assembling on and/or under treated harborages were greater than wandering bed bugs in each individual day. In the control, the number of bed bugs assembling on and/or under untreated harborages were similar to that of wandering bed bugs over DAY 1 to 6, but more than that of wandering bed bugs on DAY 5 and 7. The number of bed bugs assembling on and/or under treated harborages were similar to that on and/or under untreated harborages for each individual day. After 7 days exposure, the accumulative mortality of the MONH strain in treated harborages was 52.5±20.6 %, while that of the PARRA strain was 30±21.6 %. The control mortality of the MONH strain was 2.5±5 %, while that of the PARRA strain was 10±8.2 %.
3.3.2. Two-Choice Harborage Assays
The two-choice assay results also indicated that there was no avoidance to IMZOPS 60 treated harborages with both the MONH and PARRA strains (Figure 11 and Figure 12). In the MONH strain (Figure 11), excluding DAY 2, the daily recorded number of bed bugs assembling on and/or under treated harborages were similar to that assembling on or under untreated harborages on each individual day. In Day 2, the bed bugs from the MONH strain on and/or under treated harborages were greater than that on and/or under the untreated harborages. In the PARRA strain (Figure 12), the daily recorded number of bed bugs assembling on and/or under the treated harborages were similar to that assembling on and/or under the untreated harborages on each individual day. After 7 days exposure, the mortality of the MONH strain was 62.5±17.1 %, while mortality of the PARRA strain was 32.5±26.3 %.
4. Discussion
This study is the first report to examine the susceptibility of the common bed bug, Cimex lectularius to broflanilide. The experiments demonstrated that the resistant SYD, DARL, and RIPPO strains of the common bed bug, C. lectularius were susceptible to broflanilide, when compared to the reference susceptible MONH strain. The resistant PARRA C. lectularius strain was slightly more tolerant to broflanilide, which may be due to a form of insecticide resistance. For this strain, cuticular, metabolic, and knockdown resistance has previously been reported [16,25,26]. While it is unlikely that the knockdown resistance would not contribute to any tolerance (due to the mode of action), the presence of a thicker cuticle is likely to slow down the penetration of the insecticide into the insect, thereby leading to a higher resistance ratio [1]. However, it is also important to note that the recorded resistance ratio for the PARRA strain against broflanilide of 3.65-fold is still extremely low in comparison to other insecticides. For example, the Sydney strain of Cimex lectularius against permethrin, was found to have a resistance ratio of 1,235,000 [27].
Historically, the common bed bug C. lectularius has largely been distributed in temperate regions, while the tropical bed bug C. hemipterus, has mainly been confined to tropical and subtropical regions [15]. Recently, the distribution of two bed bug species have increasingly overlapped [1,28]. Cimex hemipterus has been recorded in temperate regions over last decade, including Central Europe [29], France [30], Korea [31], Russia (Moscow, Smolensk, Saransk, and St Petersburg [32], and Voronezh [33]), and the UK [34]. The two bed bug species display varying resistance levels and resistance mechanisms to insecticides [1,35,36], with Cimex hemipterus having higher levels of resistance to insecticides than C. lectularius [36,37,38]. Therefore, considering this, studies into the susceptibility of C. hemipterus to broflanilide are warranted prior to field use against this species.
As modern field strains of bed bugs are resistant to most classes of insecticides [6], products containing actives with new modes of action are urgently needed for bed bug management. Broflanilide appeared to be a potentially promising candidate as it is a novel insecticide with a unique mode of action (IRAC Group 30). This was confirmed with the studies herein, demonstrating that C. lectularius, including modern field strains, were highly susceptible to broflanilide, and hence the active is a viable choice for the management of these insects. Our laboratory evaluations also confirmed the efficacy of a broflanilide-based aerosol product (IMZOPS 60) to one susceptible and four resistant C. lectularius strains (both as a direct topical spray and as a dry residual residue). The experiments demonstrated that IMZOPS 60 is highly effective at killing both susceptible and resistance strains, as well as eggs and first instars. In the direct spray assays, IMZOPS 60 resulted in 100% mortality of all test bed bugs (susceptible and resistant strains) within 24 hrs post-treatment, excepting the resistant PARRA strain, which had 100% mortality after 48 hrs post-treatment. Egg hatch rates of both susceptible (MONH) and resistant (PARRA) strains were significantly reduced with IMZOPS 60 spray, and were comparable in terms of efficacy with both strains.
Previous studies have found that a broflanilide formulation (Tenebenal™) has slower residual efficacy against mosquitoes, compared with most widely employed insecticides, such as the pyrethroids [17]. Similarly, our residual assays found that residues of IMZOPS 60 on both smooth non-porous (ceramic tiles) and porous (filter paper) surfaces have a slower effect on bed bugs. For example, it took up to 3 days for all the first instars of both MONH and PARRA strains to die following residual exposure. In the case of the adult insects, the residues on tiles and filter paper resulted in high mortality to the susceptible MONH, and the resistant SYD, DARL, PARRA and RIPPO strains within 3 days of exposure. However, excluding the PARRA strain, all strains achieved 100% mortality within 7 days (MONH=2–3 days, SYD=5–6 days, DARL=4–6 days, RIPPO=6–7 days). With the PARRA strain when exposed to IMZOPS 60, mortality reached 100% after 3 days exposure on the tiles, and 97.5% on filter paper after 7 days constant exposure. Considering the slow efficacy observed, a repeated inspection after 7 days post-treatment should be considered as best practice in bed bug management using broflanilide-based products.
The residual experiments conducted herein, were all forced exposure trials. However, in the field, it may be that bed bugs will not be constantly confined to insecticide treated areas, as the insects harbor in cracks and crevices where insecticide penetration is challenging to achieve. Therefore, the influences of exposure time and mortality assessment interval, as per Leong and colleagues [39], should be investigated in future efficacy studies involving broflanilide. Fortunately, C. lectularius did not avoid surfaces treated with the insecticide, which will ensure a longer exposure with through applications.
Compared to the nymphs and adults of bed bugs, relatively little research has been undertaken on the resistance status of the egg stage [40]. However, the age of the eggs can vary in resistance to insecticides due to the combined effects of the eggshell development and resistant mechanisms [40,41,42]. Compared to the susceptible Harlan C. lectularius strain, Campbell and Miller [41] found that eggs of two pyrethroid-resistant C. lectularius strains were resistant to imidacloprid/β-cyfluthrin (Temprid SC) with 3- and 5.1-fold difference in the LC50, and to acetamiprid/bifenthrin (Transport GHP) with 39- and 1,080-fold difference in the LC50. Hinson and colleagues (2016) [40] found that Temprid SC was the most effective insecticide at preventing egg hatch (13% hatch rate) for a pyrethroid-resistant C. lectularius strain, compared to the insecticides of Bedlam (MGK-264, sumithrin [84% hatch rate]), Demand CS (lambda-cyhalothrin [91% hatch rate]), and Phantom SC (chlorfenapyr [95% hatch rate]). Compared to the susceptible Monheim C. lectularius strain, Leong and colleagues (2021) [42] also found that eggs from five pyrethroid-resistant C. hemipterus strains showed high to very high resistance against the insecticides of Tandem M (thiamethoxam/lambda-cyhalothrin), Temprid SC, Sumithion CS (fenitrothion), Pesguard FG161 EC (d-tetramethrin/cyphenothrin), and Sumthrin ME (d-phenothrin). In comparison, our studies found that the broflanilide-based aerosol (IMZOPS 60) resulted in low egg hatch rates for the resistant PARRA strain (14.9–22.6%), which was similar to the susceptible (MONH) strain (9.7–23.4%). With most aerosols, the constant pressing of the actuator (trigger or button) results in a continual flow of spray. In contrast, the flow stops once the actuator of IMZOPS 60 is fully depressed, meaning that the action is more like a pump-style aerosolizer. Perhaps this pump-style aerosolizer may have resulted in uneven spraying of the IMZOPS 60 on the bed bug eggs, leading to eegs being untreated and hence a small number of eggs from both MONH and PARRA strains hatching. However, because of the high residual efficacy, the residues of IMZOPS 60 still will kill the first instars that subsequently hatch. Consequently, IMZOPS 60 can be considered an effective ovicide for bed bug management.
The type of substrate surface (e.g., non-porous versus porous) has been found to affect the residual efficacy of insecticides against pests including bed bugs [36,43,44,45]. Generally, liquid insecticides applied on a non-porous surface provides higher performance than a porous surface. For example, Wang and colleagues [46] revealed that the residual efficacy of Tandem on vinyl (non-porous) was higher than that on fabric (porous) against C. lecularius, although the residual efficacy of Temprid SC, Transport GHP and Demand CS on vinyl was similar to that on fabric. Dang and colleagues [36] found that both Temprid SC and Tandem residues on glass surface killed resistant C. hemipterus strains significantly faster than that on filter paper. The residues of Temprid SC and Tandem on filter paper failed to control bed bugs for the duration of the experiment with mortality less than 50% after 120 h of continual exposure. Compared with liquid insecticides, the performance of aerosol formulated insecticides applied on various substrates against bed bugs can be unpredictable [46]. Furthermore, in the field, insecticide applications for bed bug management are typically applied onto porous surface substrates, particularly as bed bugs tend to avoid smooth non-porous surfaces [46]. It is worth noting that our studies found that different substrates sprayed with IMZOPS 60 had little impact on overall residual mortality, although the time to achieve complete mortality was slightly longer on the filter paper compared to the tiles.
Aging is known to impact the residual efficacy of insecticides, especially against bed bugs [36,47]. However, aging residually treated surfaces to six months had little impact with the residual efficacy of IMZOPS 60. Our studies found that the residues of IMZOPS 60 on tiles and filter paper that were aged indoors for 1 WEEK, 2 WEEKS, 4 WEEKS, 3 MONTHS, and 6 MONTHS, were all equally effective as fresh residues (1 DAY). Our results confirmed previous investigations that found broflanilide-based formulation had long residual activity on the surfaces of ceramic, wood, and cement tiles, against mosquitoes (Lee et al. 2020) [17]. As noted above, the behavioral response assays revealed that C. lectularius from the susceptible MONH and resistant PARRA strains did not avoid surfaces treated by IMZOPS 60. This means that with the long residual life of the insecticide and the non-avoidance of treated residues by C. lectularius, broflanilide will aid in reducing the need for repeated applications of the insecticide. An added benefit will be the prevention of new infestations in areas previously treated with the product. In highly infested locations such as low-income housing, broflanilide products could be applied prophylactically to prevent the establishment of new infestations, and ultimately should significantly reduce control costs.
5. Conclusions
In summary, current bed bug strains are highly resistant to most insecticide classes. Despite this, insecticide treatments remain one of the most common strategies employed to treat bed bug infestations, largely due to the lower treatment costs. Thus, insecticides with new modes of action that prove to be effective will be of great benefit for the control of modern resistant bed bug strains. The novel insecticide, broflanilide, has shown to be a highly promising candidate for bed bug management. Both susceptible and resistant strains of C. lectularius were found to be highly susceptible to technical grade broflanilide as well as a broflanilide-based aerosol product. Plus, the insecticide is highly stable and effective on different substrates. While the mode of action is relatively slow, the high efficacy (including high ovicidal activity), long residual action, and a lack of avoidance behavior, makes broflanilide a highly suitable insecticide for bed bug control.
Author Contributions
Conceptualization, K.D. and S.L.D.; methodology, K.D. and S.L.D.; investigation, K.D. and S.L.D.; data curation, K.D.; writing—original draft preparation, K.D.; writing—review and editing, K.D. and S.L.D.; supervision, S.L.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available in this article.
Acknowledgments
All insecticides including technical grade broflanilide and the broflanilide-based aerosol product (IMZOPS 60) were supplied by Earth Corporation, Japan. Earth Corporation, Japan, provided financial support to cover the publication costs of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dang, K.; Doggett, S.L.; Singham, G.V.; Lee, C.Y. Insecticide resistance and resistance mechanisms in bed bugs, Cimex spp. (Hemiptera: Cimicidae). Parasit. Vectors 2017, 10, 1–30. [Google Scholar] [CrossRef]
- oggett, S. L., D. M. Miller, and C. Y. Lee. 2018. Advances in the Biology and Management of Modern Bed Bugs; Wiley-Blackwell: Oxford, United Kingdom, 2018. [Google Scholar]
- Doggett, S.L.; Dwyer, D.E.; Penas, P.F.; Russell, R.C. Bed bugs: clinical relevance and control options. Clin. Microbiol. Rev. 2012, 25, 164–192. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Potter, M.F.; Potter, D.A.; Haynes, K.F. Insecticide resistance in the bed bug: a factor in the pest's sudden resurgence? J. Med. Entomol. 2007, 44, 175–178. [Google Scholar] [PubMed]
- Potter, M.F. 2011. The history of bed bug management – with lessons from the past. Am. Entomol. 2011, 57, 14–25. [Google Scholar] [CrossRef]
- Lee, C. Y., D. M. Miller, and S. L. Doggett. 2018. Chapter 30: Chemical control. In Advances in the Biology and Management of Modern Bed Bugs; Doggett, S.L., Miller, D.M., Lee, C.Y., Eds.; Wiley-Blackwell: Oxford, United Kingdom, 2018; pp. 311–321. [Google Scholar]
- Doggett, S.L.; Lee, C.Y. Historical and contemporary control options against bed bugs, Cimex spp. Ann. Rev. Entomol. 2023, 68, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.F.; Haynes, K.F.; Fredericks, J. Bed bugs across America: the 2015 bed bugs without borders survey. PestWorld 2015, November/December, 4–14. [Google Scholar]
- Katsuta, H.; Nomura, M.; Wakita, T.; Daido, H.; Kobayashi, Y.; Kawahara, A.; Banba, S. Discovery of broflanilide, a novel insecticide. J. Pestic. Sci. 2019, 44, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Insecticide Resistance Action Committee (IRAC). Available online: https://irac-online.org (access on 28 March 2024).
- Govoetchan, R.; Fongnikin, A.; Syme, T.; Small, G.; Gbegbo, M.; Todjinou, D.; Rowland, M.; Nimmo, D.; Padonou, G.G.; Ngufor, C. VECTRONTM T500, a new broflanilide insecticide for indoor residual spraying, provides prolonged control of pyrethroid-resistant malaria vectors. Malar. J. 2022, 21, 324. [Google Scholar] [CrossRef] [PubMed]
- van Herk, W.G.; Warren, R.L.; Bailey, T. Wireworm (Coleoptera: Elateridae) intoxication symptoms indicate the postharvest presence of broflanilide residues in soil collected from potato fields. J. Econ. Entomol. 2023, 116, 1998–2008. [Google Scholar] [CrossRef]
- Tang, T.; Hu, F.; Wang, P.; Fu, W.; Liu, X. Broflanilide effectively controls Helicoverpa armigera and Spodoptera exigua exhibiting diverse susceptibilities to chlorantraniliprole and emamectin benzoate. Pest Manag. Sci. 2021, 77, 1262–1272. [Google Scholar] [CrossRef]
- Australian Pesticides and Veterinary Medicines Authority (APVMA). Available online: https://apvma.gov.au (access on 28 March 2024).
- Usinger, R.L. Monograph of Cimicidae (Hemiptera - Heteroptera); Entomological Society of America: College Park, Maryland, USA, 1966. [Google Scholar]
- Dang, K.; Toi, C.S.; Lilly, D.G.; Bu, W.; Doggett, S. L. Detection of knockdown resistance (kdr) mutations in the common bed bug, Cimex lectularius (Hemiptera: Cimicidae) in Australia. Pest Manag. Sci. 2015, 71, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.S.; Ambrose, P.; Williams, J.; Morgan, J.; Praulins, G.; Ingham, V.A.; Williams, C.T.; Logan, R.A.E.; Ismail, H.M.; Malone, D. Tenebenal: a meta-diamide with potential for use as a novel mode of action insecticide for public health. Malar. J. 2020, 19, 398. [Google Scholar]
- Majid, A.H.A.; Zulaikha, Z. Laboratory bioassay on efficacy of dual mode of action insecticides (beta-cyfluthrin and imidacloprid) towards tropical bed bugs, Cimex hemipterus (Hemiptera: Cimicidae). J. Entomol. Zool. Stud. 2015, 3, 217–220. [Google Scholar]
- Romero, A.; Potter, M.F.; Haynes, K.F. Evaluation of chlorfenapyr for control of the bed bug, Cimex lectularius L. Pest Manag. Sci. 2010, 66, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: London, United Kingdom, 1971. [Google Scholar]
- Robertson, J.A.; Preisler, H.K.; Russell, R.M. Polo Plus: Probit and Logit Analysis, User’s Guide; LeOra Software: Petaluma, CA, USA, 2003. [Google Scholar]
- Payton, M.E.; Greenstone, M.H.; Schenker, N. Overlapping confidence intervals or standard error intervals: what do they mean in terms of statistical significance? J. Insect Sci. 2003, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.W.; Park, R.M.; Barler, A.J. Comparing median lethal concentration values using confidence interval overlap or ratio tests. Environ. Toxicol. Chemist. 2006, 25, 1441–1444. [Google Scholar] [CrossRef] [PubMed]
- Multiple comparisons of survival curves. Available online: https://www.graphpad.com/guides/prism/latest/statistics/stat_multiple_comparisons_of_surviv.htm (accessed on 28 March 2024).
- Lilly, D.G.; Latham, S.L.; Webb, C.E.; Doggett, S.L. Cuticle thickening in a pyrethroid-resistant strain of the common bed bug, Cimex lectularius L. (Hemiptera: Cimicidae). PLoS One 2016, 11, e0153302. [Google Scholar] [CrossRef] [PubMed]
- Lilly, D.G.; Dang, K.; Webb, C.E.; Doggett, S.L. Evidence for metabolic pyrethroid resistance in the common bed bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2016, 109, 1364–1368. [Google Scholar] [CrossRef] [PubMed]
- Lilly, D.G.; Zalucki, M.P.; Orton, C.J.; Russell, R.C.; Webb, C.E.; Doggett, S.L. Confirmation of insecticide resistance in Cimex lectularius (Hemiptera: Cimicidae) in Australia. Austral Entomol. 2015, 54, 96–99. [Google Scholar] [CrossRef]
- Štefka, J.; Votýpka, J.; Lukeš, J.; Balvín, O. Cimex lectularius and Cimex hemipterus (bed bugs). Trends Parasitol. 2022, 38, 919–920. [Google Scholar] [CrossRef]
- Balvìn, O.; Sasinkova, M.; Martinu, J.; Nazarizadeh, M.; Bubova, T.; Booth, W.; Vargo, E.L.; Štefka, J. Early evidence of establishment of the tropical bed bug (Cimex hemipterus) in Central Europe. Med. Vet. Entomol. 2021, 35, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Chebbah, D.; Elissa, N.; Sereno, D.; Hamarsheh, O.; Marteau, A.; Jan, J.; Izri, A.; Akhoundi, M. Bed bugs (Hemiptera: Cimicidae) population diversity and first record of Cimex hemipterus in Paris. Insects 2021, 12, 578. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Shin, E.H.; Ju, H.C.; Jeong, E.S.; Lee, S.H.; Kim, J.H. The first recent case of Cimex hemipterus (Hemiptera: Cimicidae) with super-kdr mutations in the Republic of Korea. J. Med. Entomol. 2023, 60, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Gapon, D. First records of the tropical bed bug Cimex hemipterus (Heteroptera: Cimicidae) from Russia. Zoosystematica Ross. 2016, 25, 239–242. [Google Scholar] [CrossRef]
- Golub, V.B.; Aksenenkoa, E.V.; Sobolevaa, V.A.; Kornevb, I.I. New data on the distribution of the tropical bed bug Cimex hemipterus and the western conifer seed bug Leptoglossus occidentalis (Heteroptera: Cimicidae, Coreidae) in the European Part of Russia. Russ. J. Biol. Invasions 2020, 11, 97–100. [Google Scholar] [CrossRef]
- Burgess, I. Bugs of the past—or are they on the up? Professional Pest Controller 2003, 32, 16–17. [Google Scholar]
- Dang, K.; Doggett, S.L.; Leong, X.Y.; Veera Singham, G.; Lee, C.Y. Multiple mechanisms conferring broad-spectrum insecticide resistance in the tropical bed bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2021, 114, 2473–2484. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Doggett, S.L.; Lee, C.Y. Performance of pyrethroid-neonicotinoid mixture formulations against field-collected strains of the tropical bed bug (Hemiptera: Cimicidae) on different substrates. J. Econ. Entomol. 2023, 113, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Lilly, D.G.; Dang, K.; Webb, C.E.; Doggett, S.L. Are Australian field-collected strains of Cimex lectularius and Cimex hemipterus (Hemiptera: Cimicidae) resistant to deltamethrin and imidacloprid as revealed by topical assay? Austral Entomol. 2018, 57, 77–84. [Google Scholar] [CrossRef]
- Doggett, S.L.; Cooper, R. Back with a bite: bed bugs 25 years on! FAOPMA Magazine 2024, January, 16–53. Available online: https://rb.gy/j7mnbn (access on 28 March 2024).
- Leong, X.Y.; Veera Singham, G.; Chong Shu-Chien, A.; Doggett, S.L.; Lee, C.Y. Influences of exposure time and mortality assessment interval on bioassay results of insecticide-resistant tropical bed bugs (Hemiptera: Cimicidae). Insects 2020, 11, 640. [Google Scholar] [CrossRef] [PubMed]
- Hinson, K.R.; Benson, E.P.; Zungoli, P.A.; Bridges Jr., W.C.; Ellis, B.R. Egg hatch rate and nymphal survival of the bed bug (Hemiptera: Cimicidae) after exposure to insecticide sprays. J. Econ. Entomol. 2016, 109, 2495–2499. [Google Scholar] [CrossRef]
- Campbell, B.E.; Miller, D. M. Insecticide resistance in eggs and first instars of the bed bug, Cimex lectularius (Hemiptera: Cimicidae). Insects 2015, 6, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Leong, X.Y.; Kim, D.Y.; Dang, K.; Singham, G.V.; Doggett, S.L.; Lee, C.Y. Performance of commercial insecticide formulations against different developmental stages of insecticide-resistant tropical bed bugs (Hemiptera: Cimicidae). J. Econ. Entomol. 2020, 113, 353–366. [Google Scholar] [CrossRef]
- Chadwick, P.R. Surfaces and other factors modifying the effectiveness of pyrethroids against insects in public health. Pestic. Sci. 1985, 6, 383–391. [Google Scholar] [CrossRef]
- Rust, M. K. 1995. Factors affecting control with residual insecticide deposits. In Understanding and controlling the German cockroach; Rust, M.K., Owens, J.M., Reierson, D.A., Eds.; Oxford University Press: New York City, NY, USA, 1995; pp. 149–169. [Google Scholar]
- Dang, K.; Singham, G.V.; Doggett, S.L.; Lilly, D.G.; Lee, C.Y. Effects of different surfaces and insecticide carriers on residual insecticide bioassays against bed bugs, Cimex spp. (Hemiptera: Cimicidae). J. Econ. Entomol. 2017, 110, 558–566. [Google Scholar]
- Wang, C.; Singh, N.; Zha, C.; Cooper, R. Efficacy of selected insecticide sprays and aerosols against the common bed bug. Cimex lectularius (Hemiptera: Cimicidae). Insects 2016, 7, 5. [Google Scholar] [PubMed]
- Fletcher, M.G.; Axtell, R.C. Susceptibility of the bedbug, Cimex lectularius, to selected insecticides and various treated surfaces. Med. Vet. Entomol. 1993, 7, 69–72. [Google Scholar] [CrossRef]
Figure 1.
Arenas for behavioral responses to harborages treated with IMZOPS 60. (B) Arena for one-choice assay; (A). Arena for two-choice assay.
Figure 1.
Arenas for behavioral responses to harborages treated with IMZOPS 60. (B) Arena for one-choice assay; (A). Arena for two-choice assay.

Figure 2.
Mortality (mean±SD) of the susceptible MONH strain and four resistant strains after 24 h post-treatment. Lowercase letters (a) above each column indicate no statistical difference (one-way ANOVA, Turkey’s test, P > 0.05).
Figure 2.
Mortality (mean±SD) of the susceptible MONH strain and four resistant strains after 24 h post-treatment. Lowercase letters (a) above each column indicate no statistical difference (one-way ANOVA, Turkey’s test, P > 0.05).
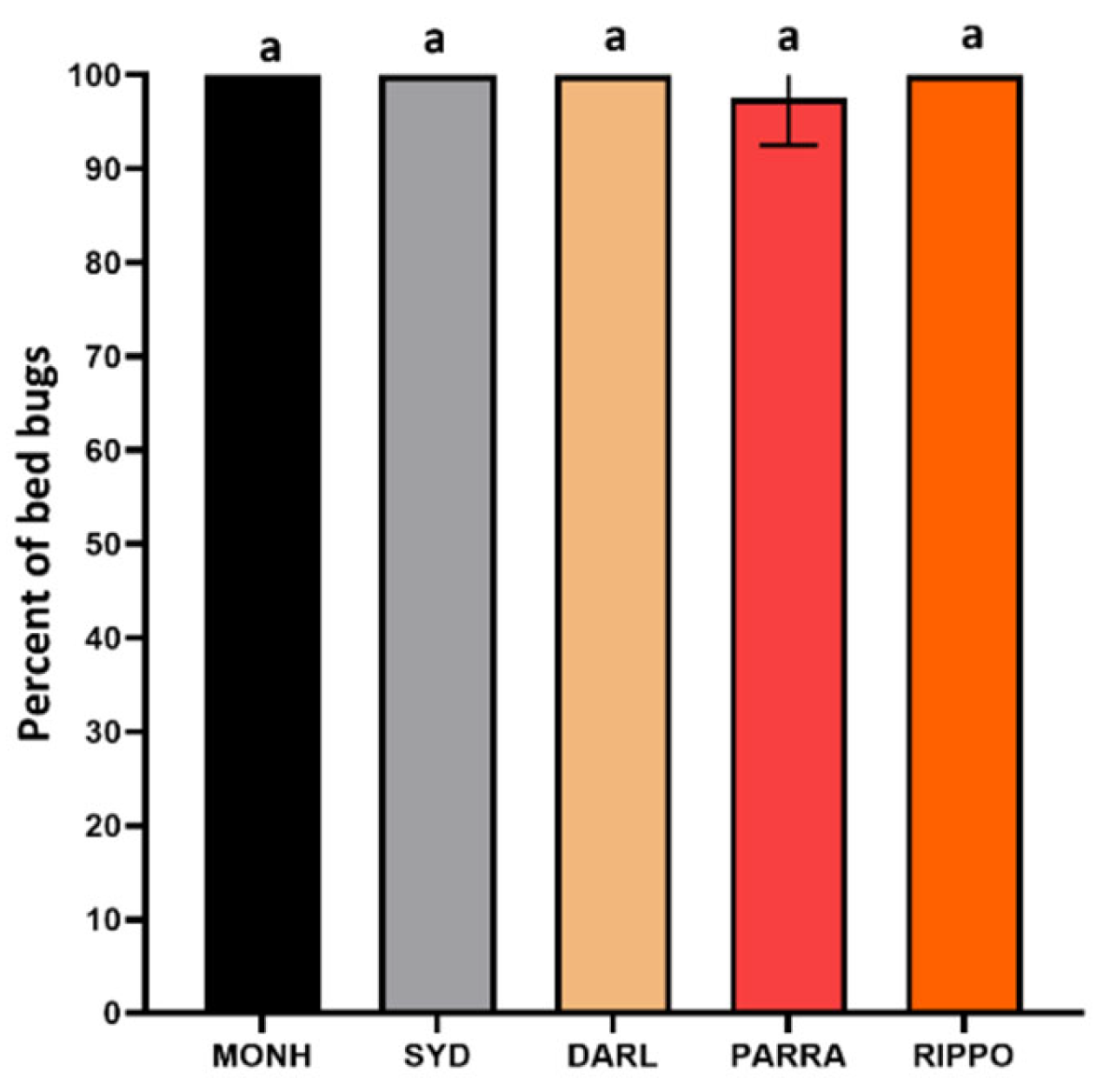
Figure 3.
Hatch rates of three groups of the MONH and PARRA eggs. GROUP ONE = control, including MONH-CONTROL and PARRA-CONTROL. GROUP TWO = 1–2 days old eggs treated by IMZOPS 60, including MONH-TREATED (1-2 DAYS) and PARRA-TREATED (1-2 DAYS). GROUP THREE = 4–5 days old eggs treated by IMZOPS 60, including MONH-TREATED (4-5 DAYS) and PARRA-TREATED (4-5 DAYS). Lowercase letters (a, b) adjacent to the error bars indicate statistical differences (one-way ANOVA, Turkey’s test, P < 0.05).
Figure 3.
Hatch rates of three groups of the MONH and PARRA eggs. GROUP ONE = control, including MONH-CONTROL and PARRA-CONTROL. GROUP TWO = 1–2 days old eggs treated by IMZOPS 60, including MONH-TREATED (1-2 DAYS) and PARRA-TREATED (1-2 DAYS). GROUP THREE = 4–5 days old eggs treated by IMZOPS 60, including MONH-TREATED (4-5 DAYS) and PARRA-TREATED (4-5 DAYS). Lowercase letters (a, b) adjacent to the error bars indicate statistical differences (one-way ANOVA, Turkey’s test, P < 0.05).
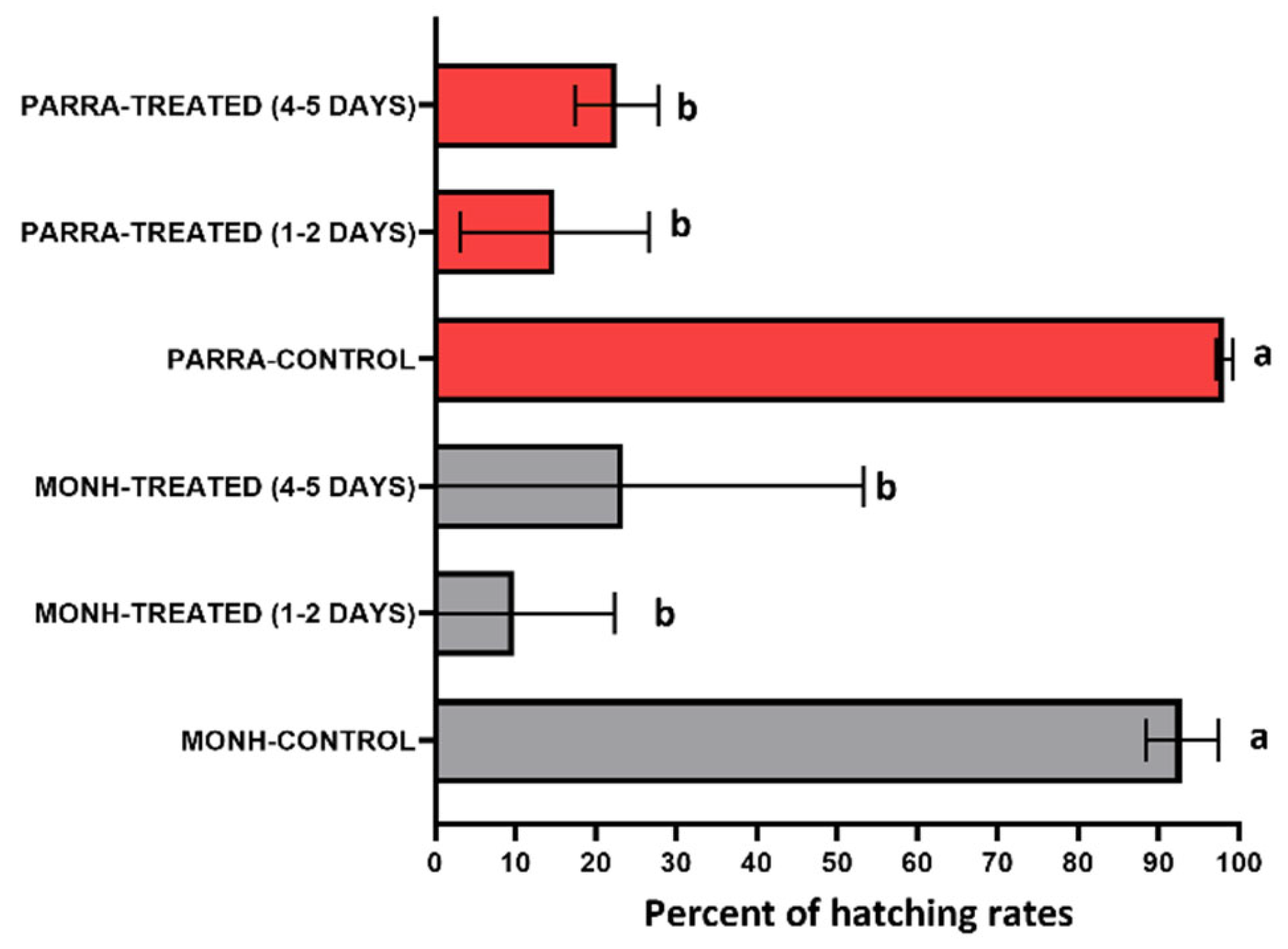
Figure 4.
Kaplan–Meier survival analyses for the residual efficacy of IMZOPS 60 on tiles (A) and filter paper (B) against the five strains of C. lectularius. MS = median survival time (day). Lowercase letters (a, b) following the values of MS indicate statistical differences between the susceptible strain and each resistant strain (Log-rank test, P value is less than α, [α = 0.05/4, Bonferroni correction, 4 is the number of pairwise comparisons (e.g., MONH vs SYD, MONH vs DARL, MONH vs PARRA, MONH vs RIPPO) of survival curves in either (A) or (B)]).
Figure 4.
Kaplan–Meier survival analyses for the residual efficacy of IMZOPS 60 on tiles (A) and filter paper (B) against the five strains of C. lectularius. MS = median survival time (day). Lowercase letters (a, b) following the values of MS indicate statistical differences between the susceptible strain and each resistant strain (Log-rank test, P value is less than α, [α = 0.05/4, Bonferroni correction, 4 is the number of pairwise comparisons (e.g., MONH vs SYD, MONH vs DARL, MONH vs PARRA, MONH vs RIPPO) of survival curves in either (A) or (B)]).
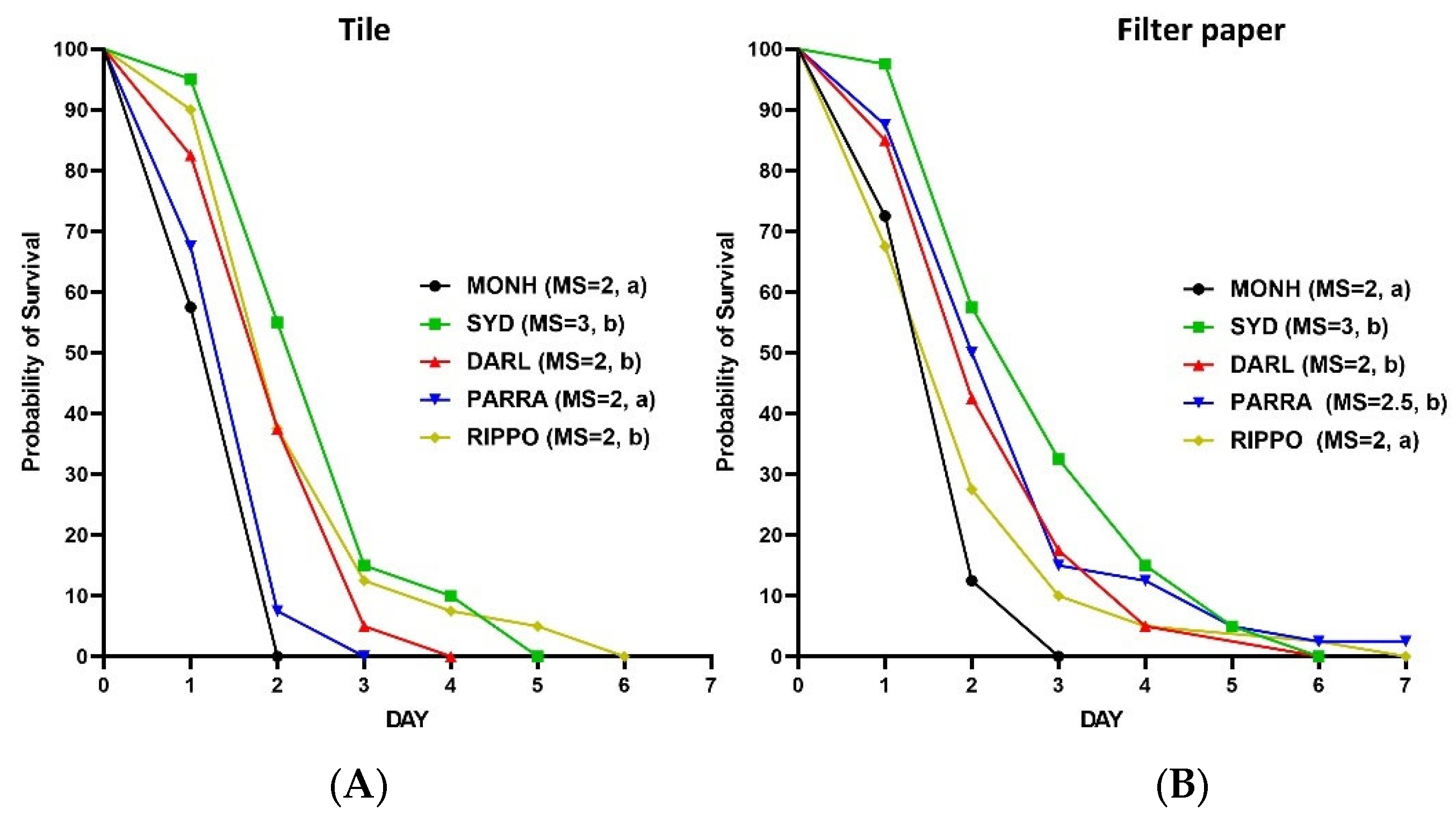
Figure 5.
Mortality (mean±SD %) of all five strains after 3 days exposure on the residues of either tiles (A) or filter paper (B). Lowercase letters (a, b) above the error bars indicate statistical differences (one-way ANOVA, Turkey’s test, P < 0.05).
Figure 5.
Mortality (mean±SD %) of all five strains after 3 days exposure on the residues of either tiles (A) or filter paper (B). Lowercase letters (a, b) above the error bars indicate statistical differences (one-way ANOVA, Turkey’s test, P < 0.05).
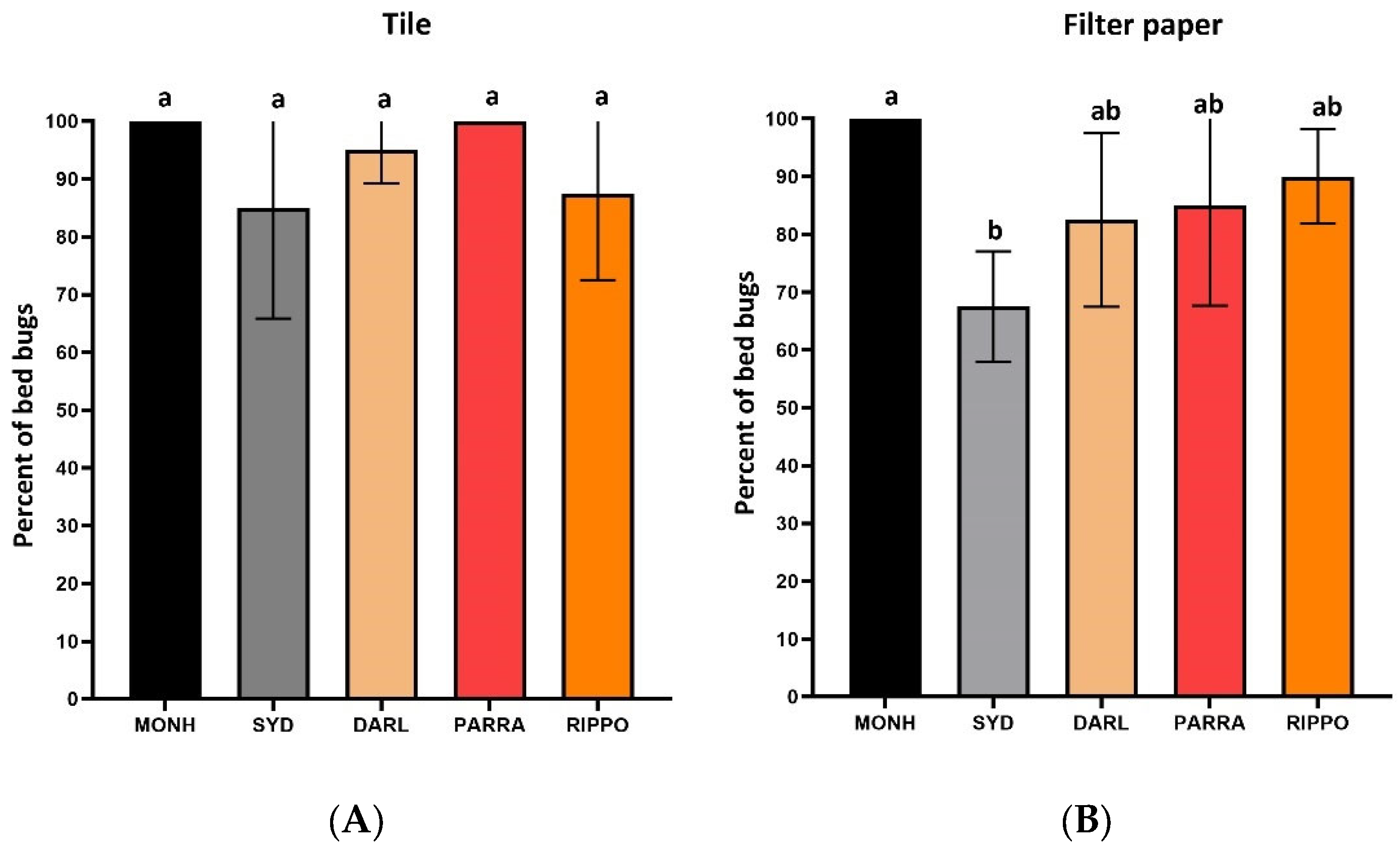
Figure 6.
Kaplan–Meier survival analyses for the residual efficacy of IMZOPS 60 on tiles (TILE) and filter paper (PAPER) against the first instars of MONH and PARRA strains. MS = median survival time (day). Lowercase letters (a, b, c) indicate statistical differences between each trial (Log-rank test, P value is less than α, [α=0.05/6, Bonferroni correction, 6 is the number of pairwise comparisons of survival curves (e.g., MONH-TILE vs PARRA-TILE, MONH-TILE vs MONH-PAPER, MONH-TILE vs PARRA-PAPER, PARRA-TILE vs MOMH-PAPER, PARRA-TILE vs PARRA-PAPER, MONH-PAPER vs PARRA-PAPER)]).
Figure 6.
Kaplan–Meier survival analyses for the residual efficacy of IMZOPS 60 on tiles (TILE) and filter paper (PAPER) against the first instars of MONH and PARRA strains. MS = median survival time (day). Lowercase letters (a, b, c) indicate statistical differences between each trial (Log-rank test, P value is less than α, [α=0.05/6, Bonferroni correction, 6 is the number of pairwise comparisons of survival curves (e.g., MONH-TILE vs PARRA-TILE, MONH-TILE vs MONH-PAPER, MONH-TILE vs PARRA-PAPER, PARRA-TILE vs MOMH-PAPER, PARRA-TILE vs PARRA-PAPER, MONH-PAPER vs PARRA-PAPER)]).
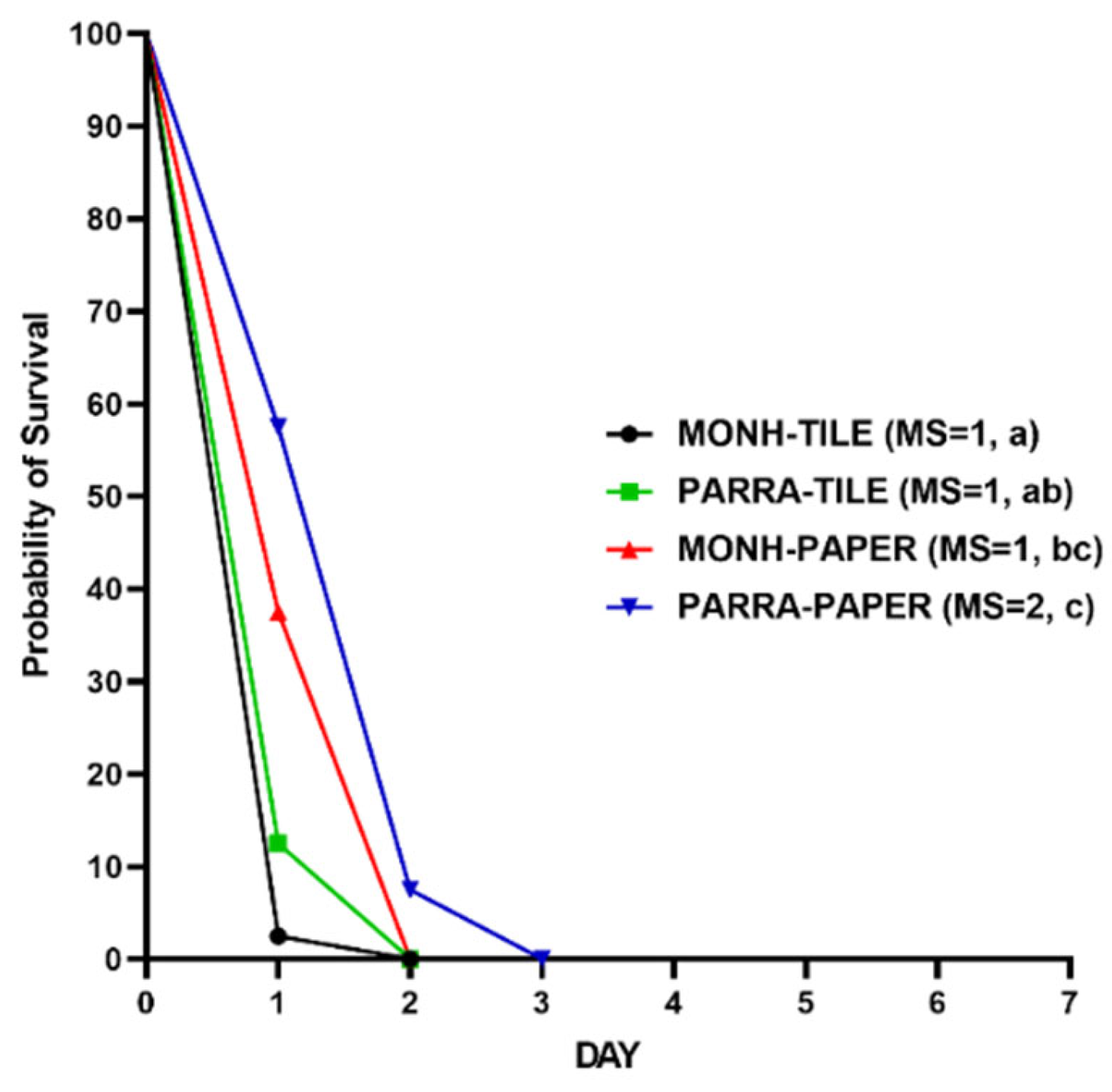
Figure 7.
Kaplan–Meier survival analyses for the efficacy of the aged residual IMZOPS 60 on tiles against MONH (A) and PARRA (B) strains. MS=median survival time (day). Lowercase letters (a,b) indicate statistical differences between the fresh residues and each aged residues (Log-rank test, P value is less than α, [α=0.05/5, Bonferroni correction, 5 is the number of pairwise comparisons (e.g., 1 DAY vs 1 WEEK, 1 DAY vs 2 WEEKS, 1 DAY vs 4 WEEKS, 1 DAY vs 3 MONTHS, and 1 DAY vs 6 MONTHS) of survival curves in either (A) or (B)]).
Figure 7.
Kaplan–Meier survival analyses for the efficacy of the aged residual IMZOPS 60 on tiles against MONH (A) and PARRA (B) strains. MS=median survival time (day). Lowercase letters (a,b) indicate statistical differences between the fresh residues and each aged residues (Log-rank test, P value is less than α, [α=0.05/5, Bonferroni correction, 5 is the number of pairwise comparisons (e.g., 1 DAY vs 1 WEEK, 1 DAY vs 2 WEEKS, 1 DAY vs 4 WEEKS, 1 DAY vs 3 MONTHS, and 1 DAY vs 6 MONTHS) of survival curves in either (A) or (B)]).
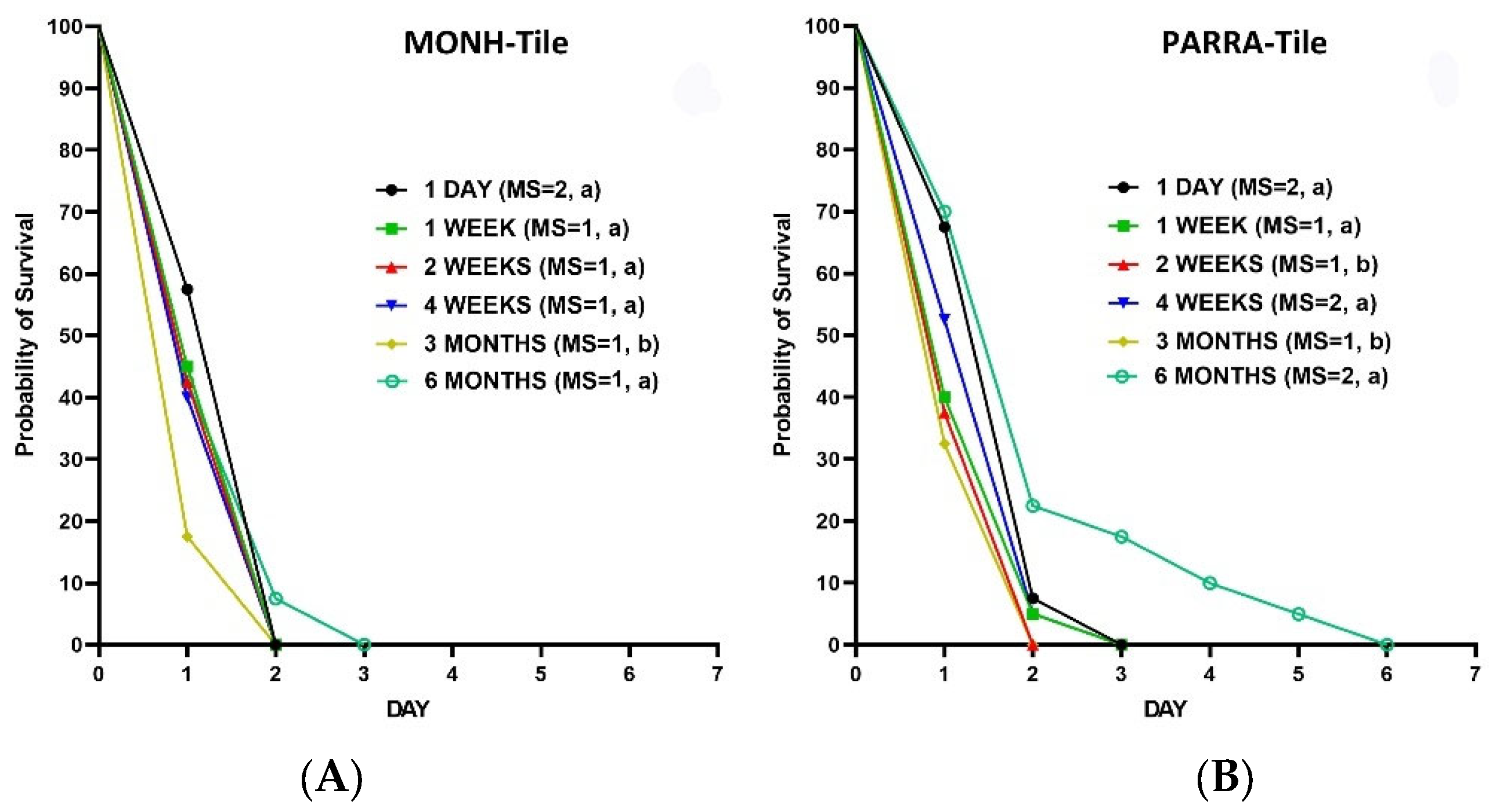
Figure 8.
Kaplan–Meier survival analyses for the efficacy of the aged residual IMZOPS 60 on filter paper against MONH (A) and PARRA (B) strains. MS = median survival time (day). Lowercase letters (a) indicate no statistical differences between the fresh residues and each aged residue (Log-rank test, P value is more than α, [α=0.05/5, Bonferroni correction, 5 is the number of pairwise comparisons (e.g., 1 DAY vs 1 WEEK, 1 DAY vs 2 WEEKS, 1 DAY vs 4 WEEKS, 1 DAY vs 3 MONTHS, and 1 DAY vs 6 MONTHS) of survival curves in either (A) or (B)]).
Figure 8.
Kaplan–Meier survival analyses for the efficacy of the aged residual IMZOPS 60 on filter paper against MONH (A) and PARRA (B) strains. MS = median survival time (day). Lowercase letters (a) indicate no statistical differences between the fresh residues and each aged residue (Log-rank test, P value is more than α, [α=0.05/5, Bonferroni correction, 5 is the number of pairwise comparisons (e.g., 1 DAY vs 1 WEEK, 1 DAY vs 2 WEEKS, 1 DAY vs 4 WEEKS, 1 DAY vs 3 MONTHS, and 1 DAY vs 6 MONTHS) of survival curves in either (A) or (B)]).
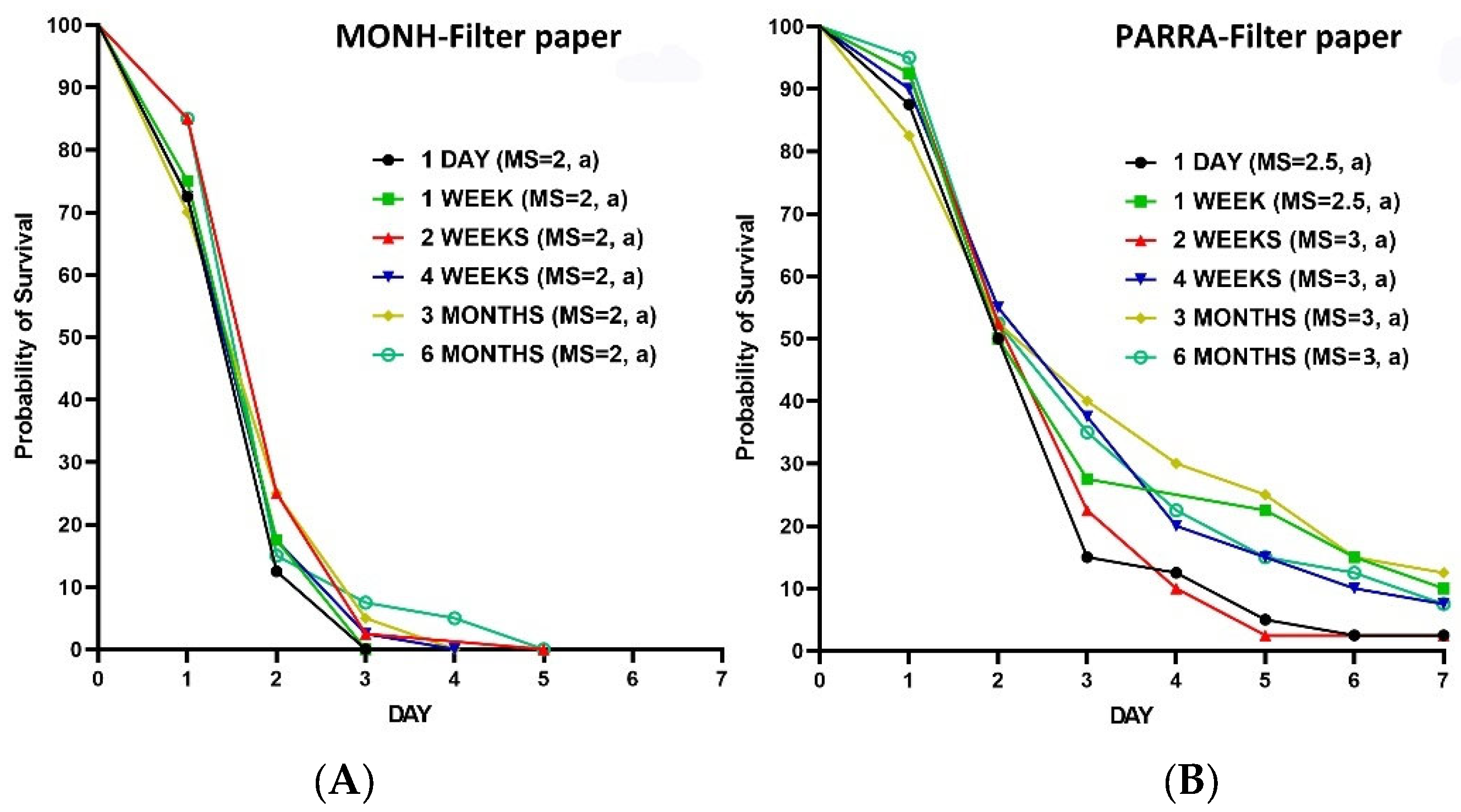
Figure 9.
Daily recorded number (mean±SD %) of the MONH C. lectularius strain on and/or under harborages, wandering, and dead in one-choice harborage assays. Either lowercase letters (a, b) or uppercase letters (A, B) above the error bars indicate statistical differences in each individual day (two-way ANOVA, Turkey’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.
Figure 9.
Daily recorded number (mean±SD %) of the MONH C. lectularius strain on and/or under harborages, wandering, and dead in one-choice harborage assays. Either lowercase letters (a, b) or uppercase letters (A, B) above the error bars indicate statistical differences in each individual day (two-way ANOVA, Turkey’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.
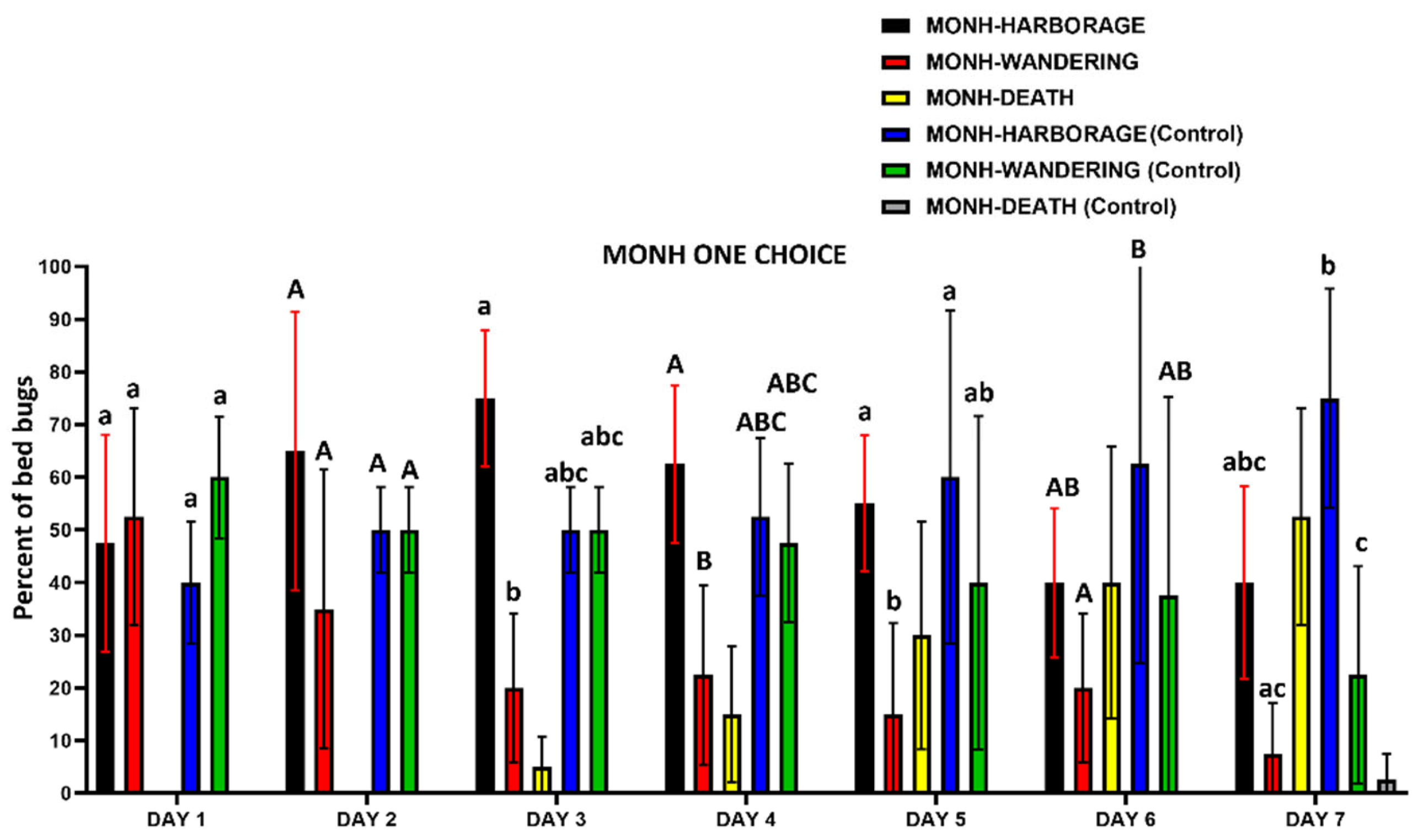
Figure 10.
Daily recorded number (mean±SD %) of the PARRA C. lectularius strain on and/or under harborages, wandering, and death in one-choice harborage assays. Either lowercase letters (a, b) or uppercase letters (A, B) above the error bars indicate statistical difference in each individual day (two-way ANOVA, Turkey’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.
Figure 10.
Daily recorded number (mean±SD %) of the PARRA C. lectularius strain on and/or under harborages, wandering, and death in one-choice harborage assays. Either lowercase letters (a, b) or uppercase letters (A, B) above the error bars indicate statistical difference in each individual day (two-way ANOVA, Turkey’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.

Figure 11.
Daily recorded number (mean±SD %) of MONH strain C. lectularius on and/or under harborages (treated and blank), wandering, and death in two-choice harborage assays. Either lowercase letters (a,b) or uppercase letters (A,B) above the error bars indicate statistic difference in each individual day (two-way ANOVA, Šídák’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.
Figure 11.
Daily recorded number (mean±SD %) of MONH strain C. lectularius on and/or under harborages (treated and blank), wandering, and death in two-choice harborage assays. Either lowercase letters (a,b) or uppercase letters (A,B) above the error bars indicate statistic difference in each individual day (two-way ANOVA, Šídák’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.
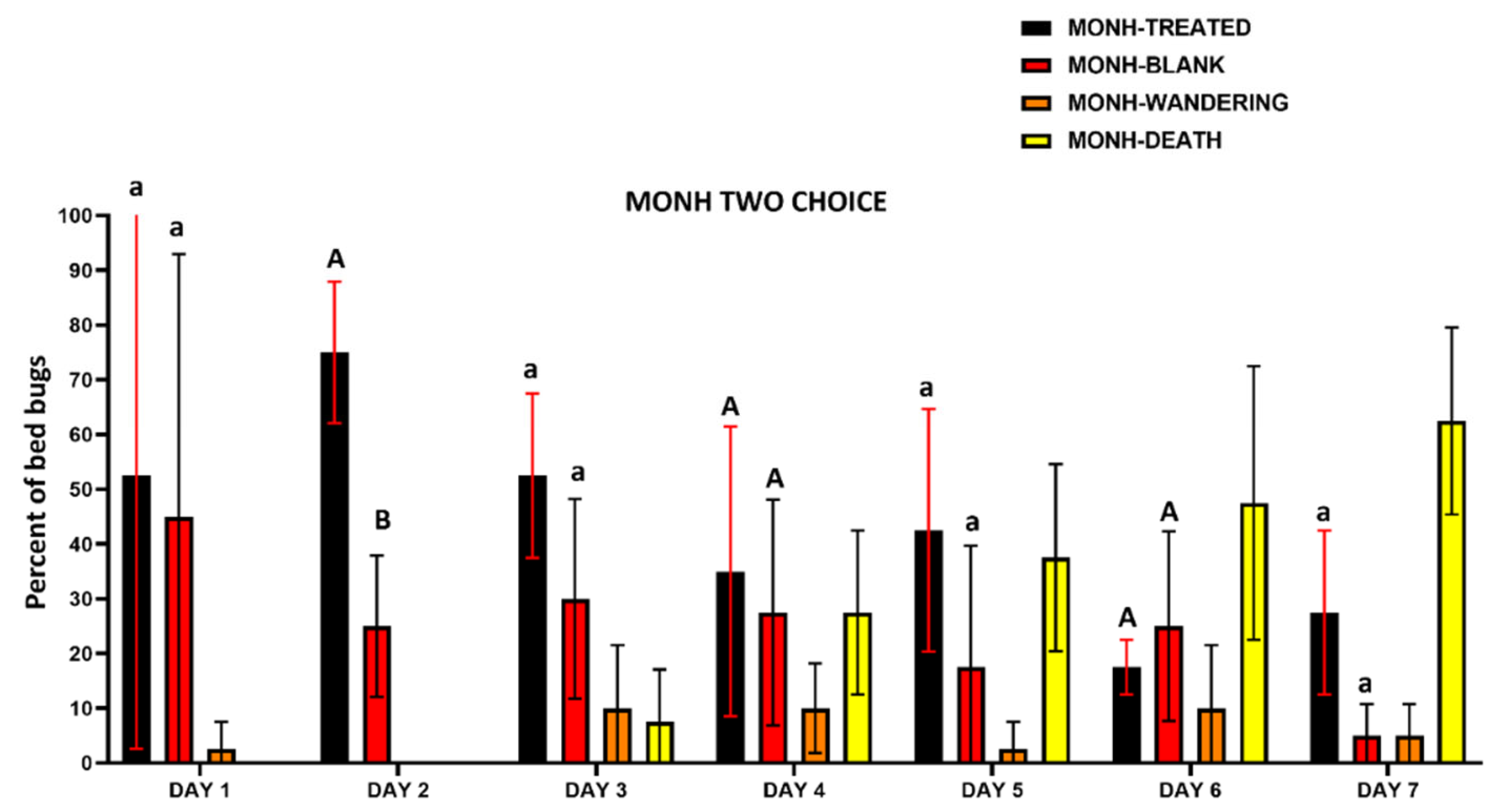
Figure 12.
Daily recorded number (mean±SD %) of PARRA strain C. lectularius on and/or under harborages, wandering, and death in two choice harborage assays. The harborages included treated harborages (TREATED) and untreated harborages (BLANK). Either lowercase letters (a, b) or uppercase letters (A, B) above the error bars indicate statistical difference in each individual day (two-way ANOVA, Šídák’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.
Figure 12.
Daily recorded number (mean±SD %) of PARRA strain C. lectularius on and/or under harborages, wandering, and death in two choice harborage assays. The harborages included treated harborages (TREATED) and untreated harborages (BLANK). Either lowercase letters (a, b) or uppercase letters (A, B) above the error bars indicate statistical difference in each individual day (two-way ANOVA, Šídák’s test, P < 0.05). As the focus of the trial was on the behavioral response of the insects to the chemical, bed bugs that died were not included in the statistical analyses.
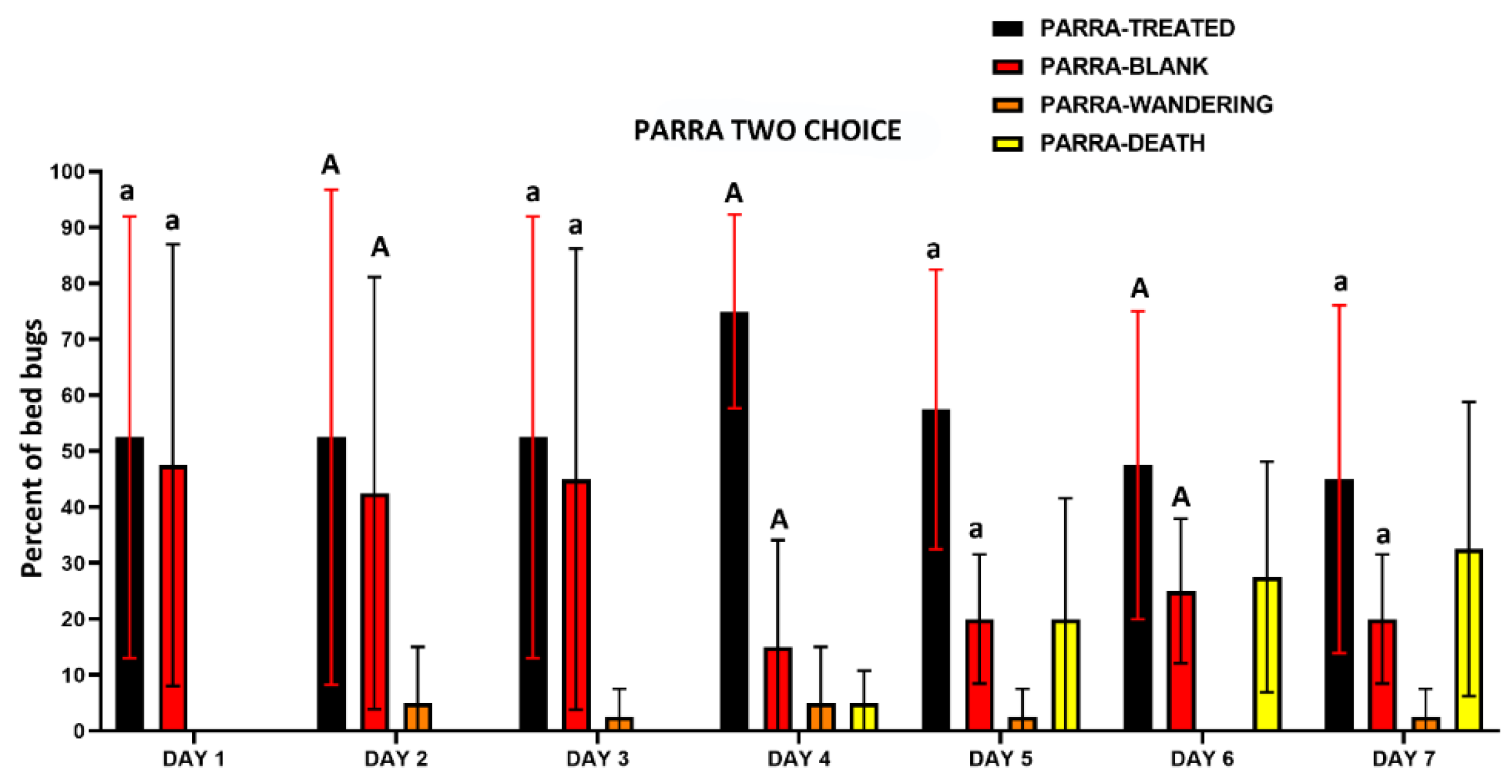

Table 1.
Bed bug strains used in the study.
| Locations | Strain code | Year | Maximum generation | Susceptibility |
kdr Haplotypeb |
|---|---|---|---|---|---|
| Monheim, Germany, 40789 | MONH | 1960s | NA | Susceptible | A |
| Sydney, NSW Australia, 2000 | SYD | 2004 | >200 | Resistant | A/B |
| Darlinghurst, NSW Australia, 2010 | DARL | 2011 | 140 | Resistant | B/C |
| Parramatta, NSW Australia, 2150 | PARRAa | 2012 | 128 | Resistant | B |
| Ripponlea, VIC Australia, 3185 | RIPPO | 2013 | 117 | Resistant | B |
a PARRA strain has been reported to possess cuticular resistance (a thickened cuticle), metabolic resistance, and kdr resistance. b kdr Haplotype according to Dang and colleagues [16].
Table 2.
Log-dose probit-mortality data for a susceptible MONH strain and four pyrethroid resistant strains tested with Broflanilide.
Table 2.
Log-dose probit-mortality data for a susceptible MONH strain and four pyrethroid resistant strains tested with Broflanilide.
| Strains | Replicatesa | Nb | LD50 ng (95%, CI)c | LD95 ng (95%, CI) | χ2 (df) | Slop ± SE | RR50 d |
|---|---|---|---|---|---|---|---|
| MONH | 8 | 640 | 1.00 (0.38 – 1.76) A | 12.49 (5.29 – 166.16) | 25.72 (5) | 1.50 ± 0.14 | 1 |
| SYD | 4 | 360 | 1.22 (0.46 – 2.17) AB | 91.69 (39.95 – 439.89) | 2.96 (4) | 0.88 ± 0.15 | 1.22 |
| DARL | 5 | 450 | 1.03 (0.11 – 2.38) AB | 40.89 (14.79 – 972.74) | 7.42 (4) | 1.03 ± 0.16 | 1.03 |
| PARRA | 8 | 720 | 3.65 (1.83 – 7.20) B | 137.58 (43.43 – 1648.30) | 26.45 (6) | 1.04 ± 0.08 | 3.65 |
| RIPPO | 5 | 450 | 2.77 (1.52 – 4.35) AB | 283.06 (113.72 – 1346.70) | 3.34 (4) | 1.50 ± 0.19 | 2.77 |
a The number of replicates was based on the population of the bed bug strain. b Total number of bed bugs used. Mortality in all control groups was below 5 % and thus no correction was required. c Different uppercase letters (A, B) in the column for LD50 indicate a statistical difference (P < 0.05) as the 95% CIs do not overlapped. d RR50 = LD50 of resistant strain/ LD50 of the susceptible MONH strain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



