
Submitted:
30 April 2024
Posted:
01 May 2024
You are already at the latest version
Abstract
The Chikungunya virus (CHIKV) presents global health challenges, with Brazil witnessing out-breaks since its introduction in 2014. In 2023, Minas Gerais (MG) experienced a CHIKV outbreak, prompting entomological investigations to identify vectors, viral lineages, and assess insecticide resistance. This study utilized social media platforms and citizen science concept to conduct a comprehensive entomological survey. Following Instagram posts, residents with CHIKV sus-pected cases were able to schedule mosquito aspirations. During the aspirations, 421 mosquitoes (165 Aedes aegypti and 256 Culex quinquefasciatus) were captured from 40 households in Salinas city (MG) and tested for Dengue, Zika and Chikungunya viruses through RT-qPCR. Twelve out 57 pools (10 of Ae. aegypti and 2 of Cx. quinquefasciatus) tested positive for CHIKV RNA. Viral RNA was also detected in the heads of 9 Ae. aegypti individuals (but not in Cx. quinquefasciatus). Genome sequencing revealed that CHIKV belonged to the East-Central-South-Africa (ECSA) genotype. Additionally, genetic analyses revealed high frequencies of kdr alleles, suggesting resistant to pyrethroids insecticides in this Ae. aegypti population. These findings emphasize the urgent need for innovative vector study and control strategies to combat arbovirus transmission, and inter-disciplinary approaches in public health interventions.
Keywords:
Mosquito
; Arbovirus
; Pyrethroid
; Social media
; Culex
1. Introduction
The Chikungunya virus (CHIKV) a member of the Togaviridae Family and Alphavirus Genus, is an arbovirus responsible for chikungunya fever, a debilitating human disease marked by high fever and severe joint pain, often leading to long-lasting sequelae [1,2]. First identified during an epidemic in Tanzania, Africa, in 1952-1953, CHIKV has since caused outbreaks across Africa, Asia, Europe, and Oceania [3]. Its introduction to the Americas occurred in 2013, reaching Brazil in 2014 and causing explosive outbreaks [4].
In Brazil, the main vector for CHIKV is the Aedes aegypti mosquito [5,6]. However, a single genetic mutation (CHIKV E1-226V) has enabled CHIKV adaptation to Aedes albopictus, expanding the potential for epidemics in areas with low Ae. aegypti infestation [7]. Brazilian populations of Ae. albopictus are highly competent for transmitting CHIKV [8,9] and have spread widely since their introduction in 1986 [10,11,12]. Additionally, urban areas in Brazil are infested with Culex quinquefasciatus, which has been found naturally infected by CHIKV [6,13,14], although its vector competence is controversial [15,16].
The coexistence of these mosquito species in Brazilian cities raises concerns for public health authorities. Despite recommendations for integrated vector control measures, including chemical control [17], the strategy predominantly relies on insecticides. However, the overuse of these chemicals has led to the selection of resistant Ae. aegypti populations [18,19]. Although national governmental campaigns detected resistance to pyrethroids in all Brazilian regions and stopped employing this insecticide class since 1989 [20], these chemicals are still highly used by household against indoors undesired insects, including mosquitoes, since they are less irritant to people and produce the rapid effect knockdown. This is likely the main reason of selection and spreading of mutations in the voltage-gated sodium channel gene (NaV), the pyrethroids target-site molecule, known as kdr (knockdown resistant mutations) [21]. There are at least two kdr alleles widespread in Ae. aegypti Brazilian populations: NaVR1 and NaVR2, respectively with one (F1534C) and three (V410L, V1016I and F1534C) [22,23] with the NaVR2 conferring higher levels of resistance [24].
In early 2023, Minas Gerais (MG) experienced its largest CHIK outbreak on record, with 69,331 confirmed cases by October, surpassing totals for 2021 (5,557) and 2022 (13,148) [25]. The Northern region of Minas Gerais, characterized by small municipalities with extensive rural areas, was severely affected, reporting 31,410 cases [26]. This region, primarily composed of small municipalities (up to 60,000 inhabitants), [27] faces economic challenges. Despite the magnitude of the outbreak, no entomological studies had been conducted in the region to identify the vectors responsible for transmission. This gap was partly due to logistical challenges and difficulties in locating infected mosquitoes [28].
Hence, this study aimed to perform an entomological investigation using social media and citizen science [29] - research conducted with participation from the general public - to identify suspected CHIKV cases. This approach optimizes efforts to determine the vectors responsible for transmission, infection rates, and the presence of insecticide resistance mutations.
2. Materials and Methods
2.1. Study Area
The study was conducted in the city of Salinas (16°09’45,8’’S; 042°17’54,2’’W), located in the Northern region of Minas Gerais (Figure 1). Salinas comprises a population of 40,178 inhabitants and a low municipal human development index (MHDI = 0.679)62 and is positioned within an ecotone between the Cerrado and Atlantic Forest biomes [30]. The area features a semi-arid climate (Aw climate type according to Köppen, 1936) [31], characterized by two well-defined seasons: an extended dry season from March to October, and a brief rainy season from November to February, aligning with the time of sample collection.
2.2. Mosquito Collection and Rearing Following Instagram Posts
At the onset of 2023, reports of individuals experiencing high fever and arthralgia began circulating in Salinas. By February 6th, eight confirmed cases of CHIK had already been reported in the city [32]. On February 8th, our laboratory's Instagram profile (@lacoi_ifnmg) posted an announcement about our entomological investigation, asking residents if they would like schedule a mosquito aspiration visit (https://www.instagram.com/p/CoarMhNvn_d/?img_index=1). Visits were arranged at the request of residents and based on their availability. Each household was visited by one municipal endemic control agent (city hall employees who work on mosquito control) and an entomologist equipped with battery-powered Nasci Aspirators, [33] oral aspirators, and field entomological cages. Sampling efforts encompassed thorough searches of all rooms within residences, with particular attention paid to hidden niches, such as under beds, tables, behind sofas, and cabinets. After, the number of captured insects was communicated to the residents.
Captured mosquitoes were separated by genera and sex and transferred via oral aspirators to field cages, which were subsequently sealed, labeled, and transported to the Insect Behavior Laboratory at the Federal Institute of Northern of Minas Gerais. Within these cages, a 10% sucrose solution soaked in cotton was provided, and the field cages were housed inside larger Bugdorm-type cages (avoiding escape risks) at room temperature (27 ± 4ºC). Mosquitoes were maintained alive for three days to allow blood digestion in potentially engorged females, a period deemed sufficient for the virus to disseminate through mosquito body and reach the salivary glands [34]. After three days, mosquitos were killed by frozen at -20ºC, transferred to cryovials and stored in liquid nitrogen (-196ºC) until further processing.
Throughout the study, additional Instagram posts were generated to update the public on the preliminary findings (e.g., https://www.instagram.com/p/CpGXmXJvQLg/ ; https://www.instagram.com/p/CpdonXJv-7u/ ).
2.3. Taxonomic Identification and CHIKV Molecular Diagnosis in Captured Mosquitoes
Mosquitoes were transferred from liquid nitrogen and subjected to identification and confirmation taxonomic on a cold table (-20ºC) under a stereoscopic microscope, following dichotomous keys [35,36]. Each mosquito was assigned a unique code and dissected (using individual scalpels to avoid cross contamination) into three parts: legs, head, and body (Figure 3). Legs were preserved in 150 µL TE Buffer 0,1x, while heads were individually stored in tubes containing 150 µL of enriched L-15 medium (20% fetal bovine serum, 0.5% non-essential amino acids, 1% penicillin, 0.1% gentamicin and 0.1% fungizone) and frozen immediately (Figure 3). Non-engorged mosquito bodies were pooled (up to 10 individuals) by species and sex (Figure 3). These pools were then crushed using a beadbeater (L-Beader 24, Loccus) in tubes containing beads and 500 µL of enriched L-15 medium (as described above) for 30 seconds at 7500 rpm [37]. After, these tubes were immediately centrifuged (12,000 rpm, 8 minutes, 4ºC) and 140 µL of the supernatant were used for RNA extraction using the Qiamp viral RNA minikit (Qiagen) following the manufacturer's instructions.
Real time RT-qPCR assays were conducted to detect the presence of Dengue (DENV), Zika (ZIKV) and CHIKV RNA using the ZDC Multiplex PCR kit (Bioclin Qibasa) according to the manufacturer's instructions. Heads corresponding to insects from positive pools were individually tested to identify the number of infected individuals per pool and assess viral dissemination following the same RNA extraction protocol. Since only CHIKV-positive body pools were obtained, corresponding head samples RNAs were subjected to RT-qPCR analysis as previously described [38]. Briefly, GoTaq® 1-Step RT-qPCR System (Promega) was used along specific set of primers and probes targeting the E1 gene, with the following sequences: CHIK F - 5’-AAGCTYCGCGTCCTTTACCAAG-3’, CHIK R - 5’- CCAAATTGTCCYGGTCTTCCT-3’, CHIK P - 5’-FAM CCAATGTCYTCMGCCTGGACACCTTT-BHQ1-3’ [38]. The RT-qPCR protocol involved reverse transcription at 50◦C for 20 min, initial denaturation at 95◦C for 2 min, followed by 45 cycles of 95◦C for 5 s and 60◦C for 1 min.
2.4. CHIKV Genome Sequencing and Phylogenomic Analyses
A representative pool (X-595) was selected due to its lowest cycle threshold (CT) value to proceed with whole genome sequencing. The extracted RNA underwent cDNA synthesis and PCR amplification using a sequencing protocol based on multiplex PCR-tiling amplicon approach [39]. Subsequently, the resulting amplicons were purified using 1x AMPure XP Beads (Beckman Coulter) and quantified using a Qubit 3.0 fluorimeter (Thermofisher Scientific) with the Qubit™ dsDNA HS Assay Kit (ThermoFisher Scientific). Genomic libraries were then prepared using the Illumina DNA Prep (Illumina) and sequenced on the MiSeq platform (Illumina) with v3 (600 cycles) cartridges, following the manufacturer's instructions.
The sequencing file were processed following an assembly pipeline previously described [40] and publicly available on GitHub (https://github.com/ filiperomero2/ViralUnity). Sample genotyping was performed using the Genome Detective Virus Tool Version 2.72 [41].
The new genome was then added to the existing dataset of complete genomes (>11,000 bp) publicly available in Genbank for subsequent analysis, resulting in a total of 877 genomes. Sequence alignment was conducted using MAFFT version 7.490 [42] and visually inspected using AliView version 1.28 [43]. Maximum likelihood (ML) trees were generated using IQ-TREE 2.2.5 [44]. Statistical robustness of the tree topology was assessed using 1000 bootstrap replicates.
2.5. Analysis of kdr as a Molecular Marker for Pyrethroid Resistance
We utilized the legs of each captured Ae. aegypti to individually genotype kdr for the three single nucleotide polymorphisms (SNPs) V410L, V1016I and F1534C. The legs were crushed in a 10% TE solution using two glass beads in a Tissue Lyser II (QIAGEN) equipment for 2 minutes at stirring speed 30. Subsequently, the samples were homogenized with the addition of 200 µL of TNES and 2 µL of Proteinase K (20 mg/mL) and left overnight at 56°C in a water bath. Following this, 100 µL of 5M NaCl were added, and the mixture was centrifuged at 15,000g for 6 minutes. The supernatant was transferred to a new microtube for washing steps with pure isopropanol, followed by washing with 70% ethanol. Once the pellet was dry, the DNA was resuspended in 50 µL of ultrapure water and stored at -20°C. We performed independent qPCR reactions for each kdr single nucleotide polymorphisms (SNPs) V410L, V1016I and F1534C, essentially as described elsewhere [22]. The reactions were carried out in the Thermofisher Real-Time Thermocycler, QuantStudio 6 Flex. The obtention of kdr genotypic and allelic frequencies considered the variation in the three SNPs of each mosquito (see Souza et al., 2023)[23].
2.6. Ethical Statements
Mosquito collections and methods were approved by local authorities (SISBIO-MMA license nº 75826-3; SISGEN nº AF40BCA). This study did not involve endangered or protected species.
3. Results
Between February 08 and March 30, 2023, we conducted visits to 40 houses across 13 neighborhoods in the city of Salinas, Minas Gerais state (Figure 1). In total, 421 mosquitoes were captured, comprising 256 Cx. quinquefasciatus (mean 6.4 ± 6.2 per house) and 165 Ae. aegypti (mean 4.1 ± 4.6 per house) (Table 1). Notably, no Ae. albopictus specimens were captured during the sampling period (Table 1). Mosquito bodies were grouped into 57 pools, all of which tested for the presence of DENV, ZIKV, and CHIKV RNA. All pools were negative for DENV and ZIKV. The total of 12 pools (10 Ae. aegypti: seven female and three male pools; and two Cx. quinquefasciatus: one female and one male pools) tested positive for CHIKV (Table 2).
The CT values of the positive pools ranged from 20.1 to 40.0 (Table 2). These 12 CHIKV positive pools comprised 71 individuals (53 Ae. aegypti and 18 Cx. quinquefasciatus), whose heads were individually examined for the presence of CHIKV. Among these, viral RNA was detected in nine heads, all in Ae. aegypti females, from six distinct pools, and with CT below 24.2 (Table 2). Conversely, the heads from the CHIKV positive pools whose CT values exceeding 28.0, including the Cx. quinquefasciatus pools (one male and one female pools), as well as four Ae. aegypti pools (three males and one female pools), tested negative for CHIKV (Table 2).
We obtained 247,073 mapped reads for the sample x-595 with coverage of 94.8% of the CHIKV genome, with a minimum depth of 20X and an average depth of 2,383.88. The Maximum Likelihood (ML) tree grouped these sequences with the CHIKV East South African (ECSA) genotype, clustering within the same clade as samples detected in São Paulo in 2021 (Figure 3).
In total, 164 Ae. aegypti were genotyped to verify kdr mutations. The most frequent genotype was the homozygous for the kdr R2 allele, which contains the three kdr SNPs (LIC) with 40.9%, followed by the heterozygous kdr R1/R2 (VVC/LIC) with 32.3%, and the homozygous for the kdr R1 (VVC/VVC) with 11.0% (Table 3). This means that at least 84.2% presented a genotype compatible with pyrethroid resistance, and almost half of which would likely display higher levels of resistance (the homozygous kdr R2/R2). Out of the 164 mosquitoes genotyped for the three SNPs, only three (1.8%) were homozygous for the wild-type NaVS allele (VVF). Taken together this reflects an ongoing selection pressure for pyrethroid resistance in Ae. aegypti from Salinas. We also observed some uncommonly observed kdr genotypes in 2% of the samples, to which allelic composition and the relationship with resistance phenotype deserve future investigation (Table 3).
Among the nine individuals with detectable CHIKV RNA in their heads, five (55.5%) were R1R2, three (33.3%) R2R2, and one SR2. Although we did not have enough samples to compare the genotypic frequencies between positive and negative CHIKV samples, it was evidenced that there was no constraint for this virus to infect and disseminate in kdr mosquitoes.
4. Discussion
The CHIKV is an arbovirus capable of triggering explosive outbreaks, leading to significant social and economic impacts due to the prolonged clinical manifestations of the disease. Despite the absence of a vaccine, vector control remains the primary preventive measure against CHIKV. Therefore, identifying the vectors responsible for virus transmission and assessing insecticide resistance levels are crucial for understanding CHIK epidemiology and implementing effective control measures. In this study, Instagram - social media platform - proved to be a valuable tool contributing to the investigation of a CHIK outbreak, pointing Ae. aegypti as the main vector in Minas Gerais, Brazil.
The low diversity of intra domiciliary mosquitoes (Cx. quinquefasciatus and Ae. aegypti only) is consistent with the essentially urban and anthropophilic habitat of these vectors [5,35]. Ae. aegypti is the main vector of urban arboviruses, including Dengue, Zika, and Chikungunya, in Brazil. Notably, Ae. aegypti was first documented with natural infection by CHIKV in 2017 in Brazil and the Americas, attributed to the ECSA genotype [5], the same found in the present study. This finding aligns with the widespread prevalence of the ECSA genotype since its introduction in 2014 [45], frequently detected in humans CHIK cases in Brazil [46,47], included those in the state of Minas Gerais [48]. Despite its importance, there are few reports on genomic CHIKV surveillance in mosquitoes from Brazil [6,12,13,14,49,50,51,52]. Although a large proportion of CHIKV cases in this country have been reported in the Southeast region, the most Brazilian urbanized and densely populated region, this is the first detection of CHIKV in vectors from this region.
The elevated MIR observed in Aedes aegypti (60.6), coupled with the low CTs obtained, are indicative of high viral RNA loads, underscoring the significant role of this species in the maintenance and transmission of CHIKV within Brazilian urban environments [5,6,49]. Notably, viral RNA was detected in the heads of nine female mosquitoes, indicating viral dissemination and highlighting their potential as vectors. Furthermore, CHIKV RNA was also found in four Ae. aegypti male pools, despite the higher CTs, suggesting potential transovarian or sexual transmission mechanisms, consistent with findings from previous studies [6,50]. Interestingly, CHIKV RNA was not detected in male heads from positive body pools, nor in female heads from one pool, suggesting limited viral dissemination in these specimens. Previous assessments of CHIKV vector competency have revealed that, despite their high vector competence and viral dissemination in secondary tissues such as wings and legs, certain individuals’ tissues or saliva may remain uninfected, potentially due to barriers in the midgut or salivary gland, which could impair viral spread [8,9,53].
Despite the greater abundance of Cx. quinquefasciatus within households, as previously demonstrated [5,6,49,54], its infection rate (MIR = 7.8) was substantially lower compared to Ae. aegypti. Additionally, the CTs were notably high, approaching the limit of the assay detection. While Ribeiro Cruz et al. [6] successfully isolated CHIKV from two pools of naturally infected female Cx. quinquefasciatus, they could not determine vector competence as the RNA was extracted from whole-bodies macerates, thus preventing the assessment of possible viral migration to the salivary glands. Consequently, to date, no compelling evidence implicates this species in the transmission of CHIKV in Brazil, as we cannot eliminate the hypothesis that those positive female pools may have been infected through blood meal residues from viremic hosts. Similarly to Ae. aegypti, the detection of a male Cx. quinquefasciatus pool positive for CHIKV RNA suggests the potential for transovarian or sexual transmission, as previously suggested [13,14]. In Kenya, Lutomiah et al. [15] proposed the involvement of Cx. quinquefasciatus in CHIKV transmission based on the discovery of naturally infected mosquitoes and evidence of high vector competency in laboratory assays.
The well-established vector competence of Brazilian Ae. albopictus populations for various CHIKV lineages [8,9], along with documented cases of natural infection [14,55], and its widespread distribution across the country [10,56], raise concerns regarding the potential for this species to serve as a vector in Brazil, as observed in other countries [7,57,58]. In Salinas, this species has previously been identified in ovitraps placed in the peridomicile, particularly in residences near the riparian forests of the Salinas River [59]. However, our study did not find Ae. albopictus specimens indoors among the 40 sampled residences, suggesting its limited adaptation to indoor environments in this locality. In Brazil, these mosquitoes typically inhabit forest edges in transition areas (ecotones) between forests and urban landscapes, which makes this species a potential bridge vector for arboviruses between these environments [60].
Insecticide resistance is a threat to the control of Ae. aegypti globally, making the monitoring of susceptibility a primary necessity for chemical control sustainability [61]. The surveillance of kdr mutations can be used as an indirect indication of pyrethroid resistance, as they partially respond to this phenotype [18]. Based on the kdr genotyping, here we evidenced that the Ae. aegypti population from Salinas is probably resistant to pyrethroids, with a high incidence of resistant genotypes. A previous study monitoring kdr mutations across Brazil revealed that the Ae. aegypti population from Montes Claros (located in the northern region of Minas Gerais, approximately 170 km away from our study area) already exhibited a high proportion (82,3%) of mosquitoes with resistant genotypes (R1R1, R1R2 and R2R2),22 quite similar to the 84.2% we currently found in Ae. aegypti from Salinas. Importantly, our research demonstrated that the nine CHIKV-infected individuals presented a kdr genotype, indicating that there are no constraints for the dissemination of this virus in kdr mosquito organisms. Following the onset of the CHIK outbreak, health authorities implemented an Ae. aegypti population control program, focusing on eliminating breeding sites and utilizing pyrethroid-based insecticides (such as Icon 2.5EW) through ultra-low volume spraying. Additionally, during mosquito sampling, we observed the widespread use of pyrethroid-based commercial insecticides, in numerous of the visited households. The sustained use of pyrethroids likely exerts selective pressure on resistant alleles/genotypes, thereby reducing the effect of chemical control efforts and potentially leading to the enhancement of outbreaks. Collectively, our findings underscore the imperative to invest in novel vector control strategies (such as Wolbachia-infected and transgenic mosquitoes), in increased efforts to stimulate entomological surveillance (including digital tools and citizen science initiatives), and in vaccine research to mitigate the risk of arboviral disease outbreaks.
Broadly defined, citizen science describes scientific investigations in which volunteers participate in scientific studies designed to answer real-world questions [29]. Several studies have been focused on public engagement and contribution to the scientific process [62,63] and the use of social media has been promising in this process [29,64,65,66]. In this study, social media platform improved collection efforts increasing the probability of arbovirus detection. Recently, the integration of citizen science principles, alongside the utilization of smartphone applications and social media platforms, have emerged as pivotal tools in mosquito surveillance, vector-borne disease monitoring, and scientific knowledge dissemination [67,68,69,70,71,72,73,74]. Given the acknowledged shortcomings of conventional arbovirus surveillance and control methods, the adoption of these innovative tools and technologies is increasingly imperative.
While our study provided valuable new findings, it's crucial to acknowledge its limitations. Firstly, the focus on entomological investigations within households may not fully capture the diversity and dynamics of vector populations in peri-domestic and sylvatic environments. Additionally, while social media platforms facilitated to aspirate houses with suspected arboviral disease, relying solely on such platforms may introduce selection biases, as individuals with access to and familiarity with these platforms may differ from those who do not participate. Lastly, the geographical scope of the study was confined to one municipality in Minas Gerais, Brazil, limiting broader generalizations about CHIKV vector ecology and insecticide resistance patterns across different regions. Despite these limitations, our findings underscore the importance of innovative approaches, such as social media-driven citizen science, in bolstering entomological surveillance efforts and advancing our comprehension of arbovirus epidemiology and control strategies.
5. Conclusions
In conclusion, our study underscores the crucial role of entomological surveillance in comprehending the epidemiology and control of the CHIKV infections. Through the utilization of employing innovative methodologies like social media and citizen science, we identified Ae. aegypti as the primary vector of CHIKV (ECSA genotype) during the 2023 outbreak in Minas Gerais, Brazil, while also revealing high frequency of kdr mutations, compatible with pyrethroid resistance in the vector population. Despite inherent limitations, such as the focus on domestic areas and potential selection bias, our findings emphasize the urgent necessity for innovative vector control strategies and the development of novel vaccines to effectively mitigate arboviral disease outbreaks. Additionally, we underscore the growing importance of interdisciplinary approaches, including collaboration among researchers, public health professionals, and community members, to bolster entomological surveillance and address the ongoing challenges posed by mosquito-borne diseases.
Author Contributions
Conceptualization, A.J.M. and F.V.S.A; methodology, P.A.A.S., C.H.O., L.P.B., T.J.T., I.A.C., I.A.F., A.O.F., G.B.P.,C.B.O., N.R.G., F.V.S.A.; software, R.M.S, F.I, N.R.G.; validation, A.J.M., B.M.O., G.B.P.; formal analysis, B.M.O., F.I, N.R.G., ; investigation, P.A.A.S., C.H.O., L.P.B., T.J.T., I.A.C., I.A.F., A.O.F., G.B.P.,C.B.O., N.R.G., W.M.M., D.C.C.C.; resources, B.M.R., F.S.C., P.M.R., A.J.M., F.I., F.V.S.A.; data curation, P.A.A.S., C.H.O., L.P.B., T.J.T., I.A.C., I.A.F., A.O.F., G.B.P., N.R.G. writing—original draft preparation, P.A.A.S.; C.H.O., F.S.C., F.V.S.A.; writing—review and editing, D.C.C.C., W.M.M., C.B.O., B.M.R., P.M.R., F.I. A.J.M.; visualization, R.M.S., N.R.G., F.I.; supervision, B.M.O., D.C.C.C., W.M.M., F.V.S.A.; project administration, P.M.R., F.S.C.,; funding acquisition, B.M.O., P.M.R. All authors have read and agreed to the published version of the manuscript.
Funding
P.M.R (CNPq Grant nº 443215/2019–7). FVSA (FAPEMIG grant nº APQ-01403-21). B.M.R., F.S.C. and P.M.R. are CNPq research fellows.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data were showed in the manuscript.
Acknowledgments
The authors extend their gratitude to Sandy Micale Aquino-Teixeira, Maria Eduarda Gonçalves-dos-Santos, and Leonardo Bernardino T. Matos for their support during the field collections and laboratory assays and to Aline Tátila Ferreira for images design. We also thanks FAPEMIG and CNPq.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Robinson, M.C. An Epidemic of Virus Disease in Southern Province, Tanganyika Territory, in 1952–1953. Trans R Soc Trop Med Hyg 1955, 49, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Pialoux, G.; Gaüzère, B.-A.; Jauréguiberry, S.; Strobel, M. Chikungunya, an Epidemic Arbovirosis. Lancet Infect Dis 2007, 7, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.E. Reemergence of Chikungunya Virus. J Virol 2014, 88, 11644–11647. [Google Scholar] [CrossRef] [PubMed]
- Leparc-Goffart, I.; Nougairede, A.; Cassadou, S.; Prat, C.; de Lamballerie, X. Chikungunya in the Americas. Lancet 2014, 383, 514. [Google Scholar] [CrossRef] [PubMed]
- Costa-da-Silva, A.L.; Ioshino, R.S.; Petersen, V.; Lima, A.F.; Cunha, M. dos P.; Wiley, M.R.; Ladner, J.T.; Prieto, K.; Palacios, G.; Costa, D.D.; et al. First Report of Naturally Infected Aedes Aegypti with Chikungunya Virus Genotype ECSA in the Americas. PLoS Negl Trop Dis 2017, 11, e0005630. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro Cruz, A.C.; Pinto Nunes Neto, J.; Patroca da Silva, S.; Vieira Pinto da Silva, E.; Juscely Galvão Pereira, G.; Maia Santos, M.; Antônio de Oliveira Monteiro, H.; Barreto dos Santos, F.; José de Paula Souza e Guimarães, R.; Fortes Aragão, C.; et al. Chikungunya Virus Detection in Aedes Aegypti and Culex Quinquefasciatus during an Outbreak in the Amazon Region. Viruses 2020, 12, 853. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.-S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya Isolates from the Outbreak of La Reunion (Indian Ocean) Exhibit Different Patterns of Infection in the Mosquito, Aedes Albopictus. PLoS One 2007, 2, e1168. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rua, A.; Zouache, K.; Girod, R.; Failloux, A.-B.; Lourenco-de-Oliveira, R. High Level of Vector Competence of Aedes Aegypti and Aedes Albopictus from Ten American Countries as a Crucial Factor in the Spread of Chikungunya Virus. J Virol 2014, 88, 6294–6306. [Google Scholar] [CrossRef] [PubMed]
- Honório, N.A.; Wiggins, K.; Câmara, D.C.P.; Eastmond, B.; Alto, B.W. Chikungunya Virus Vector Competency of Brazilian and Florida Mosquito Vectors. PLoS Negl Trop Dis 2018, 12, e0006521. [Google Scholar] [CrossRef]
- Carvalho, R.G.; Lourenço-de-Oliveira, R.; Braga, I.A. Updating the Geographical Distribution and Frequency of Aedes Albopictus in Brazil with Remarks Regarding Its Range in the Americas. Mem Inst Oswaldo Cruz 2014, 109, 787–796. [Google Scholar] [CrossRef]
- Ferreira-de-Lima, V.H.; Câmara, D.C.P.; Honório, N.A.; Lima-Camara, T.N. The Asian Tiger Mosquito in Brazil: Observations on Biology and Ecological Interactions since Its First Detection in 1986. Acta Trop 2020, 205, 105386. [Google Scholar] [CrossRef] [PubMed]
- Rocha, R. da C.; Cardoso, A. da S.; Souza, J.L. de; Pereira, E. da S.; Amorim, M.F. de; Souza, M.S.M. de; Medeiros, C. de L.; Monteiro, M.F.M.; Meneguetti, D.U. de O.; Paula, M.B. de; et al. First Official Record of Aedes (Stegomyia) Albopictus (Diptera: Culicidae) in the Acre State, Northern Brazil. Rev Inst Med Trop Sao Paulo 2023, 65. [Google Scholar] [CrossRef]
- da Silva Ferreira, R.; de Toni Aquino da Cruz, L.C.; de Souza, V.J.; da Silva Neves, N.A.; de Souza, V.C.; Filho, L.C.F.; da Silva Lemos, P.; de Lima, C.P.S.; Naveca, F.G.; Atanaka, M.; et al. Insect-Specific Viruses and Arboviruses in Adult Male Culicids from Midwestern Brazil. Infection, Genetics and Evolution 2020, 85, 104561. [Google Scholar] [CrossRef] [PubMed]
- da Silva Neves, N.A.; da Silva Ferreira, R.; Morais, D.O.; Pavon, J.A.R.; de Pinho, J.B.; Slhessarenko, R.D. Chikungunya, Zika, Mayaro, and Equine Encephalitis Virus Detection in Adult Culicinae from South Central Mato Grosso, Brazil, during the Rainy Season of 2018. Brazilian Journal of Microbiology 2022, 53, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Lutomiah, J.; Mulwa, F.; Mutisya, J.; Koskei, E.; Langat, S.; Nyunja, A.; Koka, H.; Konongoi, S.; Chepkorir, E.; Ofula, V.; et al. Probable Contribution of Culex Quinquefasciatus Mosquitoes to the Circulation of Chikungunya Virus during an Outbreak in Mombasa County, Kenya, 2017–2018. Parasit Vectors 2021, 14, 138. [Google Scholar] [CrossRef] [PubMed]
- van den Hurk, A.F.; Hall-Mendelin, S.; Pyke, A.T.; Smith, G.A.; Mackenzie, J.S. Vector Competence of Australian Mosquitoes for Chikungunya Virus. Vector-Borne and Zoonotic Diseases 2010, 10, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Brasil; Ministério da Saúde; Brasil. Ministério da Saúde. Secretaria de VigDepartamento de Articulação Estratégica de Vigilância em Saúde Guia de Vigilância Em Saúde Available online:. Available online: https://bvsms.saude.gov.br/bvs/publicacoes/guia_vigilancia_saude_5ed_rev_atual.pdf (accessed on 21 April 2024).
- Valle, D.; Bellinato, D.F.; Viana-Medeiros, P.F.; Lima, J.B.P.; Martins Junior, A. de J. Resistance to Temephos and Deltamethrin in Aedes Aegypti from Brazil between 1985 and 2017. Mem Inst Oswaldo Cruz 2019, 114. [Google Scholar] [CrossRef] [PubMed]
- Campos, K.B.; Martins, A.J.; Rodovalho, C. de M.; Bellinato, D.F.; Dias, L. dos S.; Macoris, M. de L. da G.; Andrighetti, M.T.M.; Lima, J.B.P.; Obara, M.T. Assessment of the Susceptibility Status of Aedes Aegypti (Diptera: Culicidae) Populations to Pyriproxyfen and Malathion in a Nation-Wide Monitoring of Insecticide Resistance Performed in Brazil from 2017 to 2018. Parasit Vectors 2020, 13, 531. [Google Scholar] [CrossRef] [PubMed]
- Macoris, M. de L.; Martins, A.J.; Andrighetti, M.T.M.; Lima, J.B.P.; Valle, D. Pyrethroid Resistance Persists after Ten Years without Usage against Aedes Aegypti in Governmental Campaigns: Lessons from São Paulo State, Brazil. PLoS Negl Trop Dis 2018, 12, e0006390. [Google Scholar] [CrossRef]
- Moyes, C.L.; Wiebe, A.; Gleave, K.; Trett, A.; Hancock, P.A.; Padonou, G.G.; Chouaïbou, M.S.; Sovi, A.; Abuelmaali, S.A.; Ochomo, E.; et al. Analysis-Ready Datasets for Insecticide Resistance Phenotype and Genotype Frequency in African Malaria Vectors. Sci Data 2019, 6, 121. [Google Scholar] [CrossRef]
- Melo Costa, M.; Campos, K.B.; Brito, L.P.; Roux, E.; Melo Rodovalho, C.; Bellinato, D.F.; Lima, J.B.P.; Martins, A.J. Kdr Genotyping in Aedes Aegypti from Brazil on a Nation-Wide Scale from 2017 to 2018. Sci Rep 2020, 10, 13267. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.S.; Lima, L.F.; Galardo, A.K.R.; Corbel, V.; Lima, J.B.P.; Martins, A.J. Genetic Structure and Kdr Mutations in Aedes Aegypti Populations along a Road Crossing the Amazon Forest in Amapá State, Brazil. Sci Rep 2023, 13, 17167. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.P.; Carrara, L.; Freitas, R.M. de; Lima, J.B.P.; Martins, A.J. Levels of Resistance to Pyrethroid among Distinct Kdr Alleles in Aedes Aegypti Laboratory Lines and Frequency of Kdr Alleles in 27 Natural Populations from Rio de Janeiro, Brazil. Biomed Res Int 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- SES-MG; Secretaria de Estado de Saúde de Minas Gerais Arboviroses Urbanas (Dengue, Chikungunya e Zika). Boletim Epidemiológico No 296 - Semana Epidemiológica 40/2023. Available online: https://www.saude.mg.gov.br/images/1_noticias/06_2023/1-out-nov-dez/09-10-BO_ARBO296.pdf (accessed on 21 April 2024).
- SES-MG; Secretaria de Estado de Saúde de Minas Gerais Painel de Monitoramento de Casos. Vigilância Das Arboviroses Em Minas Gerais: Chikungunya Available online:. Available online: https://www.saude.mg.gov.br/aedes/painel (accessed on 21 April 2024).
- IBGE; Instituto Brasileiro de Geografia e Estatística Áreas Urbanizadas Do Brasil 2019. Available online: https://biblioteca.ibge.gov.br/visualizacao/livros/liv101973_informativo.pdf (accessed on 21 April 2024).
- Maia, L.J.; Oliveira, C.H. de; Silva, A.B.; Souza, P.A.A.; Müller, N.F.D.; Cardoso, J. da C.; Ribeiro, B.M.; Abreu, F.V.S. de; Campos, F.S. Arbovirus Surveillance in Mosquitoes: Historical Methods, Emerging Technologies, and Challenges Ahead. Exp Biol Med 2023, 248, 2072–2082. [Google Scholar] [CrossRef] [PubMed]
- Bonney, R.; Cooper, C.B.; Dickinson, J.; Kelling, S.; Phillips, T.; Rosenberg, K. V.; Shirk, J. Citizen Science: A Developing Tool for Expanding Science Knowledge and Scientific Literacy. Bioscience 2009, 59, 977–984. [Google Scholar] [CrossRef]
- Scolforo, J.R. Mapeamento e Inventário Da Flora Nativa e Dos Reflorestamentos de Minas Gerais. ; Ed. UFLA, Ed. 2006. [Google Scholar]
- Koppen, W. Das Geographische System Der Klimat. Handbuch der Klimatologie 1936, 1–46. [Google Scholar] [CrossRef]
- SES-MG; Secretaria de Estado de Saúde de Minas Gerais Arboviroses Urbanas (Dengue, Chikungunya e Zika). Boletim Epidemiológico No 270 - Semana Epidemiológica 05/2023, 06 de Fevereiro. Available online: https://www.saude.mg.gov.br/images/1_noticias/06_2023/4-jan-fev-marc/BO_ARBO270.pdf (accessed on 21 April 2024).
- Nasci, R.S. A Lightweight Battery-Powered Aspirator for Collecting Resting Mosquitoes in the Field. Mosq News 1981, 41, 808–811. [Google Scholar]
- Dubrulle, M.; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.-B. Chikungunya Virus and Aedes Mosquitoes: Saliva Is Infectious as Soon as Two Days after Oral Infection. PLoS One 2009, 4, e5895. [Google Scholar] [CrossRef] [PubMed]
- Consoli, R.A.G.B.; Oliveira, R.L. de Principais Mosquitos de Importância Sanitária No Brasil; Editora FIOCRUZ, 1994; ISBN 8575412906.
- Forattini, O.P. Culicidologia Médica: Identificação, Biologia, Epidemiologia - Volume 2; Edusp, Ed.; 1a.; Edusp: São Paulo, 2002; ISBN 8531406994. [Google Scholar]
- de Oliveira, C.H.; Andrade, M.S.; Campos, F.S.; da C. Cardoso, J. ; Gonçalves-dos-Santos, M.E.; Oliveira, R.S.; Aquino-Teixeira, S.M.; Campos, A.A.; Almeida, M.A.; Simonini-Teixeira, D.; et al. Yellow Fever Virus Maintained by Sabethes Mosquitoes during the Dry Season in Cerrado, a Semiarid Region of Brazil, in 2021. Viruses 2023, 15, 757. [Google Scholar] [CrossRef]
- Pastorino, B.; Bessaud, M.; Grandadam, M.; Murri, S.; Tolou, H.J.; Peyrefitte, C.N. Development of a TaqMan® RT-PCR Assay without RNA Extraction Step for the Detection and Quantification of African Chikungunya Viruses. J Virol Methods 2005, 124, 65–71. [Google Scholar] [CrossRef]
- Quick, J.; Grubaugh, N.D.; Pullan, S.T.; Claro, I.M.; Smith, A.D.; Gangavarapu, K.; Oliveira, G.; Robles-Sikisaka, R.; Rogers, T.F.; Beutler, N.A.; et al. Multiplex PCR Method for MinION and Illumina Sequencing of Zika and Other Virus Genomes Directly from Clinical Samples. Nat Protoc 2017, 12, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.R.R.; Menezes, M.T. de; Salgado-Benvindo, C.; Whittaker, C.; Cox, V.; Chandradeva, N.; Paula, H.H.S. de; Martins, A.F.; Chagas, R.R. das; Brasil, R.D.V.; et al. Epidemiological and Genomic Investigation of Chikungunya Virus in Rio de Janeiro State, Brazil, between 2015 and 2018. PLoS Negl Trop Dis 2023, 17, e0011536. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, V.; Libin, P.J.K.; Theys, K.; Faria, N.R.; Nunes, M.R.T.; Restovic, M.I.; Freire, M.; Giovanetti, M.; Cuypers, L.; Nowé, A.; et al. A Computational Method for the Identification of Dengue, Zika and Chikungunya Virus Species and Genotypes. PLoS Negl Trop Dis 2019, 13, e0007231. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol Biol Evol 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol Biol Evol 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.G.; Andrade, A.M.S.; Costa, M. da C.N.; Castro, J.S.M.; Oliveira, F.L.S.; Goes, C.S.B.; Maia, M.; Santana, E.B.; Nunes, B.T.D.; Vasconcelos, P.F.C. East/Central/South African Genotype Chikungunya Virus, Brazil, 2014. Emerg Infect Dis 2015, 21, 906–907. [Google Scholar] [CrossRef] [PubMed]
- Souza, U.J.B. de; Santos, R.N. dos; Giovanetti, M.; Alcantara, L.C.J.; Galvão, J.D.; Cardoso, F.D.P.; Brito, F.C.S.; Franco, A.C.; Roehe, P.M.; Ribeiro, B.M.; et al. Genomic Epidemiology Reveals the Circulation of the Chikungunya Virus East/Central/South African Lineage in Tocantins State, North Brazil. Viruses 2022, 14, 2311. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.; Alcantara, L.C.J.; Fonseca, V.; Lima, M.; Castro, E.; Fritsch, H.; Oliveira, C.; Guimarães, N.; Adelino, T.; Evaristo, M.; et al. Increased Interregional Virus Exchange and Nucleotide Diversity Outline the Expansion of Chikungunya Virus in Brazil. Nat Commun 2023, 14, 4413. [Google Scholar] [CrossRef]
- Lessa-Aquino, C.; Trinta, K.S.; Pestana, C.P.; Ribeiro, M.O.; Sucupira, M.V.F.; Boia, M.N.; Baptista, P.A.; Cunha, R. V.; Medeiros, M.A. Detection of East/Central/South African Genotype Chikungunya Virus during an Outbreak in a Southeastern State of Brazil. Epidemiol Infect 2018, 146, 2056–2058. [Google Scholar] [CrossRef]
- Aragão, C.F.; Pinheiro, V.C.S.; Nunes Neto, J.P.; Silva, E.V.P. da; Pereira, G.J.G.; Nascimento, B.L.S. do; Castro, K. da S.; Maia, A.M.; Catete, C.P.; Martins, L.C.; et al. Natural Infection of Aedes Aegypti by Chikungunya and Dengue Type 2 Virus in a Transition Area of North-Northeast Brazil. Viruses 2019, 11, 1126. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.F.; de Brito, B.B.; Correia, T.M.L.; Viana, A.I.S.; Carvalho, J.C.; da Silva, F.A.F.; Santos, M.L.C.; da Silveira, E.A.; Neto, H.P.G.; da Silva, N.M.P.; et al. Simultaneous Circulation of Zika, Dengue, and Chikungunya Viruses and Their Vertical Co-Transmission among Aedes Aegypti. Acta Trop 2021, 215, 105819. [Google Scholar] [CrossRef] [PubMed]
- Aragão, C.F.; Cruz, A.C.R.; Nunes Neto, J.P.; Monteiro, H.A. de O.; da Silva, E.V.P.; da Silva, S.P.; Andrade, A.T. dos S.; Tadei, W.P.; Pinheiro, V.C.S. Circulation of Chikungunya Virus in Aedes Aegypti in Maranhão, Northeast Brazil. Acta Trop 2018, 186, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, F.J.C.; Mourão, F.R.P.; Ribeiro, E.S.D.; Rêgo, M.O. da S.; Frances, P.A. da C.; Souto, R.N.P.; Façanha, M. dos S.; Tahmasebi, R.; Costa, A.C. da Prevalence of Dengue, Zika and Chikungunya Viruses in Aedes (Stegomyia) Aegypti (Diptera: Culicidae) in a Medium-Sized City, Amazon, Brazil. Rev Inst Med Trop Sao Paulo 2020, 62. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Velez, D.; Lounibos, L.P.; Lord, C.C. Transmission Risk of Two Chikungunya Lineages by Invasive Mosquito Vectors from Florida and the Dominican Republic. PLoS Negl Trop Dis 2017, 11, e0005724. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-de-Brito, A.; Ribeiro, I.P.; Miranda, R.M. de; Fernandes, R.S.; Campos, S.S.; Silva, K.A.B. da; Castro, M.G. de; Bonaldo, M.C.; Brasil, P.; Lourenço-de-Oliveira, R. First Detection of Natural Infection of Aedes Aegypti with Zika Virus in Brazil and throughout South America. Mem Inst Oswaldo Cruz 2016, 111, 655–658. [Google Scholar] [CrossRef]
- de Melo Ximenes, M. de F.F.; de Araújo Galvão, J.M.; Inacio, C.L.S.; Macêdo e Silva, V.P.; Pereira, R.L. do N.; Pinheiro, M.P.G.; de Medeiros Silva, M.M.; Gomes, C.E.S. Arbovirus Expansion: New Species of Culicids Infected by the Chikungunya Virus in an Urban Park of Brazil. Acta Trop 2020, 209, 105538. [Google Scholar] [CrossRef]
- Variza, P.F.; Lorenz, C.; Oliveira, J.G. de; Fernandes, M.; Netto, S.A.; Prophiro, J.S. Updated Spatio-Temporal Distribution of Aedes (Stegomyia) Albopictus in Brazil. Acta Trop 2022, 232, 106511. [Google Scholar] [CrossRef]
- Reiter, P.; Fontenille, D.; Paupy, C. Aedes Albopictus as an Epidemic Vector of Chikungunya Virus: Another Emerging Problem? Lancet Infect Dis 2006, 6, 463–464. [Google Scholar] [CrossRef]
- Bonilauri, P.; Bellini, R.; Calzolari, M.; Angelini, R.; Venturi, L.; Fallacara, F.; Cordioli, P.; Angelini, P.; Venturelli, C.; Merialdi, G.; et al. Chikungunya Virus in Aedes Albopictus, Italy. Emerg Infect Dis 2008, 14, 852–854. [Google Scholar] [CrossRef]
- Santos, M.E.G.; Sousa, E.C.; Bitencourt, C.S.; Oliveira, C.H.; Silva, A.J.J.; Ribeiro, J.C.; Franca, A.O.; Mendes, H.H.N.; Abreu, F.V.S. Monitoramento de Mosquitos Aedes Spp. (Diptera: Culicidae) Na Região Urbana Do Município de Salinas-MG, Norte de Minas Gerais.. In Proceedings of the Anais do Congresso da Sociedade Brasileira de Parasitologia - Parasito 2021: A Parasitologia na perspectiva da Saúde Única. ; Sociedade Brasileira de Parasitologia; 2021. [Google Scholar]
- Santos, T.P. Potential of Aedes Albopictus as a Bridge Vector for Enzootic Pathogens at the Urban-Forest Interfacein Brazil - Approved with Minor Corrections. Emerg Microbes Infect 2018. [Google Scholar]
- Corbel, V.; Fonseca, D.M.; Weetman, D.; Pinto, J.; Achee, N.L.; Chandre, F.; Coulibaly, M.B.; Dusfour, I.; Grieco, J.; Juntarajumnong, W.; et al. International Workshop on Insecticide Resistance in Vectors of Arboviruses, December 2016, Rio de Janeiro, Brazil. Parasit Vectors 2017, 10, 278. [Google Scholar] [CrossRef] [PubMed]
- Bonney, R.; Shirk, J.L.; Phillips, T.B.; Wiggins, A.; Ballard, H.L.; Miller-Rushing, A.J.; Parrish, J.K. Next Steps for Citizen Science. Science (1979) 2014, 343, 1436–1437. [Google Scholar] [CrossRef] [PubMed]
- Jordan, R.; Crall, A.; Gray, S.; Phillips, T.; Mellor, D. Citizen Science as a Distinct Field of Inquiry. Bioscience 2015, 65, 208–211. [Google Scholar] [CrossRef]
- Williams, C.R.; Hawthorn-Jackson, D.; Orre-Gordon, S.; O’Sullivan, S. Some Cautions in the Use of Citizen Science: A Case Study of Urban Insect Collection. Transactions of the Royal Society of South Australia 2017, 141, 57–69. [Google Scholar] [CrossRef]
- Liberatore, A.; Bowkett, E.; MacLeod, C.J.; Spurr, E.; Longnecker, N. Social Media as a Platform for a Citizen Science Community of Practice. Citiz Sci 2018, 3, 3. [Google Scholar] [CrossRef]
- Braz Sousa, L.; Fricker, S.R.; Doherty, S.S.; Webb, C.E.; Baldock, K.L.; Williams, C.R. Citizen Science and Smartphone E-Entomology Enables Low-Cost Upscaling of Mosquito Surveillance. Science of The Total Environment 2020, 704, 135349. [Google Scholar] [CrossRef]
- Palmer, J.R.B.; Oltra, A.; Collantes, F.; Delgado, J.A.; Lucientes, J.; Delacour, S.; Bengoa, M.; Eritja, R.; Bartumeus, F. Citizen Science Provides a Reliable and Scalable Tool to Track Disease-Carrying Mosquitoes. Nat Commun 2017, 8, 916. [Google Scholar] [CrossRef]
- Abreu, F.V.S. de; Delatorre, E.; dos Santos, A.A.C.; Ferreira-de-Brito, A.; de Castro, M.G.; Ribeiro, I.P.; Furtado, N.D.; Vargas, W.P.; Ribeiro, M.S.; Meneguete, P.; et al. Combination of Surveillance Tools Reveals That Yellow Fever Virus Can Remain in the Same Atlantic Forest Area at Least for Three Transmission Seasons. Mem Inst Oswaldo Cruz 2019, 114. [Google Scholar] [CrossRef]
- Abreu, F.V.S.; dos Santos, E.; Gomes, M.Q.; Vargas, W.P.; Oliveira Passos, P.H.; Nunes e Silva, C.; Araújo, P.C.; Pires, J.R.; Romano, A.P.M.; Teixeira, D.S.; et al. Capture of Alouatta Guariba Clamitans for the Surveillance of Sylvatic Yellow Fever and Zoonotic Malaria: Which Is the Best Strategy in the Tropical Atlantic Forest? Am J Primatol 2019, 81, e23000. [Google Scholar] [CrossRef]
- Chame, M.; Barbosa, H.J.C.; Gadelha, L.M.R.; Augusto, D.A.; Krempser, E.; Abdalla, L. SISS-Geo: Leveraging Citizen Science to Monitor Wildlife Health Risks in Brazil. J Healthc Inform Res 2019, 3, 414–440. [Google Scholar] [CrossRef] [PubMed]
- Chame, M.; Abdalla, L.; Pinter, A.; Romano, A.P.M.; Krempser, E.; Ramos, D.G.; Passos, P.H. de O.; Silva, P.C.L.; Da Silva, G.M.P.; Gatti, R.R.; et al. Primates in SISS-Geo: Potential Contributions of Mobile Technology, Health Surveillance and Citizen Science to Support Species Conservation in Brazil. Neotropical Primates 2020, 26, 80–89. [Google Scholar] [CrossRef]
- Sousa, L.B.; Craig, A.; Chitkara, U.; Fricker, S.; Webb, C.; Williams, C.; Baldock, K. Methodological Diversity in Citizen Science Mosquito Surveillance: A Scoping Review. Citiz Sci 2022, 7, 8. [Google Scholar] [CrossRef]
- Craig, A.T.; Kama, N.; Fafale, G.; Bugoro, H. Citizen Science as a Tool for Arboviral Vector Surveillance in a Resourced-Constrained Setting: Results of a Pilot Study in Honiara, Solomon Islands, 2019. BMC Public Health 2021, 21, 509. [Google Scholar] [CrossRef]
- Andrade, M.S.; Campos, F.S.; Oliveira, C.H. de; Oliveira, R.S.; Campos, A.A.S.; Almeida, M.A.B. de; Fonseca, V. de S.; Simonini-Teixeira, D.; Sevá, A. da P.; Temponi, A.O.D.; et al. Fast Surveillance Response Reveals the Introduction of a New Yellow Fever Virus Sub-Lineage in 2021, in Minas Gerais, Brazil. Mem Inst Oswaldo Cruz 2022, 117. [Google Scholar] [CrossRef]
Figure 1.
Map showing the Northern Region of Minas Gerais state, highlighting the city of Salinas and the specific neighborhoods where sampling was conducted.
Figure 1.
Map showing the Northern Region of Minas Gerais state, highlighting the city of Salinas and the specific neighborhoods where sampling was conducted.
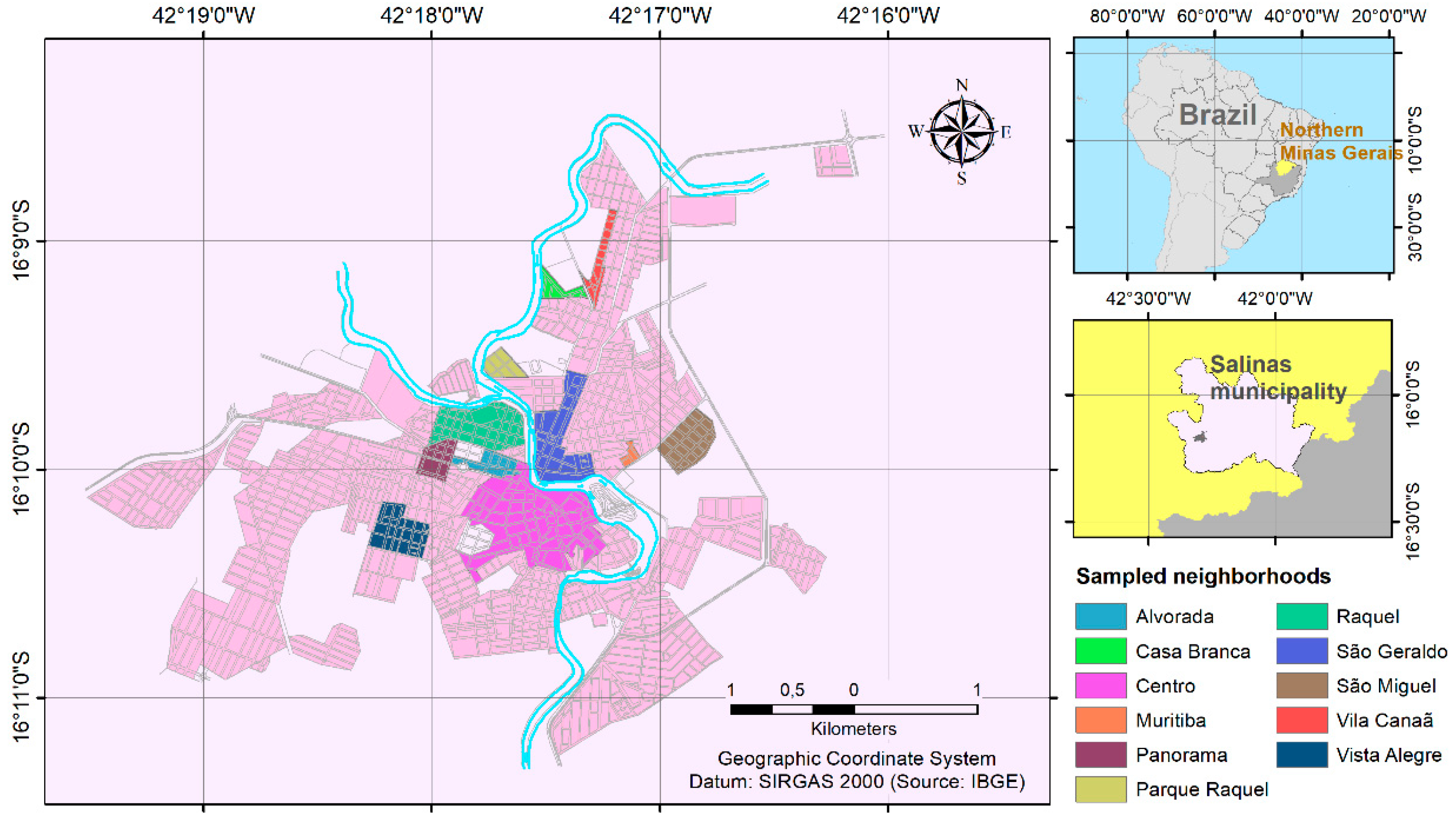
Figure 3.
Scheme showing the storage and testing carried out on each anatomical part (head, body and legs) of the mosquitoes. FBS = Fetal Bovine Serum.
Figure 3.
Scheme showing the storage and testing carried out on each anatomical part (head, body and legs) of the mosquitoes. FBS = Fetal Bovine Serum.
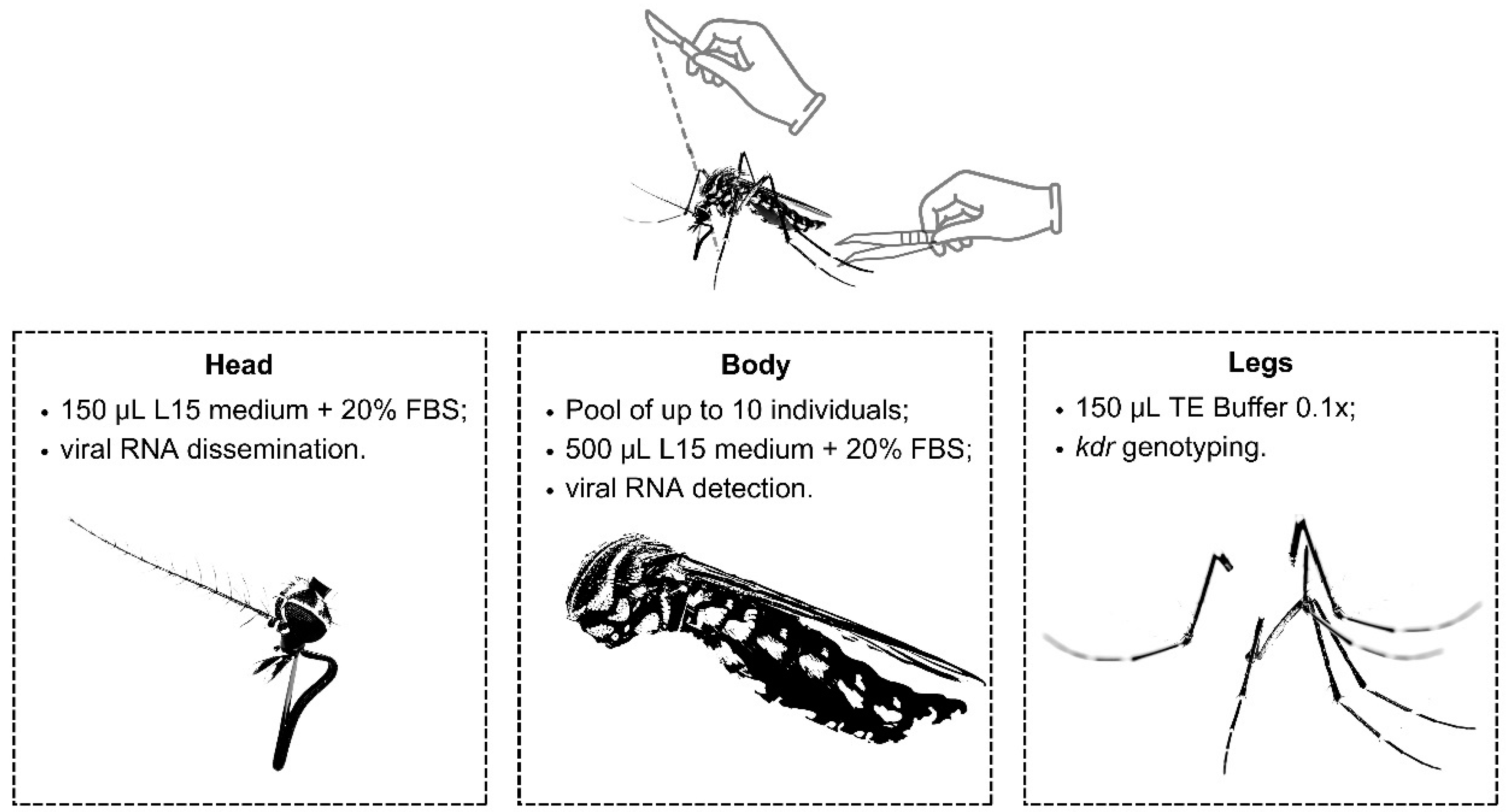
Figure 3.
Maximum likelihood tree of Chikungunya virus genomes, inferred with the genome generated in this study and 876 sequences publicly available from GenBank. The scale bar represents nucleotide substitutions per site (s/s), and the tree is mid-point rooted. The purple circle at the tip represents the genome sequence generated in this study (X-595, waiting GenBank accession number). Bootstrap values for major nodes are displayed.
Figure 3.
Maximum likelihood tree of Chikungunya virus genomes, inferred with the genome generated in this study and 876 sequences publicly available from GenBank. The scale bar represents nucleotide substitutions per site (s/s), and the tree is mid-point rooted. The purple circle at the tip represents the genome sequence generated in this study (X-595, waiting GenBank accession number). Bootstrap values for major nodes are displayed.
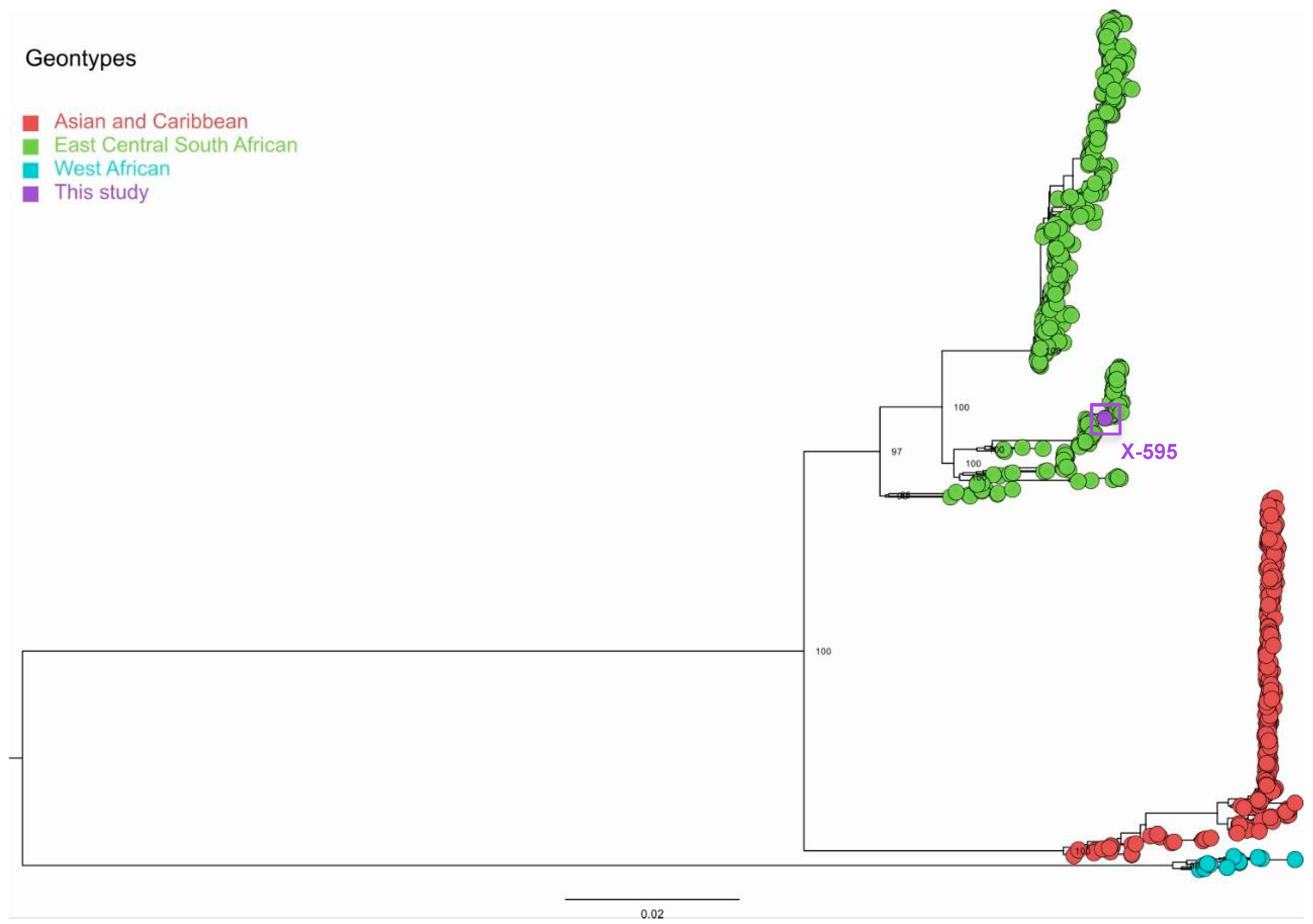

Table 1.
Captured mosquitoes and presence of CHIKV RNA in Salinas, MG, Brazil.
| Species | Male | Female | Sum (relative abundance %)# | Pools tested (CHIKV positive) | MIR* |
|---|---|---|---|---|---|
| Ae. aegypti (Linnaeus, 1762) | 70 | 95 | 165 (39.2) | 31 (10) | 60.6 |
| Cx. quinquefasciatus Say, 1823 | 143 | 113 | 256 (60.8) | 26 (2) | 7.8 |
| Total | 213 | 208 | 421 (100) | 57 (12) | 28.5 |
* Minimum Infection Rate (MIR) = nº of positive pools / nº of same species adults analyzed × 1000; #Relative abundance = nº of insects of each species / total nº of insects.
Table 2.
Description of CHIKV-positive pools and corresponding individual heads tested.
| Cod. Pool | Species | Nº of individuals | Sex | CT* | Heads | Positive heads (CT**) |
|---|---|---|---|---|---|---|
| X-595 | Ae. aegypti | 5 | F | 20.1 | c144, c145, c146, c147, c148 | c144 (21.5); c146 (22.0); c148 (30.1) |
| X-556 | Ae. aegypti | 5 | F | 21.0 | c33, c34, c35, c36, c37 | c34 (21.2) |
| X-585 | Ae. aegypti | 5 | F | 22.9 | c82, c83, c84, c88, 89 | c83 (23.6); c84 (35.8) |
| X-584 | Ae. aegypti | 2 | F | 23.1 | c81, c108 | c81 (21.6) |
| X-579 | Ae. aegypti | 5 | F | 24.1 | c51, c52, c54, c56, c58 | c58 (26.9) |
| X-594 | Ae. aegypti | 5 | F | 24.2 | c139, c140, c141, c142, c143 | c141 (23.0) |
| X-606 | Ae. aegypti | 5 | F | 28.0 | c149, c150, c151, c152, c164 | _ |
| X-555 | Ae. aegypti | 6 | M | 38.0 | c30, c32, c39, c40, c43, c44 | _ |
| X-593 | Ae. aegypti | 7 | M | 38.3 | c126, c128, c134, c135, c136, c137, c138 | _ |
| X-586 | Ae. aegypti | 8 | M | 40.0 | c85, c86, c87, c90, c91, c92, c93, c94 | _ |
| X-623 | Cx. quinquefasciatus | 9 | M | 38.1 | c296, c297, c298, c299, c300, c301, c302, c303, c304 | _ |
| X-619 | Cx. quinquefasciatus | 9 | F | 38.2 | c254, c255, c256, c257, c258, c259, c260, c261, c262 | _ |
*CT of reactions with the abdomen pools, **CT of reactions with each individualized heads from their respective positive pools.
Table 3.
NaV genotypes in Ae. aegypti from Salinas, MG, Brazil.
| Genotypes | VV+VV+ FF | VV+VV+ FC | VV+VV+ CC | VL+VI+ FC | VL+VI+ CC | LL+II+ CC | VV+Vl+ CC* | VV+VI+ FC | VL+II+ CC |
|---|---|---|---|---|---|---|---|---|---|
| Nº of individuals (Frequency) | 3 (1.8) |
2 (1.2) |
18 (11.0) |
17 (10.4) |
53 (32.3) |
67 (40.9) |
2 (1.2) |
1 (0.6) |
1 (0.6) |
Composite genotype for the three SNPs: 410 + 1016 + 1534, composed by the alleles NaVS (VVF), and the kdr R1 (VVC) and R2 (LIC). *Uncommon genotypes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



