Submitted:
30 April 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
Secondary metabolites are bioactive compounds produced by living organisms that can reveal symbiotic relationships in nature. In the work described here, soil-born entomopathogenic-nematodes associated with symbiotic bacteria (Xenorhabdus stockiae and Photorhabdus luminescens) extracted from solvent supernatant containing secondary metabolites showed significant inhibitory action against Escherichia coli, Staphylococcus aureus, Enterococcus faceium, Proteus vulgaris, Bacillus cereus, Bacillus subtilis. These secondary metabolites, characterized by Fourier transform infrared spectroscopy were found to be amine groups of proteins; hydroxyl and carboxyl groups of polyphenols; hydroxyl groups of polysaccharides, and carboxyl groups of organic acids. The major compounds identified by Gas chromatography-mass spectrometry analysis of ethyl acetate extracts of Xenorhabdus stockiae revealed the existence of Nonanoic acid derivatives, Paromycin, Pyrrolidinone, Octodecanal derivatives, Trioxa-5-aza-1-silabicyclo, 4-Octadecenal, Methyl ester, Oleic Acid and 1,2-benzenedicarboxylicacid. Additional extraction from Photorhabdus luminescens was expressed functional compounds such as Indole-3-acetic acid, Piperidinol derivatives, Phthalic acid, 1-Tetradecanol, Nemorosonol, 1-eicosanol and unsaturated fatty acids. These outcomes support the development of new alternative natural antimicrobial agents for pathogen suppression in the future.
Keywords:
Entomopathogenic nematodes
; Symbiotic bacteria
; PCR
; antimicrobial compounds
; FTIR
; GC-MS
1. Introduction
Natural compounds derived from living organisms show great chemical diversity and tremendous promise for finding new compounds with novel modes of action. Many organisms found in nature (fungi, plants and bacteria) produce antimicrobial compounds as secondary metabolites to compete with other organisms [1,2,3]. One of the biocontrol sources of Photorhabdus and Xenorhabdus genera show novel antimicrobial peptides. These bacteria encode several putative biosynthetic pathways for natural product biosynthesis and are involved in ecological functions [4,5]. Remarkably, Photorhabdus and Xenorhabdus symbiotic bacteria are associated with entomopathogenic nematodes (EPN) which are obligate and lethal insect parasites [6,7]. Once nematodes enter into insect, it releases their symbiotic bacteria into the insect hemolymph, within 2 to 3 days, the insect killed by bacterial toxins and enzymes [8,9].
Appropriate methods are required to utilize bacterial extracellular products for pest control and drug discovery applications. These symbiotic bacteria occur in the digestive tract of infective juveniles when in their primary phase but will transform into a secondary phase under in vitro conditions. Furthermore, biocontrol mechanisms of bacterial symbionts, in turn, kill the host and proliferate. To maintain suitable conditions for nematode reproduction, a nutrient supply and antimicrobial substances that inhibit the growth of a wide range of microorganisms within and outside the cadaver are required. There is space for production of supplementary bioactive compounds as uncharacterized secondary metabolites by these symbionts. The biosynthesis of antimicrobial and nematicidal metabolites strongly suggests that they inhibit consumption of the insect cadaver by competing organisms. The bacteria can be grown as free-living organisms in ideal laboratory conditions. Under unique temperature, pH and humidity conditions, they release a wide variety of virulent factors including high molecular weight toxin complexes; lipopolysaccharides; proteases; and arrays of different antibiotics, all of which can be assayed in the culture media.
Most species of Xenorhabdus and Photorhabdus produce more than one group of active secondary metabolites, and the metabolites from Xenorhabdus are more diverse than those of Photorhabdus [10,11]. These metabolites are extensively involved in a range of bioactivities in both pharmaceutical and agricultural fields [12] such as antibiotic, antimycotic, insecticidal, nematicidal, antiulcer, antineoplastic, and antiviral uses [13]. Consequently, nematode symbiotic bacteria can be considered an original source for potential biomedical applications [14]. The symbiotic bacteria have complex life cycles, and their insect hosts die rapidly due to production of insecticidal bacterial toxins [15]. Numerous studies will be needed to better understand symbiotic bacteria and their secretions. Various kinds of antibiotics against bacteria and fungi are synthesized and secreted within bacterial cultures of symbiotic bacteria [16]. Against this background, the present time is ideal for the exploration and production of bacterial metabolites for the control of bacterial pathogens. In the study described herein, the extraction of crude metabolites of Xenorhabdus and Photorhabdus is carried out with the goal of antimicrobial application development. These findings will afford better understanding of bacterial symbiont metabolites that can yield novel compounds useful for agricultural and pharmaceutical applications.
2. Results
2.1. Extraction of Symbiotic Bacteria
A total of 10 symbiotic bacteria-associated nematodes (soil beneficial) were isolated from different agriculture sites in Amazcala, Queretaro, Mexico (Table 1). Out of 10 recovered nematodes, the 2 potential symbiotic bacteria (strains 05 and 10) were found to have significant antibacterial activity against pathogens. Strains 5 and 10 were considered for bacterial isolation and molecular characterization. The symbiotic bacteria were isolated and identified through direct and indirect confirmation methods. At the primary level, identification of symbiotic bacterial colonies was observed based on dye absorption of bromothymol blue and triphenyl tetrazolium from NBTA plates. Blue-green, blue or brownish colonies resulted depending on species variation. Based on our observations of colony morphology, the two strains belonged to the genera Xenorhabdus spp. for Steinernema spp. and genera Photorhabdus spp. for Heterorhabditis spp. Figure 1 displays the general view on natural antibacterial crude secondary metabolites extracted from bacterial symbionts and identification of bioactive functional compounds by Gas chromatography-mass spectrometry (GC-MS).
2.2. Bacterial Identification by 16S rRNA
For further authentication, isolated symbiotic bacteria were identified at molecular level via 16S rRNA gene sequencing. The symbiont was amplified by using PCR primers representing regions of the 16s rRNA conserved in bacteria. Each strain produced a single band of approximately 1,450 bp. The 16s rRNA strain sequences (strains 05 and 10) were determined and showed 98 - 99% similarity with that of X. stockiae and P. luminescens respectively. Figure 2 displays the (A) Recovered soil entomopathogenic nematodes molecular characterization (strains number 05 and 10); (B) Extracted symbiotic bacteria molecular characterization (strains number 05 and 10).
2.3. Functional Group Analysis by FTIR
The FT-IR spectrum recorded for the cell free supernatant of X. stockiae, displayed peaks at 3443, 1637, 1100 and 621 cm-1. In this feature Figure 3 shows the FT-IR analysis of bacterial crude compounds of X. stockiae (A); P. luminescens (B). Table 2 shows the FT-IR spectrum of X. stockiae crude compound with assigned possible functional groups. In the resultant FT-IR characterization spectrum between bacterial culture extracts, major shifts were observed from 3436 cm-1 to 3443 cm-1. Table 3 shows the FT-IR spectrum of P. luminescens crude compound in possible assigned functional groups.
2.4. Secondary Chemical Compound Analyses for Symbiotic Bacterial in the Ethyl Acetate Crude Extract
Purified secondary metabolites were analyzed using GC–MS to obtain a basic idea of the metabolites present, and to compare the chemical profiles of each peak. Figure 4 illustrations of GC-MS chromatogram of Symbiotic bacteria (X. stockiae); Figure 5 GC-MS chromatogram of Symbiotic bacteria (P. luminescens). Interestingly, Table 4 shows summary of the fragments and retention time for significant compounds identified in X. stockiae strain from ethyl acetate crude extract. GC peaks were present corresponding to various compounds with reported biological activity including Nonanoic acid derivatives, Paromycin, Pyrrolidinone, Octodecanal derivatives, Trioxa-5-aza-1-silabicyclo, 4-Octadecenal, Methyl ester, Oleic Acid and 1,2-benzenedicarboxylicacid. Resuming that Table 5 displays the summary of the fragments and retention time for significant compounds identified in P. luminescens strain from ethyl acetate crude extract. Additional extraction from Photorhabdus luminescens was expressed functional compounds such as Indole-3-acetic acid, Piperidinol derivatives, Phthalic acid, 1-Tetradecanol, Nemorosonol, 1-eicosanol and unsaturated fatty acids.

Table 4.
Summary of the fragments and retention time for significant compounds identified in X. stockiae strain from ethyl acetate crude extract.
Table 4.
Summary of the fragments and retention time for significant compounds identified in X. stockiae strain from ethyl acetate crude extract.
| S. No. | R. Time | % of area | Compound name | Biological activities | Reference |
|---|---|---|---|---|---|
| 1 | 5.631 | 3.89 | Nonanoic acid derivatives | Antimicrobial properties | [17] |
| 2 | 5.823 | 4.27 | Paromycin | Antibacterial agents | [18] |
| 3 | 7.428 | 42.01 | Pyrrolidinone | Bioactive compounds | [19] |
| 4 | 8.848 | 3.98 | Octodecanal derivatives | Bioactive compounds | [20] |
| 5 | 13.198 | 4.53 | Trioxa-5-aza-1-silabicyclo | Anti-microbial compunds | [21] |
| 6 | 15.671 | 26.78 | 4-Octadecenal | Antifungal activity | [22] |
| 7 | 22.981 | 2.84 | Cyclopentanetridecanoic acid, Methylester | antimicrobial peptide | [23] |
| 8 | 23.978 | 4.629 | Oleic Acid | Antifungal compound | [24] |
| 9 | 31.98 | 1.876 | 1,2-benzenedicarboxylicacid | Bioactive molecules | [25] |
Table 5.
Summary of the fragments and retention time for significant compounds identified in P. luminescens strain from ethyl acetate crude extract.
Table 5.
Summary of the fragments and retention time for significant compounds identified in P. luminescens strain from ethyl acetate crude extract.
| S.No | R.Time | % of area | Compound name | Biological activities | Reference |
|---|---|---|---|---|---|
| 1 | 6.328 | 2.95 | Indole-3-acetic acid, methyl ester | Bioactive compound | [26] |
| 2 | 7.810 | 59.43 | Piperidinol derivatives | Antimicrobial agents | [27] |
| 3 | 8.302 | 5.78 | Phthalic acid | Bioactive compound | [28] |
| 4 | 12.816 | 5.91 | 1-Tetradecanol | Bioactive compound | [29] |
| 5 | 15.114 | 49.43 | Nemorosonol | Bioactive metabolites | [30] |
| 6 | 16.831 | 5.187 | octahydro-7-methyl-3-methylene | Insecticidal activity | [31] |
| 7 | 20.302 | 3.651 | 1-eicosanol | Bioactive compound | [32] |
| 8 | 25.220 | 2.284 | Octadecanoic acid, Methylester | Antimicrobial activity | [33] |
| 9 | 26.897 | 1.211 | unsaturated fatty acids | Anti-inflammation | [34] |
2.5. Antibacterial Activities of Symbiotic Bacteria
In our study revealed that the ethyl acetate extract of crude compound from X. stockiae and P. luminescens could inhibit up to 06 strains of pathogenic bacteria. Figure 6 describes the general mechanisms of antimicrobial action on bioactive functional compounds from symbiotic bacteria. In addition, Table 6 represents the antibacterial effects of symbiotic bacteria (X. stockiae and P. luminescens), illustrating the high level of inhibitory effects against tested pathogenic organisms in impregnated discs. ZOI appeared in all test organisms such as E. coli, S. aureus, E. faceium, P. vulgaris; B. cereus; B. subtilis. The highest ZOI was found for E. coli, S. aureus, E. faceium. Moderate levels of ZOI were observed for P. vulgaris; B. cereus; B. subtilis. Subsequently, Figure 7 displays the antibacterial action of symbiotic bacterial (X. stockiae and P. luminescens) crude compound against bacterial pathogen.
3. Discussion
These symbiotic bacteria are closely related to the family Enterobacteriaceae. The phylogeny of these organisms is well-defined in the sense that they are clearly placed in the gamma group of proteobacteria [35]. Each isolate was associated with a distinct bacterial genus belonging to different X. stockiae and P. luminescens. Only a few strains of the symbiotic bacteria have been described and studied in detail, and their molecular biology has been described by [36]. The primary phase is the major antibiotic producing phase, whereas the second phase produces fewer antibiotic molecules. Bacteria in this genus have also been proven to be resource-rich in natural products such as insecticidal toxins, insect immune system inhibitors, and a variety of antibiotics that facilitate the infection of nematodes [37]. These metabolites have been used in biological pesticides and therapeutic agents for many decades [38]. Admittedly, we exhibited the screening of antimicrobial crude secondary metabolites extracted from bacterial strains was carried out for further characterization.
The broad-spectrum peak at 3443 cm-1 corresponds to strong stretching vibrations of the hydroxyl functional group [39]. The bands at 1100 and 1637 cm-1 correspond to the ANAH and carbonyl (CAOA) stretching vibrations in amide linkages (amide I and amide II) of protein present in bacterial supernatant [40]. The small peak at 621 cm-1 is characteristic of carbonyl stretching vibrations in the amide II functional group [41]. The observed FT-IR spectrum results confirmed presence of the hydroxyl functional group as well as slight shift changes in all peak positions and absorption bands. These features may also be responsible for antimicrobial and anticancer properties. Xenorhabdus spp. have been reported to produce antimicrobial activity; indole compounds identified in culture broths of several Xenorhabdus spp. showed antibacterial and antifungal activity [42,43].
Xenocoumacins are polyketide-derived compounds produced by Xenorhabdus bacteria. In GC-MS outcomes pyrrolidine functional group involved in the late steps of xenocoumacin production. These compounds have been shown to possess antimicrobial activity against a variety of bacteria, including Gram-positive and Gram-negative pathogens. Xenocoumacins exert their antimicrobial effects by inhibiting bacterial RNA polymerase, leading to the suppression of bacterial growth and viability. Additionally, GC-MS results showed that P. luminescens produced secondary metabolites of 1-tetradecanol compounds. Recently, these compounds were extracted from nematode symbiotic bacteria and evaluated against insect pathogens (Pieris rapae and Pentodon algerinus) [44]. Another important and previously evaluated secondary metabolite compound; 1, 2, benzenedicarboxylic acid; represents a potential antibacterial agent. Similarly, different kinds of Xenorhabdus strains produce many bioactive compounds including antibacterial, antifungal and cytotoxicity [45]. Furthermore, octadecanoic acid and methyl ester extracted from Photorhabdus bacteria, have potential antimicrobial, anti-inflammatory, antioxidant and antibacterial activities based on earlier studies [46,47]. Photorhabdus bacteria produce phenazines, nitrogen-containing heterocyclic compounds with broad-spectrum antimicrobial activity. Phenazines act by generating reactive oxygen species within bacterial cells, causing oxidative damage to proteins, lipids, and DNA. This oxidative stress ultimately leads to bacterial cell death. Another critical extracted compound in this study, Phthalic acid, Indole-3-acetic aci and Oleic Acid was also extracted by [48]. They screened anti-protozoal activity of supernatants containing secondary metabolites produced by Photorhabdus and Xenorhabdus species against antiprotozoal compounds using the easy PACId approach (easy Promoter Activated Compound Identification) method [49]. Admittedly, Bacterial species in genera Xenorhabdus and Photorhabdus can produce various secondary metabolites to maintain their mutualistic symbiosis with the host entomopathogenic nematode. This study assessed all metabolites extracted with ethyl acetate solvents from bacterial culture broth of Xenorhabdus and Photorhabdus to identify their virulent secondary metabolites against bacterial pathogens.
Antimicrobial and growth inhibitory effects can be attributed to either the bacteria or its metabolites [50,51,52]. Our result positively correlates with [53,54]. Moreover, the antibacterial assay time for suppress the E. coli, S. aureus, E. faceium, P. vulgaris, B. cereus, and B. subtilis was inhibited bacterial pathogens. Particularly, in the present study X. stockiae could strongly inhibit the growth of 06 strains of pathogenic bacteria, compared to P. luminescences; Which is followed by E. coli, S. aureus and E. faceium. This scenario established that X. stockiae has the potential inhibition activity against bacterial pathogens. General action of antibacterial activity through different mechanisms according to [55,56]. Figure 5 displays the mechanisms of antimicrobial action against pathogens using secondary metabolites of bacterial symbionts. For instance, inhibition of bacterial cell wall membrane by alteration of amino acid sugar in linear form that cross-link through peptidoglycan layer and inhibition of biochemical pathways such as nucleic acid metabolism, translation, and transportation. On the other hand, changes in cell membrane integrity by electrostatic interaction combined with negatively charged membrane for the cell death. Additional interesting view is (1) inhibition of DNA synthesis via cross linking DNA; (2) prevention of DNA relaxation via activation DNA topoisomerase I; (3) disrupting the protein-folding cycle (4) proteolytic activity causing the degradation of DNA-RNA-Protein replication, leads to cell damages.
Based on previous reports we established antimicrobial compounds from Xenorhabdus spp. and Photorhabdus spp. may inhibit bacterial growth [57]. This is the right time finding the antimicrobial mechanism of specific target on bacterial cell membrane for the novel antimicrobial drug discovery. For example, Xenorhabdus inhibited the growth of closely related bacteria, whereas metabolic compounds were effective against a range of plant pathogens and some mammal disease pathogens [58,59,60,61]. On the other hand, such antimicrobial compounds present in the culture extract of Photorhabdus bacteria have the potential to be exploited for use against a wide range of gram-positive and gram-negative bacteria that pose challenges in medical and agricultural fields [62]. Similarly, our effects of antibacterial inhibition of P. luminescens against pathogens were certainly associated with [63]. These compounds are predominantly broad-spectrum antibacterial that are highly active against a wide range of gram-positive and gram-negative bacteria of medical and agricultural importance. According to [64], siderophore production is an Entomopathogenic characteristic of the Photorhabdus spp. through which they chelate the Fe3+ from the hemolymph, leaving the insect deprived of Fe3+ and subsequently killing it. Consequently, we studied novel antibacterial agents, predominantly bacterial products from various origins and geographic locations. Here, investigations were undertaken to find novel antibacterial agents from symbiotic bacteria (Xenorhabdus and Photorhabdus). Ideally, in the near future, their structures, biosynthesis, and mechanisms of action will be elucidated, and their production will be optimized.
4. Materials and Methods
4.1. Chemicals and Media
All the chemicals used for extraction and gas chromatography-mass spectrometry grade methanol were purchased from Merck, Mexico. Microbiological media were obtained from Hi-Media Laboratory, Querétaro, México.
4.2. Identification, Extraction and Molecular Characterization of Symbiotic Bacteria
Symbiotic bacteria (Xenorhabdus and Photorhabdus) were isolated in parallel from their nematode symbionts Steinernema and Heterorhabditis, respectively. For instance, symbiotic bacteria were extracted from newly emerged IJ nematodes, surface sterilized, and then subject to grinding. The final suspension was streaked on NBTA agar plates (0.004 nutrient agar; 2, 3, 5-triphenyltetrazolium chloride and 0.025% bromothymol blue) according to [65]. The pure bacterial colonies were obtained from both nematodes.
4.3.16S rDNA Sequencing and Phylogenetic Analysis
DNA extraction was carried out on the symbiont bacteria with a slight modification [66]. The small subunit (16s) rRNA was amplified with a PCR system, and the final products were separated by agarose gel and visualized by a transilluminator imaging system followed by previous screening of laboratory procedure [67]. Here, we used forward primer - (16s 20) 5’ AGA GTT TGA TCC TGG CTC - 3’ and reverse primer - (16s 1390) 5’ GAC GGG CGG TGT GTA CAA - 3’. The resultant high-quality sequences were analyzed with BLASTn (NCBI) to confirm authenticity of the bacterium.
4.4. Extraction of Bioactive Crude Compound from Symbiotic Bacteria Using Ethyl Acetate
Bacterial colonies were established on NBTA agar plates. Photorhabdus and Xenorhabdus occur as two-phase variants (primary and secondary) yet, for the most part, only the primary phase that produces antibiotics [68]. Thus, it was in our interest to maintain bacteria in the primary form. Soluble organic metabolites were then extracted from the bacterial cultures according to [69].
4.5. Purification of Bioactive Compound
Bacterial cultures were scaled up for metabolite isolation through liquid culture in TSY (Tryptic Soy Broth +0.5% yeast extract). A loopful of bacteria was added to 50 ml of fresh TSY in a 300 ml Erlenmeyer flask and placed on a rotary incubator shaker at 25ºC and 130 rpm for 24 hrs. The cultures were then transferred to 900 ml TSY in 2-liter flasks and placed on a rotary shaker at 25ºC for 96 hrs. Solvent extraction methods were utilized to separate bacterial metabolites based on their relative solubilities in two different immiscible liquids [70]. These steps were performed using separatory funnels and countercurrent distribution equipment. The supernatants were extracted three times with ethyl acetate, organic fractions were dried with anhydrous ammonium sulphate, concentrated in a rotary evaporator, and dissolved in acetone.
4.6. FT-IR and GC-MS Analysis
The ethyl acetate extract was characterized by a Perkin Elmer Spectrophotometer FT-IR spectrum for determination of the attached functional groups in a scanning range of 0 to 4000 cm-1 with resolution of 4 cm-1. Gas chromatography mass spectrophotometry was employed for the analysis of active constituents in Xenorhabdus spp. and Photorhabdus spp. bacterial extracts using a GCMS-QP2010 Plus gas chromatograph interfaced to a mass spectrometer. The sample was introduced into glass injector working in the split mode with helium as the carrier gas and a linear velocity pressure of 81.7 kPa. The following conditions were used: Rtx-5 MS fused silica capillary column (30 meters X 0.25 mm. i. d. X 0.25 um film thickness). The following temperatures were used: column oven temp- 80.0 °C, Injection Temp.- 270.00 °C. The constituents were identified using commercial libraries.
4.7. Antibacterial Assay
Extracts of symbiotic bacterial (Xenorhabdus spp. and Photorhabdus spp.) crude compounds were evaluated against both Gram-positive and negative bacteria. Bacterial strains such as E. coli (MTCC – 2622), S. aureus (MTCC – 3852), E. faceium (MTCC - 5153), P. vulgaris (MTCC - 1429), B. cereus (MTCC - 3159) and B. subtilis (MTCC - 4831) were obtained from Microbial type culture collection and Gene Bank, Mexico; It was sub-cultured in nutrient broth for 24 hrs at 30 ºC. For biological activity, each strain was swabbed consistently into the individual nutrient agar plates using sterile cotton swabs [71]. With a sterile micropipette, each extracted bacterial crude compound (Xenorhabdus spp. was taken 10 and 20 μg/ml; Photorhabdus spp. was taken 10 and 20 μg/ml), was loaded into an each well. Ampicillin solvent served as the positive control (10 μg/ml). After 3 min, sterilized paper disks were pressed lightly on the surface of pathogenic plates. The doses were selected based on preliminary data obtained from earlier studies. After 24 hrs incubation at 37 ºC, the different levels of zone of inhibition (ZOI) were measured.
4.5. Statistics
The results were expressed as the means standard deviation in all experiments. All the experiments were conducted in at least triplet.
5. Conclusion
In conclusion, the antimicrobial properties of natural products on Xenorhabdus and Photorhabdus symbiotic bacteria present promising avenues for further exploration in the field of microbiology and biotechnology. Furthermore, the eco-friendly nature of natural products aligns with the growing demand for sustainable solutions in combating bacterial infections and antibiotic resistance. Despite the promising findings, it is important to acknowledge the challenges associated with translating these discoveries into practical applications. Further research is needed to elucidate the mechanisms of action underlying the antimicrobial activity of natural products against Xenorhabdus and Photorhabdus symbiotic bacteria, as well as their safety profiles and potential for resistance development. Additionally, optimization of extraction methods, formulation techniques, and delivery systems will be crucial for maximizing the efficacy and stability of natural product-based antimicrobials. Overall, the exploration of natural products as antimicrobial agents against Xenorhabdus and Photorhabdus symbiotic bacteria offers exciting opportunities for innovation and development in the field of antimicrobial research. By harnessing the power of nature's chemical diversity, we may uncover novel solutions to combat bacterial infections and contribute to the ongoing efforts to address the global challenge of antibiotic resistance.
Author Contributions
Conceptualization, methodology, investigation, writing-original draft preparation G. Ch. authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
This study did not require ethical approval.
Informed Consent Statement
This study did not involve humans.
Data Availability Statement
Data will be available on request.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Chandrakasan, G.; Rodríguez-Hernández, A.-I.; López-Cuellar, M.d.R.; Palma-Rodríguez, H.-M.; Chavarría-Hernández, N. Bacteriocin encapsulation for food and pharmaceutical applications: advances in the past 20 years. Biotechnol. Lett. 2019, 41, 453–469. [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products As Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [CrossRef]
- Dongjin, J.; Youngkeun, Y.; Kang, G.H.; Choi, Y.H.; Kim, P.; Baek, N.I.; et al. Identification of an antibacterial compound, benzylideneacetone, from Xenorhabdus nematophila against major plant-pathogenic bacteria. FEMS Microbiol. Lett. 2004, 10-15.
- Bode, H.B. Entomopathogenic bacteria as a source of secondary metabolites. Curr. Opin. Chem. Biol. 2009, 13, 224–230. [CrossRef]
- Shi, Y.-M.; Hirschmann, M.; Shi, Y.-N.; Ahmed, S.; Abebew, D.; Tobias, N.J.; Grun, P.; Crames, J.J.; Poeschel, L.; Kuttenlochner, W.; et al. Global analysis of biosynthetic gene clusters reveals conserved and unique natural products in entomopathogenic nematode-symbiotic bacteria. Nat. Chem. 2022, 14, 701–+. [CrossRef]
- Boemare, N. Biology, taxonomy and systematics of Photorhabdus and Xenorhabdus. Entomopathogenic nematology, 2002, pp. 35-56. CABI Publishing, CAB International, Wallingford UK.
- Hazir, S.; Stackebrandt, E.; Lang, E.; Schumann, P.; Ehlers, R.-U.; Keskin, N. Two new Subspecies of Photorhabdus luminescens, Isolated from Heterorhabditis bacteriophora (Nematoda: Heterorhabditidae): Photorhabdus luminescens subsp. kayaii subsp. nov. and Photorhabdus luminescens subsp. thracensis subsp. nov.. Syst. Appl. Microbiol. 2004, 27, 36–42. [CrossRef]
- Loulou, A.; Mastore, M.; Caramella, S.; Bhat, A.H.; Brivio, M.F.; Machado, R.A.R.; Kallel, S. Entomopathogenic potential of bacteria associated with soil-borne nematodes and insect immune responses to their infection. PLOS ONE 2023, 18, e0280675. [CrossRef]
- Shapiro-Ilan, D.I.; Hazir, S.; Glaser, I. 2020. Advances in Use of Entomopathogenic Nematodes in Integrated Pest Management. In Integrated Management of Insect Pests: Current and Future Developments, 2020, pp. 91-105. Burleigh Dodds Science Publishing.
- Abd-Elgawad, M.M.M. Xenorhabdus spp.: An Overview of the Useful Facets of Mutualistic Bacteria of Entomopathogenic Nematodes. Life 2022, 12, 1360. [CrossRef]
- Yimthin, T.; Fukruksa, C.; Muangpat, P.; Dumidae, A.; Wattanachaiyingcharoen, W.; Vitta, A.; Thanwisai, A. A study on Xenorhabdus and Photorhabdus isolates from Northeastern Thailand: Identification, antibacterial activity, and association with entomopathogenic nematode hosts. PLOS ONE 2021, 16, e0255943. [CrossRef]
- Wang, Y.; Fang, X.; Cheng, Y.; Zhang, X. Manipulation of pH Shift to Enhance the Growth and Antibiotic Activity ofXenorhabdus nematophila. J. Biomed. Biotechnol. 2011, 2011, 1–9. [CrossRef]
- Lulamba, T.E.; Green, E.; Serepa-Dlamini, M.H. Photorhabdus sp. ETL Antimicrobial Properties and Characterization of Its Secondary Metabolites by Gas Chromatography–Mass Spectrometry. Life 2021, 11, 787. [CrossRef]
- Chandrakasan, G.; Seetharaman, P.; Gnanasekar, S.; Kadarkarai, M.; Sivaperumal, S. Xenorhabdus stockiae KT835471-mediated feasible biosynthesis of metal nanoparticles for their antibacterial and cytotoxic activities. Artif. Cells, Nanomedicine, Biotechnol. 2017, 45, 1675–1684. [CrossRef]
- Heermann, R.; Fuchs, T.M. Comparative analysis of the Photorhabdus luminescens and the Yersinia enterocolitica genomes: uncovering candidate genes involved in insect pathogenicity. BMC Genom. 2008, 9, 40–40. [CrossRef]
- Webster, J.M.; Chen, G.; Hu, K.; Li, J. Bacterial metabolites. Entomopathogenic Nematology, 2002, pp. 99-114. CABI Publishing, CAB International, Wallingford, UK.
- Sahin, N.; Kula, I.; Erdogan, Y. Investigation of antimicrobial activities of nonanoic acid derivatives. Fresenius Environmental Bulletin, 2006, 15(2), 141-143.
- Pantel, L.; Florin, T.; Dobosz-Bartoszek, M.; Racine, E.; Sarciaux, M.; Serri, M.; Houard, J.; Campagne, J.-M.; de Figueiredo, R.M.; Midrier, C.; et al. Odilorhabdins, Antibacterial Agents that Cause Miscoding by Binding at a New Ribosomal Site. Mol. Cell 2018, 70, 83–94.e7. [CrossRef]
- Reimer, D.; Luxenburger, E.; Brachmann, A.O.; Bode, H.B. A New Type of Pyrrolidine Biosynthesis Is Involved in the Late Steps of Xenocoumacin Production in Xenorhabdus nematophila. ChemBioChem 2009, 10, 1997–2001. [CrossRef]
- Gad, H.A.; Mukhammadiev, E.A.; Zengen, G.; Al Musayeib, N.M.; Hussain, H.; Bin Ware, I.; Ashour, M.L.; Mamadalieva, N.Z. Chemometric Analysis Based on GC-MS Chemical Profiles of Three Stachys Species from Uzbekistan and Their Biological Activity. Plants 2022, 11, 1215. [CrossRef]
- Taher, M.A.; Laboni, A.A.; Shompa, S.A.; Rahman, M.; Hasan, M.M.; Hasnat, H.; Khan, M. Bioactive compounds extracted from leaves of G. cyanocarpa using various solvents in chromatographic separation showed anti-cancer and anti-microbial potentiality in in silico approach. Chin. J. Anal. Chem. 2023, 51. [CrossRef]
- Baiome, B.A.; Ye, X.; Yuan, Z.; Gaafar, Y.Z.A.; Melak, S.; Cao, H. Identification of Volatile Organic Compounds Produced by Xenorhabdus indica Strain AB and Investigation of Their Antifungal Activities. Appl. Environ. Microbiol. 2022, 88, e0015522. [CrossRef]
- Karthik, Y.; Kalyani, M.I.; Krishnappa, S.; Devappa, R.; Goud, C.A.; Ramakrishna, K.; Wani, M.A.; Alkafafy, M.; Abduljabbar, M.H.; Alswat, A.S.; et al. Antiproliferative activity of antimicrobial peptides and bioactive compounds from the mangrove Glutamicibacter mysorens. Front. Microbiol. 2023, 14, 1096826. [CrossRef]
- Hawar, S.N.; Taha, Z.K.; Hamied, A.S.; Al-Shmgani, H.S.; Sulaiman, G.M.; Elsilk, S.E. Antifungal Activity of Bioactive Compounds Produced by the Endophytic Fungus Paecilomyces sp. (JN227071.1) against Rhizoctonia solani. Int. J. Biomater. 2023, 2023, 1–8. [CrossRef]
- Awori, R.M. Nematophilic bacteria associated with entomopathogenic nematodes and drug development of their biomolecules. Front. Microbiol. 2022, 13, 993688. [CrossRef]
- Joyce, S.A.; Lango, L.; Clarke, D.J. The regulation of secondary metabolism and mutualism in the insect pathogenic bacterium Photorhabdus luminescens. In Advances in applied microbiology, 2011, 76: 1-25). Academic Press.
- Breijyeh, Z.; Karaman, R. Design and Synthesis of Novel Antimicrobial Agents. Antibiotics 2023, 12, 628. [CrossRef]
- Sanda, N.B.; Hou, Y. The Symbiotic Bacteria—Xenorhabdus nematophila All and Photorhabdus luminescens H06 Strongly Affected the Phenoloxidase Activation of Nipa Palm Hispid, Octodonta nipae (Coleoptera: Chrysomelidae) Larvae. Pathogens 2023, 12, 506.
- Abdel-Wareth, M.T.A.; Ali, E.A.M.; El-Shazly, M.A. (2023). Biological Activity and GC-MS/MS Analysis of Extracts of Endophytic Fungi Isolated from Eichhornia crassipes (Mart.) Solms. Journal of Applied Biotechnology Reports, 2023, 10(1), 895-909.
- Yellamanda, B.; Vijayalakshmi, M.; Kavitha, A.; Reddy, D.K.; Venkateswarlu, Y. Extraction and bioactive profile of the compounds produced by Rhodococcus sp. VLD-10. 3 Biotech. 2016 (2):261.
- Sato, K.; Yoshiga, T.; Hasegawa, K. Involvement of Vitamin B 6 Biosynthesis Pathways in the Insecticidal Activity of Photorhabdus luminescens. Appl. Environ. Microbiol. 2016, 82, 3546–3553. [CrossRef]
- Abdel-Wareth, M.T.A.; Ali, E.A.M.; El-Shazly, M.A. Biological Activity and GC-MS/MS Analysis of Extracts of Endophytic Fungi Isolated from Eichhornia crassipes (Mart.) Solms. Journal of Applied Biotechnology Reports, 2023, 10(1), 895-909.
- Koilybayeva, M.; Shynykul, Z.; Ustenova, G.; Abzaliyeva, S.; Alimzhanova, M.; Amirkhanova, A.; Turgumbayeva, A.; Mustafina, K.; Yeleken, G.; Raganina, K.; et al. Molecular Characterization of Some Bacillus Species from Vegetables and Evaluation of Their Antimicrobial and Antibiotic Potency. Molecules 2023, 28, 3210. [CrossRef]
- Farag, M.A.; Gad, M.Z. Omega-9 fatty acids: potential roles in inflammation and cancer management. J. Genet. Eng. Biotechnol. 2022, 20, 48–11. [CrossRef]
- Sangeetha, B.G.; Jayaprakas, C.A.; Siji, J.V.; Rajitha, M.; Shyni, B.; Mohandas, C. Molecular characterization and amplified ribosomal DNA restriction analysis of entomopathogenic bacteria associated with Rhabditis (Oscheius) spp.. 3 Biotech 2016, 6, 32. [CrossRef]
- Forst, S.; Nealson, K. 1996. Molecular Biology of the Symbiotic-Pathogenic Bacteria Xenorhabdus spp. and Photorhabdus spp. Microbiol. Rev.1996, 60: 21-43.
- Clarke, D.J. Photorhabdus: a model for the analysis of pathogenicity and mutualism. Cell. Microbiol. 2008, 10, 2159–2167. [CrossRef]
- Dreyer, J.; Malan, A.P.; Dicks, L.M.T. Bacteria of the Genus Xenorhabdus, a Novel Source of Bioactive Compounds. Front. Microbiol. 2018, 9, 3177. [CrossRef]
- Priyadarshini, S.; Gopinath, V.; Priyadharsshini, N.M.; MubarakAli, D.; Velusamy, P. Synthesis of anisotropic silver nanoparticles using novel strain, Bacillus flexus and its biomedical application. Colloids Surfaces B: Biointerfaces 2013, 102, 232–237. [CrossRef]
- San-Blas, E.; Cubillán, N.; Guerra, M.; Portillo, E.; Esteves, I. Characterization of Xenorhabdus and Photorhabdus bacteria by Fourier transform mid-infrared spectroscopy with attenuated total reflection (FT-IR/ATR). Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 2012, 93, 58–62. [CrossRef]
- Gopinath, K.; Gowri, S.; Arumugam, A. 2013. Phytosynthesis of silver nanoparticles using Pterocar pussantalinus leaf extract and their antibacterial properties. J. Nanostructure Chem. 2013, 3: 68.
- Cimen, H.; Touray, M.; Gulsen, S.H.; Erincik, O.; Wenski, S.L.; Bode, H.B.; Shapiro-Ilan, D.; Hazir, S. Antifungal activity of different Xenorhabdus and Photorhabdus species against various fungal phytopathogens and identification of the antifungal compounds from X. szentirmaii. Appl. Microbiol. Biotechnol. 2021, 105, 5517–5528. [CrossRef]
- Paul, V.J.; Frautschy, S.; Fenical, W.; Nealson, K.H. Antibiotic in microbial ecology, isolation and structure assignment of several new antibacterial compounds from the insect-symbiotic bacteria Xenorhabdus spp. J. Chem. Ecol. 1981, 589-597.
- Elbrense, H.; Elmasry, A.M.A.; Seleiman, M.F.; Al-Harbi, M.S.; El-Raheem, A.M.A. Can Symbiotic Bacteria (Xenorhabdus and Photorhabdus) Be More Efficient than Their Entomopathogenic Nematodes against Pieris rapae and Pentodon algerinus Larvae?. Biology 2021, 10, 999. [CrossRef]
- Brachmann, A.O.; Bode, H.B. Identification and Bioanalysis of Natural Products from Insect Symbionts and Pathogens. In Yellow Biotechnology I; Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 135, pp. 123–155. [CrossRef]
- Parihar, R.D.; Dhiman, U.; Bhushan, A.; Gupta, P.K.; Gupta, P. Heterorhabditis and Photorhabdus Symbiosis: A Natural Mine of Bioactive Compounds. Front. Microbiol. 2022, 13, 790339. [CrossRef]
- Koilybayeva, M.; Shynykul, Z.; Ustenova, G.; Abzaliyeva, S.; Alimzhanova, M.; Amirkhanova, A.; Turgumbayeva, A.; Mustafina, K.; Yeleken, G.; Raganina, K.; et al. Molecular Characterization of Some Bacillus Species from Vegetables and Evaluation of Their Antimicrobial and Antibiotic Potency. Molecules 2023, 28, 3210. [CrossRef]
- Mollah, M.I.; Kim, Y. Virulent secondary metabolites of entomopathogenic bacteria genera, Xenorhabdus and Photorhabdus, inhibit phospholipase A2 to suppress host insect immunity. BMC Microbiol. 2020, 20, 1–13. [CrossRef]
- Gulsen, S.H.; Tileklioglu, E.; Bode, E.; Cimen, H.; Ertabaklar, H.; Ulug, D.; Ertug, S.; Wenski, S.L.; Touray, M.; Hazir, C.; et al. Antiprotozoal activity of different Xenorhabdus and Photorhabdus bacterial secondary metabolites and identification of bioactive compounds using the easyPACId approach. Sci. Rep. 2022, 12, 10779. [CrossRef]
- Isaacson, P.; Webster, J. Antimicrobial activity of Xenorhabdus sp. RIO (Enterobacteriaceae), symbiont of the entomopathogenic nematode, Steinernema riobrave (Rhabditida: Steinernematidae). J. Invertebr. Pathol. 2002, 79, 146–153. [CrossRef]
- Li, J.; Chen, G.; Webster, J.M.; Czyzewska, E. Antimicrobial Metabolites from a Bacterial Symbiont. J. Nat. Prod. 1995, 58, 1081–1086. [CrossRef]
- Thaler, J.O.; Baghdiguian, S.; Boemare, N.E. Purification and characterization of nematophilicin, phage tail like bacteriocin, from the lysogenic strain FI of Xenorhabdus nematophilus. Appl. Environ. Microbiol. 1995, 61: 2049-2052.
- Muangpat, P.; Yooyangket, T.; Fukruksa, C.; Suwannaroj, M.; Yimthin, T.; Sitthisak, S.; Chantratita, N.; Vitta, A.; Tobias, N.J.; Bode, H.B.; et al. Screening of the Antimicrobial Activity against Drug Resistant Bacteria of Photorhabdus and Xenorhabdus Associated with Entomopathogenic Nematodes from Mae Wong National Park, Thailand. Front. Microbiol. 2017, 8, 1142. [CrossRef]
- Muangpat, P.; Suwannaroj, M.; Yimthin, T.; Fukruksa, C.; Sitthisak, S.; Chantratita, N.; Vitta, A.; Thanwisai, A. Antibacterial activity of Xenorhabdus and Photorhabdus isolated from entomopathogenic nematodes against antibiotic-resistant bacteria. PLOS ONE 2020, 15, e0234129. [CrossRef]
- León-Buitimea, A.; Garza-Cárdenas, C.R.; Garza-Cervantes, J.A.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. The Demand for New Antibiotics: Antimicrobial Peptides, Nanoparticles, and Combinatorial Therapies as Future Strategies in Antibacterial Agent Design. Front. Microbiol. 2020, 11, 1669. [CrossRef]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [CrossRef]
- Hu, K.; Li, J.; Li, B.; Webster, J.M.; Chen, G. A novel antimicrobial epoxide isolated from larval Galleria mellonella infected by the nematode symbiont, Photorhabdus luminescens (Enterobacteriaceae). Bioorganic Med. Chem. 2006, 14, 4677–4681. [CrossRef]
- Booysen, E.; Dicks, L.M.T. Does the Future of Antibiotics Lie in Secondary Metabolites Produced by Xenorhabdus spp.? A Review. Probiotics Antimicrob. Proteins 2020, 12, 1310–1320. [CrossRef]
- Boszormenyi, E.; Ersek, T.; Fodor, A.; Fodor, A.M.; Foldes, L.S.; Hevesi, M.; et al. Isolation and activity of Xenorhabdus antimicrobial compounds against the plant pathogens Erwinia amylovora and Phytophthora nicotianae. J. Appl. Microbiol. 2009, 107: 764-759.
- Furgani, G.; Böszörményi, E.; Fodor, A.; Máthé-Fodor, A.; Forst, S.; Hogan, J.; Katona, Z.; Klein, M.; Stackebrandt, E.; Szentirmai, A.; et al. Xenorhabdus antibiotics: a comparative analysis and potential utility for controlling mastitis caused by bacteria. J. Appl. Microbiol. 2008, 104, 745–758. [CrossRef]
- Seo, S.; Kim, Y. 2011. Development of “Bt-Plus” biopesticide using entomopathogenic bacteria (Xenorhabdus nematophila, Photorhabdus temperata ssp. temperata) metabolites. Korean J. Appl. Entomol. 2011, 50: 171-178.
- Kanost, M.R.; Jiang, H.; Yu, X. Innate immune responses of a lepidopteran insect, Manduca sexta. Immunol. Rev. 2004, 198, 97–105. [CrossRef]
- Orozco, R.A.; Molnár, I.; Bode, H.; Stock, S.P. Bioprospecting for secondary metabolites in the entomopathogenic bacterium Photorhabdus luminescens subsp. sonorensis. J. Invertebr. Pathol. 2016, 141, 45–52. [CrossRef]
- Watson, R.J.; Millichap, P.; Joyce, S.; Reynolds, S.; Clarke, D. The role of iron uptake in pathogenicity and symbiosis in Photorhabdus luminescens TT01. BMC Microbiol. 2010, 10: 177.
- Akhurst, R.J. Morphological and Functional Dimorphism in Xenorhabdus spp., Bacteria Symbiotically Associated with the Insect Pathogenic Nematodes Neoaplectana and Heterorhabditis. J. General Microbiology 1980, 121, 303–309. [CrossRef]
- Tailliez, P.; Pagès, S.; Ginibre, N.; Boemare, N. New insight into diversity in the genus Xenorhabdus, including the description of ten novel species. Int. J. Syst. Evol. Microbiol. 2006, 56, 2805–2818. [CrossRef]
- Chandrakasan, G.; Ayala, M.T.; Trejo, J.F.G.; Marcus, G.; Maruthupandy, M.; Kanisha, C.C.; Murugan, M.; Al-Mekhlafi, F.A.; Wadaan, M.A. Bio controlled efficacy of Bacillus thuringiensis cry protein protection against tomato fruit borer Helicoverpa armigera in a laboratory environment. Physiol. Mol. Plant Pathol. 2022, 119. [CrossRef]
- Forst, S.; Clarke, D. Bacteria-nematode symbiosis. In Entomopathogenic nematology, 2002, pp. 57-77. CABI Publishing, CAB International, Wallingford, UK.
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Gardner, W.A.; Behle, R.W.; Ree, B.; Harris, M.K. Efficacy of Entomopathogenic Fungi in Suppressing Pecan Weevil,Curculio caryae(Coleoptera: Curculionidae), in Commercial Pecan Orchards. Southwest. Èntomol. 2009, 34, 111–120. [CrossRef]
- Cappuccino, J.G.; Sherman, N. Microbiology: a laboratory manual, 2002, pp. 263-264. Benjamin, Harlow.
- Bonev, B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [CrossRef]
Figure 1.
Displays the general view on natural antibacterial crude secondary metabolites extracted from bacterial symbionts and identification of functional bioactive compounds by FTIR and Gas chromatography-mass spectrometry.
Figure 1.
Displays the general view on natural antibacterial crude secondary metabolites extracted from bacterial symbionts and identification of functional bioactive compounds by FTIR and Gas chromatography-mass spectrometry.
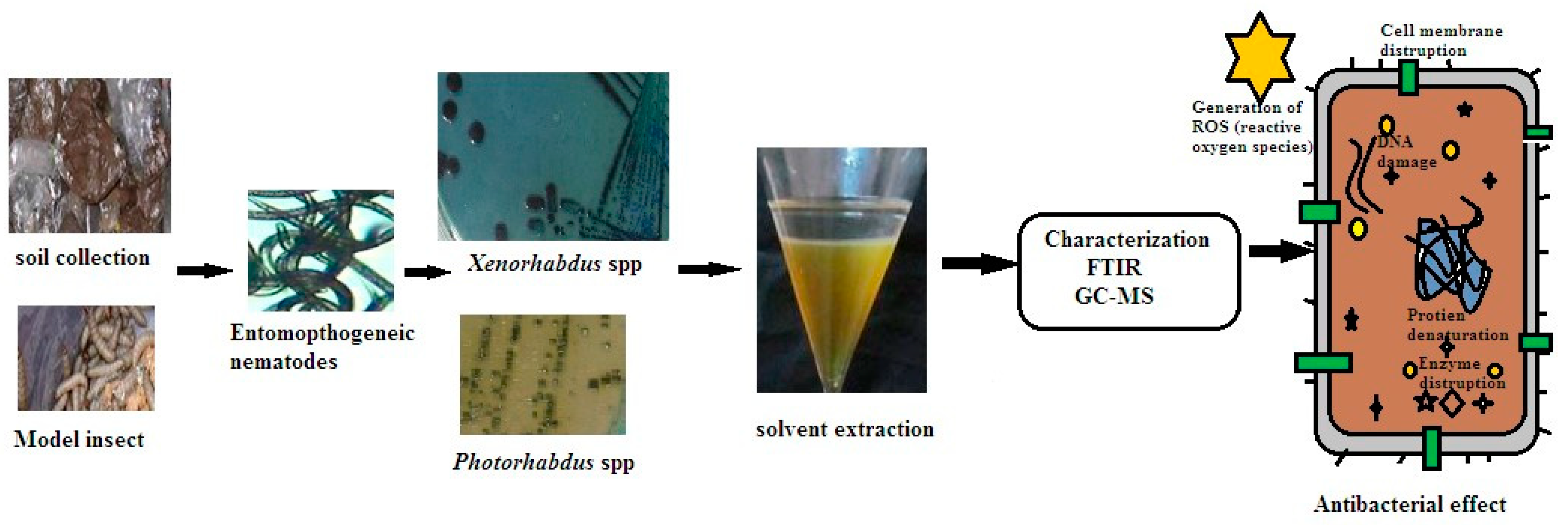
Figure 2.
displays the (A) Recovered soil entomopathogenic nematodes molecular characterization (strains number 05 and 10); (B) Extracted symbiotic bacteria molecular characterization (strains number 05 and 10).
Figure 2.
displays the (A) Recovered soil entomopathogenic nematodes molecular characterization (strains number 05 and 10); (B) Extracted symbiotic bacteria molecular characterization (strains number 05 and 10).
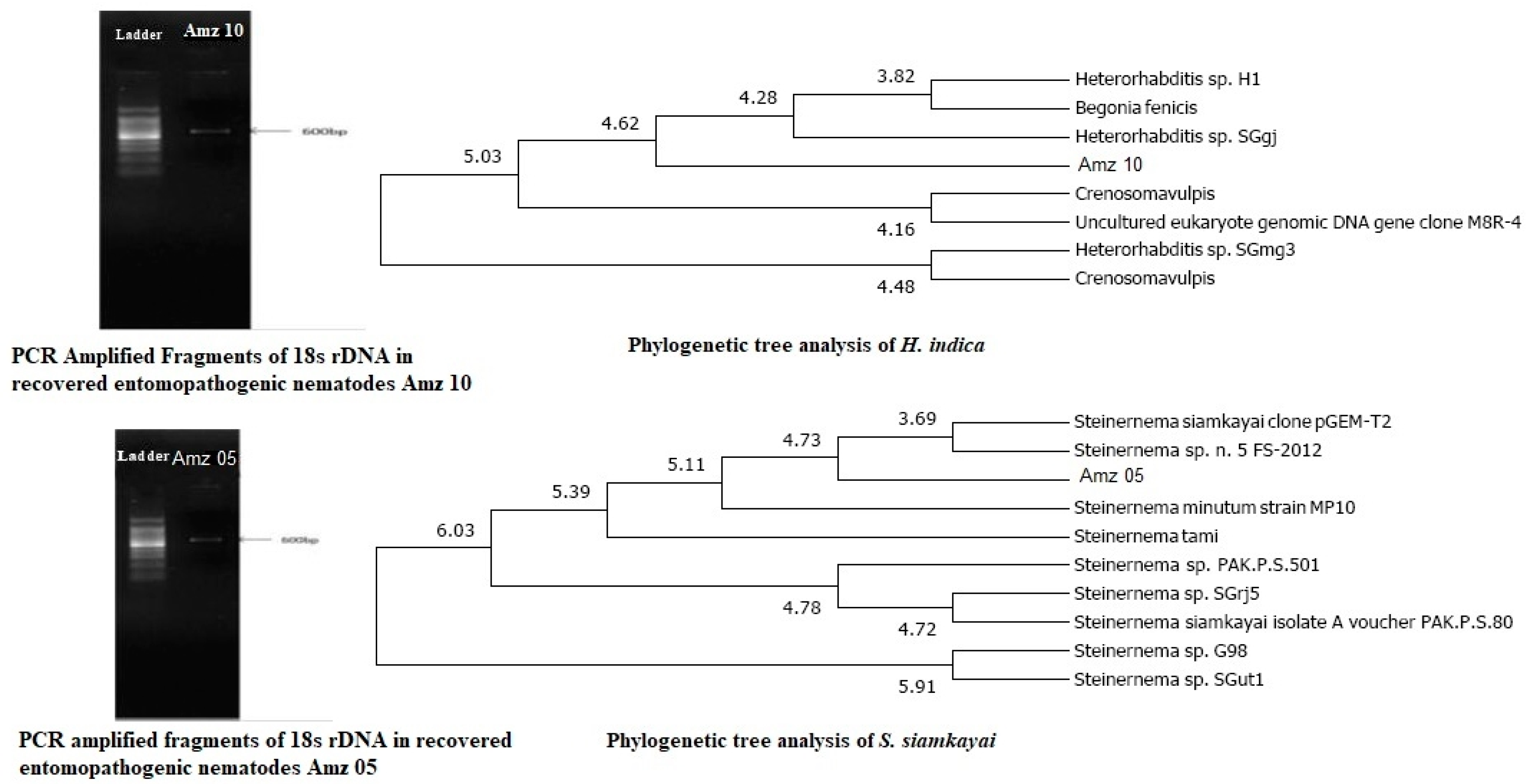
Figure 3.
shows the FT-IR analysis of bacterial crude compounds of Xenorhabdus stockiae (A); Photorhabdus lumniscenes (B).
Figure 3.
shows the FT-IR analysis of bacterial crude compounds of Xenorhabdus stockiae (A); Photorhabdus lumniscenes (B).
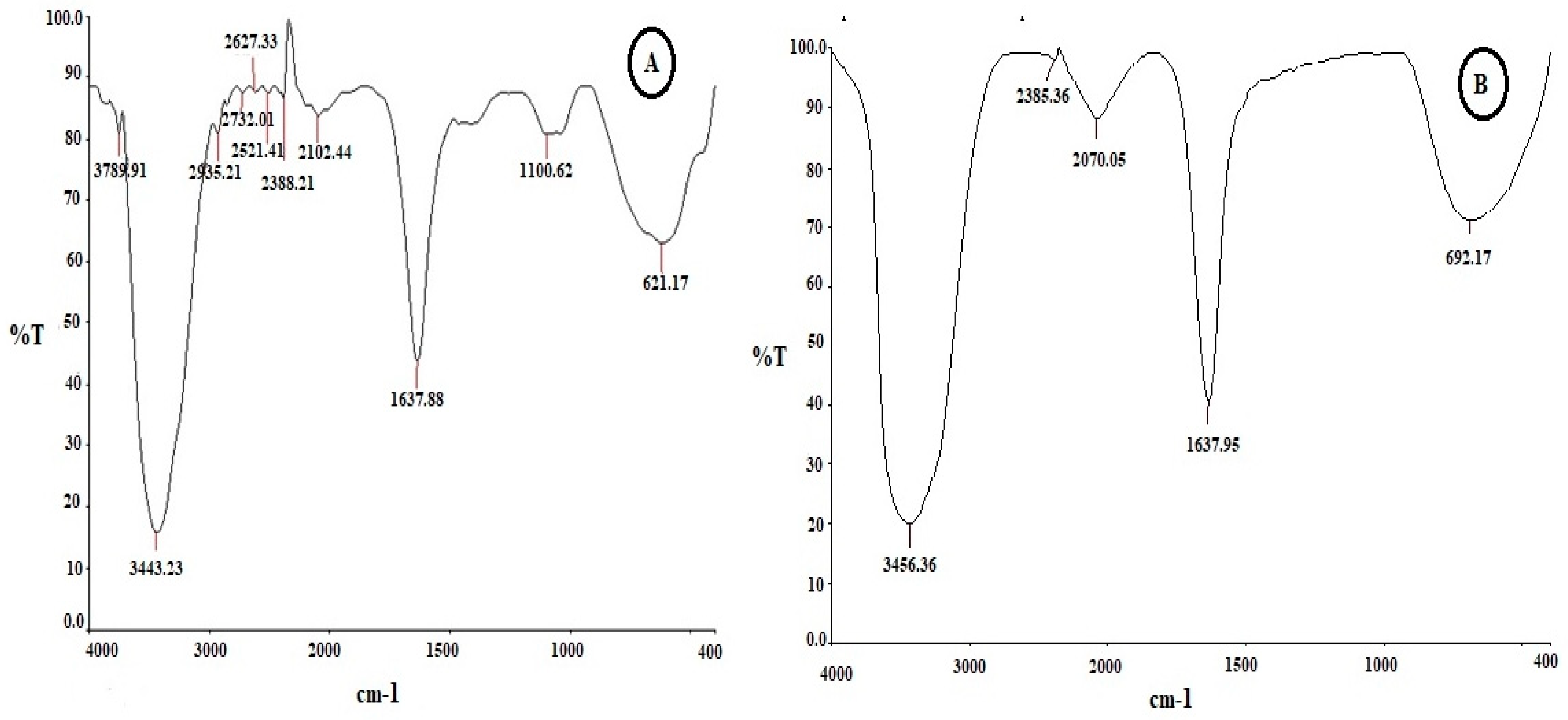
Figure 4.
illustrations of GC-MS chromatogram of Symbiotic bacteria (X. stockiae).
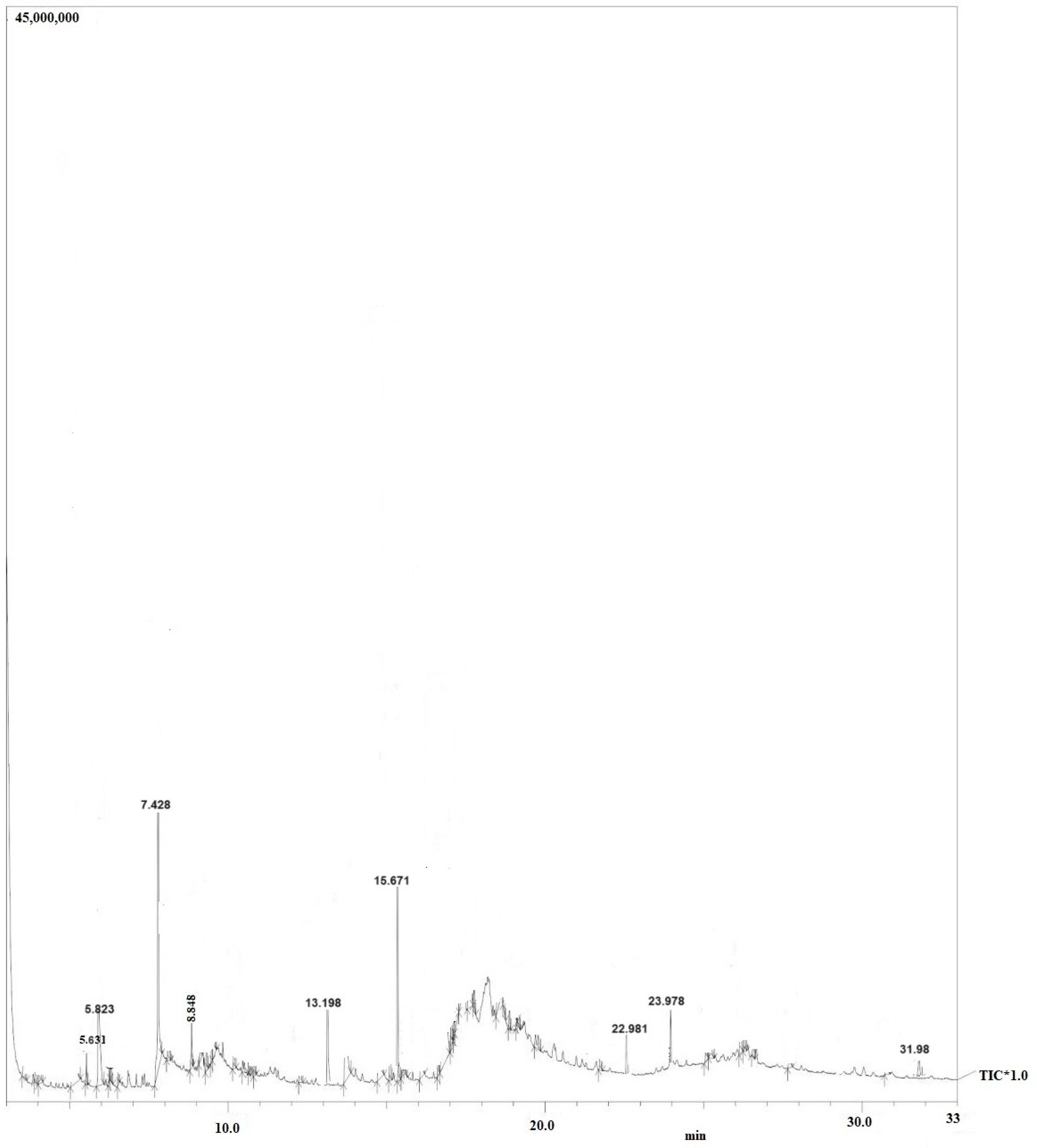
Figure 5.
illustrations of GC-MS chromatogram of Symbiotic bacteria (P. luminescens).
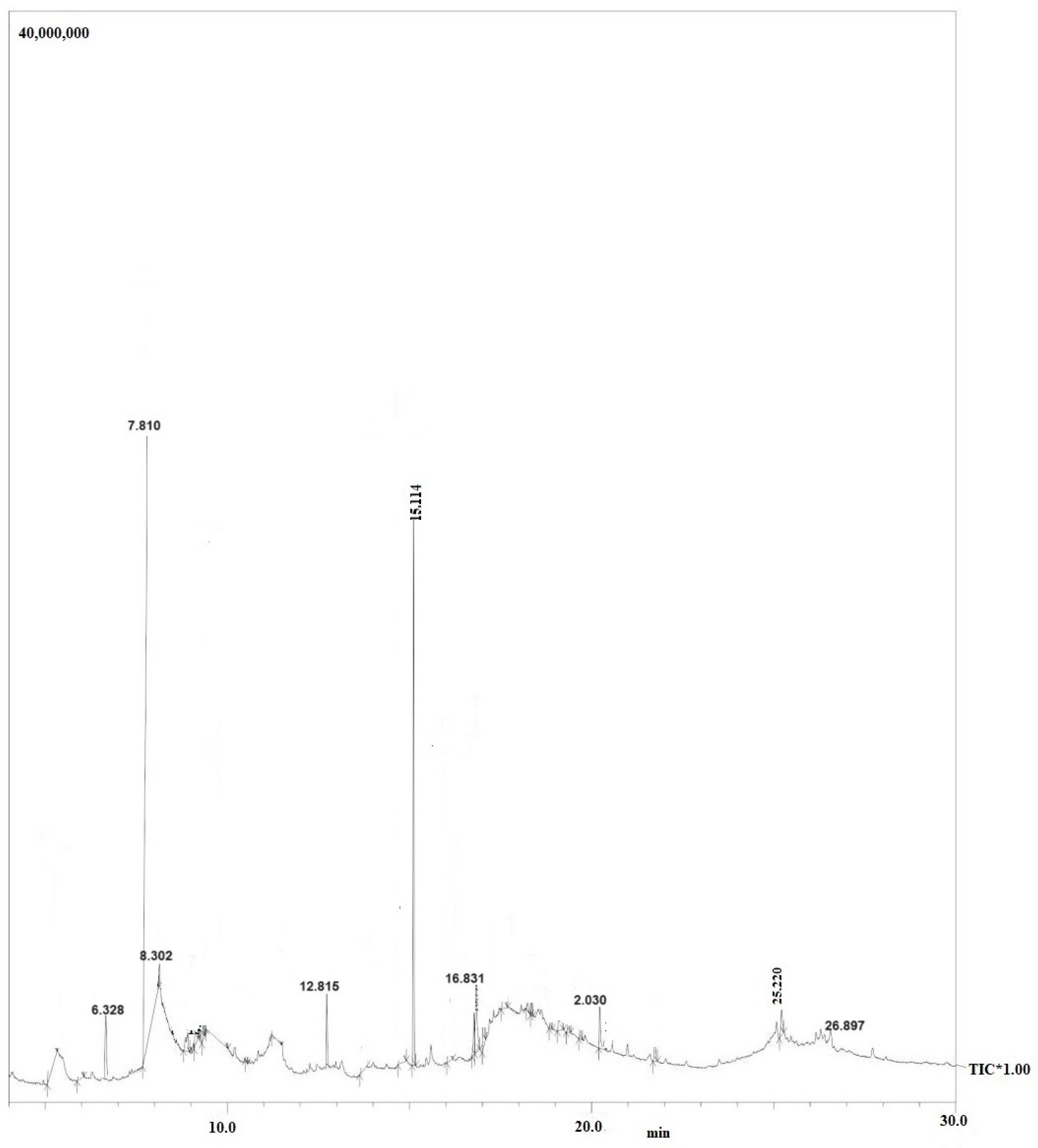
Figure 6.
describes the general mechanisms of antimicrobial action on bioactive functional compounds from symbiotic bacteria.
Figure 6.
describes the general mechanisms of antimicrobial action on bioactive functional compounds from symbiotic bacteria.
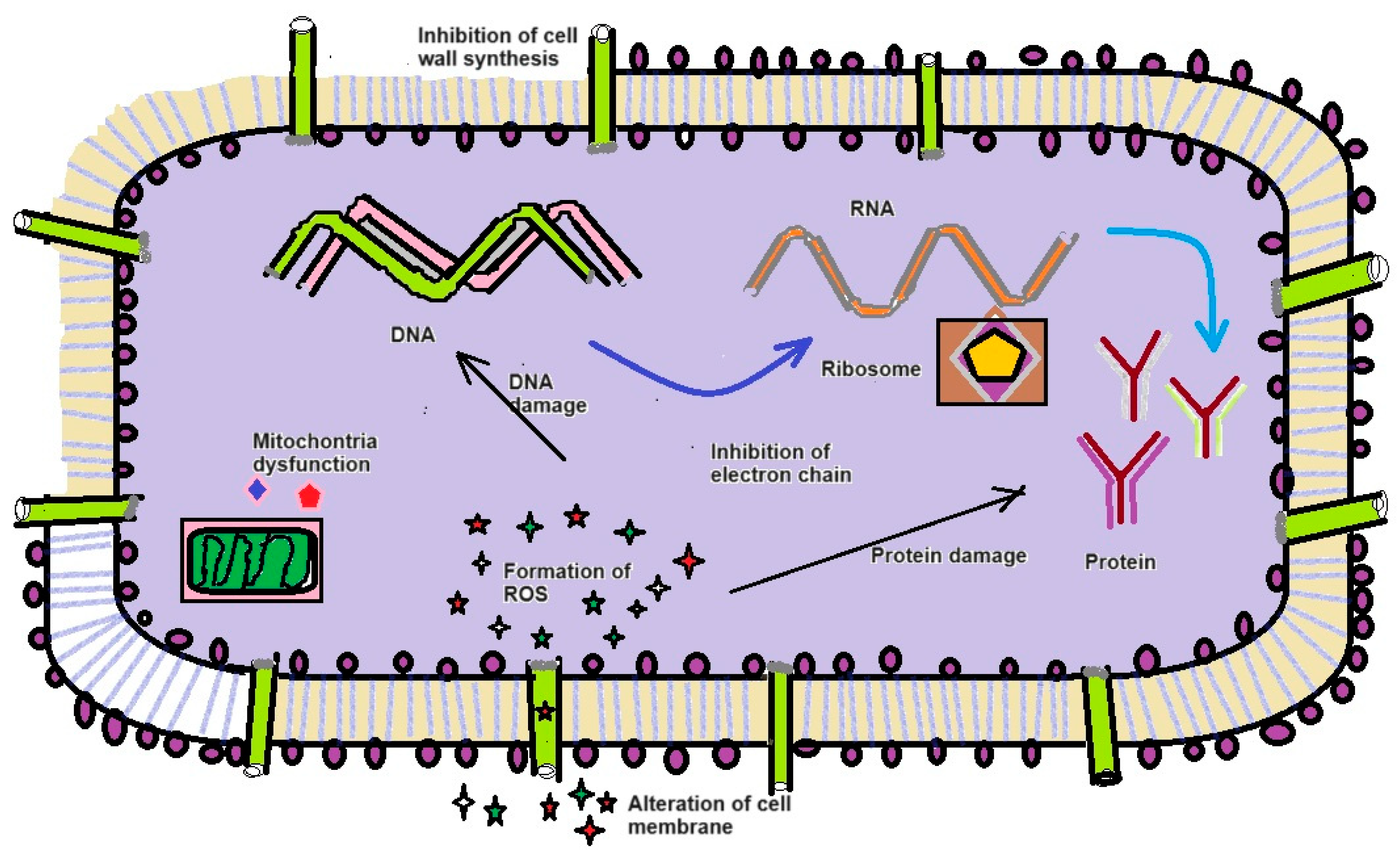
Figure 7.
displays the antibacterial action of symbiotic bacterial (X. stockiae and P. luminescens) crude compound against bacterial pathogen.
Figure 7.
displays the antibacterial action of symbiotic bacterial (X. stockiae and P. luminescens) crude compound against bacterial pathogen.
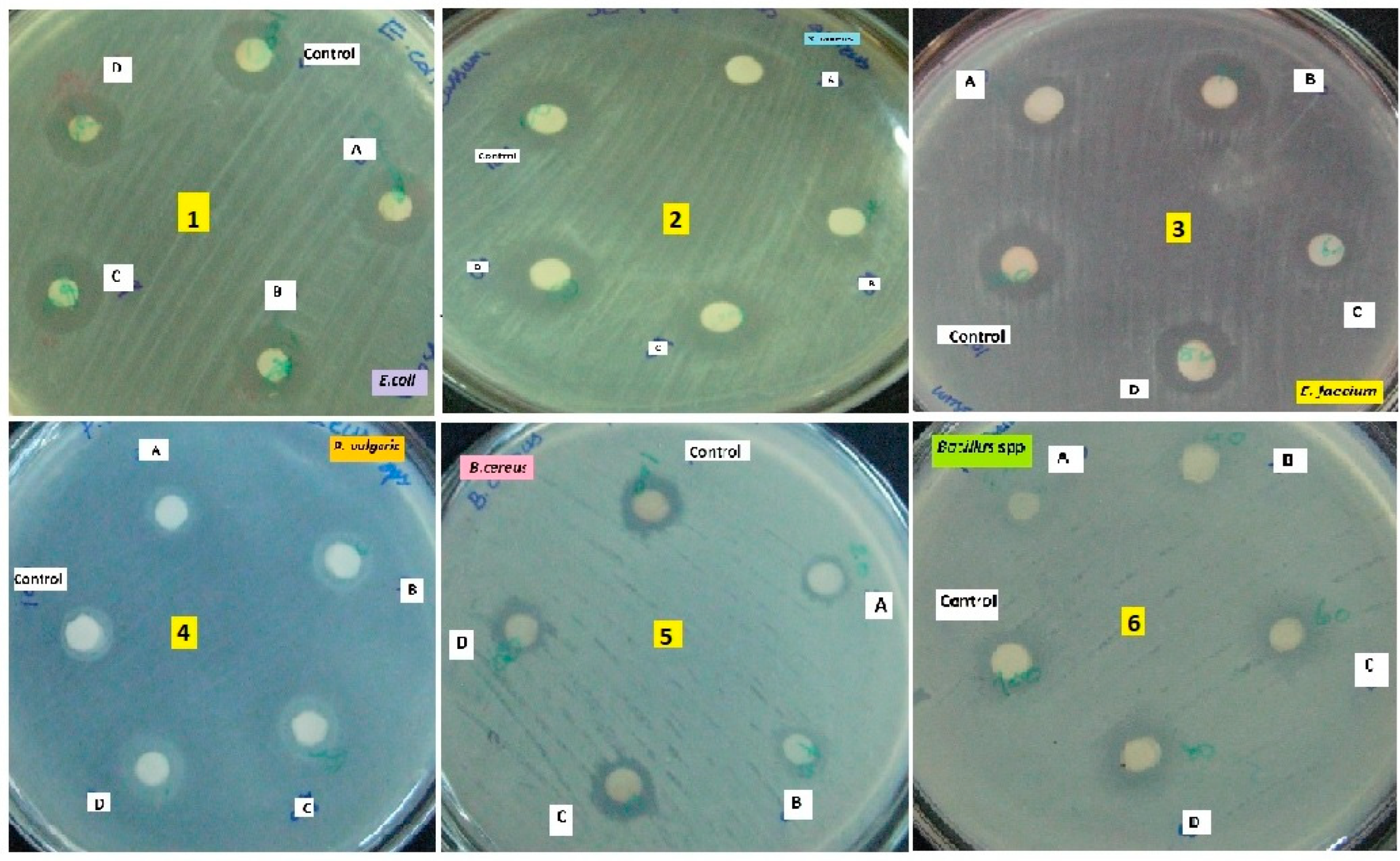
Table 1.
Distribution and extraction of soil entomopathogenic nematodes and symbiotic bacteria using insect (Galleria mellonella) bait method from (agricultural &uncultured land) Querétaro, Mexico.
Table 1.
Distribution and extraction of soil entomopathogenic nematodes and symbiotic bacteria using insect (Galleria mellonella) bait method from (agricultural &uncultured land) Querétaro, Mexico.
| Sampling site | Agriculture crops/ land |
Recovered | Soil Type | Soil Temperature (oC) | Organic content (%) | pH | Electrical conductivity (mS/cm) | Symbiotic bacteria characteristics | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Steinernema spp. | Heterorhabditis spp. | ||||||||||
| Xenorhabdus spp(Amz 05) | Photorhabdus spp(Amz 10) | ||||||||||
|
Zone I (agricultural land) |
Corn | ++ | - | sandy | 29 | 4.9 | 7.25 | 0.93 | Blue color colony | Greenish yellow color coloy | |
| Rice | - | ++ | Loam clay | 31 | 4.8 | 7.13 | 1.29 | Bioluminescence (-)tive |
Bioluminescence (+)tive | ||
| wheat | ++ | - | loam | 30 | 3.9 | 7.40 | 1.32 | Catalase (-) tive | Catalase (+) tive | ||
|
Zone II (uncultured land) |
Forest | + | - | Silt | 28 | 4.9 | 7.70 | 1.74 | Absorption of bromothymol blue | Absorption of bromothymol blue | |
| Grasses | + | - | Silt | 29 | 3.8 | 7.41 | 1.59 | Growth on 28oC | Growth on 28oC | ||
| Waste land | - | Silt | 30 | 4.1 | 7.23 | 1.03 | Insect pathogenicity | Insect pathogenicity | |||
Table 2.
FT-IR spectrum of Xenorhabdus spp. crude compound in possible assigned functional groups.
| S. No. | FT-IR peaks (cm-1) | Possible assigned functional groups |
|---|---|---|
| 1 | 3443 | Amine, N-H Stretch |
| 2 | 1637 | NH amide bend C-Br Bend |
| 3 | 1100 | ANAH and carbonyl (CAOA) |
| 4 | 621 | Carbonyl stretching (OC) |
Table 3.
FT-IR spectrum of Photorhabdus spp. crude compound in possible assigned functional groups.
| S. No. | FT-IR peaks (cm-1) | Possible assigned functional groups |
|---|---|---|
| 1 | 3436 | Amine, N-H Stretch |
| 2 | 2078 | C≡N, Nitriles |
| 3 | 1078 | Carbonyl stretch |
| 4 | 692 | Alkyl Halides |
Table 6.
Antibacterial activity of symbiotic bacterial (X. stockiae and P. luminescens) crude extracts against bacterial pathogens.
Table 6.
Antibacterial activity of symbiotic bacterial (X. stockiae and P. luminescens) crude extracts against bacterial pathogens.
| Tested organisms | Zone of Inhibition (cm) | ||||
|---|---|---|---|---|---|
| X. stockiae | P. luminescens | Positive control | |||
| A | B | C | D | ||
| 1. Escherichia coli | 2.6 ± 0.7 | 2.9± 0.7 | 2.1 ±0.8 | 2.4±0.8 | 3.1 ± 0.7 |
| 2.Staphylococcus aureus | 2.5 ± 0.6 | 2.8 ± 0.6 | 1.9 ± 0.6 | 2.0± 0.6 | 2.9 ± 0.6 |
| 3. Enterococcus faceium | 2.2 ± 0.2 | 2.5 ± 0.2 | 1.8 ± 0.4 | 2.2 ± 0.4 | 2.7 ± 0.2 |
| 4. Proteus vulgaris | 2.1 ± 0.3 | 2.4 ± 0.3 | 1.7 ± 0.8 | 2.0 ± 0.8 | 2.7 ± 0.3 |
| 5. Bacillus cereus | 2.0± 0.9 | 2.3± 0.9 | 1.6 ± 0.4 | 1.9 ± 0.4 | 2.6± 0.9 |
| 6. Bacillus subtilis | 1.9± 0.9 | 2.1± 0.9 | 1.5 ± 0.8 | 1.8 ± 0.8 | 2.6± 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



