
Submitted:
02 May 2024
Posted:
03 May 2024
You are already at the latest version
Abstract
The aim of this study was to evaluate whether alterations in food availability compromise the metabolic homeostasis of honey bees exposed to three fungicides alone or together. Ten honey bee colonies were utilized, with half receiving energy-protein supplementation for 15 weeks while another five colonies had their protein supply reduced with pollen traps. Subsequently, forager bees were collected and exposed by contact to 1 or 7 µg of bixafen, prothioconazole, and trifloxystrobin, either individually or in combination. After 48 hours, abdomens without the intestine was used for the analysis of expression of antioxidant genes (SOD-1, CAT, and GPX-1), detoxification genes (GST-1 and CYP306A1), the storage protein gene vitellogenin, and immune system antimicrobial peptide genes (Defensin-1, Abaecin, Hymenoptaecin, and Apidaecin), through real-time PCR. All pesticide treatments induced changes in gene expression, with bixafen showing the most prominent upregulation. Exposure to 1 µg of each of the three pesticides resulted in upregulation of genes associated with detoxification and nutrition processes, and downregulation of immune system genes. When the three pesticides were combined at a dose of 7 µg each, there was a pronounced downregulation of all genes. Dietary supplementation led to a slight reduction in gene expression alterations provoked by the fungicides.
Keywords:
contact exposure
; detoxification
; fungicide
; immunity
; nutrition
; RT-PCR
1. Introduction
Bees are susceptible to numerous environmental stressors, including pesticides; among these there is little or no use restriction for fungicides in many countries (National Honey Bee Health Stakeholder, 2012; Fisher et al., 2023). This is of concern because fungicides account for approximately 35% of the global pesticide market (Rondeau and Raine, 2022). The widespread use of fungicides to control and prevent damage due to pathogenic fungi often results in the development of fungicide-resistant strains, necessitating the need for new disease control strategies. An alternative to help overcome resistance has been to apply fungicide mixtures to combat one or more pathogens at various stages of the life cycle (Deising et al., 2008).
Lethal effects for bees resulting from exposure to fungicides are typically observed through exposure to relatively high doses (>100 μg/bee) (Biddinger et al., 2013; Azpiazu et al., 2021; Iwasaki and Hogendoorn, 2021). However, the actual impact on bee health needs to be evaluated using environmentally relevant doses of fungicides. Data on these doses can be difficult to obtain, especially for new products, for which there is scarce information concerning contamination levels on treated crops. Determining impact is more complicated when multiple active ingredients (a.i.) with fungicidal action are used in the same commercial product. These mixtures can lead to synergistic effects, which may not be observed when they are evaluated individually (Zubrod et al., 2019; Sgolastra et al., 2020; Schuhmann et al., 2022).
A commercial product that contains the active ingredients bixafen (bix), prothioconazole (pro), and trifloxystrobin (tri) is recommended for controlling fungal diseases in various major crops such as cotton, corn, sunflower, and soybean, with no restrictions on applications during the reproductive period of these plants (Bayer, 2019). Since these crops are visited by bees for pollen and nectar, there may be exposure of these non-target organisms to the fungicidal active ingredients by contact or ingestion of contaminated food (Giannini et al., 2015; Omar et al., 2022). In Brazil, in 2022, 1,981 t of bix, 5,193 t of pro and 5,195 t of tri were commercialized in various commercial fungicides containing one or more a.i. (IBAMA, 2023).
Bix is a fungicide belonging to the chemical group of carboxamides; it interferes with the mitochondrial respiratory chain by inhibiting the enzyme succinate dehydrogenase which is responsible for electron transfer in complex II. Consequently, with less energy available, vital fungal functions are impaired, impacting reproduction and potentially leading to mortality (Picard et al., 2011; Oliver and Hewitt, 2014). There are few studies addressing the effects of bix on pollinators. Another fungicide from the same carboxamide chemical group, boscalid, is more widespread, and known negative impacts on bee health due to exposure to this fungicide include reduced longevity (Simon-Delso et al., 2018) and decreased wing beat frequency, rendering bees lethargic (Liao et al., 2019).
The fungicide tri is a member of the strobilurin chemical group, which inhibit mitochondrial respiration at complex III of the respiratory chain. Consequently, the energy cycle within the fungus is interrupted, halting ATP production (Bartlett et al., 2002). Exposure of the Brazilian stingless bees with the common name Mandaçaia (Melipona quadrifasciata) to tri combined with the fungicide tebuconazole resulted in increased mortality; additionally, bees were repelled from sprayed tomato flowers. With fewer bees and reduced flower visits, there was a negative impact on fruit production (Silva-Neto et al., 2018).
The fungicide pro belongs to the triazolintione group and interferes with sterol biosynthesis. When pro binds to the enzyme sterol C14-demethylase, it blocks the sterol biosynthesis pathway, resulting in inhibition of production of phospholipids, accumulation of free fatty acids, and fungal death (Oliver and Hewitt, 2014). This a.i. is marketed in over 60 countries and is the third best-selling fungicide worldwide (Fang et al., 2023). There is disagreement regarding the level of toxicity of this fungicide for honey bees (Wood et al., 2020; Haas et al., 2021). However, triazole fungicides, which have the same mode of action as the triazolintione group, can affect bee behavior and susceptibility to diseases (Pettis et al., 2013; Fisher et al., 2017).
Another factor inherent to conventional agriculture that can be detrimental to bees is decreased natural food resources. Large-scale monocultures, coupled with deforestation of native vegetation, result in reduced availability of pollen and nectar (Vaudo et al., 2015). Though some crops provide pollen, the amino acid composition of pollen from a single species may not meet the nutritional demands of the bees (Behmer, 2009; Brodschneider and Crailsheim, 2010), leading to malnutrition and increased susceptibility to environmental stressors (Tadei et al., 2020; Quinlan and Grozinger, 2023).
Our objective was to determine how individual and mixed fungicide ingredients affect the expression of genes related to honey bee health and whether nutritional condition of the colonies they came from affected gene function. We treated forager bees from well fed and from nutritionally deprived colonies with the fungicides bix, pro, and tri and measured the expression of antioxidant, detoxification, storage protein, and antimicrobial protein genes.
2. Material and Methods
Location of the Experiment and Preparation of the Honey Bee Colonies
The study was conducted in the municipality of Dracena, state of São Paulo, at a latitude of 21°27’37” South and longitude of 51°33’21” West, and altitude 392 meters. Ten colonies of Africanized honey bees (Apis mellifera), headed by open mated queens, and kept in 10 frame Langstroth hives were used. They were divided into two groups of five hives: one group with supplemented feeding (SF) and the other with reduced food (RF).
Over a period of 15 weeks, the SF colonies were able to store all the food they collected in the field and were supplemented with 500 ml of energetic syrup containing water and crystal cane sugar in equal proportions twice a week, as well as 100g of protein paste composed of two parts bee pollen and one part of wild honey, provided once a week. Both food sources were obtained from the same apiary where the experiment was conducted. For the other group (RF), during the same period, there was no food supplementation, and pollen traps were installed at the hive entrances to reduce the availability of protein for the bees in this group.
Bee pollen Collection from Colonies with Reduced Feeding
In the RF group, bee pollen was collected daily and weighed using a semi-analytical balance, recording the quantities obtained weekly for each colony. To determine the efficiency of the pollen collectors (%), the entrance of forager bees through the pollen traps was observed during 30-minute intervals between 8 and 10 AM, with three repetitions per colony. The number of bees entering with pollen and the number of pollen pellets falling into the pollen trap drawer were recorded. The efficiency of the traps was calculated as the ratio of the number of pollen pellets to the number of bees entering the collectors with pollen, multiplied by two.
Exposure of Forager Bees to Isolated Fungicides or in Combination
Forager bees were collected from the hive entrances after 15 weeks of feeding management. The bees were placed in 0.25 L clear round plastic containers with perforated lids. In the laboratory, the bees were anesthetized in a freezer (-20ºC) for several minutes.
The immobilized bees were exposed by contact to bix, pro, and tri, by applying 0.5 µL of a acetone solution containing 1 or 7 µg of one or all three active ingredients in combination, using a micropipette, on the dorsal thorax of the bees (OECD, 1998). The doses were determined based on a study estimating the dose at which a bee could be exposed by contact when applying a fungicide containing the three active ingredients according to agronomic recommendations (Gomes et al., 2023). The acetone solvent alone was applied for the control. Acetone and the fungicide a.i. were purchased from Sigma-Aldrich.
Following contact exposure, 20 forager bees from colonies that received the same feeding management and exposure dose to the fungicides were placed in new plastic containers and fed ad libitum with syrup containing two parts sucrose and one part water, remaining for a period of 48 hours in an incubator with controlled temperature (33º±1ºC) and humidity (70±10%).
Gene expression Assessment
After 48 hours in the incubator, the bees were again anesthetized in a freezer for dissection, which was carried out with the aid of a stereo binocular microscope, fine tip scissors, and entomological forceps. We discarded heads, thoraxes, and intestines, leaving only the abdomens. Each sample consisted of six abdomens, resulting in four samples per treatment (two colony feeding regimes x four fungicide solutions x two doses and controls).
RNA Extraction and cDNA Library Construction
The transcriptome analysis was performed by Real-Time qPCR, using the primers described in Table 1.
For RNA extraction, 1 mL of Trizol® LS Reagent (Invitrogen™) and 200 µL of chloroform were used for each sample with six dissected abdomens (approximately 100 mg). The tubes were centrifuged at 12,000 x g for 15 minutes at 4ºC. The upper aqueous phase was transferred to another microtube, to which isopropanol was added. The samples were vortexed and centrifuged at 12,000 x g for 10 minutes at 4ºC. Finally, the supernatants were discarded, RNA pellets were washed three times with 100 µL of ethanol, centrifuged, supernatant discarded, and pellets were air-dried for 15 minutes. Subsequently, they were suspended in molecular grade water (Sigma).
After RNA isolation, quantification was made in ng by absorbance at 260 nm using a Nanodrop-1000 (Thermo Scientific). An aliquot of RNA was processed in a 1% agarose gel and post-stained with GelRed® (Sigma) to verify RNA integrity. The cDNA library was constructed from the extracted RNA by reverse transcription using 1 μg of total RNA and iScriptTM III (Life Technologies). The resulting cDNA was stored at -20°C until use. Subsequently, the sequence was amplified by polymerase chain reaction (PCR) using target primers.
Molecular Determination of Gene Expression
The molecular determination of gene expression was carried out using the real-time polymerase chain reaction (qPCR-RT) method, and the results were analyzed by relative quantification using the 2−ΔΔCt method (Pfaffl et al., 2004; Rebouças et al., 2013). qPCR reactions were performed in 96-well plates using iTaqTM Universal SYBR® Green Supermix (Bio-Rad). Reactions were conducted following the manufacturer’s instructions, with 20 µL individual reaction mixtures consisting of 10 µL of SYBRGreen PCR Master Mix (2x), 8 µL of 200 nM primers, and 2 µL of cDNA template (samples) (Rebouças et al., 2013).
All reactions were performed using the iCycler iQ™ real-time qPCR detection system (Bio-Rad Laboratories) under the following protocol: 95°C for 5 min, 40 cycles of 95°C for 30 s, 56°C for 30 s, followed by analysis of the dissociation curve (melting curve) to verify amplification of a single product. The denaturation-hybridization-synthesis cycle temperatures used were according to Pfaffl et al. (2004).
The threshold of the exponential phase, denoted as the Cycle Threshold (Ct), was detected during the temperature cycles, precisely quantifying the products of the amplification reaction by fluorescence emission. Expression data was used to calculate the Ct values. PCR efficiency and the relative expression ratio of target genes in experimental groups versus control groups were calculated according to the method of Pfaffl et al. (2004). The comparative Ct method (2−ΔΔCt method) was used to analyze the expression level of these genes relative to the RPL32 gene (Lourenço et al., 2008).
Statistical Analyses
The mean amount of bee pollen collected weekly from each colony was calculated for the RF colonies. The efficiency of the pollen traps was evaluated by the mean of 12 observations across four hives. Data dispersion was determined by calculating the standard error.
The expression of each gene was assessed by comparing the results of each treatment with the control of the SF group. Since the data did not follow a normal distribution, comparisons between medians were made using the non-parametric Kruskal-Wallis test (SAS Institute, 2013).
3. Results
Feeding Management of Colonies
During the 15-week feeding management period, each of the five colonies with food supplementation fully consumed the 100g of protein supplement and the 1000ml of sucrose syrup offered weekly. One colony from the RF group succumbed at the 11th week and was not replaced. Therefore, to maintain the same number of colonies among the groups, one colony from the SF group was randomly selected and removed from the experiment.
In the RF group, 33.8±4.9g of bee pollen was collected weekly per colony (Figure 1). By observing foraging bees with pollen in their corbiculae through the pollen traps, it was estimated that the efficiency of these collectors was 57.9±5.1%. Thus, of all the pollen collected by the bees from colonies with reduced feeding, only 42.1% remained available for the bees.
We observed a decrease in activation of all the genes when bees were exposed to 7 µg of each of the three a.i. in combination, for bees from both feeding management groups (Figure 2). With the lowest dose of each a.i. in combination (1 µg), upregulation of the genes was observed, except for those involved in the immune system, which were downregulated. When comparing the controls of bee samples from SF and RF colonies, no difference in the expression of any of the genes was observed (Figure 2).
The expression of SOD-1 was higher with both doses of bix and when the lowest dose of each of the three a.i. was used in combination, in bees from both feeding management groups. This gene was downregulated only in bees from the RF group, in individuals exposed by contact with both doses of pro and with the highest dose of tri (Figure 2).
For catalase, an increase in gene expression was observed with the use of both doses of bix, the highest dose of pro, and with the lowest dose of the three a.i. applied together in bees from both feeding management groups. Catalase was downregulated in bees treated with the highest dose of tri (Figure 2).
GPX-1 expression was increased by bix alone at both doses, with the highest dose of pro, and with the lowest dose in the combination of all three a.i., without influence from colony feeding management. Tri at a dose of 1µg promoted higher expression of this gene in bees from the RF group (Figure 2).
GST-1 was upregulated with the use of 1 or 7 µl of bix or bix + pro + tri at the lowest dose in bees from the SF and RF groups. Exclusive exposure to pro resulted in upregulation in all samples, except when the lower dose was used in bees from the SF group. Tri caused downregulation with both doses for bees from both feeding management groups, except for bees from the SF group exposed to the higher dose of this a.i. (Figure 2).
Expression of CYP306A1 was upregulated with all a.i., except for pro and tri at lower doses, in bees from the SF and RF groups, respectively. The vitellogenin gene had its expression increased when bees were exposed to one or all three active ingredients, except for the use of pro at the lowest dose, for bees from both feeding management groups, and tri at the highest dose, for bees from the RF group (Figure 2).
Defensin-1 was upregulated by exposure to bix, except for bees from the SF group exposed with the lower dose of this a.i. With the other active ingredients, either isolated or in combination, there was downregulation of this gene. All three a.i. caused downregulation of abaecin, except for pro and pri at the lowest dose, for bees from the SF group (Figure 2).
Hymenoptaecin was upregulated when bees were contaminated with bix, except for bees from the RF group that received the lowest dose of this a.i. With isolated use of the other two a.i., and the combination of the three a.i. at both doses, hymenoptaecin was downregulated in bees from both feeding management groups (Figure 2).
Apidaecin expression increased when bees from the RF group were exposed to the two doses of bix and the highest dose of tri. Bees from the SF group had increased expression of apidaecin when exposed to the highest dose of bix or tri. For the remaining comparisons, apidaecin was downregulated in bees from both feeding management groups, except for pro at the highest dose, which did not differ from the SF control (Figure 2).
4. Discussion
Colony Feeding Management
The use of pollen traps reduced the availability of this protein food for the bees. The mean total amount of bee-collected pollen per colony during the 15 weeks was 507g, which was equivalent to 57.9% of the total bee-collected pollen brought into the hives by the bees of this group. This quantity is relatively small considering that up to 20 kg per hive per year can be obtained (Campos et al., 2010). Maintaining pollen traps in hives for several weeks may result in a reduction in the quantities of collected pollen due to the population adjustment made by the bees (Mattos et al., 2015). However, in this study, the amount of bee-collected pollen harvested was relatively small from the early days of food restriction, indicating low availability of pollen in the field (Vaudo et al., 2015).
Unlike other livestock, beekeeping does not require routine feeding of colonies, except during periods of heavy rains, severe winters, and scarcity of nectar and pollen in the field (Somerville, 2005). Our study was conducted in an experimental apiary used for several years, with a known food flow, which is characterized by low availability of nectar and pollen in most months.
Among the five colonies subjected to food restriction management, one succumbed. The others survived, though with visibly reduced populations, relying on the nectar and pollen foragers managed to bring into the colonies, passing through the pollen traps. For the other group (SF), providing protein paste made of honey and bee-collected pollen, which are foods that bees naturally consume, may have contributed to the acceptance of this paste, which was completely consumed by the bees. Adequate diets can contribute to immunocompetence and resistance to pathogens (Alaux et al., 2010; Di Pasquale et al., 2013) and pesticide tolerance (Schmehl et al., 2014). Bees from the SF group consumed all the energy syrup offered twice a week. This nectar substitute, in association with the protein supplement, may promote the search for more protein food in the field, increase queen egg laying, and worker hygienic behavior (Somerville, 2005).
Gene Expression Changes in Bees from Colonies with Supplemented or Reduced Feeding
According to the safety data sheet of the commercial fungicide containing the active ingredients bix, pro, and tri (Bayer, 2022), the contact LD50 for honey bees is greater than 200 µg per individual. In our study, for treatments with the mixture of all three a.i., the doses used (1 or 7 µg per individual of each a.i.) can be considered sublethal, as they are approximately 10 and four times lower than the lethal dose reported by the manufacturer, respectively.
Christen et al. (2019) evaluated the toxic effect of azoxystrobin, a fungicide belonging to the strobilurin group, which includes tri. They observed downregulation of genes encoding enzymes involved in metabolism, oxidative phosphorylation, and hormonal regulation, which could affect energy production, ontogeny, and behavior of bees. Tri had already been detected in bee-collected pollen obtained from cultivated plants and in bee-collected pollen from nearby wild plants (David et al., 2016). Bix was detected in samples of bee bread (Bokšová et al., 2021), and pro has been detected in bee-collected pollen (Roszko et al., 2016; Prado et al., 2019; Raimets et al., 2020). Information regarding bee exposure through contact is limited.
Studying gene expression in bees can aid in understanding the effects of exposure to stressors such as pesticides. In our study, genes related to oxidative stress and detoxification (SOD-1, CAT, GPX-1, GST-1, and CYP306A1), nutrition and longevity (Vitellogenin), and immunity (Defensin-1, Abaecin, Hymenoptaecin, and Apidaecin) were chosen. The metabolism of bees, like other aerobic species, involves the formation of free radicals. When these molecules are not properly processed to favor cellular homeostasis, oxidative stress can occur (Weirich et al., 2002).
Regulation and inactivation of free radicals are carried out by the antioxidant system. This process is natural due to the metabolism of oxygen and other substances metabolized by bees; however, biotic and abiotic stressors can disrupt cellular homeostasis, generating more reactive oxygen species than the individual can neutralize and eliminate (Yan et al., 2012; Olgun et al., 2020).
Sublethal doses of insecticides such as organochlorines and organophosphates (Rehman and Waliullah, 2012; Chakrabarti et al., 2015) and neonicotinoids (Balieira et al., 2018) may lead to increased production of antioxidant enzymes such as SOD-1, CAT, and GPX, aiming to maintain cellular homeostasis. However, higher pesticide doses and specific characteristics of the xenobiotic can impair the production of these enzymes (Bus, 2017). Bix and tri are inhibitors of the respiratory chain, and their action may be unfavorable for the production of antioxidant enzymes (Casida and Durkin, 2016; D’Hose et al., 2021).
In our study, both doses of bix promoted upregulation of detoxification genes including Vg, in bees from both feeding management groups. Gene expression of SOD-1 was downregulated and remained similar to the control, respectively, for bees from the RF and SF groups. The expression of CAT and GPX-1 was similar for bees regardless of feeding management, being altered only with the highest doses of the active ingredients, through upregulation. Expression of GST-1 was upregulated when bees were contaminated with both doses of Pro, except in bees from the CAR group with the lower dose. SOD-1 and CAT did not have their expressions altered by Tri, except with the highest dose in bees from the RF group. This active ingredient promoted an increase in the expression of GPX-1 when bees from the RF group were contaminated with the lower dose. The expression of GST-1 was altered in all treatments with tri, in bees from colonies of both feeding management groups.
The gene CYP306A1 belongs to the cytochrome P450 family, which is involved in various cellular biosynthesis and detoxification processes, which are particularly important when there is exposure to xenobiotics. Upregulation of CYP6 genes is associated with resistance to pyrethroids and neonicotinoids in other insects. It is suggested that these genes may be useful for bees to positively respond to detoxification in situations of pesticide contamination (Claudianos et al., 2006). In our study, expression of the CYP306A1 gene was upregulated with all active ingredients used individually, except for pro and tri at the lowest dose, for bees from the SF and RF groups, respectively.
Vitellogenin is a storage protein, and its expression is linked to the nutritional stage of the individual. It can be considered a pleiotropic gene because, in addition to assisting in lipid transport, it is related to longevity, immunomodulation, and regulation of oxidative stress (Salmela et al., 2022). In our study, exposure to one of the three a.i. promoted upregulation of Vitellogenin, except for pro at the lowest dose and tri at the highest dose, for bees from the RF group.
Regarding the genes strictly linked to the bees immune system, there was evident alteration in gene expression. For Defensin-1, downregulation only did not occur with the use of bix, which resulted in upregulation of this gene. For abaecin, downregulation was only not observed with the use of pro and tri at lower doses in bees from the SF group, which did not differ from the SF control. For hymenoptaecin, downregulation occurred with all a.i., except bix, which resulted in upregulation, except with the lower dose, for bees from the RF group, which did not differ from the SF control. For Apidaecin, there was only no alteration in the expression of this gene with the isolated use of pro at the lowest dose.
Gene upregulation aims to rebalance cellular functioning, in order to eliminate substances or microorganisms that may disrupt homeostasis. In our study, contact exposure of bees with all three a.i. together resulted in different responses depending on the doses. With the use of 1 µg of each a.i. per bee, we observed upregulation of all genes related to the immune system, except for Abaecin; however, with the higher dose (7 µg of each active ingredient per bee), there was marked downregulation of all genes.
Gene expression is a process that occurs at the expense of energy produced in cells. With the need for increased expression of a certain gene for the cell to continue functioning in homeostasis, more energy is demanded (Frumkin et al., 2017). However, even with a higher demand for gene expression so that the harmful effects of xenobiotics do not impair cellular functioning, energy availability can be a determining factor in the response to intoxication challenge. Of the three a.i. evaluated in this study, bix and tri are known to act as inhibitors of cellular energy production (Casida and Durkin, 2016; Nicodemo et al., 2020; D’Hose et al., 2021). With the exposure of bees to these a.i. at the highest evaluated dose, cellular energy demand may not have been met, and thus, gene expression was impaired. Thus, bees may become more vulnerable to other stressors, given the fragility resulting from exposure to the fungicide with all three a.i.
The response to the challenge with bix, pro, and tri, whether isolated or in combination, was similar among bees that had supplemented or restricted feeding for most comparisons. Gene expression of bees from the SF group did not differ from the SF control group for SOD-1, with the use of pro at both doses, Catalase (tri at the highest dose), GPX-1 (tri at the lowest dose), GST-1 (pro at the lowest dose), and CYP306A1 (pro at the lowest dose). For the same comparisons, bees from the RF group had gene expression different from the SF control.
5. Conclusions
Bix, pro, and tri induced gene expression changes in honey bees. Contact exposure to all three a.i. together, with 1 µg per bee, resulted in upregulation of the genes, except for Defensin-1, Abaecin, Hymenoptaecin, and Apidaecin. The use of 7 µg per bee of all three a.i. together resulted in significant downregulation of all the genes, without attenuation of effects due to colony nutritional supplementation. Bees from colonies with restricted feeding had more gene expression alterations than bees from colonies with supplemented feeding.
Author Contributions
Aline Y. Kato: conceptualization, validation, investigation, resources, data curation. Tainá A. L. Freitas: investigation, resources, data curation. Cássia R. A. Gomes: investigation, resources, data curation. Thais R. R. Alves: investigation, resources, data curation. Yara M. M. Ferraz: investigation, resources, data curation. Matheus F. Trivellato: investigation, resources, data curation. David De Jong: conceptualization, writing – original draft, writing – review & editing. Jaqueline D. Biller: conceptualization, validation, writing – review & editing. Daniel Nicodemo: conceptualization, formal analysis, writing – original draft, writing – review & editing, project administration.
Funding
This research was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo - FAPESP (process no. 2021/00702-1) to Daniel Nicodemo.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
This research was supported by grants from the Brazilian research foundation, Fundação de Amparo à Pesquisa do Estado de São Paulo - FAPESP (process no.2021/00702-1).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Azpiazu, C.; Bosch, J.; Bortolotti, L.; Medrzycki, P.; Teper, D.; Molowny-Horas, R.; Sgolastra, F. Toxicity of the insecticide sulfoxaflor alone and in combination with the fungicide fluxapyroxad in three bee species. Sci. Rep. 2021, 11, 6821. [Google Scholar] [CrossRef] [PubMed]
- Balieira, K.V.B.; Mazzo, M.; Bizerra, P.F.V.; Guimarães, A.R.J.S.; Nicodemo, D.; Mingatto, F.E. Imidacloprid-induced oxidative stress in honey bees and the antioxidant action of caffeine. Apidol. 2018, 49, 562–572. [Google Scholar] [CrossRef]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Bayer. Fox® Xpro: fungicida, 2019. São Paulo: Bayer S. A. Available online: https://www.agro.bayer.com.br/d/fungicida-bcs-fox-xpro-br (accessed on 15 April 2024).
- Bayer. Ficha de Informações de Segurança de Produtos Químicos (FISPQ). Fox Xpro. 2022. Available online: https://www.bayerfispq.com.br/Downloads/DownloadFile?idForm=4608 (accessed on 15 April 2024).
- Behmer, S.T. Insect herbivore nutrient regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef] [PubMed]
- Biddinger, D.J.; Robertson, J.L.; Mullin, C.; Frazier, J.; Ashcraft, S.A.; Rajotte, E.G.; Joshi, N.K.; Vaughn, M. Comparative toxicities and synergism of apple orchard pesticides to Apis mellifera (L.) and Osmia cornifrons (Radoszkowski). PLoS One 2013, 8, e72587. [Google Scholar] [CrossRef]
- Bokšová, A.; Kazda, J.; Stejskalová, M.; Šubrt, T.; Uttl, L.; Mráz, P.; Bartoška, J. Findings of herbicide and fungicide residues in bee bread. Plant, Soil Environment 2021, 67, 343–352. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidol. 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Bus, J.S. “The dose makes the poison”: key implications for mode of action (mechanistic), research in a 21st century toxicology paradigm. Curr. Opin. Toxicol. 2017, 3, 87e91. [Google Scholar] [CrossRef]
- Campos, M.G.R.; Frigerio, C.; Lopes, J.; Bogdanov, S. What is the future of bee-pollen? J. ApiProd ApiMed Sci. 2010, 2, 131–144. [Google Scholar] [CrossRef]
- Casida, J.E.; Durkin, K.A. Pesticide chemical research in toxicology: lessons from nature. Chem. Res. Toxicol. 2016, 30, 94–104. [Google Scholar] [CrossRef]
- Chakrabarti, P.; Rana, S.; Sarkar, S.; Smith, B.; Basu, P. Pesticide-induced oxidative stress in laboratory and field populations of native honey bees along intensive agricultural landscapes in two Eastern Indian states. Apidol. 2015, 46, 107–129. [Google Scholar] [CrossRef]
- Christen, V.; Krebs, J.; Fent, K. Fungicides chlorothanolin, azoxystrobin and folpet induce transcriptional alterations in genes encoding enzymes involved in oxidative phosphorylation and metabolism in honey bees (Apis mellifera) at sublethal concentrations. J. Hazard. Mater. 2019, 5, 215–226. [Google Scholar] [CrossRef]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A deficit of detoxification enzymes: Pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [PubMed]
- Corona, M.; Robinson, G.E. Genes of the antioxidant system of the honey bee: annotation and phylogeny. Insect Mol. Biol. 2006, 15, 687–701. [Google Scholar] [CrossRef]
- David, A.; Botías, C.; Abdul-Sada, A.; Nicholls, E.; Rotheray, E.L.; Hill, E.M.; Goulson, D. Widespread contamination of wildflower and bee-collected pollen with complex mixtures of neonicotinoids and fungicides commonly applied to crops. Environ. Int. 2016, 88, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Deising, H.B.; Reimann, S.; Pascholati, S.F. Mechanisms and significance of fungicide resistance. Braz. J. Microbiol. 2008, 39, 286–295. [Google Scholar] [CrossRef] [PubMed]
- D’Hose, D.; Isenborghs, P.; Brusa, D.; Jordan, B.F.; Gallez, B. The short-term exposure to sdhi fungicides boscalid and bixafen induces a mitochondrial dysfunction in selective human cell lines. Molecules 2021, 26, 5842. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey bee health: do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef]
- Fang, Q.; Yan, Z.; Zhang, C.; Shi, Y.; Zhang, Z.; Gao, Q.; Xiao, J.; Liao, M.; Qi, C.; Cao, H. Insights into the fungicide prothioconazole and its metabolite in wheat: residue behavior and risk assessment. Agron. 2023, (12), 2906. [Google Scholar] [CrossRef]
- Fisher, A.; Coleman, C.; Hoffmann, C.; Fritz, B.; Rangel, J. The synergistic effects of almond protection fungicides on honey bee (Hymenoptera: Apidae) forager survival. J. Econ. Entomol. 2017, 110, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.; Degrandi-Hoffman, G.; Liao, L.H.; Tadei, R.; Harrison, J.F. The challenge of balancing fungicide use and pollinator health. Adv. Insect Physiol. 2023, 117–190. [Google Scholar]
- Frumkin, I.; Schirman, D.; Rotman, A.; Li, F.; Zahavi, L.; Mordret, E.; Asraf, O.; Wu, S.; Levy, S.F.; Pilpel, Y. Gene architectures that minimize cost of gene expression. Mol. Cell 2017, 65, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Giannini, T.C.; Cordeiro, G.D.; Freitas, B.M.; Saraiva, A.M.; Imperatriz-Fonseca, V.L. The dependence of crops for pollinators and the economic value of pollination in Brazil. J. Econ. Entom. 2015, 108, 849–857. [Google Scholar] [CrossRef]
- Gomes, C.R.A.; Freitas, T.A.L.; Kato, A.Y.; Silva, M.A.G.; Ferraz, Y.M.M.; Alves, T.R.R.; Serafim, J.A.; De Jong, D.; Prado, E.P.; Vicente, E.F.; et al. Contact contamination of honey bees in controlled fungicide spraying using fine and coarse droplets. In: 48th Apimondia, 2023, Santiago. Abstract Book - 48th Apimondia 2023, 233.
- Haas, J.; Zaworra, M.; Glaubitz, J.; Hertlein, G.; Kohler, M.; Lagojda, A.; Lueke, B.; Maus, C.; Almanza, M.T.; Davies, T.G.E.; et al. A toxicogenomics approach reveals characteristics supporting the honey bee (Apis mellifera L.) safety profile of the butenolide insecticide flupyradifurone. Ecotoxicol. Environ. Saf. 2021, 217, 112247. 217.
- Ibama. Vendas de ingredientes ativos por unidade da federação em 2022. In: Relatórios de comercialização de agrotóxicos. 2023. Available online: https://www.gov.br/ibama/pt-br/assuntos/quimicos-e-biologicos/agrotoxicos/relatorios-de-comercializacao-de-agrotoxicos (accessed on 11 April 2024).
- Iwasaki, J.M.; Hogendoorn, K. Non-insecticide pesticide impacts on bees: A review of methods and reported outcomes. Agr. Ecosyst. Environ. 2021, 314, 107423. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, H.; Cha, J.; Lee, S.H.; Kim, Y.H. Validation of quantitative real-time PCR reference genes and spatial expression profiles of detoxication-related genes under pesticide induction in honey bee, Apis mellifera. PLoS One 2022, 17, e0277455. [Google Scholar] [CrossRef]
- LIAO, L.-H.; WU, W.-Y.; DAD, A.; BERENBAUM, M.R. Fungicide suppression of flight performance in the honeybee (Apis mellifera) and its amelioration by quercetin. Proc. R. Soc. Lond. B. Biol. Sci. 2019, 286, 20192041. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, A.P.; Mackert, A.; Cristino, A.S.; Simões, Z.L.P. Validation of reference genes for gene expression studies in the honey bee, Apis mellifera, by quantitative real-time RT-PCR. Apidol. 2008, 39, 372–385. [Google Scholar] [CrossRef]
- MATTOS, I.M.; SOUZA, J.; SOARES, A.E. E. Analysis of the effects of colony population size and feeding supplementation on bee pollen production, Journal of Apicultural Research, v.54, p.411-419. 2015.
- National Honey Bee Health Stakeholder. Conference Steering Committee. Report on the National Stakeholders Conference on Honey Bee Health. US Department of Agriculture, Washington, DC. 2012.
- Nicodemo, D.; Mingatto, F.E.; De Jong, D.; Bizerra, F.V.B.; Tavares, M.A.; Bellini, W.; Vicente, E.F.; Carvalho, A. Mitochondrial respiratory inhibition promoted by pyraclostrobin in fungi is also observed in honey bees. Environ. Toxicol. Chem. 2020, 39, 1267–1272. [Google Scholar] [CrossRef]
- OECD. Test No. 214: Honeybees, Acute Contact Toxicity Test. OECD Guidelines for the Testing of Chemicals 1998, 2, 2–4. [Google Scholar]
- Olgun, T.; Dayioğlu, M.; Özsoy-Taġkiran, N. Pesticide and pathogen induced oxidative stress in honey bees (Apis mellifera L.). Mellifera 2020, 20, 32–52. [Google Scholar]
- Oliver, R.; Hewitt, H.G. Fungicides in crop protection. 2.ed. CABI: Boston, United States, 2014, 189.
- Omar, E.M.; Darwish, H.Y.A.; Othman, A.A.; El-Seedi, H.R.; Naggar, Y.A. Crushing corn pollen grains increased diet digestibility and hemolymph protein content while decreasing honey bee consumption. Apidol. 2022, 53, 52–2022. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper - Excel - based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Pettis, J.S.; Lichtenberg, E.M.; Andree, M.; Stitzinger, J.; Rose, R.; Vanengelsdorp, D. Crop pollination exposes honey bees to pesticides which alters their susceptibility to the gut pathogen Nosema ceranae. PLoS One.
- Picard, M.; Taivassalo, T.; Gouspillou, G.; Hepple, R.T. Mitochondria: isolation, structure and function. Physiol. J. 2011, 598, 4413–4421. [Google Scholar] [CrossRef] [PubMed]
- Prado, A.; Pioz, M.; Vidau, C.; Requier, F.; Jury, M.; Crauser, D.; Brunet, J.L.; Le Conte, Y.; Alaux, C. Exposure to pollen-bound pesticide mixtures induces longer-lived but less efficient honey bees. Sci. Total Environ. 2019, 650, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, G.M.; Grozinger, C.M. Honey bee nutritional ecology: from physiology to landscapes. Adv. Insect Physiol. 2023, 64, 289–345. [Google Scholar]
- Raimets, R.; Bontšutšnaja, A.; Bartkevics, V.; Pugajeva, I.; Kaart, T.; Puusepp, L.; Pihlik, P.; Keres, I.; Viinalass, H.; Mänd, M.; et al. Pesticide residues in beehive matrices are dependent on collection time and matrix type but independent of proportion of foraged oilseed rape and agricultural land in foraging territory. Chemosphere 2020, 238, 124555. [Google Scholar] [CrossRef] [PubMed]
- Rebouças, E.D.L.; Costa, J.J.D.N.; Passos, M.J.; Passos, J.R.D.S.; Hurk, R.V.D.; Silva, J.R.V. Real time PCR and importance of housekeepings genes for normalization and quantification of mRNA expression in different tissues. Braz. Arch. Biol. Technol. 2013, 56, 143–154. [Google Scholar] [CrossRef]
- Rehman, S.; Waliullah, M.I.S. Chlorpyrifos-induced neuro-oxidative damage in bee. J. Toxicol. Environ. Health Sci. 2012, 4, 30–36. [Google Scholar] [CrossRef]
- Rondeau, S.; Raine, N.E. Fungicides and bees: a review of exposure and risk. Environ. Int. 2022, 165, 107311. [Google Scholar] [CrossRef]
- Roszko, M.L.; Kamińska, M.; Szymczyk, K.; Jędrzejczak, R. Levels of selected persistent organic pollutants (PCB, PBDE) and pesticides in honey bee pollen sampled in Poland. PLoS One 2016, 11, e0167487. [Google Scholar] [CrossRef] [PubMed]
- Salmela, H.; Harwood, G.P.; Münch, D.; Elsik, C.G.; Herrero-Galán, E.; Vartiainen, M.K.; Amdam, G.V. Nuclear translocation of vitellogenin in the honey bee (Apis mellifera). Apidol. 2022, 53, 13. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS 9.2 online documentation. SAS Institute Inc., Cary, NC. 2013.
- SCHMEHL, D.R.; TEAL, P.E.A.; FRAZIER, J.L.; GROZINGER, C.M. Genomic analysis of the interaction between pesticide exposure and nutrition in honey bees (Apis mellifera). J. Insect Physiol. 2014, 71, 177–190. [Google Scholar] [CrossRef]
- Schuhmann, A.; Schmid, A.P.; Manzer, S.; Schulte, J.; Scheiner, R. Interaction of insecticides and fungicides in bees. Front. Insect Sci. 2022, 1, 808335. [Google Scholar] [CrossRef]
- SGOLASTRA, F.; MEDRZYCKI, P.; BORTOLOTTI, L.; MAINI, S.; PORRINI, C.; SIMON-DELSO, N.; BOSCH, J. Bees and pesticide regulation: lessons from the neonicotinoid experience. Biol. Conserv. 2020, 241, 108356. [Google Scholar] [CrossRef]
- Siede, R.; Meixner, M.D.; Buchler, R. Comparison of transcriptional changes of immune genes to experimental challenge in the honey bee (Apis mellifera). J. Apic. Res. 2012. [Google Scholar]
- Silva-Neto, C.M.; Ribeiro, A.C.C.; Gomes, F.L.; Neves, J.G.; Melo, A.P.C.; Calil, F.N.; Nascimento, A.R.; Franceschinelli, E.V. Interaction between biological and chemistry fungicides and tomato pollinators. Rev. Colomb. Cienc. Hortic. 2018, 12, 425–435. [Google Scholar] [CrossRef]
- Simon-Delso, N.; San Martin, G.; Bruneau, E.; Hautier, L. Time-to-death approach to reveal chronic and cumulative toxicity of a fungicide for honeybees not revealed with the standard ten-day test. Sci. Rep. 2018, 8, 7241. [Google Scholar] [CrossRef]
- SIMONE, M.; EVANS, J.D.; SPIVAK, M. Resin collection and social immunity in honey bees. Evolution 2009, 63, 3016–3022. [Google Scholar] [CrossRef]
- Somerville, D. Fat bees skinny bees: a manual on honey bee nutrition for beekeepers. GOULBURN: Australian Government Rural Industries Research and Development Corporation. 2005. 142p.
- Tadei, R.; Menezes-Oliveira, V.; Silva-Zacarin, E.C. M. Silent effect of the fungicide pyraclostrobin on the larval exposure of the non-target organism Africanized Apis mellifera and its interaction with the pathogen Nosema ceranae in adulthood. Environ. Pollut. 2020, 267, 115622. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee nutrition and floral resource restoration. Curr. Opin. Insect Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef]
- Weirich, G.F.; Collins, A.M.; Williams, V.P. Antioxidant enzymes in the honey bee, Apis mellifera. Apidol. 2022, 33, 3–14. [Google Scholar] [CrossRef]
- Wood, S.C.; Chalifour, J.C.; Kozii, I.V.; Medici De Mattos, I.; Klein, C.D.; Zabrodski, M.W.; Moshynskyy, I.; Guarna, M.M.; Wolf Veiga, P.; Epp, T.; et al. In vitro effects of pesticides on european foulbrood in honeybee larvae. Insects 2020, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Meng, F.; Jia, H.; Guo, X.; Xu, B. The identification and oxidative stress response of a zeta class glutathione S-transferase (GSTZ1), gene from Apis cerana cerana. J. Insect Physiol. 2012, 58, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: an overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef]
Figure 1.
Weekly amount of bee pollen collected from four colonies over 15 weeks (mean ± standard error).
Figure 1.
Weekly amount of bee pollen collected from four colonies over 15 weeks (mean ± standard error).
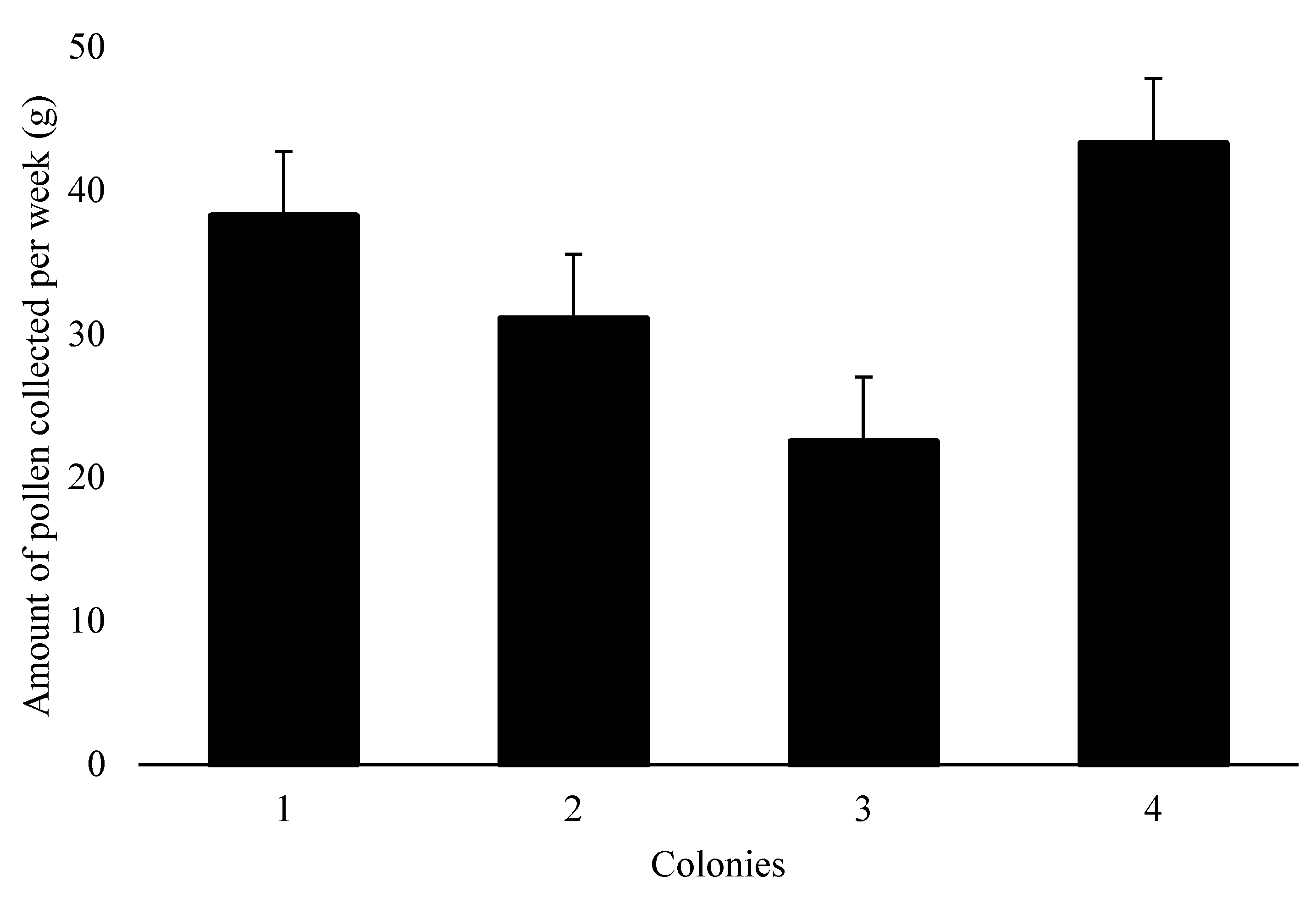
Figure 2.
Gene expression in the fat body of honey bee foragers from colonies with supplemented or restricted feeding and exposed or not to bixafen (bix), prothioconazole (pro) and trifloxystrobin (tri). Dose 1 = 1µg A. I. bee-1; dose 7 = 7µg bee-1. Shown are the medians of four biological samples per treatment in triplicate. *Different from control (C) with food supplementation.
Figure 2.
Gene expression in the fat body of honey bee foragers from colonies with supplemented or restricted feeding and exposed or not to bixafen (bix), prothioconazole (pro) and trifloxystrobin (tri). Dose 1 = 1µg A. I. bee-1; dose 7 = 7µg bee-1. Shown are the medians of four biological samples per treatment in triplicate. *Different from control (C) with food supplementation.
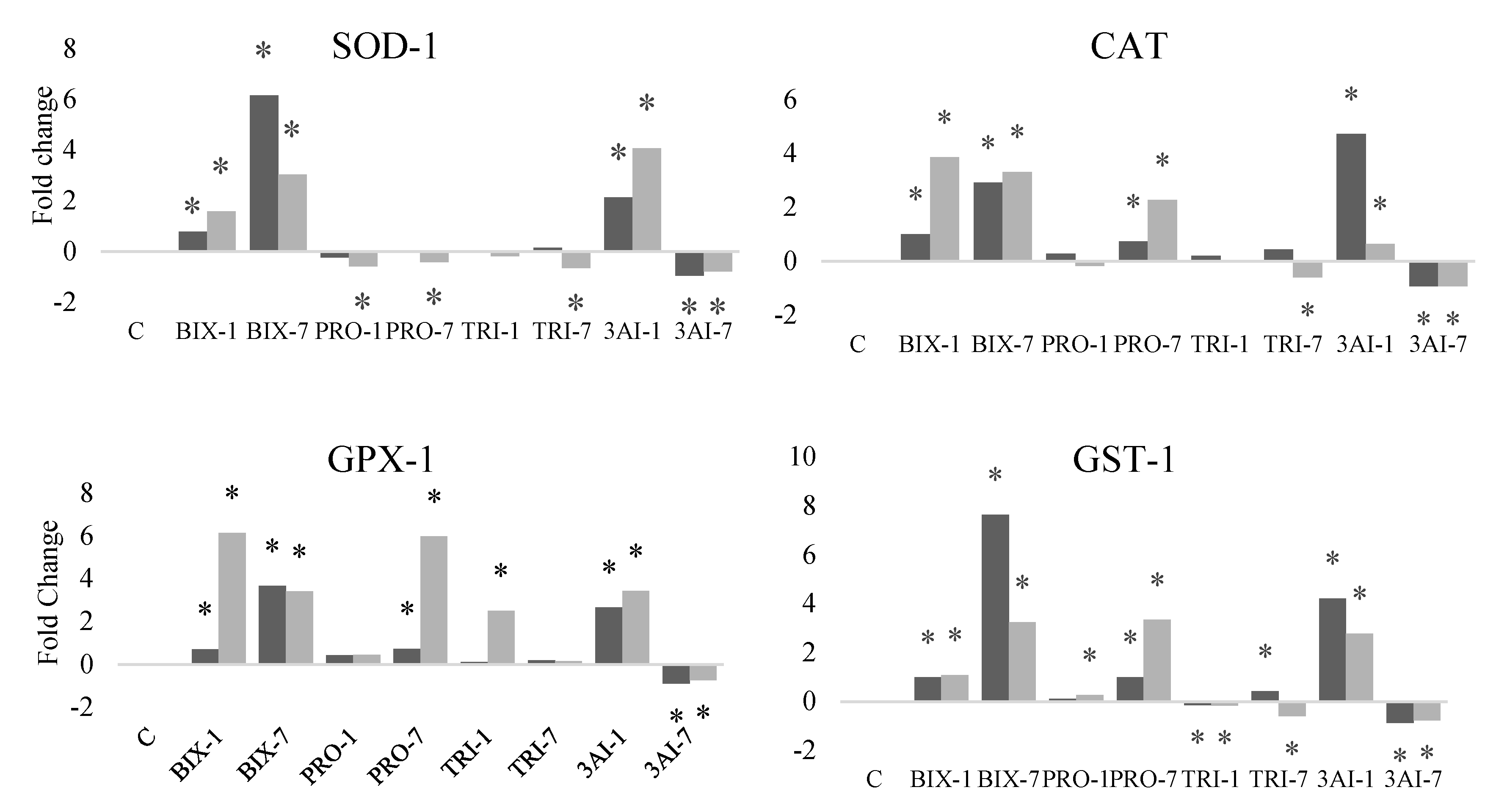
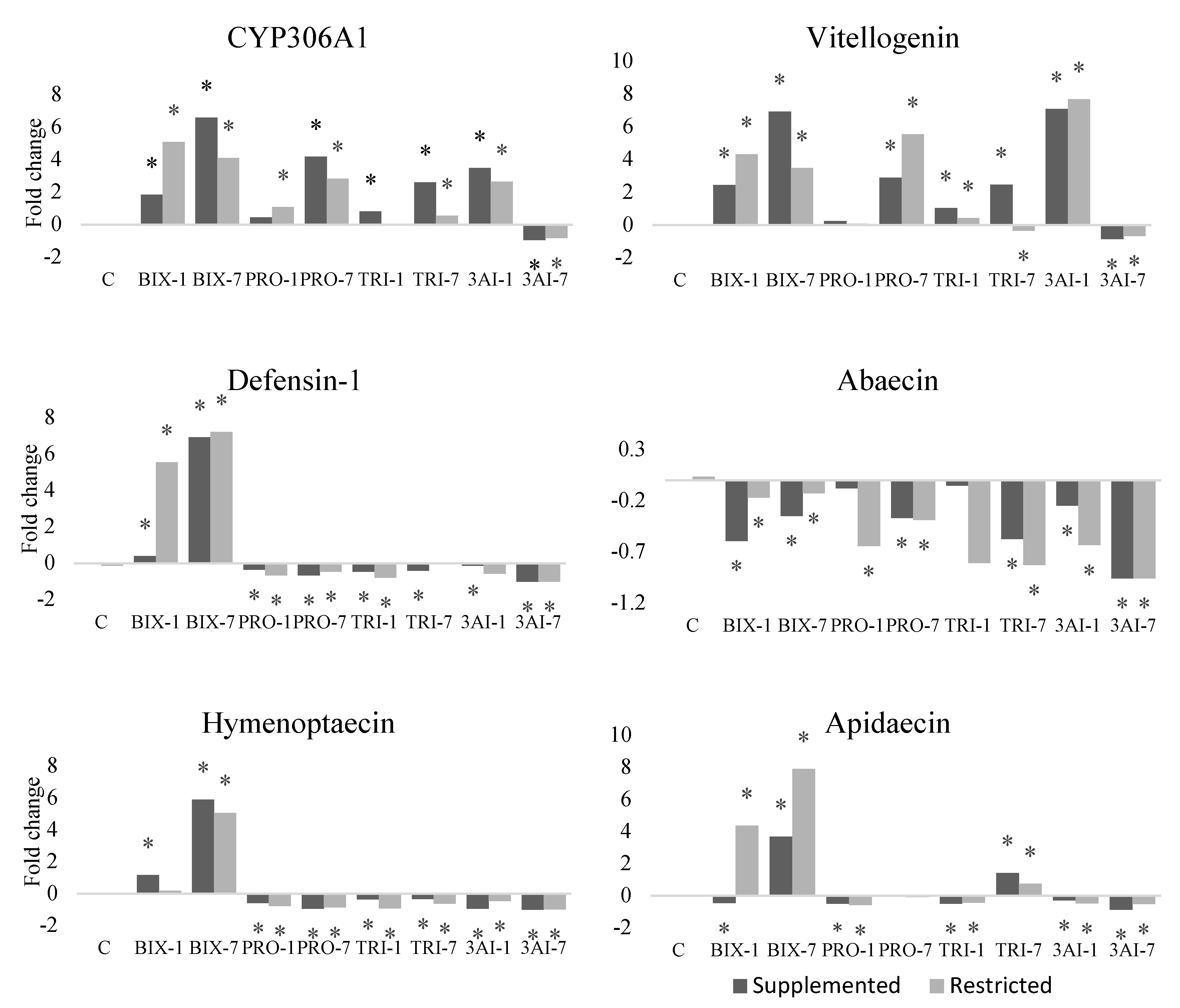

Table 1.
Reference gene and genes related to bee health that were analyzed, and their respective primer sequences (GenBank accession numbers) and references.
Table 1.
Reference gene and genes related to bee health that were analyzed, and their respective primer sequences (GenBank accession numbers) and references.
| Genes | Sequence (5’-3’) | References |
|---|---|---|
| Ribosomal Protein L32 (RPL32) (former rp49) | F: CGTCATATGTTGCCAACTGGT R: TTGAGCACGTTCAACAATGG |
Lourenço et al. (2008) |
| Superoxide Dismutase 1 (SOD-1) |
F: GGTGGTGGTCATTTGAATCATTC R: AAGAAGTGCAGCGTCTGGTTTAC |
Kim et al. (2022) |
| Catalase (CAT) |
F: TGGAGCAAGTCCTGATAAAATGC R: TGGGCCAAGACGATGTCTATG | Kim et al. (2022) |
| Glutathione Peroxidase 1 (GPX-1) |
F: CGACAACTATAAGGAAGCGAAA R: AGATAGAAAAACGTCTTCGCCT |
Corona and Robinson (2006) |
| Glutathione S-transferase 1 (GST-1) | F: TGCCGATCGATTTTTATCAACTT R: AGCCGTCAACGCAACTGC |
Corona and Robinson (2006) |
| Cytochrome P450 306A1 (CYP306A1) |
F: CGTCGATGGGAAGGATAAAA R: TCGGTGAAATATCCCGATTC |
Corona and Robinson (2006) |
| Vitellogenin | F: TCGACAACTGCGATCAAAGGA R: TGGTCACCGACGATTGGATG |
Simone et al. (2009) |
| Defensin-1 | F: TGCGCTGCTAACTGTCTCAG R: AATGGCACTTAACCGAAACG |
Siede et al. (2012) |
| Abaecin | F: AGATCTGCACACTCGAGGTCTG R: TCGGATTGAATGGTCCCTGA |
Siede et al. (2012) |
| Hymenoptaecin | F: CTCTTCTGTGCCGTTGCATA R: ′GCGTCTCCTGTCATTCCATT |
Siede et al. (2012) |
| Apidaecin | F: CTTTGTAGTCGCGGTATTTGG R: AGGCGCGTAGGTCGAGTAG |
Siede et al. (2012) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



