
Submitted:
04 May 2024
Posted:
06 May 2024
You are already at the latest version
Abstract
Antimicrobial resistance (AMR) poses a critical global health threat, leading to approximately 1.27 million deaths annually from infectious diseases resistant to existing antibiotics. Factors such as antibiotic misuse and environmental contamination have accelerated AMR, with the COVID-19 pandemic exacerbating the issue due to increased antibiotic usage. However, the development of new antibiotics has stagnated, with only a few approved in recent years, and many facing resistance. Among the priority pathogens identified by the WHO, Pseudomonas aeruginosa stands out as a significant concern due to its multidrug resistance and virulence. P. aeruginosa infections are challenging to treat due to intrinsic and acquired resistance mechanisms, as well as its ability to form biofilms. Additionally, P. aeruginosa produces a plethora of virulence factors, including proteases, siderophores, and toxins, further complicating treatment. Quorum sensing (QS) systems play a crucial role in regulating bacterial physiology and virulence factor expression. P. aeruginosa employs three QS systems, making it an attractive target for quorum quenching (QQ) strategies. Enzymes such as AHL lactonases, acylases, and oxidoreductases have shown promise in inhibiting QS, thus reducing virulence without impacting bacterial growth. Furthermore, natural QS inhibitors produced by various organisms offer potential therapeutic alternatives. Despite promising results in vitro, translating QQ strategies to clinical settings remains challenging. Nevertheless, QQ represents a novel approach in combating multidrug-resistant infections and warrants further investigation as a complementary therapeutic strategy alongside traditional antibiotics. This review summarizes enzymes with Qurorum Quenching activity evaluated against the pathogen P. aeruginosa and their potential applications in combination therapies.
Keywords:
quorum quenching
; Pseudomonas aeruginosa
Introduction
Currently, antimicrobial resistance (AMR) is one of the greatest threats to global health, food security and development. Around 1,27 millions deaths per year worldwide are caused by infectious diseases that are resistant to currently available antibiotics (Tang et al., 2023). In recent decades, AMR has been accelerated due to multiple factors, such as overuse or inappropriate use of antibiotics, contamination of water sources with antimicrobial drugs, among others. AMR has been further accelerated as a consequence of the COVID-19 pandemic, due to the increased use of antibiotics to treat patients in an attempt to avoid bacterial co-infections (Kariyawasam et al., 2022).
In addition, the development of new antibiotics is "stalled" and the effort of the scientific community appears to be insufficient to address the growing threat of AMR. In 2017 there were 31 new antibiotics in clinical development, while in 2021 that number dropped to 27. Since 2017 only 12 new antibiotics have been approved; among them, ten are already facing resistance from some pathogens (Tang et al., 2023). Within the antimicrobials that are in clinical development, about half correspond to "non-traditional" ones, including, for example, monoclonal antibodies and bacteriophages. This indicates that the current trend is the search for therapeutic agents that offer new opportunities in the treatment of infections caused by resistant bacteria, and that at the same time do not favor the development of resistances.
In 2017, the WHO published the first list of multidrug-resistant "priority pathogens", with Acinetobacter (A.) baumanni and Pseudomonas (P.) aeruginosa at the top of the list. The critical priority of these microorganisms means that urgent research and development of effective drugs is required (Lee et al., 2023). Both bacteria, together with Enterococcus (E.) faecium, Staphylococcus (S.) aureus, Klebsiella (K.) pneumoniae and Enterobacter spp. constitute the group known as ESKAPE (Lee et al., 2023), an acronym used for pathogenic bacteria recognized as multidrug-resistant (MDR). The MDR of these pathogens and their increasing prevalence as nosocomial pathogens, is of great concern, especially in critically ill and immunocompromised patients (Tigabu & Getaneh, 2021; Salim et al., 2024).
- Pseudomonas aeruginosa: the untreatable enemy of modern medicine
Among the ESKAPE, P. aeruginosa has received particular attention. This Gram-negative opportunistic human pathogen, possesses extraordinary metabolic versatility that enables the survival and proliferation under several environmental conditions (Rather et al., 2022). Together with its several virulence factors, this flexibility directly influences the relevance of this bacterium. P. aeruginosa causes infections in immunocompromised individuals with chronic obstructive pulmonary disease, burns, sepsis, cancer patients undergoing chemotherapy, urinary tract infections, cystic fibrosis, ventilator-associated pneumonia, HIV patients, among others.
In addition, P. aeruginosa has become common in ICUs (Salim et al., 2024).
Treatments to combat P. aeruginosa infections are extremely difficult due to an elevated and extensive antibiotic resistance profile, which is related to its intrinsic and acquired resistance mechanisms (Qin et al., 2022). Intrinsic resistance (IR) refers to certain characteristics like modifying the membrane permeability that prevents antibiotics from entering the cell, or the presence of efflux pumps, the best-preserved mechanism for expelling antibiotics. IR also includes enzymes, e.g., broad-spectrum ß-lactamases, that hydrolyse antibiotics, ensuring its resistance to penicillin and cephalosporin (Qin et al., 2022; Lee et al., 2023). Moreover, biofilm formation functions as a highly effective barrier that protects the cell by increasing its resistance to antibiotics (Qin et al., 2022).
Acquired resistance mechanisms involve both the horizontal genes acquisition and mutational resistance. Mobile genetic elements, e.g., plasmids, transposons, integrons, insertion sequences, among others, facilitate the horizontal acquisition of antibiotic-resistance genes. Mutational resistance refers to the acquisition of resistance through chromosomal mutations (López-Causapé et al., 2018). These resistances are important in chronic infections such as pulmonary infections in cystic fibrosis patients (Oliver et al., 2000; Lee et al., 2023). The cumulative impact of these factors in P. aeruginosa renders its infections resistant to traditional antibiotic therapy, posing a significant challenge.
- A plethora of virulence factors in P. aeruginosa
Virulence factors production and biofilm formation are important to establish and maintain the P. aeruginosa infection in host cells.
Protease production is considered the main factor contributing to the virulence of P. aeruginosa (Hoge et al., 2010; Liao et al., 2022). About 2.8% of its genome codes for different types of proteases, including metallo-proteases, serine-proteases, cysteine-proteases, threonine-proteases, among others, with diverse functions in the infection process (Rawlings 2006). For example, alkaline protease, a metalloprotease, is directly involved in causing tissue necrosis due to the degradation of laminin, a main component of the basal lamina. In addition, together with elastase B, alkaline protease can block the host immune response by different mechanisms, including the inactivation of human γ-interferon and human tumor necrosis factor-α. They also inhibit the function of neutrophils and leukocytes, allowing the bacteria to evade the phagocytosis of the host defense system (Hoge et al., 2010). Pseudolysin is another protease that acts on several host proteins, including collagens, proteoglycans, immunoglobulins, fibrinogen, cytokines and elastin among others. Its action on elastin is the most relevant since this highly cross-linked protein is very resistant to most proteases.
Siderophores pyoverdine and pyochelin are also characteristic pathogenic factors in P. aeruginosa. These low molecular-weight compounds are iron chelating agents that provide the bacteria with a survival advantage in iron-limiting conditions. It has been shown that the pyoverdine production by P. aeruginosa allows the establishment of chronic infections through the evasion of the elimination mechanisms of the host Meyer et al. (1996), Tielen et al. (2011) and Kang et al. (2019).
Similarly, several secondary metabolites, including pyocyanin, rhamnolipids and hydrogen cyanide play a role as virulence factors. Pyocyanin can interfere with the immune system of the host promoting the infection (Hall et al., 2016; Allen et al. (2005). Rhamnolipids are glycolipid reservoirs that mainly function in maintaining the cell surface hydrophobicity, but also promote hemolysis and swarming motility (Wang et al. (2005), and are important for the biofilm structure. Rhamnolipids also facilitate the evasion of the antimicrobial response of the host cell, and affect the immune system attacking leukocytes and macrophages (McClure and Schiller, 1996; Jensen et al., 2006). Hydrogen cyanide is a diffusible toxin that inhibits the mitochondrial cytochrome c oxidase and thereby blocking the aerobic respiration (Gallagher and Manoil, 2001).
On another hand, the ability of P. aeruginosa to form biofilm largely contributes during the infection of the host. The biofilm is formed by a complex matrix of bacteria surrounded mainly by self-produced extracellular polymeric substances (EPS), including proteins, polysaccharides, lipids and extracellular DNA (Thi et al., 2020). This complex matrix confers protection from the host immune responses preventing phagocytosis, and increasing their resistance to antimicrobial treatments. Thus, biofilm formation is a critical weapon to compete, survive and dominate during infections conferring a clear advantage for the colonization and long-term persistence, as in the case of chronic infections. (Thi et al., 2020). In addition, biofilm formation allows P. aeruginosa to effectively colonize different surfaces, like urinary catheters, implants, among others (Ghafoor et al., 2011; Thi et al., 2020).
- Quorum Sensing systems and the regulation of the bacterial physiology
Beyond the pathogenic nature of a microorganism like P. aeruginosa, cells have to produce virulence factors in the right place at the right time. The effectiveness of an enzyme required for the colonization of a niche, for instance, could be suboptimal when produced by a single cell, considering the quantity of enzyme a cell can synthesize. Then, metabolic costs associated with the enzyme production can be optimized if its synthesis is delayed until a population is present at an optimal density. In addition, considering that the virulence factors alert the defense mechanisms of the host, it is more profitable to associate its production to the population density. Clearly, the determination of the population density is a key aspect in the regulation of microbial physiology. In consequence, a microorganism like a bacterium, which can not “count”, necessitates a communication system in order to coordinate the gene expression with the other members of the population. Quorum Sensing (QS) is one of these systems that has received more attention in the last 20 years.
QS systems are communication mechanisms that rely on the production and detection of signaling molecules called autoinducers (AI). Produced inside the cells, these AI are extracellularly accumulated at low levels when the population density is low. As cells divide and the density increases, the AI concentration reaches a threshold concentration. The chemical signal is then bound by a ligand binding domain of a dedicated receptor protein, which changes its conformational structure after this AI-receptor complex. In the case of LuxR-family receptors, the AI-receptor complex then functions as a transcriptional regulator through a DNA binding domain. As a consequence of the increase in cell density, several genes are then repressed and/or induced. QS-regulated genes are directly or indirectly relevant for microbe-microbe and host-microbe interactions (see below for examples concerning P. aeruginosa).
The canonical QS system was originally described in Vibrio fischeri, a marine bacterium that shows both a free lifestyle with low cell densities, and a host-associated lifestyle with certain fishes and squids. When V. fischeri colonizes the symbiosis organs of the host, high cell densities are attained, allowing the activation of QS-regulated phenotypes. In this bacterium, the QS signal is N-(3-oxohexanoyl)-L-homoserine lactone (3OC6-HSL), a member of the acyl homoserine lactone (AHL) family. This group of QS molecules are characterized by a homoserine lactone ring, with an acyl side chain ranging from four to 18 carbons in length. The third position of the acyl side chain can be fully reduced or have a carbonyl or hydroxyl group. Certain long chain AHLs also present double bonds. With very few exceptions, AHL signals are synthesized by LuxI-related enzymes. In V. fischeri, LuxI is the acylhomoserine lactone synthase related to the intracellular production of 3OC6-HSL. LuxR is the V. fischeri AHL receptor protein that induces the expression of the lux operon, where the genes related to bioluminescence production are coded. To date, AHL-dependent QS systems have been described in both beneficial and pathogenic Alpha-, Beta- and Gammaproteobacteria. Other QS systems described in Gram positive or negative bacteria employ specific signal molecules, and possess dedicated enzymes and receptors.
- Three QS systems coexist in P. aeruginosa
Three interrelated QS systems arranged in a hierarchical regulatory cascade have been described in P. aeruginosa (Venturi 2006; Williams et al., 2007). Two of them, named Las and Rhl, are based on the production of AHL as signaling molecules (Figure 1). The AHL synthases LasI and RhlI synthesize N-(3-oxododecanoyl)-L-homoserine lactone (3OC12-HSL) and N-butanoyl-L-homoserine lactone (C4-HSL), respectively. When these molecules reach a threshold concentration due to increased bacterial population density, they are bound by their cognate receptors LasR and RhlR, respectively. The corresponding AHL-receptor complexes activates the expression of virulence genes and modulates specific phenotypes highly relevant for the pathogeny. For instance, LasR is involved in the expression of genes related to biofilm formation, proteases, elastases, hydrogen cyanide, among others (Rutherford and Bassler, 2012; García-Reyes et al., 2020). Meanwhile, RhlR regulates the expression of genes involved in the production of pyocyanin, hydrogen cyanide, siderophores, elastases, alkaline protease and rhamnolipids (Papenfort and Bassler, 2016; García-Reyes et al., 2020).
A third QS system depends on 2-alkyl-4-quinolones (AQs), including 2-alkyl-3,4-dihydroxyquinoline (PQS) and its precursor 4-hydroxy-2-heptylquinoline (HHQ). These QS signals interact with the protein receptor PqsR (Garcia-Reyes et al., 2021), in order to directly activate the expression of at least 35 loci, including the operon pqsABCDE that code the enzymes responsible for the synthesis of AQs. Finally, the PqsH enzyme catalyzes the conversion of HHQ to PQS and the pqsH gene is coded separately on the chromosome. The PQS system is involved in the expression of genes related to biofilm production, proteases, elastases, pyocyanin, rhamnolipids and the two siderophores pyochelin and pyoverdin (García-Reyes et al., 2020; Chadha et al., 2022).
- The therapeutic potential of Quorum Quenching in the age of antibiotic resistance
In recent years, interest in Quorum Quenching (QQ) has increased due to the different biotechnological applications, being its use as a therapeutic alternative one of the most relevant. As mentioned above, P. aeruginosa resists several antibiotics due to multiple mechanisms and, in consequence, their infections are highly problematic for combating with traditional antibiotic therapies. In this context, interfering with the regulation of the virulence expression by means of blocking or inhibiting the QS system has emerged as a promising alternative. Since the expression of genes related to virulence factors production and biofilm formation are regulated by QS, it is expectable that blocking this signaling system may lower the pathogenicity and increase the susceptibility to traditional antibiotic treatment (LaSarre & Federle, 2013). In addition, a remarkable advantage of QQ strategies is the interference with the production of virulence factors without inhibiting the growth of the pathogen. This antivirulence-based therapy strategy is a novel approach in the search for new treatments to combat infections (Sikdar & Elias, 2020).
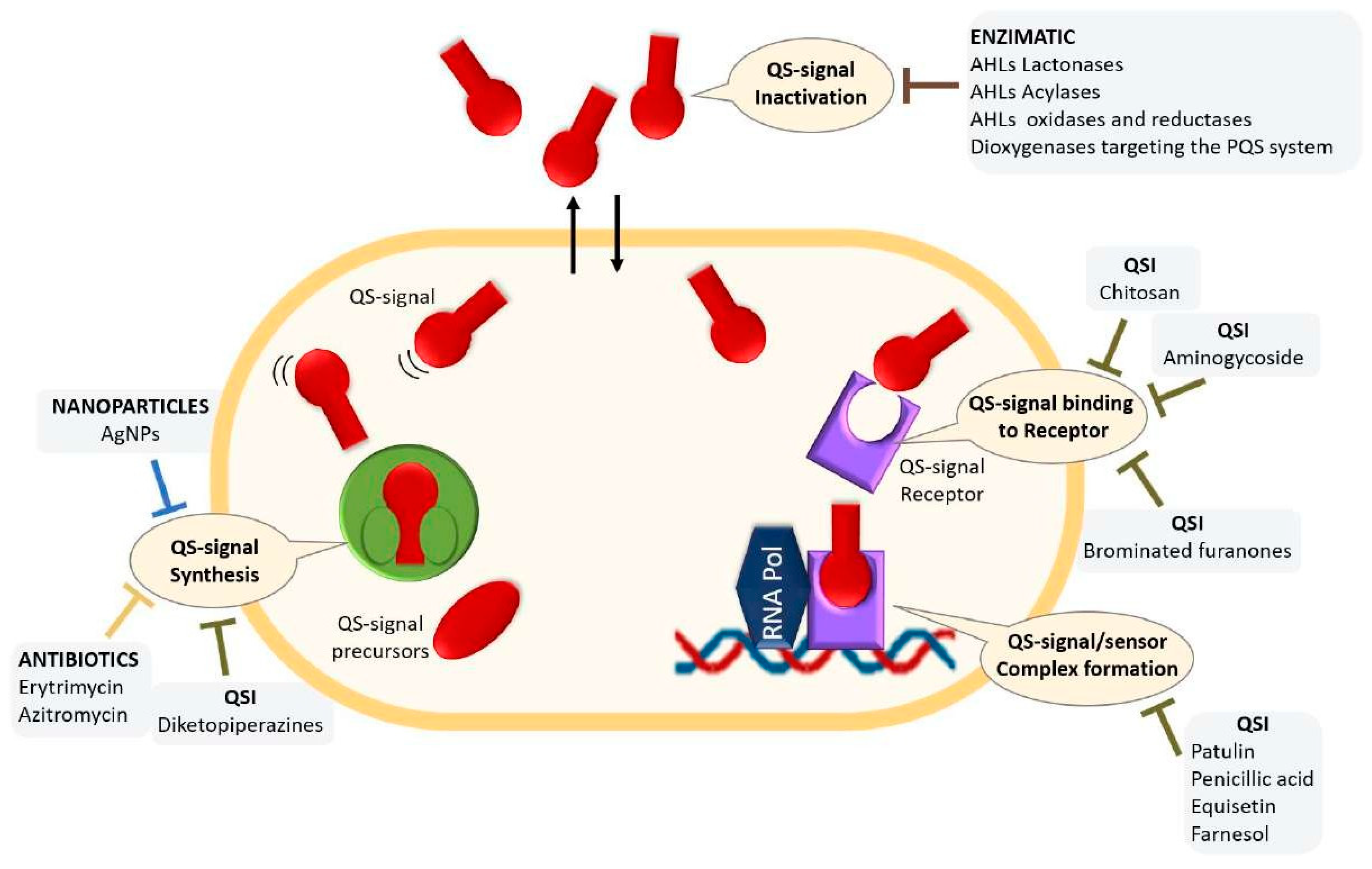
Considering the steps in the QS mechanisms (Figure 2), the interference in the communication can occur at different levels: the AI synthesis, the inactivation of IA molecules, the AI binding to the receptor, the AI efflux from the cell, or the transcription of target genes (Zhou et al., 2020; Rather et al., 2022). However, one of the best characterized QQ strategies are those related to the enzymatic degradation of AI signal molecules in general, and of AHL in particular. A large number of QQ enzymes from different sources, including bacteria, archaea and fungi, have been described that inactivate AHLs of human, animal and plant Gram-negative pathogens (Fetzner, 2015; Grandclément et al., 2016).
- Quorum Quenching enzymes
Quorum Quenching enzymes (QQE) inactivate IAs of different types and by different mechanisms of action. Production of QQE has been reported in both prokaryotic cells, including bacteria and archaea, and eukaryotic cells, including plants and mammals. The following is a brief description of different enzymes acting on Pseudomonas spp. QS systems.
- AHL lactonases
The first lactonase to be described was the AiiA from Bacillus sp. 240B1 (Dong et al., 2000). AHL lactonase enzymes inactivate the AI molecule by hydrolyzing the ester bond of the homoserine lactone ring, which produces the corresponding acyl homoserine. As a consequence, the protein receptor cannot bind this reaction product and the QS system is blocked. AHL lactonases have a broad substrate specificity, acting on AHLs with variable carbon chain length that range from 4 C to 12 C. To note, the reaction is reversible after acidification, which has diagnostic and practical implications (Yates et al., 2012). In recent years, a large number of lactonase-type enzymes active on P. aeruginosa AHL molecules have been described. These enzymes belong to one of the four families of lactonases: metallo-β-lactamase-like lactonases (MLLs), α/β hydrolase fold lactonases, phosphotriesterase-like lactonases (PLLs), and serum paraoxonases (PONs).
MLLs are characterized by a 104HXHXDH109∼H169 metal binding motif and an αβ/βα fold with two zinc cations. The first MLL member to be characterized was AiiA from Bacillus thuringiensis. When AiiA was reported, it was shown that its expression in the phytopathogen Pectobacterium carotovorum (form. Erwinia carotovora), reduced the synthesis of virulence factors (Dong et al., 2000). Produced in P. aeruginosa, AiiA completely inactivated C4-HSL and 3OC12-HSL, and reduced the production of virulence factors and biofilm formation (Reimmann et al., 2002; Anandan and Vittal 2019). Same results were obtained with crude preparations containing AiiA (Raafat, et al., 2019). Recombinant expression of AiiA even augmented the antibiotic susceptibility of P. aeruginosa, which could be related to a lower biofilm formation (Kiran et al., 2011). Other examples of MLLs effective on P. aeruginosa include the purified AiiK enzyme from Kurthia huakuii LAM0618T (Dong et al, 2018) and purified AiiM from Microbacterium testaceum (López-Jácome et al., 2019), among several others. While enzymes like AiiA inactive both AHLs from P. aeruginosa, others like YtnP from Burkholderia cepacia show more activity on 3OC12-HSL than on C4-HSL (Malešević et al., 2020). Interestingly, the characterization of MomL from Muricauda olearia cast suspicion about the true applicability of this type of enzymes against P. aeruginosa (Tang et al., 2015). Although MomL is very effective for inhibiting biofilm formation under in vitro conditions, this effect is almost absent under more real-life conditions (Zhang et al., 2017).
α/β hydrolase fold lactonases share the same fold with MLLs, but lack the characteristic metal binding motif. AiiM from Microbacterium testaceum StLB037 was the first member of this group to be identified (Wang et al., 2010). Several others have been described. However, most of them have been tested in plant pathogens but not in P. aeruginosa, like JydB from Rhodococcus sp. BH4 (Ryu et al., 2020). An exception is the Aii810 enzyme from a metagenome, reported as more active on 3OC12-HSL than C4-HSL. In addition, Aii810 also reduced protease activity, pyocyanin and alginate production and biofilm formation in P. aeruginosa (Fan et al., 2017).
PLLs are bacterial and archaeal are amidohydrolase characterized by a binuclear metal center in a (α/β)8 fold, also known as TIM barrel. First members of this group were originally described as paraoxonases, though their true lactonase nature was later confirmed. PLLs group include Gkl from Geobacillus kaustophilus HTA426 and SsoPox from Sulfolobus solfataricus, which were tested only with pure P. aeruginosa QS signals, but not on the bacterium itself (Chow et al., 2010).
PONs are enzymes structurally characterized by a six-bladed β-propeller fold with a Ca2+-dependent catalytic site. Mammal PONs like PON1, PON2 and PON3 are the best characterized members. Microbial PONs include a PON-like enzyme from Oceanicaulis alexandrii HTCC2633 (Bar-Rogovsky et al., 2013).
- AHL acylases
Acylase enzymes, also known as amidase or amidohydrolase, inactivate AHL in an irreversible manner by cleaving the amide bond, which releases the corresponding fatty acid and the homoserine lactone ring. This irreversible nature of acylase reactions is valuable, since the AHL structure cannot be recovered at acidic pH (Reina et al., 2021). Leadbetter and Greenberg suggested for the first time an acylase activity in Variovorax paradoxus VAI-C, a soil bacterium that can utilize AHLs as sole nitrogen and energy source (Leadbetter and Greenberg 2000). Later, AiiD, the first acylase to be characterized, was described in Ralstonia sp. XJ12B (Lin et al., 2003). In contrast to AHL lactonases, most AHL acylases belong to the same N-terminal nucleophile (Ntn) hydrolase-fold superfamily of enzymes with an αβ/βα fold. However, with an α/β-hydrolase fold, AiiO from Ochrobactrum sp. A44 is an exception (Czajkowski et al., 2011). AibP from Brucella melitensis 16M, AhlM from Streptomyces sp. M664 and AuAAC from Actinoplanes utahensis (Velasco-Bucheli et al., 2020) are some examples of Ntn hydrolase. Noteworthy, P. aeruginosa possesses at least three Ntn hydrolases active on AHLs: QuiP, HacB and PvdQ.
- AHL oxidases and oxidoreductases
The least numerous of QQ enzymes are AHL oxidases and oxidoreductases. These enzymes act through the modification of the AHL structure via either its oxidation or reduction. As a consequence, the resulting molecules are bound with lower affinities by the corresponding AHL receptors.
To date, the only AHL oxydase described is the cytochrome P450 monooxygenase CYP102A1, also known as P450BM3, from Priestia (form. Bacillus megaterium). However, homologs are present in both producers and non producers of AHLs (Chowdhary et al., 2007). CYP102A1 is a fatty acid monooxygenase that catalyzes the NADPH-dependent oxidation of ω-1, ω-2 or ω-3 carbons of the AHL fatty acid side chain, producing the subterminally hydroxylated AHL derivatives.
The first AHL modification through an oxidoreductase activity was described in Rhodococcus erythropolis W2, a bacterium which also shows other types of QQ enzymes (Uroz et al., 2005). This NADH-dependent AHL modification causes a reduction of the keto substitution at the third position of the acyl side chain, producing the corresponding hydroxyl derivative. To date, the only oxidoreductase enzyme to be characterized is BpiB09, a NADP-dependent dehydrogenase/reductase enzyme obtained from a soil metagenome (Bijtenhoorn et al., 2011).
- Dioxygenases targeting the PQS signals
PQS, the second class of QS signal utilized by P. aeruginosa, is also susceptible to enzymatic inactivation. Hod dioxygenase from Arthrobacter nitroguajacolicus Rü61 was the first enzyme in this group to be described (Pustelny et al., 2009). Hod is an α/β hydrolase that catalyzes the cleavage of one of the rings in the PQS structure through a 2,4-dioxygenolytic rupture, which produces N-octanoyl anthranilic acid and carbon monoxide (Pustelny et al., 2009).
Similar to Hod enzyme, AqdC2 from R. erythropolis BG43 is a dioxygenase that degrades PQS. In addition, AqdB2 from this strain also acts on HHQ, the precursor of PQS (Müller et al., 2015). Homolog enzymes acting on the same substrates were also described in Mycobacterium abscessus subsp. abscessus (Birmes et al., 2017), Nocardia farcinica and Streptomyces bingchenggensis (Wullich et al., 2020).
Treatment of P. aeruginosa PAO1 with Hod enzyme resulted in a decrease of the virulence factors lectin A, pyocyanin, and rhamnolipids, all regulated by the PQS system.
AqdC1 from Rhodococcus erythropolis and AqdC from Mycobacteroides abscessus were described by Müller et al., (2015) and Birmes et al., (2017) respectively. Both enzymes, in contrast to HodC, exhibit a higher activity towards the PQS signal, making them better candidates for therapy against Pseudomonas.
Although the PQS system appears to be specific to Pseudomonas and therefore a very specific target to combat its infections, Arranz San Martín et al. (2022) determined that HQDN.f. and HQDS.b. dioxygenases moderately interfered with P. aeruginosa virulence even in an in vivo studies in Galleria mellonella. These results suggest that at least for P. aeruginosa that regulates its virulence by a complex QS system (see XX), a combination therapy of QQ agents might be more effective (see below).
Bijtenhoorn et al., (2011) reported BpiB09 as the first NADP-dependent reductase. Its expression in P. aeruginosa PAO1 caused a significant reduction in pyocyanin production, a decrease in motility and a drop in biofilm formation. In addition, the enzyme was also tested in an in vivo model using Caenorhabditis elegans with promising results, as the worms remaining after treatment were fully mobile and viable.
Virulence factor production, including proteolytic activities, elastase, pyocyanin and biofilm formation were significantly reduced in all cases after treatment with these enzymes, promising them as possible therapeutic agents.
Despite all the preliminary trials being carried out to propose these enzymes as therapeutic candidates, there is a smaller number of publications recording progress trials using in vivo models. For example, for the previously mentioned only MomL was tested in C. elegans (Tang et al., 2015) and SsoPox was evaluated in rats and showed promising results in both cases (Hraiech et al., 2014). For both YtnP and Y2-aiiA enzymes, the cytotoxic effect was tested using human keratinocyte HaCaT cell line (Malešević et al., 2020).
Certain PONs effective against P. aeruginosa have been reported (Ozer et al., 2005; Aybey and Demirkan, 2016; Faisal et al., 2021) and considering that these PON enzymes have their origin in eukaryotic cells as mammals, could be more compatible than enzymes of prokaryotic origin for its use as therapeutics agent.
Although in vitro modes of QS inhibition have shown promising results yet the translation from bench to clinic is still challenging (Rather et al., 2022).
- QS inhibitors (QSIs)
QSIs production naturally occurred in a large number of organisms like bacteria, fungi, algae and plants. In contrast to QQEs, the wide group of QSIs can interfere with QS systems at different levels. Some examples are listed below (Grandclément et al., 2016).
- Natural QSIs
Although for bacteria the main quorum quenchers described are enzymes, some authors have reported the production of QSIs compounds (Kalia et al., 2019). However, some QSIs compounds have not been fully characterized (Holden et al., 1999; Nithya et al., 2010).
Protoanemonin (4-methylenebut-2-en-4-olide) is an example for QSIs produced by bacteria (Bobadilla Fazzini et al., 2013). This catabolite produced by both Pseudomonas sp. B13 and Pseudomonas reinekei MT1, could reduce the virulence gene expression that are under control of quorum sensing in P. aeruginosa.
Another example are diketopiperazines. These are small cyclic peptides produced mainly by Gram-negative bacteria and to a lesser extent by Gram-positive bacteria. As previously confirmed by Buroni et al. (2018), diketopiperazines act as inhibitors of acyl homoserine lactone synthase. Although there are several diketopiperazines described in the literature, only a few reports assess their effectiveness against Pseudomonas aeruginosa. Recently, Díaz et al. (2020), reported four diketopiperazines produced by Lactobacillus casei CRL 431 and Lactobacillus acidophilus CRL 730. Extracts from both strains inhibited the virulence factors production as elastase and pyocyanin and the biofilm formation against three pathogen Pseudomonas aeruginosa strains.
Fungi, algae and plants have also been described as producers of QSIs compounds. Since they are produced by eukaryotic organisms, they can be considered bio-compatible with human cells; therefore, could be excellent biological candidates for therapeutic approaches (Hemmati et al., 2020). Thus, for example, some metabolites produced by filamentous fungi are already used as antimicrobial pharmaceuticals.
Patulin, penicillic acid (Rasmussen et al., 2005), equisetin (Zhang et al., 2018) and farnesol (Cugini et al., 2007) are some examples of QSIs produced by fungi. Fungi are constantly confronted with bacteria in the environment, so they developed their own defense mechanism through their own production of chemical compounds.
All these compounds were tested against P. aeruginosa in different reports. Rasmussen et al. (2005) performed DNA microarray analysis and determined for both patulin and penicillic acid target preferentially the RhlR controlled genes. Meanwhile, for equisetin, Zhang et al. (2018) showed that this QSI acts at the three QS systems in Pseudomonas, the las, rhl and PQS system, being the last one the most significantly inhibited.
Finally, the farnesol, a sesquiterpene produced by Candida albicans, acts by down-regulating the PQS production, and therefore attenuating the pyocyanin production (Cugini et al., 2007).
Algae have also been shown to exert microbial defense mechanisms in the environment. Delisea pulchra was the first algae described as producing compounds with QQ activity, especially brominated furanones that have structural similarity to AHLs and can bind competitively to LuxR receptors, blocking the QS system (LaSarre and Federle, 2013).
In P. aeruginosa, the production of both exoprotease and pyoverdin were reduced upon treatment with furanones. In addition, these compounds penetrated and disrupted the biofilm (Hentzer et al., 2003).
Despite the furanones have been shown to inhibit QS in vitro (Martinelli et al., 2004) and in vivo (Wu et al., 2004) models, their halogenated structure may be a limitation for their use in humans. Rasch et al. (2004) and Tinh et al. (2007) tested the toxicity in fish trials corroborating these concerns.
Therefore, currently there is a new perspective to find natural products that can effectively inhibit the QS system in Pseudomonas. The plants, used since many years ago for their antibacterial properties, emerge as a source for looking for novel quorum quenchers.
In the last years, attention has been focused on several foods of plant origin searching for compounds with QQ activity. Fruits such as blueberries, vanilla and certain citrus fruits, herbs such as rosemary and turmeric, and some garlic and cinnamon extracts have shown QQ activity (Jakobsen et al., 2012; Bodini et al., 2009; Choo et al., 2006; Girennavar et al., 2008; Khan et al., 2009; Vattem et al., 2007; Vikram et al., 2010; Wang et al., 2012).
Thus, several reports showed that compounds like curcumin from turmeric (Rudrappa and Bais, 2008), iberin isothiocyanate from horseradish (Jakobsen et al., 2012a), coumarin obtained from different plants like Ceylon cinnamon (D´Almeida et al., 2017), allyl sulphide called ajoene from garlic extracts (Jakobsen et al., 2012b; Rasmussen et al., 2005), among others, showed activity against P. aeruginosa by affecting the production of virulence factors and in some cases reducing biofilm formation.
Ajoene also showed a synergistic effect with the antibiotic tobramycin in improving the clearance of P. aeruginosa from the lungs in a mouse model (Jakobsen et al., 2012 b ; Rasmussen et al., 2005).
Another plant-based compound like ginseng used in Chinese traditional medicine (Song et al., 2010), the salicylic acid and trans-ciannamaldehyde (Ahmed et al., 2019) cinnamic acid (Rajkumari et al., 2018), gingerol (Kim et al., 2015) and clove oil (Husain et al., 2013) also have shown effect on the QS-dependent virulence factors in P. aeruginosa. In addition, the cinnamic acid protected C. elegans from P. aeruginosa resulting in lower mortality in an in vivo test.
- Synthetic QSIs
The synthetic QSIs emerge as an alternative to natural QSIs showing certain advantages that we discuss below. In this group we can mention nanoparticles and some synthetic antibiotics which can affect the virulence factors expression in Pseudomonas.
Nowadays, the use of nanoparticles has become widespread, and they are candidates for use in several applications mainly due to their extensive reactivity related to their high area to volume ratio. In addition, the possibility of manufacturing the nanoparticles with the desired characteristics thanks to their chemical, magnetic and electrical properties, is another advantage (Whitesides 2005). For use in applications such as QSI, their stability, efficient delivery, enhanced biofilm penetration, among other advantages, have been shown (Fernandes et al., 2010; Nafee et al., 2014; Qais et al., 2018).
Nanoparticles used as QSIs can act by different mechanisms that function simultaneously causing membrane damage, inducing oxidative stress for generation of reactive oxygen species (ROS), releasing metal ions, among others. Thus, the bacterial resistance is difficult to achieve, as it involves simultaneous mutations in different genes (Zaidi et al., 2017; Slavin et al., 2017). Nanoparticles also cause a change in typical negative charge in the membrane cell leading to an increase in the permeability cell (Leung et al., 2014; McQuillan et al., 2014; Thill et al., 2006).
Metal nanoparticles (NPs) made from silver (AgNPs), gold (AuNPs) and zinc oxide (ZnONPs) have shown their effectiveness as antimicrobial agents. The efficacy of all these NPs in reducing biofilm formation as well as the production of certain virulence factors in Pseudomonas has been demonstrated (Ali et al., 2017; Singh et al., 2015, Samanta et al., 2017; García-Lara et al., 2015). Furthermore, they exhibited broad-spectrum bactericidal against other pathogens like S. aureus, E. coli, K. pneumoniae (Hayat et al., 2019). For AgNPs, Ali et al. (2017) determined that this can affect the biosynthesis of signaling molecules by blocking the LasI/RhlI synthase. Meanwhile, for AuNPs and ZnONPs the mechanism of their QQ function has not been clarified.
Even when ZnO is recognized as GRAS (‘generally recognized as safe’) for FDA in USA (Hayat et al., 2019), for most of the metal and metal oxide NPs toxicity issues as circulatory, neurological, hepatotoxicity (Heydrnejad et al., 2015; Ramadi et al., 2016), nephrotoxicity (Pujalté et al., 2011), and lungs problems have been described ( Tsuji et al., 2006; De Jong and Borm 2008; Zimper et al., 2010), which is a serious disadvantage. The ROS generation could be the main contributor of toxicological effects (Hayat et al., 2019).
Chitosan NPs have also been studied for their broad-spectrum antimicrobial activity with a greater selective toxicity compared with metal NPs (Hayat et al., 2019). Chitosan, a polysaccharide obtained from natural sources (Vilar Junior et al., 2016), have also been shown antifungal and antitumor activity. Vadekeetil et al., (2019) demonstrated that the chitosan NPs had an excellent in vitro antivirulence activity effect against both virulence factors and biofilm formation in P. aeruginosa. In addition, Muslin et al. (2018), also proved that chitosan as polysaccharide caused a drop in production of protease and pyocyanin, as well as the expressions of lasR and rhlR genes in P. aeruginosa.
For antibiotics, azithromycin, ciprofloxacin, ceftazidime, erythromycin and tobramycin among others, shown be effective as QSIs for reducing the expressions of virulence factors in P. aeruginosa (Bala et al., 2011; Tateda et al., 2001; Sofer et al., 1999; Goh et al., 2002; Skindersoe et al., 2008).
Several reports in this area revealed that azithromycin significantly inhibits the production of QS signals. Bala et al. (2011) demonstrated that azithromycin sub-MIC concentrations affect swimming, swarming and biofilm formation in vitro in Pseudomonas. In addition, the therapeutic evaluation of azithromycin was also tested in a urinary tract infection model. The clearance of Pseudomonas from the mouse kidneys was observed to highlight the potential effectiveness of azithromycin as QSI against P. aeruginosa. Tateda et al. (2001) showed that azithromycin also reduces the C4HSL and 3OC12HSL production in P. aeruginosa. For erythromycin, Sofer et al. (1999) reported a similar situation with a drop in AHL production in P. aeruginosa strain.
Khan et al. (2020a) and Khan et al. (2020b) described that several aminoglycoside antibiotics commonly used, exhibited biofilm inhibition by targeting the QS regulatory protein LasR in P. aeruginosa.
- Combination therapy for treatment of Pseudomonas infections
Even though some QQ activity compounds as QSIs, QQE and NPs, have proved to be effective either in vitro or in vivo tests, their exclusive use may not be enough to fight against P. aeruginosa infections.
The combined and synergistic use of QQ activity compounds has become an attractive alternative and the focus of attention in recent years. The fact that different compounds exert their action at different QS levels on the pathogen, it ensures to a large extent the efficacy of the treatment. In addition, combination therapy could prevent the emergence of resistance to a single QQ compound.
Despite the effort to try to replace traditional antibiotics, their use remains essential and, in fact, they are still used even in combination therapies with QQ compounds. It should also be considered that, since biofilm formation blocks or restricts the penetration and efficacy of traditional antibiotics, the addition of a compound with QQ activity that acts by blocking biofilm formation provides an advantage to the treatment against the pathogen. Thus, Nazari et al. (2012); Roudashti et al. (2017), Bahari et al. (2017); Li et al. (2017) and Chanda et al. (2017), reported the use of different antibiotics in combination with several QQ compounds against Pseudomonas.
Recently, a novel AHL acylase, MacQ, has been identified from a multidrug- resistant bacterium, Acidovorax sp. strain MR-S7, and it was able to degrade a wide variety of AHLs, ranging from C6 to C14 side chains with or without 3-oxo substitutions, thus interfering with the QS system in the bacterial pathogen (Kusada et al., 2017).
References
- Ahmed, S. A., Rudden, M., Smyth, T. J., Dooley, J. S., Marchant, R., & Banat, I. M. (2019). Natural quorum sensing inhibitors effectively downregulate gene expression of Pseudomonas aeruginosa virulence factors. Applied microbiology and biotechnology, 103, 3521-3535. [CrossRef]
- Ali, S. G., Ansari, M. A., Sajid Jamal, Q. M., Khan, H. M., Jalal, M., Ahmad, H., & Mahdi, A. A. (2017). Antiquorum sensing activity of silver nanoparticles in P. aeruginosa: an in silico study. In Silico Pharmacology, 5, 1-7. [CrossRef]
- Allen, L., Dockrell, D. H., Pattery, T., Lee, D. G., Cornelis, P., Hellewell, P. G., & Whyte, M. K. (2005). Pyocyanin production by Pseudomonas aeruginosa induces neutrophil apoptosis and impairs neutrophil-mediated host defenses in vivo. The Journal of Immunology, 174(6), 3643-3649. [CrossRef]
- Anandan, K., & Vittal, R. R. (2019). Quorum quenching activity of AiiA lactonase KMMI17 from endophytic Bacillus thuringiensis KMCL07 on AHL-mediated pathogenic phenotype in Pseudomonas aeruginosa. Microbial pathogenesis, 132, 230-242. [CrossRef]
- Arranz San Martín, A., Vogel, J., Wullich, S. C., Quax, W. J., & Fetzner, S. (2022). Enzyme-mediated quenching of the Pseudomonas quinolone signal (PQS): A comparison between naturally occurring and engineered PQS-cleaving dioxygenases. Biomolecules, 12(2), 170. [CrossRef]
- Aybey, A., & Demirkan, E. (2016). Inhibition of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa by human serum paraoxonase. Journal of medical microbiology, 65(2), 105-113. [CrossRef]
- Bahari, S., Zeighami, H., Mirshahabi, H., Roudashti, S., & Haghi, F. (2017). Inhibition of Pseudomonas aeruginosa quorum sensing by subinhibitory concentrations of curcumin with gentamicin and azithromycin. Journal of global antimicrobial resistance, 10, 21-28. [CrossRef]
- Bala, A., Kumar, R., & Harjai, K. (2011). Inhibition of quorum sensing in Pseudomonas aeruginosa by azithromycin and its effectiveness in urinary tract infections. Journal of medical microbiology, 60(3), 300-306. [CrossRef]
- Bar-Rogovsky, H., Hugenmatter, A., & Tawfik, D. S. (2013). The evolutionary origins of detoxifying enzymes: the mammalian serum paraoxonases (PONs) relate to bacterial homoserine lactonases. Journal of Biological Chemistry, 288(33), 23914-23927.
- Bijtenhoorn, P., Mayerhofer, H., Müller-Dieckmann, J., Utpatel, C., Schipper, C., Hornung, C., ... & Streit, W. R. (2011). A novel metagenomic short-chain dehydrogenase/reductase attenuates Pseudomonas aeruginosa biofilm formation and virulence on Caenorhabditis elegans. PloS one, 6(10), e26278. [CrossRef]
- Birmes, F. S., Wolf, T., Kohl, T. A., Rüger, K., Bange, F., Kalinowski, J., & Fetzner, S. (2017). Mycobacterium abscessus subsp. abscessus is capable of degrading Pseudomonas aeruginosa quinolone signals. Frontiers in Microbiology, 8, 339. [CrossRef]
- Bobadilla Fazzini, R. A., Skindersoe, M. E., Bielecki, P., Puchałka, J., Givskov, M., & Martins dos Santos, V. A. (2013). Protoanemonin: a natural quorum sensing inhibitor that selectively activates iron starvation response. Environmental Microbiology, 15(1), 111-120. [CrossRef]
- Bodini, S. F., Manfredini, S., Epp, M., Valentini, S., & Santori, F. (2009). Quorum sensing inhibition activity of garlic extract and p-coumaric acid. Letters in applied microbiology, 49(5), 551-555.
- Buroni, S., Scoffone, V. C., Fumagalli, M., Makarov, V., Cagnone, M., Trespidi, G., ... & Chiarelli, L. R. (2018). Investigating the mechanism of action of diketopiperazines inhibitors of the Burkholderia cenocepacia quorum sensing synthase CepI: a site-directed mutagenesis study. Frontiers in Pharmacology, 9, 400679. [CrossRef]
- Chadha, J., Harjai, K., & Chhibber, S. (2022). Revisiting the virulence hallmarks of Pseudomonas aeruginosa: a chronicle through the perspective of quorum sensing. Environmental microbiology, 24(6), 2630-2656 . [CrossRef]
- Chanda, W., Joseph, T. P., Padhiar, A. A., Guo, X., Min, L., Wang, W., ... & Zhong, M. (2017). Combined effect of linolenic acid and tobramycin on Pseudomonas aeruginosa biofilm formation and quorum sensing. Experimental and therapeutic medicine, 14(5), 4328-4338. [CrossRef]
- Choo, J. H., Rukayadi, Y., & Hwang, J. K. (2006). Inhibition of bacterial quorum sensing by vanilla extract. Letters in applied microbiology, 42(6), 637-641. [CrossRef]
- Chow, J. Y., Xue, B., Lee, K. H., Tung, A., Wu, L., Robinson, R. C., & Yew, W. S. (2010). Directed evolution of a thermostable quorum-quenching lactonase from the amidohydrolase superfamily. Journal of biological chemistry, 285(52), 40911-40920. [CrossRef]
- Chowdhary, P. K., Keshavan, N., Nguyen, H. Q., Peterson, J. A., González, J. E., & Haines, D. C. (2007). Bacillus megaterium CYP102A1 oxidation of acyl homoserine lactones and acyl homoserines. Biochemistry, 46(50), 14429-14437. [CrossRef]
- Cugini, C., Calfee, M. W., Farrow III, J. M., Morales, D. K., Pesci, E. C., & Hogan, D. A. (2007). Farnesol, a common sesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Molecular microbiology, 65(4), 896-906.
- Czajkowski, R., Krzyżanowska, D., Karczewska, J., Atkinson, S., Przysowa, J., Lojkowska, E., ... & Jafra, S. (2011). Inactivation of AHLs by Ochrobactrum sp. A44 depends on the activity of a novel class of AHL acylase. Environmental microbiology reports, 3(1), 59-68. [CrossRef]
- D'almeida, R. E., Molina, R. D. I., Viola, C. M., Luciardi, M. C., Peñalver, C. N., Bardón, A., & Arena, M. E. (2017). Comparison of seven structurally related coumarins on the inhibition of quorum sensing of Pseudomonas aeruginosa and Chromobacterium violaceum. Bioorganic chemistry, 73, 37-42. [CrossRef]
- De Jong, W. H., & Borm, P. J. (2008). Drug delivery and nanoparticles: applications and hazards. International journal of nanomedicine, 3(2), 133-149.
- Díaz, M. A., González, S. N., Alberto, M. R., & Arena, M. E. (2020). Human probiotic bacteria attenuate Pseudomonas aeruginosa biofilm and virulence by quorum-sensing inhibition. Biofouling, 36(5), 597-609.
- Dong, W., Zhu, J., Guo, X., Kong, D., Zhang, Q., Zhou, Y., ... & Ruan, Z. (2018). Characterization of AiiK, an AHL lactonase, from Kurthia huakui LAM0618T and its application in quorum quenching on Pseudomonas aeruginosa PAO1. Scientific reports, 8(1), 6013. [CrossRef]
- Dong, Y. H., Xu, J. L., Li, X. Z., & Zhang, L. H. (2000). AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proceedings of the National Academy of Sciences, 97(7), 3526-3531.
- Faisal, A. J., Said, L. A., & Ali, M. R. (2021). Quorum quenching effect of recombinant Paraoxonase-1 enzyme against quorum sensing genes produced from Pseudomonas aeruginosa. Gene Reports, 25, 101412. [CrossRef]
- Fan, X., Liang, M., Wang, L., Chen, R., Li, H., & Liu, X. (2017). Aii810, a novel cold-adapted N-acylhomoserine lactonase discovered in a metagenome, can strongly attenuate Pseudomonas aeruginosa virulence factors and biofilm formation. Frontiers in microbiology, 8, 1950. [CrossRef]
- Fernandes, R., Roy, V., Wu, H. C., & Bentley, W. E. (2010). Engineered biological nanofactories trigger quorum sensing response in targeted bacteria. Nature nanotechnology, 5(3), 213-217. [CrossRef]
- Fetzner, S. (2015). Quorum quenching enzymes. Journal of biotechnology, 201, 2-14. [CrossRef]
- Gallagher, L. A., & Manoil, C. (2001). Pseudomonas aeruginosa PAO1 kills Caenorhabditis elegans by cyanide poisoning. Journal of bacteriology, 183(21), 6207-6214. [CrossRef]
- García-Lara, B., Saucedo-Mora, M. Á., Roldán-Sánchez, J. A., Pérez-Eretza, B., Ramasamy, M., Lee, J., ... & García-Contreras, R. (2015). Inhibition of quorum-sensing-dependent virulence factors and biofilm formation of clinical and environmental Pseudomonas aeruginosa strains by ZnO nanoparticles. Letters in applied microbiology, 61(3), 299-305. [CrossRef]
- García-Reyes, S., Soberón-Chávez, G., & Cocotl-Yanez, M. (2020). The third quorum-sensing system of Pseudomonas aeruginosa: Pseudomonas quinolone signal and the enigmatic PqsE protein. Journal of medical microbiology, 69(1), 25-34.
- Ghafoor, A., Hay, I. D., & Rehm, B. H. (2011). Role of exopolysaccharides in Pseudomonas aeruginosa biofilm formation and architecture. Applied and environmental microbiology, 77(15), 5238-5246. [CrossRef]
- Girennavar, B., Cepeda, M. L., Soni, K. A., Vikram, A., Jesudhasan, P., Jayaprakasha, G. K., ... & Patil, B. S. (2008). Grapefruit juice and its furocoumarins inhibits autoinducer signaling and biofilm formation in bacteria. International journal of food microbiology, 125(2), 204-208. [CrossRef]
- Goh, E. B., Yim, G., Tsui, W., McClure, J., Surette, M. G., & Davies, J. (2002). Transcriptional modulation of bacterial gene expression by subinhibitory concentrations of antibiotics. Proceedings of the National Academy of Sciences, 99(26), 17025-17030. [CrossRef]
- Grandclément, C., Tannières, M., Moréra, S., Dessaux, Y., & Faure, D. (2016). Quorum quenching: role in nature and applied developments. FEMS microbiology reviews, 40(1), 86-116. [CrossRef]
- Hall, S., McDermott, C., Anoopkumar-Dukie, S., McFarland, A. J., Forbes, A., Perkins, A. V., & Grant, G. D. (2016). Cellular effects of pyocyanin, a secreted virulence factor of Pseudomonas aeruginosa. Toxins, 8(8), 236.
- Hemmati, F., Salehi, R., Ghotaslou, R., Samadi Kafil, H., Hasani, A., Gholizadeh, P., ... & Ahangarzadeh Rezaee, M. (2020). Quorum quenching: A potential target for antipseudomonal therapy. Infection and Drug Resistance, 2989-3005. [CrossRef]
- Hentzer, M., Wu, H., Andersen, J. B., Riedel, K., Rasmussen, T. B., Bagge, N., ... & Givskov, M. (2003). Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. The EMBO journal, 22(15), 3803-3815. [CrossRef]
- Heydrnejad, M. S., Samani, R. J., & Aghaeivanda, S. (2015). Toxic effects of silver nanoparticles on liver and some hematological parameters in male and female mice (Mus musculus). Biological trace element research, 165, 153-158. [CrossRef]
- Hiblot, J., Bzdrenga, J., Champion, C., Chabriere, E., & Elias, M. (2015). Crystal structure of Vmo lac, a tentative quorum quenching lactonase from the extremophilic crenarchaeon Vulcanisaeta moutnovskia. Scientific reports, 5(1), 8372. [CrossRef]
- Holden, M. T., Ram Chhabra, S., De Nys, R., Stead, P., Bainton, N. J., Hill, P. J., & Williams, P. (1999). Quorum-sensing cross talk: isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Molecular microbiology, 33(6), 1254-1266. [CrossRef]
- Hraiech, S., Hiblot, J., Lafleur, J., Lepidi, H., Papazian, L., Rolain, J. M., & Chabriere, E. (2014). Inhaled lactonase reduces Pseudomonas aeruginosa quorum sensing and mortality in rat pneumonia. PLoS One, 9(10), e107125. [CrossRef]
- Husain, F. M., Ahmad, I., Asif, M., & Tahseen, Q. (2013). Influence of clove oil on certain quorum-sensing-regulated functions and biofilm of Pseudomonas aeruginosa and Aeromonas hydrophila. Journal of biosciences, 38, 835-844. [CrossRef]
- Jakobsen, T. H., Bragason, S. K., Phipps, R. K., Christensen, L. D., van Gennip, M., Alhede, M., ... & Givskov, M. (2012). Food as a source for quorum sensing inhibitors: iberin from horseradish revealed as a quorum sensing inhibitor of Pseudomonas aeruginosa. Applied and environmental microbiology, 78(7), 2410-2421. [CrossRef]
- Jensen, V., Löns, D., Zaoui, C., Bredenbruch, F., Meissner, A., Dieterich, G., ... & Häussler, S. (2006). RhlR expression in Pseudomonas aeruginosa is modulated by the Pseudomonas quinolone signal via PhoB-dependent and-independent pathways. Journal of bacteriology, 188(24), 8601-8606.
- Kalia, V. C., Patel, S. K., Kang, Y. C., & Lee, J. K. (2019). Quorum sensing inhibitors as antipathogens: biotechnological applications. Biotechnology advances, 37(1), 68-90.
- Kang, D., Revtovich, A. V., Chen, Q., Shah, K. N., Cannon, C. L., & Kirienko, N. V. (2019). Pyoverdine-dependent virulence of Pseudomonas aeruginosa isolates from cystic fibrosis patients. Frontiers in microbiology, 10, 2048. [CrossRef]
- Kariyawasam, R. M., Julien, D. A., Jelinski, D. C., Larose, S. L., Rennert-May, E., Conly, J. M., ... & Barkema, H. W. (2022). Antimicrobial resistance (AMR) in COVID-19 patients: a systematic review and meta-analysis (November 2019–June 2021). Antimicrobial Resistance & Infection Control, 11(1), 45. [CrossRef]
- Khan, F., Lee, J. W., Javaid, A., Park, S. K., & Kim, Y. M. (2020). Inhibition of biofilm and virulence properties of Pseudomonas aeruginosa by sub-inhibitory concentrations of aminoglycosides. Microbial Pathogenesis, 146, 104249. [CrossRef]
- Khan, F., Pham, D. T. N., & Kim, Y. M. (2020). Alternative strategies for the application of aminoglycoside antibiotics against the biofilm-forming human pathogenic bacteria. Applied microbiology and biotechnology, 104, 1955-1976. [CrossRef]
- Khan, M. S. A., Zahin, M., Hasan, S., Husain, F. M., & Ahmad, I. (2009). Inhibition of quorum sensing regulated bacterial functions by plant essential oils with special reference to clove oil. Letters in applied microbiology, 49(3), 354-360. [CrossRef]
- Kim, H. S., Lee, S. H., Byun, Y., & Park, H. D. (2015). 6-Gingerol reduces Pseudomonas aeruginosa biofilm formation and virulence via quorum sensing inhibition. Scientific reports, 5(1), 8656. [CrossRef]
- Kiran, S., Sharma, P., Harjai, K., & Capalash, N. (2011). Enzymatic quorum quenching increases antibiotic susceptibility of multidrug resistant Pseudomonas aeruginosa. Iranian Journal of Microbiology, 3(1), 1.
- Kusada, H., Tamaki, H., Kamagata, Y., Hanada, S., & Kimura, N. (2017). A novel quorum-quenching N-acylhomoserine lactone acylase from Acidovorax sp. strain MR-S7 mediates antibiotic resistance. Applied and Environmental Microbiology, 83(13), e00080-17.
- LaSarre, B., & Federle, M. J. (2013). Exploiting quorum sensing to confuse bacterial pathogens. Microbiology and molecular biology reviews, 77(1), 73-111. [CrossRef]
- Last, D., Krüger, G. H., Dörr, M., & Bornscheuer, U. T. (2016). Fast, continuous, and high-throughput (bio) chemical activity assay for N-acyl-L-homoserine lactone quorum-quenching enzymes. Applied and Environmental Microbiology, 82(14), 4145-4154.
- Leadbetter, J. R., & Greenberg, E. P. (2000). Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. Journal of bacteriology, 182(24), 6921-6926. [CrossRef]
- Lee, J. H., Kim, N. H., Jang, K. M., Jin, H., Shin, K., Jeong, B. C., ... & Lee, S. H. (2023). Prioritization of Critical Factors for Surveillance of the Dissemination of Antibiotic Resistance in Pseudomonas aeruginosa: A Systematic Review. International Journal of Molecular Sciences, 24(20), 15209. [CrossRef]
- Leung, Y. H., Ng, A. M., Xu, X., Shen, Z., Gethings, L. A., Wong, M. T., ... & Leung, F. C. (2014). Mechanisms of antibacterial activity of MgO: non-ROS mediated toxicity of MgO nanoparticles towards Escherichia coli. Small, 10(6), 1171-1183. [CrossRef]
- Li, Y., Huang, J., Li, L., & Liu, L. (2017). Synergistic activity of berberine with azithromycin against Pseudomonas aeruginosa isolated from patients with cystic fibrosis of lung in vitro and in vivo. Cellular Physiology and Biochemistry, 42(4), 1657-1669. [CrossRef]
- Liao, C., Huang, X., Wang, Q., Yao, D., & Lu, W. (2022). Virulence factors of Pseudomonas aeruginosa and antivirulence strategies to combat its drug resistance. Frontiers in cellular and infection microbiology, 12, 926758. [CrossRef]
- Lin, Y. H., Xu, J. L., Hu, J., Wang, L. H., Ong, S. L., Leadbetter, J. R., & Zhang, L. H. (2003). Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Molecular microbiology, 47(3), 849-860.
- López-Jácome, L. E., Garza-Ramos, G., Hernández-Durán, M., Franco-Cendejas, R., Loarca, D., Romero-Martínez, D., ... & García-Contreras, R. (2019). AiiM lactonase strongly reduces quorum sensing controlled virulence factors in clinical strains of Pseudomonas aeruginosa isolated from burned patients. Frontiers in microbiology, 10, 2657. [CrossRef]
- Malešević, M., Stanisavljević, N., Novović, K., Polović, N., Vasiljević, Z., Kojić, M., & Jovčić, B. (2020). Burkholderia cepacia YtnP and Y2-aiiA lactonases inhibit virulence of Pseudomonas aeruginosa via quorum quenching activity. Microbial Pathogenesis, 149, 104561. [CrossRef]
- Martinelli, D., Grossmann, G., Séquin, U., Brandl, H., & Bachofen, R. (2004). Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC microbiology, 4, 1-10.
- McClure, C. D., & Schiller, N. L. (1996). Inhibition of macrophage phagocytosis by Pseudomonas aeruginosa rhamnolipids in vitro and in vivo. Current microbiology, 33, 109-117. [CrossRef]
- McQuillan, J. S., Groenaga Infante, H., Stokes, E., & Shaw, A. M. (2012). Silver nanoparticle enhanced silver ion stress response in Escherichia coli K12. Nanotoxicology, 6(8), 857-866. [CrossRef]
- Meyer, J. M., Neely, A., Stintzi, A., Georges, C., & Holder, I. A. (1996). Pyoverdin is essential for virulence of Pseudomonas aeruginosa. Infection and immunity, 64(2), 518-523. [CrossRef]
- Müller, C., Birmes, F. S., Rückert, C., Kalinowski, J., & Fetzner, S. (2015). Rhodococcus erythropolis BG43 genes mediating Pseudomonas aeruginosa quinolone signal degradation and virulence factor attenuation. Applied and Environmental Microbiology, 81(22), 7720-7729. [CrossRef]
- Muslim, S. N., Kadmy, I. M. A., Ali, A. N. M., Salman, B. K., Ahmad, M., Khazaal, S. S., ... & Muslim, S. N. (2018). Chitosan extracted from Aspergillus flavus shows synergistic effect, eases quorum sensing mediated virulence factors and biofilm against nosocomial pathogen Pseudomonas aeruginosa. International journal of biological macromolecules, 107, 52-58.
- Nafee, N., Husari, A., Maurer, C. K., Lu, C., de Rossi, C., Steinbach, A., ... & Schneider, M. (2014). Antibiotic-free nanotherapeutics: ultra-small, mucus-penetrating solid lipid nanoparticles enhance the pulmonary delivery and anti-virulence efficacy of novel quorum sensing inhibitors. Journal of Controlled Release, 192, 131-140. [CrossRef]
- Nazari, Z. E., Banoee, M., Sepahi, A. A., Rafii, F., & Shahverdi, A. R. (2012). The combination effects of trivalent gold ions and gold nanoparticles with different antibiotics against resistant Pseudomonas aeruginosa. Gold Bulletin, 45, 53-59. [CrossRef]
- Ng, F. S., Wright, D. M., & Seah, S. Y. (2011). Characterization of a phosphotriesterase-like lactonase from Sulfolobus solfataricus and its immobilization for disruption of quorum sensing. Applied and environmental microbiology, 77(4), 1181-1186. [CrossRef]
- Nithya, C., Devi, M. G., & Karutha Pandian, S. (2011). A novel compound from the marine bacterium Bacillus pumilus S6-15 inhibits biofilm formation in gram-positive and gram-negative species. Biofouling, 27(5), 519-528.
- Oliver, A., Cantón, R., Campo, P., Baquero, F., & Blázquez, J. (2000). High frequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lung infection. Science, 288(5469), 1251-1253. [CrossRef]
- Ozer, E. A., Pezzulo, A., Shih, D. M., Chun, C., Furlong, C., Lusis, A. J., ... & Zabner, J. (2005). Human and murine paraoxonase 1 are host modulators of Pseudomonas aeruginosa quorum-sensing. FEMS microbiology letters, 253(1), 29-37. [CrossRef]
- Papenfort, K., & Bassler, B. L. (2016). Quorum sensing signal–response systems in Gram-negative bacteria. Nature Reviews Microbiology, 14(9), 576-588.
- Pujalté, I., Passagne, I., Brouillaud, B., Tréguer, M., Durand, E., Ohayon-Courtès, C., & L'Azou, B. (2011). Cytotoxicity and oxidative stress induced by different metallic nanoparticles on human kidney cells. Particle and fibre toxicology, 8, 1-16. [CrossRef]
- Pustelny, C., Albers, A., Büldt-Karentzopoulos, K., Parschat, K., Chhabra, S. R., Cámara, M., ... & Fetzner, S. (2009). Dioxygenase-mediated quenching of quinolone-dependent quorum sensing in Pseudomonas aeruginosa. Chemistry & biology, 16(12), 1259-1267. [CrossRef]
- Qais, F. A., Khan, M. S., & Ahmad, I. (2018). Nanoparticles as quorum sensing inhibitor: Prospects and limitations. Biotechnological applications of quorum sensing inhibitors, 227-244.
- Qin, S., Xiao, W., Zhou, C., Pu, Q., Deng, X., Lan, L., ... & Wu, M. (2022). Pseudomonas aeruginosa: pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal transduction and targeted therapy, 7(1), 199. [CrossRef]
- Raafat, M. M., Ali-Tammam, M., & Ali, A. E. (2019). Quorum quenching activity of Bacillus cereus isolate 30b confers antipathogenic effects in Pseudomonas aeruginosa. Infection and Drug Resistance, 1583-1596. [CrossRef]
- Rajkumari, J., Borkotoky, S., Murali, A., Suchiang, K., Mohanty, S. K., & Busi, S. (2018). Cinnamic acid attenuates quorum sensing associated virulence factors and biofilm formation in Pseudomonas aeruginosa PAO1. Biotechnology letters, 40, 1087-1100. [CrossRef]
- Ramadi, K. B., Mohamed, Y. A., Al-Sbiei, A., Almarzooqi, S., Bashir, G., Al Dhanhani, A., ... & Al-Ramadi, B. K. (2016). Acute systemic exposure to silver-based nanoparticles induces hepatotoxicity and NLRP3-dependent inflammation. Nanotoxicology, 10(8), 1061-1074. [CrossRef]
- Rasch, M., Buch, C., Austin, B., Slierendrecht, W. J., Ekmann, K. S., Larsen, J. L., ... & Gram, L. (2004). An inhibitor of bacterial quorum sensing reduces mortalities caused by vibriosis in rainbow trout (Oncorhynchus mykiss, Walbaum). Systematic and Applied Microbiology, 27(3), 350-359 . [CrossRef]
- Rasmussen, T. B., Skindersoe, M. E., Bjarnsholt, T., Phipps, R. K., Christensen, K. B., Jensen, P. O., ... & Givskov, M. (2005). Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology, 151(5), 1325-1340. [CrossRef]
- Rather, M. A., Saha, D., Bhuyan, S., Jha, A. N., & Mandal, M. (2022). Quorum quenching: a drug discovery approach against Pseudomonas aeruginosa. Microbiological Research, 264, 127173. [CrossRef]
- Reimmann, C., Ginet, N., Michel, L., Keel, C., Michaux, P., Krishnapillai, V., ... & Haas, D. (2002). Genetically programmed autoinducer destruction reduces virulence gene expression and swarming motility in Pseudomonas aeruginosa PAO1. Microbiology, 148(4), 923-932 . [CrossRef]
- Reina, J. C., Romero, M., Salto, R., Cámara, M., & Llamas, I. (2021). AhaP, A quorum quenching acylase from Psychrobacter sp. M9-54-1 that attenuates Pseudomonas aeruginosa and Vibrio coralliilyticus virulence. Marine drugs, 19(1), 16.
- Roudashti, S., Zeighami, H., Mirshahabi, H., Bahari, S., Soltani, A., & Haghi, F. (2017). Synergistic activity of sub-inhibitory concentrations of curcumin with ceftazidime and ciprofloxacin against Pseudomonas aeruginosa quorum sensing related genes and virulence traits. World Journal of Microbiology and Biotechnology, 33, 1-8. [CrossRef]
- Rudrappa, T., & Bais, H. P. (2008). Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. Journal of agricultural and food chemistry, 56(6), 1955-1962. [CrossRef]
- Ryu, D. H., Lee, S. W., Mikolaityte, V., Kim, Y. W., Jeong, H., Lee, S. J., ... & Lee, J. K. (2020). Identification of a second type of AHL-lactonase from Rhodococcus sp. BH4, belonging to the α/β hydrolase superfamily. Journal of Microbiology and Biotechnology, 30(6), 937.
- Salim, S. A., Mohan, M. S., Forgia, N., & Busi, S. (2024). Medical Importance of ESKAPE Pathogens. In ESKAPE Pathogens: Detection, Mechanisms and Treatment Strategies (pp. 1-32). Singapore: Springer Nature Singapore.
- Samanta, S., Singh, B. R., & Adholeya, A. (2017). Intracellular synthesis of gold nanoparticles using an ectomycorrhizal strain EM-1083 of Laccaria fraterna and its nanoanti-quorum sensing potential against Pseudomonas aeruginosa. Indian journal of microbiology, 57, 448-460. [CrossRef]
- Sikdar, R., & Elias, M. (2020). Quorum quenching enzymes and their effects on virulence, biofilm, and microbiomes: a review of recent advances. Expert review of anti-infective therapy, 18(12), 1221-1233. [CrossRef]
- Singh, R., Shedbalkar, U. U., Wadhwani, S. A., & Chopade, B. A. (2015). Bacteriagenic silver nanoparticles: synthesis, mechanism, and applications. Applied microbiology and biotechnology, 99, 4579-4593. [CrossRef]
- Skindersoe, M. E., Alhede, M., Phipps, R., Yang, L., Jensen, P. O., Rasmussen, T. B., ... & Givskov, M. (2008). Effects of antibiotics on quorum sensing in Pseudomonas aeruginosa. Antimicrobial agents and chemotherapy, 52(10), 3648-3663. [CrossRef]
- Slavin, Y. N., Asnis, J., Hńfeli, U. O., & Bach, H. (2017). Metal nanoparticles: understanding the mechanisms behind antibacterial activity. Journal of nanobiotechnology, 15, 1-20. [CrossRef]
- Sofer, D., Gilboa-Garber, N., Belz, A., & Garber, N. C. (1999). ‘Subinhibitory’erythromycin represses production of Pseudomonas aeruginosa lectins, autoinducer and virulence factors. Chemotherapy, 45(5), 335-341.
- Song, Z., Kong, K. F., Wu, H., Maricic, N., Ramalingam, B., Priestap, H., ... & Mathee, K. (2010). Panax ginseng has anti-infective activity against opportunistic pathogen Pseudomonas aeruginosa by inhibiting quorum sensing, a bacterial communication process critical for establishing infection. Phytomedicine, 17(13), 1040-1046. [CrossRef]
- Tang, K. W. K., Millar, B. C., & Moore, J. E. (2023). Antimicrobial resistance (AMR). British Journal of Biomedical Science, 80, 11387.
- Tang, K., Su, Y., Brackman, G., Cui, F., Zhang, Y., Shi, X., ... & Zhang, X. H. (2015). MomL, a novel marine-derived N-acyl homoserine lactonase from Muricauda olearia. Applied and environmental microbiology, 81(2), 774-782. [CrossRef]
- Tateda, K., Comte, R., Pechere, J. C., Köhler, T., Yamaguchi, K., & Van Delden, C. (2001). Azithromycin inhibits quorum sensing in Pseudomonas aeruginosa. Antimicrobial agents and chemotherapy, 45(6), 1930-1933. [CrossRef]
- Thi, M. T. T., Wibowo, D., & Rehm, B. H. (2020). Pseudomonas aeruginosa biofilms. International journal of molecular sciences, 21(22), 8671.
- Thill, A., Zeyons, O., Spalla, O., Chauvat, F., Rose, J., Auffan, M., & Flank, A. M. (2006). Cytotoxicity of CeO2 nanoparticles for Escherichia coli. Physico-chemical insight of the cytotoxicity mechanism. Environmental science & technology, 40(19), 6151-6156.
- Tielen, P., Narten, M., Rosin, N., Biegler, I., Haddad, I., Hogardt, M., ... & Jahn, D. (2011). Genotypic and phenotypic characterization of Pseudomonas aeruginosa isolates from urinary tract infections. International Journal of Medical Microbiology, 301(4), 282-292. [CrossRef]
- Tigabu, A., & Getaneh, A. L. E. M. (2021). Staphylococcus aureus, ESKAPE Bacteria Challenging Current Health Care and Community Settings: a Literature Review. Clinical Laboratory, (7). [CrossRef]
- Tinh, N. T. N., Linh, N. D., Wood, T. K., Dierckens, K., Sorgeloos, P., & Bossier, P. (2007). Interference with the quorum sensing systems in a Vibrio harveyi strain alters the growth rate of gnotobiotically cultured rotifer Brachionus plicatilis. Journal of applied microbiology, 103(1), 194-203. [CrossRef]
- Tsuji, J. S., Maynard, A. D., Howard, P. C., James, J. T., Lam, C. W., Warheit, D. B., & Santamaria, A. B. (2006). Research strategies for safety evaluation of nanomaterials, part IV: risk assessment of nanoparticles. Toxicological sciences, 89(1), 42-50. [CrossRef]
- Uroz, S., Chhabra, S. R., Camara, M., Williams, P., Oger, P., & Dessaux, Y. (2005). N-Acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology, 151(10), 3313-3322. [CrossRef]
- Vadekeetil, A., Chhibber, S., & Harjai, K. (2019). Efficacy of intravesical targeting of novel quorum sensing inhibitor nanoparticles against Pseudomonas aeruginosa biofilm-associated murine pyelonephritis. Journal of drug targeting, 27(9), 995-1003.
- Vattem, D. A., Mihalik, K., Crixell, S. H., & McLean, R. J. (2007). Dietary phytochemicals as quorum sensing inhibitors. Fitoterapia, 78(4), 302-310. [CrossRef]
- Velasco-Bucheli, R., Hormigo, D., Fernández-Lucas, J., Torres-Ayuso, P., Alfaro-Ureña, Y., Saborido, A. I., ... & de la Mata, I. (2020). Penicillin acylase from Streptomyces lavendulae and aculeacin A acylase from Actinoplanes utahensis: Two versatile enzymes as useful tools for quorum quenching processes. Catalysts, 10(7), 730. [CrossRef]
- Venturi, V. (2006). Regulation of quorum sensing in Pseudomonas. FEMS microbiology reviews, 30(2), 274-291. [CrossRef]
- Vikram, A., Jayaprakasha, G. K., Jesudhasan, P. R., Pillai, S. D., & Patil, B. S. (2010). Suppression of bacterial cell–cell signalling, biofilm formation and type III secretion system by citrus flavonoids. Journal of applied microbiology, 109(2), 515-527. [CrossRef]
- Vilar Junior, J. C., Ribeaux, D. R., Alves da Silva, C. A., Campos-Takaki, D., & Maria, G. (2016). Physicochemical and antibacterial properties of chitosan extracted from waste shrimp shells. International Journal of Microbiology, 2016.
- Wang, W. Z., Morohoshi, T., Someya, N., & Ikeda, T. (2012). AidC, a novel N-acylhomoserine lactonase from the potato root-associated cytophaga-flavobacteria-bacteroides (CFB) group bacterium Chryseobacterium sp. strain StRB126. Applied and environmental microbiology, 78(22), 7985-7992. [CrossRef]
- Wang, X., Gong, L., Liang, S., Han, X., Zhu, C., & Li, Y. (2005). Algicidal activity of rhamnolipid biosurfactants produced by Pseudomonas aeruginosa. Harmful algae, 4(2), 433-443. [CrossRef]
- Wang, W. Z., Morohoshi, T., Ikenoya, M., Someya, N., & Ikeda, T. (2010). AiiM, a novel class of N-acylhomoserine lactonase from the leaf-associated bacterium Microbacterium testaceum. Applied and environmental microbiology, 76(8), 2524-2530. [CrossRef]
- Whitesides, G. M. (2005). Nanoscience, nanotechnology, and chemistry. Small, 1(2), 172-179. [CrossRef]
- Williams, P., Winzer, K., Chan, W. C., & Camara, M. (2007). Look who's talking: communication and quorum sensing in the bacterial world. Philosophical Transactions of the Royal Society B: Biological Sciences, 362(1483), 1119-1134. [CrossRef]
- Wu, H., Song, Z., Hentzer, M., Andersen, J. B., Molin, S., Givskov, M., & Høiby, N. (2004). Synthetic furanones inhibit quorum-sensing and enhance bacterial clearance in Pseudomonas aeruginosa lung infection in mice. Journal of Antimicrobial Chemotherapy, 53(6), 1054-1061. [CrossRef]
- Wullich, S. C., Arranz San Martín, A., & Fetzner, S. (2020). An α/β-hydrolase fold subfamily comprising Pseudomonas quinolone signal-cleaving dioxygenases. Applied and Environmental Microbiology, 86(9), e00279-20. [CrossRef]
- Yates, E. A., Philipp, B., Buckley, C., Atkinson, S., Chhabra, S. R., Sockett, R. E., ... & Williams, P. (2002). N-acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infection and immunity, 70(10), 5635-5646. [CrossRef]
- Zaidi, S., Misba, L., & Khan, A. U. (2017). Nano-therapeutics: a revolution in infection control in post antibiotic era. Nanomedicine: Nanotechnology, Biology and Medicine, 13(7), 2281-2301. [CrossRef]
- Zhang, M., Wang, M., Zhu, X., Yu, W., & Gong, Q. (2018). Equisetin as potential quorum sensing inhibitor of Pseudomonas aeruginosa. Biotechnology letters, 40, 865-870. [CrossRef]
- Zhang, Y., Brackman, G., & Coenye, T. (2017). Pitfalls associated with evaluating enzymatic quorum quenching activity: the case of MomL and its effect on Pseudomonas aeruginosa and Acinetobacter baumannii biofilms. PeerJ, 5, e3251.
- Zhou, L., Zhang, Y., Ge, Y., Zhu, X., & Pan, J. (2020). Regulatory mechanisms and promising applications of quorum sensing-inhibiting agents in control of bacterial biofilm formation. Frontiers in microbiology, 11, 589640. [CrossRef]
- Zhu, X., Chen, W. J., Bhatt, K., Zhou, Z., Huang, Y., Zhang, L. H., ... & Wang, J. (2023). Innovative microbial disease biocontrol strategies mediated by quorum quenching and their multifaceted applications: A review. Frontiers in Plant Science, 13, 1063393. [CrossRef]
- Zimper, U., Aaltonen, J., McGoverin, C. M., Gordon, K. C., Krauel-Goellner, K., & Rades, T. (2010). Quantification of process induced disorder in milled samples using different analytical techniques. Pharmaceutics, 2(1), 30-49. [CrossRef]
Figure 1.
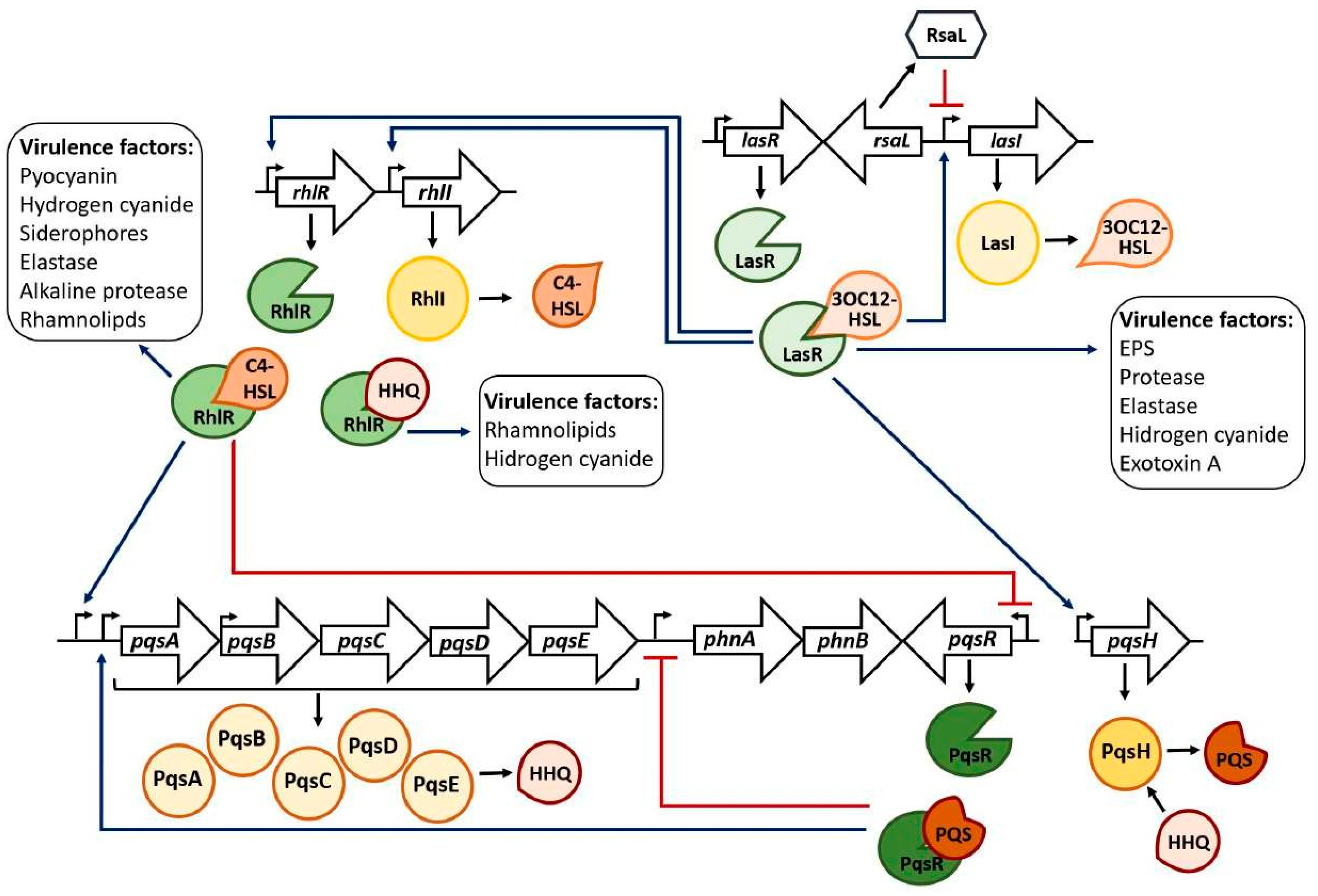
Figure 2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



