
Submitted:
06 May 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
Canine atopic dermatitis (CAD) is a common chronic allergic skin disease. In our previous study, we isolated two probiotic strains, Lactococcus cremoris subsp. cremoris MP01 and Lacticaseibacillus paracasei subsp. paracasei MP02, from Mongolian fermented milk. These strains demonstrated potential in alleviating symptoms similar to AD and reducing specific immunoglobulin E (IgE) levels in a mouse model of AD. Building upon this discovery, our current investigation sought to evaluate the therapeutic potential of these probiotics for treating CAD. We conducted an open-label, single-arm clinical trial where dogs diagnosed with AD were administered LCP capsules containing Lc. cremoris subsp. cremoris MP01 and L. paracasei subsp. paracasei MP02 once daily for 60 days. Stool, skin swab, and venous blood specimens were collected at three intervals: before, and after 30 and 60 days of LCP intervention. The skin and fecal microbiomes were analyzed using full-length 16S rRNA sequencing. After 60 days of LCP intervention, notable improvements were observed in skin lesions and scratching behaviors, accompanied by reductions in both the Canine Atopic Dermatitis Extent and Severity Index (CADESI) and the Pruritus Visual Analogue Scale (PVAS) scores. Additionally, LCP intervention resulted in significant reductions in IgE levels and modulation in type 1 T helper (Th1)/ type 2 T helper cells (Th2)-related cytokine secretion. Microbiota analysis unveiled changes in the composition of both skin and gut microbiotas, including reductions in Shigella flexneri on the skin and Romboutsia in the gut. Levels of short-chain fatty acid (SCFA) increased following the 60-days of LCP intervention. These effects were likely mediated by alterations in gut microbiota and the upregulation of SCFA levels, which subsequently reduced the secretion of allergy-related IgE. Collectively, these mechanisms contributed to alleviating CAD symptoms, regulating skin microbiota composition, and reducing the adherence and invasion of environmental microbes and pathogens. Overall, our findings highlight the promising therapeutic potential of the Lc. cremoris subsp. cremoris MP01 and L. paracasei subsp. paracasei MP02 mixture as an effective strategy for managing CAD in dogs.
Keywords:
Canine atopic dermatitis
; Skin microbiome
; Fecal microbiome
; Probiotics
; Allergy
1. Introduction
Canine atopic dermatitis (CAD) is a common chronic allergic skin disease in clinical practice, characterized by frequent itching and scratching. It is the second most prevalent type of allergic dermatitis in dogs after flea infections, with about 10–15% of global incidence. Current medications can only alleviate symptoms; however, this disease significantly impacts the quality of life for both dogs and their owners [1,2,3].
CAD is a type I hypersensitivity reaction, primarily driven by an imbalance of immune response between Type 1 T helper cells (Th1 cells) and Type 2 T helper cells (Th2 cells). When the host encounters antigens, the Th2 cell secrete cytokines such as interleukin (IL)-4, IL-5, IL-13, and IL-31, which stimulate B cells to release large amounts of immunoglobulin E (IgE). Upon re-exposure to allergens, the release of IL-3, IL-4, IL-5, and pro-inflammatory factors such as histamine and prostaglandin lead to increased vascular permeability and typical inflammatory responses such as redness, swelling, heat, and pain [4,5,6]. Repetitive scratching disrupts the epidermal barrier and tissue fluid exudation. Coupled with licking behaviors in dogs, this makes allergen penetration easier and facilitates the colonization of pathogens, which can even include commensal bacteria that typically prevent pathogen invasion [7,8,9].
The skin and gut share many similar functions, including a dense distribution of nerves and blood vessels, and play critical roles in the overall immune and endocrine systems, also serving as barriers against external environments [10,11]. Studies have shown that many skin diseases are accompanied by changes in the gut microbiota. Patients with atopic dermatitis exhibit reduced microbial diversity, decreased abundance of beneficial bacteria such as short-chain fatty acid producers, and an increased proportion of pathogenic bacteria [12,13,14]. Therefore, there is a significant correlation between gut health and skin, indicating mutual influence between dysbiosis of gut and skin microbiota and immune imbalance [15].
Numerous studies have indicated that probiotics can regulate gut microbiota and maintain the integrity of the gut epithelium, impacting overall immune balance by stimulating Th1 cells or modulating regulatory T cells (Treg) cytokine levels, thus improving allergic symptoms [16,17,18]. However, clinical trials investigating the role of probiotics in improving CAD by analyzing both gut and skin microbiota are still notably lacking. Previously, our laboratory used an AD mouse model to confirm that a mixed strain combination of Lactococcus cremoris subsp. cremoris and Lacticaseibacillus paracasei subsp. paracasei could reduce the ratio of Th1 and Th2 cells in the spleen and decrease serum IgE, showing positive effects on skin inflammation, epidermal proliferation, and skin damage [19]. Thus, this study evaluates the efficacy of the mixed strain combination on improving CAD clinical outcomes, and integrates gut and skin microbiome analysis to explore the impacts of probiotics on the pathophysiology of CAD.
2. Results
2.1. Administrating LCP Reduced the Symptoms of Atopic Dermatitis in Dogs via Modulating the Immune System
After 60 days of LCP treatment, most tested dogs showed a significant improvement in skin symptoms, characterized by reduced erythema and shedding (Figure 1B). Assessment using CADESI and PVAS scores revealed that up to 75% of the dogs experienced lower severity scores (Figure 1C).
Total IgE levels, a crucial indicator of atopic dermatitis, showed a significant decrease after LCP intervention (Figure 2A). Individual analysis demonstrated a consistent decrease in total IgE levels across all dogs. Given that atopic dermatitis is associated with type I allergy and regulated by Th1/Th2 responses, levels of interferon-γ (IFN-γ) and IL-4 were measured to assess the expression of these pathways (Figure 2B). While IFN-γ levels increased and IL-4 levels decreased on day 60 compared to baseline, these changes did not reach statistical significance.
2.2. LCP Intervention Altered the Composition of Specific Bacterial Species in the Fecal Microbiota Of Dogs Suffering from Atopic Dermatitis
To identify potential bacterial biomarkers, we conducted third-generation sequencing analysis on fecal samples from dogs classified into mild to severe groups. Figure 3 illustrates the alterations in fecal microbiota before and after the trial. Compared to day 0, the Shannon index (p = 0.16) and richness index (p = 0.41) showed no significant differences, while Pielou evenness decreased (p = 0.06) on day 60 (Figure 3A). PCoA plot represented beta diversity (Figure 3B), with PC1 and PC2 accounting for 16.7% and 18.6% of the total variance, respectively. Similar patterns in the PCoA plot before and after LCP treatment suggested minimal changes in the composition of the core fecal bacteria.
In Figure 3C, a total of 5 main phyla was found in AD canine fecal samples in this study, including Firmicutes (93.03%), Proteobacteria (3.84%), Actinobacteria (2.74%), Bacteroidetes (0.36%), and Fusobacteria (0.04%), and 10 dominant families, with Lachnospiraceae (from 56.70% to 60.55%), followed by Peptostreptococcaceae (from 13.04% to 14.64%), Clostridiaceae (from 11.91% to 2.27%), Streptococcaceae (from 0.51% to 10.58%), Erysipelotrichaceae (from 5.38% to 3.08%), Enterobacteriaceae (from 3.53% to 4.15%), Coriobacteriaceae (from 3.6% to 1.65%), Enterococcaceae (from 2.43% to 0.53%), Lactobacillaceae (from 1.31% to 0.08%), and Oscillospiraceae (from 0.19% to 0.88%). In the genus level, the top 10 bacteria were Blautia (from 38.07% to 46.07%), Peptacetobacter (from 10.22% to 12.09%), Faecalimonas (from 11.85% to 7.88%), Sarcina (from 10.65% to 0.00%), Mediterraneibacter (from 5.88% to 4.75%), Lactococcus (from 0.39% to 7.06%), Shigella (from 2.91% to 2.91%), Erysipelatoclostridium (from 3.44% to 2.02%), Collinsella (from 3.62% to 2.02%), and Romboutsia (from 2.57% to 1.71%).
One hundred and thirty-seven bacterial species were found in the fecal samples. The highest amount of species was Peptacetobacter hiranonis (from 10.22% to 12.09%), and the following 9 species were Blautia caecimuris (from 11.78% to 9.85%), Blautia schinkii (from 8.16% to 11.83%), Faecalimonas umbilicata (from 11.85% to 7.88%), Blautia coccoides (from 4.30% to 6.72%), Mediterraneibacter [Ruminococcus] gnavus ATCC 29149 (from 5.22% to 4.35%), Blautia glucerasea (from 3.40% to 5.61%), Blautia argi (from 4.67% to 4.28%), Lactococcus cremoris (from 0.38% to 7.06%), and Blautia hansenii DSM 20583 (from 3.42% to 3.15%). Using Wilcoxon signed rank test to analyze the bacterial relative abundance before and after the treatment at different levels (Figure 3D), we found that Erysipelotrichaceae significantly declined (p < 0.05) at the family level, and Romboutsia reduced (p = 0.09) at the genus level as well. As for the bacteria in the LCP capsule, Lc. cremoris was detected in four-sixth of tested dogs after the LCP intervention.
2.3. LCP Intervention Altered the Gut Microbial Function
To clarify the functional differences resulting from changes in gut microbiota composition after LCP treatment, as well as the influence of gut environment changes on skin symptoms, this study compared functional differences related to carbohydrate metabolism and immune system-associated inflammation (Figure 3E). We observed a trend towards differences (p < 0.1) between groups in the Pentose phosphate pathway (ko00030), Pyruvate metabolism (ko00620), and Propanoate metabolism (ko00640). Pyruvate metabolism and Propanoate metabolism showed higher functionality in the LCP-treated group, while the Pentose phosphate pathway exhibited higher functionality in the group before LCP treatment. Additionally, Butanoate metabolism (ko00650) showed increased functionality after LCP treatment, possibly associated with short-chain fatty acid synthesis. Moreover, although no significant differences were found in immune system-related pathways, there was a decrease in functionality of nucleotide oligomerization domain (NOD) -like receptor signaling pathway (ko04621) after LCP treatment.
Subsequent short-chain fatty acid analysis was conducted to understand potential pathways through which changes in the gut environment affect the skin condition. According to the results in Figure 4, although there was no statistically significant difference in SCFA levels before and after the use of LCP capsules, after 60 days of probiotic intervention, increased levels of acetic acid, propionic acid, and butyric acid were observed in the feces of most of the tested dogs.
2.4. LCP Intervention Altered the Composition of Specific Bacterial Species in the Skin Microbiota of Dogs Suffering from Atopic Dermatitis
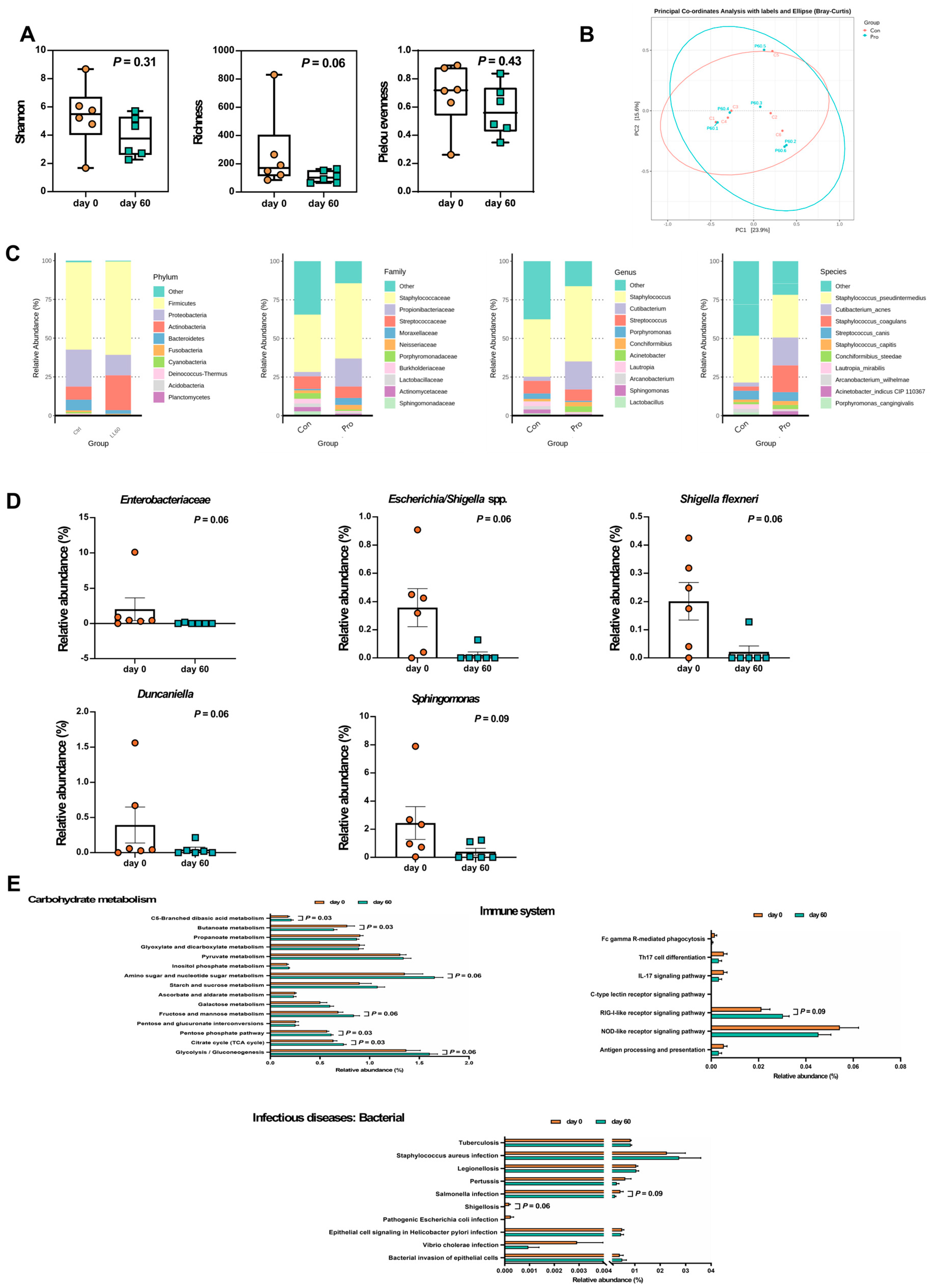
Alpha diversity of the skin microbiota in dogs with atopic dermatitis (AD) is illustrated in Figure 5A. The richness index (p = 0.06) decreased after 60 days of LCP capsule intake, although the differences were not statistically significant. In the PCoA plot, beta diversity exhibited a shift, with PC1 and PC2 accounting for 23.9% and 15.6%, respectively, of the total skin microbiota composition (Figure 5B).
In the skin sample microbiota composition (Figure 5C), the predominant phyla on the tested dogs' skin were Firmicutes (58.3%), Proteobacteria (18.59%), Actinobacteria (15.43%), Bacteroidetes (4.65%), Fusobacteria (0.71%), Cyanobacteria (0.54%), Deinococcus-Thermus (0.51%), Acidobacteria (0.37%), and Planctomycetes (0.12%). Among the 178 identified families, the top ten families, which collectively represented 75.59% of the overall microbiota composition in both groups, were notable. Staphylococcaceae exhibited the highest prevalence, comprising approximately 37.20% before probiotic administration and 48.71% afterward. Additionally, Propionibacteriaceae (3.63% before and 0.89% after probiotic intake) and Streptococcaceae (8.06% before and 7.31% after) were also prominent. In terms of dominant bacterial genera, 411 genera were identified. The top ten genera accounted for 73.09% of the overall microbiota composition. Staphylococcus had the highest abundance, approximately 37.20% before and 48.69% after probiotic intake, followed by Cutibacterium, Streptococcus, Porphyromonas, Conchiformibius, Acinetobacter, Lautropia, Arcanobacterium, and Sphingomonas. At the species level, 886 species were identified. The top ten species accounted for 65.04% of the overall microbiota composition. Staphylococcus pseudintermedius had the highest abundance, approximately 30.29% before and 27.70% after probiotic intake, followed by Cutibacterium acnes, Staphylococcus coagulans, Streptococcus canis, Staphylococcus capitis, Conchiformibius steedae, Lautropia mirabilis, Arcanobacterium wilhelmae, Acinetobacter indicus CIP 110367, and Porphyromonas cangingivalis.
Subsequent non-parametric statistical analysis using the matched-pairs Wilcoxon signed rank test compared the relative abundance differences in skin microbiota composition before and after probiotic use. The results indicated a decreasing trend (p < 0.1) in the genera Duncaniella, Escherichia/Shigella spp., and Sphingomonas, and at the species level in Shigella flexneri (p < 0.1) (Figure 5D).
2.5. LCP Intervention Altered the Skin Microbial Function
To assess the impact of changes in skin microbiota composition following LCP treatment on immune-related functions, this study compared functional differences in Carbohydrate metabolism and the Immune system at level two. According to the results in Figure 5E, a trend towards differences (p < 0.1) were observed between the two groups in Carbohydrate metabolism at level two, specifically in Glycolysis/Gluconeogenesis (ko00010), Citrate cycle (TCA cycle) (ko00020), Pentose phosphate pathway (ko00030), Fructose and mannose metabolism (ko00051), Amino sugar and nucleotide sugar metabolism (ko00520), Butanoate metabolism (ko00650), and C5-Branched dibasic acid metabolism (ko00660). Glycolysis/Gluconeogenesis, Citrate cycle (TCA cycle), Pentose phosphate pathway, Fructose and mannose metabolism, Amino sugar and nucleotide sugar metabolism, and C5-Branched dibasic acid metabolism exhibited higher functionality in the group after LCP use, while Butanoate metabolism showed higher functionality in the group before LCP use. In the Immune system at level two, a marginal difference (p < 0.1) was observed between the two groups in the RIG-I-like receptor signaling pathway (ko04622), which showed higher functionality in the group after LCP use. Additionally, functionalities of Th17 cell differentiation (ko04659), IL-17 signaling pathway (ko04657), and NOD-like receptor signaling pathway (ko04621) were also observed to decrease after LCP use. Furthermore, given that skin damage in atopic dermatitis often accompanies secondary infections, comparisons were made for pathways related to infection at level two. It was found that the functionalities of Shigellosis (ko05131) and Salmonella infection (ko05132) under Infectious diseases: Bacterial levels tended to be lower after LCP use (p < 0.1).
Figure 5.
Skin microbiota analyses in dogs with atopic dermatitis (AD) throughout the experiment. (A) Alpha diversity, (B) Beta diversity, (C) Taxonomic shifts from the phylum to species levels, (D) Relative abundance of specific bacteria, and (E) Relative abundance of KEGG pathways. Statistical analyses of alpha diversity and relative abundance were conducted using the matched-pairs Wilcoxon signed-rank test. Each dot represented a dog. The results were presented as mean ± SEM (n = 6). Con indicated before the LCP supplement; Pro indicated after the LCP supplement for 60 days.
Figure 5.
Skin microbiota analyses in dogs with atopic dermatitis (AD) throughout the experiment. (A) Alpha diversity, (B) Beta diversity, (C) Taxonomic shifts from the phylum to species levels, (D) Relative abundance of specific bacteria, and (E) Relative abundance of KEGG pathways. Statistical analyses of alpha diversity and relative abundance were conducted using the matched-pairs Wilcoxon signed-rank test. Each dot represented a dog. The results were presented as mean ± SEM (n = 6). Con indicated before the LCP supplement; Pro indicated after the LCP supplement for 60 days.
2.6. Correlation Analysis between Fecal and Skin Microbiota and Atopic Dermatitis Indicators
Due to the complexity and diversity of both the gut and skin environments, microbial communities may interact via host-derived metabolites or biochemical substances. In order to understand the relationship between these microbial communities and indicators of atopic dermatitis, this study examined the correlation between previously identified risk microbes from microbiota analysis and atopic dermatitis indicators using Spearman's rank correlation coefficient. The analysis results were presented using a heatmap. According to the results in Figure 6, Erysipelotrichaceae, Romboutsia, Enterobacteriaceae, Escherichia/Shigella spp., and Shigella flexneri were all positively correlated with CADESI and total IgE concentration. Concentrations of IFN-γ were found to be positively correlated with Erysipelotrichaceae and Romboutsia, while IL-4 secretion showed a positive correlation with Erysipelotrichaceae and Enterobacteriaceae.
2.7. The Effectiveness of LCP Intervention in Alleviating Canine Atopic Dermatitis Involves Changes in the Abundance of Biomarkers Indicated in Both Skin and Fecal Samples
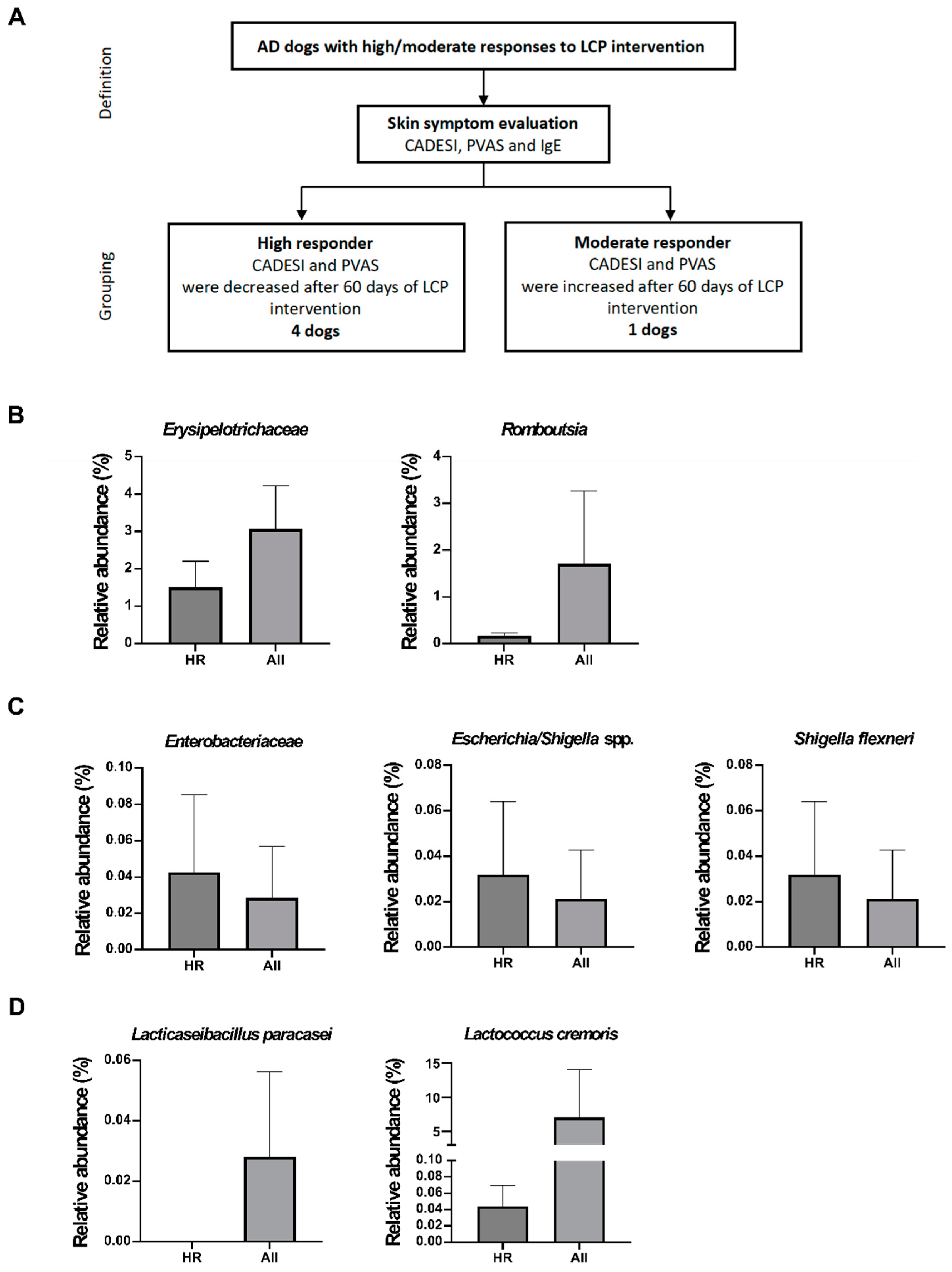
Finally, we investigated the factors contributing to the varying individual dog responses to LCP intervention against AD, as well as the effect of the gut and skin microbiome on LCP efficacy. The dogs were grouped based on clinical symptoms (IgE, CADESI, and PVAS), but since all dogs showed a decrease in IgE concentration after using LCP capsules, CADESI and PVAS were primarily used for grouping. Dogs with decreases in both CADESI and PVAS were classified as high responders (HR), while those with increases in both scores were classified as moderate responders (MR). Finally, 4 dogs were classified as high responders to AD and 1 as a moderate responder. However, the sample size for the moderate responder group (n = 1) was insufficient for statistical analysis. Therefore, subsequent comparisons were made between high-responding dogs (n = 4) and the overall group of dogs (n = 6) (Figure 7A).
In Figure 7B, the results indicate that 60 days after LCP use, the relative abundance ratios of Erysipelotrichaceae and Romboutsia in the gut of high responder (HR) dogs were lower than those in the overall group of dogs. Specifically, there was a greater decrease in Erysipelotrichaceae in the HR group compared to the overall group, while Romboutsia showed a slight increase. Additionally, in the comparison of skin microbiota (Figure 7C), higher relative abundances of Enterobacteriaceae, Escherichia/Shigella spp., and Shigella flexneri were observed on the skin of high-responding dogs. However, further analysis of the change ratios revealed that the decrease in these three specific bacteria was greater in the HR group than in the overall group of dogs. Subsequent analysis of two probiotic strains found in the feces, Lc. cremoris subsp. cremoris and L. paracasei subsp. paracasei, showed that they were not higher in abundance in the HR group compared to the overall group of dogs (Figure 7D).
3. Discussion
In this study, significant improvements were noted in the condition of the dogs' skin following a 60-day regimen of LCP capsule administration, with visible enhancements in symptoms such as redness, hair loss, and skin lesions. Around 75% of the dogs exhibited reduced CADESI and PVAS scores after the 60-day period, suggesting that this probiotic combination could effectively mitigate both the severity of skin lesions and scratching behavior. Chronic itching problems in dogs can induce psychological stress and lead to abnormal behavior [20], making the reduction of scratching behavior not only beneficial for minimizing skin damage and the risk of secondary infections but also for enhancing the overall quality of life for the dogs [21].
Further analysis of cytokines and total IgE in dog serum revealed that with increased duration of probiotic intake, total IgE concentration gradually decreased, reaching a significant difference after 60 days. Instances of decreased IL-4 and increased IFN-γ were observed in the serum of the tested dogs, consistent with our previous findings in an AD mouse study [19]. Following the use of the LCP capsule, both skin lesions and specific IgE levels were significantly lower compared to those in the AD group mice (p < 0.05), along with modulation of the ratio of Th1 and Th2 cells in the spleen. Atopic dermatitis is a complex inflammatory skin condition characterized by immune response imbalance, involving various cytokines and immunoglobulins. Elevated IL-4 levels drive IgE production, leading to allergic sensitization and AD symptom development. Concurrently, decreased IFN-γ levels weaken regulatory mechanisms that typically suppress the Th2 response and limit IgE production. This imbalance perpetuates the inflammatory cascade, exacerbating AD symptoms [22]. Reduction in IgE levels and alteration of the Th1/Th2 balance explain the improvement in skin symptoms and reduction in itching behavior. Additionally, although total IgE levels can be influenced by various factors and exhibit significant individual differences [23,24], during the evaluation of atopic dermatitis, other skin inflammation symptoms and parasitic infections were considered simultaneously for exclusion, and a paired-sample statistical analysis method was adopted to minimize the impact of individual differences.
Sequence analysis of the fecal microbiome revealed a reduction in microbial diversity following LCP administration. Currently, there is no definitive conclusion regarding whether atopic dermatitis leads to a decrease in gut microbiota diversity. While most literature suggests that AD patients exhibit reduced gut microbiota diversity compared to healthy individuals [25,26,27], some studies show no difference between the two groups [28,29]. Based on the analysis of weighted UniFrac distance, α-diversity, and β-diversity, it is inferred that LCP use did not significantly affect the core microbiota of dogs. However, there were changes in the composition of certain gut microbiota, resulting in a reduction in the relative abundance of some taxa, thereby decreasing evenness and affecting diversity.
Comparing dominant bacterial taxa before and after the trial revealed differences in microbial composition. Consistent with other canine studies, Firmicutes predominated, followed by Proteobacteria, Actinobacteria, Bacteroidetes, and Fusobacteria [26,30]. The family Erysipelotrichaceae significantly decreased (p < 0.05), which is associated with increased expression of the NOD2 gene in human studies [31]. NOD2, a cytoplasmic protein highly expressed in epithelial cells, is involved in the activation of pro-inflammatory pathways and regulation of Th1 responses mediated by toll-like receptor (TLR2) [32,33,34,35]. Studies in mice have shown that deletion of the NOD2 gene inhibits the production of Th2-related cytokines (IL-4, IL-5, and IL-13), consistent with our finding of a positive correlation between Erysipelotrichaceae and IL-4 and IgE. Furthermore, a decrease in the genus Romboutsia (p < 0.1) was observed. Previous research in humans found higher relative abundance of Romboutsia in AD patients compared to those with eczema and allergic rhinitis [36]. Romboutsia has been positively correlated with pro-inflammatory cytokines such as IL-6, IL-1β, TNF-α, and IL-23 [37,38], which may promote differentiation of Th17 cells. Th17 cells stimulate keratinocyte proliferation and differentiation, and their activation may contribute to skin barrier dysfunction. Therefore, Romboutsia may modulate inflammation and affect skin barrier structure via circulation. It is worth to notice that, when comparing the effects of LCP capsules on highly responsive AD dogs, the proportions of Erysipelotrichaceae and Romboutsia in the gut were lower than in the overall group of dogs.
Interestingly, prior to the trial, only one dog exhibited detectable levels of Lc. cremoris and L. paracasei. However, after 60 days, there was an increase in the proportion of Lc. cremoris observed in over half of the tested dogs, although this change was not statistically significant. On the other hand, L. paracasei exhibited a trend of increase only in dogs where it was initially detected before the trial commenced. These findings suggest that the levels of Lc. cremoris and L. paracasei may play a crucial role in alleviation of AD in dogs. Apart from the indigenous gut microbiota, individualized colonization patterns may stem from host factors, including variations in the expression of mucosal immune-related genes within the gastrointestinal organs [39].
Changes in gut microbial composition were noted alongside significant alterations in KEGG microbial functions. Although no significant variances were detected in pathways linked to the immune system, a decline in the function of the NOD-like receptor (NLR) signaling pathway was observed following LCP use, consistent with prior findings. The intervention with LCPs appeared to reduce the abundance of Erysipelotrichaceae, subsequently downregulating the NLR signaling pathway, thereby diminishing inflammasome formation and reducing the release of pro-inflammatory cytokines such as IL-18 [40]. Moreover, the LCP-treated group exhibited increased functionality in pyruvate metabolism and propanoate metabolism, correlating with the synthesis of short-chain fatty acids, which aligns with our findings. Elevated concentration of acetic acid, propionic acid, and butyric acid was detected in the feces of most tested dogs after 60 days of LCP intervention. The previous research highlighted the capacity of gut-derived SCFAs to impact epidermal keratinocytes, modifying their metabolism to enhance skin barrier function and mitigate allergen penetration [41].
In the examination of skin microbiota, both the Simpson index and α-diversity showed a declining trend post-probiotic use, mirroring findings in gut microbiota analysis. While skin microbiota diversity is pivotal, it can be influenced by various factors including environmental conditions, humidity, skin structure, and sebum distribution. Sebum secretion may foster lipophilic bacteria, thus favoring their dominance, potentially increasing microbiota diversity in dry skin environments. Moreover, compromised skin barriers may render wounds more susceptible to environmental microbial colonization, or tissue fluid secretion may elevate skin microbiota diversity by creating a moist environment [15]. Further analysis revealed reductions in Duncaniella, Escherichia/Shigella spp., and Sphingomonas at the genus level, with S. flexneri exhibiting a species-level decrease. Duncaniella, previously known as S24-7, belongs to the Muribaculaceae family, a major bacterial group in laboratory mice gut microbiota [42]. Certain Duncaniella species have demonstrated potential in ameliorating DSS-induced intestinal damage, Sphingomonas, part of the Sphingomonadaceae family, was found in higher proportions on the skin of healthy cattle in regions with greater rainfall [43,44]. However, the effects of Duncaniella and Sphingomonadaceae on dogs and skin aspects require clarification.
Regarding S. flexneri, it is categorized within the Escherichia/Shigella spp. genus and Enterobacteriaceae family. Limited research exists on its relevance to canine atopic dermatitis, but in human studies, S. flexneri has been implicated in activating the NLR of epithelial cells and triggering inflammatory responses, particularly via NOD1 recognition. NLRP10 expression in epidermal and dermal fibroblast-like cells can amplify NOD1 signaling, thus affecting innate immune responses, with potential associations with atopic dermatitis and allergic contact dermatitis [45,46,47,48]. Comparisons between highly responsive AD dogs and the overall population revealed a higher proportion of decreases in Enterobacteriaceae, Escherichia/Shigella spp., and S. flexneri in the highly responsive group.
In the results of gene functional prediction analysis, pathways related to SCFA metabolism including Glycolysis/Gluconeogenesis, Citrate cycle (TCA cycle), Pentose phosphate pathway, Fructose and mannose metabolism, Amino sugar and nucleotide sugar metabolism, and C5-Branched dibasic acid metabolism exhibited higher functionality in the probiotic-treated groups. Besides the intestinal-derived short-chain fatty acids, short-chain fatty acids on the skin can also serve as inhibitors of histone deacetylases (HDACs), thereby reducing the production of pro-inflammatory cytokines by inhibiting the NF-κB pathway, or by regulating Treg cells and promoting keratinocyte differentiation through HDAC inhibition, thereby influencing skin health [49,50,51]. Within the immune system, the retinoic acid-inducible gene (RIG)-I-like receptor signaling pathway showed higher functionality in the probiotic-treated groups. Additionally, given that skin damage associated with atopic dermatitis often accompanies secondary infections, pathways related to bacterial infection were compared. It was found that the functionality of Shigellosis within Infectious diseases: Bacterial showed a decrease after LCP use, consistent with the earlier decrease in relative abundance of S. flexneri. Within the immune system, functions related to Th17 cell differentiation, IL-17 signaling pathway, and NLR signaling pathway also exhibited lower functionality after probiotic use. These results indicate that changes in the skin barrier status can influence the composition of microbiota, and these compositional changes in different functional microbiota can also modulate skin environment health.
4. Materials and Methods
4.1. Bacterial Strains
Lc. cremoris subsp. cremoris MP01 and L. paracasei subsp. paracasei MP02 were isolated from traditional fermented milk. The probiotic mixture was cultured and activated in Lactobacilli MRS broth (Neogen Corporation, Lansing, MI, USA) at 30 oC for 24 hours.
4.2. Preparation of Probiotic Capsule
The dosage of AD dogs was decided by previous study [19]. The freeze-dried probiotic powder, consisting of Lc. cremoris subsp. cremoris MP01 and L. paracasei subsp. paracasei MP02, was produced by Grape King Bio, Ltd. (Taoyuan, Taiwan) and filled in capsule No.4 as LCP capsules. Each capsule contained 3.1 × 1010 CFU and was stored at -20 oC.
4.3. Clinical Canine Trial
This study was approved by the Institutional Animal Care and Use Committee of National Taiwan University (IACUC approval no: NTU-109-EL-000146). All owners were required to sign a consent form to participate in this study. Recruitment occurred from August to November 2021. The criteria for preliminary selection were adapted from the previous research [52,53]. There were no restrictions regarding the dogs' age, weight, gender, breed, and sterilization status. Eligible dogs had to exhibit itching and erythema on their skin and meet at least four of the following conditions:
- The initial onset of symptoms must have occurred at or before 3 years of age.
- The dog primarily resided indoors. Itching symptoms were significantly improved by steroids
- Skin lesions were primarily located on the ears, interdigital spaces, around the mouth, eyes, dorsal surfaces of the joints, and inguinal areas.
- The edge skin of the ears appeared normal
- The skin of the lower back was normal
Dogs were excluded if they were pregnant or nursing. Those with infectious dermatitis were required to discontinue medication to meet the inclusion criteria, and those with food-induced dermatitis had to eliminate the allergen before joining the trial. Additionally, dogs could not have used antibiotics, steroids, or immune suppressants within two weeks prior to the experiment.
Forty-five dogs were initially enrolled in this study. Following initial examinations and assessments by veterinarians, 11 dogs remained eligible. Eight dogs ultimately completed this clinical trial (Figure 1A). The median body weight of the participating dogs was 11.42 kg, ranging from 2.7 to 32 kg, with a female-to-male ratio of 1:1. Age distribution was as follows: two dogs were 1–3 years old; three dogs each belonged to the 3–7, and >8 years age groups. Based on the Canine Atopic Dermatitis Extent and Severity Index (CADESI)-04 of day 0, two dogs were classified into the normal or reducing AD symptoms group (CADESI score < 10), three into the mild group (score = 10–34), two into the moderate group (score = 35–59), and one dog into the severe group (score ≥ 60). The dogs were required to continuously intake one LCP capsule daily for 60 days. Blood, skin, and fecal samples were collected on day 0, 30, and 60.
4.4. Assessment of the Degree of Atopic Dermatitis and Pruritus
To evaluate the severity level of atopic dermatitis, the CADESI-04 [54] and the Pruritus Visual Analog Scale (PVAS) [55] were applied. For CADESI-04, the level of erythema, lichenification, excoriation, and alopecia were inspected and graded in 20 body areas on a scale from 0 (none) to 3 (severe), with a total score of 180. The PVAS was administered by the owners.
4.5. Cytokine Production from Peripheral Blood Mononuclear Cells (PBMCs)
Blood samples were mixed with Dulbecco’s phosphate-buffered saline (DPBS; Thermo Fisher Scientific Inc., MA, USA) and centrifuged at 400 ×g, 18 oC for 35 mins. The monocellular cells were collected and suspended in Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma-Aldrich Inc., St. Louis, MO, USA) with heated-inactivated fetal bovine serum (FBS) and antibiotic-antimycotic solution (Corning Inc., Corning, NY, USA). Cells were cultured at 37 oC for 24 hours with 5% of CO2. Then, the cell-cultural suspension was centrifuged at 400 ×g, 4 oC for 10 mins to obtain supernatant for cytokine quantification.
The commercial enzyme-linked immunoassay (ELISA) kits for canines were used to measure interleukin (IL)-4 (BioassayTechnology Laboratory, China), interferon (INF)-γ (R&D systems, MN, USA) from PBMC.
4.6. IgE Production in Serum
Immunoglobulin E (IgE) in serum was measured through the canine ELISA kit (LifeSpan BioScience Inc., WA, USA).
4.7. Microbiota Analyses in Skin and Fecal
Fecal samples were collected as described in the previous study described [56]. Skin bacteria from the areas around the mouth, ears, armpits, and groin of the tested dogs were collected using a swabbing method. The cotton swabs were then soaked in 0.85% sterilized sodium chloride solution, and DNA was extracted.
Third-generation sequencing was performed by BIOTOOLS Co., Ltd. (Taipei, Taiwan). Universal primers 27F: 5’-AGRGTTYGATYMTGGCTCAG-3’, and 1492R: 5’ RGYTACCTTGTTACGACTT-3’ was used to amplify the full length of the 16S rRNA gene (V1–V9). After a series of quality controls to eliminate noise, reads with a quality score (RQ) greater than 30 were retained. These reads were then processed using the DADA2 package (dada2_1.20) in R software to denoise to generate amplicon sequence variants (ASVs), with each ASV defined as one species cluster. The ASVs were classified to their taxonomy using QIIME2 (v2021.4; http://qiime2.org) and the NCBI 16S ribosomal RNA database (2021.1).
In this study, we analyzed α diversity (observed species richness), Shannon-Wiener diversity index, and Pielou’s evenness), β diversity (Principal Coordinates Analysis, PCoA), PICRUSt (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States), and KEGG (Kyoto Encyclopedia of Genes and Genomes) to determine the change in composition or predicted function in the gut or skin microbiota. The Spearman correlation was used to evaluate the relationship between specific bacterial taxonomies and atopic dermatitis-related indicators.
4.8. Short-Chain Fatty Acids Analysis
The analysis was slightly modified from the previous study [57]. The obtained SCFAs were dissolved in 250 μL of methanol. The PU-2089 Quaternary HPLC pump (Jasco International Co. Ltd, Japan) with Reprosil 100 C18 5 μΜ, 250 × 4.6 mm (Dr. Maisch GmbH, Germany) was used to quantify the concentration of fecal acetic acid, propionic acid, and butyric acid. The mobile phase contained acetonitrile (C2H3N, AE-0627; Aencore, Australia), 99% methanol (CH3OH, CAS 67-56-1; Duksan, Korea), and ultrapure water with a volume ratio of 30, 16, and 54, respectively, and then adjusted the pH value to 4.5 by trifluoroacetic acid (J.T.Baker, NJ, USA). The flow rate was set as 1.1 mL/min; the column temperature was 50℃; the detected wavelength was 400 nm.
4.9. Statistical Analysis
All the data were presented as mean ± SEM. According to the results of the Shapiro-Wilk normality test, the statistical analyses were performed by the Wilcoxon signed-rank test or ratio paired t-test by GraphPad Prism v9.3.1 (GraphPad Software Inc., MA, USA) and Statistical Analysis System v9.4 (SAS Institute Inc., NC, USA). Since the data from microbiota analyses were performed in relative abundance, we applied nonparametric statistics in all microbiota studies in this research.
5. Conclusions
In summary, the LCP probiotic combination used in this study demonstrated efficacy in improving clinical symptoms of canine skin, reducing itching behavior, and regulating the immune system. The possible mechanisms involve an increase in the proportion of Lc. cremoris while decreasing the proportions of Erysipelotrichaceae and Romboutsia, coupled with an upregulation in SCFA levels. Additionally, it modulated the intestinal immune system towards regulating the Th1/Th2 balance and decreasing the secretion of allergy-related immunoglobulin E. Through circulation in the bloodstream, it further improved systemic immune responses, thereby contributing to alleviating symptoms of atopic dermatitis, regulating skin microbiota composition, and reducing adherence and invasion of environmental microbes and pathogens. However, the small sample size in the microbiota analysis section affected some statistical results, and the mechanisms underlying these changes in the microbiota in canine atopic dermatitis remain unclear, necessitating further investigation in the future.
Author Contributions
Conceptualization, Chen, M.J. and Lee, Y.J.; methodology, Hsieh, J.C., Huang, H.W.; Yeh, T.C.; software, Huang, H.W.; Yeh, T.C.; validation, Huang, H.W.; Hsieh, J.C.; formal analysis, Yeh, T.C.; Huang, H.W. investigation, Yeh, T.C.; Hsieh, J.C. resources, Chen, M.J.; data curation, Yeh, T.C.; writing—original draft preparation, Yeh, T.C.; Huang, H.W.; Tsai, C.W.; Chen, M.J. writing—review and editing, Huang, H.W.; Chen, M.J. visualization, Yeh, T.C.; supervision, Chen, M.J.; Lee, Y.J. project administration, Chen, M.J.; funding acquisition, Chen, M.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research is funded by Council of Agriculture of Taiwan, Executive Yuan, Republic of China (110AS-10.1.7-AD-U1).
Institutional Review Board Statement
The animal study was approved by the Institutional Animal Care and Use Committee of National Taiwan University (IACUC approval no: NTU-109-EL-000146).
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article and its Supplementary materials. Raw data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We express our gratitude to all the dog owners, Dr. Tsu-Cheng Hsu, Dr. Jung-Chih Chen, Dr. Shr-Ping Teng, Dr. Yi-Wen Lin, Dr. Yi-Fan Lee and Dr. Yin-Chih Liu for their participation and cooperation.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ASVs: amplicon sequence variants; CAD, canine atopic dermatitis; CADESI, Canine Atopic Dermatitis Extent and Severity Index; DPBS, Dulbecco’s phosphate-buffered saline; FBS, fetal bovine serum; HR, high responders; HDACs, histone deacetylases; IgE, immunoglobulin E; IACUC, Institutional Animal Care and Use Committee; IFN-γ, interferon-γ; IL, interleukin; KEGG, Kyoto Encyclopedia of Genes and Genomes; MR, moderate responders; NOD, nucleotide oligomerization domain; PICRUSt, Phylogenetic Investigation of Communities by Reconstruction of Unob-served States; PCoA, principal coordinates analysis; PVAS, Pruritus Visual Analogue Scale; Treg, regulatory T cells; RIG, retinoic acid-inducible gene; SCFA, short-chain fatty acid; Th1 cells, type 1 T helper cells; Th2 cells, type 2 T helper cells.
References
- Gedon, N.K.Y.; Mueller, R.S. Atopic dermatitis in cats and dogs: a difficult disease for animals and owners. Clin. Transl. Allergy 2018, 8, 1–12. [CrossRef]
- Outerbridge, C.A.; Jordan, T.J. Current knowledge on canine atopic dermatitis: pathogenesis and treatment. Adv. Small Anim. Care 2021, 2, 101–115.
- Couceiro, G.A.; Ribeiro, S.M.M.; Monteiro, M.M.; Meneses, A.M.C.; Sousa, S.K.S.; Coutinho, L.N. Prevalence of canine atopic dermatitis at the Veterinary Hospital of the “Universidade Federal Rural da Amazônia” in Belém/Pará, Brazil. Pesqui. Vet. Bras. 2021, 41.
- Humeniuk, P.; Dubiela, P.; Hoffmann-Sommergruber, K. Dendritic Cells and Their Role in Allergy: Uptake, Proteolytic Processing and Presentation of Allergens. Int. J. Mol. Sci. 2017, 18, 1491. [CrossRef]
- Marsella, R.; Sousa, C.A.; Gonzales, A.J.; Fadok, V.A. Current understanding of the pathophysiologic mechanisms of canine atopic dermatitis. J. Am. Veter- Med Assoc. 2012, 241, 194–207. [CrossRef]
- Nguyen, H.L.; Anderson, K.R.; Tollefson, M.M. New and Emerging Therapies for Pediatric Atopic Dermatitis. Pediatr. Drugs 2019, 21, 239–260. [CrossRef]
- Gonzales, A.J.; Bowman, J.W.; Fici, G.J.; Zhang, M.; Mann, D.W.; Mitton-Fry, M. Oclacitinib (APOQUEL®) is a novel Janus kinase inhibitor with activity against cytokines involved in allergy. J. Veter- Pharmacol. Ther. 2014, 37, 317–324. [CrossRef]
- Bağci, I.S.; Ruzicka, T. IL-31: A new key player in dermatology and beyond. J. Allergy Clin. Immunol. 2018, 141, 858–866. [CrossRef]
- Chrobak-Chmiel, D.; Golke, A.; Kwiecień, E.; Biegańska, M.J.; Dembele, K.; Dziekiewicz-Mrugasiewicz, M.; Czopowicz, M.; Kizerwetter-Świda, M.; Rzewuska, M. Is Vitamin D3 a Worthy Supplement Protecting against Secondary Infections in Dogs with Atopic Dermatitis?. Pathogens 2023, 12, 145. [CrossRef]
- De Pessemier, B.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut–Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions. Microorganisms 2021, 9, 353. [CrossRef]
- Pfefferle, P.I.; Keber, C.U.; Cohen, R.M.; Garn, H. The Hygiene Hypothesis – Learning From but Not Living in the Past. Front. Immunol. 2021, 12. [CrossRef]
- Dou, J.; Zeng, J.; Wu, K.; Tan, W.; Gao, L.; Lu, J. Microbiosis in pathogenesis and intervention of atopic dermatitis. Int. Immunopharmacol. 2019, 69, 263–269. [CrossRef]
- E Fujimura, K.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [CrossRef]
- Fang, Z.; Li, L.; Zhang, H.; Zhao, J.; Lu, W.; Chen, W. Gut Microbiota, Probiotics, and Their Interactions in Prevention and Treatment of Atopic Dermatitis: A Review. Front. Immunol. 2021, 12, 720393. [CrossRef]
- Ellis, S.R.; Nguyen, M.; Vaughn, A.R.; Notay, M.; Burney, W.A.; Sandhu, S.; Sivamani, R.K. The Skin and Gut Microbiome and Its Role in Common Dermatologic Conditions. Microorganisms 2019, 7, 550. [CrossRef]
- Kwon, H.-K.; Lee, C.-G.; So, J.-S.; Chae, C.-S.; Hwang, J.-S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.-C.; Im, S.-H. Generation of regulatory dendritic cells and CD4 + Foxp3 + T cells by probiotics administration suppresses immune disorders. Proc. Natl. Acad. Sci. 2010, 107, 2159–2164. [CrossRef]
- Shin, J.-H.; Chung, M.-J.; Seo, J.-G. A multistrain probiotic formulation attenuates skin symptoms of atopic dermatitis in a mouse model through the generation of CD4+Foxp3+T cells. Food Nutr. Res. 2016, 60, 32550. [CrossRef]
- Kim, W.-K.; Jang, Y.J.; Han, D.H.; Seo, B.; Park, S.; Lee, C.H.; Ko, G. Administration of Lactobacillus fermentum KBL375 Causes Taxonomic and Functional Changes in Gut Microbiota Leading to Improvement of Atopic Dermatitis. Front. Mol. Biosci. 2019, 6, 92. [CrossRef]
- Chen, H.-Y.; Chen, Y.-T.; Li, K.-Y.; Huang, H.-W.; Lin, Y.-C.; Chen, M.-J. A Heat-Killed Probiotic Mixture Regulates Immune T Cells Balance and IgE Production in House Dust Mite Extraction-Induced Atopic Dermatitis Mice. Microorganisms 2022, 10, 1881. [CrossRef]
- Harvey, N.D.; Craigon, P.J.; Shaw, S.C.; Blott, S.C.; England, G.C. Behavioural Differences in Dogs with Atopic Dermatitis Suggest Stress Could Be a Significant Problem Associated with Chronic Pruritus. Animals 2019, 9, 813. [CrossRef]
- Linek, M.; Favrot, C. Impact of canine atopic dermatitis on the health-related quality of life of affected dogs and quality of life of their owners. Veter- Dermatol. 2010, 21, 456–462. [CrossRef]
- Udoye, C.C.; Rau, C.N.; Freye, S.M.; Almeida, L.N.; Vera-Cruz, S.; Othmer, K.; Korkmaz, R..; Clauder, A.-K.; Lindemann, T.; Niebuhr, M.; et al. B-cell receptor physical properties affect relative IgG1 and IgE responses in mouse egg allergy. Mucosal Immunol. 2022, 15, 1375–1388. [CrossRef]
- Carballo, I.; Alonso-Sampedro, M.; Gonzalez-Conde, E.; Sanchez-Castro, J.; Vidal, C.; Gude, F.; Gonzalez-Quintela, A. Factors Influencing Total Serum IgE in Adults: The Role of Obesity and Related Metabolic Disorders. Int. Arch. Allergy Immunol. 2021, 182, 220–228. [CrossRef]
- Chaudhary, S.K.; Singh, S.K.; Kumari, P.; Kanwal, S.; Soman, S.P.; Choudhury, S.; Garg, S.K. Alterations in circulating concentrations of IL-17, IL-31 and total IgE in dogs with atopic dermatitis. Vet. Dermatol. 2019, 30, 383–e114.
- Reddel, S.; Del Chierico, F.; Quagliariello, A.; Giancristoforo, S.; Vernocchi, P.; Russo, A.; Fiocchi, A.; Rossi, P.; Putignani, L.; El Hachem, M. Gut microbiota profile in children affected by atopic dermatitis and evaluation of intestinal persistence of a probiotic mixture. Sci. Rep. 2019, 9, 1–10. [CrossRef]
- Sugita, K.; Shima, A.; Takahashi, K.; Ishihara, G.; Kawano, K.; Ohmori, K. Pilot evaluation of a single oral fecal microbiota transplantation for canine atopic dermatitis. Sci. Rep. 2023, 13, 1–11. [CrossRef]
- Ye, S.; Yan, F.; Wang, H.; Mo, X.; Liu, J.; Zhang, Y.; Li, H.; Chen, D. Diversity analysis of gut microbiota between healthy controls and those with atopic dermatitis in a Chinese population. J. Dermatol. 2020, 48, 158–167. [CrossRef]
- Uchiyama, J.; Osumi, T.; Mizukami, K.; Fukuyama, T.; Shima, A.; Unno, A.; Takemura-Uchiyama, I.; Une, Y.; Murakami, H.; Sakaguchi, M. Characterization of the oral and faecal microbiota associated with atopic dermatitis in dogs selected from a purebred Shiba Inu colony. Lett. Appl. Microbiol. 2022, 75, 1607–1616. [CrossRef]
- Wang, Y.; Hou, J.; Tsui, J.C.-C.; Wang, L.; Zhou, J.; Chan, U.K.; Lo, C.J.Y.; Siu, P.L.K.; Loo, S.K.F.; Tsui, S.K.W. Unique Gut Microbiome Signatures among Adult Patients with Moderate to Severe Atopic Dermatitis in Southern Chinese. Int. J. Mol. Sci. 2023, 24, 12856. [CrossRef]
- You, I.; Kim, M.J. Comparison of Gut Microbiota of 96 Healthy Dogs by Individual Traits: Breed, Age, and Body Condition Score. Animals 2021, 11, 2432. [CrossRef]
- Turpin, W.; Bedrani, L.; Espin-Garcia, O.; Xu, W.; Silverberg, M.S.; Smith, M.I.; Garay, J.A.R.; Lee, S.-H.; Guttman, D.S.; Griffiths, A.; et al. Associations ofNOD2polymorphisms with Erysipelotrichaceae in stool of in healthy first degree relatives of Crohn's disease subjects. BMC Med Genet. 2020, 21, 1–8. [CrossRef]
- Duan, W.; Mehta, A.K.; Magalhaes, J.G.; Ziegler, S.F.; Dong, C.; Philpott, D.J.; Croft, M. Innate signals from Nod2 block respiratory tolerance and program TH2-driven allergic inflammation. J. Allergy Clin. Immunol. 2010, 126, 1284–1293.e10. [CrossRef]
- Girardin, S.E.; Boneca, I.G.; Viala, J.; Chamaillard, M.; Labigne, A.; Thomas, G.; Philpott, D.J.; Sansonetti, P.J. Nod2 Is a General Sensor of Peptidoglycan through Muramyl Dipeptide (MDP) Detection. J. Biol. Chem. 2003, 278, 8869–8872. [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9.
- Maeda, S.; Hsu, L.C.; Liu, H.; Bankston, L.A.; Iimura, M.; Kagnoff, M.F.; Eckmann, L.; Karin, M.. Nod2 mutation in Crohn's disease potentiates NF-κB activity and IL-1ß processing. Science 2005, 307, 734–738.
- Su, Y.-J.; Luo, S.-D.; Hsu, C.-Y.; Kuo, H.-C. Differences in gut microbiota between allergic rhinitis, atopic dermatitis, and skin urticaria. Medicine 2021, 100, e25091. [CrossRef]
- Li, R.; Yao, Y.; Gao, P.; Bu, S. The Therapeutic Efficacy of Curcumin vs. Metformin in Modulating the Gut Microbiota in NAFLD Rats: A Comparative Study. Front. Microbiol. 2021, 11. [CrossRef]
- Wang, H.; Zhang, M.; Wen, X.; He, L.; Zhang, M.; Zhang, J.; Yang, X. Cepharanthine ameliorates dextran sulphate sodium-induced colitis through modulating gut microbiota. Microb. Biotechnol. 2022, 15, 2208–2222. [CrossRef]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z.; et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388–1405.e21. [CrossRef]
- Platnich, J.M.; Muruve, D.A. NOD-like receptors and inflammasomes: A review of their canonical and non-canonical signaling pathways. Arch. Biochem. Biophys. 2019, 670, 4–14. [CrossRef]
- Trompette, A.; Pernot, J.; Perdijk, O.; Alqahtani, R.A.A.; Domingo, J.S.; Camacho-Muñoz, D.; Wong, N.C.; Kendall, A.C.; Wiederkehr, A.; Nicod, L.P.; et al. Gut-derived short-chain fatty acids modulate skin barrier integrity by promoting keratinocyte metabolism and differentiation. Mucosal Immunol. 2022, 15, 908–926. [CrossRef]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 1–15. [CrossRef]
- Glaeser, S.P.; Kämpfer, P. The family sphingomonadaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin, Germany, 2014; Volume 8, pp. 641–707.
- Gamage, H.K.; Vuong, D.; Minns, S.A.; Chen, R.; Piggott, A.M.; Lacey, E.; Paulsen, I.T. The composition and natural variation of the skin microbiota in healthy Australian cattle. Research Square 2022, submitted.
- Imamura, R.; Wang, Y.; Kinoshita, T.; Suzuki, M.; Noda, T.; Sagara, J.; Taniguchi, S.; Okamoto, H.; Suda, T. Anti-Inflammatory Activity of PYNOD and Its Mechanism in Humans and Mice. J. Immunol. 2010, 184, 5874–5884. [CrossRef]
- Lautz, K.; Damm, A.; Menning, M.; Wenger, J.; Adam, A.C.; Zigrino, P.; Kremmer, E.; Kufer, T.A. NLRP10 enhancesShigella-induced pro-inflammatory responses. Cell. Microbiol. 2012, 14, 1568–1583. [CrossRef]
- Mirza, N.; Sowa, A.S.; Lautz, K.; Kufer, T.A. NLRP10 Affects the Stability of Abin-1 To Control Inflammatory Responses. J. Immunol. 2019, 202, 218–227. [CrossRef]
- Fritz, J.H.; Le Bourhis, L.; Sellge, G.; Magalhaes, J.G.; Fsihi, H.; Kufer, T.A.; Collins, C.; Viala, J.; Ferrero, R.L.; Girardin, S.E.; et al. Nod1-Mediated Innate Immune Recognition of Peptidoglycan Contributes to the Onset of Adaptive Immunity. Immunity 2007, 26, 445–459. [CrossRef]
- Aoyama, M.; Kotani, J.; Usami, M. Butyrate and propionate induced activated or non-activated neutrophil apoptosis via HDAC inhibitor activity but without activating GPR-41/GPR-43 pathways. Nutrition 2010, 26, 653–661.
- Carrion, S.L.; Sutter, C.H.; Sutter, T.R. Combined treatment with sodium butyrate and PD153035 enhances keratinocyte differentiation. Exp. Dermatol. 2014, 23, 211–214. [CrossRef]
- Wang, Y.; Kao, M.-S.; Yu, J.; Huang, S.; Marito, S.; Gallo, R.L.; Huang, C.-M. A Precision Microbiome Approach Using Sucrose for Selective Augmentation of Staphylococcus epidermidis Fermentation against Propionibacterium acnes. Int. J. Mol. Sci. 2016, 17, 1870. [CrossRef]
- Favrot, C.; Steffan, J.; Seewald, W.; Picco, F. A prospective study on the clinical features of chronic canine atopic dermatitis and its diagnosis. Veter- Dermatol. 2010, 21, 23–31. [CrossRef]
- Terada, Y.; Nagata, M.; Murayama, N.; Nanko, H.; Furue, M. Clinical comparison of human and canine atopic dermatitis using human diagnostic criteria (Japanese Dermatological Association, 2009): Proposal of provisional diagnostic criteria for canine atopic dermatitis. J. Dermatol. 2011, 38, 784–790. [CrossRef]
- Olivry, T.; Saridomichelakis, M.; Nuttall, T.; Bensignor, E.; Griffin, C.E.; Hill, P.B. Validation of the Canine Atopic Dermatitis Extent and Severity Index (CADESI)-4, a simplified severity scale for assessing skin lesions of atopic dermatitis in dogs. Veter- Dermatol. 2014, 25, 77–e25. [CrossRef]
- Rybníček, J.; Lau-Gillard, P.J.; Harvey, R.; Hill, P.B. Further validation of a pruritus severity scale for use in dogs. Veter- Dermatol. 2009, 20, 115–122. [CrossRef]
- Watanabe, K.; Fujimoto, J.; Sasamoto, M.; Dugersuren, J.; Tumursuh, T.; Demberel, S. Diversity of lactic acid bacteria and yeasts in Airag and Tarag, traditional fermented milk products of Mongolia. World J. Microbiol. Biotechnol. 2007, 24, 1313–1325. [CrossRef]
- Torii, T.; Kanemitsu, K.; Wada, T.; Itoh, S.; Kinugawa, K.; Hagiwara, A. Measurement of short-chain fatty acids in human faeces using high-performance liquid chromatography: specimen stability. Ann. Clin. Biochem. Int. J. Biochem. Lab. Med. 2010, 47, 447–452. [CrossRef]
Figure 1.
(A) Study design. (B) Schematic diagram of the skin lesions in the perilabial area (left) and inguinal areas (right) of the recruited dog before and after taking LCP capsules for 60 days. (C) Changes in CADESI and PVAS score. Statistical analysis of the CADESI scores was performed using a paired t-test, while the PVAS score was analyzed using the Wilcoxon signed-rank test. The results were presented as mean ± SEM (n = 8). Change = (post-treatment score on day 60 – pretreatment score on day 0)/ pretreatment score on day 0.
Figure 1.
(A) Study design. (B) Schematic diagram of the skin lesions in the perilabial area (left) and inguinal areas (right) of the recruited dog before and after taking LCP capsules for 60 days. (C) Changes in CADESI and PVAS score. Statistical analysis of the CADESI scores was performed using a paired t-test, while the PVAS score was analyzed using the Wilcoxon signed-rank test. The results were presented as mean ± SEM (n = 8). Change = (post-treatment score on day 60 – pretreatment score on day 0)/ pretreatment score on day 0.
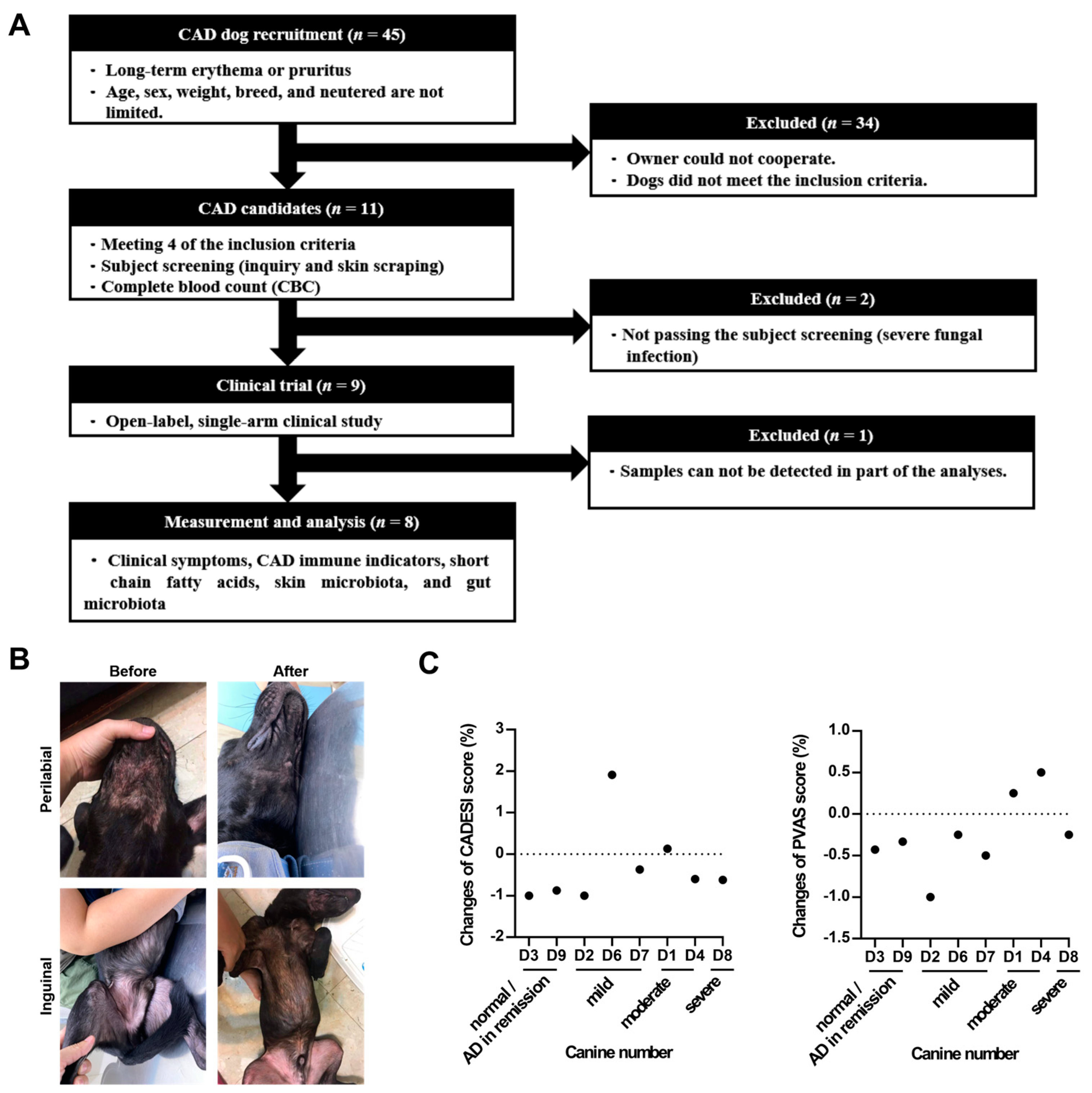
Figure 2.
Effects of (A) total IgE and (B) Th1/Th2-related cytokines in serum after 60 days of LCP capsule intervention in dogs with atopic dermatitis (AD). Statistical analysis was performed using the Wilcoxon signed-rank test for total IgE analysis and a paired t-test for cytokine analyses. Each dot represented a dog. The results were presented as mean ± SEM (n = 8). Change in total IgE = (post-treatment value on day 60 - pretreatment value on day 0) / pretreatment value on day 0. The Th1/Th2 ratio is calculated as IFN-γ concentration (pg/μg)/ IL-4 concentration (pg/μg). ***p < 0.001.
Figure 2.
Effects of (A) total IgE and (B) Th1/Th2-related cytokines in serum after 60 days of LCP capsule intervention in dogs with atopic dermatitis (AD). Statistical analysis was performed using the Wilcoxon signed-rank test for total IgE analysis and a paired t-test for cytokine analyses. Each dot represented a dog. The results were presented as mean ± SEM (n = 8). Change in total IgE = (post-treatment value on day 60 - pretreatment value on day 0) / pretreatment value on day 0. The Th1/Th2 ratio is calculated as IFN-γ concentration (pg/μg)/ IL-4 concentration (pg/μg). ***p < 0.001.
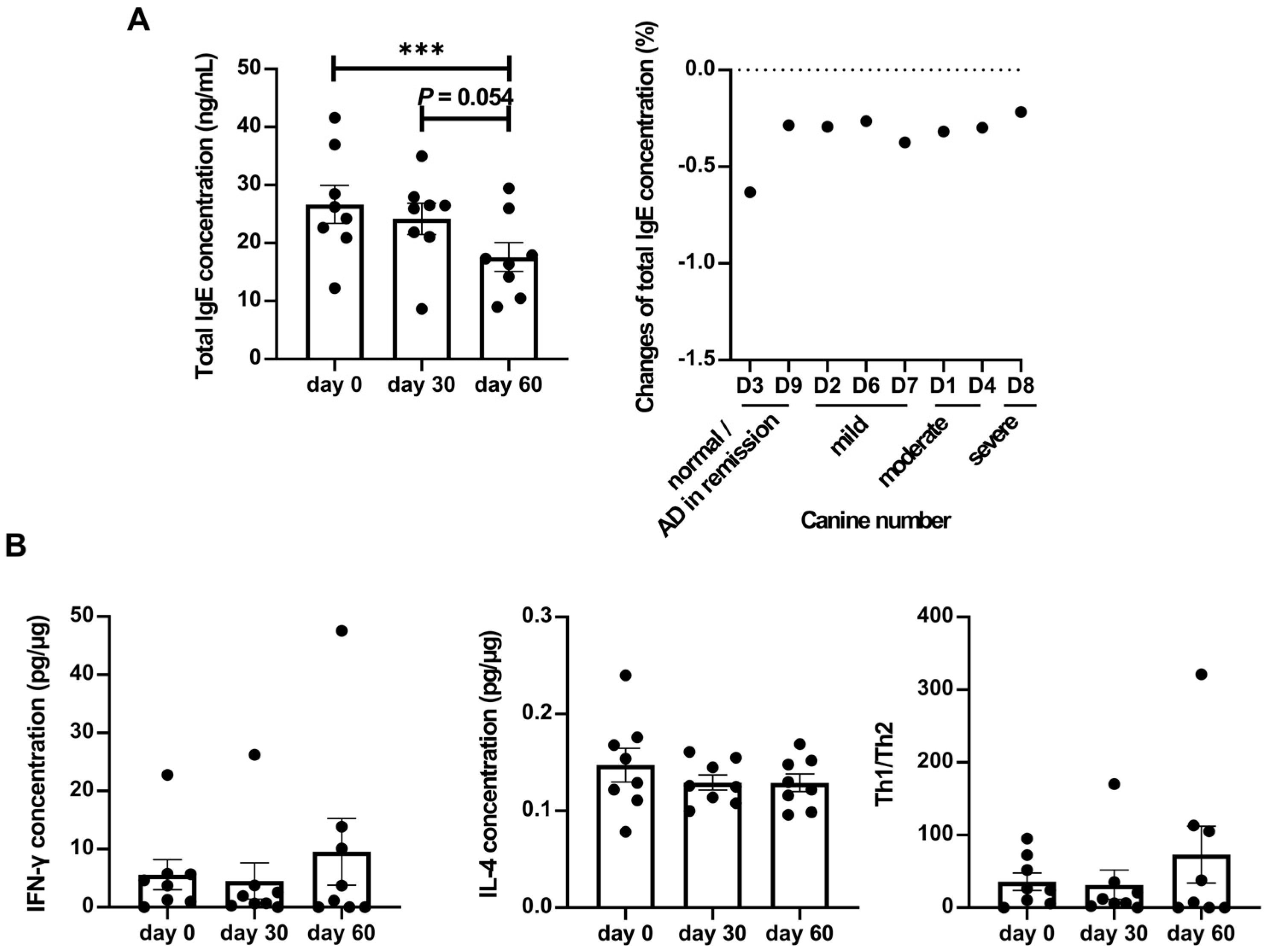
Figure 3.
Fecal microbiota analyses of dogs with atopic dermatitis (AD) throughout the experiment. (A) Alpha diversity, (B) Beta diversity, (C) Taxonomic shifts from the phylum to species levels, (D) Relative abundance of specific bacteria, and (E) Relative abundance of KEGG pathways. Statistical analyses of alpha diversity and relative abundance were performed using the Wilcoxon signed-rank test. Each dot represented a dog. The results were presented as mean ± SEM (n = 6). Con indicated before the LCP supplement; Pro indicated after the LCP supplement for 60 days.
Figure 3.
Fecal microbiota analyses of dogs with atopic dermatitis (AD) throughout the experiment. (A) Alpha diversity, (B) Beta diversity, (C) Taxonomic shifts from the phylum to species levels, (D) Relative abundance of specific bacteria, and (E) Relative abundance of KEGG pathways. Statistical analyses of alpha diversity and relative abundance were performed using the Wilcoxon signed-rank test. Each dot represented a dog. The results were presented as mean ± SEM (n = 6). Con indicated before the LCP supplement; Pro indicated after the LCP supplement for 60 days.
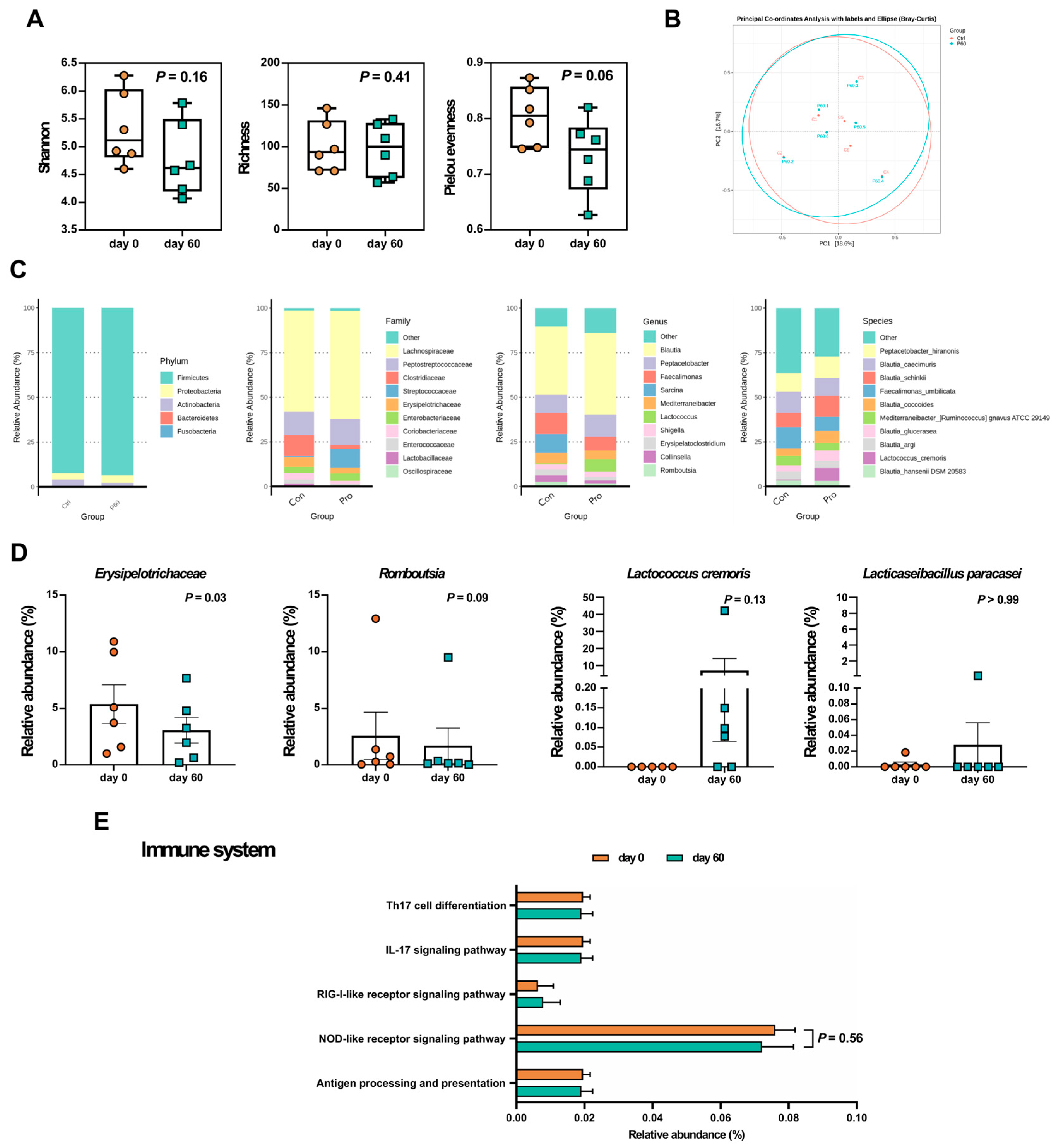
Figure 4.
Changes in fecal short-chain fatty acids (SCFAs) in dogs with atopic dermatitis (AD) following a 60-day probiotics intervention. Statistical analysis was performed using a paired t-test. Each dot represented a dog. The results were presented as mean ± SEM (n = 6).
Figure 4.
Changes in fecal short-chain fatty acids (SCFAs) in dogs with atopic dermatitis (AD) following a 60-day probiotics intervention. Statistical analysis was performed using a paired t-test. Each dot represented a dog. The results were presented as mean ± SEM (n = 6).
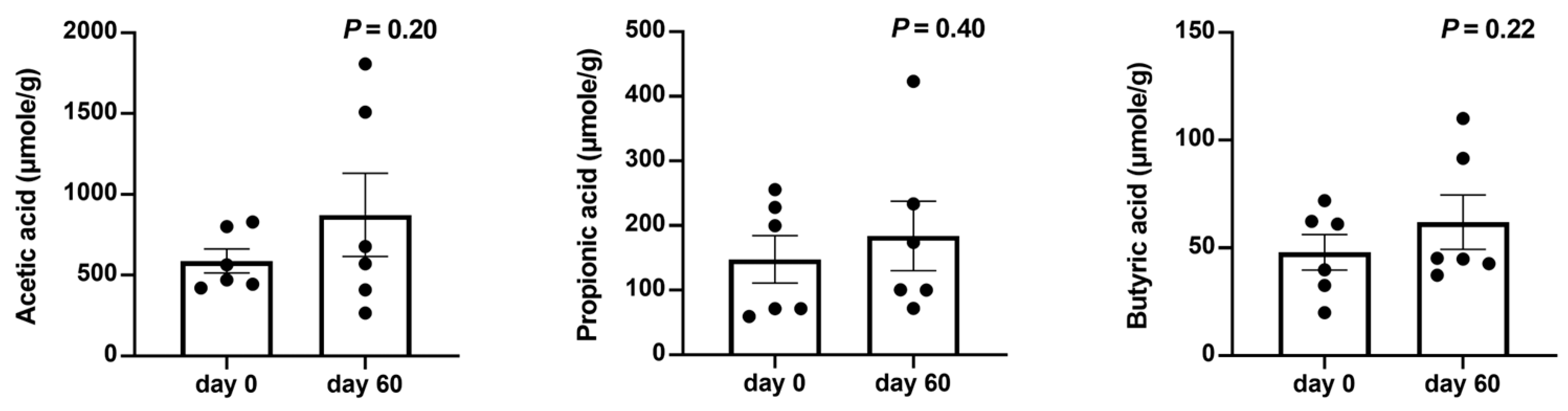
Figure 6.
Spearman’s rank correlation. Each cell was colored according to the correlation results. CADESI represneted Canine Atopic Dermatitis Extent and Severity Index. *p < 0.1.
Figure 6.
Spearman’s rank correlation. Each cell was colored according to the correlation results. CADESI represneted Canine Atopic Dermatitis Extent and Severity Index. *p < 0.1.
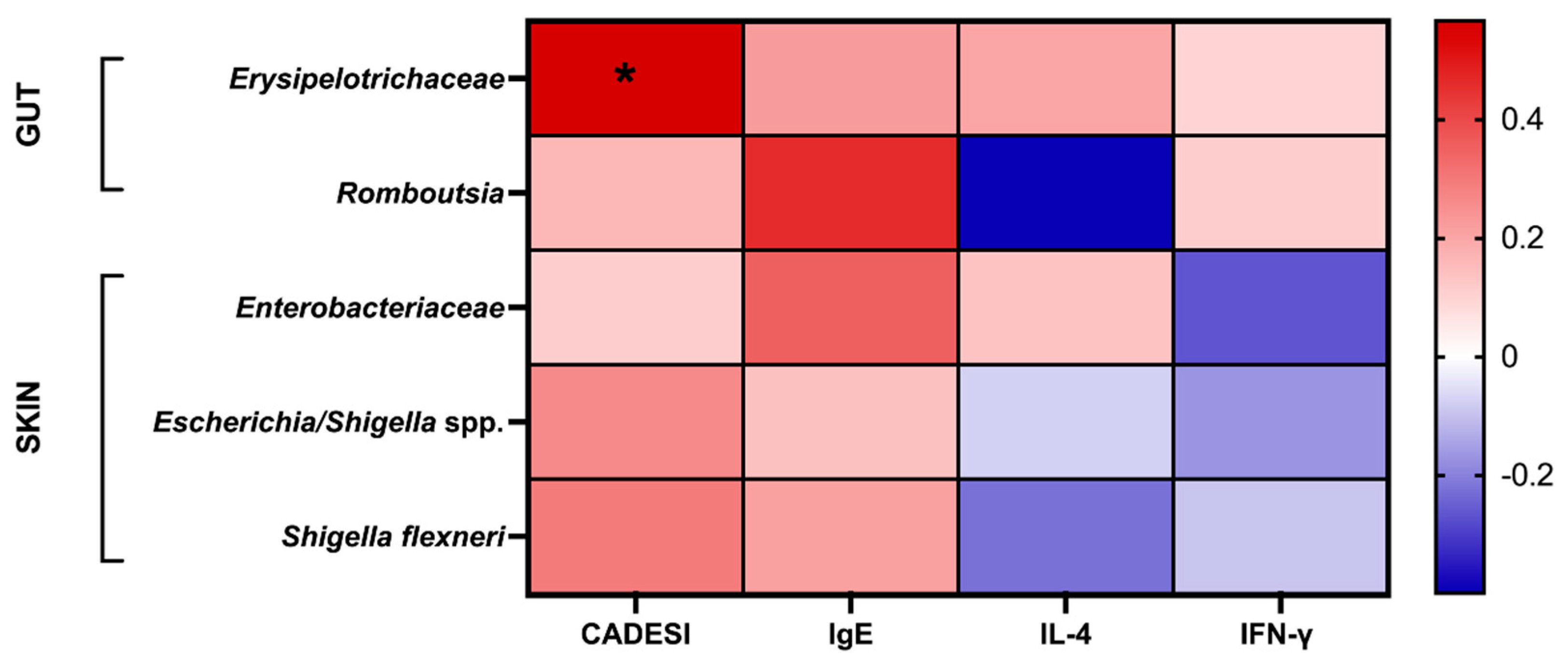
Figure 7.
Effects of LCP on the microbiota of high responder (HR) dogs with atopic dermatitis (AD). (A) Grouping criteria for the HR and moderate responder (MR) groups, (B) Fecal microbiota analysis, (C) Skin microbiota analysis, and (D) The relative abundance of LCP.
Figure 7.
Effects of LCP on the microbiota of high responder (HR) dogs with atopic dermatitis (AD). (A) Grouping criteria for the HR and moderate responder (MR) groups, (B) Fecal microbiota analysis, (C) Skin microbiota analysis, and (D) The relative abundance of LCP.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



