
Submitted:
03 May 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
The objective of this study was to evaluate the genetic effects involved on pepper fruit post-harvest water loss, and correlated to morphology traits. Fruits of eight landraces of C. baccatum, and their 28 hybrids, were evaluated in a randomized complete block design. Analysis of variance, diallel analysis, phenotypic and genotypic correlation were performed to compare fruit water loss (WL), width (FW), length (FL), total soluble solids (TSS), dry matter content (DMC), pericarp thickness (PT), cuticle thickness (CT), and exocarp thickness (ET). WL varied from 14 to 68% during storage. Non-additive effects played more important role than additive, including CT, EP and TSS. Effective strategies to reduce WL should be achieved by repeated backcrosses followed by selection in segregating populations (SP). Therefore, the selection of hybrids should follow high specific combining ability traits. Selecting fruits with higher CT and lower TSS values are indirectly selecting fruits with lower WL. Varieties with low WL can be breed using landraces 04 and 58 as parents or by the hybrid 04 x 44 in open lines in SP. Brazilian C. baccatum landraces are a source of genetic variability, and the SP emerging from crossing lines with reduced WL should develop new ways for conventional breeding.
Keywords:
shelf life
; genetic effects
; diallel
; narrow sense heritability
1. Introduction
Worldwide only five of the 31 species of Capsicum are commonly cultivated for commercial purposes [1]. The five domesticated species are C. annuum, C. baccatum, C. chinense, C. frutescens, and C. pubescens [2]. These species are commercialized as fresh fruits, as raw material for the processing food industry, or as ornamental and medicinal plants [3,4,5,6]. However, pepper grown for fresh market is, particularly, constantly affected by fruit water loss, which is the major factor limiting the shel-life of the fruit [7,8].
Water loss on pepper is naturally controlled by a hydrophobic cuticle barrier presented in the exocarp of the fruits [7,8]. Studies reported that pepper fruit cuticle and/or exocarp thickness varies among cultivars, which directly influence fruit quality, and the during storage [9,10,11,12,13,14].
Previous analysis of 50 accessions from diverse sources of peppers around the world indicated that water loss is reduced in accessions belonging to C baccatum, particularly those from Brazil [16]. C. baccatum and C. chinense are the predominant commercial hot pepper species sold in this country [17]. The great variability on fruit traits including color, shape, length and pungency, between and within these two species, still unknown [2,12,17]. The understanding of the relationships between post-harvest traits and specific chemical components and/or structural features of fruit cuticles is still very preliminary [9].
Populations from a biparental cross, such as recombinant inbred lines (RIL), backcross inbred lines (BIL), and near-isogeneic inbred lines (NIL), has been generated to the study of quantitative traits such as cuticle-associate traits in different environmental conditions [18,20]. Conversely, random intercrossing of multiple founder genotypes produces multiparent advanced population, as diallel cross-followed by successive selfing [18,19]. The resultant populations of a multiparent advanced generation intercross (MAGIC) have multiallelic states of each gene, consequently, higher genetic and phenotypic variability, as well as higher resolution for QTL mapping compared to biparental populations [19].
Wild and/or landraces are important sources to provide information about the diversity of cuticle morphology. Several germplasm studies have been conducted in pepper to evaluate natural variation in water loss rates and to understand its relationship with cuticle components and properties [15]. However, few genetic studies have worked with multiway cross to evaluate postharvest water loss and to characterize the variability found within and among populations of Capsicum species [6,19]. Despite all efforts made by researchers around the world, genetic studies on post-harvest water loss are still limited in pepper and more effort are necessary to understand the genetic effects involved in determination of this trait and to allow germplasm screening for accessions with enhanced postharvest characters [5,8,9,15].
Expanding germplasm surveys to identify new beneficial genetic sources and QTL studies is necessary to expedite postharvest breeding. Water loss rate is one of the most important factors limiting post-harvest storage of Capsicum fruit and improved shel life is a major target for pepper production. Thus, by reducing water loss rate of the fruit it should improve peppers storage potential. Significant progress has been made in enhancing pepper-fruit yield and quality, but not for postharvest improvement of pepper fruit [15].
This study is the first one using mutiparental population to elucidate the inheritance of water loss and provide new material for use in breeding programs for extending fruit shelf life.
Thus, the objective of this study was to evaluate the genetic effects involved on postharvest water loss of C, baccatum fruits and to correlate fruit morphologic traits to water loss.
2. Materials and Methods
Eight landraces of C. baccatum, such as UFV 04, UFV 24, UFV 38, CB 44, CB 46, CB 50, CB 56 and CB 58, from the Federal University of Viçosa - Horticultural Germplasm Bank, were selected according to genetic background and phenotypic diversity (Table 1) and crossed in a complete diallel at the Garden Field of Federal University of Viçosa, Minas Gerais, Brazil. The F1 seeds of the 28 hybrids and eight parents were planted in the field in a randomized complete block design (r = 3). Experimental plots were comprised by 10 pepper plants with an in-row spacing of 1.0 m and a row spacing of 1.0 m.
In this study, pepper fruit were harvested at maturity and four fresh healthy fruits per replication, totalizing 12 fruits per treatment. The water loss experiment was conducted twice in time. Once significance was not detected to interaction (genotype x time) the following analyses was performed using the mean of the two experiments. Then, each mean from 24 fruits per treatment were used. The fruits were washed with distilled water, air dried, and packed in low-density polystyrene (17.5 × 20 × 0.5 cm) and unpacked kept at 20±1ºC for 9 days in the Laboratório de Análise de Progênies of the Federal University of Viçosa.
Water loss of each fruit was measured as the difference of fruit weight before and after storing. In addition, the fruit width, fruit length, total soluble solids, fruit dry matter content, and pericarp thickness was measured in each fruit of all plots following the Capsicum descriptors [21]. The cuticle thickness and exocarp thickness were measured using a light microscopy and a 5 mm diameter cork borer, in which longitudinal sections were cut on a manual microtome and measurements performed under the light microscopy with a ocular-micrometer scale.
Statistical Analysis
Data were subjected to analysis of variance using the software Genes38, when the F value was significant, a multiple means comparison was performed using the Scott–Knott test adjusted at a P value of 0.01.
Genetical analyses were performed using the software Genes [22], in which the diallel analysis was performed to estimate general combining ability (GCA) and specific combining ability (SCA) effects using the procedure described by Griffing [23], Model II, fixed model. The following statistical model was used: Xijk = u + gi + gj + sij+ + eijk, where Xijk is the observation value for a cross between the parent ith and jth in the kth replication; u is the general population means; gi and gj are the GCA values of the ith and jth parents, respectively; sij is the SCA value for the hybrid between the parent ith and jth; and eijk is the error. Significant differences among GCA effects and SCA effects were tested using F values.
A one-way multivariate analysis of variance model was also fitted to estimate genotypic covariance components using the method of moments. Phenotypic and genotypic correlation matrices were obtained and displayed in a weighted correlation network diagram [24]. To determine direct and indirect effects of fruit descriptors over the fruit water loss, a path analysis model was fitted. Analyses were carried with the software R.
3. Results
Based on analysis of variance (Table 2), there was significant difference among landraces and/or hybrids for all evaluated traits, while the general combining ability (GCA) of traits was significant for all variables, except for exocarp thickness. The Specific combining ability (SCA) variance was also significant for all traits, except for water loss and fruit dry matter content. Particularly, the non-additive effects played more important role than additive effects, including cuticle thickness, exocarp thickness and total soluble solids. According to the Scott-Knott test, the highest variability was measured for the exocarp thickness and fruit length (six groups), followed by the fruit width (five groups), total soluble solids and fruit wall thickness (four groups), dry matter content (three groups), water loss and cuticle thickness (two groups) (Table 3).
Water loss:parents 46 and 56 had significant positive general combining ability for water loss according to estimates for the combining ability effects (ĝi). Contrarily, parents 4 and 58 had a negative general combining ability for water loss. The minimum specific combining ability effect values (negative) for water loss were obtained by the hybrid 4 x 44. This result was enhanced by the SCA analyses, in which the hybrid 4 x 44 had the most reduced water loss (Table 4).
Cuticle thickness: parents 4 and 58 had significant positive values of ĝi, and parents 38 and 46 negative values of ĝi. Particularly, the Sij values for this trait had the hybrids 4 x 38, 4 x 44, 24 x 38 and 24 x 56 as the major significant positive values and the hybrid 24 x 58 as the major significant negative values (Table 4).
Exocarp thickness: major significant positive values of ĝi for the exocarp thickness was measured for parents 4, 46, and 50, while the parent 38 had a significant negative value of ĝi for the thickness of exocarp. In general, hybrids had a significant positive Sij (Table 4).
Fruit width: parents 4 and 50 had significant positive ĝi effects for fruit width, contrarily parents 44, 46, 56, and 58 had significant negative ĝi effects for fruit width. Hybrids 24 x 46, 44 x 50, 50 x 56, and 50 x 58 were positives, except by the parents 4 x 38 that had negatives values of Sij (Table 4).
Fruit length: parents 24 and 38 had significant positive values for fruit length, contrarily, parents 46 and 56 had significant negative values. The major positive values of Sij were found in the hybrids 4 x 56, 4 x 58, 24 x 38, and 44 x 58, while the hybrids 24 x 56, 50 x 58, 24 x 46, and 38 x 58 had negative values.
Total soluble solids: significant positive value of total soluble solids were measured for parents 24, 46, and 56. All other parents had significant negative values. Significant values of Sij were measured positive in the hybrids 4 x 44, 4 x 56, 24 x 44, 24 x 56, 46 x 56, 50 x 56 and 50 x 58 and 4 x 46, 56 x 58 and 24 x 58 (negative) (Table 4).
Fruit dry matter content: parents 46 and 56 showed significant positive values, while parents 4, 24, 38 and 50 had negative effect. The significant Sij values were found for hybrids 24 x 56, 46 x 56, 46 x 58 and 4 x 50 (positive) and hybrids 50 x 56 and 50 x 58 (negative) (Table 4).
Overall, there was a cluster of phenotypic and genotypic correlations between dry matter content, total of soluble solids, wall thickness, fruit width, and water loss (Figure 1; Table 5). The fruit width and fruit wall thickness had strong negative correlations (-0.5 and -0.52) with water loss, respectively. Cuticle thickness and water loss had a moderate negative correlation (-0.38). Dry matter content and water loss had strong positive correlations (0.94).
The coefficient of determination of the model for the path analysis was high (R2 = 0.79) and despite the high positive correlation with water loss, dry matter content had a low negative direct effect (path coefficient) indicating no cause/effect correlation (Table 5). Total soluble solids had high positive path coefficient, contrarily, the cuticle thickness and fruit width had a high negative direct effect value, which classify these traits as the main determinant of water loss (Table 5).
4. Discussion
Genetic diversity among genotypes of different landraces can be used to improve the fruit quality of commercial pepper [12,25]. The variation among C. baccatum landraces and their hybrids for fruit width, fruit length, total soluble solids, fruit dry matter content, and pericarp thickness measured in the present study substantiate this statement, particularly for fruit water loss. The high variability among genotypes for quantitative traits can be used to introduce variability in breeding programs (Table 3). Furthermore, significant differences were found for water loss (WL) showing selection for good progenitors and to explore the hybrid vigor, which can be efficient and much less expensive on the reduction of fruit postharvest water loss. Coupled with the high narrow sense heritability values observed for WL, the existing variability detected among genotypes allows for gains when selection is practiced in an early generation [12).
Previous studies showed variation for pepper fruit length is controlled by genes acting additively and non-additively [26,27]. Other works showed that additive variation is predominant for fruit length [3,26,28,29,30], fruit width [27,30] and for fruit wall thickness [29]. On the other hand studies showing dominant variation is predominant gene effect for fruit width [12], TSS [3,12], dry matter content and fruit wall thickness [3,10,27] in peppers.
The loss in relative water content differs in each cultivar examined in pepper fruit. [31]. According with previous study developing pepper types with greater amounts of epicuticular wax will provide an approach for extending their postharvest shel-life [32]. Significant correlations between water loss in Capsicum sp. fruit with wax and cutin monomers composition also provides cultivar-related differences in cuticle functionality [33].
The higher GCA ()/SCA () ratio indicates the predominance of additive gene effect for all traits, except for the cuticle thickness, exocarp thickness and Total Soluble Solids (Table 2), showing that non-additive (dominance or epistasis) gene action was important for controlling these three last traits. The magnitudes of GCA and SCA effects are indicative of the relative importance of additive and non-additive (dominance or epistasis), gene action in the inheritance of a trait, respectively [23]. Previous studies showed that variation for pepper fruit length controlled by genes acting additively and non-additively [26,27]. On the other hand additive variation is predominant for fruit length [3,27,28,29,30] and for fruit wall thickness [29]. instead a dominant variation is predominant for genes effecting fruit width [12], TSS [3,12], dry matter content and fruit wall thickness [3,12,27] in peppers.
High values of narrow sense heritability measured for water loss can be useful in breeding programs, particularly when selection is carried on in early generations [34]. Furthermore, the GCA () / SCA () ratio indicated the predominance of additive gene effect for all traits, except for the cuticle thickness, exocarp thickness, and total soluble solid, which required a non-additive (dominance or epistasis) gene action. Magnitudes of GCA and SCA effects were an indicative of the relative importance of additive and non-additive (dominance or epistasis) gene action in the inheritance of a trait [23]. The significant additive gene effects in this study demonstrated that an effective way to reduce water loss and improve fruit width, fruit length, dry matter content, and fruit wall thickness should be achieved through repeated backcrosses and selection of desirable recombinants from segregating populations. Therefore, the selection of hybrids should follow the high specific combining ability effects coming from a parent with high GCA effects [12,23]. Two additive QTL controlling fruit post-harvest water loss in a NIL F2 population were identified [20]. Traits determined by additive gene effects make its selection effective in initial segregating generations of breeding programs based on hybridization methods [12]. Several studies have revealed how variation in cuticle properties in pepper fruits is under genetic control and others have implicated cuticular wax thickness as the main barrier to transpiration [8,9,34,35,36,37,38].
To consider cuticle thickness, exocarp thickness, and TSS, the results suggest the possibility of vigor hybrid exploitation based on the significant non-additive effects found for these traits [12] Hybrids with good sij to these traits (Table 1) and at least one of the parents with elevated significant GCA (Table 4), must be selected to C. baccatum breeding. Selecting hybrids with high specific combining ability effects, and at least one parent with high or average GCA effects for a particular trait, is a good strategy for plant breeding [12,23].
The traits, fruit length and exocarp thickness, with low correlation and/or coefficient path values cannot be used to obtain satisfactory genetic gains in WL. On the other hand it is possible to obtain gains selecting those traits with significant correlation and with the same sign of path coefficient. This fact evidential that selecting fruits with higher cuticle thickness and fruit width values and lower TSS values we are indirectly selecting fruits with lower WL, which can be used as a criterion to help in indirect selection. Previous work with a biparental population of C. annuum did not determinate the inheritance of water loss because it was used non divergent parents for this trait [39]. On the other hand, a QTL study mapped genes that control natural variation in post-harvest fruit water loss in an inter-specific cross of C. annuum × C. chinense [20] showing FWL is a quantitative inherited trait corroborating the findings of this work.
The genetic diversity within these domesticated species has been explored in plant breeding programs. Tropical deforestation is among the most massive and urgent environmental problems facing Capsicum germplasm resources [40]. The expansion of agribusiness in many locations around the world, as in some states of Brazil, could lead to extinction of landraces of several chili endemic species, like C. baccatum. Furthermore, the indiscriminate expansion of sugar cane, cotton, coffee and soybean plantations led to the reduction of the Atlantic Forest in Brazil, reducing the genetic pool for the non-domesticated Capsicum species before researchers even had the chance to evaluate them [6,41].
This study was the first one screening the fruit water loss in Brazilian C. baccatum landraces and it showed low water loss varieties could be developed from the original landraces held in UFV germplasm bank. Overall, breeding programs seeking for reduce water loss on post harvesting management of pepper should indirectly select for fruit with thicker cuticle, larger width, and higher total soluble solids. Contrarily, fruit length and exocarp thickness will have no effect on water loss. Varieties with low water loss can be breed using the landraces 04 and 58 as parents or using the hybrid 04 x 44 for open lines in segregate populations.
5. Conclusions
Fruit water loss is a major concern on post-harvest longevity for pepper fresh market. The multiparent populations developed in this study represent a unique material to use in breeding programs with the goal of extend shelf life of Capsicum fruits. The identification of lines and hybrids with genetically elevated total soluble solids and cuticle thickness provide genetic variability for improve pepper varieties shelf life. In addition, the Brazilian pepper landraces of C. baccatum species are a source of genetic variability for plant breeders, and the new segregating populations emerging through the crossing of pepper lines with reduced water loss should be developed, opening new ways for conventional breeding. Therefore, the pepper industry and consumers could benefit significantly from newly developed varieties with improved postharvest qualities.
Author Contributions
Rego ER and Finger FL designed the field research. Rego ER also conducted the field experiments and did the genetic analysis. Silva AR performed the correlation and path statistical analysis, Azevedo AA, Meira RMSA, and Rego MM did the anatomy analysis. Rego ER, Pessoa AMS, da Silva ALBR, Silva, R.S, and Rego MM1 analyzed the data and wrote the manuscript. All authors read and approved the manuscript.
Funding
This work was supported by the National Council for Scientific and Technological Development-Brazil (CNPq).
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Acknowledgments
Authors gratefully acknowledge the Federal University of Viçosa - Horticultural Germplasm Bank for providing the landraces and the National Council for Scientific and Technological Development-Brazil (CNPq).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moscone, E.A.; Scaldaferro, M.A.; Grabiele, M.; Cecchini, N.M.; García, Y.S.; Jarret, R.; Davinã, J.R.; Ducasse, D.A.; Barbosa, G.E.; Ehrendorfer, F. The evolution of the chili pepper (Capsicum – Solanaceae): a cytogenetic perspective. Acta Hortic. 2007, 745, 137–169. [Google Scholar] [CrossRef]
- Pickersgill, B. Genetic resources and breeding of Capsicum spp. Euphytica 1997, 96, 129–133. [Google Scholar] [CrossRef]
- Nascimento, N.F.F.; Rêgo, E.R.; Nascimento, M.F.; Bruckner, F.L.; Finger, F.L.; Rêgo, M.M. Combining ability for yield and fruit quality in the pepper Capsicum annuum. Genet. Mol. Res. 2014, 13, 3237–3249. [Google Scholar] [CrossRef] [PubMed]
- Rêgo, E.R.; Rêgo, M.M.; Finger, F.L. Methodological basis and advances for ornamental pepper breeding program in Brazil. Acta Hortic. (ISHS) 2015, 1087, 309–314. [Google Scholar] [CrossRef]
- Rêgo, ER.; Rêgo, M.M. Genetics and Breeding of Chili Pepper Capsicum spp., in Production and Breeding of Chilli Peppers (Capsicum spp.). ed. Rêgo, E.R.; Rêgo, M.M.; Finger, F. L. Springer International Publishing, Switzerland, 2016, 1-129.
- Rêgo, E.R.; Rêgo, M.M. Ornamental Pepper, in Ornamental Crops. Ed. Van Huylenbroeck, J. Springer International Publishing, Switzerland, 2018, 529-565.
- Tafolla-Arellano, J.C.; Báez-Sañudo, R.; Tiznado-Hernández, M.E. The cuticle as a key factor in the quality of horticultural crops. Scientia Horticulturae 2018, 232, 145–152. [Google Scholar] [CrossRef]
- Wang, J.; Shan, Q.; Yi, T.; Ma, Y.; Zhou, X.; Pan, L.; Liu, F. Fine mapping and candidate gene analysis of CaFCD1 affecting cuticle biosynthesis in Capsicum annuum L. Theoretical and Applied Genetics 2023, 136, 46. [Google Scholar] [CrossRef] [PubMed]
- Lara, I.; Heredia, A.; Domínguez, E. Shelf life potential and the fruit cuticle: the unexpected player. Frontiers in Plant Science 2019, 10, 460894. [Google Scholar] [CrossRef]
- Banaras, M.; Lownds, N.K.; Bosland, P.W. Relationship of physical properties to postharvest water loss in pepper fruits (Capsicum annuum L.). Pakistan J. Bot. 1994, 26, 321–326. [Google Scholar]
- Maalekuu, K.; Elkind, Y.; Tuvia-Alkalai, S.; Shalom, Y.; Fallik, E. Quality evaluation of three sweet pepper cultivars after prolonged storage. Advances in horticultural science 2003, 17, 1000–1005. [Google Scholar]
- Rêgo, E.R.; Rêgo, M.M.; Finger, F.L.; Cruz, C.D.; Casali, V.W.D. A diallel study of yield components and fruit quality in chilli pepper (Capsicum baccatum). Euphytica 2009, 168, 275–287. [Google Scholar] [CrossRef]
- Martínez, S.; Curros, A.; Bermúdez, J.; Carballo, J.; Franco, I. The composition of Arnoia peppers (Capsicum annuum L.) at different stages of maturity. International journal of food sciences and nutrition 2007, 58, 150–161. [Google Scholar] [PubMed]
- Ilić, Z.S.; Sunic, L.; Mirecki, N.; Fallik, E. Cultivars differences in keeping quality and bioactive constituents of bell pepper fruit during prolonged storage. Journalof Advances in Biotechnology 2014, 31, 313–318. [Google Scholar] [CrossRef]
- Ziv, C.; Lers, A.; Fallik, E.; Paran, I. Genetic and biotechnological tools to identify breeding targets for improving postharvest quality and extending shelf life of peppers. Current Opinion in Biotechnology 2022, 78, 102794. [Google Scholar] [CrossRef] [PubMed]
- Parsons, E.P.; Popopvsky, S.; Lohrey, G. T.; Alkalai-Tuvia, S.; Perzelan, Y.; Bosland, P.; Bebeli, P. J.; Paran, I.; Fallik, E.; Jenks, M. Fruit cuticle lipid composition and water loss in a diverse collection of pepper (Capsicum). Physiologia plantarum 2013, 149, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Lannes, S.D.; Finger, F.L; Schuelter, A.R.; Casali, V.W.D. Growth and quality of Brazilian accessions of Capsicum chinense fruits. Scientia Horticulturae 2007, 112, 266–270. [Google Scholar] [CrossRef]
- Bazakos, C.; Hanemian, M.; Trontin, C.; Jiménez-Gómez, J.M.; Loudet, O. New strategies and tools in quantitative genetics: how to go from the phenotype to the genotype. Annual review of plant biology 2017, 68, 435–455. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Bres, C.; Mauxion, J.; Bakan, B.; Rothan, C. Breeding for cuticle-associated traits in crop species: traits, targets, and strategies. Journal of experimental botany 2017, 68, 5369–5387. [Google Scholar] [CrossRef] [PubMed]
- Popovsky-Sarid, S.; Borovsky, Y.; Faigenboim, A.; Parsons, E.P.; Lohrey, G.T.; Alkalai-Tuvia, S.; Fallik, E.; Jenks, M.A.; Paran, I. Genetic and biochemical analysis reveals linked QTLs determining natural variation for fruit postharvest water loss in pepper (Capsicum). Theor Appl Genet. 2017, 130, 445–459. [Google Scholar] [CrossRef] [PubMed]
- IPGRI. Descriptors for Capsicum. Rome, International plant genetic resources institute, 1995; 49p.
- Cruz, C. D. Genes: a software package for analysis in experimental statistics and quantitative genetics. Acta Scientiarum Agronomy 2013, 35, 271–276. [Google Scholar] [CrossRef]
- Griffing, B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Silva, A.R.; Rêgo, E.R.; Pessoa, A.M.S.; Rêgo, M.M. Correlation network analysis between phenotypic and genotypic traits of chili pepper. Pesquisa Agropecuária Brasileira 2016, 51, 372–377. [Google Scholar] [CrossRef]
- Rêgo, E.R.; Rêgo, M.M.; Cruz, C.D.; Finger, F.L.; Casali, V.W. Phenotypic diversity, correlation and importance of variables for fruit quality and yield traits in Brazilian peppers (Capsicum baccatum). Genet. Resour. Crop. Evol. 2011, 58, 909–918. [Google Scholar] [CrossRef]
- Legg, P.D.; Lippert, L.F. Estimates of genetic and environmental variability in a cross between two strains of pepper (Capsicum annuum L.). Proc. Am. Soc. Hort. Proc. Sci. 1966, 89, 443–448. [Google Scholar]
- Marin, O.; Lippert, L.F. Combining ability analysis of anatomical components of the dry fruit in chili pepper. Crop Sci. 1975, 15, 326–329. [Google Scholar]
- Ahmed, N.; Tanki, M.I.; Jabeen, N. Heterosis and combining ability studies in hot pepper (Capsicum annuum L.). Applied Biological Research 1999, 1, 11–14. [Google Scholar]
- Santos, R.M.C.; Rêgo, E.R.; Borém, A.; Nascimento, M.F.; Nascimento, N.F.F.; Funger, F.L.; Rêgo, M.M. Epistasis and inheritance of plant habit and fruit quality traits in ornamental pepper (Capsicum annuum L.). Genetics and Molecular Research 2014, 13, 8876–8887. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.Q.; Rodrigues, R.; Bento, C. S.; Pimenta, S. Heterosis and combining ability for ornamental chili pepper. Horticultura Brasileira 2017, 35, 349–357. [Google Scholar] [CrossRef]
- Lownds, N.K.; Banaras, M.; Bosland, P.W. Postharvest water loss and storage quality of nine pepper (Capsicum) cultivars. HortScience 1994, 29, 191–193). [Google Scholar] [CrossRef]
- Banaras, M.; Lownds, N.K.; Bosland, P.W. Relationship of physical properties to postharvest water loss in pepper fruits (Capsicum annuum L.). Pakistan Journal of Botany 1994, 26, 321–326. [Google Scholar]
- Parsons, E.P.; Popopvsky, S.; Lohrey, G.T.; Lu, S.; Alkalai-Tuvia, S.; Perzelan, Y.; Paran, I.; Fallik, E.; Jenks, M.A. Fruit cuticle lipid composition and fruit post-harvest water loss in an advanced backcross generation of pepper (Capsicum sp.). Physiologia plantarum 2012, 146, 15–25. [Google Scholar] [CrossRef]
- Silva-Neto, J.J.; Rêgo, E.R.; Nascimento, M.F.; Silva Filho, V.A.L.; Almeida Neto, J. X.; Rêgo, M.M. Variabilidade em população base de pimenteiras ornamentais (Capsicum annuum L.). Rev. Ceres 2014, 61, 84–89. [Google Scholar] [CrossRef]
- Lownds, N.K., Banaras, M.; Bosland, P.W. Relationships between postharvest water loss and physical properties of pepper fruit (Capsicum annuum L.). HortScience 1993, 28, 1182-1184.
- Kissinger, M.; Tuvia-Alkalai, S.; Shalom, Y.; Fallik, E.; Elkind, Y.; Jenks, M.A.; Goodwin, M.S. Characterization of Physiological and Biochemical Factors Associated with Postharvest Water Loss in Ripe Pepper Fruit during Storage. J. Amer. Soc. Hort. Sci. 2005, 130, 735–741. [Google Scholar] [CrossRef]
- Kissinger, M. et al. Characterization of physiological and biochemical factors associated with postharvest water loss in ripe pepper fruit during storage. Journal of the American Society for Horticultural Science. 130, 735-741 (2005).
- Arumugam, V.; Balamohan, T.N. Wax coating affects postharvest shelf-life of non-cooled sweet pepper. Journal of Spices and Aromatic Crops 2014, 23, 98–101. [Google Scholar]
- Vilarinho, L.B.O. Silva, D.J.H.; Greene, A.; Salazar, K.D.; Alves, C.; Eveleth, M.; Nichols, B.; Tehseen, S.; Khoury Jr, J. K.; Johnson, J.V.; Sargent, S.A.; Rathinasabapathi, B. Inheritance of fruit traits in Capsicum annuum: Heirloom cultivars as sources of quality parameters relating to pericarp phape, color, thickness, and total soluble solids. Journal of the American Society for Horticultural Science. 140, 597-604 (2015).
- Bosland, P.W. Capsicums: innovative use of an ancient crop. In: Janick, J. (Ed.) Progress in New Crops. ASHS Press, Arlington, 2016, 479-487.
- Rêgo, E.R.; Finger, F.L.; Rêgo, M.M. “Types, Uses and Fruit Quality of Brazilian Chili Peppers”, in Spices: Types, Uses and Health Benefits. Ed. Johnathan F. (Nova Sci. Pub.), 2012, 1-70.
Figure 1.
Correlation network for phenotypic (A) and genotypic(B) for water loss of chili peppers. (CT = cuticle thickness; ET = exocarp thickness; FW = fruit width; FL = fruit length; TSS = total soluble solids; DM = dry matter content; FWT = fruit wall thickness; WL = water loss).
Figure 1.
Correlation network for phenotypic (A) and genotypic(B) for water loss of chili peppers. (CT = cuticle thickness; ET = exocarp thickness; FW = fruit width; FL = fruit length; TSS = total soluble solids; DM = dry matter content; FWT = fruit wall thickness; WL = water loss).
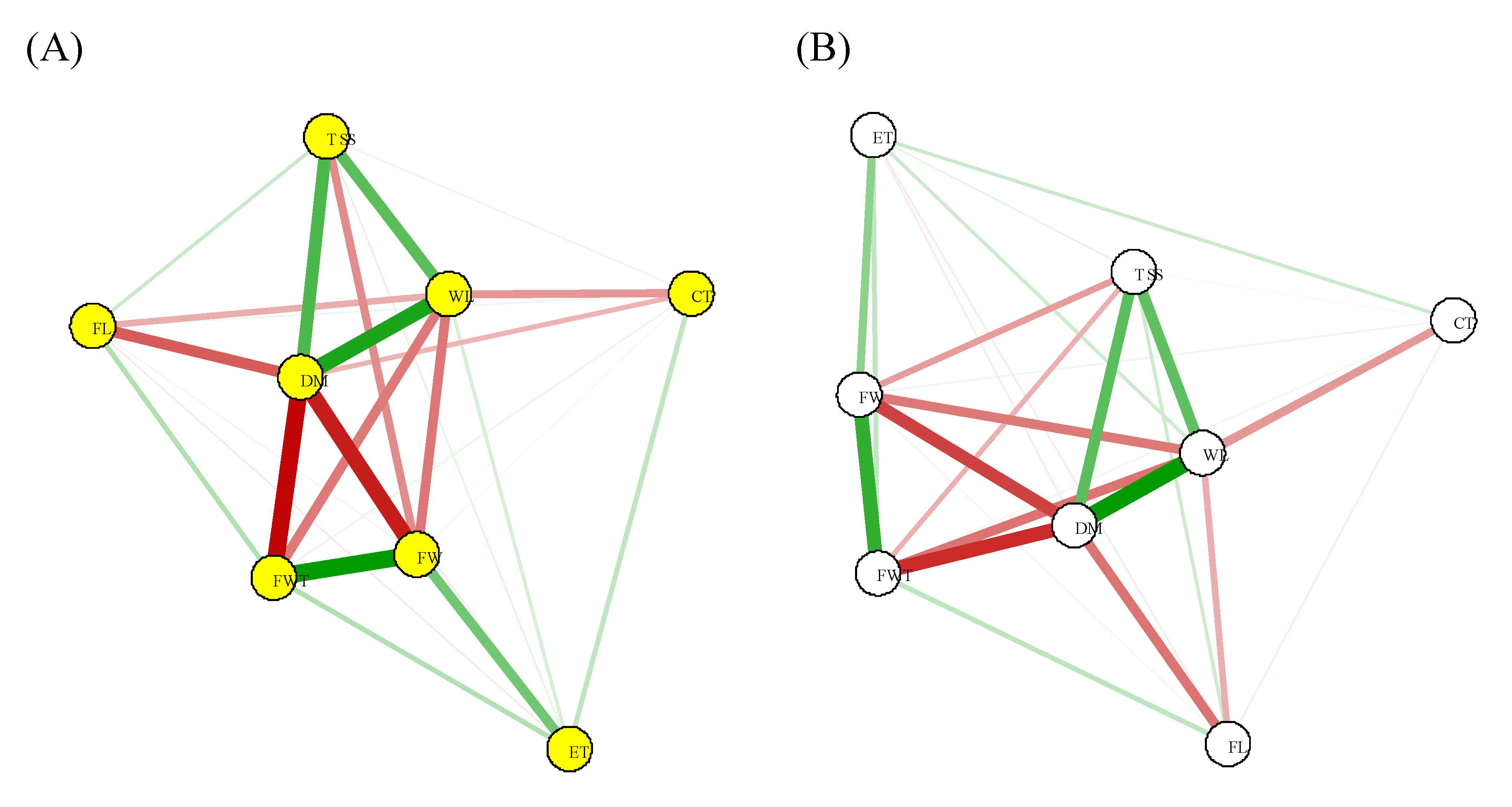

Table 1.
Fruit traits for eight parents of Capsicum baccatum var pendulum used in the diallel cross.
Table 1.
Fruit traits for eight parents of Capsicum baccatum var pendulum used in the diallel cross.
| Parents | Color | Cuticle thickness | Exocarp thickness | Width | Length | Total soluble solids | Dry matter content | Wall thickness |
|---|---|---|---|---|---|---|---|---|
| µm | µm | mm | mm | % | % | mm | ||
| UFV - 04 | red | 25 | 145 | 53 | 46 | 9.6 | 14 | 3 |
| UFV - 24 | red | 22 | 110 | 32 | 142 | 10.2 | 15 | 2.3 |
| UFV - 38 | red | 22 | 65 | 19 | 69 | 8.7 | 15 | 1.7 |
| UFV - 44 | yellow | 25 | 95 | 15 | 74 | 9.4 | 17 | 2.3 |
| UFV - 46 | red | 20 | 90 | 14 | 57 | 9.6 | 23 | 1.5 |
| UFV - 50 | red | 25 | 130 | 37 | 44 | 8.8 | 18 | 2.9 |
| UFV - 56 | red | 25 | 95 | 10 | 47 | 12.9 | 28 | 0.9 |
| UFV - 58 | red | 35 | 105 | 17 | 66 | 10.7 | 16 | 1.9 |
| CV (%) | 18.1 | 23.6 | 59.8 | 46.8 | 13.5 | 24.4 | 34.3 |
CV = Coeficient of variation.
Table 2.
Analysis of variance (mean squares) and quadratic components of GCA ability () and SCA ability () for fruit traits of 8 x 8 diallel cross in pepper (Capsicum baccatum).
Table 2.
Analysis of variance (mean squares) and quadratic components of GCA ability () and SCA ability () for fruit traits of 8 x 8 diallel cross in pepper (Capsicum baccatum).
| SV | DF | Water loss | Cuticle thickness | Exocarp thickness | Fruit width | Fruit length | Total soluble solids | Dry matter content | Fruit wall thickness | |
|---|---|---|---|---|---|---|---|---|---|---|
| % | µm | µm | mm | mm | % | % | mm | |||
| Treatment | 35 | 373.97** | 43.83** | 1,073.92** | 202.83** | 1,355. 95** | 3.83** | 26.86** | 1.20** | |
| GCA | 7 | 1,222. 04** | 85.89* | 1369.19ns | 769.60** | 5,890. 07** | 10.04** | 107.65** | 4.97** | |
| SCA | 28 | 161.95ns | 33.31** | 1,000.10** | 61.13** | 222.43** | 2.35** | 6.67 ns | 0.26** | |
| Error | 36 | 131.05 | 12.67 | 50.35 | 6.91 | 55.75 | 0.32 | 3.94 | 0.05 | |
| 2g | 54.55 | 10.93 | 3,035.35 | 38.13 | 291.71 | 0.48 | 5.18 | 0.24 | ||
| 2s | 15.45 | 48.55 | 35,862.19 | 27.11 | 83.33 | 1.01 | 1.34 | 0.1 | ||
| 2g/ 2s | 3.53 | 0.22 | 0.08 | 1.4 | 3.5 | 0.47 | 3.86 | 2.4 | ||
| Mean | 37.04 | 25.55 | 87.43 | 21.57 | 72.59 | 8.88 | 17.63 | 2.14 | ||
| h2b (%) | 94.01 | 71.08 | 95.31 | 93.2 | 95.88 | 91.73 | 85.32 | 95.19 | ||
| h2n (%) | 63.26 | 21.8 | 21 | 74.6 | 81.7 | 48 | 71.8 | 80.27 |
ns and ** = non-significant and significant (p≤0.01) by F test, respectively. h2b and h2n= broad sense heritability and narrow sense heritability, respectively.
Table 3.
Means of fruit traits evaluated in 8 parents and 28 hybrids F1 of pepper (Capsicum baccatum var pendulum).
Table 3.
Means of fruit traits evaluated in 8 parents and 28 hybrids F1 of pepper (Capsicum baccatum var pendulum).
| Parents/Hybrids | Water Loss | Cuticle thickness | Exocarp thickness | Width | Length | Total soluble solids | Dry matter content | Wall thickness |
|---|---|---|---|---|---|---|---|---|
| % | µm | µm | mm | mm | % | % | mm | |
| 4 | 31.47 b† | 25.00 b | 145.00 a | 62.15 a | 51.10 f | 7.50 c | 13.70 c | 2.80 a |
| 04 x 24 | 18.64 b | 27.50 a | 45.00 f | 25.80 c | 95.25 c | 8.65 d | 14.45 c | 3.15 a |
| 04 x 38 | 33.31 b | 32.50 a | 85.00 d | 31.70 b | 60.75 f | 8.50d | 14.50 c | 2.75 a |
| 04 x 44 | 17.66 b | 35.00 a | 105.00 c | 19.85 d | 61.10 f | 9.50 c | 15.05 c | 2.35 b |
| 04 x 46 | 45.44 a | 25.00 b | 100.00 c | 20.25 d | 55.10 f | 7.60 d | 17.35 c | 2.80 a |
| 04 x 50 | 36.99 b | 30.00 a | 115.00 b | 33.20 b | 52.40 f | 7.90 d | 16.20 c | 3.25 a |
| 04 x 56 | 31.27 b | 30.00 a | 60.00 e | 13.50 e | 59.00 f | 10.20 c | 20.15 b | 1.10 d |
| 04 x 58 | 14.47 b | 30.00 a | 65.00 e | 25.65 c | 84.50 d | 8.30 d | 14.45 c | 2.60 b |
| 24 | 49.25 a | 22.50 b | 110.00 c | 23.75 d | 173.50 a | 11.90 b | 13.70 c | 2.85 a |
| 24 x 38 | 22.60 b | 30.00 a | 90.00 d | 19.70 d | 128.30 b | 7.80 d | 12.60 c | 1.86 c |
| 24 x 44 | 32.74 b | 27.50 a | 70.00 e | 20.55 d | 107.00 c | 10.55 c | 15.80 c | 2.45 b |
| 24 x 46 | 58.28 a | 30.00 a | 70.00 e | 21.35 d | 88.95 d | 8.85 d | 16.80 c | 2.40 b |
| 24 x 50 | 25.17 b | 23.75 b | 120.00 b | 32.50 b | 105.65 c | 9.80 c | 14.45 c | 3.25 a |
| 24 x 56 | 57.77 a | 30.00 a | 95.00 d | 12.20 e | 70.75 e | 11.40 b | 23.25 b | 1.55 c |
| 24 x 58 | 25.67 b | 20.00 b | 75.00 e | 21.30 d | 99.30 c | 7.70 d | 16.00 c | 2.35 b |
| 38 | 31.98 b | 22.50 b | 65.00 e | 19.20 d | 85.40 d | 9.05 c | 16.00 c | 1.70 c |
| 38 x 44 | 46.10 a | 17.50 b | 65.00 e | 16.25 e | 73.05 e | 7.35 d | 18.70 c | 1.65 c |
| 38 x 46 | 43.01 a | 21.25 b | 92.50 d | 17.15 d | 70.60 e | 9.35 c | 19.75 b | 2.30 b |
| 38 x 50 | 25.63 b | 20.00 b | 65.00 e | 26.7 c | 78.40 e | 7.65 d | 14.70 c | 2.25 b |
| 38 x 56 | 36.75 b | 20.00 b | 65.00 e | 14.4 e | 61.75f | 8.70 d | 21.00 b | 1.00 d |
| 38 x 58 | 30.23 b | 22.50 b | 40.00 f | 18.35 d | 68.85 e | 7.50 d | 16.0 c | 1.65 c |
| 44 | 45.58 a | 25.00 b | 95.00 d | 14.00 e | 56.70 f | 7.00 d | 18.60 c | 1.80 c |
| 44 x 46 | 41.63 a | 20.00 b | 82.50 e | 13.80 e | 60.05 f | 9.60 c | 21.35 b | 1.65 c |
| 44 x 50 | 35.11 b | 22.50 b | 102.50 c | 30.75 b | 56.90 f | 7.65 d | 15.35 c | 3.05 a |
| 44 x 56 | 44.48 a | 22.50 b | 85.00 d | 13.30 e | 64.05 f | 9.35 c | 22.75 b | 1.15 d |
| 44 x 58 | 26.46 b | 32.50 a | 90.00 d | 15.50 e | 84.60 d | 8.10 d | 17.10 c | 1.85 c |
| 46 | 62.85 a | 20.00 b | 90.00 d | 12.40 e | 48.15 f | 10.35 c | 22.30 b | 1.70 c |
| 46 x 50 | 40.33 a | 22.50 b | 110.00 c | 25.75 c | 56.65 f | 9.40 c | 18.10 c | 2.90 b |
| 46 x 56 | 62.41 a | 22.5 b | 92.50 d | 10.50 e | 45.70 f | 13.30 a | 27.55 a | 0.85 d |
| 46 x 58 | 27.22 b | 27.50 a | 97.50 d | 13.30 e | 63.45 f | 8.15 d | 22.80 b | 1.40 c |
| 50 | 32.35 b | 25.00 b | 130.00 b | 36.05 b | 66.65 f | 8.20 d | 13.65 c | 3.10 a |
| 50 x 56 | 48.52 a | 20.00 b | 55.00 f | 26.10 c | 47.05 f | 8.35 d | 16.00 c | 3.20 a |
| 50 x 58 | 13.75 b | 27.50a | 75.00 e | 31.40 b | 54.90 f | 8.40 d | 15.85 c | 3.05 a |
| 56 | 67.67 a | 25.00 b | 95.00 d | 7.85 e | 42.40 f | 10.25 c | 26.05 a | 0.50 d |
| 56 x 58 | 36.21 b | 30.00 a | 100.00 c | 13.40 e | 61.80 f | 7.85 d | 16.05 c | 1.10 d |
| 58 | 34.71 b | 35.00 a | 105.00 c | 17.00 d | 73.5 e | 8.20 d | 16.70 c | 2.00 c |
†Values followed by different letters within fruit traits (column) indicate significant difference among Parents/hybrids according to Scott-Knott’s criteria (p ≤ 0.01).
Table 4.
Estimates of general combining effects (gi) and specific combining effects (Sij) for fruit traits of parents and hybrids, respectively, of an 8 x 8 diallel cross in peppers (Capsicum baccatum var. pendulum).
Table 4.
Estimates of general combining effects (gi) and specific combining effects (Sij) for fruit traits of parents and hybrids, respectively, of an 8 x 8 diallel cross in peppers (Capsicum baccatum var. pendulum).
| Water loss | Cuticle thickness | Exocarp thickness | Width | Length | Total soluble solids | Dry matter content | Wall thickness | |
|---|---|---|---|---|---|---|---|---|
| % | µm | µm | mm | mm | % | % | mm | |
| Parents | ||||||||
| 4 | -7.26** | 3.00** | 7.81** | 10.01** | -8.30** | -0.43** | -1. 92** | 0.43ns |
| 24 | 0.59ns | 0.37ns | -0.18ns | 0.67ns | 38.88** | 0.85** | -1.80** | 0.33ns |
| 38 | -3.18ns | -2.12* | -15.43** | -1.15* | 5.92** | -0.50** | -0.95* | -0.25ns |
| 44 | 0.19ns | 0.00ns | 0.31ns | -3.61** | -3.31* | -0.38** | 0.46ns | -0.15ns |
| 46 | 11.05** | -2.12* | 3.81** | -4.72** | -11.65** | 0.70** | 2.96** | -0.16ns |
| 50 | -4.32ns | -1.62ns | 11.56** | 8.43** | -6.81** | -0.44** | -2.10** | 0.78ns |
| 56 | 11.93** | -0.50ns | -4.43** | -7.51** | -15.84** | 0.97** | 4.01** | -0.84* |
| 58 | -9.00** | 3.00** | -3.43* | -2.12** | 1.11ns | -0.76** | -0.71ns | -0.13ns |
| Hybrids | ||||||||
| 4 x 24 | -11.73ns | -1.43ns | -50.05** | -6.45** | -7.92ns | -0.66** | 0.53ns | 0.23ns |
| 4 x 38 | 6.71ns | 6.06** | 5.19ns | 1.26ns | -9.45** | 0.54ns | -0.27ns | 0.42** |
| 4 x 44 | -12.30* | 6.44** | 9.44* | -8.11** | 0.13ns | 1.43** | -1.13ns | -0.06ns |
| 4 x 46 | 4.61ns | -1.43ns | 0.94ns | -6.61** | 2.46ns | -1.55** | -1.33ns | 0.38** |
| 4 x 50 | 11.53ns | 3.06ns | 8.19* | -6.82** | -5.08ns | -0.11ns | 2.56* | -0.11ns |
| 4 x 56 | -10.43ns | 1.94ns | -30.80** | -10.57** | 10.55* | 0.78* | 0.42ns | -0.63** |
| 4 x 58 | -6.30ns | -1.55ns | -26.80** | -3.81* | 19.10** | 0.60ns | -0.56ns | 0.15ns |
| 24 x 38 | -1.86ns | 6.19** | 18.19** | -1.39ns | 10.90* | -1.44ns | -2.29* | -0.38 |
| 24 x 44 | -5.09ns | 1.56 | -17.55** | 1.91ns | -1.16ns | 1.19** | -0.50ns | 0.12ns |
| 24 x 46 | 9.59ns | 6.19** | -21.05** | 3.82* | -10.87* | -1.59ns | -1.99ns | 0.07ns |
| 24 x 50 | -8.15ns | -0.55ns | 21.19** | 1.81ns | 0.97ns | 0.50ns | 0.69ns | -0.01ns |
| 24 x 56 | 8.19ns | 4.56* | 12.19** | -2.54ns | -24.89** | 0.69* | 3.40** | -1.10ns |
| 24 x 58 | -2.95ns | -8.93** | -8.80* | 1.17ns | -13.29** | -1.29** | 0.87ns | -0.002ns |
| 38 x 44 | 12.03ns | -3.43ns | -7.30ns | -0.56ns | -2.14ns | -0.65* | 1.55ns | -0.10ns |
| 38 x 46 | -1.90ns | -0.05ns | 16.69** | 1.45ns | 3.74ns | 0.27ns | 0.10ns | 0.56** |
| 38 x 50 | -3.91ns | -4.30* | -18.55** | -2.13ns | 6.70ns | -0.29ns | 0.08ns | -0.43** |
| 38 x 56 | -9.05ns | -2.93ns | -2.55ns | 1.48ns | -0.92ns | -0.65* | 0.30ns | -0.06ns |
| 38 x 58 | 5.37ns | -3.93* | -28.55* | 0.05ns | -10.77* | -0.13ns | 0.02ns | -0.12ns |
| 44 x 46 | -6.65ns | -3.43ns | -9.05* | 0.56ns | 2.43ns | 0.40ns | 0.30ns | -0.18ns |
| 44 x 50 | 2.18ns | -1.43ns | 3.19ns | 4.35** | -5.57ns | -0.41ns | -0.67ns | 0.27* |
| 44 x 56 | -4.69ns | -2.55ns | 1.69ns | 2.84ns | 10.61ns | -0.11ns | 0.64ns | -0.001ns |
| 44 x 58 | -1.76ns | 3.94* | 5.69ns | -0.33ns | 14.21** | 0.36ns | -0.29ns | -0.006ns |
| 46 x 50 | -3.44ns | 0.69ns | 7.19ns | 0.46ns | 2.52ns | 0.26ns | -0.42ns | 0.13ns |
| 46 x 56 | 2.37ns | -0.43ns | 5.69ns | 1.15ns | 0.60ns | 2.75** | 2.94* | -0.29* |
| 46 x 58 | -11.98ns | 1.06ns | 9.69* | -1.42ns | 1.40ns | -0.68* | 2.91* | -0.45** |
| 50 x 56 | 3.85ns | -3.43ns | -39.55** | 3.59* | -2.89ns | 1.06** | -3.57** | 1.11** |
| 50 x 58 | -9.95ns | 0.56ns | -20.55** | 3.51* | -11.99** | 0.71* | 0.99ns | 0.25** |
| 56 x 58 | -3.76ns | 1.94ns | 20.44** | 1.45ns | 3.94ns | -1.24** | -4.89** | -0.07ns |
ns and ** = non-significant and significant (p≤0.01), by t test, respectively.
Table 5.
Path coefficients of different fruit characters for fruit water loss on pepper.
| CT | ET | FW | FL | TSS | DM | FWT | Genetic Correlation Coefficient | |
| CT | -0.56 | 0.09 | -0.03 | -0.04 | -0.01 | 0.18 | -0.01 | -0.38 |
| ET | -0.11 | 0.47 | -0.25 | 0.04 | 0.06 | 0.03 | -0.05 | 0.19 |
| FW | -0.03 | 0.2 | -0.57 | -0.02 | -0.24 | 0.34 | -0.18 | -0.50 |
| FL | -0.04 | -0.03 | -0.02 | -0.51 | 0.12 | 0.25 | -0.06 | -0.29 |
| TSS | 0.01 | 0.04 | 0.21 | -0.09 | 0.65 | -0.29 | 0.07 | 0.60 |
| DM | 0.21 | -0.03 | 0.4 | 0.27 | 0.39 | -0.48 | 0.18 | 0.94 |
| FWT | -0.03 | 0.11 | -0.44 | -0.12 | -0.19 | 0.38 | -0.23 | -0.52 |
R2 = 0.79. Residue = 0.46 (CT = cuticle thickness; ET = exocarp thickness; FW = fruit width; FL = fruit length; TSS = total soluble solids; DM = dry matter content; FWT = fruit wall thickness).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



